Sharks and skates of Scotland: An overview of the elasmobranch fauna of Scottish seas
SIORC Report
Contents
- Background
- Acknowledgements
- Keywords
- Abbreviations
- Introduction
- Elasmobranchs
- International and National Policy context
- UK elasmobranch fisheries management
- Additional UK conservation measures for Elasmobranchs
- Spatial Management
- Codes of conduct
- Addressing knowledge gaps to aid marine planning
- Purpose of this report
- Data sources
- Elasmobranch ecology and biology
- Species list and Appendices
Background
The SIORC (Sharks, skates and rays In the Offshore Region and Coastal Zone of Scotland and also gaelic for shark, pronounced ‘shirk’) is a Working Group within the Marine Alliance for Science and Technology for Scotland (MASTS), launched in spring 2014. SIORC brings together expertise and research capacity to address knowledge gaps in the understanding of sharks, skates, and rays (elasmobranchs) in Scottish waters. The goal of SIORC is to provide a strong scientific evidence base for stakeholders and policymakers by prioritising and delivering on a research conservation agenda for elasmobranchs in Scottish waters, from the coastal zone to the deep seas.
SIORC held a series of workshops during MASTS’ Annual Science Meetings from 2014 – 2019 to synthesise current knowledge of elasmobranch biodiversity and distribution, life histories, and ecology in Scottish waters. In addition to the workshops, SIORC has also undertaken targeted fieldwork; overhauled and re-launched Scottish elasmobranch datasets; supported an international exchange of researchers between Scotland and Peru; attended and helped coordinate public outreach events, including Celebrating Scotland’s Sharks, an event run by the Scottish Sea Angling Conservation Network (SSACN) in 2014, and the Stirling International Angling Festival in the same year.
This report is a key output from SIORC activities, aimed at enhancing our baseline knowledge of elasmobranch fishes in Scottish waters to inform their conservation and management. It is intended to be a reference document that provides baseline information on each species and identifies important and potential data sources. These data are visualised in their raw form as ‘presence only’; this report did not analyse presence/absence, population status or abundance from these data sources.
To describe the status and population trends of Scottish elasmobranchs, the authors reviewed the IUCN global and European assessments of population trends for the elasmobranch species recorded in Scottish waters. Species with fewer than 10 records in Scottish waters were classified as ‘rare’. Any existing literature describing Scotland-specific population trends was also reviewed (see individual species descriptions for references); the population status of any species with no existing literature describing its population trend was classified as ‘unknown’. A summary of the review is provided below and in Table 1. More detail is contained in the individual species descriptions linked below. The individual species descriptions aim to summarise recent trends and current knowledge on the range and numbers of elasmobranch species recorded in Scottish waters, and to guide readers to additional available data.
Main findings
- This inventory has recorded a total of 64 elasmobranchs (28 skates and rays, and 36 sharks) reported in Scottish waters
- Of the species reported, 20 are found in shelf/coastal waters, 40 are deep-sea species, and four are oceanic
- Twenty-three species were recognised as having low amounts of data with less than 10 records in Scottish waters; of these, nineteen species were identified as ‘rare visitors’ to Scottish waters
Acknowledgements
This work received funding from NatureScot and MASTS and is a product of SIORC.
Suggested citation: Thorburn, J., Cole, G., Bortoluzzi, J., Bromley, C., Burns, F., Burrett, I., Clarke, L., Hardman, S., Henderson, S., Neat, F., Orr, K., Pinn, E., Williams, A., Shedden, L., Donnan, D., Dodd, J., Henry, L-A., (2025). Sharks, skates, and rays of Scotland. An overview of the elasmobranch fauna of Scottish seas.
Keywords
Elasmobranch; Shark; Skate; Ray; Conservation; Marine; Mobile species
Abbreviations
- CFP – Common Fisheries Policy
- DATRAS – Database of trawl surveys hosted by ICES
- ICES – International Council for the Exploration of the Sea
- ISRA – International Union for the Conservation of Nature Important Shark and Ray Areas
- IUCN – International Union for the Conservation of Nature
- JNCC – Joint Nature Conservation Committee
- MASTS – Marine Alliance for Science and Technology in Scotland
- MCZ – Marine Conservation Zone
- MPA – Marine Protected Area
- PMF – Priority Marine Feature
- RSA – Recreational Sea Angling
- SAC – Special Area of Conservation
- SIAMISS – Scottish-Irish anglerfish and megrim industry-science survey
- SIORC – Sharks, skates and rays in the offshore region and coastal zone of Scotland
- SSTP – Scottish Shark Tagging Programme
- TAC – Total Allowable Catch
- UKSTP – UK Shark Tagging Programme
Introduction
Scotland’s Marine Environment
High levels of biodiversity and geodiversity characterise the Scottish marine environment. Geological features that support biologically rich, abundant, or unique ecosystems include Scotland’s fjordic loch systems, its island chains, including the Inner and Outer Hebridean archipelagos, as well as estuaries and deep-sea features such as the continental slope, three seamounts, channels, ridges, banks, cold seeps, abyssal plains, and troughs. These geological systems interact with regional, mesoscale, and local-scale oceanographic and hydrographic features that often promote both nutrient upwelling and downwelling, thereby increasing surface productivity and exporting carbon production to deeper waters.
The waters around Scotland represent an intersection of the warm, saline, northward-flowing surface North Atlantic Current modified by the fresher, cooler Scottish coastal currents. In deeper waters around Scotland, water masses are also characterised by layers of cooler, highly oxygenated Labrador Sea Water and Wyville Thomson Ridge Overflow Water, e.g., across the Rockall Trough and its three seamounts, Rosemary Bank, Anton Dohrn, and the Hebrides Terrace Seamount. In very deep sections, e.g., in the Faroe-Shetland Channel, bottom temperatures can reach sub-zero, where overflow waters mix with North Atlantic Deep Water.
The intersection of cool and warmer waters, combined with the high diversity of ecosystem types and productivity regimes, helps explain the biogeographic mixture of elasmobranch fauna observed in Scotland’s coastal zone and deep sea. Seasonal visitors, such as basking sharks and the occasional southern migrant, such as blue shark and common thresher shark, are seen in the summer. In contrast, boreal and Arctic species, such as Arctic skate, have also been observed. The biology, ecology and behaviour of elasmobranchs is closely coupled to oceanography and hydrography, and these are expected to alter in the coming decades as a result of climate change. Therefore, it is likely that the biodiversity of Scotland’s elasmobranchs and thus their roles in the ecosystems will also change. Decision-makers and managers must have the best information possible available to allow conservation policies to be adapted as new information comes to light.
Safeguarding Scotland’s seas amid rapid climate change requires transdisciplinary efforts to guide sustainable development that balances socioeconomic needs with conservation and protection (e.g., Jouffray et al., 2020). Over the next decade and century, the cumulative impact of multiple stressors arising from human activities and climate change on Scotland’s biodiversity requires urgent attention.
Elasmobranchs
Elasmobranch extinction risk is substantially higher than for other fish species due to strong K-strategist life history traits, and they have already experienced significant, global declines (Dulvy et al. 2021). Longevity, late age of maturity, low fecundity and large, well-developed offspring (Stevens et al., 2000; Ellis et al., 2005; Jabado et al., 2024) make elasmobranchs particularly vulnerable to population reductions (Jabado et al., 2024). Elasmobranchs play an essential role in marine ecosystems as mesoconsumers (consumers at intermediate trophic levels) and top predators (Jabado et al., 2024), and their loss may substantially alter ecosystem structure and function (Myers & Worm, 2003; Heithaus et al., 2008).
Several species of elasmobranchs in the Northeast Atlantic, including those found in Scotland, have suffered severe population declines (Ellis et al., 2005; De Oliveira et al., 2013), with at least one species being declared locally extinct (Brander, 1981).
Scotland has a high proportion of endangered species, with over half of the species listed by the IUCN as near threatened (Figure 1). Despite this conservation concern, the population trends of many species are still very poorly understood, with over a quarter lacking any population assessment, recorded here as “unknown” (Figure 2). We must understand the stresses on elasmobranch populations, as these can have significant indirect effects on elasmobranch communities by exerting selection pressures (Jennings & Kaiser, 1998). Pressures on elasmobranch populations (e.g. over-fishing) can lead to changes in the age structure and size of individuals within a population (Stevens et al., 2000; Hutchings, 2005), as well as changes to the community structure of elasmobranchs within a given area (Walker & Hislop, 1998), which may impact the broader ecosystem (Heithaus et al., 2008). Consequently, the appropriate management of remaining populations is vital to ensure Scotland’s diverse elasmobranch communities and their ecosystems have a chance to re-stabilise and replenish, helping to support the societies and communities that depend on the ecosystem services that flow from them.
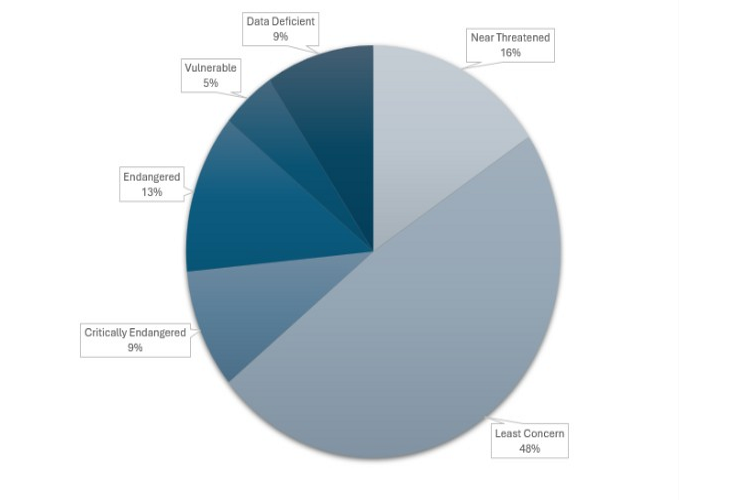
Figure 1. Number of elasmobranch species recorded in Scotland in each IUCN Red List category, including 'rarities'.
Click for a full description
Pie chart showing the number of elasmobranch species recorded in Scotland in each of the IUCN Red List categories:
- Least Concern - 31 (48%)
- Near Threatened - 10 (16%)
- Data Deficient - 6 (9%)
- Vulnerable - 3 (5%)
- Endangered - 8 (13%)
- Critically Endangered 6 (9%), including 'rarities'
IUCN categories use European assessment where possible; for species with no European assessment, the global assessment is used.
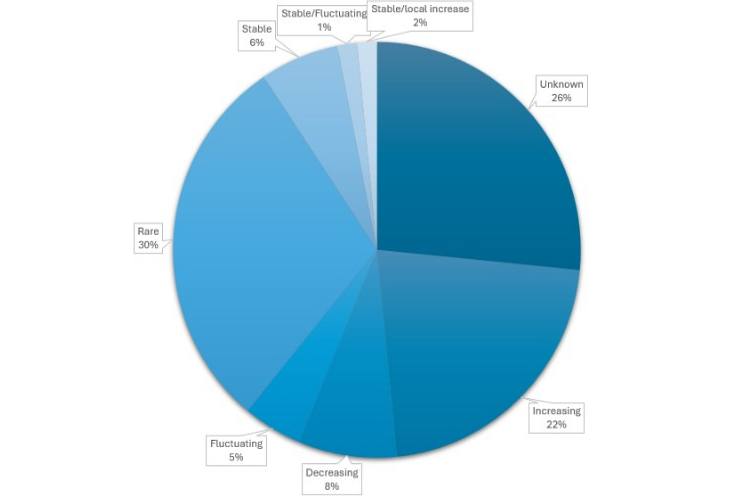
Figure 2. Population trends of elasmobranchs recorded in Scottish waters.
Click for a full description
Pie chart showing population trends of elasmobranchs recorded in Scottish waters:
- increasing 22%
- decreasing 8%
- fluctuating 5%
- rare 30%
- stable 6%
- stable/fluctuating 1%
- stable/local increase 2%
- unknown 26%
International and National Policy context
Scotland and the wider UK are parties to various multilateral agreements that directly support the conservation management of elasmobranchs. These include the United Nations Fish Stocks Agreement (UNFSA), the Food and Agriculture Organization (FAO), the Code of Conduct for Responsible Fisheries (CRF) and FAO’s voluntary International Plan of Action for Conservation and Management of Sharks. Notably, the Port State Measures Agreement (PSMA) is the first binding international instrument that specifically targets illegal, unreported and unregulated (IUU) fishing. Directly relevant to Scotland’s vast deep-sea biome, which covers 80% of Scotland’s Exclusive Economic Zone out to its 12 nm limit, are the UN General Assembly Resolutions 61/105 and 64/72 to manage deep-sea fisheries with minimal impact on the environment or prohibit such fishing in areas meeting the criteria for Vulnerable Marine Ecosystems (VMEs; Weaver et al., 2011).
International conventions also contributing to elasmobranch conservation include:
- OSPAR Convention: Protects the marine environment of the North-East Atlantic. Sharks and skates are included in the OSPAR List of Threatened and/or Declining Species. These are Angel shark, Basking shark, Common skate, Gulper shark, Leafscale gulper shark, Porbeagle, Portuguese dogfish, Spotted skate, Spurdog, Thornback skate, White skate.
- Convention on Biological Diversity (CBD): Mandates biodiversity conservation, including elasmobranchs.
- Global Biodiversity Framework (GBF): set global targets to increase the coverage of protected areas to 30% of land, water, and sea; promote sustainable fisheries management, integrating conservation with fishing practices to halt and reverse biodiversity loss and halt human-induced extinction of known threatened species by 2030.
- Convention on Migratory Species (CMS): The UK signed the Memorandum of Understanding on the Conservation of Migratory Sharks in 2012. The MoU aims to achieve and maintain a favourable conservation status for migratory sharks based on the best available scientific information, taking into account the socio-economic value of these species for people. Several elasmobranch species are listed for protection, including basking shark, porbeagle, tope and spurdog.
- The EDGE list comprises the most evolutionarily distinct and threatened species on Earth (EDGE Species), the loss of which would represent the loss of long branches of the Tree of Life. The list includes basking shark (ranked 7 in the world), tope and porbeagle.
- Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) is a multilateral treaty that aims to ensure that international trade does not threaten the survival of species in the wild by providing varying degrees of protection through restrictions on trade of species placed on one of three Appendices. Appendix 1 makes the commercial trade of species in this Appendix illegal, except in exceptional, licensed circumstances. Appendix II imposes strict regulations on trade in species and requires an export permit, but not necessarily an import permit. Appendix III species are only permitted with an appropriate export permit and a certificate of origin. Scottish species of elasmobranch currently on CITES Appendix II are Basking sharks (Cetorhinus maximus), Porbeagle (Lamna nasus) and thresher sharks (Alopias species).
UK elasmobranch fisheries management
Elasmobranch fisheries, including deepwater species, are managed in a combined national Total Allowable Catch (TAC) for ‘skates and rays’. Catches of cuckoo skate (Leucoraja naevus), thornback skate (Raja clavata), blonde skate (Raja brachyura), spotted skate (Raja montagui), smalleyed skate (Raja microocellata), sandy skate (Leucoraja circularis), shagreen skate (Leucoraja fullonica) are required to be reported separately. A total allowable catch (TAC) is agreed for the North-east Atlantic for spurdog (Squalus acanthias) at the UK - EU fisheries negotiations but only as result of fisheries bycatch, targeted fisheries is not allowed.
Shark finning (the removal of shark fins at sea and the discard of the carcass overboard) has been illegal in UK waters since 2003. This was further reinforced with a Fins Naturally Attached (FNA) policy in 2009 which required all retained sharks to be landed with their fins still attached to the carcass. The Shark Fins Act 2023 built on these, banning the import and export of detached shark fins and products containing them, including items such as shark fin soup. The act reflected growing public and political support for shark conservation and aligned the UK with international efforts to protect declining shark populations.
The Tope (Prohibition of Fishing) Order 2008 was introduced to conserve tope (Galeorhinus galeus) stocks and prevent the development of a “directed” tope fishery in English waters.
In 2012, Scotland introduced the Sharks, Skates and Rays (Prohibition of Fishing, Trans-shipment and Landing) (Scotland) Order 2012. This order prohibits the landing of listed species caught in Scottish waters, giving several species a higher level of protection than they were subject to in other European waters. The Order also specifically set out to protect tope, banning their landing by either commercial or recreational angling vessels, and the fishing of tope with any gear other than by rod and line or hand-line. Scotland also has a ‘Return Unharmed Policy’ that refers to the legal requirement to return certain species, including the elasmobranchs flapper skate, blue skate, basking shark, porbeagle, angel shark and undulate skate, and all species below the minimum conservation reference size, back to the sea. This policy is mandated for both commercial and recreational fishers to prevent the overexploitation of vulnerable populations and to protect endangered species.
Species listed as Prohibited under The Sharks, Skates and Rays (Prohibition of Fishing, Trans-shipment and Landing) (Scotland) Order 2012
- Angel shark (Squatina squatina)
- Common skate (Dipturus batis)
- White skate (Rostroraja alba)
- Spurdog (Squalus acanthias)
- Knifetooth dogfish (Scymnodon ringens)
- Sailfin roughshark (Sharpback shark) (Oxynotus paradoxus)
- Greater lanternshark (Etmopterus princeps)
- Undulate ray (Raja undulata)
- Leafscale gulper shark (Centrophorus squamosus)
- Portuguese dogfish (Centroscymnus coelolepis)
- Blackmouth catshark (Blackmouth dogfish) (Galeus melastomus)
- Longnose velvet dogfish (Centroscymnus crepidater)
- Black dogfish (Centroscyllium fabricii)
- Greenland shark (Somniosus microcephalus)
- Six-gilled shark (Hexanchus griseus)
- Velvet belly (Etmopterus spinax)
- Deep-water catsharks (Apristurus spp.)
- Frilled shark (Chlamydoselachus anguineus)
- Birdbeak dogfish (Deania calcea)
- Kitefin shark (Dalatias licha)
- Tope (Galeorhinus galeus)
Additional UK conservation measures for Elasmobranchs
Bespoke protection is afforded to a few species through the Wildlife and Countryside Act 1981. The Listings of elasmobranch under this act differ between England, Wales and Scotland. Under Schedule V of the Wildlife and Countryside Act 1981, it is an offence to: (1) intentionally or recklessly kill, injure or take fish; (2) possess or sell fish; (3) intentionally or recklessly disturb or harass basking shark (Cetorhinus maximus) in Scottish waters (out to 12nm).
The International Plan of Action for the Conservation and Management of Sharks, a voluntary, non-binding international guideline was adopted by the FAO in 1999. The aim of the IPOA-Sharks is to promote the long-term sustainability of shark, skate, ray, and chimaera populations. It provides a framework for countries to develop their own National Plans of Action (NPOAs) by improving data collection, implementing responsible fishing practices, and managing shark fisheries to prevent overexploitation and ensure sustainable use of these species. JNCC report 360 Plan of Action for the Conservation and Management of Sharks in UK Waters was written in 2004.
Spatial Management
The Marine (Scotland) Act 2010 provides powers for Scottish Ministers to designate Marine Protected Areas (MPAs). The primary purpose of an MPA is to provide a refuge for a specific species or habitat, preventing or alleviating stock collapse, while also facilitating population growth or maintenance. The Scottish MPA network comprises sites for nature conservation, biodiversity protection, demonstrating sustainable management, and preserving our heritage. In total, the network now covers approximately 37% of our seas and oceans (Figure 3), comprising:
- 65 Sites of Special Scientific Interest
- 58 Special Areas of Conservation
- 58 Special Protection Areas
- 36 Nature Conservation MPAs
- 16 RAMSAR sites
- 8 Historic MPAs
- 5 Other Area Based Measures
- 1 Demonstration & Research MPA
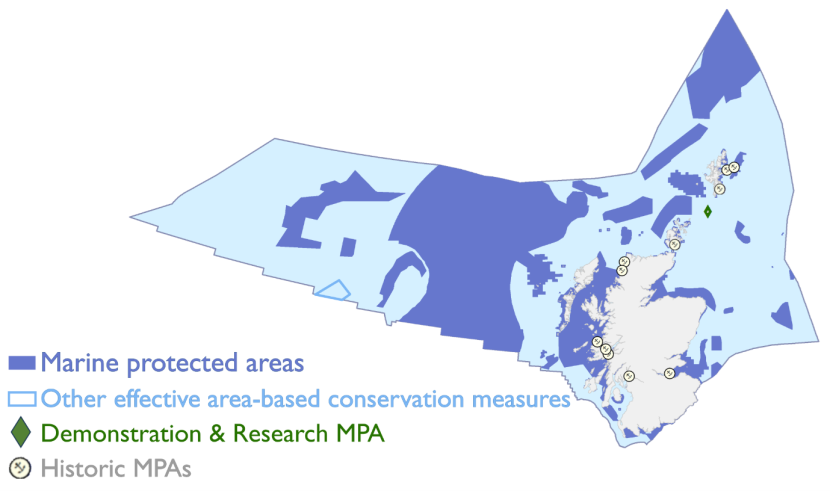
Figure 2. Scotland’s Marine Protected Area (MPA) network as of 2025
Click for a full description
A map showing Scotland’s economic exclusion zone (EEZ, the maritime area defined by the United Nations Convention on the Law of the Sea extending up to 200 nautical miles from Scotland's coast, where Scotland has special rights to explore and use marine resources like oil, fish, and minerals, light blue) highlighting designated MPAs (dark blue), areas managed under other area-based measures (outlined in mid blue), and demonstration and research MPAs (green). Collectively, these areas form a network aimed at conserving marine biodiversity, safeguarding habitats and species, and promoting sustainable use of marine resources.
Image from Scottish Government Marine Environment website.
Marine Protected Areas (MPAs) are considered a key management tool to aid elasmobranch conservation (Barker and Schluessel, 2005; Heupel and Simpfendorfer, 2005; MacKeracher, 2019). Scotland has three MPAs that directly support elasmobranch conservation; for the flapper skate, there is the Loch Sunart to the Sound of Jura Nature Conservation MPA designated for the protection of adult flapper skate and the Red Rocks and Longay Nature Conservation MPA designated for the protection of a flapper skate egg nursery, while the Basking shark is listed as a designated feature for the Sea of the Hebrides Nature Conservation MPA.
Wider conservation measures beyond the MPA network are achieved through Scotland’s Priority Marine Features (PMF). These represent key marine habitats and species of conservation importance in Scottish waters. Priority Marine Features are protected under Scotland’s National Marine Plan Policy 9b and any development and use of the marine environment must not result in significant impact on the national status of PMFs. Seven elasmobranch species are included in the PMF list (Table 1).
| Priority Marine Feature | Species name | Scottish Marine Area |
|---|---|---|
| Basking shark | Cetorhinus maximus | Both |
| Flapper skate and blue skate (formerly common skate) | Dipturus intermedius and D. batis | Both |
| Leafscale gulper shark | Centrophorus squamosus | Offshore waters |
| Porbeagle shark | Lamna nasus | Offshore waters |
| Portuguese dogfish | Centroscymnus coelolepis | Offshore waters |
| Sandy ray | Leucoraja circularis | Offshore waters |
| Spiny dogfish | Squalus acanthias | Both |
Codes of conduct
Beyond the international and national policies, sectoral management of commercial-scale operations, technical and area-based measures, Scotland also practices a range of codes of conduct, some of which are rooted in legislation.
Marine tourism brings in £3.7 billion to the Scottish economy each year, with the Recreational Sea Angling sector bringing in £140 million alone. The potential for interactions with elasmobranchs, among other species, has necessitated various codes of conduct, both voluntary and legally binding.
Under Section 51 of the Nature Conservation (Scotland) Act 2004, NatureScot developed the Scottish Marine Wildlife Watching Code (SMWWC). This code outlines the types of activities likely to disturb marine wildlife, the circumstances under which marine wildlife may be approached, and recommendations for viewing marine wildlife with minimal disturbance. The WiSE (Wildlife Safe) Scheme offers a standard for commercial skippers to ensure safe wildlife watching that will also reduce the risk of disturbance to wildlife, including basking sharks. The Green Blue and RYA publicised voluntary measures to reduce boat speeds and to keep a lookout in basking shark hotspots to reduce collision risks in areas where it is more likely to find groups of sharks at the surface.
There are also numerous codes of conduct to advise best practice on Angling equipment and handling protocols to maximise the survival rate in catch and release fisheries, e.g., angling groups such as the Scottish Sea Angling Conservation Network (SSACN) and NatureScot through the SkateSpotter website.
Addressing knowledge gaps to aid marine planning
The 2018 review of Scotland’s National Marine Plan (NMP) acknowledged that extensive monitoring of marine taxa and ecosystems supported the principle of “Living within environmental limits”. However, elasmobranchs remain underrepresented in Scotland’s marine evidence base. The Marine Scotland Maps NMPi portal, for example, lists data for only 19 elasmobranch species.
Scotland’s NMP envisions “clean, healthy, safe, productive and diverse seas, managed to meet the long-term needs of nature and people.” Yet this vision cannot be fully realised without robust baseline knowledge of elasmobranch biodiversity, distribution, biology and ecology. The absence of such data constrains effective governance, particularly the integration of elasmobranch conservation into Marine Protected Areas (MPAs), technical fisheries measures, and broader sectoral planning.
Scotland’s long history of commercial fishing has likely had significant direct and indirect impacts on elasmobranch populations. Beyond reducing absolute population size, fishing can drive ecological change through selective pressures (Jennings & Kaiser, 1998), altering age and size structures (Stevens et al., 2000; Hutchings, 2005), and disrupting community composition (Walker & Hislop, 1998). These changes can degrade functional ecosystem roles of elasmobranchs, including through loss of key habitats (Heithaus et al., 2008). These ecological pressures are increasingly overlaid with climate-driven shifts in distribution and resilience (Sguotti et al., 2016).
Scotland’s ambition to expand its marine sectors, such as aquaculture, offshore renewable energy, and tourism, presents further risks to elasmobranchs. Despite increased efforts in spatial planning, elasmobranchs are often poorly addressed in Strategic Environmental Assessments (SEAs) and Environmental Impact Assessments (EIAs). Notably, the 'Sustainability Appraisal for Scotland’s pilot regional Marine Spatial Plan (MSP)' in the Pentland Firth and Orkney Waters (PFOW) identified the lack of baseline data as a critical barrier to evaluating impacts, particularly in an area slated for rapid renewable energy development and known to be used by critically endangered species such as the porbeagle shark.
Without comprehensive baseline data, most elasmobranch species in Scottish waters remain effectively invisible in national planning processes. This knowledge gap undermines efforts to assess and mitigate the environmental impacts of policies, plans, programmes and projects under the NMP and associated strategies.
Purpose of this report
This work brings together multiple stakeholders and diverse knowledge systems to update our understanding of elasmobranch biology and ecology in Scottish waters.
This includes:
- An updated species inventory of the species for which there are records of presence in Scottish waters (up to 2020)
- Bathymetric distribution of species as of 2025
- Regional conservation status of each species in 2025
- Population status in Scottish waters from pre-existing analysis (up to 2020)
- Life history traits, trophic ecology, and remaining knowledge gaps in 2025
This report aims to summarise available data for each elasmobranch species in Scottish waters, guiding researchers and policymakers toward key data sources to support sustainable management against the backdrop of climate change and increasing sectoral pressures. It also reflects Scotland’s commitment to leading a cross-stakeholder approach to address data gaps and, over time, build a robust, evidence-based conservation framework that showcases national research capacity. While not exhaustive, the report highlights available datasets through references to relevant publications, presence-only surveys, fisheries data, and tagging information.
Data sources
Elasmobranch management has, until recently, been based on limited knowledge, as species-specific data have historically been scarce (Stevens et al., 2000). In order to generate a baseline of knowledge for Scottish elasmobranchs, this report synthesises a variety of data sources that could be exploited to provide the critical information needed by policymakers to improve the conservation management of elasmobranchs in Scotland, including:
- ICES DATRAS datasets: BTS, DWS, DYFS, IE-IGFS, NS-IBTS, ROCKALL, SCOWCGFS, SWC-INTS
- Marine Directorate Monkfish Surveys (SIAMISS)
- Marine Directorate Deep water surveys
- Marine Directorate landings data 2011-2019
- Marine Directorate discard data 2014-2018
- Scientific literature and commissioned reports
- The Scottish Sea Angling Conservation Network, which assimilated data from: The Scottish Shark Tagging Programme, Glasgow Museums Tagging Programme and the UK Shark Tagging Programme
- Basking shark sightings
This report does not present an exhaustive account of all elasmobranch records ever reported in Scottish waters. However, it provides a comprehensive overview of the majority of available data from scientific surveys, fisheries, and recreational angling activities up to 2020.
Elasmobranch ecology and biology
Traditionally, many species of elasmobranchs are viewed as highly migratory (Hyrenbach et al., 2000; Mucientes et al., 2009), with some species capable of performing trans-ocean movements (e.g. Templeman, 1976; Gore et al., 2008). Long life spans and a propensity to migrate mean an individual elasmobranch can make use of many different geographic locations and habitats during its lifetime (Bres, 1993; Thorburn et al., 2019, 2024). Habitat use varies between the sexes and life history stage, leading to population segregation (McMillan & Morse 1999, Sims et al., 2001, Daan et al., 2005; Fordham et al., 2006; Wearmouth & Sims, 2008), with different population groups showing different movements (Mucientes et al., 2009) and seasonal variation (Wearmouth and Sims, 2010; Thorburn et al., 2019, 2024). Despite the wide-ranging behaviour of some elasmobranchs, it has been shown that many species make use of the same areas repeatedly, termed ‘site fidelity’ (Goldman & Anderson 1999; Speed et al., 2010). Site fidelity varies considerably between species and habitats, with some species displaying residency in specific areas (Papastamatiou et al., 2009; Andrews et al., 2009; Thorburn et al., 2015, 2024; Lavender et al., 2021).
A fundamental challenge to the conservation of marine organisms is defining an appropriate spatial scale to manage a species, which has led to the grouping of exploited marine organisms into stocks. As a general rule, stocks are split into regional populations that have no genetic or demographic overlap (Waples 1998). There are numerous tools available to gain insight into the movement patterns of elasmobranchs. Straight line movement can be inferred from the detection of an individual’s presence in an area through methods such as mark recapture (MR) (e.g. Holden & Horrod, 1979; Hurst et al., 1999) or acoustic telemetry (e.g. Heupel & Simpfendorfer, 2005; Papastamatiou et al., 2013). Higher resolution movements may be re-created from environmental data recorded by electronic tags (e.g. Sims et al., 2003; Gore et al., 2008; Lavender et al., 2021; Thorburn et al., 2024) or from precise information on position, such as data gained through active acoustic tracking (e.g. Nakano et al., 2003) or satellite GPS locations (e.g. Teo et al., 2004; Papastamatiou et al., 2013). Movement may also be surmised from molecular markers, which provide information on population-wide movements and allow inferences regarding the maximum extent of a population as well as reproductive mixing (Chapman et al., 2015).
Embryo nutrition strategies
All elasmobranchs exhibit internal fertilisation (Carrier et al., 2004), with females retaining the fertilised embryos internally during the early stages of development, when the embryos are most vulnerable. Elasmobranchs can be broadly categorised into two reproductive modes based on the duration of embryonic retention by the female: (1) viviparous (live-bearing) and (2) oviparous (egg-laying) species (Carrier et al., 2004). Across different elasmobranch species, all major forms of vertebrate embryonic nutrition are represented, including two oviparous strategies and at least five distinct forms of viviparity.
Oviparous (e.g. flapper skate)
Extended: Almost all oviparous species have extended oviparity, in which ova are fertilised, enclosed in an egg case, deposited and hatch after a period of embryo development. This development period is highly variable and largely dependent on temperature. In deep-water skate species, development times of up to 1290 days (3.5 years) have been reported (Hoff, 2008).
Retained: Egg cases are retained in the oviduct to allow further development. This limits the amount of time they are left in the open before hatching. Multiple eggs can be retained in the oviduct at once (Compagno, 1990).
Viviparous
Placental viviparity (e.g. blue shark):
Placental nutrition. The yolk sac, once exhausted, becomes highly vascularised and attaches to the uterine surface and forms a placenta, carrying nutrients to the embryo via the mother’s blood. It is worth noting that some elasmobranchs combine several of the following strategies to provide nutrition to their young (Worums 1977).
Aplacental viviparity (e.g. spurdog and porbeagle):
Yolk dependency: (e.g. spurdog): the embryos receive nutrients solely from a yolk sac, a finite resource, resulting in young being born relatively small.
Uterine Milk: (also known as Placental Analogues); an area of the uterine epithelium produces nutritional ‘milk’ that is ingested by the young. This form of nutrition is common in rays.
Oophagy: (e.g. porbeagle): The female elasmobranch continually produces eggs, which are ingested by embryos in the uterus.
Adelphophagy (intra-uterine cannibalism): In some cases of oophagy, there will be several developing embryos in the uterus to begin with. When energy demands outgrow the nutritional value of the supplied eggs, the dominant embryo will then kill the other developing embryos and feed on them (Wourms 1977).
In all cases, large, well-developed young are produced, maximising survival potential. This does, however, mean fewer young are produced per reproductive cycle and, due to the length and energy demands of the gestation period, many female elasmobranchs can only reproduce once every two years. Large, well-developed young are also almost immediately vulnerable to fisheries mortality (Brander 1981; Smith et al., 2008).
Species list and Appendices
Sharks
Requiem sharks - Carcharhinidae
- Blue shark - Prionace glauca
Ground sharks - Carcharhiniformes
Catsharks - Pentanchidae and Scyliorhinindae
Demon catsharks
- White ghost catshark - Apristurus aphyodes
- Ghost catshark - Apristurus manis
- Iceland catshark - Apristurus laurussonii
- Black roughscale catshark - Apristurus melanoasper
- Smalleye catshark - Apristurus microps
Sawtail catsharks
- Blackmouth catshark - Galeus melastomus
- Mouse catshark - Galeus murinus
Scyliorhinus species
- Small spotted catshark - Scyliorhinus canicula
- Nursehound - Scyliorhinus stellaris
Houndsharks – Triakidae
- Tope – Galeorhinus galeus
- Starry smoothhound – Mustelus asterias
Cow and frilled sharks - Hexanchiformes
- Bluntnose sixgill shark - Hexanchus griseus
Basking sharks - Cetorhinidae
- Basking shark - Cetorhinus maximus
Mackerel sharks - Lamniformes
- Porbeagle - Lamna nasus
Gulper sharks - Centrophoridae
- Leafscale gulper shark - Centrophorus squamosus
- Birdbeak dogfish - Deania calceus
Kitefin sharks - Dalatiidae
- Kitefin shark - Dalatias licha
Lanternsharks – Etmopteridae
- Great lanternshark- Etmopterus princeps
- Velvet belly lanternshark - Etmopterus spinax
- Black dogfish - Centroscyllium fabricii
Sleeper sharks – Somniosidae
- Portuguese dogfish - Centroscymnus coelolepis
- Longnose velvet dogfish - Centroselachus crepidater
- Knifetooth dogfish - Scymnodon ringens
Dogfish sharks – Squalidae
- Spurdog - Squalus acanthias
Skates
Softnose skates – Arhynchobatidae
Bathyraja species
- Richardson’s skate – Bathyraja richardsoni
Hardnose skates – Rajidae
Amblyraja species
- Arctic skate - Amblyraja hyperborea
- Jensen’s skate - Amblyraja jenseni
- Starry skate - Amblyraja radiata
Dipturus species
- Flapper skate - Dipturus intermedius
- Common blue skate - Dipturus batis
- Norwegian skate - Dipturus nidarosiensis
- Longnosed skate - Dipturus oxyrinchus
Leucoraja species
- Sandy skate - Leucoraja circularis
- Shagreen skate - Leucoraja fullonica
- Cuckoo skate - Leucoraja naevus
Neoraja species
- Blue pygmy skate - Neoraja caerulea
Raja species
- Blonde skate - Raja brachyura
- Thornback skate - Raja clavata
- Spotted skate - Raja montagui
Rajella species
- Deep-water skate - Rajella bathyphila
- Bigelow's skate - Rajella bigelowi
- Round skate - Rajella fyllae
- Mid-Atlantic skate Rajella kukujevi
- White skate - Rostroraja alba
Illustrations
Dietrich Burkel, a pioneer of shark and skate tagging
The illustrations in the Sharks and Skates of Scotland: An overview of the elasmobranch fauna of Scottish seas report were painted by Dr Dietrich Burkel, a geologist by training who worked as Assistant Keeper of Natural History and later Depute Keeper at Kelvingrove Museum.

Dietrich Burkel with a fibreglass model of the Porbeagle he caught in 1970, the first one landed on rod and line in Scotland.
Click for a full description
The model is still on display at the Kelvingrove Museum today along with others made by Dietrich during his time as Assistant Keeper of Natural History .
Dietrich was a keen angler and talented artist. He made fibreglass casts of many of the fish he caught which he then painstakingly painted them, resulting in models of several Scottish record specimens being added to the collection at Kelvingrove. He is credited with catching the first Porbeagle on road and line in Scotland on the 1st August 1970. The shark was caught off the Mull of Galloway and weighed 173lb (7.5kg) and measured over 6’ (2m), a fibreglass model of the shark made by Dietrich still holds pride of place at Kelvingrove Museum today.
Thornback skate
Dietrich painted several versions of the thornback skate because their colour and spot patterns vary so much. This is thought to help the thornback skate camouflage itself over a variety of substrates from light sand to dark mud. While they do not change colour instantly they use pigment changes to match their background.










Dietrich was a keen advocate of catch and release angling, once racing ashore to quickly weigh a Tope he had caught so that he could release it alive rather than killing it to get the species record as was the custom at the time. After attending the Ullapool skate angling competition and being horrified that the skate were being brought ashore and killed to be weighed he began encouraging anglers to tag and return fish and started the Glasgow Museum database of skate captures in 1974. This was thought to be the first tagging of any sharks, skate or rays in the UK.

Dietrich visited Scotland in 2009 after retiring as curator at the Zoological Museum at the University of Hamburg to fish for and tag skate in the Sound of Jura.
The project grew under supervision from Richard Sutcliffe, Dietrich’s successor at Kelvingrove Museum with volunteer anglers such as Davy Holt, Bill Little, Brian Swanbanks and Duncan Swinbacks who developed weight charts so that skate no longer had to be weighed but their weight could be estimated from length and width. The Scottish Sea Angling Conservation Network took custody of the data in 2007. The project continued as the Scottish Shark Tagging Programme until 2021 until the data was included in SkateSpotter, a flapper skate tag recapture online photodatabase and app managed by NatureScot and The Scottish Association for Marine Science. The Glasgow Museum data collected for flapper skate, common blue skate, nursehound, spurdog, cuckoo skate, thornback skate, spotted skate and round skate is included in the Sharks and Skates of Scotland: An overview of the elasmobranch fauna of Scottish seas report alongside fishery survey data and fisheries landings and discards data.
In 2013 the Scottish Sea Angling Conservation Network used the data relating to flapper skate to propose and justify designation of the Loch Sunart to the Sound of Jura MPA for the protection of flapper skate. The site was designated in 2014 and the site was closed to mobile gear fishing in 2016. Anglers continue to voluntarily tag and report recaptures of skate today to help NatureScot monitor the MPA.
Thanks to Ian Burrett, Richard Sutcliffe and Robyn Haggard for help compiling this tribute.
References
Barker, M. J., & Schluessel, V. (2005). Managing global shark fisheries: Suggestions for prioritizing management strategies. Aquatic Conservation: Marine and Freshwater Ecosystems, 15(4), 325–347.
Brander, K. (1981) Disappearance of common skate Raia batis from Irish Sea. Nature 290:48–49.
Bres, M. (1993). The behaviour of sharks. Reviews in Fish Biology and Fisheries 3:133–159.
Carrier, J., Pratt, H., Castro, J. (2004). Reproductive Biology of Elasmobranchs. In: Musick, J., Carrier, J., Heithaus, M. (eds) Biology of Sharks and Their Relatives. CRC Press, p 269–286.
Chapman. D.D., Feldheim, K.A., Papastamatiou, Y.P., Hueter, R.E. (2015). There and Back Again: A Review of Residency and Return Migrations in Sharks, with Implications for Population Structure and Management. Annual Review of Marine Science 7:547–570.
Compagno, L.J.V. Alternative life-history styles of cartilaginous fishes in time and space. Environmental Biology of Fish 28, 33–75 (1990)
De Oliveira, J. A. A., Ellis, J. R., & Dobby, H. (2013). Incorporating density dependence in pup production in a stock assessment of Northeast Atlantic spurdog Squalus acanthias. ICES Journal of Marine Science, 70(7), 1341–1353
Dulvy, N. K., Pacoureau, N., Rigby, C. L., et al., (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Current Biology, 31(20), 4773–4787
Edgar, G. J., Russ, G. R., & Babcock, R. C. (2007). Marine protected areas. Marine Ecology, 28(Suppl. 1), 533–555.
Ellis, J., Cruz-Martinez, A., Rackham, B., & Rogers, S. (2005). The distribution of chondrichthyan fishes around the British Isles and implications for conservation. Journal of Northwest Atlantic Fisheries Science, 35, 195–213
Goldman, K.J., Anderson, S.D. (1999). Space utilization and swimming depth of white sharks, Carcharodon carcharias, at the South Farallon Islands, central California. Environmental Biology of Fishes 56:351–364.
Gore, M.A., Rowat, D., Hall, J., Gell, F.R., Ormond, R.F. (2008). Transatlantic migration and deep mid-ocean diving by basking shark. Biology Letters 4:395–398.
Heithaus, M. R., Frid, A., Wirsing, A. J., & Worm, B. (2008). Predicting ecological consequences of marine top predator declines. Trends in Ecology & Evolution, 23(4), 202–210
Heupel, M. R., & Simpfendorfer, C. A. (2005). Using acoustic monitoring to evaluate MPAs for shark nursery areas: The importance of long-term data. Marine Technology Society Journal, 39(1), 10–18
Hoff, G. R. (2008). A nursery site of the Alaska skate (Bathyraja parmifera) in the eastern Bering Sea. Fishery Bulletin, 106(3), 233–244.
Hutchings, J. A. (2005). Life history consequences of overexploitation to population recovery in Northwest Atlantic cod (Gadus morhua). Canadian Journal of Fisheries and Aquatic Sciences, 62(4), 824–832
Hyrenbach, K.D., Forney, K.A., Dayton, P.K. (2000). Marine protected areas and ocean basin management. Aquatic Conservation: Marine and Freshwwater Ecosystems 10:437–458.
Jabado, R. W., Morata, A. Z. A., Bennett, R. H., Finucci, B., Ellis, J. R., Fowler, S. L., Grant, M. I., Barbosa Martins, A. P., & Sinclair, S. L. (Eds.). (2024). The global status of sharks, rays, and chimaeras. IUCN.
Jennings, S., & Kaiser, M. J. (1998). The effects of fishing on marine ecosystems. Advances in Marine Biology, 34, 201–352
Jouffray, J.-B., Blasiak, R., Norström, A. V., Österblom, H., & Nyström, M. (2020). The Blue Acceleration: The trajectory of human expansion into the ocean. One Earth, 2(1), 43–54
Lavender, E., Aleynik, D., Dodd, J., Illian, J., James, M., Wright, PJ., Smout, S. & Thorburn, J. (2021) Movement patterns of a Critically Endangered elasmobranch (Dipturus intermedius) in a Marine Protected Area. Aquatic Conservation. 32(2).
MacKeracher, T., Diedrich, A., & Simpfendorfer, C. A. (2019). Sharks, rays and marine protected areas: A critical evaluation of current perspectives. Fish and Fisheries, 20(2), 255–267
McMillan, D.G. & Morse, W.W. (1999). Essential Fish Habitat Source Document: Spiny
Dogfish, Squalus acanthias, Life History and Habitat Characteristics. NOAA Technical Memorandum, NMFS-NE-150.
Mucientes, G.R., Queiroz, N., Sousa, L.L., Tarroso, P., Sims, D.W. (2009). Sexual segregation of pelagic sharks and the potential threat from fisheries. Biological Letters 5:156–159.
Myers, R. A., Baum, J. K., Shepherd, T. D., Powers, S. P., & Peterson, C. H. (2007). Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315(5820), 1846–1850
Myers, R.A., Worm, B. (2003). Rapid worldwide depletion of predatory fish communities. Nature 423:280–283.
Nakano, H., Matsunaga, H., Okamoto, H., Okazaki, M. (2003). Acoustic tracking of bigeye thresher shark Alopias superciliosus in the eastern Pacific Ocean. Marine Ecology Progress Series 265:255–261.
Papastamatiou, Y.P., Lowe, C.G., Caselle, J.E., Friedlander, A.M. (2009). Scale-dependent effects of habitat on movements and path structure of reef sharks at a predator-dominated atoll. Ecology 90:996–1008.
Papastamatiou, Y.P., Meyer, C.G., Carvalho, F., Dale, J.J., Hutchinson, M.R., Holland, K.N. (2013). Telemetry and random-walk models reveal complex patterns of partial migration in a large marine predator. Ecology 94:2595–2606.
Sguotti, C., Lynam, C. P., García‐Carreras, B., Ellis, J. R., & Engelhard, G. H. (2016). Distribution of skates and sharks in the North Sea: 112 years of change. Global Change Biology, 22(8), 2729–2743.
Sims, D., Nash, J., Morritt, D. (2001). Movements and activity of male and female dogfish in a tidal sea lough: alternative behavioural strategies and apparent sexual segregation. Marine Biology 139:1165–1175.
Sims, D.W., Southall, E.J., Richardson, A.J., Reid, P.C., Metcalfe, J.D. (2003). Seasonal movements and behaviour of basking sharks from archival tagging: no evidence of winter hibernation. Marine Ecology Progress Series 248:187–196.
Smith, S. E., Au, D. W., & Show, C. (2008). Intrinsic rates of increase in pelagic elasmobranchs. Sharks of the open ocean: biology, fisheries and conservation, 288-297.
Speed, C.W., Field, I.C., Meekan, M.G., & Bradshaw, C.J (2010). Complexities of coastal shark movements and their implications for management. Marine Ecology Progress Series, 408:275–293.
Stevens, J. D., Bonfil, R., Dulvy, N. K., & Walker, P. A. (2000). The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES Journal of Marine Science, 57(3), 476–494.
Thorburn, J., Neat, F., Bailey, DM., Noble, LR. & Jones, CS. (2015) Winter Residency and Site Fidelity in the Critically Endangered NE Atlantic Spurdog (Squalus acanthias). Marine Ecological Progress Series, 526.
Thorburn, J., Neat, F., Burrett, I., Henry, L. A., Bailey, D. M., Jones, CS. & Noble, L. R. (2019) Ontogenetic and seasonal variation in movements and depth use, and evidence of partial migration in a benthopelagic elasmobranch. Frontiers in Ecology special edition: Flexibility in the Migration Strategies of Animals, 7, 353
Thorburn, J., Wright, PJ., Lavender, E., Dodd, J., Neat, F., Martin, JGA., Lynam, C., James, M. (2021) Seasonal and ontogenetic variation in depth use by a Critically Endangered benthic elasmobranch and its implications for spatial management. Frontiers in Marine Science, 8, 656368.
Thorburn, J., Gagnon, É., Martin, J., Henry, L.-A., Wright, P.J., Dodd, J., James, M. & Neat F. (2024) Fine-scale residency and temperature-driven habitat selection in a migratory shark species Environmental Biology of Fishes, 108(4), 483-500.
Templeman, W. (1976). Transatlantic Migrations of Spiny Dogfish (Squalus acanthias). Journal of the Fisheries Research Board of Canada, 33:2605–2609.
Teo, S.L.H., Boustany, A., Blackwell, S., Walli, A., Weng, K.C. & Block, B.A. (2004). Validation of geolocation estimates based on light level and sea surface temperature from electronic tags. Marine Ecology Progress Series, 283:81–98.
Thorburn, J., Gagnon, É., Martin, J., Henry, L.-A., Wright, P.J., Dodd, J., James, M. & Neat F. (2024) Fine-scale residency and temperature-driven habitat selection in a migratory shark species. Environmental Biology of Fishes, Environmental Biology of Fishes, 108(4), 483–500.
Walker, P., & Hislop, J. (1998). Sensitive skates or resilient rays? Spatial and temporal shifts in ray species composition in the central and north-western North Sea between 1930 and the present day. ICES Journal of Marine Science, 55(3), 392–402.
Waples, R.S. (1998). Separating the wheat from the chaff: patterns of genetic differentiation in high gene flow species. Journal of Hereditry, 89:438–450.
Wearmouth, V.J., Sims, D.W. (2008). Sexual segregation in marine fish, reptiles, birds and mammals: behaviour patterns, mechanisms and conservation implications. Advances in Marine Biology, 54:107–170.
Wearmouth, V.J., Sims, D.W. (2010). Sexual Segregation in Elasmobranchs. Biologia Marina Mediterranea, 17:236–239.
Weaver, P. P. E., Benn, A., Arana, P. M., Ardron, J. A., Bailey, D. M., Baker, K., Billett, D. S. M., Clark, M. R., Davies, A. J., Durán Muñoz, P., Fuller, S. D., Gianni, M., Grehan, A. J., Guinotte, J., Kenny, A., Koslow, J. A., Morato, T., Penney, A. J., Perez, J. A. A.,
Priede, I. G., Rogers, A. D., Santos, R. S., & Watling, L. (2011). The impact of deep-sea fisheries and implementation of the UNGA Resolutions 61/105 and 64/72: Report of an international scientific workshop. National Oceanography Centre, Southampton.
Wourms, J.P. (1977). Reproduction and Development in Chondrichthyan Fishes, American Zoologist, Volume 17, Issue 2, pages 379–410.
Annex 1: Note on Chimaeras
Class: Chondrichthyes Order: Chimaeriformes Family: Chimaeridae
Synonyms: Ratfish, ghostfish, ghost sharks
Chimaeras are closely related to elasmobranchs, both elasmobranchs and Chimaeriformes are subclasses in the class Chondrichthyes. Chimaeras, like elasmobranchs, have a cartilaginous skeleton and share many life history traits. They are associated with deep water habitats and are rarely found in shelf waters. As a subclass, they typically extend from 200 to 2500 m depth.
Conservation Listings
Eight species of Chimaera have been recorded in Scottish waters, their status and population trend (as assessed by the IUCN) is listed in Table 1.
Species | Common name | IUCN status | Population trend |
|---|---|---|---|
Chimaera monstrosa (Linnaeus, 1758) | Rabbitfish, rattail | Global – Vulnerable | Decreasing |
Chimaera opalescens (Luchetti, Iglésias & Sellos, 2011) | Opal chimaera | Global – Least Concern | Stable |
Harriotta haeckeli (Karrer, 1972) | Smallspine spookfish | Global – Least Concern | Stable |
Harriotta raleighana (Goode & Bean, 1895) | Narrownose chimaera | Global – Least Concern | Stable |
Hydrolagus affinis (De Brito Capello, 1868) | Small-eyed eyed rabbitfish | Global – Least Concern | Stable |
Hydrolagus mirabilis (Collett, 1904) | Big-eyed rabbitfish | Global – Least Concern | Stable |
Hydrolagus pallidus (Hardy & Stehmann, 1990) | Pale ghost shark | Global – Least Concern | Stable |
Rhinochimaera atlantica (Holt & Byrne, 1909) | Straightnose rabbitfish | Global – Least Concern | Stable |
CITES: Not listed
CMS: Not listed
OSPAR: Not listed
Biology and Ecology
Chimaeras, like elasmobranchs, are “K-selected” species. This means they are long-lived, have low fecundity, late age at maturity and slow growth. This makes them exceptionally vulnerable to overfishing, and stocks that are overfished may take a long time to recover (Koslow et al., 2000).
Chimaeras reach from around 40 to 150cm in length at maturity. The maximum age is estimated at 30 years for Rabbitfish (C. Monstrosa); however, this is likely an underestimate (Calis et al., 2005).
In common with many elasmobranchs, most studied species of chimaeras exhibit sexual dimorphism with males reaching a smaller overall size and size at maturity than females (Moura et al., 2004; Finucci et al., 2017). As well as paired claspers, male chimaeras possess paired pelvic, and a single frontal tenaculum. These are denticle-covered appendages that are used for grasping the female during mating (Didier et al., 2012). Chimaeras differ from elasmobranchs in having upper jaw cartilage that is completely fused to the head. They also have permanent, hypermineralised tooth plates that grow throughout life.
Reproduction: Oviparous (egg-laying). Eggs laid on the seabed may take up to 12 months to gestate (Walker 2020). Sperm storage has been confirmed in Rhinochimaeridae and is assumed to occur in all chimaeras (Finucci et al., 2017). Fecundity for chimaerids is estimated at between 6 and 30 eggs per year (Finucci et al., 2017). Sexual segregation by depth has been identified in 4 species in the Northeast Atlantic (C. monstrosa, H. mirabilis, C. opalescens n. sp. and Harriotta raleighana) with females occurring deeper than males. Fisheries exploitation, therefore, may affect the sexes differently (Holt et al., 2013).
Diet: The diet of chimaeriforms is generally presumed to consist of benthic invertebrates, including crustaceans, molluscs, and echinoderms; however, fish and soft-bodied invertebrates have also been detected in diet studies (Didier et al., 2012). Diets may vary by geographic region; for example, Rabbitfish (C. Monstrosa) sampled in the Mediterranean were found to feed almost exclusively on the decapod Monodaeus couchii (Tamayo et al., 2021).
Protective measures
Conservation measures apply to chimaeras within the NEAFAC Regulatory Area (covering all UK waters). Similar to the regulations pertaining to deep-water sharks, all NEAFAC parties are prohibited from directed fishing for deep-sea chimaeras and must submit all available data on chimaeras to ICES for further stock evaluation (NEAFC Recommendation 09:2025); however, unlike deep-sea sharks, they are not subject to a zero TAC and therefore may be kept as bycatch. Landings into the United Kingdom of any quantity of any mixture of deep-sea species in excess of 100 kgs, live weight, shall be landed only into a port designated under the North-East Atlantic Fisheries Commission (NEAFC) designated ports scheme.
Regulations that exist to protect deep sea species more generally such as the ban on gillnets at depths below 200 m in the North East Atlantic Fisheries Commission (NEAFC) Regulatory Area (Rec 03 2006: Gill Nets 2006), as well as prohibition on fishing with bottom-set gillnets, entangling nets, and trammel nets below 200m (under Regulation (EU) 2019/1241 as amended by S.I 2019/1312 and S.I. 2020/1542) and prohibition of bottom trawls below 800 m (EU 2016/2336 as amended by S.I 2019/753) will provide some protection for these primarily deep dwelling species.
Human interactions
Chimaeras are often caught as bycatch in deep-sea fisheries that are targeting other species. Chimaeras have a dorsal spine that is considered venomous and C. monstrosa injuries requiring medical intervention are reported from fishermen in Scotland (Hayes et al., 2011). This suggests they may be regularly encountered by trawlers in Scottish waters.
Rabbitfish (C. monstrosa) are listed by the IUCN as vulnerable, and landings of the species in the Northeast Atlantic more than doubled between 2006-2014. This may be due to zero total allowable catch limits for other deepwater species being in place for ICES member countries, resulting in the retention of rabbitfish (ICES 2024; IUCN 2020).
References
Calis, E., Jackson, E. H., Nolan, C. P. and Jeal, F. 2005. Preliminary Age and Growth Estimates of the Rabbitfish, Chimaera monstrosa, with Implications for Future Resource Management. Journal of Northwest Atlantic Fisheries Science 35: 21
Didier, D.A., Kemper, J.M., Ebert, D.A., 2012. Phylogeny, biology, and classification of extant holocephalans. In: Carrier, J.C., Musick, J.A., Heithaus, M.R. (Eds.), Biology of Sharks and Their Relatives 2nd edn.. CRC Press, New York, 97–124
Finucci, B. 2020. Chimaera monstrosa. The IUCN Red List of Threatened Species 2020: e.T63114A124459382. Accessed on 17 January 2025.
Finucci, B., et al (2017). Reproductive biology of the two deep-sea chimaerids, longnose spookfish (Harriotta raleighana) and Pacific spookfish (Rhinochimaera pacifica). Deep Sea Research Part I: Oceanographic Research Papers, 120, pp.76-87
Hayes AJ, Sim AJW. Ratfish (Chimaera) spine injuries in fishermen. Scottish Medical Journal. 2011;56(3):161-163. doi:10.1258/smj.2011.011115
Holt, R. E., Foggo, A., Neat, F. C., & Howell, K. L. (2013). Distribution patterns and sexual segregation in chimaeras: Implications for conservation and management. ICES Journal of Marine Science, 70(6), 1198–1205
King, J.R., McPhie, R.P., 2013. Preliminary age, growth and maturity estimates of spotted ratfish (Hydrolagus colliei) in British Columbia. . Deep-Sea Res
ICES (2024). Report of the Working Group on Elasmobranch Fishes (WGEF). ICES Scientific Reports. Report
Moura, T., Figueiredo, I., Machado, P.B. and Gordo, L.S., 2004. Growth pattern and reproductive strategy of the holocephalan Chimaera monstrosa along the Portuguese continental slope. Journal of the Marine Biological Association of the United Kingdom, 84(4), pp.801-804
Koslow J. A. Boehlert G. W. Gordon J. D. M. Haedrich R. L. Lorance P. Parin N., Continental slope and deep-sea fisheries: implications for a fragile ecosystem, ICES Journal of Marine Science, 2000, vol. 57 (pg. 548-557)
Tamayo, M., Barría, C., Coll, M. and Navarro, J., 2021. Highly specialized feeding habits of the rabbitfish Chimaera monstrosa in the deep sea ecosystem of the northwestern Mediterranean Sea. Journal of Applied Ichthyology, 37(6), pp.868-874
Walker, T.I. (2020). Chapter 10 Reproduction of Chondrichthyans. In: Yoshida, M., Asturiano, J. (eds) Reproduction in Aquatic Animals. Springer, Singapore
Annex 2: Protection of deep sea elasmobranchs
Protection of deep sea elasmobranchs
Deep-water elasmobranch species in Scotland have been managed collectively since 2003 through a combination of measures that apply to deep-water fisheries and specifically target deep-water elasmobranch species. In Scotland, domestic protection is provided via The Sharks, Skates and Rays (Prohibition of Fishing, Transshipment and Landing) (Scotland) Order 2012, which prohibits the landing of several elasmobranch species. The deep-sea species included on The Sharks, Skates and Rays (Prohibition of Fishing, Transshipment and Landing) (Scotland) Order 2012 are listed below.
- Scymnodon ringens (Knifetooth dogfish)
- Oxynotus paradoxus (Sailfin roughshark / Sharpback shark)
- Etmopterus princeps (Greater lanternshark)
- Centrophorus squamosus (Leafscale gulper shark)
- Centroscymnus coelolepis (Portuguese dogfish)
- Galeus melastomus (Blackmouth catshark)
- Centroscymnus crepidater (Longnose velvet dogfish)
- Centroscyllium fabricii (Black dogfish)
- Somniosus microcephalus (Greenland shark)
- Hexanchus griseus (Six-gilled shark)
- Etmopterus spinax (Velvet belly)
- Apristurus spp. (Deep-water catsharks)
- Chlamydoselachus anguineus (Frilled shark)
- Deania calcea (Birdbeak dogfish)
- Dalatias licha (Kitefin shark)
At the time of writing (January 2025), deep-sea sharks listed in NEAFC Recommendation 09:2024 (see below) ‘for which catches should be regulated in ICES sub areas I to XIV (NEAFC Category 2: ‘no directed fisheries and minimise all bycatch’) are protected under EU Regulation 2018/2025 Annex I. This regulation enforces a zero TAC in Union waters and applicable international waters of ICES subareas 5–10 and 12, meaning no targeting, retention (even bycatch), transhipment, or landing is allowed, except under strict bycatch monitoring. The protection has been retained in the UK following Brexit.
- Centrophorus granulosus, C. squamosus (Gulper sharks)
- Centroscyllium fabricii (Black dogfish)
- Centroscymnus coelolepis (Portuguese dogfish)
- Centroscymnus crepidater (Longnose velvet dogfish)
- Dalatias licha (Kitefin shark)
- Etmopterus princeps (Greater lanternshark)
- Apristurus laurussonii (Iceland catshark)
- Chlamydoselachus anguineus (Frilled shark)
- Deania calcea (Birdbeak dogfish)
- Galeus melastomus (Blackmouth dogfish)
- Galeus murinus (Mouse catshark)
- Hexanchus griseus (Bluntnose six-gill shark)
- Etmopterus spinax (Velvet belly)
- Oxynotus paradoxus (Sailfin roughshark)
- Scymnodon ringens (Knifetooth dogfish)
- Somniosus microcephalus (Greenland shark)
In addition, Regulation 2023/194 (Article 18) extends the zero TAC to all international waters for EU vessels, ensuring complete protection across the EU and ICES regions. Protection is also provided by NEAFC’s Recommendation on Conservation and Management Measures for Deep Sea Sharks in the NEAFC Regulatory Area from 2024 to 2027 (NEAFC Recommendation 09 2024), which prohibits directed fishing for deep-sea sharks in the NEAFC area. Contracting parties must submit all data on deep-sea sharks (i.e. bycatch data) to ICES for further evaluation of the state of the stocks.
The following general EU and UK regulations for deep-sea fisheries (ICES 2024) are also likely to protect deep-sea elasmobranchs.
Trawl and gillnets are banned in waters greater than 200m in the Azores, Madeira and Canary Islands areas since 2005.
- Since 2006, gillnets have been prohibited at depths below 200 m in the Northeast Atlantic Fisheries Commission (NEAFC) Regulatory Area (Rec 03 2006: Gill Nets 2006). This encompasses all international waters within the ICES area.
- Gillnets banned below 600 m in ICES areas 6.a-b, 7.b-c, 7.j-k and 12 since 2007 (however, a maximum bycatch of 5% of deep-water sharks in hake and monkfish gillnet catches was allowed (Council Regulation (EC) No 41/2007)).
- Ban on the “rasco (gillnet)” fishing gear at depths below 600 m since 2009 (EC Regulation 43/2009). However, this applies to a very small number of boats (4-6) in the Basque Country.
- Ban on bottom trawling at depths below 800 m in all EU waters since 2016 (Article 8, EU Regulation 2016/2336, also incorporated into UK law (as amended by S.I 2019/753)).
- Fishing with static nets (bottom-set gillnets, entangling nets, and trammel nets) in EU waters in some regions of the Eastern North Sea, Celtic Seas and Oceanic Northeast Atlantic (ICES divisions 3a, 6a, 6b,7b, 7c, 7j and 7k and ICES subareas 8, 9, 10 and 12 east of 27° W in waters with a charted depth of more than 200 m); is prohibited (Article 9, EU 2019/1241, also incorporated into UK law (as amended by S.I 2019/1312 and S.I. 2020/1542)).
- In 2022, the EU closed 87 VME’s (vulnerable marine ecosystems) that are between 400 - 800 m depth to all bottom contact fishing (Implementing Regulation (EU) 2022/1614).
References
ICES (2024). Report of the Working Group on Elasmobranch Fishes (WGEF). ICES Scientific Reports.







