NatureScot Research Report 1388 - Review of Scottish Seagrass Declines
Note: Published 20th January 2026 and updated 8th June 2026 with additional historical records
Published: 2026
Authors: Max Carstairs & Siân Carstairs
Cite as: Max Carstairs & Siân Carstairs. Review of Scottish Seagrass Declines. NatureScot Research Report 1388.
Contents
- Keywords
- Background
- Main findings
- Acknowledgements
- Introduction
- Seagrass Ecology
- Threats to Seagrass Populations
-
Data Sources (Availability, Quality and Comparability)
- Data gathering
- Assessment of national databases
-
Quality assessment of early seagrass bed maps
- Butcher, 1941 (International Wildfowl Inquiry)
- Ackroyd, Montrose Basin, 1972
- Nature Conservancy Council, Moray Firth studies, 1972 & 1978
- University of Aberdeen, Moray Firth, 1970-1981
- Minto, Clyde Estuary, BSc, 1974
- Rae, Nigg Bay, PhD, 1979
- Sphere Environmental Consultants, Nigg Bay, 1981
- McLusky and Roddie, Montrose Basin, 1982
- Fox, Yost and Gilbert, Northern Firths, 1986
- Marine Nature Conservation Review, 1987-1998
- Duncan, Nigg Bay, MSc, 1991
- Atkins, Caudwell and Herbert, Montrose Basin, 1991
- Hawker, Solway Firth, 1993 & 1994
- RSPB, Udale Bay and Nigg Bay, 1993, 2000 & 2011
- Quality assessment of contemporary seagrass bed maps
- Seagrass bed definitions and assessment of data comparability
- OSPAR water quality data set 1990-2008
- Assessment of Previous Studies of Seagrass Declines
- Seagrass Population Dynamics by Scottish Marine Region
- Discussion and Recommendations
Keywords
Seagrass; Zostera; Status; Ecology; Threats; Declines; Recovery; Restoration
Background
Declines of seagrass have been reported worldwide, dating back to the late 19th century. The aim of this study was to assess the functional ecology and current status of seagrass populations in Scottish waters, with a particular focus on reviewing evidence of seagrass declines, recoveries, and their potential causes. A review of population dynamics was carried out for each of the 11 Scottish Marine Regions. The findings are discussed in the context of future monitoring, conservation management (including seagrass restoration projects), and carbon sequestration.
Main findings
- A review of literature and available data revealed significant historical declines of seagrass across Scotland.
- The Zostera marina population in the upper Firth of Clyde appears to have suffered a serious historical decline, to the extent that its continued presence is uncertain. A significant historical decline of Z. marina also appears to have occurred in the Firth of Forth. Water quality issues have been linked to seagrass declines at these and other locations.
- Significant historical losses have been documented for subtidal Z. marina beds in remote rural areas such as the Outer Hebrides and Shetland.
- Reasons for declines were varied. In the Outer Hebrides and Shetland, the largest declines have been linked to the 1930s wasting disease epidemic but with subsequent losses also reported. Vast historical losses due to wasting disease were also reported in the Firth of Tay.
- The intertidal seagrass populations of the northern firths (Dornoch, Cromarty, Beauly and Moray) and the Solway Firth appeared to have remained unaffected by the wasting disease epidemic of the 1930s.
- Possible causes of recent declines at Berneray include suction dredging for razor clams and construction of a causeway between Berneray and Uist. Construction of the Eriskay Causeway has also been linked to Z. marina declines.
- Scallop dredging has reportedly been a cause of seagrass declines at Islay, Arran and Kintyre.
- A substantial historical reduction in the size of the Zostera noltii beds within Southannan Sands SSSI is attributable to land reclamation in the 1970s.
- Substantial recent recoveries of intertidal seagrass populations have been recorded in Montrose Basin, Loch Ryan and the Firth of Forth, associated with water quality improvements.
- The cessation of mechanical cockling in the Solway Firth during the early 1990s is considered to have facilitated noticeable seagrass population increases. Water quality improvements may also have contributed to recorded increases in this estuary.
Acknowledgements
We would like to thank NatureScot and in particular Brodie Thomas, Sarah Cunningham, Kelly Saunders, Ben James and Paul Longbottom for providing advice, documents and useful comments on the draft.
Our thanks is also given to the following for provision of useful shapefiles or survey reports:
- Rebecca Giesler, University of Highland and Islands
- Francis Williams and Isla MacLeod, Mossy Earth
- Katherine Knight, Scottish Marine Environmental Enhancement Fund
- Lucy Kay and Howard Wood, The Community of Arran Seabed Trust
- Clair McFarlan, Solway Firth Partnership
- Ethan Ross, University of Aberdeen
- Mark Hartl, Heriot Watt University
- Andrew Rodger, Namarra Projects, Berneray
- James Donaldson, The Botanist Foundation
- Steph Elliott, Royal Society for the Protection of Birds
- Duncan Donald, Botanical Society of Britain and Ireland
- We would also like to thank Clair McFarlan, Solway Firth Partnership for kindly giving permission to reproduce much material published in Carstairs, 2024.
Introduction
Seagrass beds are important habitats, providing shelter for a variety of marine life as well as having a role in carbon storage and coastal protection. In Scotland, seagrass beds are recorded from the Solway to Shetland, on both the east and west coasts. Seagrass beds are of national and international importance, protected as a Priority Marine Feature through Scotland’s National Marine Plan and as an OSPAR Threatened and Declining Habitat.
Significant declines in Scotland’s seagrass bed extent were noted during the 20th and 21st centuries. Declines have been attributed to poor water quality, the seagrass wasting disease caused by the protist Labyrinthula zosterae, and coastal development, such as the construction of causeways. Recent recoveries have been noted in the Firth of Forth, Solway Firth and Loch Ryan. In many areas, seagrass population trends, and reasons for changes in extent, are unknown.
With the growing interest in seagrass restoration, it is crucial to understand where and why declines are occurring, as well as where and why healthy beds continue to thrive, to inform enhancement related work effectively. This project aimed to understand more about these factors by compiling a review of available evidence from Scotland.
Historical context
The seagrass populations of Britain and Europe were evidently much more abundant in historical times than at present. Ørsted described seagrass beds as “savannahs of the sea” (Forbes, 1859). Zostera marina beds along the continental European and north-west Atlantic coasts were prolific enough for a diversity of uses, including upholstery and fertilising fields (Green & Short, 2003).
Accounts in historical literature indicate that extensive populations of Z. marina existed during the 19th and early 20th centuries in Scotland. Sonntag (1894) described Z. marina as occurring frequently at several locations in the Firth of Forth while Hennedy (1865) reported that the species occurred commonly in the Firth of Clyde. Miller (1857) described “submarine meadows” of Z. marina in the Cromarty Firth. In Shetland, Z. marina was called “marlie”, and its abundance gave rise to the name Marlee Loch, where it was once so plentiful that angling was hampered (Scott & Palmer, 1987). Z. marina also filled the head of Weisdale Voe so densely that a channel sometimes had to be cut to allow boat passage (Scott & Palmer, 1987). Large amounts of Z. marina were washed up on Shetland during autumn gales and were used for cattle bedding and stuffing mattresses (Scott & Palmer, 1987). In Orkney, it was used for thatching roofs and enriching fields (Urquhart, 1824). An advertisement appeared in The Farmer’s Magazine of 1822 for harvesting seagrass off the Scottish West Highlands by the tonne.
Historical declines of seagrass in Scotland appear to have occurred on a vast scale, mirroring those across the wider biogeographic area (Blois et al., 1961; Boström et al., 2003; Green & Short, 2003; Petersen, 1914, 1918; Blegvad, 1935). Localised declines of seagrass beds were recorded in the western North Atlantic during 1854, 1889, 1894, 1908, and 1915 (Cottam, 1934, 1935; Cotton, 1933; Martin, 1954). Declines in England occurred during 1920, 1921 and 1922, followed by a catastrophic loss - estimated at 90% - of Z. marina across Europe and the western North Atlantic during the early 1930s (Butcher, 1934, 1935, 1941; Cottam, 1935; Giesen et al., 1990; Ralph & Short, 2002; Tutin, 1942; Waycott et al., 2009). The scale of loss across Denmark (93%) was particularly evident, as beds had been mapped in detail by Petersen and co-workers in the early 20th century (Ostenfeld, 1908; Petersen, 1914, 1918; Blegvad, 1935; Green & Short, 2003). Further, less acute seagrass declines, as well as recoveries, have since occurred across eastern North America and Europe (Dexter, 1985; Green & Short, 2003; Jones & Unsworth, 2016; Waycott et al., 2009).
During the 1930s decline event, it was noted that seagrass mortality was associated with a high prevalence of the pathogenic organism Labyrinthula sp. (Renn, 1935, 1936). Labyrinthula macrocystis was confirmed as the causative agent of wasting disease outbreaks along the east coast of North America during the 1980s (Muehlstein et al., 1988; Muehlstein et al., 1991; Short et al., 1987).
It is now widely regarded that this pathogen, and other related ones, are a natural feature of seagrass beds at relatively low levels (Tutin, 1938; Sullivan et al., 2018). However, adverse environmental factors are likely to stress seagrasses, enabling pathogens to proliferate (Rasmussen, 1977). A growing body of evidence indicates that environmental deterioration, particularly the eutrophication of coastal environments, is responsible for large scale global losses of seagrass beds (Green & Short, 2003; Jones & Unsworth, 2016; Lillebø et al., 2011; Ralph et al., 2006; Short & Burdick, 1995).
Conservation value of seagrass beds
Seagrass beds are of high conservation value and perform many beneficial physical functions, including reducing tidal energy (Figure 1) and coastal erosion, stabilising coastal sediments, aiding the establishment and protection of saltmarshes, and improving water quality (Bos et al., 2007; Gambi et al., 1990; Ganthy et al., 2015; Gutiérrez et al., 2011; Hughes, 2001; Hughes & Paramor, 2004; Lillebø et al., 2011). Seagrasses are valued for their relatively high rates of carbon sequestration (Cunningham & Hunt, 2023; Duarte et al., 2005; Pidgeon, 2009), though further research is required at the genus, species and infraspecific level to improve understanding of sequestration rates for seagrasses across north-west Europe.

Figure 1. Wave development due to raised patch of Zostera noltii, The Dhoon, Solway Firth. Image © Max Carstairs.
Seagrass beds significantly enhance the structural diversity of marine environments, particularly within large estuaries containing extensive sand and mudflats. Intertidal and subtidal seagrass beds provide food resources and cover for a variety of invertebrates and fish. The pipefish Syngnathus typhle is almost entirely restricted to seagrass beds (BRIG, 2008). Seagrass beds may also act as nursery grounds for juvenile fish (Bertelli & Unsworth, 2014; Lilley & Unsworth, 2014; Pihl et al., 2006).
Seagrass beds also provide food through leaves, seeds, and rhizomes, for migratory and overwintering wildfowl (Fox, 1996). On the east coast of Scotland, seagrass beds provide feeding grounds for overwintering pale-bellied brent geese, which breed on Svalbard. This is the smallest population of geese in Europe and is classified as having unfavourable conservation status (Fox et al., 2010) due to a lack of recovery following a population crash related to severe declines of Zostera in the 1930s.
Seagrass protection in Scotland
Legislative protection is afforded to UK seagrass populations that occur within statutory protected areas, such as Sites of Special Scientific Interest (SSSI), Special Areas of Conservation (SAC), Special Protection Areas (SPA), Nature Conservation Marine Protected Areas (NCMPA), and Ramsar sites. In Scotland, seagrass beds appear in the citations for one NCMPA, five Ramsar sites, 11 SACs and 28 SSSIs.
The vast majority of Scottish intertidal seagrass beds fall within the EU Habitats Directive Annex I habitats “estuaries” and “mudflats and sandflats not covered by sea water at low tide”. Many sublittoral stands of Z. marina will fall within the Annex I Habitats “large shallow inlets and bays” and “sandbanks which are slightly covered by sea water all the time”. Zostera also occurs within the Annex I habitat “coastal lagoons”. Seagrasses are not listed as Annex II species.
A network of Marine Protected Areas (MPA) has been designated around the UK to meet international and national commitments, such as the OSPAR Convention and the UN Convention on Biodiversity, and to assist in achieving Good Environmental Status under the UK Marine Strategy Regulations. MPAs are designed to protect Scotland’s seas, marine life and habitats from damage caused by human activities. Fishing restrictions are in place within some Scottish MPAs to prevent mechanical disturbance of the seabed, and the Scottish Government plan to introduce fisheries management measures in existing MPAs where such measures are not already in place (Scottish Government, 2024).
Seagrass beds are a Scottish Priority Marine Feature (PMF) and a Scottish Biodiversity List Habitat. Seagrass beds have also been added to the OSPAR Convention’s list of Threatened and/or Declining Species and Habitats.
There is a policy requirement to conserve PMFs as “wider seas” biodiversity. General policy 9b of the National Marine Plan states that:
“Development and use of the marine environment must not result in significant impact on the national status of Priority Marine Features”.
Therefore, PMFs such as seagrass beds must be considered within the planning process for proposed development projects.
The water quality of Scottish coasts is currently managed through the Water Environment and Water Services (Scotland) Act 2023. The aim of the Act is to protect, and, where necessary, restore waterbodies to “good” status. This requires both good ecological and good chemical status to be attained. Seagrasses are a key component used to monitor ecological status of transitional and coastal waters (Foden et al., 2010).
Species coverage, taxonomy and recording accuracy
Species coverage
English-language names for seagrass include eelgrass, grass-wrack and wigeon grass. The term seagrass generally refers to members of the genus Zostera, though it can also be extended to include tasselweeds of the genus Ruppia. For example, Zostera sp. and Ruppia sp. are both encompassed within the seagrass bed PMF. Wigeon grass generally refers to Ruppia sp., which can be found in estuaries and brackish waters such as coastal pools and lagoons. Species of Zostera occur much more extensively in coastal environments and are the subject of this study.
Taxonomy
Two species of seagrass, Z. marina and Zostera noltii, occur in Scottish coastal waters, though three species were recognised until recently (Table 1).
| Common name | Habitat | Latin name (WoRMS, 2025) | Latin name (Stace, 2019) | Historical pseudonyms |
|---|---|---|---|---|
| Broad-leaved seagrass | Subtidal | Zostera marina
| Zostera marina var. marina | Zostera marina var. genuina |
| Narrow-leaved seagrass | Intertidal | Zostera marina
| Zostera marina var. stenophylla | Zostera angustifolia, Zostera marina var. angustifolia*, Zostera hornemanniana |
| Dwarf seagrass | Intertidal | Zostera noltii | Zostera noltei | Zostera nana |
Z. marina occurs in two different forms, a long, broad-leaved form that occurs subtidally and a smaller, narrow-leaved form that inhabits the intertidal zone.
The taxonomy of the two different types of Z. marina has long been debated within the scientific community. The intertidal form of Z. marina was historically considered a separate species, called Zostera angustifolia. However, recent genetic studies did not support this classification (Becheler et al., 2010; Coyer et al., 2013; Gagnon et al., 1980; Olsen et al., 2013). The taxon is currently considered to be either an ecophene or ecotype of Z. marina (Stace, 2019). An ecophene is a growth form which merely exhibits phenotypic variations in relation to habitat differences. In an ecotype, the difference in appearance is heritable i.e. it is underpinned by genetic adaptations to a particular habitat (Stace, 1989). Ecotypes remain fully interfertile within the sphere of the species and related ecotypes (Stace, 1989) and may evolve into separate species (Begon et al., 1990).
Several workers have referred to Z. marina ecotypes (Beca-Carretero et al., 2024; Olsen et al., 2013). Olsen et al. (2013) considered that the two European Z. marina forms were ecotypes though genetic differences likely occurred at a higher resolution than their methodology could detect. Genetic differences in Z. marina ecotypes along the west coast of North America were inferred by Backman (1991). Rasmussen (1973) reported that intertidal Z. marina plants grew wider leaves when planted sub-tidally. However, during seed-based restoration exercises in the UK, Z. marina plants have overwhelmingly grown in the form associated with the donor rather than the recipient habitat e.g. seeds from subtidal sites have expressed as perennials when planted intertidally (R.J. Lilley, pers. comm.).
The narrow-leaved, intertidal form of Z. marina occurring in Scottish waters may therefore be a genuine ecotype with distinct heritable characteristics rather than merely an ecophene. However, given the uncertainty, different forms of Z. marina are referred to as varieties (which encompasses both ecotypes and ecophenes) in this report, following Sell & Murrell (1996) and Stace (2010, 2019). Discussion of taxa to infraspecific level was considered essential in this report, given the marked ecological differences (e.g. in distribution, habitat, and growth form). These differences assist in assessing the nature of population declines, natural recovery, and managed restoration.
Identification and recording issues
Historical recording issues were noted for the intertidal Zostera marina variety. The name Zostera marina var. angustifolia was published in 1816 by Hornemann with the taxon subsequently upgraded to species level by Reichenbach in 1845 under the name Z. angustifolia. However, the plant was variously referred to as a separate species, a variety of Z. marina and in some cases was not mentioned at all within the British floras spanning from 1850 to 2019 (Hooker & Arnott, 1850; Bentham, 1858; Clapham et al., 1952; Butcher, 1961; Sell & Murrell, 1996; Stace, 1991, 2019). As many botanists only record plants to species level, a number of occurrences of the intertidal variety have historically been documented merely as Z. marina.
Seagrasses can be difficult to identify to genus, species and variety, and identification issues are evident in the historical literature and survey reports. These included differentiation of the following taxon pairings: Ruppia sp. and Z. noltii; intertidal Z. marina and Z. noltii; intertidal and subtidal Z. marina varieties. Accounts in which the variety is not given or apparent are referred to in this report as Z. marina.
The accuracy of records used in this project was assessed through accompanying habitat descriptions and specimen photographs, where available. Geographic location was also taken into account.
Seagrass Ecology
Taxon distributions and habitat preferences
Zostera noltii
Z. noltii typically occurs in the mid to upper intertidal zone. The upper distribution appears to be controlled by the position of the average groundwater line (near mean high water neaps) below which tidal flats remain wet or moist during low tide (Carstairs, 2024). In the Solway Firth, Z. noltii occurs densely up to this seepage line but not above it (Carstairs, 2024). The species also occurs in saline lagoons.
Z. noltii is a characteristic species of the east coast estuaries and the Solway Firth, with a small population in the Firth of Clyde. It also occurs within sea lochs such as Loch Ryan in Dumfries and Galloway and those on the eastern side of the Sound of Jura in Argyll. Sparsely distributed historical records extend up the west coast from Islay to the Hebrides, as far north as Lewis (Annex 1). The species was reported from Sandi Sand, Deerness and Finstown in Orkney during 2022 (Unsworth et al., 2025), although these records may well be misidentifications. Otherwise, these would represent a northwards expansion of the species in Scotland. Plants identified as Z. noltii from the Loch of Hellister, Shetland in 1993 are considered most likely to be misidentifications of Ruppia maritima following genetic testing of samples collected from the same location in 2007 (BSBI, 2024).
Zostera marina
The intertidal variety of Z. marina typically occupies the mid and lower shore. It can occur up to the average groundwater line, particularly where there is standing water e.g. in permanent pools (Carstairs, 2024). It is predominantly found in the sand and mudflats of large Scottish estuaries on the east coast and the Solway Firth (Carstairs, 2024; Rae, 1979). Z. marina can also thrive on rocky shores where fine sediments accumulate in the interstitial spaces between larger sediments such as gravels and pebbles that have collected within depressions and fissures in the bedrock. It is restricted to relatively sheltered areas of rocky shores which remain water logged at low tide (Carstairs, 2024). There are scattered historical records in the Firth of Clyde and up the west coast to Loch Carron, Wester Ross. In the Outer Hebrides, Z. marina occurs intertidally from Barra to North Uist (Annex 1).
Many of the records of Z. marina from the east coast are considered likely to represent occurrences of the intertidal variety (Cleator, 1993). Similarly, historical records of Z. marina in the Solway Firth appear in several publications, although no habitat descriptions signifying the presence of subtidal Z. marina were encountered during the current project. A report of Z. marina between Arbigland and Southerness on the Solway Firth has been repeated in the literature, though the New Statistical Account of Scotland (1845), states that the plant occurred in “flat rocky pools” which clearly describes a habitat of the intertidal variety. Plants present in rock pools may be more closely related to permanently submerged subtidal populations than intertidal beds on mudflats.
Subtidal Z. marina is occasionally exposed during extremely low tides. It is primarily distributed within relatively sheltered waters along the coasts of western Scotland, the Inner and Outer Hebrides, Orkney and Shetland (Annex 1). These coastlines are structurally diverse and indented with coastal embayments, as well as sea lochs along the west coast and Hebrides and voes in Shetland. Z. marina populations can also occupy coastal lagoons, shallow sounds between small islands and waters sheltered by a diversity of features including volcanic dykes. A few small stands also occur along the east coast, e.g. at Rosemarkie Bay and Burghead in the outer Moray Firth.
Around the British Isles Z. marina typically occurs down to 4 m but can extend down to 9 m (Stace, 2019). The species has been recorded growing at greater depths in Ventry Bay, Ireland, where Whelan & Cullinane (1985) found continuous beds from 0.5 m to 10 m, with patches extending to 13 m below chart datum. This is the deepest recorded contemporary Z. marina population in north-west Europe, although the species has been historically recorded at 17 m depth in the German Baltic Sea (Boström et al., 2014).
The depth to which Z. marina grows is primarily governed by the amount of light available for photosynthesis. High nutrient concentrations in the water column may promote the growth of phytoplankton and algal epiphytes, which intercept light (Figure 2). Water clarity may also be reduced by suspended sediments.

Figure 2. Algal epiphytes on Zostera marina in Wig Bay, Loch Ryan. Image © Max Carstairs.
Sediment composition and associated chemical status are also influential. High concentrations of hydrogen sulphide and low oxygen concentrations can be deleterious to seagrass survival, even where sufficient light is available for growth (Holmer & Bondgaard, 2001). Low amounts of silt and clay in predominantly sandy sediments with relatively low concentrations of hydrogen sulphide and high oxygen concentrations facilitate seagrass proliferation at the greatest depths (Krause-Jensen et al., 2011). The depth at which seagrass grows at any given location therefore reflects both light availability and sediment conditions (Krause-Jensen et al., 2011). A historical reduction in the depth at which Z. marina grows has been reported from Danish and German waters and is evidence of a deterioration of environmental conditions (Boström et al., 2014).
Growth, reproduction, and dispersal mechanisms
Z. marina and Z. noltii reproduce asexually via the lateral spread of clonal rhizomes and sexually via seed production (Marbà & Duarte, 1998; Ackerman, 1983). Rhizomatous growth is primarily responsible for the maintenance of local populations while sexual reproduction improves genetic diversity and facilitates the colonisation of distant areas.
Plants can survive in salinities of around 3‰ to 40‰ and in sea temperatures from close to zero up to around 30°C with variation occurring in relation to locally adapted populations (Blok et al., 2018; Setchel, 1929; McRoy, 1969; Salo et al., 2014; den Hartog, 1970; Schou et al., 2023). Fonseca and Uhrin (2009) considered that photosynthesis may be limited where salinity is below 10‰. The optimum temperature range for growth, flowering, and germination appears to be approximately 10°C to 20°C (Nejrup & Pedersen, 2008; Davison & Hughes, 1998; Moore et al., 2014; Setchel, 1929; Blok et al., 2018). De Cock (1981) found that the rate of male flowering increased with temperature. The relationship between flowering and temperature may account for the occurrence of large Z. marina beds comprised of single clones in northern latitudes.
Pollination of Z. marina occurs at and beneath the surface (Cox et al., 1992). Pollen rafts form on the water’s surface at low tide and are driven by currents and wind into seagrass stigmas. Pollen grains are adhesive, filiform, and of the same specific gravity as seawater, maximising the chance of contact with stigmas present around the same elevation in the water column (De Cock, 1980). Pollination may also involve pollen transportation by invertebrates, as seen in the red seaweed Gracilaria gracilis by the isopod Idotea balthica (Lavaut et al., 2022) and the seagrass Thalassia testudinum by crustaceans (van Tussenbroek et al., 2012). Zostera beds are known to contain abundant mobile amphipod and mysid crustaceans (Davison & Hughes, 1998), meaning that pollen transportation by invertebrates may also occur in Zostera beds.
Zostera species can exhibit a variety of growth and reproduction strategies. For example, Z. noltii populations around the eastern Scottish coasts are perennial and primarily reproduce via vegetative growth (Rae, 1979). This is also the case for subtidal Z. marina populations (Davison & Hughes, 1998; Thomson et al., 2014).
Rae (1979) found that intertidal Z. marina populations along the east coast of Scotland predominantly exhibited an annual lifestyle, with reproduction facilitated overwhelmingly via seed dispersal. An annual habit was observed at Montrose Basin, Beauly Firth, Loch Fleet, Cromarty Firth and Dornoch Firth. Populations on the south shore of the Tyne Estuary, East Lothian, also utilised this strategy, though the population on the north shore was perennial and maintained by rhizomatous growth (Rae, 1979). Jackson (1999) reported that the intertidal Z. marina population at Finstown, Orkney was annual.
The most likely reasons for an annual habit in Scotland are sensitivity to frosts, and environmental perturbations in the lower intertidal zone including winter wave action and associated sediment redistribution. Z. noltii appears to be a hardier plant, and this may account for a perennial life cycle in a challenging environment.
The perennial life history strategy of Z. marina is related to habitat stability. Perennial subtidal Z. marina populations in Scotland occur in relatively clear waters. A perennial intertidal Z. marina bed was observed at Dunure, Ayrshire, occupying habitat with similar environmental conditions to subtidal populations (Figure 3). The mid and lower parts of the bed were situated in large, extensive rockpools with stable mixed-sediment bottoms in many places, facilitating the growth of seagrass. These pools held water during low tide. Upper shore depressions in small mixed sediment over bedrock drained slowly, with many retaining permanent water for most of the low tide period. The water was clear with no visible turbidity (Carstairs, 2024).

Figure 3. Perennial Zostera marina in an intertidal pool at Dunure, Ayrshire, 28th February 2025. Zostera on mixed sediments in foreground with bedrock encrusted by coralline algae in the background. Image © Max Carstairs.
The western fringe of Auchencairn Bay, Solway Firth, contains a dense cockle bed, which stabilises the sediments, giving a firm surface with poor drainage. In this habitat, perennial Z. marina occurs frequently, with occasional patches in the nearby mudflats (Figure 4). However, only very sparsely distributed perennial plants occur across the vast majority of the bay, which predominately supports an annual Z. marina population.

Figure 4. Zostera marina rhizomes and rootmass exposed at naturally eroding bed edge. Several leaves also visible, Auchencairn Bay, Solway Firth. 26th February 2025. Image © Max Carstairs.
Perennial Z. marina, therefore, occurs in relatively stable habitats, particularly in terms of permanent submergence, sediment stability and light penetration. Permanent standing water in the intertidal zone is likely to reduce the risk of heat damage, desiccation, and damage from frosts, while sediment stability prevents deep burial or exposure of rhizomes.
Seed dispersal, longevity and germination
There has been extensive research into the reproduction of Z. marina, including seed dispersal, longevity and germination. In contrast, few studies have been published for Z. noltii and therefore the species is not reviewed in this section.
Deposition rates for seeds of Z. marina have been calculated between 1,500 to 6,000 seeds deposited per m2 per year, with local rates reaching up to 15,000 seeds deposited per m2 per year (Costa, 1988). Dooley et al. (2013) found that the viability of seeds stored in seawater decreased over time, from 78% in one-year-old seeds to 32% in four-year-old seeds. Viability of sediment seed banks appears to last less than 12 months (Jarvis & Moore, 2010). Germination rates of Z. marina seeds are naturally low but increase with decreasing salinity (Phillips, 1983). Conversely, seedling development and survival are inhibited at low salinities; Xu et al. (2016) found that in northern China, most seedlings failed to establish in brackish conditions and that the optimal salinity for germination, seedling growth and survival appeared to be ≥20‰.
Seeds of aquatic plants typically sink close to the parent plant, which limits their ability to colonise new areas (Davis, 1985). High proportions of Z. marina seeds have been recorded settling near the parental source: 95% within 30 m (Ruckelshaus, 1996) and 80% within 5 m (Orth et al., 1994). Churchill et al. (1985) reported that an oxygen bubble formed in the rhipidium, and when the capsule ruptured to release its contents, gas bubbles attached to 5‑13% of seeds, facilitating an average dispersal distance of 21 m and a maximum of 200 m.
Z. marina is also capable of dispersing seeds over longer distances through diaspores i.e. broken-off spathes, rhipidia and reproductive shoots, which can be transported on currents at the water surface (See Figure 5). Källström et al. (2008) modelled a potential dispersal distance of rhipidia of up to 150 km in one season along the Swedish west coast. Hosokawa et al. (2015) found that only a small proportion of Z. marina seeds were dispersed via positively buoyant diaspores. The presence of Z. marina washed up on Fair Isle on four separate occasions in 1982, 2017 and 2018 (BSBI, 2024; Quinteros Peñafiel et al., 2017) may represent long-distance dispersal of plant material in Scottish waters, as seagrass beds have never been recorded around the island. Rhizome fragments transported via coastal currents can also be a mechanism of reproductive dispersal (Hemminga & Duarte, 2000).
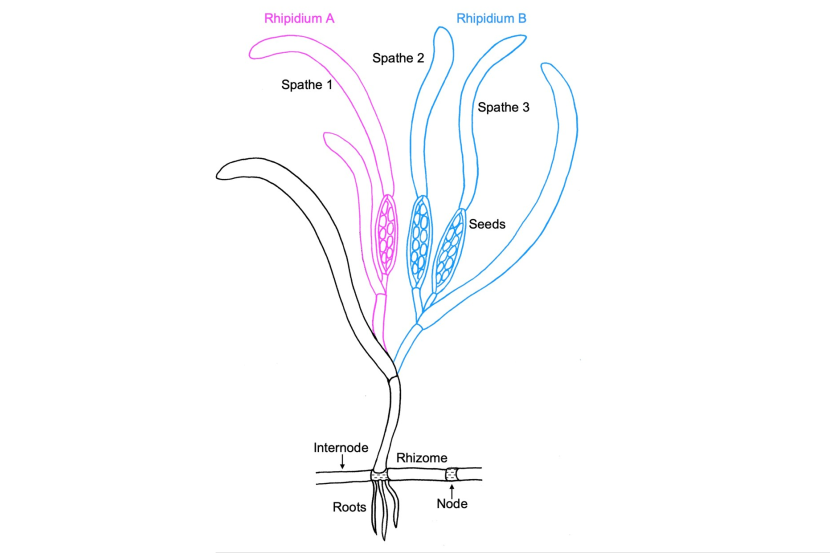
Figure 5. Schematic diagram of Zostera marina reproductive structures and their terminology. Image adapted from Sgambelluri et al. (2024) by Emma Carstairs.
Click for a full description
In the diagram, there are two rhipidiums. A lateral rhizome comprised of longitudinal sections separated by nodes where short roots sprout into the substratum is shown at the bottom of the plant. The shoot with leaves, rhipidia and spathes is depicted arising from a rhizome node.
Dispersal of plant material by waterbirds is a well-documented phenomenon and is often implicated in the discovery of plant species far from their known distribution (Reynolds et al., 2015). Diaspore (seeds, shoots and other reproductive structures) transportation by animals can be either internal (e.g. through ingestion) or external (e.g. by adhering to feathers or skin). Sumoski & Orth (2012) demonstrated that Z. marina seeds can remain viable after passage through the digestive systems of fish and waterfowl. This is likely due to the robust seed coatings, which can remain present in marine sediments for centuries (Costa, 1988) and even longer, potentially becoming sub-fossils (Godwin (1975) gave details of an unpublished record of Z. noltii fruit from a Cornish estuarine bed “possibly of late Pliocene age”.)
Evidence for bird dispersal comes from a detached specimen of Zostera found floating in Loch Tarff, Scotland, in 1972 (McCallum Webster, 1978). The freshwater loch is landlocked and situated around 50 miles from the nearest coastline (Annex 1).
Natural population dynamics
Seagrass beds naturally go through periods of growth and retraction. Beds can change in extent and density seasonally as well as from year to year. Changes may occur because of seagrass life cycles and physiology, or due to environmental conditions. The distribution of intertidal seagrass populations is strongly governed by sediment dynamics. Natural population dynamics can potentially confound assessments of anthropogenic change in seagrass bed extent and density between years.
Seasonal development and senescence
The extent and density of seagrass beds can change throughout the year and can vary between species and varieties. For example, Z. noltii in Nigg Bay (Cromarty Firth), flowered and produced seed in July and August (Rae, 1979). The leaves of Z. noltii were found throughout the winter, though exhibited a dark green colour, in contrast to the bright green of summer (Rae, 1979).
Rae noted that the seedlings of intertidal Z. marina in Nigg Bay were visible in February and numerous by March. Plants remained small well into May, when rapid growth commenced. Flowering began in June. A small number of plants survived from the previous year. By the end of July, some intertidal Z. marina plants had started to degrade, and the bed became progressively denuded from September onwards, with few green plants present in January (Rae, 1979).
The leaves of sublittoral Z. marina are evergreen with shorter, narrower winter leaves typically replacing summer leaves which shed during autumn (Scott & Palmer, 1987; Davison, 1997).
Inter-annual population fluctuations
Variations in bed distribution can occur between years. In annual populations, these are related to seed productivity, survival, and germination rates. In perennial beds, variations are determined by the survival of rhizomes over winter and their ability to produce new shoots in spring and summer. Underlying factors influencing these parameters include rates of sediment accretion and erosion, pollution levels and degree of sediment anoxia. Harlin & Thorne-Miller (1982) attributed inter-annual patch dynamics to patterns of sediment accretion. Blois et al. (1961) found that patches of sand could be deposited within a day over Z. marina beds, causing mortality if they were not washed away relatively quickly. Gaps were eventually colonised by rhizomes of nearby surviving plants. More recent evidence suggests cyclical processes in sediment anoxia may also create patchiness (Borum et al., 2005, 2014).
Many researchers have assessed changes in seagrass populations by comparing single year historical data with contemporary studies, largely due to the limitations of historical data. More recently, studies tracking changes over successive years have been undertaken, though their results typically show either a trend of decline or recovery in response to gradients of environmental pressure (Benmokhtar et al., 2023). While these studies are valuable, they provide little insight into natural population fluctuations. A better understanding of the natural variation within populations would enhance the robustness of long-term assessments of population change. However, the opportunity to undertake studies over a number of successive years in pristine environments is limited by the widespread prevalence of anthropogenic pressures.
Given these considerations, it is not surprising that a wide range of results have been reported for inter-annual variation of seagrass populations. For example, a Swedish study showed a 3% total increase between 2000 and 2004, with large inter-annual variation in some areas i.e. some individual beds were virtually absent in 2003 but extensive in 2004 (OSPAR, 2009).
The life cycle strategies of different seagrass species and varieties are likely to play a significant role in inter-annual variations in population size. Mature populations of perennial seagrass occupying all suitable habitat within a sheltered estuarine embayment would be expected to remain relatively stable in terms of size and location, in the absence of disturbance or pollution events. Stability is facilitated by the survival of rhizomes over the winter, from which new shoot growth arises in spring. Data from Pembrokeshire appears to support this, showing an expanding Z. noltii population at Angle Bay, and a relatively stable population at Pembroke Dock (Figure 6).
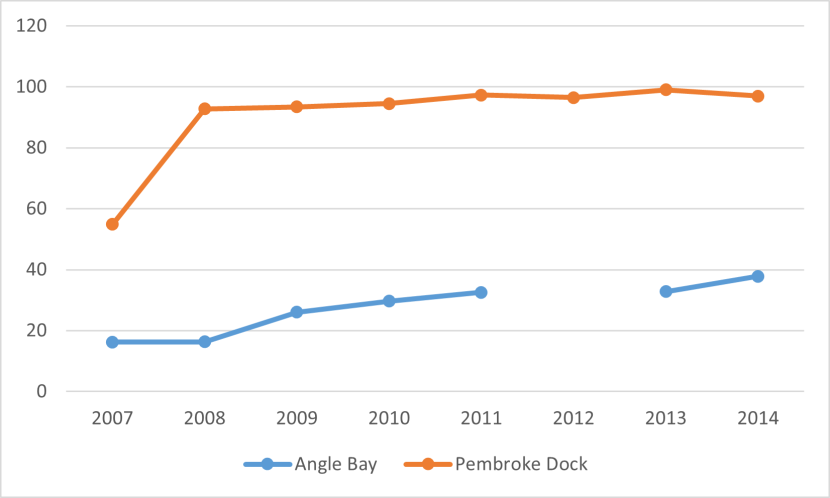
Figure 6. Time series data of Zostera noltii bed size (ha) from two sites in Pembrokeshire, Wales. Constructed from data in Bertelli et al. (2018).
Click for a full description
A graph of time series data depicting the coverage in hectares of Zostera noltii at Angle Bay and Pembroke Dock between 2007 and 2014. Seagrass coverage at Pembroke Dock is depicted in red and shows a steep increase between 2007 and 2008 after which coverage stabilised with the line flattening out. Seagrass coverage at Angle Bay is depicted by a blue line which is flat between 2007-2008 but then rises into 2014. No data was recorded for Angle bay in 2012.
In contrast, annual populations of intertidal Z. marina could be anticipated to fluctuate more widely due to a range of biological and environmental factors. These factors are likely to include the amount of seed production, the degree of rough weather during the seed dispersal phase, the depth of seed burial by sediments over the winter period, and the rates of germination and seedling survival during spring. Support for this assertion comes from limited studies at Auchencairn Bay on the Solway Firth. The large seagrass bed at Auchencairn Bay contains a Z. noltii population in the upper and mid-shore, with Z. marina present in the mid shore and extending far into the lower shore (Figure 7). A comment entered in to the Seagrass Spotter database (Unsworth et al., 2025) stated that:
“It is the most dynamic field site I visit. Every year the bed is different, sometimes it’s incredibly vast and goes out into the distance for 30 mins easily and other years it’s much more patchy”.
This statement indicates that the Z. marina component of the seagrass bed varies extensively due to an annual lifecycle strategy. A brief field visit was made by the current authors to Auchencairn Bay on 26th February 2025. Rhizomatous Z. noltii plants were observed in substantial quantities. In contrast, Z. marina plants were generally absent on the open mudflats with only small, sparsely distributed patches of rhizomatous plants observed. The intertidal Z. marina bed at Auchencairn is therefore predominantly annual.
Most of the larger intertidal seagrass beds in the Solway Firth are mixed, with an extensive Z. marina component extending from the mid to lower shore. This appears to be feature of recovering or relatively healthy beds (Carstairs, 2024). The frequent occurrence of mixed species beds presents a significant challenge when assessing intertidal population changes, particularly as many monitoring studies do not produce maps differentiating between zones of Z. noltii and Z. marina.
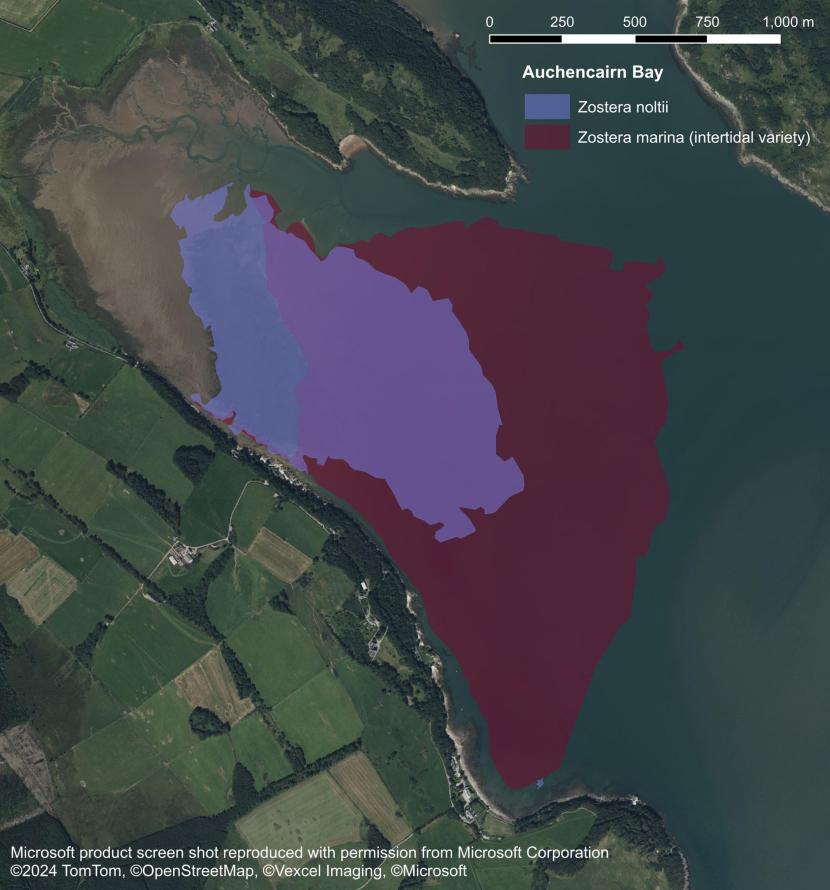
Figure 7. Intertidal Zostera noltii and Zostera marina distribution in Auchencairn Bay, 2023, Carstairs (2024). Areas where both Zostera noltii and Zostera marina are found are shown where the two polygons overlap (lilac coloured polygon).
Die-off events
Seagrass die-off events can occur as a result of high temperatures and calm conditions, hyper-salinity, and a combination of low oxygen and high hydrogen sulphide concentrations in benthic sediments (Holmer et al., 2005; Pedersen et al., 2004). Several of these factors are interconnected. For example, as temperature rises, oxygen concentrations fall, increasing the potential for development of anoxic conditions in the water column and especially in benthic sediments. These conditions may weaken seagrass to the point where wasting disease can proliferate.

Figure 8. Wasting disease lesions on Zostera marina, Solway Firth. Image © Max Carstairs.
Subtidal populations of Z. marina intercept fine particles in the water column, causing their accumulation on the seabed, where they are bound and retained by seagrass roots (Wilson, 1949). The development of fine sediment environments is associated with an increase in hypoxia and concentrations of toxic metabolites, particularly hydrogen sulphide. Z. marina exhibits structural and functional adaptions to low oxygen levels in the sediment (Borum et al., 2012; Penhale & Wetzel, 1983) though extremely low oxygen levels are detrimental (Penhale & Wetzel, 1983; Smith et al., 1988; Zimmerman & Alberte, 1996) and have been linked to seagrass die-off events (Baden et al., 2003; Borum et al., 2005, 2014).
These observations raise the possibility that long-term cyclical die-off events may be a natural feature of seagrass populations. This process occurs during population development of another angiosperm, the invasive cord grass Sporobolus anglicus, which modifies intertidal sediment profiles over decades, creating anaerobic conditions. It is possible that seagrass populations experience similar cyclical long-term die-off events, recolonising once sediment quality has recovered naturally. However, the frequency of occurrence and spatial extent of these events within seagrass beds is unclear. Historical descriptions and data from Scotland presented in this report demonstrate continuity of bed presence at intertidal and subtidal sites over decades and centuries suggesting that die-off events under natural circumstances do not affect entire beds.
Meteorological events
Damage to seagrass populations can occur naturally due to meteorological events, with the extent of bed loss proportional to the intensity of the event. Hurricanes and storms have caused damage and complete seagrass bed losses (Costa, 1988; den Hartog, 1987; Orth & Moore, 1983). Storm-damaged beds were found to recover within ten years (Costa, 1988). Storms are also associated with increases in suspended sediments, which intercept light and can potentially settle on and smother seagrass beds. The frequency and intensity of UK storms has recently increased, and this is projected to increase further over the coming century with climate change (Met Office, 2024).
Frost, snow, and ice may cause seagrass mortality via temperature-induced tissue damage, and ice may also lead to mortality via scouring. Miller (1854) related that subtidal beds of Z. marina on the Cromarty Firth were very susceptible to frost and that he had observed damage to large quantities of seagrass:
“Wherever they rise to the surface, seem very susceptible of frost. I have seen the shores strewed with a line of green Zostera, with its spikes charged with seed, after a smart October frost, that had been coincident with the ebb of a low spring-tide.”
Frost has been reported to have damaged intertidal seagrass beds in Wester Ross, Cornwall and the Isle of Wight (Covey & Hocking, 1987; Critchley, 1980). Ice damage to Z. marina beds in Denmark and eastern Canada was reported by Blegvad (1929) and Robertson & Mann (1984) respectively. A Z. marina bed at Midtown, Loch Ewe, Wester Ross succumbed to frost during the extremely cold winter of 2010 after which it never recovered. The bed became badly frosted on 2nd March when a very cold spell coincided with a remarkably low spring tide (D. Donald, pers. comm.).
Ice formation is a fairly rare occurrence in Scottish coastal waters, particularly along the western seaboard, due to the influence of the Gulf Stream, though frost, snow, and ice on the intertidal shore are more frequent (M. Carstairs, pers. obs.). No accounts were found of damage to seagrass beds in Scotland from ice-scouring, though this may occur inconspicuously. Opportunistic studies on leaf, rhizome, and seed survival during prolonged periods of cold winter weather would enhance the limited understanding of these factors in Scotland.
Hydro-morphodynamics
The upper limits of Z. marina and Z. noltii in the intertidal zone are set by the position of the average groundwater line (Carstairs, 2024). This line (also referred to in the literature as the groundwater seepage line and the effluent line) is defined as the juncture between the slope near the top of the beach and the level ground which covers most of the sand/mud flat (McLachlan & Defeo, 2018). The line is situated close to mean high water neaps (Jackson & Short, 2020). Water derived from groundwater seepages in combination with tidal drainage back through sediments generally remains present at or very close to the surface below this line on tidal flats during low tides and thereby minimises the risk of seagrass mortality through desiccation (Carstairs, 2024).
Sediment accretion and erosion
Seagrass populations occur in a variety of habitats around Scotland. All of these contain relatively fine sediment either at or near the surface of the substrate. For example, Z. marina may grow in tide-washed rockpools and in subtidal channels with moderate flows where at least small amounts of fine sediments accumulate within features such as gravel beds. The larger sediment types prevent fine sediments from being washed away (Carstairs, 2024). Seagrass is unsuited to highly mobile sediment environments and has not been observed by the authors on high energy dissipative beaches with mobile sandbars which migrate seasonally. Lower energy habitats comprised of extensive accumulations of soft sediments have great potential for slower natural migration and also exploitation by seagrass. Seagrasses stabilise and may modify habitats, promoting favourable conditions for future growth and expansion. The dynamics of these processes are discussed below.
The rates of intertidal accretion and erosion in sand and mudflats are significant natural factors governing seagrass populations. Ranwell et al. (1974) found that intertidal Zostera occurred where annual fluctuations in sediment level were less than approximately 7 cm. Rae (1979) found that the main distribution of intertidal Z. marina and Z. noltii in Nigg Bay occurred where surface levels varied by less than 1 cm.
Seagrass leaves intercept fine sediment and deflect currents over beds so that within dense subtidal Z. marina beds, flow velocity is low, facilitating sediment accretion and retention (Fonseca et al., 1983; Gambi et al., 1990; Peterson et al., 2004; Reidenbach & Thomas, 2018). Accreted sediments may also be bound by the mass of rhizomes and roots which can lead to development of substantial sediment banks which may extend into the lower intertidal zone. As rhizome growth can only proceed at a slight angle to the horizontal, seagrasses are unable to cope with substantial amounts of sediment deposited over relatively short timeframes and die if these are not washed away soon after deposition (Blois et al., 1961).
Several authors stated that sediment banks were washed away by currents once Z. marina was killed during the 1930s wasting disease epidemic (den Hartog, 1970; Rasmussen, 1973; Wilson, 1949). Photographic evidence presented by Wilson (1949), demonstrated an approximately 60 cm reduction in height of sediment banks around mean low water springs, following seagrass mortality during the 1930s. Rasmussen (1973) also described a lowering of beach elevation profiles in Denmark following the wasting disease epidemic.
The observations made by Wilson (1949), Blois et al. (1961) and Rasmussen (1973) indicate that Z. marina accretes sediment over long periods of time. Orth (1977) noted the stabilising effect of Z. marina during storms in Chesapeake Bay, with banks covered with Z. marina remaining stable, whereas unvegetated banks eroded by up to 20 cm. The loss of fine sediments and subsequent development of either shifting coarser sands, or stony shores with seaweeds, appears to be a factor contributing to the slow recovery of perennial Z. marina populations (den Hartog, 1970; Rasmussen, 1973; Wilson, 1949). Isolated developing plants are susceptible to being washed out of relatively coarse, loose sediments, in comparison to those developing in established beds with slow flows and an accumulation of fine benthic sediments (Moore, 1958).
Channel migration
Estuarine channels are naturally dynamic, and their positions characteristically migrate over time within the ecosystem. Anthropogenic changes to catchment flow regimes may, however, influence estuarine hydrology and channel movements.
Hydro-morphological changes, particularly relating to the course of the main channels flowing through estuarine embayments, have led to significant changes in the distribution of seagrass beds at Skyreburn Bay and Rough Firth in the Solway Firth (Carstairs, 2024). The main channel at Rough Firth migrated closer to Rough Island between the early 1990s and 2023, according to OS maps. This increased the amount of intertidal area available for seagrass colonisation west of the Urr Water, which, at least in part, facilitated the development of a large bed below Glen Isle and Castle Hill.
The edge of the Z. noltii bed east of Urr Water contained isolated clumps, which had been separated from the main bed by strong currents, eroding the plants and their supporting clayish muddy sediments.

Figure 9. Eroding Zostera noltii bed edge near Urr Water, Rough Firth, 2023. Image © Max Carstairs.
Relationships with other biota
Bioturbation
Physical movements of sediments by the polychaete invertebrates Arenicola marina and Hediste diversicolor can restrict seagrass distribution (Hughes et al., 2000; Philippart, 1994; Philippart & Dijkema, 1995; Reise, 1985). Z. noltii and Z. marina share intertidal sedimentary habitat preferences with the burrowing polychaete A. marina (Figure 10). Research by Philippart (1994) indicated that seagrass beds are unable to expand into areas of mudflat with dense colonies of A. marina due to the amount of sediment reworking by the invertebrate. Conversely, large colonies of A. marina are unable to establish within seagrass beds, as the network of rhizomes likely impedes burrowing. Similarly, the polychaete cannot inhabit shallow muds overlying a firm clay layer, which precludes burrowing. Oncken et al. (2022) found that environmental stress tipped the balance in favour of A. marina, which exhibited more resilience to physical and chemical pressures. Declines of seagrass may facilitate colonisation by A. marina, where suitable sediments occur, after which re-establishment of seagrass may be inhibited.

Figure 10. Arenicola marina casts in recovering Zostera noltii bed, Stranraer, Loch Ryan. Image © Max Carstairs.
Delefosse & Kristensen (2012) reported that shallow burial of seagrass seeds by the polychaete Hediste diversicolor was likely beneficial to seed survival through concealment from potential consumers. Conversely, germination was delayed or failed where seeds accumulated at depth greater than 6 cm in A. marina burrows.
Bioturbation can also be undertaken by bivalves such as the common cockle Cerastoderma edule (Meysick et al., 2022).
Filter-feeding invertebrates
Filter-feeding invertebrates which form dense beds such as the European oyster (Ostrea edulis), blue mussel (Mytilus edulis), and common cockle (Cerastoderma edule), may influence seagrass populations. The inter-relationships between filter feeders and seagrass are complex and dynamic and differ between intertidal and subtidal sites (Meysick et al., 2022; Reusch & Williams, 1998). For example, reduced flows within dense subtidal seagrass beds limit opportunities for filter feeding while feeding opportunities may be enhanced within intertidal beds (Reusch & Williams, 1998). Larval settlement may increase around seagrass patches and bed edges due to collision with leaf blades. Once established, developed bivalves may inhibit further seagrass expansion, while conversely, dense seagrass rhizome mats may exclude burrowing filter feeders (Meysick et al., 2022; Reusch & Williams, 1998). Competition for space has been noted in the eastern pacific between Z. marina and the invasive mussel Musculista senhousia (Reusch & Williams, 1998).
Thelarvae of Ostrea edulis attach to hard substrates, especially other oysters, and the species historically engineered large-scale biogenic reefs (Thurstan et al., 2024; zu Ermgassen, 2025). Oysters stabilise sediments and build reefs within estuaries, creating shallow sills over which relatively fast currents develop (Jackson, 2001; Rothschild et al., 1994; Yates, 1913), enhancing structural heterogeneity and the diversity of flow conditions.
Oysters played an important historical role in cleansing coastal waters (Jackson, 2001; Newell, 1988; Perkins, 1973) by filtering suspended sediments and plankton from the water column and producing mucous-bound deposits on the sea floor (Haven & Morales-Alamo, 1966; Newell, 1988; Yonge 1960).
Historical accounts from Chesapeake Bay illustrate that large populations of the American oyster (Crassostrea virginica) and Z. marina coexisted, with oysters typically occupying areas of firmer ground and faster flows (Jackson, 2001; Yates, 1913). Mosaics of Mytilus edulis and Z. marina beds have also been recorded in Canada (Barrel & Grant, 2015). Changes in the balance of environmental factors may lead to distributional shifts. For example, Crassostrea virginica colonised areas where Z. marina had occurred in Chesapeake Bay prior to the 1930s wasting disease epidemic (Hopkins, 1957).
Studies by Newell (1988) indicate that the abundance of Crassostrea virginica in Chesapeake Bay before the 1870s facilitated filtration equivalent to all the water in the bay within one week. Following population collapse, filtration of the same volume of water would have taken 46 weeks. Therefore, removal of filter feeding invertebrates, especially oysters, is likely to significantly elevate turbidity levels in coastal waters.
Beds of native oyster (Ostrea edulis) were once commonly distributed around Scotland, with notable beds in the Solway Firth, Firth of Clyde off Kintyre, Firth of Forth, Firth of Tay, Moray Firth, and Cromarty Firth (Thurstan et al., 2024; UMBSM, 2007; Smout & Stewart 2012). Substantial populations also occurred along the west coast and around Orkney and Shetland (Thurstan et al., 2024; UMBSM, 2007). Exploitation is known to have occurred for centuries, and the collapse of beds due to overfishing occurred during the 19th century and in the early 20th century (UMBSM, 2007).
Blue mussels (Mytilus edulis) can tolerate a wide range of salinities. They inhabit the open coast and estuaries with accounts of large historical beds present in most Scottish estuaries. Mussels were extensively harvested from the intertidal and subtidal zones as the preferred bait for the whitefish handline fishery during the 18th, 19th and early 20th centuries. This led to significant population declines around Scotland including the serious depletion of extensive beds such as Montrose Basin, Dornoch Firth, Firth of Forth and Firth of Clyde (Fishery Board for Scotland, 1887; Scottish mussel and bait beds committee, 1889; Fenton, 1992; Coull, 1996; MBHS, 2004; Smout & Stewart, 2012). The issue of bait collecting diminished as the 20th century progressed and bottom trawling for whitefish steadily replaced handlining (Coull, 1996; Smout & Stewart, 2012). During this period, mussel beds were also reported to have been destroyed by pollution (MBHS, 2004; Scottish mussel and bait beds committee, 1889).
More recent declines in Scottish blue mussel populations have been recorded (Burrows et al., 2017, 2025). The reasons behind these declines are unclear and further research is ongoing though increased ocean acidification and warming may be influential.
Acidification of the marine environment is greatest in surface waters, with coastal areas and estuaries acidifying rapidly (Garner et al., 2022). Macrophytes have the potential to buffer acidification at the local scale through the uptake of carbon dioxide during photosynthesis, thereby ameliorating the effects of ocean acidification on calcifying organisms such as oysters (Garner et al., 2022). The association between seagrass and filter feeding bivalves such as blue mussels and oysters is therefore co-beneficial.
Epiphyte grazing by marine invertebrates
The mud snail Peringia ulvae can attain extremely high densities on soft intertidal sediments. P. ulvae grazes epiphytic algal growths on seagrasses, therefore high densities of the mollusc assist in bed maintenance. High densities of P. ulvae are characteristic of healthy to moderately-enriched intertidal mudflats and Z. noltii beds (Cardoso et al., 2005). Gastropod density may increase with increases in nutrient concentrations (Philippart, 1995), but the snail cannot tolerate high nutrient concentrations. P. ulvae is sensitive to relatively fast water currents, so increases in flow velocity may facilitate higher rates of epiphyte persistence in eutrophic waters (Schanz et al., 2002).
Subtidal Z. marina beds characteristically contain high densities of invertebrates, particularly mysid and amphipod crustacea, which graze on epiphytic algae. Reductions in grazer numbers may facilitate blooms in epiphytic algae (Andersson et al., 2009; Whalen et al., 2013). Populations of invertebrate grazers within seagrass beds may be reduced by increased predation rates, in a trophic cascading effect whereby densities of small fish increase in response to depletion of larger predators through overfishing (Williams & Heck, 2001). For example, the collapse of Swedish cod (Gadus morhua) stocks was associated with increased abundances of predatory small fishes and decapods, the loss of epiphyte grazers and decline of Z. marina (Duffy et al., 2014; Svedäng & Bardon, 2003).
Grazing and relationship between seagrass and wildfowl populations
Several species of wildfowl feed on seagrass in Scotland, especially wigeon and brent geese. Brent geese can feed to a depth of 40 cm, and as Z. marina can grow to over two metres in length, the water depth in which the geese can feed can be substantial. Both wigeon and brent geese can also take advantage of plants dislodged by other foraging species (Ganter, 2000; Ogilvie, 1992). Brent geese have been recorded feeding on floating Zostera and washed-up material (Ganter, 2000; M. Ogilvie, pers. comm.), and though this is a sub-optimal means of acquiring food (Ganter, 2000), it demonstrates that Zostera beds growing in relatively deep water can be exploited by wildfowl. Whooper swans have been observed upending to feed on Z. marina in Loch Indaal, and in doing so, they dislodged plants which were consumed by wigeon (Ogilvie, 1992).
Wigeon grazing on Zostera is common along the east coast of Scotland (Rae, 1979). However, Rae found no evidence that grazing in the Cromarty Firth and Tyne Estuary affected overwintering plant biomass, although she related that the scope of her studies was limited. Grazing by wildfowl can remove significant amounts of both above-ground and below-ground seagrass biomass (Jacobs et al., 1981; Madsen, 1988; Tubbs & Tubbs, 1983). However, grazing is not typically detrimental to seagrass beds (Charman, 1977; Ganter, 2000; Ranwell & Downing, 1959; Tubbs & Tubbs, 1982) unless beds decline in size below a critical threshold due to environmental stress, at which point the remaining plants may succumb to intensive grazing (Berry, 1939; den Hartog, 1994).
Grazing by geese has been shown to increase seagrass density by way of increasing growth vigour and disturbing surface sediments, providing openings for seed development and temporarily lowering the anoxia horizon (Nacken & Reise, 2000; Zipperle et al., 2009). Jacobs et al. (1981) considered that activities of overwintering wildfowl may increase sediment mobility, thereby regulating the build-up of sediments around seagrass plants.
Brent geese and wigeon have a strong association with Zostera, to the extent that changes in Zostera abundance can have marked effects on their distribution and population sizes (Atkinson-Willes & Matthews, 1960; Berry, 1939). Three sub-species of brent goose are currently recognised: dark-bellied brent goose (Branta bernicla bernicla); light-bellied brent goose (Branta bernicla hrota); and the black brant goose (Branta bernicla nigricans).
The overwhelming majority of brent geese in Scotland are the light-bellied form, with small numbers of the dark-bellied form also occurring (Forrester et al., 2007). Light-bellied brent geese have a much stronger association with Zostera than the dark-bellied form. Birds wintering on the east coast of Scotland are from the Svalbard breeding population, while those on the west coast arrive from the Eastern Canadian Arctic (Forrester et al., 2007). Branta bernicla hrota is hereafter referred to as brent goose or geese, unless otherwise stated.
Prior to the wasting epidemic, brent geese primarily fed on Zostera in their wintering grounds in Britain, Ireland and continental Europe before flying north to breed in the Arctic. The brent goose is the smallest European goose and therefore requires a high-energy source to build up sufficient body weight for winter survival, migration flights, and spring breeding (Ganter, 2000). Brent geese also have a high wing loading, which necessitates refuelling stop-overs during migrations between overwintering and breeding grounds (Ganter, 2000). Zostera has a relatively high nutrient content (McRoy, 1970; Ranwell & Downing, 1959) with easily accessible rhizomes, and as beds occurred regularly along European coasts prior to the wasting disease epidemic, they were fundamentally important to brent goose populations.
The brent goose population of Northern Europe (sub-species Branta bernicla bernicla and Branta bernicla hrota) suffered a dramatic population crash, estimated between 75-90%, following the 1930s Zostera wasting disease epidemic (Ogilvie & Mathews, 1969). Concurrent post-wasting disease declines of 90% were estimated in the western Atlantic Branta bernicla hrota population (Cottam et al., 1944). The brent goose population crash was largely attributed to loss of Zostera beds (Butcher, 1941; Ogilvie & Mathews, 1969). However, other workers have connected overshooting with declines since the 19th century. This element was apparently glossed over during the post-wasting-disease International Wildfowl Inquiry in order to appease the wildfowling contingent of the International Committee for Bird Preservation (Tubbs, 1996). Overshooting was undoubtedly responsible for significant declines in the early in the 20th century, but this does not diminish the importance of Zostera in terms of supporting healthy brent goose populations.
Similar declines occurred in the number of wigeon in Scotland after the 1930s epidemic (Atkinson-Willes & Matthews, 1960), though these birds can overwinter in fresh as well as coastal waters. Despite the losses, Zostera, where available, remains the preferred coastal food resource for wigeon and brent geese (Ganter, 2000; Murray et al., 2019).
The association between wildfowl and seagrass weakened somewhat following the wasting disease outbreak (Cottam et al., 1944; Cottam & Munro, 1954). Wigeon and brent geese adapted to an extent to less nutritious alternatives such as green algae, short saltmarsh plants, and inland crops and pastures (Ranwell & Downing, 1959). Records of abundant coastal populations of these two bird species in the early 1900s are therefore indicative of abundant historical seagrass populations.
Displacement by invasive species
The invasive brown algae Sargassum muticum was first recorded in Scotland in Loch Ryan in 2004 (Harries et al., 2007a; Davison, 2009). The seaweed is now well established within intertidal areas with mixed sediments in the Firth of Clyde (M. Carstairs, pers. obs.) and has recently spread as far north as Orkney (Want et al., 2023). Sargassum muticum inhabits shallow, sheltered pools and channels, where it typically attaches to hard substrates via a holdfast. Due to the large size of the fronds, which can reach up to 8 m (den Hartog, 1997), it can extend over areas of finer sediments which lack suitable substrates for attachment, thereby shading out seagrass. den Hartog (1997) observed the ability of Sargassum muticum to supress the recovery of intertidal Z. marina beds over extensive areas of the Brittany coast at Roscoff. The seaweed occurs in intertidal pools alongside Z. marina on mixed substrate at Dunure, Ayrshire (Carstairs, 2024; Figure 11) and in subtidal beds at Kildonan, Arran (Thomas, 2016). Tweedley et al. (2008) provided evidence that the species is also able to colonise dense seagrass beds in soft sediments.

Figure 11. Sargassum muticum growing alongside Zostera marina in rockpools at Dunure, Ayrshire. Image © Max Carstairs.
Click for a full description
The protruding rocks support a dense population of barnacles as well as a variety of molluscs including dog whelk and common limpet with sparse patches of stunted fucoid seaweeds.
The invasive kelp Undaria pinnatifida, a native of east Asia, was first recorded in the UK along the south coast of England in 1994 and has since spread as far north as Northern Ireland and the Firth of Forth (Bunker et al., 2017). The potential ramifications of this species for native flora and fauna, including seagrass beds, are not currently understood. The species is tolerant of low salinities and can proliferate within estuaries (Farrell & Fletcher, 2000), although it may pose a greater threat to subtidal than intertidal seagrasses.
The invasive hybrid cord-grass Sporobolus anglicus (formerly Spartina anglica) is a new species created when Spartina alterniflora was accidentally introduced from North America to the south of England in the late 19th century, most likely via ship ballast water (McCorry et al., 2003; Raybould et al., 1991). This species hybridised with the native Spartina maritima to produce a sterile hybrid, Spartina townsendii. A new fertile species, Spartina anglica, was created by 1872 when chromosome doubling occurred in Spartina townsendii (Cope & Gray, 2009). Given these taxonomic complexities, Spartina anglica now Sporobolus anglicus was not recognised as a new species until 1968.
Sporobolus anglicus favours deep, water-logged mud and can grow vigorously across intertidal mudflats ( Figure 12). The planting of this species was encouraged across northern Europe from the 1920s onwards to assist with intertidal mud stabilisation and reclamation, due to its potential for rapid growth (Cabot, 1999; McCorry et al., 2003). Encroachment has occurred over large intertidal areas (Cope & Gray, 2009) at the expense of native species including seagrass populations in Ireland (Cabot, 1999; Madden et al., 1993; McCorry et al., 2003).

Figure 12. Sporobolus anglicus growing in Zostera bed, Airds Bay, Solway Firth. Image © Max Carstairs.
A rapid expansion of S. anglicus in Great Britain, covering over 12,000 ha, had occurred by the mid-1960s (Hubbard & Stebbings, 1967). Following an invasive phase lasting many decades, several populations of S. anglicus have undergone substantial declines once they have altered soil conditions and drainage patterns to the extent that they have become detrimental to the species (Grayand & Benham, 1990). The effect on seagrass populations appears not to have been studied in detail, hampering assessment of the degree of risk posed by new expansions of S. anglicus. Similarly, information is lacking regarding seagrass recovery rates following large-scale declines of S. anglicus.
Threats to Seagrass Populations
Land use
Major changes in land use have significant effects on coastal hydrology and sediment dynamics. These changes therefore pose a threat to seagrass beds. Following the disappearance of the last ice sheet, forest and peatland developed into climax communities that dominated the Scottish landscape (Edwards & Whittington, 2003; Porley & Hodgetts, 2009). Natural forms of these habitats are typically rich in structural heterogeneity, to which a diversity of vascular plants and bryophytes contribute, facilitating greater water retention relative to modified habitats.
Changes in the hydrology of the Scottish landscape likely began when forests were cleared to provide farmland. Peatlands, which accumulate and retain great amounts of water, were later subjected to a range of pressures, including harvesting for fuel, drainage, and conversion to farmland and forestry.
Commercial forestry, which progressed extensively from 1940 to the present time (Mackey et al., 1998), is associated with increased rates of suspended sediment run-off, nutrient enrichment, and acidification (Gee & Stoner, 1988; Ratcliffe, 2007; Stevens et al., 1988). Hydrological regimes are significantly altered, with reduced absorbency and storage capacity of soils and vegetation, leading to flashier watercourses which rise and fall more rapidly (Ratcliffe, 2007). Changes in river flows led to corresponding changes in coastal water parameters, including current strength, sediment dynamics, and salinity. The most detrimental of these changes to seagrass beds would be from any resulting eutrophication and elevated levels of sediments held in suspension in coastal seas.
Forestry practices have improved in the UK in recent decades, with a view to reducing soil erosion, surface water run-off, and associated suspended sediment concentrations (Forest Research, 2023). However, current UK Government policy is to rapidly increase the rate of tree planting as a means of ameliorating climate change (Environment, Food and Rural Affairs Committee, 2022).
Farming practices in Scotland have intensified steadily since the 1940s and are associated with increased nutrient inputs to watercourses and alteration of their hydrological regimes. Conversion of water-retaining wetland habitats such as raised bog, blanket bog, acid flushes, wet heath, and fens into grassland has been achieved via a variety of means, including the installation of surface and subsurface drainage systems. These changes have led to larger hydrological peaks and erosional forces within riverine systems (Blanc et al., 2012).
Water quality issues are associated with run-off from intensively fertilised “improved” grasslands, as well as soil exposure and erosion from rotational ploughing, tilling, and reseeding of improved grasslands. Livestock fouling in watercourses and poaching of banks can also lead to significant sediment inputs into watercourses. Arable land, as well as being ploughed, may also be subject to herbicide and pesticide applications.
Many areas of the coastal lowlands of eastern Scotland are designated as Nitrate Vulnerable Zones (NVZ). In addition, two NVZs occur along the coastal plain of the Solway Firth with one along the eastern banks of the River Nith below Dumfries and a second above Luce Bay. Coastal waters receiving hydrological inputs from NVZs are potentially vulnerable to eutrophication.
Land reclamation and habitat channelisation
Land reclamation has occurred extensively within large Scottish estuaries. Approximately 50% of the intertidal area of the Firth of Forth has been converted to farmland or developed (McLusky, 1989). This has led to a significant decrease in saltmarsh and almost certainly seagrass beds, although no direct evidence of the latter was found. Seagrass populations prosper in shallow estuarine habitats, particularly in sheltered embayments (Rae, 1979; Carstairs, 2024; Rayer, 2024; Rodgers, 2024) which are easier to reclaim.
Physical changes to the structure of a flowing waterbody will alter its hydrological regime (Chu et al., 2022; de Jong et al., 2014; Zhang et al., 2021). The effects of estuarine reclamation vary depending on specific conditions, but it can generally lead to a narrowing of estuaries, restricting the lateral areas where water can disperse. This likely increases average current speeds and wave strengths, particularly during periods of high flow. When reclamation removed the vast majority of intertidal habitats in the Tees, Northern England, all that remained was a narrow channel flowing between the confines of straightened shorelines. This process is similar to that which occurs in lowland rivers due to floodplain development and construction of flood defence structures. These processes convert structurally diverse rivers into simple trapezoidal channels, which further contribute to the flashy flows within catchments (Purseglove, 1989). Gilvear (1994) stated that most of the flood embankment and channelisation schemes in Scotland had not taken into account the effects on downstream hydrology. It is likely that historical land reclamation and channel modifications have had a significant effect on Scottish seagrass populations, although before and after data is unfortunately lacking.
Causeways and dams
Causeways are solid structures, typically with occasional gaps, usually located between land masses (e.g. inter-island or island to mainland connections). These structures can have significant ecological effects on a large geographic scale relative to their immediate area (Vilks et al., 1975; Sutherland et al., 2013). The habitat in the immediate vicinity of causeways can change entirely, transforming from a single, often relatively fast-flowing sound into two poorly connected or completely disconnected embayments. The reduction in throughflow can cause sediment accumulation in embayments adjacent to the new structure. Water masses redirected from their natural topographical course may generate twisting currents with associated scouring action (Hugget, 1996; Waldichuk, 1985).
The environmental impacts of estuarine barrages and river dams are similar to those of causeways (Hugget, 1996). These structures require a means of regulating flows, which are bidirectional within estuaries (D’Alpaos & Defina, 1996). Disruption of the normal salinity regime is therefore possible (Evans & Rogers, 1996). Oxygen levels can also shift from those of a frequently flushed, oxygenated system to a more stagnant, ponded environment, increasing the potential for the development of algal blooms (Hugget, 1996).
Waste water and diffuse pollution
Assessing historical water quality status and trends for the UK is hampered by the lack of routine monitoring data prior to 1980 (Whelan et al., 2022). Industrial pollutants and crude sewage catastrophically affected many Scottish rivers from the 19th century until recent times (Hammerton, 1994). This was highlighted by the absence of fish for long periods from stretches of the River Clyde (Henderson & Hamilton, 1986). Substantial recovery followed improvements to the Scottish wastewater infrastructure in the latter half of the 20th century (Hammerton, 1994; Marsden & Mackay, 2001).
Nutrient and pesticide concentrations increased in UK rivers after farming intensification during the 1940s (Mackey et al., 1998; Whelan et al., 2022). However, concentrations of ammonia and phosphate decreased while nitrate stabilised during the 1990s, following implementation of EU directives, including the Nitrate Directive (Whelan et al., 2022). Run-off from farmland remains the largest source of organic nutrients entering UK waterbodies (The Rivers Trust, 2024).
Eutrophication of coastal waters is primarily driven by nitrogen enrichment, though phosphorus can also play a role in estuaries (Environment Agency, 2019). Sources of nutrients include wastewater discharges and diffuse runoff from agricultural land treated with substances such as slurry and artificial fertilisers. Nutrients can enter coastal waters via rivers, direct discharges, and via groundwater (Short et al., 1996; Stigter, 2011) and can affect seagrass both directly and indirectly. Mesocosm experiments by Burkholder et al. (2007) demonstrated that low levels of chronic nitrate enrichment were directly lethal to Z. marina.
Millier et al. (2010) stated that:
“it is generally accepted that the prevalent source of P[hosphorus] to the majority of UK lowland rivers is STW [Sewage Treatment Works] wastewaters and that this type of discharge has greatest potential to cause eutrophication, being … primarily discharged in the most bioavailable form.”
Nutrient enrichment also elevates levels of bacteria, phytoplankton, and epiphytes, which intercept light, reducing the amount available to seagrass for photosynthesis (Burkholder et al., 1992, 2007). This reduces the depth at which seagrass can grow. Therefore, subtidal populations of Z. marina are more at risk from this form of pollution than the other Scottish taxa. Eutrophication also facilitates the development of floating filamentous algal mats (Raffaelli et al., 1989; See Figure 13), which can shade seagrass and cause direct mortality by settling on top of beds for prolonged periods of time (den Hartog, 1994; Carstairs, 2024; van den Hoek et al., 1983). Severe nutrient enrichment can also lead to anoxia of the lower water layers (Raffaelli & Hawkins, 1996) and sediments.
A recent symptom of deteriorating environmental conditions (including nutrient loading within estuaries) is the expansion of Ruppia beds at the expense of Zostera populations (Cho et al., 2011; Hensel et al., 2023). This genus is more tolerant of environmental fluctuations and stress than Zostera and can rapidly proliferate via sexual reproduction (Cho et al., 2011; Moore et al., 2014; Hensel et al., 2023). Monitoring of this genus across Scotland would provide a useful additional perspective on ecosystem health.

Figure 13. Green algal mats in Zostera bed, Solway Firth. Image © Max Carstairs
Aquaculture
Salmon farming in Scotland began in the 1970s. Farms are mainly distributed along the west coast, Hebrides and Northern Isles with sheltered locations such as sea lochs historically favoured. More recently, there has been a progressive move for site expansion to more dispersive sites. Concerns have been expressed regarding the environmental impacts of salmon farms given the industry’s ambition for expansion (Scotland Food and Drink, 2016). However, potential impacts on seagrass populations could be reduced by siting of fish farms in locations with strong, repeating, water currents rather than in sheltered bays and sea lochs.
Fish farms have the potential to adversely affect seagrass populations, particularly via nutrient enrichment and pesticide use. Open pen fish farms release excreta, uneaten food, fish medicines and other chemicals into the environment. As point sources of pollution, these can have deleterious effects on the surrounding waters including increased siltation/sedimentation and nutrient enrichment. SEPA, under a site’s Controlled Activities Regulations (CAR) licence, regulate the discharge into the marine environment.
Finfish aquaculture is now the primary source of direct nutrients to the OSPAR regions, contributing approximately as much nitrogen as industry and wastewater treatment combined (OSPAR, 2023). Navarro et al. (2008) detected significantly higher concentrations of ammonium and dissolved organic nitrogen concentrations and heterotrophic microbial abundance at water sampling stations near a fish farm in Loch Fyne though changes in chlorophyll levels were not observed.
There have been no Scottish studies undertaken on the impacts of fish farms on seagrass, however Cullain et al. (2018) found impacts on Z. marina beds concentrated within 1 km of a fish farm in Port Mouton Bay, Canada. However, areas of impact are related to local hydrodynamics and the production levels of the farm, with the fish farm used in the Cullain et al. (2018) being a relatively small operation, with annual production levels estimated at 760 t. For context, current applications for fish farms in Scotland are often for annual production levels of 3000 t. Tett et al. (2018) related that nutrient enrichment in Scottish waters was likely to be more pronounced in sheltered areas with gentle currents and especially in sea lochs with a sill at the mouth due to limited water exchange. Fish farms have been implicated in the disappearance of seagrass beds in some Scottish waters (James, 2004) though lack of data precluded definitive conclusions.
Potential impacts on water quality from an expanding industry are highlighted in the Marine Scotland Assessment 2020 (Scottish Government, 2020a) which found that for both total nitrogen and total phosphorus, statistically significant increasing trends for the Orkney Islands and Outer Hebrides SMRs between 2007 and 2017 were likely a result of increases in loads from marine pen fish farms due to increasing farmed fish biomass over that time period.
There is currently a moratorium against open pen fish farms on the north and east coasts of Scotland. This prevents salmon farms being developed in these regions to protect migration routes as recognised in Nation Planning Framework 4 Policy 32a and Section 3.9 of the Wild Salmon Strategy: Implementation Plan 2023–2028 (Scottish Government, 2023).
Studies on current and future impacts are required to support the National Marine Plan objectives regarding protecting PMFs. Future research should include an assessment of how pesticides used in the aquaculture industry may potentially impact grazers of seagrass epiphytes, and invertebrates that could be involved in seagrass pollination. The depth of the euphotic zone should also be monitored for change.
Other pollutants from industry, agricultural and urban environments
Oil pollution
The presence of seagrass confers natural protection to coastal habitats and benthic fauna from oil pollution. Jacobs (1980) found that the thick rhizome mat of Z. marina prevented oil from the “Amoco Cadiz” spill from penetrating sediments and therefore minimised direct contact with benthic organisms. Oil pollution has the potential to affect seagrass by preventing photosynthesis and causing tissue necrosis (Jacobs, 1980). Experimental studies and post-spill monitoring indicate that seagrass is fairly resistant to oil pollution incidents though oil may halt or reduce growth (Fonseca et al., 2017; Howard et al., 1989; Jacobs, 1980; Rae, 1979). In contrast, the chemical dispersants used to break down and disperse spilled oil have a greater adverse effect on seagrass (Fonseca et al., 2017; Howard et al., 1989; Jacobs, 1980; Rae, 1979). Therefore, consideration should be given to their use where seagrass beds are present (Rae, 1979; Scarlett et al., 2005). Howard et al. (1989) recommended that where oil settles on seagrass beds, it should be left to disperse naturally through tidal action rather than using chemical dispersants.
Heavy metals
Heavy metals can enter coastal waters from a variety of sources and have a range of adverse effects on seagrass (Qiao et al., 2022). Brackup & Capone (1985) found that mercury, nickel and lead as well as the organic substances aldicarb, naphthalene, pentachlorophenol, and Kepone reduced nitrogen fixation in seagrass roots. Qiao et al. (2022) found that copper and cadmium adversely affected Z. marina though short-term effects below EC50 (half maximal effective concentration) were reversible.
Due to their high stability and ability to penetrate seagrass tissues, heavy metals can bioaccumulate in seagrass beds (Ryumina et al., 2024). As seagrass plants die and decay, accumulated metals are transferred to the sediments, leading to higher metal concentrations than in surrounding bare areas (Ryumina et al., 2024).
The levels of heavy metals occurring in European coastal waters are generally lower than those found experimentally to produce adverse phytotoxic effects (Lyngby & Brix, 1984; De Casabianca et al., 2004). However, long-term accumulation of metals in the sediments, combined with other stressors may be detrimental to seagrass beds. The ability of seagrass beds to accumulate heavy metals may improve water quality in nearby areas, and their use in coastal remediation has been suggested (Qiao et al., 2022). Preventing heavy metals from entering coastal environments is preferable wherever feasible, given the potential for bioaccumulation of heavy metals in wildfowl feeding on seagrass.
Microplastics
Large sheets of plastic and moderately sized plastic litter wash onto beaches, though they typically accumulate higher on the shore than the Zostera zone i.e. above the average groundwater line (M. Carstairs, pers. obs.). Microplastics (<5 mm), due to their small particle size and high concentrations in Scottish waters, may settle in significant quantities within both subtidal (Jones et al., 2020) and intertidal areas (Blumenröder et al., 2017). Blumenröder et al. (2017) found that concentrations of microplastics in Orcadian waters were similar to those from the Clyde and Firth of Forth, which are heavily industrialised areas. The transportation of microplastics within coastal currents likely accounts for their widespread distribution (Jones et al., 2020).
The degree of microplastic settling is enhanced by seagrass, which intercept particles in the water column causing them to settle within the seagrass bed (Jones et al., 2020). Seagrass beds can act as a microplastic sink, with higher concentrations recorded in the benthic sediments of seagrass beds compared to surrounding areas (Jones et al., 2020). Microplastic intercepted by seagrass can become attached to seagrass leaf biofilms, and elevated concentrations have been recorded within the tissues of epiphytic grazers (Jones et al., 2020).
The study of the effects of microplastics on seagrass is at an early stage. Molin et al. (2023) observed a slight decrease in photosynthetic activity and respiration rates of seagrass in relation to short-term (14 days) microplastic exposure. Further studies are required to assess the degree to which light availability and photosynthesis may be affected by microplastics, as well as their potential impact on populations of epiphyte grazers.
Biocides - herbicides and pesticides
Glyphosate is the most widely used herbicide globally and in the UK. It is used to control weeds in arable land and urban areas. A recent study of seasonal concentrations in European rivers by Schwientek et al. (2024) indicated that herbicides are not the predominant source of glyphosate in the water environment. The authors linked patterns of occurrence to waste waters and suggested a potential source: the formation of glyphosate from aminopolyphosphonates present in laundry detergents. Further research is currently being conducted on this topic (Schwientek et al., 2024).
Nielsen & Dahllöf (2007) found that the growth rates of phytoplankton and Z. marina were affected by the herbicides glyphosate, bentazone, and MCPA (2-methyl-4-chlorophenoxyacetic acid). Delistraty & Hershner (1984) found that the herbicide Atrazine inhibited Z. marina growth rates while Hershner et al. (1983) linked it to declines of Z. marina in Chesapeake Bay.
Pesticides are applied to arable land in Scotland and can reach coastal waters via run-off from land and transport through river systems. Pesticides are also used to control sea lice numbers in salmon farms. These specifically target crustacea but can also have an impact on non-target species (Hall-Spencer & Bamber, 2007; Page & Burridge, 2014). Recent studies indicate that seagrass pollination may be enhanced by crustacea and that loss of small grazing crustacea facilitates blooms of algal epiphytes (Andersson et al.,2009; van Tussenbroek et al., 2012, 2016)
A number of studies have highlighted causal pathways and adverse impacts of insecticides on marine invertebrates (Urbina et al., 2019; Burridge et al., 2010; Page & Burridge, 2014; Bechmann et al., 2019; Hall-Spencer & Bamber, 2007). The potential role of subsurface invertebrate pollinators has not been investigated for seagrass taxa inhabiting Scottish waters, therefore the possible impacts of pesticides on Scottish seagrass reproduction and gene flow cannot currently be assessed.
Sources of physical disturbance and damage
Demersal mobile fishing gear
Mobile, bottom-contacting fishing gear is used to exploit demersal finfish and shellfish in Scottish waters. Toothed dredges are used to target queen scallop Aequipecten opercularis which occurs from the intertidal zone down to 100 m, and great scallop Pecten maximus which occurs from around 10 m to depths of 100-200 m. Trawls are used to harvest demersal finfish and Norway lobster Nephrops norvegicus which typically inhabits depths of 20-200 m, though it can occur in shallower waters in Scottish sea lochs (Hill & Sabatini, 2008).
VMS pings from scallop boats >12 m in length in the Firth of Clyde indicated that scallop dredging is concentrated along the coast (McIntyre et al., 2012). Eigaard et al. (2017) studied Scottish logbook statistics and Vessel Monitoring System (VMS) data from 2010–2012 and found that fishing with mobile bottom-contacting gear occurred at a variety of depths, including depths in which seagrass beds can proliferate.
These operations pose a direct potential threat to seagrass beds through damage to plants and/or uprooting of plants. Indirect threats include sediment and nutrient re-suspension caused by towed gear and damage to the seabed structure, which may add to pressures on seagrass populations.
A number of published studies from the western North Atlantic demonstrate adverse impacts of scallop dredging on Z. marina beds in terms of reduced plant density and biomass (Thayer & Stewart, 1974; Fonseca et al., 1984; Bishop et al., 2005; Fonseca & Uhrin, 2009).
Formal scientific studies on the impact of scallop dredging on seagrass beds have not occurred in Scottish waters. Scallop dredging has been reported to have historically damaged seagrass beds in Loch Indaal, Islay, leaving lines of bare sediments in the beds where gear had been towed (Waring, 2024). Similarly, there has been an anecdotal report that scallop dredging caused extensive damage to a seagrass bed in Skipness Bay, Kintyre (Wood, 2006). The Community of Arran Seabed Trust also reported that the extent of seagrass populations around Arran has been negatively impacted by abrasive fishing gear (COAST, 2012a).
Intertidal shellfish harvesting
Mechanical harvesting of common mussel (Mytilus edulis) and common cockle (Cerastoderma edule) was found to have damaged intertidal seagrass beds in the Wadden Sea. This was cited as a reason for the lack of long-term recovery following catastrophic historical declines during the 1930s (de Jonge & de Jong, 1992). Suction dredging of cockles in the Solway Firth also resulted in damage to intertidal seagrass beds (Perkins, 1988).
Other sources of physical disturbance
Physical disturbance and damage to seagrass beds may also be caused by a variety of other means including coastal defence works, dredging of ports and channels, boat propellors, mooring and anchor chain scour (Figure 14), bait digging and trampling (Carstairs, 2024; Egerton 2011; Erftemeijer & Lewis, 2006).

Figure 14. Anchor chain damage to intertidal Zostera marina, Solway Firth. Image © Max Carstairs.
Climate change
Climate change threatens seagrass through increased water temperatures changes in precipitation dynamics and sea level rise. Scotland lies centrally within the biogeographic zone of Z. marina and is at the very northern edge of the range of Z. noltii with both species extending southwards to North Africa (Green & Short, 2003). Both species could therefore be anticipated to remain present around Scotland under various climate change scenarios though these may potentially involve a degree of population decline and readjustment in relation to local and regional adaptations. Reproductive changes may also occur, for example, temperature increases may lead to earlier flowering and germination dates and may increase capacity for sexual reproduction in northern latitudes (Blok et al., 2018).
Seagrass may also be indirectly affected via temperature increases which reduce oxygen concentrations and increase the rate of biological reactions, potentially creating anaerobic conditions in the sediment and lower water layers. Phytoplankton concentrations and the frequency of algal blooms may also increase as a result of higher temperatures (Gobler, 2020). This may be further exacerbated by a greater frequency of storms and greater rainfall intensity with associated increased run-off rates and nutrient inputs from terrestrial habitats in addition to elevated levels of resuspended nutrients within the water column. There is some evidence to suggest that the photosynthetic apparatus of seagrass plants may be damaged during periods of hypoxia resulting in altered rates of photosynthesis and respiration which can persist following the return of normal oxygen levels (Holmer & Bondgaard, 2001; Rasmusson et al., 2020; Che et al., 2023). All of these effects are potentially detrimental to seagrass populations and may increase their susceptibility to outbreaks of wasting disease (Groner et al., 2021; Schenck et al., 2023).
Seagrass beds can be very sensitive to storm damage, although this depends on the location of beds, with those in sheltered bays less vulnerable. Storms can cause mechanical damage to leaf blades, and in some cases remove whole plants (Costa, 1988; den Hartog 1987; Orth & Moore 1983). Sediment suspension and deposition can preclude photosynthesis and cause extensive seagrass mortalities (Blois, et al., 1961). Storminess and associated sediment reworking in coastal habitats is projected to increase as a result of climate change (Burden et al., 2020).
Predicted rises in sea level (McKirdy, 2007; Met Office, 2018) due to melting of snow and ice are likely to become a major threat to Scottish seagrass populations. The extent of this rise will depend on global glacial melting rates and will vary geographically across Scotland, influenced by the former thickness of glacial ice. The degree of crust rebound is now modest and will not offset rising sea levels to the degree previously considered by the scientific community (Rennie & Hansom, 2011).
The highest rise in levels is projected to occur in Shetland, where McKirdy (2007) estimated an increase of 60 cm by the year 2100. In Orkney and the Western Isles, sea level rise is expected to be 20 cm less. The west coast of Scotland is predicted to experience an increase of just over half that of Shetland (around 30 cm), with even lower rises anticipated in central Scotland, including the Firth of Forth and Moray Firth.
Rising sea levels will reduce levels of light available to existing beds, potentially causing losses in deeper areas. This may be partially offset by the inland migration of coastline, creating potential new habitat for seagrass. Unique geomorphological features such as coastal lagoons may experience change or extirpation (Angus et al., 2011). Landward migration of the high tide line will be severely restricted where moderately steep rock strata occur or where coastal infrastructure including sea defences are present. For example, the majority of sheltered estuarine embayments containing seagrass along the Scottish coastline of the Solway Firth are fringed by rock with low lying land limited to the narrow floodplains of incoming freshwater streams and rivers. In addition, glacially derived sediment sources for beach formation and replenishment have already been exhausted in places such as Shetland, and sea level rise is projected at a rate that will far out-pace the hydro-morphological development of stable marine habitats (Morris, 2010).
Data Sources (Availability, Quality and Comparability)
Data gathering
The following institutions and groups were contacted for data on seagrass beds:
- All Scottish universities with a Marine Biology department
- Scottish Wildlife Trust (SWT)
- Scottish Environment Protection Agency (SEPA)
- Royal Society for the Protection of Birds (RSPB)
- Wildfowl and Wetlands Trust (WWT)
- Seagrass conservation organisations
- Independent seagrass consultants
Assessment of national databases
The assessment in this report utilises two databases which are considered to contain high quality data. These data sets primarily contain data collected by professional scientists with data quality checked prior to acceptance.
The Botanical Society of Britain and Ireland’s distributional database (BSBI DDb) contains data collected primarily by competent botanists along with submissions made by developing botanists and also members of the public. All records are submitted to vice-county recorders and are then assessed for plausibility prior to being entered into the database. Questionable submissions and/or records of rarities are typically followed up with either a request for a specimen or a field visit. Specimens of particularly difficult taxa can also be sent to nationally recognised referees for verification.
The Geodatabase of Marine features adjacent to Scotland (GeMS) database contains data primarily collected by professional marine biologists. In addition, data may be provided by experienced volunteers and community groups. Data quality checking procedures are similar to those of the BSBI.
Several online records from the herbarium of the Royal Botanical Gardens Edinburgh (RBGE) are included in the report along with their unique Herbarium Specimen Codes. The original notes accompanying herbarium specimens in many cases did not state if the material was found washed up or taken from a seagrass bed. Other herbarium sheets gave fairly detailed accounts.
Quality assessment of early seagrass bed maps
To assess seagrass population changes in detail at specific locations, an attempt was made to find the earliest historical maps for sites and compare these with subsequent surveys. Early mapping was undertaken using a variety of techniques with the exact method often undocumented. The quality and limitations of these older maps in relation to the current project aims are assessed below.
Butcher, 1941 (International Wildfowl Inquiry)
A seagrass distribution map for Great Britain was published in 1941 (Butcher, 1941). The methodology used to generate this map is unclear as the author stated in the text that the map represents the “maximum distribution” and then expanded “that is all the areas are shaded where any form of the plant has been found at any time.” The map legend states “data for England based on county floras and personal observation” and “data for Scotland are incomplete and are based on topographical botany”. Butcher (1941) also related that Zostera was not found on “wave-swept, shingly and rocky shores on the west”. These statements indicate that the Scottish Zostera distributions were substantially derived from published geological or admiralty maps containing soft sediment distributions. Butcher’s map of seagrass distribution extends across large areas of the Solway Firth for which no other historical or contemporary records are known to the current authors. Butcher’s map was therefore not used in the current study.
Ackroyd, Montrose Basin, 1972
The report contains mapped distributions of intertidal Z. noltii and Z. marina (Ackroyd, 1972). Unfortunately, the original report which evidently contained colour maps could not be located and only a black and white scan was available which was considered be a second or third generation copy. The mapped distribution of Z. noltii could not be distinguished from background artifacts introduced during the copying process though a description of the distribution is present in the text. The key map for intertidal Z. marina states “scale inaccurate”. The seagrass distribution maps were therefore not reproduced for the current project.
Nature Conservancy Council, Moray Firth studies, 1972 & 1978
A document entitled “A Prospectus for Nature Conservation within the Moray Firth” was produced by the Nature Conservancy (NC, 1972) with a revised edition published in 1978 (NCC, 1978). A survey methodology was not given, nor was a date for the surveys. The surveyor(s) are not named but it is thought that the work for the first edition was predominantly undertaken by Andrew Currie of the NCC as Fox et al. (1986) state that he undertook extensive surveys of all vegetation types in the area between 1968 and 1974. This gives a potential date range for the maps in the first edition of 1968-1972.
The 1972 prospectus contains maps of Zostera for the Cromarty Firth and Munlochy Bay with Dornoch Firth and Beauly Firth (upper section only) additionally included in the 1978 prospectus. The maps appear of suitably high quality for inclusion in the current project.
Mapping to species level is presented in the second edition published in 1978 (with the exception of Munlochy Bay which is reproduced from 1972). Unfortunately, not all of the reported seagrass beds are mapped, with additional bed locations, presumably of relatively insignificant populations, described in the text.
University of Aberdeen, Moray Firth, 1970-1981
A series of ecological studies were carried out for the Highlands and Islands Development Board and Cromarty Petroleum during the 1970s and early 1980s. A seagrass map is presented in the 1978 report (University of Aberdeen, 1978). The map is undated though surveying is likely to have been undertaken in the late 1970s. The mapped seagrass distributions are very similar to those in the 1978 Nature Conservancy Council prospectus and as the map was obtained at the end of the current project it is not reproduced here.
Minto, Clyde Estuary, BSc, 1974
A BSc study of the relationships between invertebrates and seagrass was undertaken on the Clyde by Minto in 1974 (Minto, 1974: Cited in Cleator, 1993). Unfortunately, the thesis was not available for this project despite attempts to track it down through a variety of avenues. However, details of the survey given in Cleator (1993) indicate that only Z. noltii was recorded and that bed mapping was not undertaken. The table of locations in Cleator (1993) states “precise location not available” in relation to the Minto study.
Rae, Nigg Bay, PhD, 1979
Zostera beds were mapped in detail to species level showing bed extent and denser areas of seagrass distribution (Rae, 1979). Field work was conducted between 1975-1978 though no precise dates are given for the seagrass distribution maps. The maps appear of suitably high quality for inclusion in the current project.
Sphere Environmental Consultants, Nigg Bay, 1981
Seagrass surveys were caried out at Nigg Bay by Sphere Environmental Consultants in 1980 and 1981. The report for 1980 was not available for the current project therefore the survey methodology is not understood. Maps of 1980 and 1981 seagrass distribution appear in the 1981 report. These are comprised of 15 A4 (1:8,000) maps on transparent sheets and 3 A0 (1:10,000) maps on various paper types. The survey methodology for 1981 was primarily based on aerial photographs with extensive ground truthing.
The 1981 map is “a preliminary assessment of the position of the boundaries and was made by free hand scaling onto a 1:10,000 map” (Sphere Environmental Consultants, 1981). The aerial photos were printed at 1:8,000 scale, traced and subsequently transferred freehand to a 1:10,000 basemap. The transfer to a different scale by eye has introduced at least a moderate degree of error. This is noticeable when comparing the tracings of seagrass beds and the coastline made from aerial photographs to the seagrass beds mapped on the 1:10,000 basemaps.
The report is described as a preliminary assessment and states that the data will be used in preparation of a colour 1:10,000 map, although this is not presented in the report. (Sphere Environmental Consultants, 1981). Despite issues of scaling and georeferencing, the Sphere maps provide an indication of the extent and position of seagrass beds in 1981. However, the maps have not been digitised in the current project as derived calculations of bed area are likely to be inaccurate.
A map of seagrass distribution in Nigg Bay purported to be from the work undertaken by Sphere in 1981 is presented in Trendall et al. (2011). This is however incorrect and the source of the map is in fact the 1978 NCC prospectus. A map of the 1978 distribution is present in Sphere (1981) though Sphere clearly state that it is from the NCC prospectus. Unfortunately, Sphere (1981) stated the map was from approximately 1970, although the map exactly matches that in the 1978 NCC prospectus and there is therefore no doubt regarding its origin.
McLusky and Roddie, Montrose Basin, 1982
This study presented maps of seagrass density (Domin Scale) within quadrats stationed at intervals along transects and was the first survey to cover the entire basin (McLusky & Roddie, 1983).
The authors expressed difficultly with the identification of seagrass species. Their identifications were considered erroneous given the species distributions presented in the maps therefore the data has been consolidated into a combined Zostera map for the current study.
The original presence/absence data was georeferenced and displayed over the 2017 SEPA dataset allowing a visual spatial comparison of seagrass population change.
Fox, Yost and Gilbert, Northern Firths, 1986
Surveys of seagrass distribution were undertaken in the Dornoch Firth, Cromarty Firth, Beauly Firth and Moray Firth. Abundance of Zostera was mapped within 250 m x 250 m squares. The data was semi-quantitative with presence/absence and density data recorded using the following scale:
0 = Absent
1 = Present
2 = Occasional to frequent
3 = Total cover
Curiously, data presented in the report appeared in larger, 500 m x 500 m squares, presumably to allow each estuary to be displayed on an individual A4 page. This resulted in a loss of accuracy and meant that significant areas of numerous beds extended over land. This became problematic when these maps, incorrectly digitised as polygons, were published as Scottish seagrass bed areas on the OSPAR website, attributed to Fox et al., 1986 (see figure 15). These polygons have subsequently been used erroneously to derive estimates of historical seagrass bed size and estimations of historical seagrass decline.
Figure 15. Comparison of publicly available OSPAR polygon versus newly constructed polygons near Redcastle, Beauly Firth. Both polygons are derived from data in Fox et al., 1986
Click for a full description
Showing an aerial photograph near Redcastle, Beauly Firth. In the map, seagrass polygons from OSPAR are shown as coarse resolution grid squares which expand from dry land area to the sea. More than half the area of the OSPAR polygons covers dry land. Reconstructed polygons (which have taken to account dry land area) are also present in the map. These cover a smaller area and do not expand into dry land areas.
The Fox et al. (1986) report mentions that surveys were not confined to the spring tidal cycle and therefore the seaward edge of the beds were not always reached. Furthermore, an area of the Beauly Firth was not surveyed due to time constraints.
Despite these limitations, this data set was informative regarding seagrass status and new polygons were reconstructed for the current project using the original 250 m x 250 m squares shown in the field survey maps in the report appendix. Also, dry land areas were removed to more usefully assess seagrass bed areas from 1986. This still leaves a theoretical margin of error in terms of bed edge capture of up to 250 m at any given point particularly along the seaward edge and therefore the main value of the data is as a visual comparator and quantitative area figures provided (even for the newly constructed polygons) should be treated with caution (See Table 2).
| Site | Fox et al., 1986 polygons as published by OSPAR (ha) | Fox et al., 1986 polygons reconstructed from original survey data and clipped to high water mark (ha) |
|---|---|---|
| Dornoch Firth | 2,852 | 1,837 |
| Cromarty Firth | 3,241 | 2,184 |
| Beauly & Inner Moray Firths | 1,140 | 541 |
| Total | 7,232 | 4,562 |
Marine Nature Conservation Review, 1987-1998
During the 1980s and 1990s large areas of the Scottish coast were surveyed in order to provide a comprehensive overview of habitats for the Marine Nature Conservation Review (MNCR). The review was augmented by Nature Conservancy Council data spanning back to the 1970s. Area summaries for most marine sectors were published in the 1990s. The reviews contained broadscale habitat maps generated from site reports.
Seagrass beds were reported but were amalgamated with other soft sediment habitats to create larger polygons. For example, the MNCR review of Loch Ryan is based on maps presented in Howson (1989) which have pooled several separately mapped biotope polygons into one larger polygon. In other areas the map legends contain ambiguous information such as “Muddy sand and pebbles with sparse Zostera sp. and Arenicola marina” from the report on lagoons in Shetland and Orkney.
Furthermore, the original data in Howson (1989) used in the MNCR review for Loch Ryan is a rather crudely drawn schematic and while this gives a rough idea of bed position and proportions it is not accurate enough to use in comparative studies of population change.
Therefore, the published MNCR summary maps cannot be used for assessing seagrass population changes without reference to the original survey map from which they have been generated. Unfortunately, many of the original survey maps were not included in scanned copies of the reports e.g. Hiscock (1989) and Covey & Emblow (1992).
Duncan, Nigg Bay, MSc, 1991
A copy of this report was not available and therefore the methodology and conclusions could not be studied. A photocopy of the Zostera distribution map presented in Davison (1997) was digitised to allow area calculations. Field mapping appears to have been of high quality and the map provides a useful addition to the historical collection for this site.
Atkins, Caudwell and Herbert, Montrose Basin, 1991
Mapping was undertaken using the approximately the same grid system and methodology as McLusky and Roddie in 1982 which aided comparative analysis between the two surveys.
Hawker, Solway Firth, 1993 & 1994
Mapping was undertaken with a compass and sextant with species of the genus Zostera the subject of a dedicated survey. Mapping occurred at a detailed level with seagrass distribution around channels displayed. These maps of individual beds are therefore of high quality. Beds were located using transect routes across estuaries and therefore some beds potentially could have been missed (Hawker, 1993 & 1994; Carstairs, 2024).
RSPB, Udale Bay and Nigg Bay, 1993, 2000 & 2011
Udale Bay was surveyed by the RSPB in 1993, 2000 and 2011 (Hodge, 1993; Stoneman, 2000; Foot, 2012). Quadrat data was collected along eight transects though these did not cover the entire area occupied by seagrass beds which extended beyond the RSPB reserve boundary. Transect bearings were established using a prismatic compass and quadrats sampled at intervals of ten double paces (Hodge, 1993). Unfortunately, the survey methodology was implemented prior to widespread use of hand-held GPS units and therefore co-ordinates for the quadrats are not available. A map of continuous Zostera distribution up to the reserve boundary is presented in the 1993 report with patchy and sparse Zostera excluded. Quadrat data are presented graphically in the three reports. 17 transects were surveyed in Nigg Bay in 2011 though sampling did not occur here in 1993 or 2000. (Foot, 2012). The documents were obtained at the end of the current project which precluded exploration of the feasibility of constructing distribution maps from the raw data.
Quality assessment of contemporary seagrass bed maps
Recent studies using hand held GPS have not been described individually in this quality assessment. GPS studies appear to have been predominantly undertaken using two methods (1) taking individual GPS points and (2) using a tracking function which gathers multiple points as the surveyor walks, gathering data along a seagrass bed periphery but also any exploratory ventures into the surrounding mudflats to search for seagrass plants.
The quality of polygons can be considered high where multiple GPS points e.g. one per metre are gathered to delineate complex bed edges. However, the GPS points used to construct polygons are rarely shown in mapping outputs and it was typically not possible to assess how accurately these studies were conducted.
Dornoch Firth SAC Site Condition Monitoring Report. 2004. Unpublished report.
At the time of writing, only an unfinished draft was available which described the methodology but did not include the results, which were only available as field notes. Data from the survey appears in the GeMS database and OSPAR website map as both points and polygons.
The survey included the intertidal zone of the lower firth as follows:
“The intertidal component of the survey work involved the collection of observations and samples from a series of transects within the Dornoch Firth cSAC. These transects were located within the intertidal part of the cSAC from Newtown Point to the mouth of the Dornoch Firth.”
The surveyors deviated from the transects to record Zostera beds as follows:
“Where the boundaries of Zostera spp. or mussel beds were defined, surveyors mapped the extent of the bed in the area by using the track mapping facility of the GPS to record the route they followed while walking along the bed margins. This action was recorded in target notes for that transect.”
This apparently accounts for two of the four polygons in GeMS. The other two appear to have been produced by drawing around transect point records, resulting in two long, narrow polygons which do not resemble typical seagrass bed shapes and are therefore unsuitable for use in any comparative analysis. The point data does however establish that Zostera was present in several locations in the lower Dornoch Firth in 2004.
Seagrass bed definitions and assessment of data comparability
A variety of survey techniques have been used to survey Scottish seagrass populations. Mapping seagrass presence using point data taken along transects such as McLusky & Roddie (1983) provides a useful visual representation of seagrass distribution (Figure 28). The accuracy at which seagrass distribution is captured is a function of both transect spacing and sampling intervals along transects.
Directly mapping the position of bed edges can be a useful way to assess seagrass population changes, however differences in how bed structure is defined and surveyed can lead to significant differences in recorded bed size.
Seagrass bed structure
Carstairs (2024) described zones present within a large Z. noltii bed in the Solway Firth (See Figure 16) from which the description below has been adapted:
- A core area with a continuous connection between plants occurring at high density;
- A secondary core area where plants remained continuously connected, albeit with a lower overall percentage cover as plants occurred in mosaics with large patches and strips of mud;
- A peripheral zone where plants occurred fairly regularly though patchily and remained a significant ecological component of the beds;
- Isolated individual plants occurring distantly from and often not in a relatable pattern to the main bed.
Bed structure and shape can vary greatly as can the relative sizes of each zone. Furthermore, not all zones may be present in individual beds (Carstairs, 2024). It follows that different methodologies for mapping seagrass beds are likely to result in significant differences in reported bed sizes.

Figure 16. Zostera noltii bed at Skyreburn Bay, Solway Firth. Image © Siân Carstairs.
Littoral Z. noltii beds can have a well-defined bed edge with an abrupt change from dense seagrass to bare sediments (M. Carstairs, pers. obs.). However, the transition from dense seagrass cover to bare sediments often spans relatively large distances, particularly at the seaward edge of intertidal estuarine beds of Z. marina and Z. noltii (M. Carstairs, pers. obs.). At Skyreburn Bay, Solway Firth, core and secondary core zones of Z. noltii were mapped together, and the peripheral zone mapped separately (Figure 17). The dense core area was 7.74 ha and the peripheral zone covered an additional 9.43 ha. Therefore, inclusion of the peripheral zone more than doubled the size of the recorded bed area. The data highlights the potential for differences in what is considered the bed edge to confound comparisons of change over time when different survey techniques are used. For example, some surveys included sparsely distributed plants within maps while others did not.
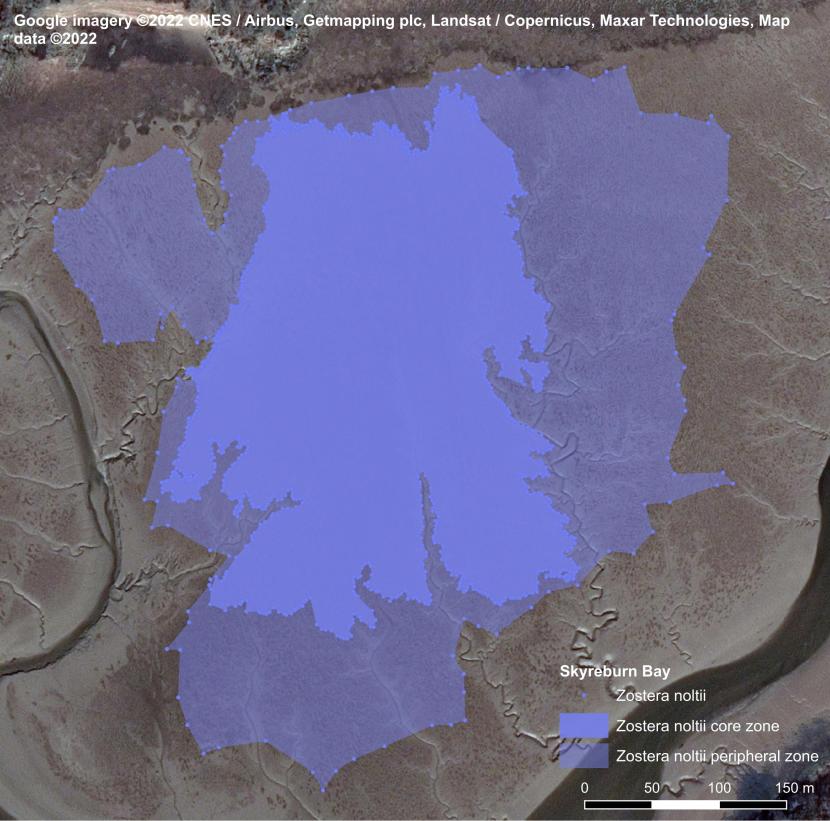
Figure 17. Zostera noltii bed mapping at Skyreburn Bay, Solway Firth, from Carstairs, 2024.
OSPAR (2008) related that:
“to qualify as a Zostera ‘bed’, plant densities should provide at least 5% cover (although when Zostera densities are this low, expert judgement should be sought to define the bed). More typically, however, Zostera plant densities provide greater than 30% cover”
This approach is adopted within the Water Framework Directive methodology used by SEPA which defines the spatial extent of a seagrass bed by the ≥5% cover boundary (WFD-UKTAG, 2014). The 5% cover boundary is also used to define the extent of seagrass PMFs (Tyler-Walters et al., 2016).
Even where a survey methodology is defined, application in the field could be anticipated to lead to significant variation between surveys. Although quadrats are taken to assess seagrass density under the WFD methodology, they cannot be deployed along the entire bed edge and therefore judgment of the ≥5% cover boundary line may be somewhat subjective.
The exact survey techniques used were not always provided or clearly defined in reports. Polygons of seagrass beds are also sometimes available online though an accompanying report describing the survey methodology is not available. Furthermore, the use of aerial photography is considered unlikely to be able to differentiate between bare sediments and sediments with sparsely distributed seagrass plants though we did not encounter any comparative studies on this subject. During WFD assessments, SEPA have used aerial imagery to supplement hand held GPS assessments e.g. at Montrose Basin where thixotropic muds precluded surveying on foot (Scottish Government, 2020b). The intricacy of maps published online for Nigg Bay by SEPA also indicate aerial imagery has been utilised.
Mapping bare patches within beds
Most surveyors did not map the numerous small bare patches which may occur within a seagrass bed. However, polygons published online by SEPA for Nigg Bay show that these patches (some just a few centimetres across) have been subtracted from the total bed size (Figure 18) which according to our calculations has reduced the reported bed size by approximately a third. It would not be appropriate to compare the area quoted by SEPA with historical surveys by other workers at Nigg Bay. Therefore, we have traced the outline of SEPA bed edges for inclusion in data tables (Figure 19) The SEPA data for Udale Bay has the same issue so an outline has also been created for that bed (not shown).

Figure 18. Close-up of SEPA 2015 mapping of Nigg Bay showing area included in reported bed size (green) and area excluded (white).
Figure 19. New outline for SEPA Nigg Bay data to facilitate comparison with historical surveys
Mapping bed presence
Recording differences may also extend to entire beds and may affect whether sparsely distributed seagrass populations are mapped at all during survey programmes. Sparsely distributed seagrass plants, at well under 1% cover, can occur, for example over extensive areas within estuarine embayments (M. Carstairs, pers. obs.) These plants would not be considered to comprise a bed under the OSPAR or PMF habitat definitions, and it would seem extremely unlikely that satellite imagery or aerial photography would be able to detect plants at this low density.
Survey methods, coverage & effort (non-aerial)
Seagrass surveys have been undertaken across Scotland by different organisations to fulfil a range of objectives. Consequently, a variety of methodologies have been used to conduct surveys which in many cases precludes direct comparison of results.
Accurately mapping the entirety of seagrass beds within an entire ecosystem e.g. an estuary requires substantial planning, effort, diligence, skill, and resources. This is also true of individual large beds within these ecosystems. Several of the intertidal reports reviewed stated that not all beds within estuaries were mapped, which hampers comparative studies. Deep mud precluded surveying on foot in a few locations. Inability to survey the full extent of the seaward edge of intertidal beds due to tidal constraints was declared in three reports.
A range of methods have been used for surveying subtidal beds including waders, snorkelling, paddle board, drop down camera, and diving. However, methods restricting survey depth to very shallow waters may not facilitate comprehensive surveying.
Summary
Caution has been applied in this study when comparing results obtained in different studies. This is particularly the case for beds where different survey techniques have been used e.g. mapping sparsely distributed plants on foot versus the Water Framework Directive methodology or aerial imagery.
OSPAR water quality data set 1990-2008
It was necessary to attempt some comparison between water quality trends and seagrass population dynamics as part of the project. This provided a challenge as to which water quality data to use. The suite of water quality parameters sampled during national SEPA monitoring is limited in relation to the potential mixture of chemicals that may occur. Furthermore, water quality sampling is relatively infrequent and usually restricted to the water column whereas pollution can be retained for significant periods of time within settled sediments. Therefore, water quality sampling programmes and assessments can miss pollution on both spatial and temporal scales for the following reasons: position of sampling points being too distant from local sources of pollution to detect effects; failure to detect pollutants at high concentrations in benthic sediments (Fraser & Kendrick, 2017); failure to detect pollutants not on the scheduled monitoring list (Whelan et al., 2022) and failure to detect acute episodes of pollution.
OSPAR water quality data (Scottish Government, 2020a) was used to indicate recent trends in coastal water quality in order to compare with seagrass population trends. This data set contains numerical values for phosphate and nitrate concentrations over time, however the sampling locations and mapping frequency for each region are not provided and therefore conclusions are given tentatively.
Assessment of Previous Studies of Seagrass Declines
Assessment of seagrass declines in UK waters by Green, Unsworth, Chadwick and Jones (2021)
Two methods were used to assess the decline of seagrass in the UK, a comparative analysis of past and present surveyed seagrass areas, and the creation of a modelled theoretical maximum seagrass extent across Great Britain (Northern Ireland was excluded) with which to compare current seagrass extent.
Both approaches relied heavily on data from Scottish sites and this review of their paper is based on our knowledge of that data and those sites. Data for England, Wales and Northern Ireland was not examined in the current review.
To produce a historical baseline for the UK seagrass population, and a contemporary extent to compare with it and assess the decline, Green et al. (2021) gathered polygons representing seagrass beds from various sources, with 1998 chosen as a cut-off date between the two periods.
The review of Scottish sites was almost entirely based on intertidal data. The data used in the ‘historical’ period spanned from 1986-1997 and data in the ‘contemporary’ period ranged from 2006-2018. Therefore, their analysis of Scottish beds is essentially an assessment of potential intertidal seagrass declines between the late 20th century and the early 21st century.
Where data was found in both time periods for the same site, the contemporary area was subtracted from the historical area and the difference labelled as a ‘high certainty loss’, meaning that the authors had a high certainty that the historical bed area had reduced in size to the contemporary area.
Where no contemporary data was found for a site with historical data, the whole historical area was labelled as a ‘medium certainty loss’ meaning that the authors had only a medium certainty that the historical bed area had been lost in its entirety because the possibility existed that it was still there but had not been resurveyed in the contemporary period.
For Scotland, all of the historical data came from the OSPAR dataset, captured by the authors in 2017 (Table 3), with the polygons for Dornoch Firth, Cromarty Firth and Moray Firth contributing 97% of the total area. The source for this data is the report produced by Fox et al., 1986 and, as discussed earlier, these polygons as published by OSPAR are erroneous. They are a large overrepresentation of what was actually recorded during the surveys with polygons drawn partly over dry land. They are not reliable as a measurement of bed extent and have produced a severely exaggerated historical baseline.
| Date | Place Name | Dataset | Data Owner | Historic area (ha) | Contemporary area (ha) | High certainty loss (ha) | Medium certainty loss (ha) |
|---|---|---|---|---|---|---|---|
| 1986 | Cromarty Firth | OSPAR | Fox et al. 1986 | 3,241.47 | 1,200 | 2,041.47 | 2,041.47 |
| 1986 | Dornoch Firth | OSPAR | Fox et al. 1986 | 2,862.26 | 116.98 | 2,745.28 | 2,745.28 |
| 1986 | Moray Firth | OSPAR | Fox et al. 1986 | 1,139.56 | - | - | 1,139.56 |
| 1995 | Islay | OSPAR | JNCC | 43.58 | - | - | 43.58 |
| 1995 | Loch Caolisport | OSPAR | JNCC | 4.38 | - | - | 4.38 |
| 1995 | Loch Carron | OSPAR | JNCC | 3.3264 | - | - | 3.3264 |
| 1995 | Loch Creran | OSPAR | JNCC | 1.16 | - | - | 1.16 |
| 1997 | Shetland | OSPAR | JNCC | 6.67 | - | - | 6.67 |
| 1995 | Loch Sheildaig | OSPAR | JNCC | 11.18 | - | - | 11.18 |
| 1995 | Loch Sunart | OSPAR | JNCC | 3.33 | 0.03 | 3.3 | 3.3 |
| 1995 | Loch Sween | OSPAR | JNCC | 28.31 | - | - | 28.31 |
| 1995 | Loch Torridon | OSPAR | JNCC | 8.85 | - | - | 8.85 |
| 1990 | Lochgolihead | OSPAR | JNCC | 2.14 | - | - | 2.14 |
| 1995 | Longa Island | OSPAR | JNCC | 24.45 | - | - | 24.45 |
| 1995 | Oban | OSPAR | JNCC | 5.04 | - | - | 5.04 |
| 1995 | Skye | OSPAR | JNCC | 38.83 | - | - | 38.83 |
| 1991 | Dumfries area | OSPAR | JNCC | 36.55 | - | - | 36.55 |
| 1990 | Stranraer | OSPAR | JNCC | 3.86 | - | - | 3.86 |
The ‘contemporary’ Scottish data in the study came from a variety of sources, mainly NatureScot, Foster & Davidson (2018) and Community of Arran Seabed Trust (COAST). The largest area by far was 1,200 ha for the Cromarty Firth, representing 55% of the total ‘contemporary’ data for Scotland. The source listed for this in the supplementary data was described as a ‘paper’ by ‘Chapman, 2008’. Instead of a research paper containing seagrass polygon data, it appears to be an updated UK Biodiversity Action Plan Priority Habitat Description for seagrass, described as ‘Edited by Nikki Chapman’ and dated 2008 which contains the following paragraph:
“The Cromarty Firth supports what is most probably the largest total area of dwarf eelgrass and narrow leaved eelgrass in Britain (approximately 1,200 ha), while the Maplin Sands is estimated to be the largest surviving continuous population of dwarf eelgrass in Europe (covering around 325 ha).”
The 1,200 ha figure is also quoted in the original UK BAP Habitat Description for seagrass published in 1995 (UK Government, 1995). This figure is taken from a coastal directory published in 1988 (Gubbay, 1988) and the accompanying text has been slightly modified and incorporated into the BAP.
The ultimate origin of the 1,200 ha figure is unclear but digitisation and analysis of the Cromarty Firth polygons in the 1978 NCC prospectus for the Moray Firth gave a similar area and shape to polygons reproduced in Gubbay (1988). The cumulative figure calculated in QGIS for the 1978 polygons digitised during the current study was 1,505 ha which is within the margin of error which could be expected to be obtained from historical calculations of polygon size. It is also stated in Gubbay (1988) that the NCC 1978 prospectus was used as a source of information.
It seems likely then that the Green et al. (2021) analysis for Cromarty Firth has compared a ‘historic’ seagrass area of 3,241 ha from 1986 with a ‘contemporary’ seagrass area of 1,200 ha originating from circa 1978 and erroneously concluded a ‘high certainty loss’ of 2,041 ha.
For the Dornoch Firth, an inflated historic area has been compared with a contemporary area derived from a limited 2004 SNH survey of part of the SAC. As the authors acknowledge, point records exist outside of bed areas but because these do not provide an indication of extent, they have been ignored for the purpose of their analysis. In the Dornoch Firth there are point records from 2004 that show presence of seagrass outwith the polygons which would indicate a conclusion of high certainty of loss in these areas is misplaced. Green et al. (2021) concluded that:
“The meadows where historic and contemporary data are available show mass declines. The once huge meadows in the Cromarty and Dornoch Firths have been reduced from 3,241 to 1,200 ha and 2,546 to 117 ha, respectively.”
Considering the challenges with both the historical and contemporary data for these sites, coupled with the undue certainty about the declines, this conclusion is incorrect.
For the Moray Firth, an inflated polygon was used for the historical 1986 baseline and no contemporary bed data was found. The authors therefore ascribed a ‘medium certainty loss’ of the entirety of the 1,139.56 ha. Widespread point records from 2008 demonstrate that this was not the case.
Overall, the Green et al. (2021) paper tabulated a ‘High certainty, known seagrass loss’ of 4,790 ha, or 58%, in Scotland since 1936. Since this is based on inaccurate data, which in any case only dates back to 1986, this is an erroneous assertion.
The medium certainty analysis performed on the other sites relied on OSPAR polygons derived from broadscale MCNR surveys, conducted during the 1990s, which were not dedicated seagrass surveys. Limited surveying has occurred in Scotland since and a systematic resurvey of MNCR areas has not been undertaken. Ascribing a ‘medium certainty loss’ where no contemporary data exists is therefore inappropriate in Scotland. For example, a ‘medium certainty loss’ was ascribed for the ‘Dumfries area’ (Table 3) though these beds, namely Auchencairn, Orchardton and Rough Firth, were recorded in 2023 (Carstairs, 2024) and had undoubtedly remained present since they were last surveyed in the early 1990s (Covey & Emblow, 1992; Hawker, 1993, 1994). The bed at Stranraer was described as a ‘medium certainty loss’ though was actually present during the contemporary period (Foster & Davidson, 2018).
The modelled analysis of seagrass losses across Great Britain used the inflated OSPAR polygons for the Dornoch, Cromarty and Moray Firths to populate the model and these are anticipated to have skewed the results.
Seagrass Population Dynamics by Scottish Marine Region
Moray Firth
Figure 20. Moray Firth Scottish Marine Region.
The Moray Firth Scottish Marine Region (SMR) extends from Duncansby Head in the north to the Beauly Firth near Inverness and eastwards to Fraserburgh (Figure 20). The outer exposed coastlines of the Moray Firth SMR are fairly rocky and uniform, grading through shingle and sand to fine sediments in the central area where four firths and Loch Fleet discharge water from vast areas of the Highlands into the sea. These large waterbodies contain extensive sand and mudflats and are variously protected at their mouths from incoming wind and waves by spits and headlands. They are therefore highly suitable for the development of seagrass populations which have been documented within this region as early as 1837 (Watson, 1837). The seagrass beds of the Moray Firth SMR appeared to remain unaffected by the 1930s wasting disease (Berry, 1939).
Loch Fleet
Seagrass was plentiful in Loch Fleet at the turn of the 19th century (Marshall & Shoolbred, 1898) and until recently the beds had remained abundant and were an important source of food for overwintering wildfowl. The Loch Fleet seagrass beds have been declining steadily since at least 2012 and are now at risk of extinction. The upper end of Loch Fleet was truncated during construction of The Mound which is a tidal barrier built between 1814-1816 for the dual purpose of allowing stagecoach passage across the valley and reclaiming land in the upper estuarine reaches. The structure contains a series of sluice gates at the northern end to manage the river water flowing into Loch Fleet and the ingress of tidal water to areas above the gates. In 1983 a bridge was constructed over the River Fleet at the northern end of the barrier to improve traffic flow but this has had a virtually negligible effect on the hydrology of the ecosystem as the original sluice gates have remained in place.
The seagrass beds were assessed as being in favourable condition following monitoring during September 2000 (Scottish Natural Heritage, 2011).However, subsequent monitoring later revealed that the beds had reduced in extent by 82% between 2000 and 2012 (Figure 21) with a further decline between 2012 and 2016 (NatureScot, Cycle 3 - undated internal site check notes). The species composition had also changed significantly, with intertidal Z. marina becoming virtually absent in the Creag Bheag bed. A few sparsely distributed seagrass plants were all that was encountered during a walkover in summer 2025 (F. Williams, pers. comm.).
The decline appears to have occurred following changes in the sluice gate operations. Until late 2000, the gates had been operated manually during spates, but after this, reliance shifted solely to a one-way valve system. Potential causal pathways are outlined below:
- Elevated or fluctuating groundwater level upstream of the A9 due to backed-up freshwater flows, potentially mobilising nutrients from the floodplain and/or any contaminants present in the artificially constructed ‘mound’;
- Increased water depth in Loch Fleet caused by saline water backing up at the sluice gates, reducing light penetration;
- Changes in flow regime potentially leading to changes in duration of seagrass immersion;
- Changes in flow regime leading to changes in scouring and rates of sediment deposition.
The first factor is a theoretical possibility but cannot be assessed further here due to absence of data. Any increase in water depth in the main body of the loch is likely to be small, with effects on seagrass beds limited to the lower shore, which is not reflected in the data. Therefore, even if factors 2 and 3 occur, they are unlikely to be significant. If a change in flow regime was responsible for seagrass declines, factor 4 would be the most probable cause. The loss of virtually all seagrass near the sluice gates between 2000 and 2012 suggests the gates may have been a causal factor, though significant declines also occurred across most of Loch Fleet (Figure 21).
![A map showing seagrass distribution in Loch Fleet during 2000 depicted by a red polygon and in 2012 depicted by a green polygon. Seagrass significantly declined on the north eastern shore and had almost disappeared from the other areas by 2012. © Crown copyright [and database rights] 2024 OS 100017908](/sites/default/files/styles/embedded/public/2025-12/figure-21-higher-resolution.png?itok=NHGUBMn5)
Figure 21. Loch Fleet seagrass distribution during 2000 and 2012 from Axelsson & Dewey (2013).
The close timings of the change in sluice gate operation and seagrass decline may be coincidental, and the large spatial scale of the decline points more towards a water quality issue. Distillery waste has been spread on fields in addition to livestock manure, and blanket weed was reported as being common in 2012 (Axelsson & Dewey, 2013).
It is also possible that factors such as hydrological changes and water quality issues combined to increase pressures on seagrass beyond a critical threshold.
Other potential contributing factors include bioturbation by marine invertebrates and wildfowl grazing. High densities of Arenicola marina have been reported, though population distributions and densities have not been mapped. In any case, high densities of bioturbating invertebrates in former seagrass beds areas are more likely to be a symptom rather than a cause of decline (Philippart, 1994).
The relationship between wildfowl populations and Zostera across the biogeographic region appears to be relatively stable. For example, Ganter (2000) found no evidence of long term adverse impacts on seagrass beds due to grazing by brent geese. However, other species of wildfowl such as wigeon, mallard and pintail (Jacobs et al., 1981), greylag geese (Pistorius et al., 2006), mute swans (Berry, 1939; Binnie et al., 2012) and whooper swans (Ogilvie, 1992) can also exploit Zostera beds and these may have become problematic once the Zostera beds declined below a critical size. Axelsson & Dewey (2013) reported the presence of hundreds of greylag geese at Loch Fleet in addition to other birds. Berry (1939), while discussing the decline of wigeon in the Firth of Tay, indicated that mute swans were responsible for the decline of the remaining Zostera beds following the wasting disease epidemic:
“The disappearance of Zostera was blamed for their scarcity [wigeon] and this in turn was attributed by local gunners to the hundreds of Mute Swans which finally collected on the last remaining Zostera beds near Tayport.”
It is understood that the sluice gates have been repaired, and monitoring could be conducted to determine whether seagrass populations show a corresponding improvement. However, even if the cause of the decline has been mitigated, the establishment of dense populations of Arenicola marina may inhibit recovery. Restoration of a near-natural flow regime e.g. by way of removal of the sluice gated causeway section would be beneficial for marine habitats and migratory fish. For example, this would prevent potential long-term impacts on the protected features of the sites, including Atlantic salmon, caused by changes in the management or breakdown in operational functionality of the sluice gates. However, negative impacts on the Mound Alderwoods SSSI would be anticipated and therefore a detailed feasibility study of the proposal would be required.
Dornoch Firth
The only comprehensive survey that was found for the Dornoch Firth was Fox et al. (1986). Since then, no dedicated seagrass surveys have been systematically conducted for the entire firth with published maps. Scottish Natural Heritage (SNH) carried out site condition monitoring in 2004, which included transects to assess seagrass distribution along with very limited bed mapping. The area of seagrass recorded was substantially lower than that recorded by Fox et al. in 1986 (Table 4) however, it seems that only a few areas were surveyed in 2004. The presence of two seagrass transects in the southern firth suggest that beds were present in the area but not mapped. Polygons around these transects are considered to have been inappropriately drawn at a later stage (Figure 22).
| Sites | Fox et al. 1986 (reconstructed polygons) (ha) | SNH 2004 (ha) |
|---|---|---|
| Several locations | 1,837 | - |
| Cuthill Sands | - | 102 |
| Edderton Sands | - | 5 |
| Tain | - | 10 |
| Total | 1,837 | 117 |
Green et al. (2021) asserted that a reduction in bed size from 2,546 ha to 117 ha occurred in the Dornoch Firth between 1986 and 2004. However, this analysis used inflated figures erroneously derived from the Fox et al. (1986) OSPAR polygons to represent the ‘historical’ position, while the ‘current’ extent was based on the limited data from the 2004 SNH survey.
In summary, there may have been a substantial decline in the seagrass population of the Dornoch Firth though lack of dedicated surveys preclude firm conclusions.
Figure 22. Scottish Natural Heritage, Dornoch Firth site condition monitoring Zostera survey 2004.
Cromarty Firth
A herbarium specimen of Z. marina was collected at Cromarty in 1827 (RBGE E00762029) while Watson (1837) documented a record of the taxa from Invergordon. Vast intertidal beds are described in modern nature conservation literature (NCC, 1978; Fox et al., 1986: Gubbay, 1988) although it is clear from earlier descriptions that extensive subtidal beds were also historically present.
St. John (1863) described intertidal beds and how these are exploited by brent geese:
“There is a kind of short grass in the sheltered bays on parts of this coast, which at high water is covered, but which is bare at low tide, and this seems to commence throwing out young shoots early in March. The brent geese are very fond of this, and leave the bay of Cromarty and other places where they have fed during the winter on the roots of the long waving seagrass, in order to feed on this young short herbage which the ebb tide leaves uncovered here.”
The geologist Hugh Miller, who grew up on the shores of the Cromarty Firth, also described the occurrence of extensive subtidal Zostera beds (Miller, 1854; Miller, 1857). A selection of Miller’s descriptions are given below:
- “Zostera marina abounds on a series of sand banks, partially uncovered by the larger stream tides, which lie directly opposite the town of Cromarty.” (Miller, 1854)
- “Zostera marina, such as I had often seen on the Cromarty beach thrown up from the submarine meadows of the Firth beyond.” (Miller, 1857)
- “I felt a good deal of interest in marking, about this time, how certain belts of marine vegetation occurred on a vast boulder situated in the neighbourhood of Cromarty, on the extreme line of the ebb of spring-tides….But far beyond its base, where the sea never falls, green meadows of Zostera flourish in the depths of the water.” (Miller, 1854).
The boulder described by Miller is almost certainly Clach Mhallaichte, one of the ancient boundary stones of the township of Cromarty.
Mapping undertaken in the 1970s indicates that around 1,350 ha to 1,500 ha of seagrass was present in the Cromarty Firth (see Table 5). Fox et al. (1986) found that these beds were still abundant though the figures for the Fox et al. (1986) reconstructed polygons are still considered to represent an overestimate of bed size and are not discussed further here. Large-scale surveys of the estuary were not conducted during the 1990s though results from Nigg Bay (Duncan, 1991) are similar to previous studies at the site.
Udale Bay was surveyed by the RSPB in 1993, 2000 and 2011 (Hodge, 1993; Stoneman, 2000; Foot, 2012). While an overall change in distribution of seagrass was not detected changes in species distribution and density were reported.
The extent of seagrass mapped within the RSPB reserve in Udale Bay in 1993 is noticeably smaller than the distribution in the reserve recorded during the 1970s (Figure 48, Annex 2). Similarly, the areas of seagrass mapped by SEPA during 2015 in Nigg Bay and Udale Bay are smaller than those obtained during the 1970s. This may be a function of low density plants being recorded within beds during the 1970s but not during 1993 and 2015. Hodge stated that his map excluded ‘areas of very sparse, discontinuous cover’. Davies et al. (2024) detected 800 ha across the Cromarty Firth between 2017 and 2023 using satellite imagery (Table 5). This figure is approximately half of that recorded during the 1970s and may indicate a decline in bed size. However, the differences in methodology between surveys in this case, full field mapping, including low density areas vs. satellite imagery could account for a significant proportion of the variation.
A small amount of the variation can be attributed to the construction of the Port of Nigg in 1979, which directly led to the loss of 27 ha of seagrass bed habitat recorded in the 1970s (NC 1972; NCC, 1978).
| Bay | NC 1972 | NCC 1978 | Rae 1979 | Duncan 1991 | Fox et al., 1986 reconstructed polygons | SEPA 2015 (new outlines) | Mossy Earth 2023 | Davies et al., 2024 |
|---|---|---|---|---|---|---|---|---|
| Alness Bay | 155 | 284 | - | - | 258 | - | - | - |
| Nigg Bay | 534 | 629 | 511 (821)* | 554 | 752 | 353 | - | - |
| Udale Bay | 377 | 369 | - | - | 436 | 165 | N/A** | - |
| Upper Cromarty Firth | 250 | 217 | - | - | 499 | - | 189 | - |
| Cromarty Bridge to River Sgitheach | 18 | 6 | - | - | 129 | - | 6 | - |
| Yankee Pier | 17 | - | - | - | 111 | - | - | - |
| Total | 1,351 | 1,505 | - | - | 2,185 | 518 | 195 | 800 |
The huge loss in the Cromarty Firth reported by Green et al. (2021) was merely a result of comparing inflated figures for 1986 with ‘contemporary’ data which appears to have preceded the ‘historical’ data.
Nigg Bay
Historical seagrass maps from the 1970s (NC, 1972; NCC, 1978) show similar bed sizes and distributions to surveys undertaken by Fox et al. (1986). However, recent surveys by SEPA (unpublished data from GeMS) show smaller bed sizes in Nigg Bay, which do not extend as far down the shore (See Figure 23; Annex 2). There are several potential reasons for the difference in mapped areas, including differences in survey methodology, and changes in seagrass distribution potentially resulting from changes in the beach elevation profile or sea level rise.
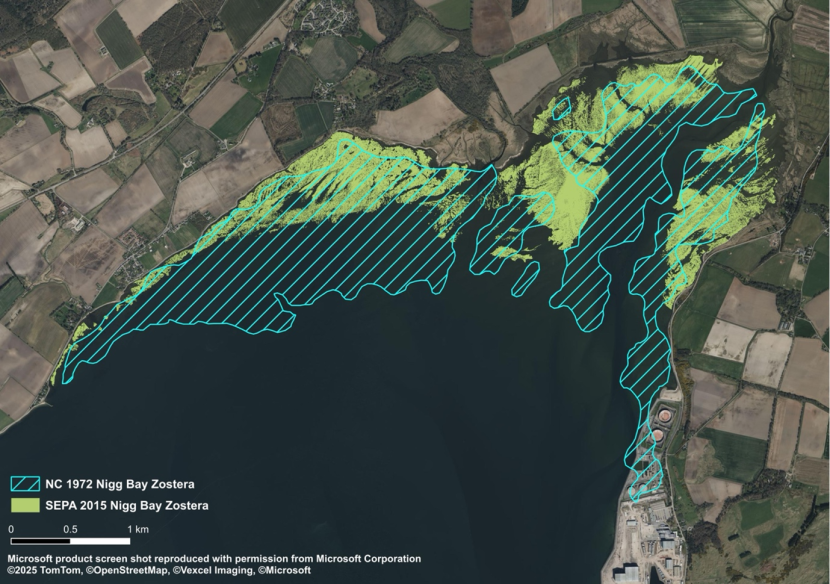
Figure 23. Zostera distribution at Nigg Bay ~1972 and 2015.
Rae (1979) and Fox et al. (1986) mapped sparsely distributed plants, whereas the Water Framework Directive methodology for seagrass monitoring does not require mapping of low density (<5%) seagrass cover. Large intertidal seagrass beds can have a substantial area of sparsely distributed plants, particularly at the seaward edge (M. Carstairs, pers. obs.). This may have significantly affected the validity of comparisons. However, it should be noted that, at the time of writing, SEPA’s report had not been published, so exact details of their survey methodology were not available to the authors.
There is a possibility that significant changes in the elevation profile, and therefore the extent of the intertidal zone have occurred. Historical and contemporary Ordnance Survey (OS) maps show substantial differences in the position of the mean low water spring line, indicating that the intertidal zone has been reduced by up to 2 km in places (Figure 24). This shift may suggest that anthropogenic activities have contributed to a net loss of sediments from the beach over time. This process is typically associated with beaches which have extensive coastal developments or defence structures at the beach head, though could also conceivably be associated with dredging of main channels.
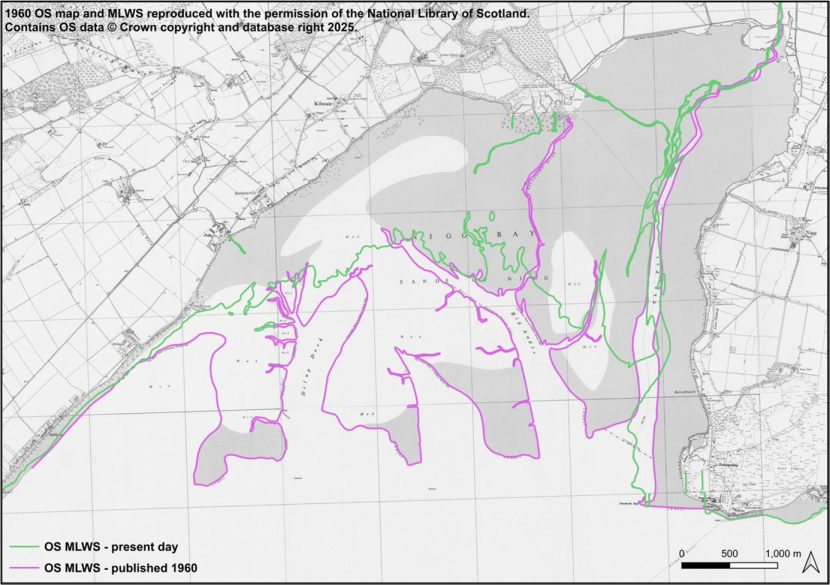
Figure 24. Historical and contemporary MLWS in Nigg Bay.
OS maps are not updated frequently enough to determine when this change happened or whether it is a gradual process. Furthermore, details of the intertidal zone appear to have been copied from one edition to another over extended time periods, with only one change occurring between circa 1890 and the present. The mapping methodology of the OS MLWS is loosely defined and has historically involved the average of two low spring tides. More recently, it may rely on one aerial photography survey. As a result, prevailing conditions at the time of surveying may lead to substantial differences in where the MLWS line is mapped, particularly within the low gradient setting of an extensive intertidal mudflat.
For example, the Lowest Astronomical Tide at Nigg Bay is 70 cm lower than MLWS, which could easily ‘widen’ the exposed area by hundreds of metres. Variations in atmospheric pressure also affect sea levels, therefore it is not currently feasible to conclude, on the basis of two published OS MLWS lines that changes in elevation profile and beach extent have occurred. It is likely that the apparent change merely reflects aspects of the tidal survey methodology.
Investigations should be initiated to establish if a historical change in elevation profile can be corroborated from different sources, such as navigational charts and LiDAR data.
Beauly Firth
Marshall noted an abundance of the intertidal variety of Z. marina at Lentran in 1892 (RBGE E00762057).
Losses appear to have occurred in the Beauly Firth (Table 6).
Several beds also appear to have declined in size since 1978 and 1986 (Figure 25). However, it should be noted that the bed extent from 1986 is likely inflated due to survey methodology. In addition, the 1978 NCC survey maps only covered the western half of Beauly Firth while surveying in 2023 did not extend west of Lentran on the southern shore due to the presence of deep mud (F. Williams, pers. comm.).
| Beauly Firth | NCC, 1978 | Fox et al., 1986 reconstructed polygons | Mossy Earth, 2023 |
|---|---|---|---|
| Total | 95 | 308 | 14 |
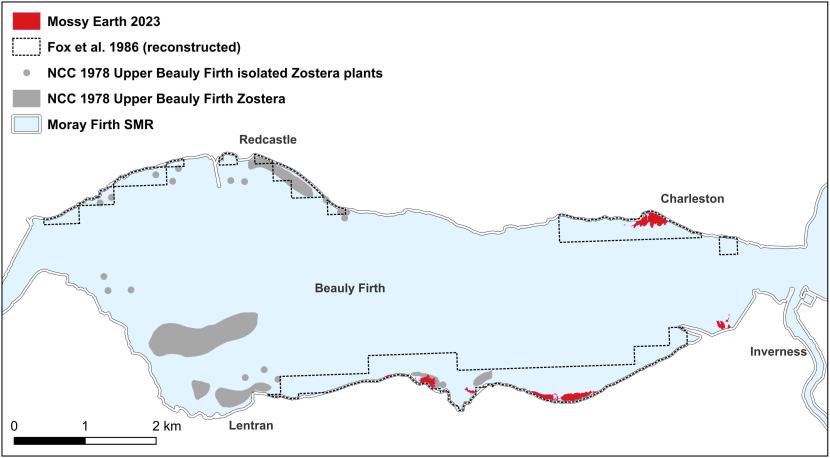
Figure 25. Beauly Firth historical and contemporary seagrass areas.
Inner Moray Firth (Inverness Firth)
A herbarium specimen of Z. marina was collected from sands east of Fort George in 1832 (RBGE E00762066).
The seagrass bed at Munlochy Bay appears to have remained relatively stable between the 1970s and 2023 (Table 7). The bed at Longman Bay may have declined in size between 1986 and 2023 though the 1986 figures are likely to be inflated.
| Bay | NC, 1972 | NCC, 1978 | Fox et al. 1986 reconstructed polygons | NatureScot, 2021 | Mossy Earth, 2023 |
|---|---|---|---|---|---|
| Munlochy Bay | 68 | 68 | - | - | 60 |
| Longman Bay | - | - | 234 | - | 99 |
| Rosemarkie Bay | - | - | - | 0.6 | 0.7 |
| Castle Stuart Bay | - | - | - | - | 66 |
| Total | 68 | 68 | 234 | 0.6 | 226 |
Outer Moray Firth
Seagrass has been recorded at a variety of locations including Culbin Sands, Findhorn, Duffus and Lossiemouth (Annex 1). A herbarium specimen of Z. marina was collected by G. Gordon at Burghead in 1836 (RBGE E00762024). Subtidal Z. marina beds have recently been recorded at Rosemarkie and Burghead and there has been a recent report of a bed at Spey Bay though this has not been confirmed (F. Williams, pers. comm.). Washed up Z. marina plants have been historically recorded further eastwards. Craib (1912) stated that the plant was “Washed in on rocks near Banff Harbour and near Gardenstown”.
Several specimens of the subtidal variety of Zostera marina were washed up in Sinclair’s Bay, north of Wick in 2025 (M. Carstairs, pers. obs.).
Summary
The recent status of seagrass beds in the Moray, Beauly, Dornoch and Cromarty Firths is unclear due to a relative lack of surveying, and the use of different survey methodologies. Further research is urgently required to establish whether recently recorded substantial declines are real. The seagrass population in Loch Fleet declined significantly (99%) between 2001 and 2016. The beds have not been surveyed since 2016 therefore their current status is unknown.
North East
Figure 26. North East Scottish Marine Region.
The North East SMR is located in the northern North Sea (Figure 26). The north east coast from Fraserburgh in the north to St Cyrus in the south has sandy beaches but few indentations and only one small estuary, the Ythan, half way between Peterhead and Aberdeen. The land along the coast is farmed and all land north of Aberdeen is designated as an NVZ. No definite records of Z. marina growing in situ were found. Therefore, this region will not be discussed further.
Forth and Tay
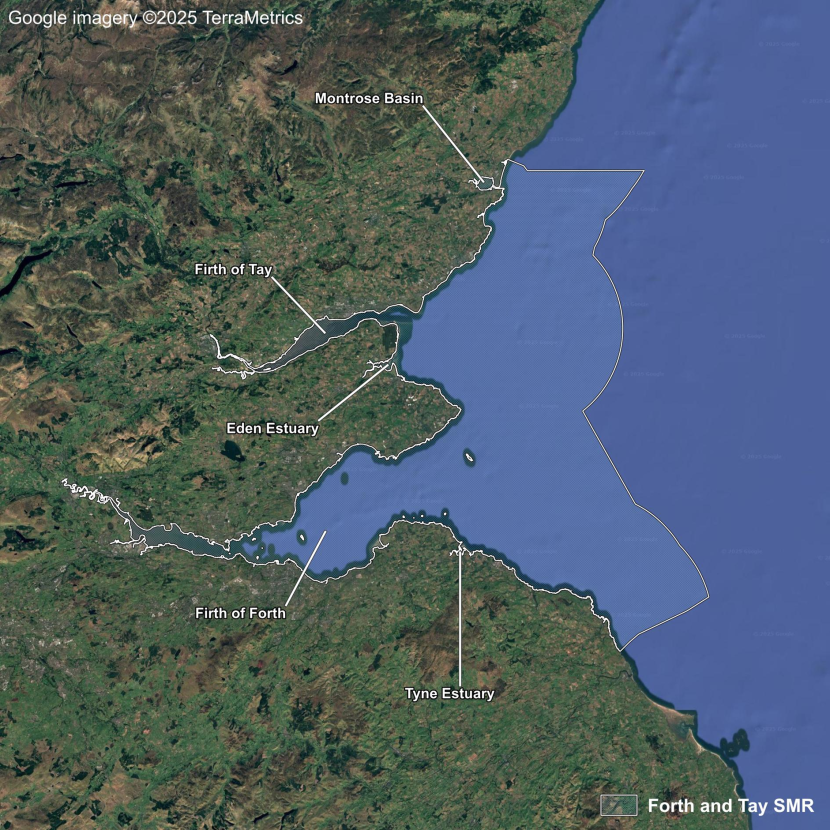
Figure 27. Forth and Tay Scottish Marine Region.
The Forth and Tay SMR extends from St Cyrus in the north to the Scottish border (Figure 27). The coastal lowlands are intensively farmed with large swaths designated as NVZs. This means that small coastal catchments are prone to eutrophication. The Firth of Tay is fringed by NVZs on both sides but the catchment extends westwards beyond these zones through a variety of land uses and eventually into the Grampian Mountains. The catchment of the Firth of Forth is heavily urbanised in places and covers a large area of the rural central lowlands. The land east of Edinburgh is designated as an NVZ.
Seagrass beds occur in a number of estuaries described individually in sections below. In addition, Z. marina has been recorded at Johnshaven, the estuary mouth at St Cyrus, Lunan Bay and Victoria Park Beach (Ingram & Noltie, 1981; BSBI, 2024). Intertidal Z. marina was historically recorded at Kinkell, St Andrews in 1937 and 1957 (BSBI, 2024).
Montrose Basin
Don (1813) first described intertidal Z. marina beds at Montrose Basin using the local name “Back Sands”. The first comprehensive survey occurred in 1982 (McLusky & Roddie, 1983). A survey by Ackroyd in 1972 did not cover all of the basin, and a copy of the report showing the seagrass beds in detail could not be obtained for the current project. However, McLusky & Roddie (1983) stated that the distribution of Zostera in 1982 was similar to that found by Ackroyd.
All three British taxa were recorded by McLusky & Roddie (1983), though the authors expressed difficulty with identification. The locations of records, with reference to shore position, strongly suggest that only Z. noltii and intertidal Z. marina variety were likely to have been present (all subsequent surveys only record the latter two taxa). Zostera was found to cover approximately half of the basin and was distributed towards the north and west (Figure 28).
Seagrass distribution had declined by 1991 and was absent in the eastern half of the basin (Atkins et al., 1992) while Enteromorpha sp. increased in the same area indicating nutrient enrichment. This was linked to a change in the location of untreated sewage outfalls in 1980 (Atkins et al., 1992) with adverse effects on seagrass potentially talking several years to manifest. Outfalls were moved further away from the seagrass beds towards the basin mouth though incoming tidal currents, which were observed carrying plumes of sewage to the seagrass beds, appear to have confounded the attempt to improve the basin’s water quality (Atkins et al., 1992).
A waste water treatment works was constructed in 2002 and studies conducted by SEPA in relation to the Water Framework Directive between 2008 to 2017 showed seagrass recovery within the eastern basin (Foster & Davidson, 2018) with the 2017 distribution being very similar to that recorded in 1982 (Figure 28).
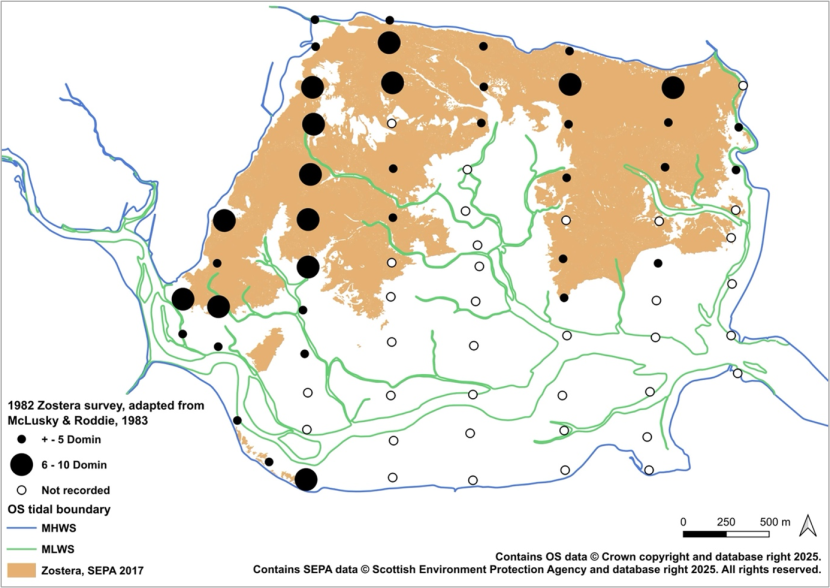
Figure 28. Zostera distribution in 1982 and 2017 at Montrose Basin.
Firth of Tay
A number of 19th century records for Z. marina in the Firth of Tay are held in the herbarium of the RBGE and the BSBI database (BSBI, 2024) dating back to 1835 at Ferry-Port-on-Craig (now Tayport; RBGE E00762117).
Accounts in the wildfowl literature indicate that Zostera populations in the Firth of Tay were severely affected by the 1930s wasting disease epidemic with the last remaining beds occurring at Tayport (Berry, 1930; 1939). The event was considered the cause of a dramatic concomitant decline in the wigeon population (Atkinson-Willes, 1963). Favourable habitats for seagrass in the Firth of Tay are concentrated in the outer parts of the estuary from Tayport and Broughty Ferry seawards with some suitable intertidal habitat in Invergowrie Bay, west of Dundee (M. Carstairs, pers. obs.). However, seagrass has not been recorded from the inner estuary. Salinities at Tayport range from approximately 11‰ to 32‰ (Bates et al., 2004). The seagrass population in the outer Firth of Tay would therefore have been vulnerable to wasting disease which is only considered to be virulent in salinities exceeding around 12‰ to 15‰ (Rasmussen, 1973).
Sparse occurrences of intertidal Z. noltii and Z. marina were found in the Tay during 2002 (Bates et al., 2004). Specific locations were given only for Z. noltii which was distributed across Tayport and in the western end of Monifieth Sands. Intertidal patches and individual plants of these species were present at Tayport during 2025 (M. Carstairs, pers. obs.) though Monifieth Sands was not visited.
Eden Estuary
Wilson (1910) reported extensive Z. marina beds in the Eden Estuary. These beds were almost certainly composed of the intertidal variety, as indicated in a photograph of several specimens provided by the author (Wilson, 1910). Only sparse occurrences of intertidal Zostera were found during surveys in the mid-1980s, 1994 and during 2002 (Caudwell & Jones, 1994; Bates et al., 2004). Extensive algal mats were discernible in an aerial photograph taken in 1988 (Caudwell & Jones, 1994) and large beds of algae in the estuary were recorded during a survey in 1994 (Caudwell & Jones, 1994). Algal mats covered approximately 60% of the northern shores and 40% of the southern shores of the outer estuary with small patches of algae on the northern shores of the inner estuary. Algal mats were absent from the southern shore of the inner estuary though Z. noltii was present. The high abundance of algae and the inverse relationship between seagrass and algal distribution suggests that nutrient enrichment is likely to have been responsible for historical declines of seagrass in the Eden Estuary.
Foster & Davidson (2018) found that seagrass cover over the whole estuary was 17.22 ha in 2013, increasing to 55.88 ha in 2015, though this may have been partly due to increased survey effort. Percentage cover remained stable at 28-29% during the same period.
A significant increase in Z. noltii occurred along the south shore of the inner bay between 2002 and 2015 with some decline in the distribution of intertidal Z. marina (Bates et al., 2004; Foster & Davidson, 2018).
Firth of Forth
A number of 19th century records for Z. marina in the Firth of Forth are held in the herbarium of the RBGE and the BSBI database (BSBI, 2024) dating back to 1825 near Burntisland (RBGE E00762004).
Z. marina was listed by Watson (1837) as:
“on shore of the Forth nearly opposite Cramond Island and at the Black Rocks at Leith”
This description suggests the presence of the intertidal variety. Elsewhere, Watson (1837) described Z. marina as occurring at Aberlady Bay and abundantly between Burntisland and Pettycur.
Balfour (1863) listed Z. marina as occurring at ‘Granton, Burntisland, Aberlady, Pettycur, Black Rocks at Leith’. Sonntag (1894) gave the same locations and noted the species abundance as being frequent. Martin (1934) documented intertidal Z. marina at Aberlady, Leith and Granton. Smith et al. (2002) reported that the intertidal form of Z. marina had declined markedly in Aberlady bay since 1991 “due to algal bloom”, though this is not apparent in data from 1991 and 2000 (Finlay, 2014).
Zoutenbier et al. (2016) determined that coverage of intertidal Z. marina was <0.5 ha, with the authors classifying the species as rare. This indicates that a significant decline of Z. marina occurred in the Firth of Forth since Sonntag described the species as frequent in 1894.
Z. noltii was observed by Evans (1889) on mudflats at Aberlady Bay and ‘in abundance on the mudflats near Torryburn and also at a short distance to the west of the mouth of the Almond at Cramond’. Martin (1934) also documented Z. noltii at Aberlady.
New populations of Z. noltii were recorded at Brucehaven (Limekilns) and Ironmill Bay (Zoutenbier, 2011; Zoutenbier et al., 2016).
Finlay (2014) gave a figure of 70 ha for the total area of seagrass coverage in the Forth Estuary, which was almost exclusively Z. noltii (Table 8). Rogers (2024) and Rayer (2024) surveyed a selection of beds in 2024, and a summary of results is presented in Table 8 alongside data for 2014. Substantial increases in bed size were noted at most sites, though a significantly reduced amount of seagrass was recorded at Tyninghame (see section below).
| Site | SEPA 2014 (ha) | Finlay 2014 (ha) | Rayer / Rogers 2024 (ha) |
|---|---|---|---|
| Culross | 0.26 | 2.04 | 4.55 |
| Torry Bay | 1.88* | 20.94 | 25.94 |
| Ironmill | - | 0.31 | - |
| Brucehaven | 1.74 | 2.55 | 2.50 |
| Dalgety Bay | - | 0 | - |
| Burntisland | - | 5 plants | 0.25 |
| Kincardine Bridge | - | 0 | - |
| Carriden | 8.14 | 7.34 | - |
| Blackness | 25.05 | 25.63 | - |
| Cramond | - | 5.46 | 14.02 |
| Aberlady | - | 1.64 | - |
| Tyninghame | - | 4.37 | 2.33 |
Tyne Estuary
A herbarium specimen of the intertidal variety of Z. marina was collected at Tyninghame in 1869 (RBGE E00762077). Seagrass bed distributions were available as polygons dating back to 1976 with surveys carried out by different workers. These appear to show a successive retreat since 1991. This coincides with several hydro-morphological changes which can be seen in online Google aerial imagery and OS maps. The main channel of the River Tyne has migrated towards the two areas containing Zostera beds i.e. one bed in the upper reaches and one at the river mouth. Parts of the beds have been directly lost where the channel has cut into them. Drainage patterns of mudflats around channels typically differ to those of continuous flats with seagrass generally absent near large watercourses (M. Carstairs, pers. obs.). The effect of channel movements on seagrass at Tyninghame may therefore have extended into adjacent mudflats. Migration of the main channel at the estuary mouth appears to have occurred in relation to the accretion of sediments derived from Belhaven Bay via longshore drift. This may be a natural change, however there isn’t enough evidence of changes in the upstream catchment to conclude this.
The catchment is located within an NVZ in the east coast lowlands and therefore the seagrass beds may also be subject to adverse water quality issues.
Summary
Seagrass beds in the Forth and Tay have experienced historical declines due to factors such as the 1930s wasting disease epidemic and pollution. The seagrass beds in the Firth of Tay suffered a severe historical decline most likely due to their location in the outer estuary where high salinities would have left them vulnerable to wasting disease infection. Current seagrass status in the Firth of Tay is unclear due to paucity of historical and current data, though recovery appears to be limited. Seagrass beds in the Firth of Forth are recovering significantly following historical losses, most likely due to improvements in water quality. The declines at Tyninghame are, at least in part, due to hydro-morphological changes relating to the position of the River Tyne.
The seagrass population in Montrose Basin appears to have been relatively stable between 1972 and 1982 though declined afterwards potentially due to relocation of sewage outfalls. Subsequent population recovery followed the installation of a wastewater treatment works in 2002.
Solway
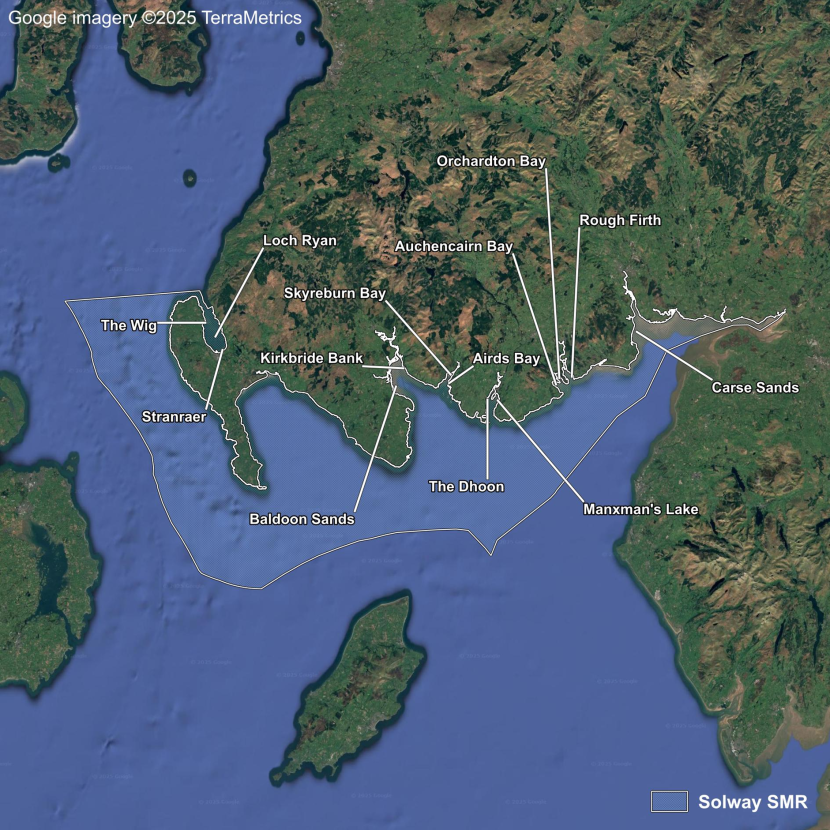
Figure 29. Solway Scottish Marine Region.
The Solway is the most southerly SMR (Figure 29). The firth is estuarine and has a large tidal range with extensive intertidal mud and sand flats. The coastline is diverse with a number of large sheltered embayments which support extensive seagrass populations between Rough Firth and Baldoon Sands. The inner firth between Dumfries and Carlisle is characterised by a fairly linear coastline, shifting sands and relatively poor historical water quality. Seagrass beds have not historically been recorded within the inner firth.
Loch Ryan is a large, relatively shallow sea loch with little freshwater input. The loch’s ecosystem is of national importance, containing the last extensive native oyster beds in Scotland and both intertidal and subtidal seagrass populations.
Solway Firth
A number of 19th century records for Z. marina along the Solway Firth are listed in the BSBI database. The earliest record is from 1831 at Kirkcudbright Bay (BSBI, 2024).
According to wildfowlers, Zostera remained abundant in the Solway Firth despite the 1930s wasting disease epidemic (Berry, 1939).
During the late 1980s, concern was expressed regarding the impacts on sediments and Zostera beds from unregulated mechanical cockle harvesting, which had developed on an unprecedented scale (Perkins, 1988). Preliminary investigations revealed that intertidal Z. marina had been virtually eliminated from areas where dredging had occurred (Perkins, 1988).
These concerns prompted Scottish Natural Heritage to commission a survey of the larger Solway seagrass beds (Hawker, 1993, 1994), leading to prohibitions on boat dredging in 1992 and tractor dredging in 1994 (Southall & Tully, 2014). A managed hand-cockling fishery has opened and closed in recent years, based on stock levels and catch returns. The fishery is currently closed and would require a Habitats Regulations Appraisal to restart.
Recoveries of Z. noltii and Z. marina seagrass beds were detected in the Solway Firth between 1993/94 and 2022/23, with the total area of resurveyed beds recorded increasing by 68% (Carstairs, 2024; Table 9; Annex 2).
| Site | 1993/94 seagrass bed area (all species) (ha) | 2022/23 seagrass bed area (all species) (ha) | Increase (ha) | Increase % |
|---|---|---|---|---|
| Kirkbride Bank | - | 6.44 | - | - |
| Baldoon Sands | 325.24 | 535.93 | 210.69 | 65 |
| Skyreburn (East) | 9.97 | 17.66 | 7.69 | 77 |
| Skyreburn (West) | - | 11.34 | - | - |
| Airds Bay | - | 2.73 | - | - |
| The Dhoon | - | 1.85 | - | - |
| Manxman’s Lake | 27.24 | 57.26 | 30.02 | 110 |
| Auchencairn Bay | 122.07 | 196.51 | 74.44 | 61 |
| Orchardton Bay | - | 43.34 | - | - |
| Rough Firth | 29.71 | 57.99 | 28.28 | 95 |
| Carse Sands | - | 0.004 | - | - |
| Total | 514.22 | 931.05 | 351.12 | 68 |
Beds were found to have expanded into areas formerly harvested for cockles, such as the northern half of Baldoon Sands. Additionally, general improvements in water quality were considered to be the most likely factor facilitating bed expansion into other areas (Carstairs, 2024). This is supported by OSPAR data, which shows an overall decrease in nitrogen levels from 2001 onwards following implementation of the Water Framework Directive in 2000 (Scottish Government, 2020a). In 2008, water quality improved from moderate to good status at Manxman’s Lake, Auchencairn, Orchardton Bay, Rough Firth, Skyreburn and Airds Bay (SEPA, 2024). Furthermore, Carstairs (2024) found that the relationship between green algal cover and seagrass cover in the Solway Firth was inverse and statistically significant i.e. high algal cover was associated with low or no seagrass cover.
A small proportion of the bed increase was undoubtedly due to greater survey coverage during the 2022/23 surveys. It is also likely that differences in survey methodology accounted for a portion of the difference between the two sets of surveys with sparsely distributed seagrass plants mapped by Carstairs (2024).
At Skyreburn Bay, encroaching Sporobolus anglicus beds appear to have displaced 0.4 ha of sparse seagrass bed in the upper shore since 1994. Extensive baseline S. anglicus distribution data was collected during 2023 which will facilitate future comparisons (Carstairs, 2024).
Loch Ryan
A herbarium specimen of Z. marina was collected from Loch Ryan near Stranraer in 1833 (RBGE E01005004). Records for both Z. noltii and intertidal Z. marina were recorded at Stranraer in 1963 (BSBI, 2024). Loch Ryan was surveyed in 1988 by Howson (1989), who noted small Z. noltii beds at Wig Bay, a long, relatively narrow Z. noltii bed at Stranraer and subtidal Z. marina beds. Despite a paucity of historical data for the site, it is evident that the vast intertidal Z. noltii beds which now abound were heavily suppressed by long-term chronic water quality issues prior to 2014 (Carstairs, 2024). The distribution of subtidal Z. marina is also likely to have been restricted (Carstairs, 2024).
A substantial recovery of intertidal seagrass occurred in Loch Ryan between 2014 and 2022/23. The recovery coincided with two major infrastructure changes at Stranraer: the relocation of Irish ferry sailings from Stranraer to Cairnryan in November 2011 and the removal of the majority of existing wastewater discharges from Loch Ryan between 2011 and 2013.
The relocation of the Cairnryan ferry terminal to six miles further up the coast of Loch Ryan (to exploit the deep water at this location) reduced the need for dredging deep channels to Stranraer, decreased displacement waves generated by ferries, and minimised sediment disturbance caused by jet propulsion used in vessel manoeuvres (Carstairs, 2024).
The impact of these changes was not apparent in a 2013 survey conducted by Foster and Davidson (2018), though it should be noted that seagrass recovery can be a relatively slow process taking years, decades, or even centuries (Walker et al., 2006). However, Z. noltii is considered to be the most resilient of the Northern European species to environmental change and can exhibit relatively fast rates of recolonisation (Yonge, 1949; Rasmussen, 1977).
The total recorded area of intertidal seagrass beds in Loch Ryan increased from 10.48 ha to 79.21 ha between 2013 and 2022/23 (Carstairs, 2024; Table 10). The increase in the Stranraer bed is shown in Figure 30 & Figure 31.
| Site | 2013 seagrass bed area (all species) (ha) | 2022/23 seagrass bed area (all species) (ha) | Increase (ha) | Increase % |
|---|---|---|---|---|
| Wig Bay | 2.11 | 9.46 | 7.35 | 349 |
| Stranraer | 8.37 | 69.74 | 61.37 | 733 |
| Total | 10.48 | 79.21 | 68.73 | 656 |
The distribution and extent of the subtidal Z. marina beds has never been fully explored (Howson 1989). During the late 1980s subtidal Z. marina was located at McCulloch’s Point and at McMeckan’s Rocks, and south of Wig Bay near The Spit and near St Mary’s Croft (Crosby, 1989; Howson, 1989). A significant amount of washed-up material was recorded at Stranraer in 2022 (Carstairs, 2024) and the bed near The Spit was observed again in 2024 (De Lisle, 2024).
Summary
Seagrass beds in the Solway Firth have increased in size following cessation of mechanical cockle harvesting in the early 1990s. Seagrass populations may also have increased in relation to water quality improvements.
Seagrass beds in Loch Ryan have substantially increased in size following relocation of waste water discharges out of the loch and relocation of the ferry terminal away from the head of the loch between 2011 and 2013.
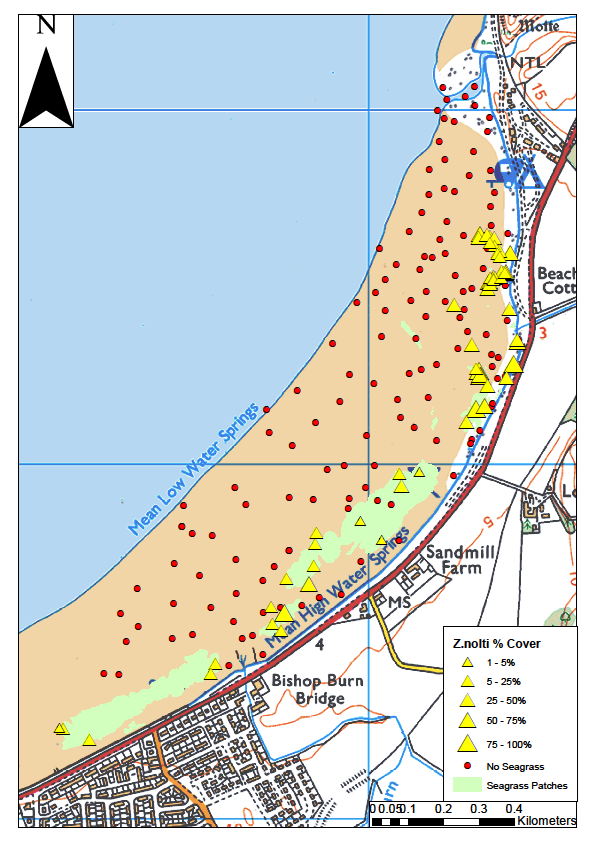
Figure 30. Zostera noltii data collected in 2013 at Stranraer (Foster & Davidson, 2018).
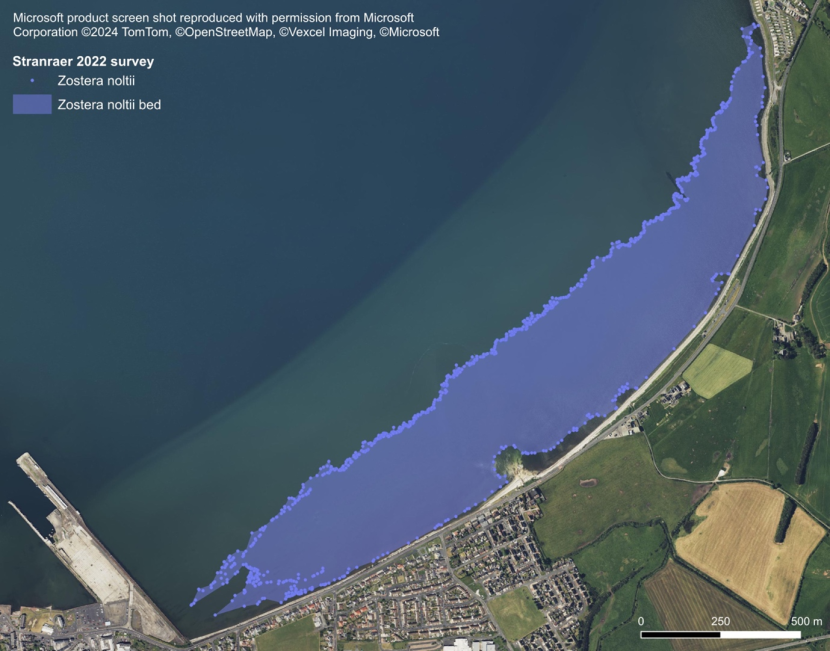
Figure 31. Zostera noltii bed at Stranraer surveyed in 2022 (Carstairs, 2024).
Clyde
Figure 32. Clyde Scottish Marine Region.
The Clyde SMR comprises the Clyde Estuary, the Firth of Clyde, and several sea lochs, including Loch Fyne and Loch Long (Figure 32). Z. marina beds were recorded in this region as early as 1813 (Hopkirk, 1813).
Hennedy’s Clydesdale Flora (1865) describes Z. marina as being “very common. In the sea and on banks at low water” and “Z. angustifolia” as “on mud banks and in saltwater ditches”. This appears to describe both subtidal and intertidal varieties of Z. marina.
Lee (1933) described Z. marina as “in the sea, often exposed at low tide; common”, and lists the plant as occurring in the following general areas (the text does not imply that seagrass was present throughout these areas): the portion of Ayrshire draining to the Firth of Clyde; most of Renfrewshire, excluding the portion subsumed into the City of Glasgow; Dumbartonshire; the Clyde Isles, including Arran, Bute, Inchmarnock, and Greater and Lesser Cumbrae; the eastern part of Cantyre draining towards Loch Fyne; and Kilbrannan Sound.
Inner Firth of Clyde (Glasgow to Bute and Great Cumbrae)
Distributional data from BSBI and GEMS databases (Annex 1) indicate that Z. marina has experienced a dramatic reduction in population distribution since the 19th century. Declines in the inner Firth of Clyde can be attributed to historical poor water quality which extended south into Ayrshire (Henderson & Hamilton, 1986; Hammerton, 1994).
Z. marina was listed for Renfrewshire by Watson (1837) and was reported from Langbank in 1885 (Watson, 2013), though the plant is now considered to be extinct in the county (Watson, 2013). Subtidal beds of Z. marina historically occurred in Fintry Bay and Kames Bay in Great Cumbrae (Batters, 1892) though no contemporary records were encountered during the current project.
Hopkirk (1813) described seagrass thrown on the shore at Helensburgh. Several 1990s BSBI records for Z. marina near Helensburgh indicate that beds may still occur in the upper Firth of Clyde (BSBI, 2024). However, details given for two of the five records state that the material has been washed up, and none of the records specifically describe plants growing in situ.
Beds of Z. noltii were reported in the Clyde estuary in 1974 though precise locations were not given (Minto, 1974; cited in Cleator, 1993). Small beds of Z. noltii occur along the shore at Cardross and Ardoch. In 2016, SEPA recorded that the total area of these beds was 10.8 ha with a coverage of 33% (Scottish Government, 2020b).The paucity of data available for these sites precluded assessment of whether the seagrass beds at Cardross and Ardoch have changed.
During 1855, Arnott reported an extensive bed of Z. noltii at Hunterston:
“Between Fairlie and Hunterston Point, covering hundreds of acres” (Hooker & Arnott, 1855).
While Hennedy (1865) added:
“On the opposite shores of Cumbrae not so common.”
The Hunterston Construction Yard was built on reclaimed areas of Hunterston and Southannan Sands in the 1970s. The marine construction yard and the coal terminal conveyor now extend beyond MLWS and sub-divide the sandflats and mudflats into three areas: Hunterston Sands, Southannan Sands and Fairlie Sands (Figure 33). These areas comprise Southannan Sands SSSI.
In 2016, SEPA reported that the total area of the Z. noltii beds found in the Southannan Sands SSSI was 19.3 ha with a coverage of 33% (Scottish Government, 2020b). It would appear that a huge decline of seagrass has occurred since Arnott’s 1855 account given that one hundred acres is equal to 40 ha. Although no maps of the bed exist prior to the 1970s, the declines can be attributed directly to land reclamation and associated changes in the hydro-morphological regime. Figure 33 shows the 2016 beds, together with the present-day Ordnance Survey (OS) MHW superimposed over an OS map published in 1960, to show the reclaimed area which amounts to 130 ha or 321 acres.
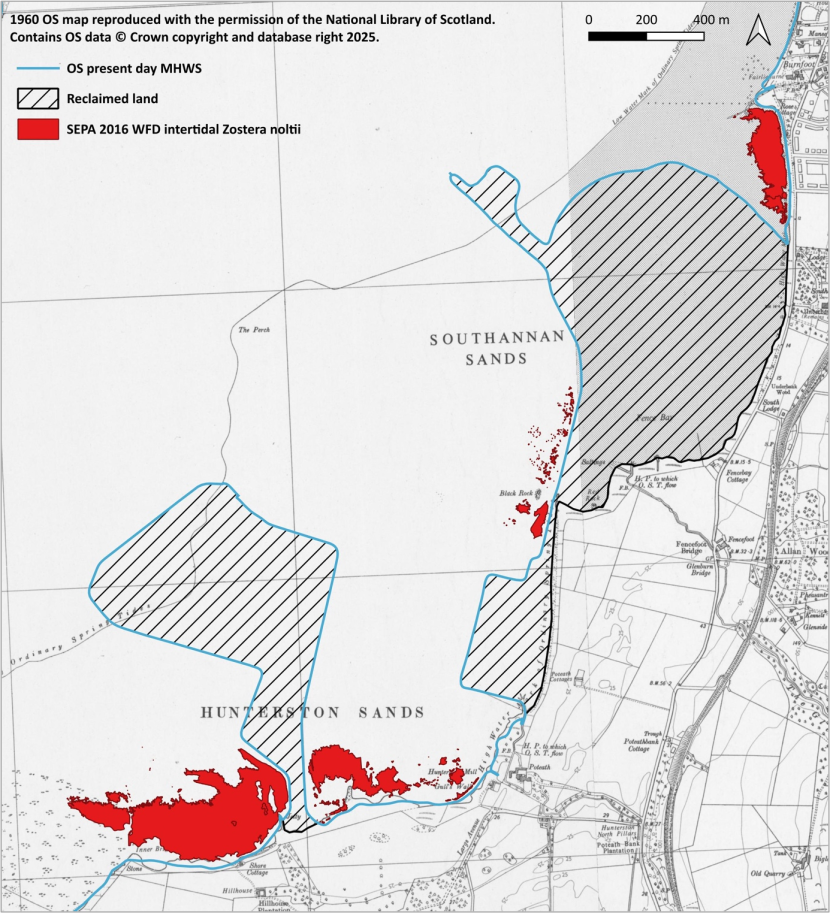
Figure 33. Reclaimed area at Hunterston and Southannan Sands and SEPA 2016 Zostera noltii beds.
More detailed surveying of intertidal and sub-tidal habitats in the upper Firth of Clyde between Glasgow and Bute may reveal the presence of additional beds. However, whilst limited comparative data makes a quantitative assessment difficult, there appears to have been a serious decline of both Z. marina and Z. noltii.
Clyde sea lochs
The sea lochs of the northern part of the Clyde Sea are a series of long, deep fjordic lochs with generally steep rocky sides and relatively little shallow mudflat or sandflat habitat suitable for Zostera. Watson (1837) documented the occurrence of Z. marina in Loch Fyne at Inverary. Graham (1890) noted “a rich pasturage of seagrass” at Ardrishaig in Loch Gilp. The MNCR surveys recorded sparse Z. noltii in sandy areas at the head of Loch Goil and Z. noltii in areas of standing water over muddy sand at the head of Holy Loch (Dipper & Beaver, 1999). Z. noltii was also recorded in Holy Loch in 1996 and 2022 (BSBI, 2024). It is not possible to assess whether the Z. noltii bed at Holy Loch has changed in extent or remained stable.
The MNCR surveys did not report the presence of Z. noltii in Loch Gilp (Dipper & Beaver, 1999), however a Z. noltii bed at Lochgilphead surveyed in 2023 was found to be 19 ha in size (Knight & Holden, 2024).
Loch Riddon flows into the Kyles of Bute which flow down the west and east of the island of Bute into the Firth of Clyde. The River Ruel flows into the head of the loch, which is very sheltered, and this has allowed the development of the extensive Ruel estuary, a rarity on the west coast of Scotland. The estuary is designated as a SSSI with “dwarf eel-grass” (Z. noltii) mentioned in the SSSI citation. The MCNR Clyde sea loch survey did not record Z. noltii in Loch Riddon, however, there are records from 1911, 1970 and 1997 in the BSBI database (BSBI, 2024). No extent data exists for this area, so it is not possible to assess whether the Z. noltii bed in Loch Riddon has changed in extent or remained stable.
In a report of the Fishery Board for Scotland (1890), Thomas Scott, one of the naturalists of the board wrote:
“I obtained one specimen of this species in the vicinity of Skeirvuie—a small island near the head of East Loch Tarbert (Loch Fyne)—where Z. marina grows in considerable abundance.”
No current records were found for this location.
Z. marina was historically recorded in the parish of Kingarth on Bute (Ballantyne, 1911). The site location is not given, though Kilchattan Bay would appear be a strong candidate and worthy of a survey. The plant has not been recorded on Bute since (Hannah, 2019).
Outer Firth of Clyde (Arran, east coast of the Kintyre peninsula and Ayrshire Coast)
A herbarium specimen of Z. marina was collected at Brodick, Arran, in 1847 (RBGE E00762039). Bryce (1859), in a list comprising “only the rarer plants of Arran”, described Z. marina as “abundant”. Beds of Z. marina were reported to occur at:
“Lamlash, on the sandy shore opposite the village; also, on Holy Island, not far from St. Molios’s cave”
This description is augmented by three historical records collated by Cowley (2012). These are: Norman, 1855, “seagrass beds near Lamlash Pier”; Miles, 1856, “beds at Lamlash Bay and Holy Isle”; and Robertson, 1896, “beds exposed at low tide in many spots at Lamlash”. Bryce later described extensive seagrass meadows in “sandy districts” (Bryce, 1872).
Z. marina has been found in Lamlash Bay by Seasearch surveyors in 2003 (Duncan, 2003), 2016, 2017, 2018 and 2020 (GeMS).
Scattered Z. marina beds occur within the dyke swarm (parallel linear igneous rock features) along the south coast of Arran between Kilmory and Kildonan. The seagrass beds were surveyed in 2012 and 2016 (Binnie et al., 2012; Thomas, 2016) with a large decline reported between the two surveys.
The 2012 and 2016 surveys did not cover the exact same area and neither survey covered the entire coastal area containing suitable seagrass habitat. However, where surveying did occur at the same places the recorded seagrass areas were significantly smaller in 2016.
The frequency of GPS point capture varies immensely between the two surveys with relatively few points taken during 2012. This affects accuracy and may signify differences in how isolated patches were mapped and bed edges defined. Therefore, there are potential issues with data comparability.
Thomas (2016) recorded extensive Sargassum muticum and concluded that it had recently proliferated at the site and was responsible for substantial seagrass declines as the seaweed was not mentioned in the 2012 survey report. However, it is uncertain if Sargassum muticum was already present at the site in 2012, as it has been present at locations around Arran since at least 2007 (Davison, 2009).
Subtidal Z. marina in Whiting Bay has a patchy distribution but extends for around 4 km and forms the largest known Zostera bed in the Clyde (NatureScot, 2024). Seagrass has also been recorded off Pirnmill to the northwest of the island. Two small subtidal Z. marina beds have recently been recorded at Corrie on the east side of Arran with unconfirmed reports of beds south of Corrie and around the King’s Cave area (L. Kay, pers. comm.).
The Community of Arran Seabed Trust (COAST) stated that seagrass beds at Pirnmill and Whiting Bay have long been subject to damage from dredging activity which had limited their extent (COAST, 2012a, 2012b). The historical literature also indicates substantial declines have occurred at Lamlash Bay where dredging damage to seagrass was observed in 2009 (H. Wood, pers. comm.). Only two small patches of seagrass have been recorded in recent years, whereas in 1896 Robertson reported seagrass at many spots which extended into the lower intertidal zone (Cowley, 2012; Łękawska & Kay, in prep.). Furthermore, seagrass no longer occurs on the sandy shore at Lamlash as described by Bryce (1859).
Elsewhere in the Outer Firth of Clyde, there are historical records of Z. marina from 1932 and 1961 at The Dhorlin at the mouth of Campbeltown Loch. The MNCR surveys of sea lochs in the Clyde Sea reported that Campbeltown Loch was not surveyed (Dipper & Beaver, 1999), and no contemporary records were found. There has been an anecdotal report that seagrass declines in the Skipness Bay, Kintyre have been caused by scallop dredging (Wood, 2006).
There are two historical records of Z. marina at Dunure, Ayrshire from 1961 and 1968 (BSBI, 2024). A bed of Z. marina occurs in the mid-upper intertidal zone at Dunure (Carstairs, 2024). The bed was present during visits made in 2022, 2023, and 2025 with no apparent visual change in distribution (M. Carstairs, pers. obs.). Basic mapping was conducted in 2023 (Figure 32) and seagrass was found to occur within an area of 1 ha. Z. marina occurred extensively in large permanent pools though was absent from substantial areas within the polygon containing exposed bedrock and boulders. The patchy nature of the bed means that seagrass coverage was much less than 1 ha. The bed is sheltered by a rock platform supporting occasional lines of boulders along the lower shore which dissipate wave energy (M. Carstairs, pers. obs.). S. muticum grows in pools at the lower edge of the bed (Figure 11) with a few small plants in pools at the upper shore. Mapping was not undertaken during the 1960s, so it was not possible to determine whether the extent of the bed has changed.

Figure 34. Intertidal Zostera marina bed at Dunure, Ayrshire in 2023 (Carstairs, unpublished).
Summary
Z. marina has historically declined in the inner Firth of Clyde and does not appear to have recovered. These declines are likely to be largely attributable to poor water quality within the Clyde (Henderson & Hamilton, 1986). The Z. noltii population at Southannan Sands SSSI most likely experienced significant declines in relation to extensive land reclamation in the 1970s.
Historical seagrass declines have occurred on Arran at Lamlash Bay and suppression of seagrass extent by bottom-towed fishing gear has been reported at Pirnmill and Whiting Bays. Similarly, damage from bottom towed gear has been reported off Kintyre. Further monitoring is required to assess any impacts of S. muticum particularly within the intertidal Z. marina beds at Kildonan, Kilmory and Dunure.
Argyll
Figure 35. Argyll Scottish Marine Region.
The Argyll SMR spans from the island of Coll in the north to the Mull of Kintyre in the south and has a diverse coastline containing numerous islands and sea lochs (Figure 35). There are many sheltered bays and inlets in this region in which seagrass beds occur.
West coast of the Kintyre peninsula
Cunningham & Kenneth (1979) commented that subtidal Z. marina has historically been recorded from Machrihanish and the south east shore of Gigha and that intertidal Z. marina has been recorded near the head of West Loch Tarbert. Z. noltii has also been recorded from West Loch Tarbert (Cunningham & Kenneth, 1979).
The MNCR sea loch surveys located beds of Z. noltii at the head of West Loch Tarbert in 1989 (Dipper et al., 2008). In 2022 the loch was found to contain seven Z. marina beds covering an area of 31 ha (Knight & Holden, 2022). A Z. noltii bed at Crinan was 13 ha in size (Knight & Holden, 2022).
Extent was not recorded during historical surveys, however contemporary records show that seagrass beds still cover large areas around Kintyre.
Sound of Jura
Z. noltii was recorded at the head of Loch Sween (RBGE E00762107) and both varieties of Z. marina off the shore of Loch Sween near Kilmichael of Inverlussa in 1899 (Salmon, 1900; RBGE E00762064). Cunningham & Kenneth (1979) commented that subtidal Z. marina was usually seen in drift and gave the following locations for historical occurrence: Loch Stornoway; Loch Coalisport; Barrahormid; and Danna. Z. noltii has been recorded from the Fairy Isles, Ulva, and Danna (Cunningham & Kenneth, 1979).
The MNCR study of lagoons found subtidal Z. marina in Craiglin lagoon, off Loch Sween. The report noted previous studies suggesting the fluctuating nature of the seagrass population. Z. marina was reported to be abundant adjacent to the shore in 1982 but was replaced by green algae by 1985. Zostera was reported to be absent in June 1993 but abundant in May 1994 (Covey et al., 1998).
The MNCR study of sea lochs in the west of Scotland reported patches of Z. noltii at the head of Loch na Cille and beds of Z. marina in the Loch Sween complex within Linne Mhuirich, Caol Scotnish and off Castle Sween (Dipper et al., 2008). In particular:
“Linne Mhuirich is also of interest for the extensive and well-developed beds of seagrass Z. marina (Zmar) which occur throughout its length, in shallow water of 3 m depth or less. The largest beds occur at the southern end, in the shallow water to the south of the rapids and a short distance to the north of them. Dense Zostera beds also occur at the head of Linne Mhuirich, continuing southward on the western shore to below Tighavullin. Further beds occur just beyond the main basin where it narrows to form the inner basin.”
Loch Craignish lies on the mainland coast at the northern end of the Sound of Jura, in an area of low hills. It is a complex fjordic sea loch which is now the subject of a seagrass research and restoration project led by Seawilding. Several seagrass beds were mapped by them during 2022, with a total area of 5.25 ha of Z. marina and 0.25 ha of Z. noltii (Knight & Holden, 2022). Anecdotal evidence obtained from community members indicated that seagrass historically occurred much more extensively within the loch (Knight & Holden, 2022). Restoration work within Loch Craignish has not been successful in many areas of the loch, which has been attributed to the presence of anoxic sediment (Seawilding, 2025).
Larger historical quantities of seagrass were reported for Dunvullaig Bay in Loch Craignish, while beds were described as being extensive in "the lagoon". A family of recreational swimmers, snorkelers, and divers who purchased a lochside property in 1994 subsequently witnessed a significant decline in the environmental and ecological quality of the loch. They attributed the degradation to dredging (Adkins, 2023) but the report does not specifically mention that dredging occurred through seagrass beds.
Seagrass populations were also reported to have occurred in the adjacent Loch Beag, where seagrass is currently considered absent (Knight & Holden, 2022). The authors estimated that the potential historical abundance of seagrass beds may have been up to 92 ha in Loch Craignish and Loch Beag. This assessment was undertaken using data from habitat suitability modelling, as well as sediment core and eDNA analysis (Knight & Holden, 2022).
Jura
Sublittoral Z. marina was recorded off the northeast coast of Jura in 1957 by Knox (1959) at Eilean Buidhe Mhor, north of Ardlussa and Eilean a’Bhorra, near Lussagiven. The notes for a herbarium specimen (RBGE E00762005) collected from the latter location state:
“Sandy substrate, depth 4ft. at L.T. Forming large spreading pure colonies in centre of straits. Many epiphytic algae on stems.”
The MNCR study of sea lochs in the west of Scotland reported Z. noltii from the shallow freshwater-influenced intertidal inner basin of Loch Tarbert which almost bisects the island of Jura (Dipper et al., 2008).
Islay
Z. marina has been recorded at the head of Loch Indaal (Ogilvie, 1992). Further down the loch, one Z. marina bed surveyed in 2014 via snorkelling at Gartbreck was estimated to cover 0.61 ha (Courcier, 2024). Elsewhere on Islay, Z. marina meadows were noted at Kildalton (0.04 ha) and at Port Ellen (1.21 ha) (Courcier, 2024).
Z. noltii has been recorded on the Loch Indaal flats north of Bowmore (Ogilvie, 1992; Project Seagrass, 2024; GeMS) and also Carraig Fhada and Loch an t-Sailein (Ogilvie, 1992, 1995).
The MNCR study of sea lochs in the west of Scotland reported extensive beds of Z. marina growing on the clean sands in the sheltered northern half of Loch Indaal, which were surveyed in 1982 (Dipper et al., 2008). It was noted that there was a sewage outfall at Bowmore discharging untreated effluent directly into the loch.
Ogilvie (1992) also highlighted potential sources of pollution, including a few small watercourses flowing through agricultural land before entering the loch. Pollutants affecting watercourses on Islay included silage liquor, sheep dip, wastes from farm byres and milking parlours, and run-off from fertilised fields (Elliot, 1989). The waste products from two local distilleries were considered to rival those from sewage in terms of their impact on the loch (Ogilvie, 1992). The author described the distillery discharges as a “mix of nitrates, nitrites, and phosphates together with particles of crushed barley”. A few years prior to 1992, one of the distilleries ceased discharging waste directly into the loch (Ogilvie, 1992).
Changes in Zostera abundance and overwintering wildfowl populations have been noted on separate occasions in Loch Indaal. Brent geese wintered in the loch until 1938 (Elliot, 1989). Observations of an abundant population were made in 1878 (Scot-Skirving, 1878) and 1886 (Elwes, 1930). Elliot (1989) attributed the disappearance of overwintering brent geese to the impact of wasting disease on Zostera populations.
Ogilvie (1992) reported extensive Z. marina beds at or below low water mark in Loch Indaal and noted that substantial increases in the amount of Z. marina beach cast had occurred in recent years. In addition, large patches of Z. noltii growing on intertidal sandflats were discovered in 1991. This species had not previously been recorded in the loch and was only known to occur in a single bay in the southeast of the island. An increase in wigeon numbers coincided with the observed increases in Zostera, while numbers at neighbouring Loch Gruinart remained stable over the same period (Ogilvie, 1992). Ogilvie later reported that up to around 20 light-bellied brent geese often overwintered in Loch Indaal, with ones and twos present in flocks of other bird species (Langsbury & Ogilvie, 2006), and that 40-50 now winter in the inner parts of the loch (M. Ogilvie, pers. comm.).
Observations of concomitant increases in seagrass beach cast and overwintering numbers of wigeon and brent geese suggest that the seagrass population at the head of the loch is increasing, however, a lack of comparative data precludes definitive conclusions.
Scallop dredging has been reported to have historically damaged Z. marina beds in Loch Indaal, leaving lines of bare sediments in the beds where gear had been towed (Waring, 2024).
Colonsay and Oronsay
Seagrass features conspicuously in the history of Colonsay and Oronsay. Eilean na Bilearach, which in Gaelic means Island of Seagrass, lies off the Ardskenish peninsula in south-west Colonsay (Byrne, 1993). A local seagrass tale comes from the narrowest point between Colonsay and Oronsay, known as The Strand. The islands are connected by dry land at low tide though Am Poll Gorm ‘the green pool’ does not drain and is said to have contained seagrass. John Hart is reputed to have been in the habit of swimming The Strand at high tide to visit his sweetheart on Oronsay but eventually became entangled in seagrass and drowned (King & Scammel, 2017).
McNeill (1910) provided the following description of Z. marina:
“Abundant growing where sediment has been deposited in the sea and frequently washed ashore. The long grass-like leaves, when dried, are used for packing and for stuffing mattresses.”
There are records of Z. marina around Colonsay and Oronsay from the 1930s onwards with beds located at the Ardskenish peninsula, The Strand, Liath Eilean and Port na h-Àtha (BSBI, 2024, Unsworth et al., 2025).
North Argyll sea lochs
William Anderson Smith wrote extensively about the flora and fauna of the north Argyll sea lochs in the late 19th century. Originally published as serialised newspaper articles, his writings were subsequently compiled into two books, one on the district of Benderloch and one specifically on Loch Creran, where he lived. He gave many interesting descriptions of sometimes quite extensive Z. marina beds in the Benderloch area and although he often neglected to state which sealoch the accounts were from, most appear to be from Loch Creran.
For example, Smith (1882) reported that in September 1879 “At the head of Loch Creran, ere entering the narrows to the inner loch, is a well-sheltered sandy bay. This is covered with Zostera marina, even far above low water of ordinary tides, the ground under it being unstable. Such an instance is very rare, as it is always considered to be a very low tide that shows the sea-grass”. This appears to be a reference to a partly intertidal seagrass population.
Smith also observed a bed of Z. marina in Loch Creran in March 1882, at least partly exposed at low tide, which extended for “several hundred yards” (Smith, 1887) and which contained serpulid reefs.
During a survey of the loch for the MNCR review in June 1989, a dense bed of Z. marina was found on the north shore, east of North Ardnaclach (Dipper, 2008). In November and December 1994, Moore et al., (1998) surveyed Loch Creran for serpulid reefs and reported three seagrass beds along the north shore – one along the south edge of the spit near South Ardnaclach, one east of North Ardnaclach in the same location as the MNCR bed, and one further north, below Invernahyle.
The last reported sighting of Z. marina in Loch Creran was made in 2002 by Colin Moore (Scottish Government, 2020c; C. Moore, pers. comm.). Seagrass has not subsequently been recorded during a range of surveys on the loch including an excursion in 2019 undertaken on a low spring tide along the north shore from Creagan well past the westernmost record of a seagrass bed (C. Moore, pers. comm.).
Therefore, it would seem that the seagrass population in Loch Creran has declined severely to the point that it may well have become extinct (C. Moore, pers. comm.).
Mull and Iona
Bangerter & Cannon (1978) collated several subtidal Z. marina records from Mull including one from 1877 from Dòirlinn a’Chailbhe (between Mull and Calve Island), one from Loch Mingary from 1879, and others from the north end of the Sound of Iona, Fionnphort, Sgeir na Cille, Inch Kenneth, South Ardmeanach, Carsaig and Loch Buie. Millar (1993) reported subtidal Z. marina at Sligneach shore, Iona. Z. marina was recorded again at Dòirlinn a’Chailbhe in 1983 during the UCS Mull sublittoral survey (GeMS). There are more recent records of subtidal Z. marina near Fionnphort from 2016 (GeMS; BSBI), with the GeMS SeagrassSpotter record showing a lush seagrass bed present (Unsworth et al., 2025).
Contemporary records of Z. marina also exist at Port Uisken (as recorded on seagrass spotter in 2017; GeMS), and off the Treshnish Isles (as recorded in 2016 during a Seasearch survey; GeMS).
Dipper & Naylor (2025) reported sublittoral beds of Z. marina from Little Colonsay (undated) and Ulva (2016) with the latter growing in maerl gravel and becoming partially exposed at extreme low tide.
Intertidal Z. marina was recorded at the north east side of Loch Don in 1966 (RBGE E00762049; Bangerter & Cannon, 1978) and Lochdonhead (Bangerter & Cannon, 1978).
Tiree and Coll
Mitchell et al. (1983) reported that Z. marina grew in coarse sands off Tiree. Pearman & Preston (2000) reported that there are no known populations of Z. marina in situ although there are occasional records of washed-up material from many beaches.
Summary
An assessment of population change was not possible due to lack of comparative data. However, the number of records, both historical and contemporary, suggests that there are many locations that could be favourable for seagrass beds. Further surveys in this area would be valuable.
West Highlands
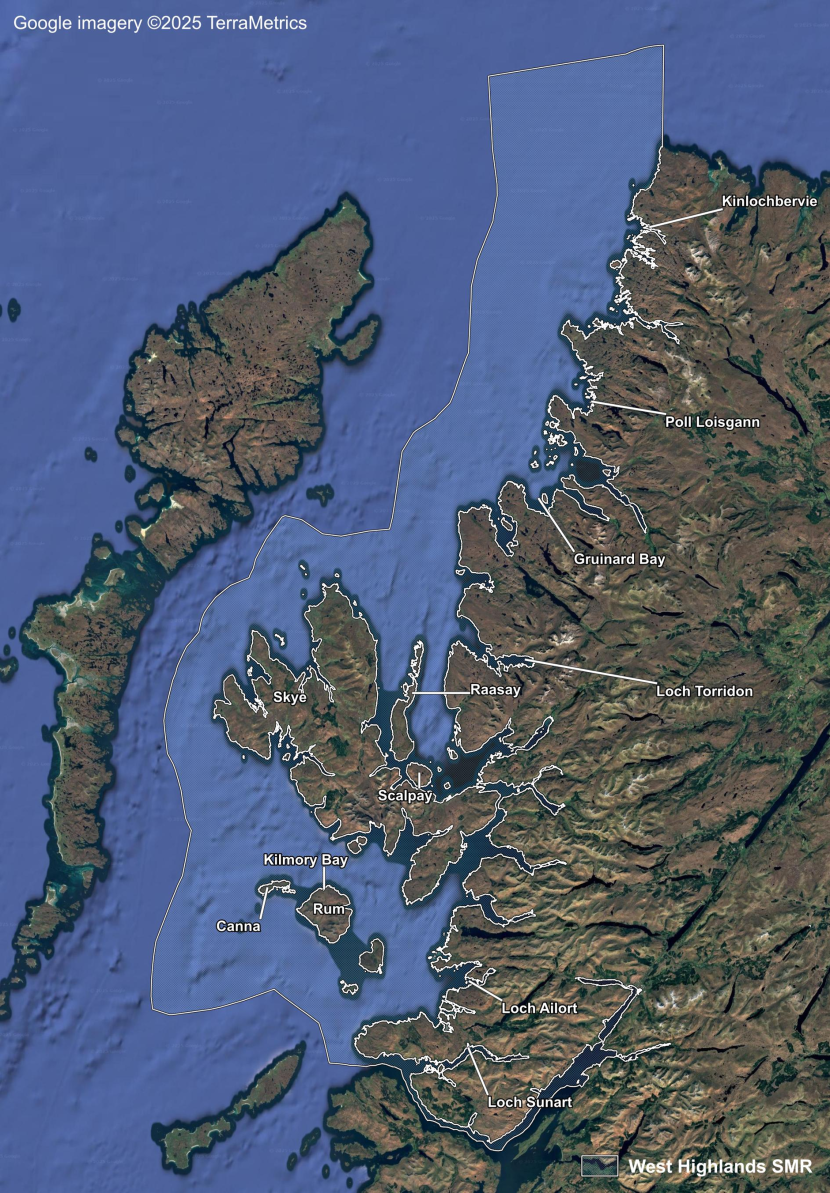
Figure 36. West Highlands Scottish Marine Region.
The West Highlands SMR extends from Cape Wrath in the north to the Sound of Mull in the south and has a diverse coastline containing numerous islands, inlets, bays, lagoons and sea lochs (Figure 36). Seagrass beds are found in numerous places in this region. The main points of interest are discussed below roughly in order from south to north, using the historical county names where appropriate.
Inverness-shire
Loch Sunart is a narrow fjordic sea loch, the second-longest in Scotland after Loch Fyne. The MCNR study of sea lochs in the west of Scotland mapped small beds of subtidal Z. marina at the mouth of the loch and at the head of Salen Bay in 1995 (Dipper et al., 2008). In 2006, a small 0.03 ha bed of Z. noltii was mapped further upstream near Eilean Mòr during SNH site condition monitoring (GeMS).
The MNCR study of sea lochs of north-west Scotland reported varying amounts of Z. marina in several of the lochs of the mainland coast (Dipper et al., 2002). One of the most apparently suited to Zostera is Loch Moidart which contains some of the largest areas of littoral sediments found in Scottish sea lochs. Almost the whole of the inner loch from around Eilean an Fheidh eastwards consists of fine muddy sandflats. Much of the upper loch consists of intertidal mud and sandflats and the intertidal area makes up 75% of the loch. This is in contrast to the majority of Scottish west coast sea lochs, which mostly have very small intertidal areas. Z. marina was recorded but not mapped (Dipper et al., 2002). This area should warrant further investigation, given the large expanse of intertidal muds and sandflats which Zostera species favour.
Dipper et al. (2002) noted that “Some of the first installations and research into fish farming began in Loch Ailort” and reported extensive fish-farming operations within the loch. Sparse Z. marina along the north side of the loch to the west of Peanmeanach Bay were reported from the surveys (Dipper et al., 2002). In 2003, 2014 and 2016, site condition monitoring surveys of Sound of Arisaig SAC found several Z. marina beds on sandbanks in lower Loch Ailort (GeMS). No comparative data for extent exists in Loch Ailort, meaning that an assessment of population change is not possible.
Scattered patches of Z. marina were recorded in Loch nan Uamh (Dipper et al., 2002), however the site has not been revisited.
One of the shortest and shallowest sea lochs in the area, Loch nan Ceall, lies in the Arisaig area to the north-east of the Ardnamurchan peninsula. Z. marina was reported in sheltered sandy inlets amongst the skerries, including on the north coast of Luinga Mhor (Dipper et al., 2002).
Rum and Canna
Mitchell et al. (1983) reported that Z. marina grew in coarse sands of the east side of Rum. Heslop-Harrison (1958) indicated Z. marina was present in the vicinity of Kilmory Bay on Rum. It was recorded in 1986 but was not re-found in 2004, despite a botanist swimming in the bay as part of the search (Pearman et al., 2008). Professor J.W. Heslop-Harrison’s seagrass records are included in this report following guidance in Pearman & Walker (2004).
In 2010, during a SNH survey of the Sound of Canna, subtidal Z. marina beds were found in Kilmory Bay and in a sheltered bay off Rubha Carr-innis on the island of Canna (Howson et al., 2012). Z. marina had been previously found off the south of Canna in 1993 (GeMS). The survey locations were not the same so could not be compared.
Subtidal Z. marina beds have recently been recorded in the Galmisdale area of Eigg and the species was also recorded near the harbour during the late 19th century (Macvicar, 1897). The record relates to intertidal plants though these may have represented the upper parts of an otherwise subtidal population.
Skye, Raasay, Scalpay and Longay
A herbarium specimen of Z. marina was collected in Skye in 1958 at Fiskavaig Bay, below Aird an t-Sabhail (RBGE E00762031). Z. marina has been reported growing in coarse sands in the south of Skye (Mitchell et al., 1983). The plant has been recorded in a number of sheltered sea lochs though is generally absent from the northern parishes of Kilmuir, Snizort, and the far west coast of Durnish (BSBI, 2024).
Loch Ainort on the east coast of Skye is a small, sheltered sea loch. Z. marina was recorded in patches of inshore muddy sand on the southern side of the loch (Dipper et al., 2002). Scattered patches of Z. marina were recorded in Loch Slapin and Loch Brittle (Dipper et al., 2002). A photograph of Z. marina at Ord in Loch Eishort is present in Baxter et al. (1999). Z. marina has been recorded in several locations within and around Lochs Eishort and Slapin between 2014 and 2019 by SNH, Seasearch and the South Skye Seas Initiative (GeMS).
In 2020 Z. marina was reported from Loch Bracadale on Skye, and a small bed of Z. marina was reported from a bay on the island of Soay, off the south coast of Skye (GeMS).
Heslop-Harrison (1937) reported that Z. marina was uncommon though occurred on Raasay, above Balachuirn at the mouth of the burn, on the shores of Longay, and at the mouth of Allt Liath on Scalpay. There is a record of a 4 m x 4 m patch of Z. marina off North Fearns at the south end of Raasay from March 2024 (BSBI, 2024).
Wester Ross
Z. marina has been recorded at a number of sites including Red Point off South Beach, Loch Broom, Loch Carron, Gruinard Bay, Loch Torridon, Morar, Gairloch Bay and Loch Ewe (Druce, 1903, 1904, 1929; D. Donald, unpublished data).
Lochs Carron and Kishorn form a sea loch complex that opens into the Inner Sound between Skye and the Wester Ross mainland. Dipper et al. (2002) reported that:
“The sandy areas of Loch Carron west of Plockton … support beds of eelgrass Z. marina in shallower water (Zmar). Zostera is present at around 1 m depth inshore of A' Ghlas-leac and, like maerl, may be common along this coast in suitably sheltered areas.”
The mouth of Loch Torridon opens into the northern end of the Inner Sound. A patchy but extensive bed of Z. marina was reported off the north shore, south-west of Inveralligin, and from Ob Mheallaidh, a shallow sheltered lagoonal inlet on the south side of Upper Loch Torridon. Sublittoral Zostera was also located in fine muddy sand in Loch Shieldaig, an offshoot of Loch Torridon (Dipper et al., 2002).
Scattered patches of Z. marina were recorded in Loch Alsh and Loch Gairloch (Dipper et al., 2002). Z. marina was recorded from the north shore of the mouth of Loch Gairloch in 2010 during SNH benthic surveys (GeMS), suggesting that the beds were still present in this area. This is supported by observations that masses of fragments with roots were washed up in 2024 at Gaineamh Mhòr (D. Donald, unpublished data).
An intertidal population of Z. marina was discovered in 2007 at Loch Ewe near Midtown at NG 823 851 however this population appeared to succumb to severe frost during the exceptionally cold winter of 2010 and has not been recorded since (D. Donald, pers. comm.). An offshore bed has since been discovered near Mellon Charles (D. Donald, unpublished data).
Four Z. marina beds were mapped in 2004 during surveys of strategically chosen locations between Greenstone Point in Gruinard Bay and Sheigra at Kinlochbervie (James, 2004). A number of locations were not visited due to adverse weather conditions, though beds were recorded off Second Coast and a shallower bed further east within Gruinard Bay at Fraoch Eilean Mòr (James, 2004). Z. marina appears to still be present in Gruinard Bay where it was recorded in 2010 during SNH benthic surveys (GeMS), in 2020 by Project Seagrass (GeMS), and in subsequent years including 2025 (D. Donald & P. Cunningham, pers. comm.).
James (2004) reported apparent losses of two seagrass beds in Enard Bay, Loch Garvie. These beds had previously been located in Poll Loisgann, a small, sheltered sea loch, and at Cùil Lochain in a coastal embayment further north (White, 1987). The reasons for the disappearance of these two beds is unclear; however, the loss in Poll Loisgann, in particular, may have been linked to a development of new fish farm, which became operational in 1992. The fish farm was located approximately 100 m from the seagrass bed (James, 2004; White, 1987). The initial Atlantic salmon stocking biomass of 75 tonnes within Poll Loisgann was increased to 275 tonnes in 1995, which could be anticipated to have an adverse effect on seagrass within the small, sheltered sea loch (James, 2004).
West Sutherland
A bed of Z. marina was located at Port Dhrombaig in 2001 (Evans et al., 2002). Two small Z. marina stands were recorded within Loch Dhrombaig (James, 2004). Subtidal Z. marina was reported from the same place in 2020 by Project Seagrass (GeMS).
Subtidal Z. marina was recorded at Oldany Island during 1955 (Anthony, 1976).
Summary
An assessment of population change was not possible in many locations due to lack of comparative data. In Wester Ross, two subtidal Z. marina beds in Enard Bay, Loch Garvie appear to have been lost between 1987 and 2004. The disappearance of a bed at Poll Loisgann may have been linked to a development of a new fish farm. In other places, both contemporary and historical records exist, suggesting that populations may be stable.
The number of historical and contemporary records in this area, suggests that there are many potentially favourable locations for seagrass beds. Further surveys would be valuable.
Outer Hebrides
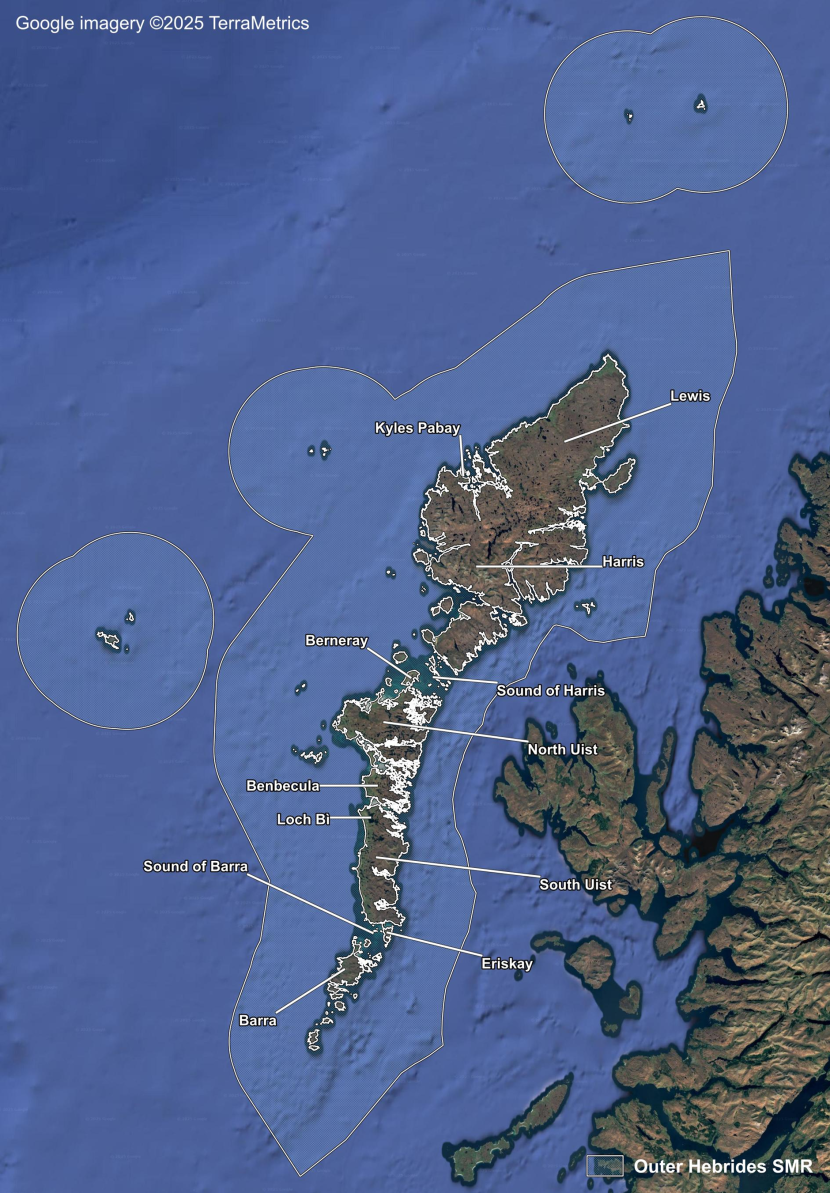
Figure 37. Outer Hebrides Scottish Marine Region.
The Outer Hebrides SMR covers the Outer Hebrides Archipelago (Figure 37). The Outer Hebrides have an extensive shallow continental shelf extending westwards into the Atlantic with deeper water in The Minch which lies to the east. Seagrass is absent from the northern shores of Lewis which are relatively exposed and characterised by a fairly uniform coastline. Seagrass beds are predominantly distributed along the western seaboard especially in lagoons and sheltered bays. Seagrass also occurs in sounds which lie in a north west to south east direction between the islands. Anecdotal accounts from wildfowlers in the Outer Hebrides indicate that the 1930s wasting disease badly affected some Z. marina populations but not others (Berry, 1939).
Great Bernera
In 1875, Smith described the substantial subtidal beds in the sound between Great Bernera and Little Bernera:
“As we proceed to the narrowest part between abrupt cliffs, with just room enough to enable a good boat to pass, under our keel may be observed extensive beds of Zostera marina, covering that portion of the sound bottom that lies opposite the snug, half-concealed, little farmhouse of the kindly King of Bernera.”
There is a record of Z. marina growing in this area in 1982 (BSBI, 2024).
Sound of Harris
Atkinson-Willes (1963) reported that the Z. marina beds at Ensay and Killegray in the Sound of Harris were as abundant as they had been 20 years earlier. The sound contained flourishing populations in many places, which were considered to be unaffected by the wasting disease epidemic.
More recently, well developed Z. marina beds were noted during an extensive survey of the Sound of Harris in 2005 which estimated the extent of the beds to be ~280 ha (Malthus et al., 2006). The report noted:
“The beds typically take the form of a series of bands of dense Zostera alternating with bands of open sand with each band being several metres wide. The sward height of the Zostera is typically 1–1.5 m and the plants are densely spaced to give continuous coverage within the band.”
Z. marina beds were also recorded within the Sound of Harris during a MarPAMM survey in 2021 (GeMS). As no historical records of extent exist, it is not possible to do a quantitative assessment of population changes.
Berneray
Andrew Rodger is currently assessing beds south of Berneray and has reported a >50% (180 ha) reduction in area over the last 34 years (pers. comm.). Potential causes of decline include suction dredging of razor clams (Tuck et al., 2000) and the construction of a causeway between Berneray and North Uist.
Sound of Barra
In 2006, a survey of the Sound of Barra was carried out by Harries et al. (2007b). Z. marina was estimated at 360 ha. The report found similar patterns to those in the Sound of Harris:
“The beds typically take the form of a series of bands of dense Zostera alternating with bands of open sand with each band being several metres wide. Within a band the Zostera fronds are densely packed and the sward height can be ~1.5 m in well-developed beds. The Zostera rhizomes act to stabilise the mobile sand and at some locations the sediment surface of the Zostera bands is several cm higher than the adjacent areas of loose sand.”
In 2015, diver surveys of six seagrass beds in the Sound of Barra showed the beds to be locally dense. Previously known beds were all re-found but seagrass populations at sites to the east of the Eriskay Causeway appeared to have declined presumably due to a change in water flows and increased sedimentation following construction of the causeway in 2001 (Bunker et al. 2018; NatureScot, 2025).
Outer Hebrides sea lochs
One Zostera bed was found during the MNCR surveys of sea lochs in the Outer Hebrides in the 1980s although large areas remained unsurveyed (Beaver & Dipper, 2002). These areas are exposed or very exposed with sheltered areas towards the heads of lochs and there is a wide range of seabed depth and substrate character. A healthy bed of Z. marina was reported at Tràigh na Beirgh, Kyles Pabay, West Loch Roag in 1982. Recent surveys in 2020 and 2024 by Project Seagrass have confirmed that a Z. marina bed is still found within this site.
Outer Hebrides lagoons
The lagoons of the Outer Hebrides are much more suited to Zostera than the sea lochs, being more sheltered and generally very shallow with only a few deeper than 5 m (Thorpe et al., 1998). A herbarium specimen of the intertidal variety of Z. marina was collected from Loch an Strumore in 1894 (Bennet, 1895; RBGE E00762046). The plant was also found in Loch Bì in 1933 (Nicol, 1936). The 72 lagoons surveyed for the MNCR ranged in size from the 0.06 ha Loch Portain lagoon to the 850 ha Loch Bì, the second largest lagoon in the UK.
During the surveys in 1993, 1994 and 1995, Z. noltii was recorded in five lagoons in Lewis, North Uist, and South Uist, including in the northern end of Loch Bì, near the culverts which connect the lagoon to the sea. Subtidal Z. marina was recorded in 11 lagoons in Lewis, North Uist and Benbecula. Notably, in Oban a’ Chlachain, North Uist, on coarse sand and small boulders at depths of three to six metres, Z. marina was recorded growing to almost two metres long in large beds with Ruppia sp. and Chorda filum. In Oban Sponish, North Uist, the basin was found to be “dominated by luxuriant beds” of the seagrasses Z. marina and Ruppia sp. (Thorpe et al., 1998).
In 1995, Entec (1996) carried out a more intensive survey of Loch nam Madadh, a complex fjardic system in North Uist. Large beds of the seagrasses Ruppia sp. and Zostera sp. were found in Ob nan Stearnan, Loch an Strumore and Loch an Duin, and in the small lagoons west of Loch Minish, west of Loch Voiskinish and south of Loch an Duin.
Summary
Seagrass declines in relation to the 1930s wasting disease were noted by wildfowlers, although the beds in the Sound of Harris appeared to be unaffected by the epidemic. In some locations, extensive Z. marina beds still exist, however, the lack of comparative historical data means that an assessment of population trends is not possible. A recent decline appears to have occurred south of Berneray with potential causes being suction dredging for shellfish and construction of a nearby causeway. Seagrass populations east of the Eriskay Causeway also appear to have declined as a response to changes in hydrodynamic conditions.
North Coast
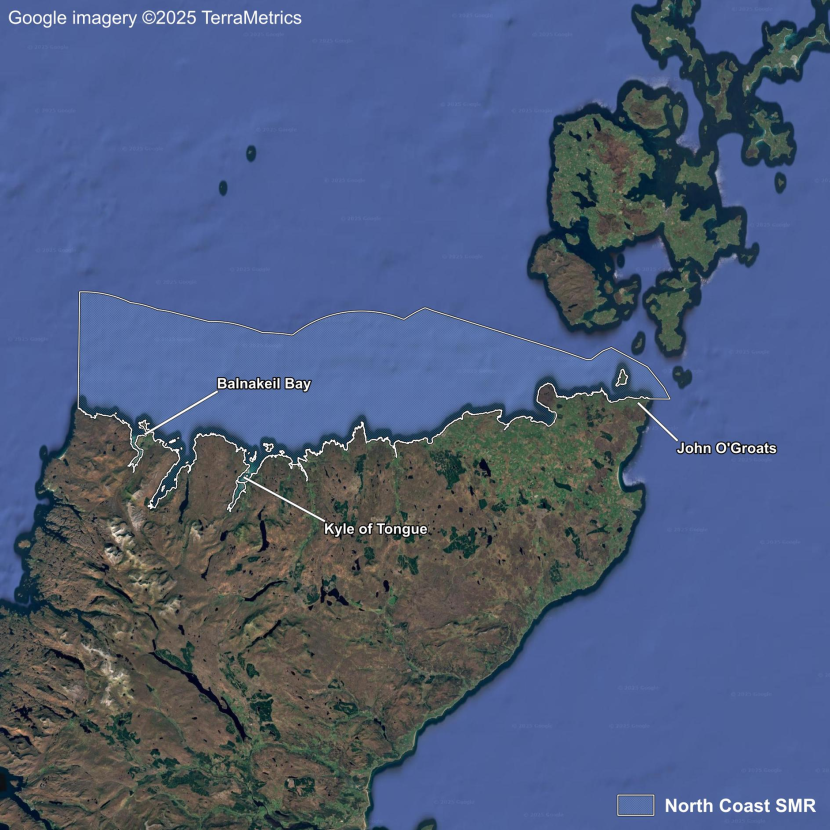
Figure 38. North Coast Scottish Marine Region.
The North Coast SMR comprises the northern coastlines of Sutherland and Caithness (Figure 38). This is predominantly rocky, and indentations are generally limited to narrow geos cut into the cliffs by highly erosive wave action. A few large inlets and sandy beaches are present though the coast is mostly inhospitable for seagrass. Six records were found: a herbarium specimen of Z. marina collected near Durness in 1856 (RBGE E00762018); three of Z. marina intertidal variety from Kyle of Tongue in 1900, 1919 and 1921; one of Z. marina from Balnakeil Bay in 1993; and one of washed-up Z. marina on a beach nearby in 2008 (RBGE E00762058; Marshall, 1901; Anthony, 1976; BSBI, 2024). The historical records do not indicate the presence of any sizeable seagrass beds in this region, for example, Marshall reported that the intertidal variety of Z. marina was only present very locally in the Kyle of Tongue (Marshall, 1901). It was not possible to quantify any declines or recoveries in this area due to data limitations.
Orkney Islands
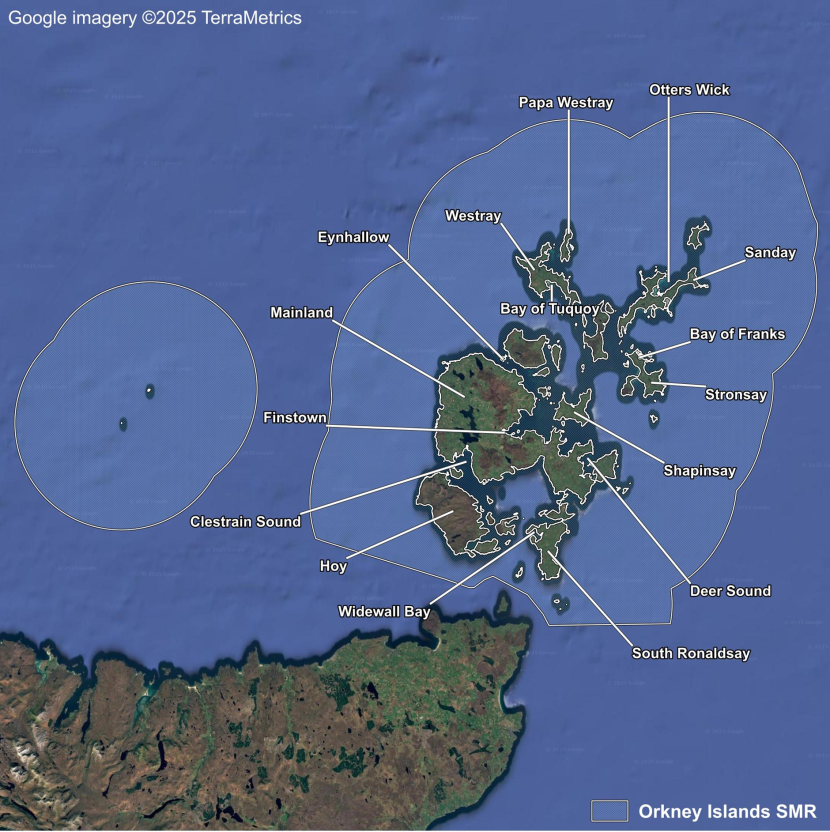
Figure 39. Orkney Islands Scottish Marine Region.
Orkney Islands SMR covers the Orkney Islands (Figure 39). Deep water occurs along the western edge of Orkney which is exposed to large Atlantic swells. The western parts of several islands have high cliffs which buffet wave action. The topography grades eastwards into a predominantly gentle, flat landscape which extends into the coastal waters. There are many large sheltered and relatively shallow bays which are ideal for seagrass.
Historical accounts of seagrass beds on Orkney indicate that they occurred profusely. Spence (1914) described the plant as common while Urquhart (1824) gave the following descriptions:
“The marine plant Zostera or Sea-grass is found in abundance in all those bays of the Orkney Islands which are not exposed to the immediate fury of the ocean; and is there known by the name of Mella or Mallow. Wherever the bays are land-locked, banks of sand and mud accumulate, which appear to be held together principally by the roots of this plant, which are strong and succulent, and throw out numerous lateral fibres”;
“Should a sufficient demand arise for this grass, at a fair price, any quantity could be collected in the Orkney Islands that the market could require”.
A local farmer recalled that during the 1920s over a hundred tonnes of seagrass washed ashore at harvest time near St Peter’s Pool in Deerness, though in 1974 “a good ton was washed up after many years of minimal amounts” (Smith, 1975).
In the MNCR Orkney Lagoons summaries, Thorpe (1998) reported dense patches of subtidal Z. marina in the Oyce of Huip, Stronsay, and scattered Z. marina in the mid-shore and sublittoral zones of The Ouse at Finstown on the Mainland of Orkney. The survey was conducted in 1994.
The MNCR area summaries for Orkney (Murray et al., 1999) reported a large, very dense bed of Z. marina at St. Peter's Pool in the inner Deer Sound, surveyed in 1996, and dense patches of Z. marina in the southern, most sheltered part of Otters Wick, Sanday, surveyed in 1997. It also reported beds off the islands of Papa Westray and Eynhallow surveyed in 1996 and 1997, respectively, and sparse Z. marina off south Sanday and the north-west coast of Hoy. The island of Westray was not included in the summaries.
In 2010, a SNH-commissioned study combined predictive modelling (using bathymetry and wind-driven wave exposure data) with boat-based surveys to confirm the existence of known Z. marina beds and find new ones (Thomson et al., 2014). Extensive subtidal beds were confirmed in Deer Sound and Widewall Bay. New large subtidal beds were found along the north shore of Wyre in stronger currents than is usual and were associated with maerl habitat.
Subtidal Z. marina was also found at: Bay of Isbister; Longhope; Burray; Graemsay; Rousay Sound; Gairsay Sound; Mill Bay, Stronsay; and Veantrow Bay, Shapinsay.
Small patches of what appeared to be the intertidal variety of Z. marina were found on the extensive mud flats of the Finstown Ouse lagoon, with intertidal and subtidal Z. marina found outside it, which is the same as was found in 1994.
The survey very likely underestimated the extent of Z. marina in Orkney as it was carried out from January to March 2010 when perennial Z. marina is harder to see than in summer. In addition, the islands of Westray, Papa Westray and Sanday, which are known to have Z. marina beds, were not surveyed.
Project Seagrass data from surveys in 2021 and 2022 (GeMS), show subtidal Z. marina beds in Clestrain Sound and the outer Deer Sound on the Mainland, in the Bight of Quoyolie off Papa Westray (in the same place as in 1996), in the Bay of Tuquoy off Westray, and in the Bay of Franks off Stronsay. Both Z. marina and Z. noltii were reported from Finstown, although the latter record may be a misidentification.
Summary
Numerous subtidal Z. marina beds exist around Orkney though historical descriptions indicate that populations in the 19th and early 20th centuries were much larger. However, a lack of comparative data precludes detailed assessment of population change.
Shetland Isles
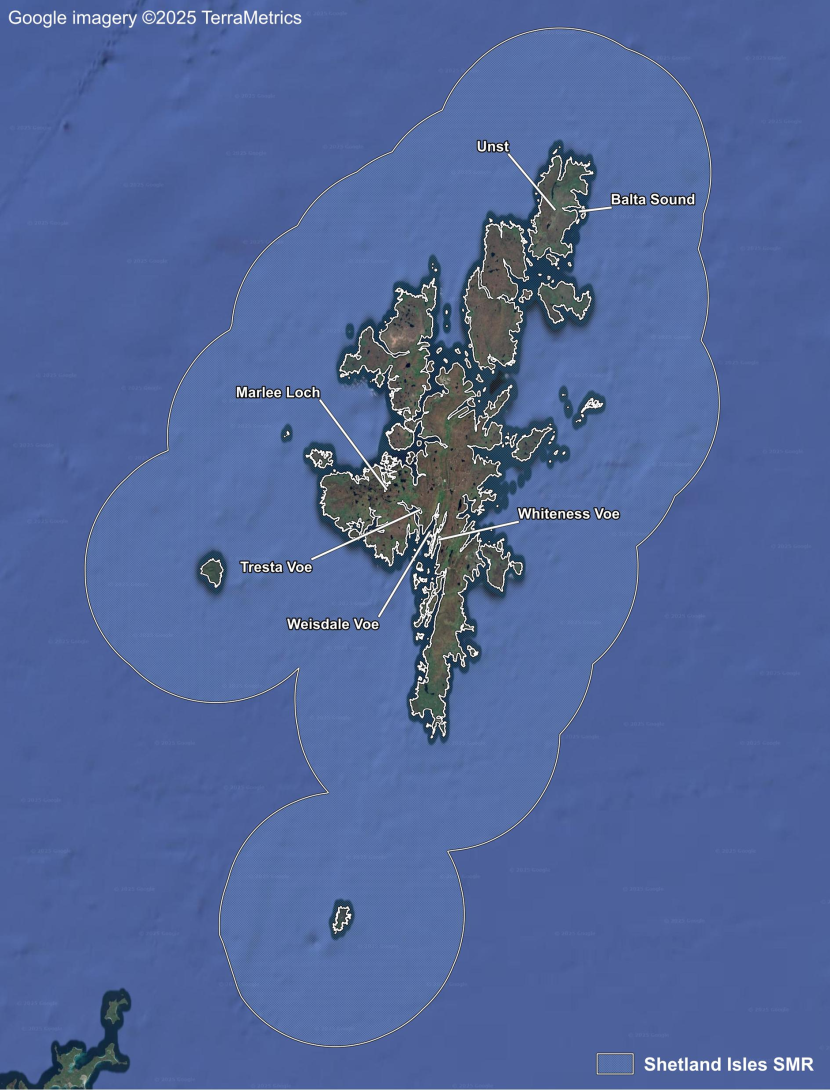
Figure 40. Shetland Isles Scottish Marine Region.
The Shetland Isles SMR covers the Shetland Isles (Figure 40). The rocky, exposed coasts of the Shetland Isles contain a number of long fjards and voes which provide a range of sheltered conditions in which seagrass can proliferate. Z. marina was considered common in Shetland during the 19th century and early 20th century (Edmondston, 1845; Saxby, 1903). It grew densely enough to be collected for bedding on Unst, while in Weisdale Voe and Marlee Loch the species was abundant until the 1930s wasting disease epidemic (Scott & Palmer, 1987). The disease is considered to have caused the dramatic reductions at these locations and the extinction of the species in Tresta Voe (Scott & Palmer, 1987; Scott et al., 2002; Scott, 2011).
Z. marina is also assumed to be extinct in Balta Sound. It was last seen there in 1887 (Scott & Palmer, 1987) indicating that it disappeared before the 1930s wasting disease epidemic. A herring fishing boom commenced in Shetland in 1880 with Balta Sound becoming a major herring harbour for over 30 years (Coull, 1996). Balta Sound supported over 10,000 fishermen and land-based workers with up to 50 curing stations along the shore (Coull, 1996; Goodlad, 1971; Halcrow, 1950). In the 1901 season, 130,870 crans of herring were landed there (The Shetland News, 1901). A cran typically contains about 1,200 herring but can vary from 700 to 2,500 depending on the size of the fish (ICH Scotland, 2025). The small sound is naturally sheltered, and the effluent from the fish gutting, cleaning and curing process as well as the sanitary requirements of thousands of workers would have seriously affected water quality, potentially leading, over many seasons, to the disappearance of the seagrass population.
Describing the MCNR survey series, Hiscock, (1989) reported that:
“Seagrass, Zostera marina has been especially looked for during our surveys but has been recorded from only five out of 236 sites … surveyed during 1986, 1987 and 1988.”
Z. marina was reported in several small beds around the upper Whiteness Voe during a MNCR survey in 1989 which despite their size were considered by Hiscock (1989) to be
“the most extensive known in Shetland and occupy several different sediment types there, although elsewhere have been recorded from mud or muddy sand only.”
Whiteness Voe was then designated as a Marine Consultation Area. Subsequent housing developments led to an increased volume of sewage entering the head of Whiteness Voe. A 1994 survey found this had created eutrophic conditions (Bunker et al., 1995):
“Whereas the Z. marina beds around the edges of the Voe seemed healthy, those by Mid Whiteness Rock had declined and in places were covered by filamentous algal growth and surrounded by anoxic sediments.”
In 2000, a wastewater treatment scheme for Wormadale and Nesbister was completed which was said to have improved the water quality in the head of the voe which is where the Zostera grows (Shetland Islands Council, 2004).
Marlee Loch and the Vadills lagoon system it is a part of appears to have lost its Z. marina. Hiscock (1986, cited in Thorpe, 1998) is reported to have described beds of the seagrass Z. marina in the southern basin of the Vadills, but subsequent MNCR surveys in 1987 and 1993 failed to find it (Thorpe, 1998). Nor was Z. marina found during a SNH commissioned site condition survey in 2003 (ERT (Scotland) Ltd., 2007).
The MNCR lagoon survey (Thorpe, 1998) did report extensive areas of dense subtidal Z. marina in the Loch of Hellister, on the Mainland, and also reported that freshwater-influenced muddy sand in the shallow north-east of the loch was colonised by sparse stands of Z. noltii. Samples of plant material from this area of the loch were collected in 2007 but these were identified as Ruppia maritima (BSBI, 2024). Therefore it is likely that the MNCR observation was a misidentification.
The MNCR area summaries for Shetland reported two small beds of Z. marina off West Burra, on soft flocculent mud in the northern end of the voe and on firmer sandy mud behind the island of Papil (Howson, 1999).
Survey work by the University of the Highlands and Islands in 2023 and 2024 has located several beds of Z. marina in upper Whiteness Voe in much the same locations as Hiscock in 1989, although the distribution differs, as does the methodology. One small Z. marina bed was also found in Upper Weisdale Voe. Z. marina beds were not relocated in the Loch of Hellister though visibility was suboptimal at the time of the survey. (Geisler et al., 2025).
Summary
Historical accounts indicate that dense populations of subtidal Z. marina occurred in several locations. These were reported to have disappeared as a result of the wasting disease epidemic and have not been recorded since. The previously declining population of subtidal Z. marina in Whiteness Voe appears to have stabilised following implementation of a wastewater improvement scheme.
Discussion and Recommendations
Seagrass declines in Scottish waters
Seagrass bed declines were evident across Scotland. Factors implicated in declines include poor water quality, land reclamation, mechanical damage and ecosystem degradation. The historical literature indicated that wasting disease caused significant declines in some places including the Firth of Tay, Shetland and the Outer Hebrides. The findings of this report are therefore in agreement with those from other parts of Europe and eastern North America in that significant declines occurred at the time of the wasting disease epidemic while other factors have also adversely affected seagrass populations (Blegvad, 1935; Cottam, 1944; Rasmussen, 1977; Baden et al., 2003; Green & Short, 2003).
Subtidal Z. marina appears to occur much more patchily than it did formerly around Scotland. Historical descriptions from Orkney and the West Highlands indicate that much greater biomasses occurred in the 19th and early 20th centuries. Substantial subtidal beds also appear to have been lost from the Cromarty Firth. The largest known population declines of subtidal Z. marina have occurred in Shetland in response to the wasting disease epidemic.
Shetland is the most northerly and geographically isolated Scottish archipelago. The relatively low summer temperatures may limit sexual reproduction, and a population bottleneck effect appears to have occurred here with low genetic diversity recently recorded. These factors may explain the lack of resilience of the Shetland population to withstand adverse environmental stress.
Significant historical seagrass declines were detected for Z. marina and Z. noltii in estuaries with the largest conurbations. The most widespread occurred in the upper Firth of Clyde, which receives wastewater and industrial effluents from Glasgow, the largest city in Scotland. Historically, the Clyde was the most polluted river in the country with severe water quality issues also occurring in the Forth catchment (Henderson & Hamilton, 1986; Hammerton, 1994; Smout & Stewart 2012). Comparison of the historical botanical literature with contemporary data indicates that Z. marina diminished and possibly died out in the upper Firth of Clyde and became seriously depleted in the Firth of Forth.
Seagrass recovery does not appear to have begun in the upper Firth of Clyde estuary, potentially reflecting the water quality status, which has not shown the reductions in nutrient levels seen in the Firth of Forth following implementation of the Water Framework Directive in 2000. Contrastingly, the intertidal seagrass populations of the Firth of Forth have recently exhibited a marked recovery, although this involves only Z. noltii. Z. marina is less hardy than Z. noltii in terms of general environmental conditions and seagrass populations situated further down the shore could be anticipated to have less capacity to tolerate a reduction in available light. This likely contributed significantly to the historical decline of Z. marina, especially the subtidal populations, in historically polluted Scottish estuaries.
Small catchments situated within the east coast lowlands (including the catchments of the Rivers Ythan, Lunan and Eden) are relatively prone to water quality issues from agriculture and sewage (Marsden & Mackay, 2001). They contrast, for example, with the large northern firths and Solway Firth which receive vast amounts of water from low nutrient upland environments. These have the capacity to dilute nutrient inputs from the agriculturally improved lowlands. Water quality issues were most likely the cause of historical seagrass declines in the Eden Estuary and may account for the apparent absence of seagrass in the Ythan Estuary. However, the lack of any seagrass records for the Ythan Estuary (including prior to the wasting disease epidemic and the onset of farming intensification) is difficult to understand, though may just reflect a lack of historical recording effort.
Nutrient inputs from aquaculture have increased along the west coast of Scotland and around the Northern Isles. The seagrass population in Poll Loisgann disappeared following installation of a salmon farm although lack of data precluded definitive conclusions regarding causative factors.
Land reclamation and coastal development have also played a part in seagrass declines around Scotland. Land reclamation directly removed a portion of the Zostera bed at Nigg Bay, Cromarty Firth and led to large-scale declines at Hunterston in Ayrshire. Such direct losses, combined with indirect losses from associated changes in hydrology, have undoubtedly played a significant role in the historical decline of Scottish seagrass beds, particularly in heavily urbanised estuaries.
Seagrass declines at Eriskay and Berneray have been linked to construction of inter-island causeways. Detailed mapping of subtidal seagrass beds south of Berneray has revealed a greater than 50% decline in seagrass over the last 34 years. Extensive seagrass bed declines occurred in Loch Fleet following a change in sluice gate management for a tidal barrage. The extent of beds reduced by 82% between 2000 and 2012 with further declines occurring up to the present.
There are anecdotal reports that bottom-towed fishing gear has supressed seagrass populations off the coast of Arran, Islay, and Kintyre. Damage from mechanical shellfish harvesting has also caused declines to seagrass populations in some areas. For example, intertidal Z. marina was believed to have been virtually eliminated from areas in the Solway Firth where suction dredging for cockles had occurred (Perkins, 1988). In Berneray, suction dredging for razor clams may have contributed to declines in seagrass bed extent.
Historical harvesting of mussel beds, including subtidal raking and dredging to supply bait for the whitefish handline fishery, in addition to overharvesting of oyster beds, is likely to have had a significant adverse effect on seagrass both in terms of reduced water filtration and physical disturbance from harvesting operations.
Seagrass recoveries in Scottish waters
In common with other European countries, the Scottish population of subtidal Z. marina does not appear, from limited data and observations, to have substantially recovered from the wasting disease epidemic. This can be seen in the loss of beds from specific locations around Shetland, which have never repopulated or have only recovered to a small fraction of their former status. The lack of recovery of subtidal population Z. marina around Scotlandmay partly be related to water quality issues and loss of sediment banks historically accreted by seagrass (Wilson, 1949; Rasmussen, 1977).
Substantial efforts to improve water quality towards the end of the 20th century and into the 21st century have resulted in significant recoveries in seagrass bed populations. This is particularly evident for Z. noltii populations at Loch Ryan and the Firth of Forth. Seagrass populations within Loch Indaal may also be recovering following water quality improvement.
Recoveries are also evident in the Solway Firth seagrass populations, encompassing both intertidal Z. marina and Z. noltii. Z. marina is recovering in areas where mechanical disturbance from cockling has ceased, suggesting that the management measures in this area are effective.
Recoveries of some seagrass populations in Scotland demonstrate what can be achieved if pressures are identified and removed.
Seagrass populations of unknown status
The historical literature indicates that profuse populations occurred around Orkney in the early 19th century and many beds have recently been recorded. Unfortunately, a lack of regular systematic and comprehensive mapping precluded quantitative assessments of population change. There are a number of survey programmes underway in Orkney to aid understanding of distribution and extent of seagrass beds and these studies, in combination with past surveys, may enable future population trends to be detected.
Seagrass population changes in the Moray, Beauly and Cromarty Firths are unknown. There is insufficient regular, comparative survey data to assess potential changes. At Udale Bay and Nigg Bay in the Cromarty Firth for example there are few differences in the mapped distribution within the period 1970-1991, however, in recent surveys recorded areas were markedly lower. The change in recorded areas may indicate large scale declines and these sites should be monitored intensively over a number of successive years to establish whether long term declines are occurring. However, as substantially different survey techniques were used between the two time periods the differences in areas mapped may be a function of different survey techniques.
Historical records indicate that seagrass habitats occurred widely within the Argyll and West Highlands Scottish Marine Regions. This area warrants further survey work to determine the status of historical beds, and to locate new beds.
Assessment of limitations
The current study did not locate any maps of Scottish seagrass beds drawn prior to the 1930s. Butcher’s 1941 map of Zostera around the UK coast appears to be an estimate of likely distribution before 1930, extrapolated from limited presence/absence seagrass data likely referenced against soft sediment distributions (Carstairs, 2024). As a result, it was not used in this project.
The earliest mapped polygon data for seagrass beds in Scotland found during this project dates to around 1972 in the Cromarty Firth. Unfortunately, the relatively late mapping of beds in Scotland and lack of decline estimates from the time of the wasting disease epidemic preclude accurate quantitative assessments of historical seagrass distributions in Scottish waters.
The variety of techniques used to undertake historical mapping of seagrass beds has reduced data comparability, limiting conclusions in many areas. In particular, survey methodologies varied in how plant density thresholds were used to define bed edges. Large intertidal beds often have extensive areas of sparsely distributed plants, particularly at their seaward edge. As a result, differences in mapping methodology regarding bed definitions can confound temporal analyses of population change.
Lack of mapping to species and variety level in mixed beds also significantly affects assessments of temporal population change. Inter-annual stability is likely to be greater in perennial Z. noltii populations than in annual intertidal Z. marina populations.
Several of the historical surveys did not provide a methodology, and most did not comprehensively map all areas containing seagrass within large estuaries. Some surveyors did not allow enough time to reach the lower bed due to tidal limitations and did not return at a later date. Additionally, most survey reports did not explicitly indicate where seagrass was confirmed to be absent.
The uncertainty regarding the historical and current distributions of types of Z. marina around Scotland hampers assessments of ecosystem decline and recovery. This uncertainty stems from botanists historically recording at different taxonomic levels, changes in taxon classifications, and difficulties with field identification (Cleator, 1993). The extent of subtidal Z. marina beds is likely to be disproportionally under-recorded due to their presence in permanently submerged waters in remote locations. Similarly, lack of visibility and paucity of data preclude assessment of the extent to which particular activities, such as scallop dredging, affect subtidal seagrass beds.
Attributing causality to seagrass declines or lack of recovery across large swaths of subtidal habitat is further confounded due to the historical concomitant development of industries that have the potential to impact seagrass populations. For example, the three-mile fishing limit for trawling was rescinded in 1984 after a period of 95 years (Thurstan & Roberts, 2010), allowing access into shallow coastal waters, including sea lochs, during a period when salmon farming was also developing within coastal waters.
Future prospects
There is evidence of a recovery of seagrass populations in response to water quality improvements in Loch Ryan, the Eden Estuary, Montrose Basin and the Firth of Forth. Recent substantial increases in seagrass populations within the heavily industrialised catchment of the Firth of Forth are encouraging and vindicate the effort and expense associated with gradual water quality improvements, which commenced towards the end of the 20th century. In contrast, with the fish farm industry expanding, areas adjacent to farms may experience an increase in nutrient loading (Scottish Government, 2020a). Increased run-off associated with climate change could also increase nutrient loading in coastal habitats. This in combination with potential losses of epiphyte grazers will require careful monitoring in terms of cascading trophic effects on seagrass population dynamics. Losses of potential invertebrate pollinators should also be monitored.
Fishing activity using towed bottom contacting gear has been restricted within certain MPAs, and additional management measures are currently being considered, including protection for seagrass outside of MPAs. Current and future management of bottom-towed fishing activity are likely to facilitate the recovery of seagrass populations. The recovery of seagrass within the Solway Firth shows that beds can recover following the implementation of management measures for shellfish harvesting.
Sargassum muticum is now abundant in places in the Firth of Clyde (Davison, 2009). The seaweed may have caused significant seagrass declines on Arran and is present within the seagrass bed at Dunure. Further spread of S. muticum could lead to extensive declines of Z. marina beds on predominantly rocky shores as have been reported at Roscoff, France (den Hartog, 1997). Monitoring the presence and spread of S. muticum in relation to individual seagrass beds may allow targeted management intervention in advance of further declines.
Many seagrass beds are vulnerable to sea level rise. In particular, the seagrass beds in Shetland are potentially at risk as the island archipelago has the highest projections in Scotland (McKirdy, 2007). The remoteness of the island group from other coastal areas containing seagrass beds reduces the potential for viable plant material to reach the islands, thereby limiting opportunities for recolonisation and gene flow.
Other climate related pressures, including increased storm frequencies may also impact seagrass beds. Shallow beds in exposed locations are particularly vulnerable to storms and may cease to exist under future climate scenarios.
Restoration and carbon sequestration implications
There are a variety of natural factors that affect the distribution and population dynamics of seagrass beds. Considering these in restoration projects could ultimately improve the outcomes. For example, site suitability studies for potential seagrass restoration should, where data is available, include assessments of rates of sediment accretion and erosion, degree of sediment anoxia, wave exposure and water quality including levels of light penetration.
Furthermore, the current project demonstrates that the two varieties of Z. marina occupy very different ecological niches in Scotland and generally have different life history strategies. The degree to which this is underpinned by genetics is not currently known, though significant differences are likely to occur in line with the process of evolution by natural selection. Restoration projects should therefore match donor plant material to potential recipient sites in terms of variety, geographical location and habitat type.
Small seagrass populations in isolated island archipelagos are vulnerable to environmental stress events and may benefit from the introduction of fresh genetic material. The extinction of seagrass from the Faroe Islands in the 20th century exemplifies this scenario (Jensen et al., 2023). There is only one remaining substantial subtidal Z. marina population in Shetland, in Whiteness Voe, which sub-sampling indicates is comprised of a single clone (R.J. Lilley, pers. comm.) and therefore genetically impoverished. Extensive clonal subtidal Z. marina beds are a feature of northern European waters with one recently recorded in the Baltic Sea being more than 1,400 years old (Reusch et al., 1999; Yu et al., 2024). The population at Whiteness Voe may benefit from the introduction of fresh genetic material via the introduction of pollen, seeds, or rhizomes, to increase resilience against environmental stress events including outbreaks of wasting disease (Reusch et al., 2005). However, the bed is likely to be a relic of great antiquity and conservation value in its own right. It may be more appropriate to improve conditions naturally rather than undertake genetic augmentation which should be a last resort in the event that the bed significantly declines.
Restoration work could also focus in areas where seagrass used to occur but has disappeared. There is evidence that recovery of seagrass beds is possible if pressures are removed. For example, elsewhere in Shetland, many historical seagrass beds appear to have been completely wiped out by potential water quality issues and the wasting disease epidemic. These areas, such as Balta Sound, Marlee Loch and Tresta Voe may be good candidates for the reintroduction of subtidal Z. marina populations. Restoration at these sites would further enhance the species survival prospects around the archipelago. Rhizomes from populations thriving in similar environmental conditions, with similar sediment characteristics and tidal regimes would be most likely to survive, although much will depend on the present state of the recipient sites. For sites in Shetland, Whiteness Voe should be used if stock levels are sufficient in order to provide locally adapted material. Additional material from Orkney could be used to enhance genetic diversity and therefore population resilience to stress events, although consideration should be given to the appropriateness of introducing genetic material from another archipelago. This report has identified a variety of other locations where seagrass beds used to occur. The restoration potential in these areas warrants further investigation.
Seagrass restoration exercises in areas which historically contained oyster beds would benefit from concomitant reestablishment of filter feeder populations in suitable habitats.
Rates of carbon sequestration are likely to vary between seagrass species, as well as in relation to life history strategies and in relation to the degree of sediment accretion facilitated by seagrass. Carbon storage could be anticipated to be higher in perennial rather than annual seagrass beds, particularly in subtidal Z. marina beds. Subtidal Z. marina beds accrete sediments over substantial periods of time (Wilson 1949; Blois et al., 1961, Rasmussen 1973; Rasmussen 1977) while sediment accretion in intertidal beds may only occur seasonally (den Hartog & Phillips, 2001) or not at all (Wilkie, 2011). Large subtidal sediment banks developed by Z. marina were washed away following plant mortalities in relation to the 1930 wasting disease. Natural redevelopment of these banks following re-introduction of seagrass may offer the greatest potential in terms of carbon sequestration by seagrass in Scotland.
Future research and recommendations
This study identified several areas that warrant future research:
- The conservation of Scottish seagrass beds would significantly benefit from a greater understanding of their historical distribution. It is recommended that a detailed study of historical aerial photography is undertaken to establish if the assessment of declines of subtidal Z. marina in particular can be improved. Historical photographs have been used to great effect in assessing such declines along the east coast of North America and have successfully highlighted substantial seagrass declines south of Berneray in Scotland.
- The seed coats of seagrass are extremely tough and can persist for decades in marine sediments. Analysis of sediment core sections could be undertaken to further elucidate former seagrass distribution and identify past periods of nutrient enrichment and algal blooms. Cores can be dated using a variety of techniques including pollen profiles, sedimentation rates, radioisotopes and archaeological evidence.
- Monitoring for potential cascading trophic effects should regularly occur in subtidal Z. marina habitats in relation to aquacultural operations. Similarly, studies on the potential processes involved and extent of invertebrate-mediated subsurface pollination of Scottish seagrass taxa would be valuable in terms of conservation management.
Furthermore, this report has several recommendations with regards to monitoring seagrass beds, including where to focus survey effort:
- Greater knowledge of the current distribution of seagrass beds around the Scottish coast is urgently required. This would enhance understanding of their vulnerability and provide contemporary baseline data against which the impacts of ongoing and future stressors could be assessed. An appropriate methodology combined with diligent field work would be required to obtain high integrity data, allowing valid comparisons with past and future surveys. This is particularly important in terms of sea level rise, which may already be having an effect, especially on intertidal populations. The status of seagrass beds on the west coast of Scotland, in particular, is poorly understood. A better understanding of the current distribution of seagrass beds on the west coast would be beneficial.
- Despite the recent downgrading of intertidal Z. marina in the British taxonomic literature from a species to a conspecific of subtidal Z. marina, it is important that recorders continue to delineate this taxon from subtidal Z. marina where possible, given the significant differences in their ecology, habitat and distribution. Further knowledge regarding their ecology is likely to be invaluable in restoration efforts.
- It is recommended that the Cromarty Firth is surveyed using techniques similar to those employed in the 1970s to ascertain whether recent declines in recorded seagrass distributions are real or a function of different survey techniques. Surveys should be carried out over several consecutive years to take account of natural inter-annual variations in population size. The Dornoch Firth has not been surveyed comprehensively since 1986 and should be the focus of a comparative assessment.
- Baseline distribution mapping of Ruppia sp. should be undertaken to monitor any expansion of this genus which would likely signify deteriorating conditions for Zostera sp.
- Hand removal of Sargassum muticum from seagrass beds along the west coast, where feasible, may be necessary to prevent further deterioration.
References
Ackerman, J.D. 1983. Current flow around Zostera marina plants and flowers: implications for submarine pollination. The Biological Bulletin, 165, p. 504.
Ackroyd, J. 1972. Montrose Basin - Zostera. Report to JNCC. Unpublished.
Adkins, F. 2023. Seawilding: the Scottish community reviving a loch (Accessed on 25th September 2025).
Andersson, S., Presson, M., Moksnes, P., & Baden, S. 2009. The role of the amphipod Gammarus locusta as a grazer on macroalgae in Swedish seagrass meadows. Marine Biology, 156, pp. 969–981.
Angus, S., Hansom, J. & Rennie, A. 2011. Habitat change on Scotland’s coasts. In: S. J. Marrs, S. Foster, C. Hendrie, E.C. Mackey & D.B.A. Thompson (eds.) The Changing Nature of Scotland. The natural heritage of Scotland series (17). Edinburgh: TSO Scotland. pp. 183-198.
Anthony, J. 1976. John Anthony’s Flora of Sutherland. Edited and compiled by J.B. Kenworthy. Botanical Society of Edinburgh.
Atkins, S.M., Caudwell, C.M. & Herbert, R.A. 1992. Montrose Basin Tidal Flats, Environmental Survey - September 1991. Report to Scottish Wildlife Trust. Environmental Advisory Unit, University of Dundee.
Atkinson-Willes, G.L. & Matthews, G.V.T. 1960. The past status of the brent goose. British Birds, 53, p. 352.
Atkinson-Willes, G.L. 1963. Wildfowl in Great Britain. Monograph of the Nature Conservancy Council No. 3. HMSO.
Axelsson, M.B. & Dewey, S. 2013. Loch Fleet eelgrass study – the extent and abundance of eelgrass in the Loch Fleet SSSI in 2012. Scottish Natural Heritage Commissioned Report No. 702.
Backman, T.W.H. 1991. Genotypic and phenotypic variability of Zostera marina on the west coast of North America. Canadian Journal of Botany, 696, pp. 1361-1371.
Baden, S., Gullström, M., Lundén, B., Pihl, L. & Rosenberg, R. 2003. Vanishing seagrass (Zostera marina, L.) in Swedish coastal waters. Ambio, 32, pp. 374-377.
Balfour, J.H. 1863. Flora of Edinburgh being A List of Plants found in the vicinity of Edinburgh. Edinburgh: Adam & Charles Black.
Ballantyne, J. 1911. List of Bute Plants. Transactions of the Natural History Society of Glasgow, 8, pp. 219-226. [Read to the society in 1906].
Bangerter, E.B. & Cannon, J.F.M. 1978. Flowering plants and conifers. In: A.C. Jermy & J.A. Crabbe (eds.) The Island of Mull, a survey of its flora and environment. London: British Museum.
Bates, C.R., Moore, C.G., Malthus, T., Mair, J.M. & Karpouzli, E. 2004. Broad scale mapping of habitats in the Firth of Tay and Eden Estuary, Scotland. Scottish Natural Heritage Commissioned Report No. 007.
Batters, E.A.L. 1892. Additional Notes on the Marine Algae of the Clyde Sea Area. Journal of Botany, 30, pp. 170-177.
Baxter, J.M., Duncan, K., Atkins, S.M. & Lees, G. (eds.) 1999. Scotland’s living coastline. London: TSO.
Beaver, R. & Dipper, F.A. 2002. Marine Nature Conservation Review Sector 14. Sealochs in the Outer Hebrides: area summaries. Peterborough: JNCC.
Beca-Carretero, P., Marín, C., Azcárate-García, T., Cara, C.L., Brun, F. & Stengel, D.B. 2024. Ecotype-specific and correlated seasonal responses of biomass production, non-structural carbohydrates, and fatty acids in Zostera marina. Plants, 13, no. 3: p. 396.
Becheler, R., Diekmann, O., Hily, C., Moalic, Y. & Arnaud-Haond, S. 2010. The concept of population in clonal organisms: mosaics of temporally colonized patches are forming highly diverse meadows of Zostera marina in Brittany. Molecular Ecology, 19, pp. 2394-2407.
Bechmann, R.K., Arnberg, M., Gomiero, A., Westerlund, S., Lyng, E., Berry, M., Agustsson, T., Jager, T. & Burridge, L.E. 2019. Gill damage and delayed mortality of Northern shrimp (Pandalus borealis) after short time exposure to anti-parasitic veterinary medicine containing hydrogen peroxide. Ecotoxicology and Environmental Safety, 180, pp. 473-482.
Begon, M., Harper, J.L. & Townsend, C.R. 1990. Ecology: Individuals, Populations and Communities. 2nd edition. Oxford: Blackwell Scientific Publications.
Benmokhtar, S., Robin, M., Maanan, M., Boutoumit, S., Badaoui, B. & Bazairi, H. 2023. Monitoring the spatial and interannual dynamic of Zostera noltei. Wetlands Wilmington, 43(5), p. 43.
Bennet, A. 1895. Contributions towards a flora of the Outer Hebrides. Annals of Scottish Natural History, No. 13, pp. 240-247.
Bentham, G. 1858. Handbook of the British Flora A Description of the Flowering Plants and Ferns indigenous to, or naturalised in, The British Isles. 1st edition. London: Lovell Reeve.
Berry, J. 1930. Notes on the Movement of Duck off Tentsmuir Point, Fife. The Scottish Naturalist. No 18, 43-46.
Berry, J. 1939. The distribution and status of wild geese and wild duck in Scotland, International Wildfowl Inquiry, Volume II. Cambridge: Cambridge University Press.
Bertelli, C.M. & Unsworth, R.K.F. 2014. Protecting the hand that feeds us: Seagrass Zostera marina serves as commercial juvenile fish habitat. Marine Pollution Bulletin, 83(2), pp. 425-429.
Bertelli, C.M., Robinson, M.T., Mendzil, A.F. Pratt, L.R. & Unsworth, R.K.F. 2018. Finding some seagrass optimism in Wales, the case of Zostera noltii. Marine Pollution Bulletin, 134, pp. 216-222.
Binnie, A., Beecher, P. & Gordon, D. 2012. GIS Mapping of Zostera Seagrass Beds within the Proposed South Arran MPA. Community of Arran Seabed Trust report.
Bishop, M.J., Peterson, C.H., Summerson, H.C. & Gaskill, D. 2005. Effects of harvesting methods on sustainability of a bay scallop fishery: dredging uproots seagrass and displaces recruits. Fishery Bulletin, 103, pp. 712-719.
Blanc, J., Arthur, S., Wright, G. & Beevers, L. 2012. Natural flood management NFM knowledge system: Part 3 - The effect of land drainage on flood risk and farming practice. Heriot-Watt University.
Blegvad, H. 1929. Mortality among animals of the littoral region in ice winters. Report of the Danish Biological Station, 35, pp. 51-62.
Blegvad, H. 1935. An epidemic disease of the eelgrass Zostera marina L.. Report of the Danish Biological Station 39, pp. 1-8.
Blois, J.C., Francaz, J.M., Gaudichon, S. & Lebris, L. 1961. Observations sur les herbiers de zostères de la région de Roscoff. Cahiers de Biologie Marine, 2, pp. 223-262.
Blok, S.E., Olesen, B. & Krause‐Jensen, D. 2018. Life history events of eelgrass (Zostera marina L.) populations across gradients of latitude and temperature. Marine Ecology Progress Series, 590, pp. 79-93.
Blumenröder, J., Sechet, P., Kakkonen, J.E. & Hartl, M.G.J. 2017. Microplastic contamination of intertidal sediments of Scapa Flow, Orkney: a first assessment. Marine Pollution Bulletin, 124(1), pp. 112–120.
Borum, J., Pedersen, O., Greve, T.M., Frankovich, T.A., Zieman, J.C., Fourqurean, J.W. & Madden, C.J. 2005. The potential role of plant oxygen and sulfide dynamics in die-off events of the tropical seagrass, Thalassia testudinum. Journal of Ecology, 93, pp. 148–158.
Borum, J., Gruber, R.K. & Kemp, W.M. 2012. Seagrass and related submersed vascular plants. In: J.W. Day, B.C. Crump, W.M. Kemp & A. Yáñez-Arancibia (eds.) Estuarine Ecology. 2nd edition. Hoboken, New Jersey: John Wiley & Sons.
Borum, J., Raun, A.L., Hasler-Sheetal, H., Pedersen, M.O., Pedersen, O. & Holmer, M. 2014. Eelgrass fairy rings: sulfide as inhibiting agent. Marine Biology, 161, pp. 351–358.
Bos, A.R., Bouma, T.J., de Kort, G.L.J. & van Katwijk, M.M. 2007. Ecosystem engineering by annual intertidal seagrass beds: sediment accretion and modification. Estuarine, Coastal and Shelf Science, 74, 1-2, pp. 344-348.
Boström, C., Baden, S., Krause-Jensen, D. 2003. The seagrasses of Scandinavia and the Baltic Sea. In: E.P. Green, F.P. Short & M.D. Spalding (eds.) World Atlas of Seagrasses: Present Status and Future Conservation. Berkeley: University of California Press.
Boström, C., Baden, S., Bockelmann, A.-C., Dromph, K., Fredriksen, S., Gustafsson, C., Krause-Jensen, D., Möller, T., Nielsen, S.L., Olesen, B., Olsen, J., Pihl, L. & Rinde, E. 2014. Distribution, structure and function of Nordic eelgrass (Zostera marina) ecosystems: implications for coastal management and conservation. Aquatic Conservation: Marine and Freshwater Ecosystems, 24, pp. 410-434.
Brackup, I. & Capone, D.G. 1985. The effects of several metal and organic pollutants on nitrogen fixation by the roots and rhizomes of Zostera marina. L. Environmental and Experimental Botany, 25(2), pp. 145-151.
BRIG (A. Maddock, ed.). 2008. UK Biodiversity Action Plan Priority Habitat Descriptions.
Bryce, J. 1859. Geology of Clydesdale and Arran: embracing also the marine zoology and the flora of Arran, with complete list of species, notes on the rarer insects of Arran, and notices of its scenery and antiquities. London & Glasgow: Richard Griffin & Co.
Bryce, J. 1872. The Geology of Arran and the other Clyde Islands with an Account of the Botany, Natural History, and Antiquities, Notices of the Scenery and an Itinerary of the Routes. 4th edition. Glasgow: William Collins, Sons, & Company.
BSBI. 2024. Botanical Society of Britain and Ireland, Distributional Database DDb. Accessed October 2024 - March 2025.
Bunker, F.St.P.D., Rostron, D. & Perrins, J.M. 1995. An assessment of the impact of eutrophication by domestic sewage effluent in Whiteness Voe Marine Consultation Area 9th to 13th August 1994. Contractor: Marine Seen, Hundleton, Dyfed. SNH Research, Survey and Monitoring Report, No. NE94/006.
Bunker, F., Brodie, J.A., Maggs, C.A., & Bunker, A.R. 2017. Seaweeds of Britain and Ireland. 2nd edition. Plymouth: Wild Nature Press.
Bunker, F.StP.D., Mercer, T.M., Howson, C.M., Moore, J.M., Diaz, P., Maggs, C.A. & Kamphausen, L. 2018. Site condition monitoring of maerl beds and seagrass beds in the Sound of Barra SAC 2015 – diving survey. Scottish Natural Heritage Research Report No. 924.
Burden, A., Smeaton, C., Angus, S., Garbutt, A., Jones, L., Lewis, H.D. & Rees. S.M. 2020. Impacts of climate change on coastal habitats relevant to the coastal and marine environment around the UK. MCCIP Science Review, pp. 228–255.
Burkholder, J.M., Mason, K.M. & Glasgow, H.B. 1992. Water-column nitrate enrichment promotes decline of eelgrass Zostera marina: evidence from seasonal mesocosm experiments. Marine Ecology Progress Series, 812, pp. 163–178.
Burkholder, J.M., Tomasko, D.A. & Touchette, B.W. 2007. Seagrasses and eutrophication. Journal of Experimental Marine Biology and Ecology, 350, pp. 46–72.
Burridge, L., Weis, J.S., Cabello, F., Pizarro, J. & Bostick, K. 2010. Chemical use in salmon aquaculture: A review of current practices and possible environmental effects. Aquaculture, 306, pp. 7-23.
Burrows, M.T., Twigg, G., Mieszkowska, N. & Harvey, R. 2017. Marine Biodiversity and Climate Change (MarClim): Scotland 2014/15. Scottish Natural Heritage Commissioned Report No. 939.
Burrows, M.T., Brook, T. & Twigg, G. 2025. Marine Biodiversity and Climate Change (MarClim): Scotland 2020-2022. NatureScot Research Report 1371.
Butcher, R.W. 1934. Zostera. Report on the present condition of eel grass on the coasts of England, based on a survey during August to October, 1933. ICES Journal of Marine Science, 9, pp. 49–65.
Butcher, R.W. 1935. Wasting disease of Zostera marina. Nature, 135, p. 545.
Butcher, R.W. 1941. The Distribution of Zostera eel grass, wigeon grass and other seashore plants in relation to the migrations of wildfowl. International Wildfowl Inquiry, Volume I. Cambridge: Cambridge University Press.
Butcher, R.W. 1961. A New Illustrated British Flora. London: Leonard Hill.
Byrne, K. 1993. Placenames of Colonsay and Oransay. Privately Published.
Cabot, D. 1999. Ireland: A Natural History. London: HarperCollins.
Cardoso, P.G., Brandão, A., Pardal, M.A., Raffaelli, D. & Marques, J.C. 2005. Resilience of Hydrobia ulvae populations to anthropogenic and natural disturbances. Marine Ecology Progress Series, 289, pp. 191-199.
Carstairs, M. 2024. The ecology and status of seagrass populations in the Solway Firth and Loch Ryan. Solway Firth Partnership Commissioned Report.
Caudwell, C.M. & Jones, A.M. 1994. Survey of green algal mats on the Eden Estuary Local Nature Reserve. Environmental Advisory Unit, University of Dundee.
Charman, K. 1977. The grazing of Zostera by wildfowl. Aquaculture, 12, pp. 229-233.
Che, X., Zhang, T., Li, H., Zhang, L. & Liu, J. 2022. Effect of hypoxia on photosystem II of tropical seagrass Enhalus acoroides in the dark. Photochemistry and Photobiology, 98, pp. 421–428.
Cho, H., Biber, P. & Nica, C. 2011. The Rise of Ruppia In Seagrass Beds: Changes In Coastal Environment and Research Needs. Seagrass: Ecology, Uses and Threats, pp. 209-224.
Chu, N., Yao, P., Ou, S., Wang, H., Yang, H. & Yang, Q. 2022. Response of tidal dynamics to successive land reclamation in the Lingding Bay over the last century. Coastal Engineering, 173, No. 104095.
Churchill, A.C., Nieves, G. & Brenowitz, A.H. 1985. Floatation and dispersal of eelgrass seeds by gas bubbles. Estuaries, 8, pp. 352-354.
Clapham, A.R., Tutin, T.G. & Warburg, E.F. 1952. Flora of the British Isles. 1st edition. Cambridge: Cambridge University Press.
Cleator, B. 1993. The Status of the Genus Zostera in Scottish Coastal waters. SNH Review No. 22, ISSN 1350 3111.
COAST. 2012a. Proposed Marine Protected Area South Coast of Arran. Community of Arran Seabed Trust.
COAST. 2012b. 2012 Seagrass mapping for the MPA. Community of Arran Seabed Trust.
Cope, T. & Gray, A. 2009. Grasses of the British Isles. London: Botanical Society of the British Isles.
Costa, J.E. 1988. Eelgrass in Buzzards Bay: distribution, production, and historical changes in abundance. United States Environmental Protection Agency.
Cottam, C. 1934. Past periods of eelgrass scarcity. Rhodora, 36, pp. 261–264.
Cottam, C. 1935. Further notes on past periods of eelgrass scarcity. Rhodora, 37, 440, pp. 269–271.
Cottam, C., Lynch, J.J. & Nelson, A.L. 1944. Food habits and management of American sea brant. The Journal of Wildlife Management, 8, pp. 36–56.
Cottam, C. & Munro, D.A. 1954. Eelgrass status and environmental relations. The Journal of Wildlife Management, 18, pp. 449–460.
Cotton, A.D. 1933. Disappearance of Zostera marina. Nature, 132, p. 277.
Coull, J.R. 1996. The Sea Fisheries of Scotland: A Historical Geography. Edinburgh: John Donald.
Courcier, P.A. 2024. Exploring Seagrass Meadows and Biodiversity around the Isle of Islay. MSc Thesis, The University of Edinburgh.
Covey, R. & Hocking, S. 1987. Helford River Survey. Unpublished report to Helford River Steering Group.
Covey, R. & Emblow, C.S. 1992. Littoral survey of the inner Solway Firth and additional sites in Dumfries and Galloway. JNCC Report, No. 33. Marine Nature Conservation Review Report, No. MNCR/SR/20.
Covey, R., Fortune, F., Nichols, D.M., Thorpe, K. 1998. Marine Nature Conservation Review Sectors 3, 4, 12, 13 & 15. Lagoons in mainland Scotland and the Inner Hebrides: area summaries. Peterborough: JNCC.
Cowley, C. 2012. The Marine Species of Lamlash Bay in the Nineteenth Century. Lamlash, Isle of Arran.
Cox, P.A., Laushman, R.H. & Ruckleshaus, M.H. 1992. Surface and submarine pollination in the seagrass Zostera marina L. Botanical Journal of the Linnean Society, 109(2), pp. 281–291.
Coyer, J.A., Hoarau, G., Kuo, J., Tronholm, A., Veldsink, J. & Olsen, J.L. 2013. Phylogeny and temporal divergence of the seagrass family Zosteraceae using one nuclear and three chloroplast loci. Systematics and Biodiversity, 11(3), pp. 271-284.
Craib, W.G. 1912. The Flora of Banffshire. Aberdeen University Studies: No. 54.
Critchley, A.T. 1980. The further spread of Sargassum muticum. British Phycological Journal, 15, p. 194.
Crosby, R. 1989. Report of Seasearch Survey Loch Ryan, S.W. Scotland, June 1989. Marine Conservation Society.
Cullain, N., McIver, R., Schmidt, A.L., Milewski, I. & Lotze, H.K. 2018. Potential impacts of finfish aquaculture on eelgrass Zostera marina beds and possible monitoring metrics for management: a case study in Atlantic Canada. PeerJ, 6, No. e5630.
Cunningham, M.H. & Kenneth, A.G. 1979. The Flora of Kintyre. Wakefield: EP Publishing.
Cunningham, C. & Hunt, C. 2023. Scottish Blue Carbon - a literature review of the current evidence for Scotland’s blue carbon habitats. NatureScot Research Report 1326.
D’Alpaos, L. & Defina, A. 1996. Tidal flow control by moveable gates: the case of the Po River Delta Lagoons. In: N. Burt & J. Watts (eds.) Barrages, engineering design and environmental impacts. Chichester: John Wiley & Sons.
Davies, B.F.R., Oiry, S., Rosa, P., Zoffoli, M.L., Sousa, A.I., Thomas, O.R., Smale, D.A., Austen, M.C., Biermann, L., Attrill, M.J., Roman, A., Navarro, G., Barillé, A.L., Harin, N., Clewley, D., Martinez-Vicente, V., Gernez, P. & Barillé, L. 2024. A sentinel watching over inter-tidal seagrass phenology across Western Europe and North Africa. Communications Earth and Environment, 5(1), p. 382.
Davis, F.W. 1985. Historical changes in submerged macrophyte communities of upper Chesapeake Bay. Ecology, 66(3), pp. 981-993.
Davison, D.M. 1997. The genus Zostera in the UK: A literature review, identifying key conservation, management and monitoring requirements. Environment and Heritage Service Research and Development Series. No.97/12.
Davison, D.M. & Hughes, D.J. 1998. Zostera biotopes: An overview of dynamics and sensitivity characteristics for conservation management of marine SACs, Vol. 1. Scottish Association for Marine Science UK Marine SACs Project.
Davison, D.M. 2009. Sargassum muticum in Scotland 2008: a review of information, issues and implications. Scottish Natural Heritage Commissioned Report No. 324.
De Casabianca, M.L., Tari, P.S., Gauchet, R., Raynaud, C. & Rigollet, V. 2004. Relationships between heavy metal concentrations in sediments and eelgrass and environmental variables Zostera marina, Thau Lagoon, France. Vie et Milieu, 54(4), pp. 231-238.
De Cock, A.W.A.M. 1980. Flowering, pollination and fruiting in Zostera marina L. Aquatic Botany, 9, pp. 201-220.
De Cock, A.W.A.M. 1981. Influence of temperature and variations in temperature on flowering in Zostera marina L. under laboratory conditions. Aquatic Botany, 10, pp. 125-131.
de Jonge, V.N. & de Jong, D.J. 1992. Role of tide, light and fisheries in the decline of Zostera marina L. in the Dutch Wadden Sea. In: N.M.J.A. Dankers, C.J. Smit & M. Scholl (eds.) Present and future conservation of the Wadden Sea: Proceedings of the 7th International Wadden Sea Symposium, Ameland 1990. Publication Series. Netherlands Institute for Sea Research NIOZ, 20: pp. 161-176.
de Jonge, V.N., Schuttelaars, H.M., van Beusekom, J.E.E., Talke, S.A. & de Swart, H.E. 2014. The influence of channel deepening on estuarine turbidity levels and dynamics, as exemplified by the Ems estuary. Estuarine, Coastal and Shelf Science, 139, pp. 46–59.
De Lisle, L. 2024. Report on the Discovery of a Seagrass Bed Zostera marina in Loch Ryan. Report to Solway Firth Partnership and Dumfries & Galloway Council by Newton Stewart Aqua Club.
Delefosse, M. & Kristensen, E. 2012. Burial of Zostera marina seeds in sediment inhabited by three polychaetes: Laboratory and field studies. Journal of Sea Research, 71, pp. 41-49.
Delistraty, D.A. & Hershner, C. 1984. Effects of the herbicide Atrazine on adenine nucleotide levels in Zostera marina Eelgrass. Aquatic Botany, 18, pp. 353-369.
den Hartog, C. 1970. The Seagrasses of the World. Amsterdam: North-Holland Publishing.
den Hartog, C. 1987. ‘Wasting disease’ and other dynamic phenomena in Zostera beds. Aquatic Botany, 27, pp. 3-14.
den Hartog, C. 1994. Suffocation of a littoral Zostera bed by Enteromorpha radiata. Aquatic Botany, 47, pp. 21-28.
den Hartog, C. 1997. Is Sargassum muticum a threat to eelgrass beds? Aquatic Botany, 58(1), pp. 37-41.
den Hartog, C. & Phillips, R.C. 2001. Common structures and properties of seagrass beds fringing the coasts of the world. In: K. Reise (ed.) Ecological comparisons of sedimentary shores. Ecological Studies, vol. 151. Berlin: Springer.
Dexter, R.W. 1985. Changes in the standing crop of eelgrass, Zostera marina L., at Cape Ann, Massachusetts, since the epidemic of 1932. Rhodora, 87(851), pp. 357-366.
Dipper, F.A. & Beaver, R. 1999. Marine Nature Conservation Review Sector 12. Sealochs in the Clyde Sea: area summaries. Peterborough: JNCC.
Dipper, F.A. & Johnston, C.M. 2002. Marine Nature Conservation Review Sector 15. Sealochs in north-west Scotland: area summaries. Peterborough: JNCC.
Dipper, F. & Naylor, P. 2025. Coastal Seas. London: Bloomsbury Publishing.
Dipper, F.A., Howson, C.M. & Steele, D. 2008. Marine Nature Conservation Review Sector 13. Sealochs in west Scotland: area summaries. Peterborough: JNCC.
Don, G. 1813. Account of Native Plants in the county of Forfar, and the Animals found there. In: J. Headrick A General View of the Agriculture in the County of Angus, or Forfarshire. Edinburgh: P. Neill.
Dooley, F.D., Wyllie-Echeverria, S. & van Volkenburgh, E. 2013. Long-term seed storage and viability of Zostera marina. Aquatic Botany, 111, pp. 130-134.
Druce, G.C. 1903. Notes on the flora of Western Ross-shire. The Annals of Scottish Natural History, 47, pp. 166-175.
Druce, G.C. 1904. Notes on the Flora of Westerness. The Annals of Scottish Natural History, 49, pp. 36-42.
Druce, G.C. 1929. The Flora of West Ross. Arbroath: T. Buncle & Co.
Duarte, C.M., Middelburg, J.J. & Caraco, N. 2005. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences, 2, pp. 1–8.
Duffy, J.E., Hughes, A.R. & Moksnes, P.-O. 2014. Ecology of Seagrass Communities. In: M.D. Bertness, J.F. Bruno, B.R. Silliman & J.J. Stachowicz (eds.) Marine Community Ecology and Conservation. Sunderland: Sinauer Associates.
Duncan, K. 1991. The impact of land reclamation on the distribution of Zostera spp. in Nigg Bay. MSc Thesis, University of Aberdeen.
Duncan, C. 2003. Lamlash Bay Seasearch 2003 - Interim Report to Scottish Natural Heritage. Marine Conservation Society.
Edmondston, T. 1845. A Flora of Shetland: Comprehending a list of the flowering and cryptogamic plants of the Shetland Islands. Aberdeen: G. Clark.
Edwards, K.J. & Whittington, G. 2003. Vegetation Change. In: K.J. Edwards & I.B.M. Ralston (eds.) Scotland After the Ice Age: Environment, Archaeology and History 8000 BC - AD 1000. Edinburgh: Edinburgh University Press.
Egerton, J. 2011. Management of the seagrass bed at Porth Dinllaen. Initial investigation into the use of alternative mooring systems. Report for Gwynedd Council.
Eigaard, O.R., Bastardie, F., Hintzen, N.T., Buhl-Mortensen, L., Buhl-Mortensen, P., Catarino, R., Dinesen, G.E., Egekvist, J., Fock, H.O., Geitner, K., Gerritsen, H.D., González, M.M., Jonsson, P., Kavadas, S., Laffargue, P., Lundy, M., Gonzalez-Mirelis, G., Nielsen, J.R., Papadopoulou, N., Posen, P.E., Pulcinella, J., Russo, T., Sala, A., Silva, C., Smith, C.J., Vanelslander, B. & Rijnsdorp, A.D. 2017. The footprint of bottom trawling in European waters: Distribution, intensity, and seabed integrity. ICES Journal of Marine Science, vol. 74, no. 3, pp. 847-865.
Elliot, R.E. 1989. Birds of Islay. London: Christopher Helm.
Elwes, H.J. 1930. Memories of Travel, Sport, and Natural History. London: Ernest Benn.
Entec. 1996. Broad scale habitat mapping of intertidal and subtidal coastal areas: Loch Maddy, North Uist. Scottish Natural Heritage Research, Survey and Monitoring Report. No 76.
Environment Agency. 2019. Phosphorus and Freshwater Eutrophication Pressure Narrative.
Environment, Food & Rural Affairs Committee. 2022. Tree planting: Third Report of Session 2021–22. House of Commons.
Erftemeijer, P.L.A. & Lewis, R.R. 2006. Environmental impacts of dredging on seagrasses: A review. Marine Pollution Bulletin, 52, pp. 1553-1572.
ERT Scotland Ltd. 2007. Site Condition Monitoring: surveys of lagoons in the Vadills Lagoon Special Area of Conservation, July–August 2003. Scottish Natural Heritage Commissioned Report No. 209.
Evans, W. 1889. On the Occurrence of Zostera nana, Roth, in the Firth of Forth. Transactions of the Botanical Society of Edinburgh, 17(1-4), pp. 415-416.
Evans, D.M. & Rogers, A.P. 1996. The Effect of the Tawe Barrage on Estuarine Water Quality. In: N. Burt & J. Watts (eds.) Barrages, engineering design and environmental impacts. Chichester: John Wiley & Sons.
Evans, P.A., Evans, I.M. & Rothero, G.P. 2002. Flora of Assynt. Evans.
Farrell, P. & Fletcher, R. 2000. The biology and distribution of the kelp, Undaria pinnatifida Harvey Suringar, in the Solent. In: M. Collins & K. Ansell (eds.) Solent Science - A Review. Amsterdam: Elsevier Science.
Fenton, A. 1992. Shellfish as Bait: the Interface between Domestic and Commercial Fishing. In: T.C. Smout (ed.) Scotland and the Sea. Edinburgh: John Donald.
Finlay, S. 2014. Mapping Seagrass Distribution in the Firth of Forth. MSc Thesis, Heriot Watt University.
Fishery Board for Scotland. 1887. Summary of Reports by Officers of the Fishery Board on mussel beds in tidal waters and drying grounds for fishermen’s nets in Scotland. London: HMSO.
Fishery Board for Scotland. 1890. Annual report being for the year 1889 Vol. 8. Edinburgh: HMSO.
Foden, J., Brazier, P., Best, M., Scanlan, C. & Wells, E. 2010. Water Framework Directive Development of Classification Tools for Ecological Assessment: Intertidal Seagrass Marine Angiosperms. UKTAG Report: Seagrass v3.
Fonseca, M.S., Zieman, J.C., Thayer, G.W. & Fisher, J.S. 1983. The role of current velocity in structuring eelgrass Zostera marina L. meadows. Estuarine, Coastal and Shelf Science, 17, pp. 367-380.
Fonseca, M.S., Thayer, G.W. & Chester, A.J. 1984. Impacts of Scallop harvesting on Eelgrass (Zostera marina) meadows: Implications for management. North American Journal of Fish Management, 4, pp. 286-293.
Fonseca, M.S. & Uhrin, A.V. 2009. The status of eelgrass, Zostera marina, as bay scallop habitat: consequences for the fishery in the western Atlantic. Marine Fisheries Review, 71(3), pp. 20-33.
Fonseca, M., Piniak, G.A. & Cosentino-Manning, N. 2017. Susceptibility of seagrass to oil spills: A case study with eelgrass, Zostera marina in San Francisco Bay, USA. Marine Pollution Bulletin, 115(1), pp. 29-38.
Foot, C. 2012. Zostera Survey 2011 Udale and Nigg Bays. RSPB internal report.
Forbes, E. 1859. The Natural History of the European Seas. London: John van Voorst.
Forest Research. 2023. The UK Forestry Standard. Farnham: Forest Research.
Forrester, R.W., Andrews, I.J., McInerny, C.J., Murray, R.D., McGowan, R.Y., Zonfrillo, B., Betts, M.W., Jardine, D.C. & Grundy, D.S. 2007. The Birds of Scotland. Aberlady: The Scottish Ornithologists' Club.
Foster, R. & Davidson, K. 2018. Angiosperm Monitoring for the EU Water Framework Directive 2013 – 2015: Baseline seagrass surveys of the Montrose Basin, Eden Estuary, Forth Estuary and Loch Ryan. SEPA Report No. MB-01.2018.
Fox, A. 1996. Zostera exploitation by Brent Geese and Wigeon on the Exe Estuary, southern England. Bird Study, 43, pp. 257-268.
Fox, A.D, Yost, L. & Gilbert, G. 1986. A preliminary appraisal of the intertidal sea-grass resource in the Moray Firth. Nature Conservancy Council.
Fox, A.D., Ebbinge, B.S., Mitchell, C., Heinicke, T.A., Colhoun, K., Clausen, P., Dereliev, S., Farago, S., Koffijberg, K., Kruckenberg, H., Loonen, M.J.J.E., Madsen, J., Mooij, J., Musil, P., Nilsson, L., Pihl, S. & van der Jeugd, H. 2010. Current estimates of goose population sizes in western Europe, a gap analysis and an assessment of trends. Ornis Svecica, 20, pp. 115–127.
Fraser, M.W. & Kendrick, G.A. 2017. Belowground stressors and long-term seagrass declines in a historically degraded seagrass ecosystem after improved water quality. Nature: Scientific Reports, 7, No. 14469.
Gagnon, P.S., Vadas, R.L., Burdick, D.B. & May, B. 1980. Genetic identity of annual and perennial forms of Zostera marina L. Aquatic Botany, 8, pp. 157–162.
Gambi, M.C., Nowell, A.R.M. & Jumars, P.A. 1990. Flume observations on flow dynamics in Zostera marina eelgrass beds. Marine Ecology Progress Series, 61, pp. 159-169.
Ganter, B. 2000. Seagrass Zostera spp. as food for brent geese Branta bernicla: an overview. Helgoland Marine Research, 54, pp. 63–70.
Ganthy, F., Soissons, L., Sauriau, P., Verney, R. & Sottolichio, A. 2015. Effects of short flexible seagrass Zostera noltei on flow, erosion and deposition processes determined using flume experiments. Sedimentology, 62(4), pp. 997-1023.
Garner, N., Ross, P.M., Falkenberg, L.J., Seymour, J.R., Siboni, N. & Scanes, E. 2022. Can seagrass modify the effects of ocean acidification on oysters? Marine Pollution Bulletin, 177, No. 113438.
Gee, A.S. & Stoner, J.H. 1988. The effects of afforestation and acid deposition on the water quality and ecology of upland Wales. In: M.B. Usher & D.A.B. Thompson (eds.) Ecological Change in the Uplands. Oxford: Blackwell Scientific Publications.
Giesen, W.B.J.T., van Katwijk, M.M. and den Hartog, C. 1990. Temperature, salinity, insolation and wasting disease of eelgrass (Zostera marina L.) in the Dutch Wadden Sea in the 1930's. Netherlands Journal of Sea Research, 25, pp. 395-404.
Giesler, R.J., Allan, K. & Shucksmith, R. 2025. Searching for Shetland’s lost seagrass: establishing the baseline distribution and abundance of seagrass in the Shetland Islands. Shetland UHI report. p54.
Gilvear, D.J. 1994. River flow regulation. In: P. S. Maitland, P. J. Boon & D. S. McLusky (eds.) The Fresh Waters of Scotland. Chichester: John Wiley & Sons.
Gobler, C.J. 2020. Climate change and harmful algal blooms: insights and perspective. Harmful Algae, 91, No. 101731.
Godwin, H. 1975. The History of the British Flora: a factual basis for phytogeography. 2nd edition. Cambridge: Cambridge University Press.
Goodlad, C.A. 1971. Shetland Fishing Saga. Lerwick: The Shetland Times.
Graham, H.D. 1890. The Birds of Iona & Mull. Edinburgh: David Douglas.
Grayand, A.J. & Benham, P.E.M. 1990. Spartina anglica: a research review. ITE research publication no. 2. HMSO.
Green, E.P. & Short, F.T. (eds.) 2003. World Atlas of Seagrasses. Berkeley: University of California Press.
Green, A.E., Unsworth, R.K.F., Chadwick, M.A. & Jones, P.J.S. 2021. Historical analysis exposes catastrophic seagrass loss for the United Kingdom. Frontiers in Plant Science, 12: Article 629962
Groner, M.L., Eisenlord, M.E., Yoshioka, R.M., Fiorenza, E.A., Dawkins, P.D., Graham, O.J., Winningham, M., Vompe, A., Rivlin, N.D., Yang, B., Burge, C.A., Rappazzo, B., Gomes, C.P. & Harvell, C.D. 2021. Warming sea surface temperatures fuel summer epidemics of eelgrass wasting disease. Marine Ecology Progress Series, 679, pp. 47-58.
Gubbay, S. 1988. A Coastal Directory for Marine Nature Conservation. Ross-on-Wye: Marine Conservation Society.
Gutiérrez, J.L., Jones, C.G., Byers, J.E., Arkema, K.K., Berkenbusch, K., Commito, J.A., Duarte, C.M., Hacker, S.D., Lambrinos, J.G., Hendriks, I.E., Hogarth, P.J., Palomo, M.G. & Wild, C. 2011. Physical Ecosystem Engineers and the Functioning of Estuaries and Coasts. In: E. Wolanski & D.S. McLusky (eds.) Treatise on Estuarine and Coastal Science, Volume 7. Waltham: Academic Press.
Halcrow, A. 1950. The Sail Fishermen of Shetland. Lerwick: T & J Manson.
Hall-Spencer, J. & Bamber, R. 2007. Effects of salmon farming on benthic crustacea. Ciencias Marinas, 334, pp. 353-366.
Hammerton, D. 1994. Domestic and industrial pollution. In: P. S. Maitland, P. J. Boon & D. S. McLusky (eds.) The Fresh Waters of Scotland. Chichester: John Wiley and Sons.
Hannah, A. 2019. Isle of Bute Flora. Trollius Publications.
Harlin, M.M. & Thorne-Miller, B. 1982. Seagrass-sediment dynamics of a flood-tidal delta in Rhode Island USA. Aquatic Botany, 14, pp. 127-138.
Harries, D.B., Harrow, S., Wilson, J.R., Mair, J.M. & Donnan, D.W. 2007a. The establishment of the invasive alga Sargassum muticum on the west coast of Scotland: a preliminary assessment of community effects. Journal of the Marine Biological Association of the United Kingdom, 875, pp. 1057-1067.
Harries, D.B., Malthus, T.J., Karpouzli, E., Moore, C.G., Lyndon, A.R., Mair, J.M., Service, M., Mitchell, A. 2007b. Biotope mapping of the Sound of Barra, Scotland. Scottish Natural Heritage Commissioned Report No.258.
Haven, D.S. & Morales-Alamo, R. 1966. Aspects of biodeposition by oysters and other invertebrate filter feeders. Limnology and Oceanography, 11, pp. 487-498.
Hawker, D. 1993. Eelgrass Zostera in The Solway Firth. Scottish Natural Heritage Commissioned Report.
Hawker, D. 1994. Solway Firth Zostera Survey Monitoring Report. Scottish Natural Heritage Commissioned Report.
Hemminga, M.A. & Duarte, C.M. 2000. Seagrass Ecology. Cambridge: Cambridge University Press.
Henderson, A.R. & Hamilton, J.D. 1986. The status of fish populations in the Clyde Estuary. Proceedings of the Royal Society of Edinburgh, 90B, pp. 157-170.
Hennedy, R. 1865. The Clydesdale flora: a description of the flowering plants and ferns of the Clyde district. Glasgow: David Robertson.
Hensel, M.J.S., Patrick, C.J., Orth, R.J., Wilcox, D.J., Dennison, W.C., Gurbisz, C., Hannam, M.P., Landry, J.B., Moore, K.A., Murphy, R.R., Testa, J.M., Weller, D.E. & Lefcheck, J.S. 2023. Rise of Ruppia in Chesapeake Bay: Climate change-driven turnover of foundation species creates new threats and management opportunities. Proceedings of the National Academy of Sciences of the United States of America. 120 23: e2220678120.
Hershner, C., Ward, K., Illowsky, J., Delistraty, D. & Martorana, J. 1983. The effects of Atrazine on Zostera marina in the Chesapeake Bay, Virginia. Final Report. US EPA, Chesapeake Bay Program, Virginia Institute of Marine Science.
Heslop-Harrison, J.W. 1937. The Natural History of the Isle of Raasay and of the adjoining Islands of South Rona, Scalpay, Fladday, and Longay. Proceedings of the University of Durham Philosophical Society, 10(5), pp. 246-351.
Heslop-Harrison, J.W. 1958. Botanical investigations in the Isles of Lewis, Harris and Great Bernera v-c 110 and in the Isle of Rhum v-c 104 in 1957. Proceedings of the University of Durham Philosophical Society, 13A, pp. 80-88.
Hill, J.M. & Sabatini, M. 2008. Nephrops norvegicus Norway lobster. In: H. Tyler-Walters Marine Life Information Network: Biology and Sensitivity Key Information Reviews. Plymouth: Marine Biological Association of the United Kingdom (Accessed on 2nd December 2024).
Hiscock, K. 1989. Marine biological survey of upper Whiteness Voe, Shetland Islands. 13 & 14 June 1989. Nature Conservancy Council, CSD Report, No 973. Marine Nature Conservation Review, Report No MNCR/SR/009/89.
Hodge, J.G.W. 1993. A method for Surveying the Distribution of Zostera at Udale Bay. RSPB Internal Report.
Holmer, M. & Bondgaard, E.J. 2001. Photosynthetic and growth response of eelgrass to low oxygen and high sulfide concentrations during hypoxic events. Aquatic Botany, 70, pp. 29–38.
Holmer, M., Frederiksen, M.S. & Møllegaard, H. 2005. Sulfur accumulation in eelgrass Zostera marina L. and effect of sulfur on eel grass growth. Aquatic Botany, 81, pp. 367–379.
Hooker, W.J & Arnott, G.A.W. 1850. The British Flora: Comprising the Phaenogamous or Flowering Plants and the Ferns. 6th edition. London: Longman, Brown, Green & Longmans.
Hooker, W.J & Arnott, G.A.W. 1855. The British Flora: Comprising the Phaenogamous or Flowering Plants and the Ferns. 7th edition. London: Longman, Brown, Green & Longmans.
Hopkins, S.H. 1957. Parasitism. In: J.W. Hedgepeth (ed.) Treatise on Marine Ecology and Paleoecology, GSA Memoir 67, Vol. 1. New York: Geological Society of America.
Hopkirk, T. 1813. Flora Glottiana: A Catalogue of the Indigenous Plants on the Banks of the River Clyde, and in the Neighbourhood of the City of Glasgow. Glasgow: John Smith & Son.
Hosokawa, S., Nakaoka, M., Miyoshi, E. & Kuwae, T. 2015. Seed dispersal in the seagrass Zostera marina is mostly within the parent bed in a protected bay. Marine Ecology Progress Series, 523, pp. 41–56.
Howard. S, Baker, J.M. & Hiscock, K. 1989. The Effect of Oil and Dispersants on Seagrass in Milford Haven. In: B. Dicks (ed.) Ecological Impacts of the Oil Industry. London: Wiley & Sons.
Howson, C.M. 1989. Surveys of Scottish Sealochs, Loch Ryan. Contractor: University Marine Biological Station, Millport Nature Conservancy Council, CSD Report, No. 953.
Howson, C.M. 1999. Marine Nature Conservation Review Sector 1. Shetland: area summaries. Peterborough: JNCC.
Howson, C. M., Clark, L., Mercer, T.S. & James, B. 2012. Marine biological survey to establish the distribution and status of fan mussels Atrina fragilis and other Marine Protected Area MPA search features within the Sound of Canna, Inner Hebrides. Scottish Natural Heritage Commissioned Report No. 438.
Hubbard, J.C.E. & Stebbings, R.E. 1967. Distribution, dates of origin and acreage of Spartina townsendii s.l. marshes in Great Britain. Proceedings of the Botanical Society of the British Isles, 7, pp. 1-7.
Hugget, D. 1996. The Habitats Directive: a barrier to barrages? In: N. Burt & J. Watts (eds.) Barrages, engineering design and environmental impacts. Chichester: John Wiley & Sons.
Hughes, R., Lloyd, D., Ball, L. & Emson, D. 2000. The effects of the polychaete Nereis diversicolor on the distribution and transplanting success of Zostera noltii. Helgoland Marine Research, 54, pp. 129-136.
Hughes, R.G. 2001. Biological and physical processes that affect saltmarsh erosion and saltmarsh restoration: Development of hypothesis. In: K. Reise (ed.) Ecological comparisons of sedimentary shores. Ecological Studies, vol. 151. Berlin: Springer.
Hughes, R.J. & Paramor, O.A.L. 2004. On the loss of saltmarshes in south-east England and methods for their restoration. Journal of Applied Ecology, 41, pp. 440-448.
ICH Scotland. 2025. Intangible Cultural Heritage in Scotland Inventory [online] (Accessed 21st October 2025).
Ingram, R. & Noltie, H.J. 1981. The Flora of Angus Forfar, v.c. 90. Dundee: Dundee Museum & Art Gallery.
Jackson, E. 1999. The underwater meadows of Orkney. Bulletin of the Orkney Field Club, pp. 10-13.
Jackson, J.B.C. 2001. What was natural in the coastal oceans? PNAS, 98(10), pp. 5411-5418.
Jackson, D.W.T. & Short, A.D. 2020. Sandy Beach Morphodynamics. London: Elsevier.
Jacobs, R.P.W.M. 1980. Effects of the ‘Amoco Cadiz’ oil spill on the seagrass community at Roscoff with special reference to the benthic infauna. Marine Ecology Progress Series, 2, pp. 207–212.
Jacobs, R.P.W.M., den Hartog, C., Braster, B.F. & Carriere, F.C. 1981. Grazing of the sea grass Zostera noltii by birds at Terschelling Dutch Wadden Sea. Aquatic Botany, 10, pp. 241-259.
James, B. 2004. North-west Scotland subtidal seagrass bed survey 2004. Scottish Natural Heritage Commissioned Report No. 76.
Jarvis, J. & Moore, K. 2010. The role of seedlings and seed bank viability in the recovery of Chesapeake Bay, USA, Zostera marina populations following a large-scale decline. Hydrobiologia, 649, pp. 55-68.
Jensen, J.-K., Thorning-Lund, F. & Hammer, S. 2023. Supplement to Faroe Islands botanical list with 64 species or subspecies including rare, new and potentially invasive species with comments. Nordic Journal of Botany, 2023(7): e03586.
Jones, B.L. & Unsworth, R.K.F. 2016. The perilous state of seagrass in the British Isles. Royal Society Open Science, 3, No. 150596.
Jones, K.L., Hartl, M.G.J., Bell, M.C. & Capper, A. 2020. Microplastic accumulation in a Zostera marina L. bed at Deerness Sound, Orkney, Scotland. Marine Pollution Bulletin, 152, No. 110883.
Källström, B., Nyqvist, A., Åberg, P., Bodin, M. & André, C. 2008. Seed rafting as a dispersal strategy for eelgrass Zostera marina. Aquatic Botany, 88(2), pp. 148-153.
King, J. & Scammell, E. 2017. Place-names of Colonsay and Oronsay. Perth: Scottish Natural Heritage.
Knight, K. & Holden, E. 2022. Seagrass Restoration 2022 Project Update. Seawilding.
Knight, K. & Holden, E. 2024. Seagrass Restoration 2023 Project Update. Seawilding.
Knox, R.B. 1959. Flora of the Isle of Jura. 1. Flowering Plants and Ferns. Transactions and Proceedings of the Botanical Society of Edinburgh vol 37, part 4: pp 251–256.
Krause-Jensen, D., Carstensen, J., Nielsen, S.L., Dalsgaard, T., Christensen, P.B., Fossing, H. & Rasmussen, M.B. 2011. Sea bottom characteristics affect depth limits of eelgrass Zostera marina. Marine Ecology Progress Series, 425, pp. 91-102.
Langsbury, G. & Ogilvie, M.A. 2006. The Birds of Islay A Celebration in Photographs. Lochindaal Press.
Lavaut, E., Guillemin, M.-L., Colin, S., Faure, A., Coudret, J., Destombe, C. & Valero, M. 2022. Pollinators of the sea: A discovery of animal-mediated fertilization in seaweed. Science, 377(6605), pp. 528-530.
Lee, J.R. 1933. Flora of the Clyde Area. Glasgow: John Smith & Sons.
Łękawska, M. & Kay, L. In prep. Overview of Arran seagrass bed data. COAST draft report.
Lillebø, A.I., Flindt, M.R., Cardoso, P., Leston, S., Dolbeth, M., Pereira, M.E., Duarte, A.C. & Pardal, M.A. 2011. Restoration of Seagrass Community to Reverse Eutrophication in Estuaries. In: E. Wolanski & D.S. McLusky (eds.) Treatise on Estuarine and Coastal Science, Volume 10. Waltham: Academic Press.
Lilley, R.J. & Unsworth, R.K.F. 2014. Atlantic Cod Gadus morhua benefits from the availability of seagrass Zostera marina nursery habitat. Global Ecology and Conservation, 2, pp. 367-377.
Lyngby, J.E. & Brix, H. 1984. The uptake of heavy metals in eelgrass Zostera marina and their effect on growth. Ecological Bulletins, 36, pp. 81–89.
Mackey, E.C., Shewry, M.C. & Tudor, G.J. 1998. Land Cover Change: Scotland from the 1940s to the 1980s. Scottish Natural Heritage. Edinburgh: TSO.
Macvicar, S.M. 1897. On the Flora of Eigg. Annals of Scottish Natural History. No 21: 110-117.
Madden, B., Jennings, E. & Jeffrey, D.W. 1993. Distribution and ecology of Zostera in Co. Dublin. Irish Naturalist Journal, 24(8), pp. 303-310.
Madsen, J. 1988. Autumn feeding ecology of herbivorous wildfowl in the Danish Wadden Sea and impact of food supplies and shooting on migration. Danish Review of Game Biology, 13, pp. 1-32.
Malthus, T.J., Harries, D.B., Karpouzli, E., Moore, C.G., Lyndon, A.R., Mair, J.M., Foster-Smith, B., Sotheran, I. & Foster-Smith, D. 2006. Biotope mapping of the Sound of Harris, Scotland. Scottish Natural Heritage Commissioned Report No. 212.
Marbà, N. & Duarte, C.M. 1998. Rhizome elongation and seagrass clonal growth. Marine Ecology Progress Series, 174, pp. 269–280.
Marsden, M.W. & Mackay, D.W. 2001. Water quality in Scotland: the view of the regulator. Science of the Total Environment, 265(1-3), pp. 369-386.
Marshall, E.S. 1901. Plants of North Scotland 1900. Journal of Botany, 39: pp. 266-275.
Marshall, E.S. & Shoolbred, W.A. 1898. Notes of a tour in N. Scotland 1897. Journal of Botany, 36, pp. 166-177.
Martin, I.H. 1934. The Field-Club Flora of The Lothians. Edinburgh: William Blackwood & Sons.
Martin, A.C. 1954. A clue to the eelgrass mystery. Transactions of the 19th North American Wildlife and Natural Resources Conference, 19, pp. 441-449.
MBHS. 2004. Ebb and Flow: Aspects of the History of Montrose Basin. Angus: The Pinkfoot Press.
McCallum Webster, M. 1978. Flora of Moray, Nairn and East Inverness. Aberdeen: Aberdeen University Press.
McCorry, M.J., Curtis, T.G.F. & Otte, M.L. 2003. Spartina in Ireland. In: M.L. Otte (ed.) Wetlands of Ireland Distribution, Uses and Economic Value. Dublin: University College Dublin Press.
McIntyre, F., Fernandes, P.G. & Turrell, W.R. 2012. Scottish Marine and Freshwater Science Report Volume 3 Number 3 Clyde Ecosystem Review. Scottish Government.
McKirdy, A. 2007. Land of Mountain and Flood: The Geology and Landforms of Scotland. Edinburgh: Birlinn.
McLachlan, A. & Defeo, O. 2018. The Ecology of Sandy Shores. 3rd edition. London: Academic Press.
McLusky, D.S. & Roddie, K. 1983. Montrose Basin Survey 1982. Unpublished report to Nature Conservancy Council.
McLusky, D.S. 1989. The Estuarine Ecosystem. 2nd edition. Glasgow: Blackie.
McNeill, M. 1910. Colonsay; one of the Hebrides; its plants: their local names and uses; legends, ruins, and place-names; Gaelic names of birds, fishes, etc.; climate, geological formation, etc. Edinburgh: David Douglas.
McRoy, C.P. 1969. Eelgrass under the Arctic winter ice. Nature, 224, pp. 818-819.
McRoy, C.P. 1970. On the biology of eelgrass in Alaska. PhD Thesis, University of Alaska.
Met Office. 2018. UKCP18 Factsheet: Sea level rise and storm surge (Accessed 23rd February 2025).
Met Office. 2024. Climate change drives increase in storm rainfall (Accessed 16th May 2025).
Meysick, L., Infantes, E., Rugiu, L., Gagnon, K. & Boström, C. 2022. Coastal ecosystem engineers and their impact on sediment dynamics: Eelgrass-bivalve interactions under wave exposure. Limnology and Oceanography, 67(3), pp. 621-633.
Millar, J.M. 1993. Flowers of Iona. 2nd edition. Iona: The New Iona Press.
Miller, H. 1854. My School and Schoolmasters. Boston: Gould and Lincoln.
Miller, H. 1857. The Testimony of the Rocks or, Geology in its Bearings on the two Theologies, Natural and Revealed. Boston: Gould and Lincoln.
Millier, H.K.G.R., Hooda, P.S. & Downward, S.R. 2010. The impact of treated sewage wastewater discharges on the phosphorus levels and hydrology of two second order rivers flowing into the Thames. Journal of Environmental Monitoring, 12(6), pp. 1307-1314.
Mitchell, R., Earll, R.C. & Dipper, F.A. 1983. Shallow sublittoral ecosystems in the Inner Hebrides. Proceedings of the Royal Society of Edinburgh. Section B. Biological Sciences, 83, pp. 161–183.
Molin, J.M., Groth-Andersen, W.E., Hansen, P.J., Kühl, M. & Brodersen, K.E. 2023. Microplastic pollution associated with reduced respiration in seagrass Zostera marina L. and associated epiphytes. Frontiers in Marine Science, 10, 1216299.
Moore, H.B. 1958. Marine Ecology. New York: Wiley.
Moore, C.G., Saunders, G.R. and Harries, D.B. 1998. The status and ecology of reefs of Serpula vermicularis L. (Polychaeta: Serpulidae) in Scotland. Aquatic Conservation of Marine and Freshwater Ecosystems, 8: pp. 645-656.
Moore, K.A., Shields, E.C. & Parrish, D.B. 2014. Impacts of Varying Estuarine Temperature and Light Conditions on Zostera marina (Eelgrass) and its Interactions With Ruppia maritima (Widgeongrass). Estuaries and coasts, 37(1), pp. 20-30.
Morris, R. 2010. Sea-level rise: implications for people and wildlife. British Wildlife, 21(3), pp. 170-176.
Muehlstein, L.K., Porter, D. & Short. F.T. 1988. Labyrinthula sp. a marine slime mould producing the symptoms of wasting disease in eelgrass, Zostera marina. Marine Biology, 99, pp. 465-472.
Muehlstein, L.K., Porter, D. & Short, F. T. 1991. Labyrinthula zosterae sp. nov., the causative agent of wasting disease of eelgrass, Zostera marina. Mycologia, 83(2), pp. 180-191.
Murray, E., Daikin, M.J., Fortune, F. & Begg, K. 1999. Marine Nature Conservation Review Sector 2. Orkney: area summaries. Peterborough: JNCC.
Murray, R.D., Andrews, I.J. & Holling, M. 2019. Birds of South Scotland-east Scotland 2007-13: a tetrad atlas of the birds of Lothian and Borders. Aberlady: The Scottish Ornithologists Club.
Nacken, M. & Reise, K. 2000. Effects of herbivore birds on intertidal seagrass beds in the northern Wadden Sea. Helgoland Marine Research, 54, pp. 87-94.
NatureScot. 2024. South Arran MPA Conservation and Management Advice. Published in November 2024, Version 6.
NatureScot. 2025. Sound of Barra SAC Conservation and Management Advice. Published in April 2025, Version 12.
Navarro, N., Leakey, R.J.G. & Black, K.D. 2008. Effect of salmon cage aquaculture on the pelagic environment of temperate coastal waters: seasonal changes in nutrients and microbial community. Marine Ecological Progress Series, 361, pp. 47-58.
NC (Nature Conservancy). 1972. A Prospectus for Nature Conservation within the Moray Firth. Natural Environment Research Council.
NCC (Nature Conservancy Council). 1978. Nature Conservation within the Moray Firth Area. Natural Environment Research Council.
Nejrup, L.B. & Pedersen, M.F. 2008. Effects of salinity and water temperature on the ecological performance of Zostera marina. Aquatic Botany, 88, pp. 239-246.
New Statistical Account of Scotland. 1845. The new statistical account of Scotland, Volume IV, Part 2, p. 583. Edinburgh: William Blackwood & Sons.
Newell, R.I.E. 1988. Ecological changes in Chesapeake Bay: are they the result of overharvesting the American oyster, Crassostrea virginica. In: M.P. Lynch & E.C. Krome (eds.) Understanding the Estuary: Advances in Chesapeake Bay Research. Baltimore: Chesapeake Bay Research Consortium.
Nicol, E.A.T. 1936. The Fauna of Loch Bee. The Scottish Naturalist. No. 217, pp. 131-134.
Nielsen, L & Dahllöf, I. 2007. Direct and indirect effects of the herbicides Glyphosate, Bentazone and MCPA on eelgrass Zostera marina. Aquatic Toxicology, 82(1), pp. 47-54.
Ogilvie, M.A. 1992. Recent status change of some birds on Islay. Scottish Birds, 1, pp. 240-259.
Ogilvie, M.A. 1995. The Wild Flowers of Islay: A Checklist. Lochindaal Press.
Ogilvie, M.A. & Matthews, G.V.T. 1969. Brent Geese, Mudflats and Man. Wildfowl, 20, pp. 119-125.
Olsen, J.L., Coyer, J.A., Stam, W.T., Moy, F.E., Christie, H. & Jørgensen, N.M. 2013. Eelgrass Zostera marina populations in northern Norwegian fjords are genetically isolated and diverse. Marine Ecology Progress Series, 486, pp. 121-132.
Oncken, N.S., Kristensen, E. & Quintana, C.O. 2022. Spatial overlap between lugworm Arenicola marina and eelgrass Zostera marina distribution in coastal waters: The role of environmental stressors. Estuarine, Coastal and Shelf Science, Volume 272, e107886.
Orth, R.J. 1977. The importance of sediment stability in seagrass communities. In:B.C. Coull (ed.) Ecology of marine benthos. Columbia: University of South Carolina Press.
Orth, R.J., Moore, K.A. 1983. Chesapeake Bay: An Unprecedented Decline in Submerged Aquatic Vegetation. Science, 222, pp. 51-53.
Orth, R., Luckenbach, M. & Moore, K. 1994. Seed Dispersal in a Marine Macrophyte: Implications for Colonization and Restoration. Ecology, 75, pp. 1927-1939.
OSPAR Commission. 2008: Case Reports for the OSPAR List of Threatened and/or Declining Species and Habitats.
OSPAR Commission. 2009. Background Document for Zostera beds, Seagrass beds.
OSPAR Commission. 2023. QSR Synthesis Report 7. Preventing Pollution to Achieve Clean Seas Accessed on 23rd September 2025
Ostenfeld, C.H. 1908. On the ecology and distribution of the grass wrack Zostera marina in Danish waters. Report of the Danish Biological Station, 16.
Page, F.H. & Burridge, L. 2014. Estimates of the effects of sea lice chemical therapeutants on non-target organisms associated with releases of therapeutants from tarped net-pens and well-boat bath treatments: a discussion paper. DFO Canadian Science Advisory Secretariat Research Document 2014/103. Ottawa: Fisheries and Oceans Canada.
Pearman, D.A. & Preston, C.D. 2000. A Flora of Tiree, Gunna and Coll. Privately published by the authors.
Pearman, D.A., Preston, C.D., Rothero, G.P. & Walker, K.J. 2008. The Flora of Rum An Atlantic Island Reserve. Privately published by the authors.
Pearman, D.A. & Walker, K.J. 2004. An examination of J.W. Heslop Harrison’s unconfirmed plant records from Rum. Watsonia, 25, pp. 45–63.
Pedersen, O., Binzer, T. & Borum, J. 2004. Sulphide intrusion in eelgrass Zostera marina L. Plant, Cell & Environment, 27, pp. 595–602.
Penhale, P.A. & Wetzel, R.G. 1983. Structural and functional adaptations of eelgrass Zostera marina L. to the anaerobic sediment environment. Canadian Journal of Botany, 615, pp. 1421-1428.
Perkins, E.J. 1973. The Marine Fauna and Flora of the Solway Firth. Dumfries: The Dumfriesshire and Galloway Antiquarian Society.
Perkins, E.J. 1988. The Impact of Suction Dredging Upon the Population of Cockles, Cerastoderma Edule, in Auchencairn Bay, 1988. Internal NCC report.
Petersen, C.G.J. 1914. IX Om bændeltangens (Zostera marina) Aars-produktion i de danske farvande. In: H.F.E Jungersen & E. Warming (eds.) Mindeskrift i anledning af hundredâret for Japetus Steenstrups fodsel. Kobenhavn: G.E.C. Gad.
Petersen, C.G.J. 1918. The sea bottom and its production of fish food: a summary of the work done in connection with valuation of Danish waters from 1883 to 1917. Report of the Danish Biological Station, 25, pp. 1-82.
Peterson, C.H., Luettich, J.R.A., Micheli, F. & Skilleter, G.A. 2004. Attenuation of water flow inside seagrass canopies of differing structure. Marine Ecology Progress Series, 268, pp. 81–92.
Philippart, C.J.M. 1994. Interactions between Arenicola marina and Zostera noltii on a tidal flat in the Wadden Sea. Marine Ecology Progress Series, 111, pp. 251-257.
Philippart, C.J.M. 1995. Seasonal variation in growth and biomass of an intertidal Zostera noltii stand in the Dutch Wadden Sea. Netherlands Journal of Sea Research, 33, pp. 205-218.
Philippart, C.J.M. & Dijkema, K.S. 1995. Wax and wane of Zostera noltii Hornem. in the Dutch Wadden Sea. Aquatic Botany, 49(4), pp. 255-268.
Phillips, R.C., Grant, W.S. & McRoy, C.P. 1983. Reproductive strategies of eel grass (Zostera marina L.). Aquatic Botany, 16: pp. 1-20.
Pidgeon, E. 2009. Carbon sequestration by coastal marine habitats: Important missing sinks. In: D.d’A. Laffoley & G. Grimsditch (eds.) The Management of Natural Coastal Carbon Sinks. Gland, Switzerland: IUCN.
Pihl, L., Baden, S., Kautsky, N., Rönnbäck, P., Söderqvist, T., Troell, M. & Wennhage, H. 2006. Shift in fish assemblage structure due to loss of seagrass Zostera marina habitats in Sweden. Estuarine, Coastal and Shelf Science, 67(1–2), pp. 123-132.
Pistorius, P.A., Follestad, A. & Taylor, F.E. 2006. Temporal changes in spring migration phenology in the Norwegian Greylag Goose, Anser anser, 1971-2004. Wildfowl, 56, pp. 23-36.
Porley, R. & Hodgetts, N. 2009. Mosses and Liverworts. London: Collins.
Project Seagrass. 2024. Project Bilearch Report for The Botanist Foundation.
Purseglove, J. 1989. Taming the Flood: History and Natural History of Rivers and Wetlands. Oxford: Oxford University Press.
Qiao, Y., Zhang, Y., Xu, S., Yue, S., Zhang, X., Liu, M., Sun, L., Jia, X. & Zhou, Y. 2022. Multi-leveled insights into the response of the eelgrass Zostera marina L to Cu than Cd exposure. Science of The Total Environment, 845, 157057.
Quinteros Peñafiel, C.V., Riddiford, N.J. & Twyford, A.D. 2017. A floristic survey of Fair Isle. New Journal of Botany, 7(2–3), pp. 101–111.
Rae, P. 1979. The seagrasses of the Moray Firth: their ecology and relationship to adjacent industrial development. PhD Thesis, University of Aberdeen.
Raffaelli, D., Hull, S. & Milne, H. 1989. Long-term changes in nutrients, weed mats and shorebirds in an estuarine system. Cahiers de Biologie Marine, 30, pp. 259-270.
Raffaelli, D. & Hawkins, S. 1996. Intertidal Ecology. London: Chapman and Hall.
Ralph, P. & Short, F.T. 2002. Impact of the wasting disease pathogen, Labryrinthula on the photobiology of eelgrass Zostera marina. Marine Ecology Progress Series, 226, pp. 265-271.
Ralph, P.J., Tomasko, D., Moore, K., Seddon, S. & Macinnis-Ng, C. 2006. Human Impacts on Seagrasses: Eutrophication, Sedimentation, and Contamination. In: A.W.D. Larkum, R.J. Orth & C.M. Duarte (eds.) Seagrasses: Biology, Ecology and Conservation. Dordrecht: Springer.
Ranwell, D.S. & Downing, B.M. 1959. Brent goose Branta bernicla L. winter feeding pattern and Zostera resources at Scolt Head Island, Norfolk. Animal Behaviour, 7(1–2), pp. 42-48.
Ranwell, D., Wyer, D., Boorman, L., Pizzey, J. & Waters, R. 1974. Zostera transplants in Norfolk and Suffolk, Great Britain. Aquaculture, 4, pp. 185-198.
Rasmussen, E. 1973. Systematics and ecology of the Isefjord marine fauna Denmark: with a survey of the eelgrass Zostera vegetation and its communities. Ophelia, 111, pp. 1–507.
Rasmussen, E. 1977. The wasting disease of eelgrass Zostera marina and its effects on environmental factors and fauna. In: C.P. McRoy & C. Helfferich (eds.) Seagrass Ecosystems: A Scientific Perspective. New York: Marcel Dekker.
Rasmusson, L.M., Buapet, P., George, R., Gullström, M., Gunnarsson, P.C. & Björk, M. 2020. Effects of temperature and hypoxia on respiration, photorespiration, and photosynthesis of seagrass leaves from contrasting temperature regimes. ICES Journal of Marine Science, 77, pp. 2056-2065.
Ratcliffe, D. 2007. Galloway and the Borders. London: HarperCollins.
Raybould, A.F., Gray, A.J., Lawrence, M.J. & Marshall, D.F. 1991. The evolution of Spartina anglica C.E. Hubbard (Gramineae): genetic variation and status of the parental species in Britain. Biological Journal of the Linnean Society, 44(4), pp. 369-380.
Rayer, C. 2024. Mapping of Intertidal Seagrass Beds in the Firth of Forth. MSc Thesis, Heriot Watt University.
Reidenbach, M.A. & Thomas, E.L. 2018. Influence of the seagrass, Zostera marina, on wave attenuation and bed shear stress within a shallow coastal bay. Frontiers in Marine Science, 5, Article 397.
Reise, K. 1985. Tidal flat ecology - An experimental approach to species interactions. Ecological Studies, 54, pp. 1-191.
Renn, C.E. 1935. A mycetozoan parasite of Zostera marina. Nature, 135, pp. 544-545.
Renn, C.E. 1936. The wasting disease of Zostera marina: a phytological investigation of the diseased plant. The Biological Bulletin, 70, pp. 148-158.
Rennie, A.F. & Hansom, J.D. 2011. Sea level trend reversal: Land uplift outpaced by sea level rise on Scotland’s coast. Geomorphology, 125, pp. 193-202.
Reusch, T.B. & Williams, S.L. 1998. Variable responses of native eelgrass Zostera marina to a non-indigenous bivalve Musculista senhousia. Oecologia, 113(3), pp. 428-441.
Reusch, T., Boström, C., Stam, W.T. & Olsen, J. 1999. An ancient eelgrass clone in the Baltic Sea. Marine Ecology Progress Series, 183, pp. 301-304.
Reusch, H., Ehlers, A., Hämmerli, A. & Worm, B. 2005. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proceedings of the National Academy of Sciences of the United States of America, 102(8), pp. 2826-2831.
Reynolds, C., Miranda, N.A.F. & Cumming, G.S. 2015. The role of waterbirds in the dispersal of aquatic alien and invasive species. Diversity and Distributions, 21, pp. 744-754
Robertson, A.I. & Mann, K.H. 1984. Disturbance by ice and life-history adaptations of the seagrass Zostera marina. Marine Biology, 80, pp. 131–141.
Rogers, H.A. 2024. Mapping of Intertidal Seagrass Beds in the Forth Estuary. MSc Thesis, Heriot Watt University.
Rothschild, B.J., Ault, J.S., Goulletquer, P. & Heral, M. 1994. Decline of the Chesapeake Bay oyster population: a century of habitat destruction and overfishing. Marine Ecology Progress Series, 111, pp. 29-39.
Ruckelshaus, M.H. 1996. Estimation of genetic neighborhood parameters from pollen and seed dispersal in the marine angiosperm Zostera marina L. Evolution, 50(2), pp. 856-864.
Ryumina, A., Tishenko, P., Shkirnikova, E. & Barabanshchikov, Y. 2024. Heavy metal content in the seagrass Zostera marina Linnaeus, 1753 in Voevoda Bay Amur Bay, Sea of Japan. Russian Journal of Marine Biology, 50, pp. 156-163.
Salmon, C.E. 1900. Plant notes from Sutherland and Cantire. Journal of Botany. 38, pp 299-303.
Salo, T., Pedersen, M.F. & Boström, C. 2014. Population specific salinity tolerance in eelgrass (Zostera marina). Journal of Experimental Marine Biology and Ecology, 461, pp. 425−429.
Saxby, A. 1903. Edmonston's flora of Shetland : comprehending a list of the prevalent wild-flowers, horse-tails, club-mosses and ferns of the Shetland Isles. Edinburgh: Oliphant, Anderson & Ferrier.
Scarlett, A., Galloway, T.S., Canty, M., Smith, E.L., Nilsson, J. & Rowland, S.J. 2005. Comparative toxicity of two oil dispersants, superdispersant-25 and corexit 9527, to a range of coastal species. Environmental Toxicology and Chemistry, 24, pp. 1219–1227.
Schanz, A., Polte, P. & Asmus, H. 2002. Cascading effects of hydrodynamics on an epiphyte–grazer system in intertidal seagrass beds of the Wadden Sea. Marine Biology, 141, pp. 287–297.
Schenck, F.R., DuBois, K., Kardish, M.R., Stachowicz, J.J. & Hughes, A.R. 2023. The effect of warming on seagrass wasting disease depends on host genotypic identity and diversity. Ecology, 104(3), e3959.
Schou, J.C., Moeslund, B., van de Weyer, K., Wiegleb, G., Lansdown, R.V., Holm, P., Baastrup-Spohr, L. & Sand-Jensen, K. 2023. Aquatic Plants of Northern and Central Europe including Britain and Ireland. Princeton: Princeton University Press.
Schwientek, M., Rügner, H., Haderlein, S.B., Schulz, W., Wimmer, B., Engelbart, L., Bieger, S. & Huhn, C. 2024. Glyphosate contamination in European rivers not from herbicide application? Water Research, 263(1): 122140.
Scot-Skirving. 1878. Notes on the natural history of Islay. Proceedings of the Royal Physical Society of Edinburgh, 5, pp. 35-43.
Scotland Food and Drink. 2016. Aquaculture Growth to 2030: A Strategic Plan for Farming Scotland’s Seas.
Scott, W. 2011. Some Aspects of the Botany of the Shetland Islands. Scalloway: Privately published.
Scott, W. & Palmer, R. 1987. The Flowering Plants of the Shetland Islands. Lerwick: The Shetland Times.
Scott, W., Harvey, P., Riddington, R. & Fisher, M. 2002. Rare Plants of Shetland. Lerwick: The Shetland Times.
Scottish Government, 2011. Scotland's Marine Atlas: Information for The National Marine Plan (Accessed 8th January 2025).
Scottish Government, 2020a. Marine Scotland Assessment. Nutrient inputs in water (Accessed 8th January 2025).
Scottish Government, 2020b. Marine Scotland Assessment. Intertidal seagrass (Accessed 8th January 2025).
Scottish Government, 2020c. Marine Scotland Assessment. Biogenic Habitats. Accessed 3rd May 2026).
Scottish Government, 2023. Wild Salmon Strategy: Implementation Plan 2023–2028 (Accessed 8th January 2025).
Scottish Government, 2024. Marine Protected Areas MPAs [online] (Accessed 11th December 2024).
Scottish mussel and bait beds committee. 1889. Report of the committee appointed by the Secretary for Scotland to inquire into the condition of the Scottish mussel and bait beds, together with evidence and appendix. C. 5660. London: HMSO.
Scottish Natural Heritage. 2011. Loch Fleet: Site of Special Scientific Interest. Site Management Statement.
Seawilding. 2025. Seagrass Restoration (Accessed 23rd September 2025)
Sell, P.D. & Murrell, G. 1996. Flora of Great Britain and Ireland: Volume 5, Butomaceae - Orchidaceae. Cambridge: Cambridge University Press.
SEPA. 2024. Water Classification Hub (Accessed 12th December 2024).
Setchel, W.A. 1929. Morphological and phenological notes on Zostera marina. University of California Publications on Botany, 14, pp. 185-201.
Sgambelluri, L.R., Jarvis, J.C. & Kamel, S.J. 2024. Multiple paternity, fertilization success, and male quality: Mating system variation in the eelgrass, Zostera marina. Ecology and Evolution,14, e11608.
Shetland Islands Council. 2004. Shetland Local Plan 2004, Tingwall, Whiteness and Weisdale Community Council Area Statement.
Short, F.T., Muehlstein, L.K. & Porter, D. 1987. Eelgrass wasting disease cause and recurrence of a marine epidemic. Biological Bulletin of the Marine Biology Laboratory, Woods Hole, 173, pp. 557-562.
Short, F.T. & Burdick, D.M. 1995. Mesocosm experiments quantify the effects of eutrophication on eelgrass, Zostera marina. Limnology and Oceanography, 40(4), pp. 740-749.
Short, F.T., Burdick, D.M., Granger, S. and Nixon, S.W. 1996. Long-term decline in eelgrass, Zostera marina L., linked to increased housing development. In: J. Kuo, R.C. Phillips, D.I. Walker & H. Kirkman (eds.) Seagrass biology: proceedings of an international workshop, Rottnest Island, Western Australia, 25-29 January 1996. Nedlands, Western Australia: Sciences UWA.
Smith, W.A. 1875. Lewsiana or Life in the Outer Hebrides. London: Daldy, Isbister, & Co.
Smith, W.A. 1882. Benderloch: Notes from the West Highlands. Paisley: Alexander Gardner.
Smith, W.A. 1887. Loch Creran: Notes from the West Highlands. Paisley: Alexander Gardner.
Smith, I. 1975. St. Peter’s Pool, Deerness. Bulletin of the Orkney Field Club, pp. 1-2.
Smith, R.D., Pregnall, A.M. & Alberte, R.S. 1988. Effects of anaerobiosis on root metabolism of Zostera marina eelgrass: implications for survival in reducing sediments. Marine Biology, 98, pp. 131–141.
Smith, P.H., Dixon, R.O.D. & Cochrane, M.P. 2002. Plant Life of Edinburgh and the Lothians. Edinburgh: Edinburgh University Press.
Smout T.C. & Stewart, M. 2012. The Firth of Forth, An Environmental History. Edinburgh: Birlinn
Sonntag, C.O. 1894. A Pocket Flora of Edinburgh and the Surrounding District: a collection and full description of all phanerogamic and the principal cryptogamic plants classified after the natural system, with an artificial key and a glossary of botanical terms. Edinburgh: Williams and Norgate.
Southall, T.D. & Tully, O. 2014. Solway Cockle Fishery Study. A review of management options for the Solway Firth Cockle Fishery. Solway Firth Partnership Commissioned Report.
Spence, M. 1914. Flora Orcadensis. Kirkwall: Spence.
Sphere Environmental Consultants.1981. The Beatrice Project Environmental Monitoring Programme. Zostera spp. and other plant communities Nigg Bay 1981.
St. John, C. 1863. Natural History and Sport in Moray. Edinburgh: Edmonston and Douglas.
Stace, C.A. 1989. Plant Taxonomy and Biosystematics. 2nd edition. Cambridge: Cambridge University Press.
Stace, C.A. 1991. New Flora of the British Isles. 1st edition. Cambridge: Cambridge University Press.
Stace, C.A. 2010. New Flora of the British Isles. 3rd edition. Cambridge: Cambridge University Press.
Stace, C.A. 2019. New Flora of the British Isles. 4th edition. C & M Floristics.
Stevens, P.A., Adamson, J.K., Anderson, M.A. & Hornung, M. 1988. Effects of clearfelling on surface water quality and site nutrient status. In: M.B. Usher & D.A.B. Thompson (eds.) Ecological Change in the Uplands. Oxford: Blackwell Scientific Publications.
Stigter, T. 2011. Restoration of groundwater quality to sustain coastal ecosystems productivity. Treatise on Estuarine and Coastal Science, 10, pp. 245–262.
Stoneman, J. 2000. Udale Bay Reserve Zostera Survey, 2000. RSPB internal report.
Sullivan, B.K., Trevathan-Tackett, S.M., Neuhauser, S. & Govers, L.L. 2018. Review: Host-pathogen dynamics of seagrass diseases under future global change. Marine Pollution Bulletin, 134, pp. 75-88.
Sumoski, S.E. & Orth, R.J. 2012. Biotic dispersal in eelgrass Zostera marina. Marine Ecology Progress Series, 471, pp. 1–10.
Sutherland, T.F., Elner, R.W. & O’Neill, J.D. 2013. Roberts Bank: Ecological crucible of the Fraser River estuary. Progress in Oceanography, 115, pp. 171-180.
Svedäng, H. & Bardon, G. 2003. Spatial and temporal aspects of the decline in cod (Gadus morhua L.) abundance in the Kattegat and eastern Skagerrak. ICES Journal of Marine Science, 60(1), pp. 32-37.
Tett, P., Benjamins, S., Black, K., Coulson, M., Davidson, K., Fernandes, T.F., Fox, C., Hart, M., Hicks, N., Hughes, A., Hunter, D.C., Nickell, T., Risch, D., Tocher, D., Vare, L., Verspoor, E., Wilding, T., Wilson, B. & Wittich, A. 2018. Review of the environmental impacts of salmon farming in Scotland. [Report for the Environment, Climate Change and Land Reform (ECCLR) Committee]. Oban: SAMS Research Services.
Thayer, G.W. & Stuart, H.H. 1974. The bay scallop makes its bed of seagrass. Marine Fisheries Review, 36, pp. 27-30.
The Farmer’s Magazine. 1822. Zostera or Sea-grass. February, pp. 27-28.
The Rivers Trust. 2024. The State of Our Rivers Report (Accessed 17th April 2024).
The Shetland News. 1901. The Season at Baltasound. Saturday 21st September, p. 5.
Thomas, O. 2016. Ecology of contrasting infralittoral seagrass beds in the South Arran MPA: temporal changes in extent and comparisons of associated biota. MSc Thesis, Heriot-Watt University.
Thomson, M., Jackson, E. & Kakkonen, J. 2014. Seagrass Zostera beds in Orkney. Scottish Natural Heritage Commissioned Report No. 765.
Thorpe, K. 1998. Marine Nature Conservation Review Sectors 1 & 2. Lagoons in Shetland and Orkney: area summaries. Peterborough: JNCC.
Thorpe, K., Dalkin, M.J., Fortune, F. & Nichols, D.M. 1998. Marine Nature Conservation Review Sector 14. Lagoons in the Outer Hebrides: area summaries. Peterborough: JNCC.
Thurstan, R.H. & Roberts, C.M. 2010. Ecological meltdown in the Firth of Clyde, Scotland: Two centuries of change in a coastal marine ecosystem. PLoS ONE, 5(7), e11767.
Thurstan, R.H., McCormick, H., Preston, J., Ashton, E.C., Bennema, F.P., Bratoš Cetinić, A., Brown, J.H., Cameron, T.C., da Costa, F., Donnan, D.W., Ewers, C., Fortibuoni, T., Galimany, E., Giovanardi, O., Grancher, R., Grech, D., Hayden-Hughes, M., Helmer, L., Jensen, K.T., Juanes, J.A., Latchford, J., Moore, A.B.M., Moutopoulos, D.K., Nielsen, P., von Nordheim, H., Ondiviela, B., Peter, C., Pogoda, B., Poulsen, B., Pouvreau, S., Roberts, C.M., Scherer, C., Smaal, A.C., Smyth, D., Strand, A., Theodorou, J.A. & zu Ermgassen, P.S.E. 2024. Records reveal the vast historical extent of European oyster reef ecosystems. Nature Sustainability, 7, pp. 1719–1729.
Trendall, J., Bedford, G., Tarrant, D., Fortune, F. & Saunders, G. 2011. Baseline Survey and mapping of intertidal features within selected Scottish East Coast SSSI and Ramsar Sites. Scottish Natural Heritage Commissioned Report No. 447.
Tubbs, C.R. & Tubbs, J.M. 1982. Brent geese Branta bernicla and their food in the Solent, southern England. Biological Conservation, 23, pp. 33-54.
Tubbs, C.R. & Tubbs, J.M. 1983. The distribution of Zostera and its exploitation by wildfowl in the Solent, southern England. Aquatic Botany, 15, pp. 223-239.
Tubbs, C.R. 1996. Estuary Birds – Before the counting began. British Wildlife, 7(4), pp. 226-235.
Tuck, I.D., Bailey, N., Harding, M., Sangster, G., Howell, T., Graham, N. & Breen, M. 2000. The impact of water jet dredging for razor clams, Ensis spp., in a shallow sandy subtidal environment. Journal of Sea Research, 43(1), pp. 65-81.
Tutin, T.G. 1938. The autecology of Zostera marina in relation to its wasting disease. New Phytologist, 37(1), pp. 1-84.
Tutin, T.G. 1942. Biological Flora of the British Isles. No. 7. Zostera genus L. Journal of Ecology, 30, pp. 217–226.
Tweedley, J., Jackson, E. & Attrill, M. 2008. Zostera marina seagrass beds enhance the attachment of the invasive alga Sargassum muticum in soft sediments. Marine Ecology Progress Series, 354, pp. 305-309.
Tyler-Walters, H., James, B., Carruthers, M. (eds.) Wilding, C., Durkin, O., Lacey, C., Philpott, E., Adams, L., Chaniotis, P.D., Wilkes, P.T.V., Seeley, R., Neilly, M., Dargie, J. & Crawford-Avis, O.T. 2016. Descriptions of Scottish Priority Marine Features PMFs. Scottish Natural Heritage Commissioned Report No. 406.
UK Government. 1995. Biodiversity: The UK Steering Group Report Volume 2: Action Plans. London: HMSO.
UMBSM. 2007. Conservation of the native oyster in Scotland. Scottish Natural Heritage Report 251.
University of Aberdeen. 1978. Moray Firth development ecological survey. Continuing studies of intertidal plant communities in the Moray Firth sub-region. Unpublished Interim Report, No. 9 to Highlands and Islands Development Board and Cromarty Petroleum.
Unsworth, R.K.F., Jones, B.L.H, Rogers, A., Cullen-Unsworth, L.C. & Lilley, R.J. 2025. SeagrassSpotter. Project Seagrass, Bridgend, UK (Accessed 23rd February 2025).
Urbina, M.A., Cumillaf, J.P., Paschke, K. & Gebauer, P. 2019. Effects of pharmaceuticals used to treat salmon lice on non-target species: Evidence from a systematic review. Science of the Total Environment, 649, pp. 1124-1136.
Urquhart, J.T. 1824. On the preparation of the Zostera or sea-grass in Orkney.Prize-essays and Transactions of the Highland Society of Scotland, 6, pp. 588-593.
van den Hoek, C., Admiraal, W. & de Jonge, V.N. 1983. The role of algae and seagrasses in the ecosystem of the Wadden Sea: A Review. In: W.J. Wolff (ed.) Ecology of the Wadden Sea. Rotterdam: A.A. Balkema.
van Tussenbroek, B.I., Monroy-Velazquez, L.V. & Solis-Weis, V. 2012. Meso-fauna foraging on seagrass pollen may serve in marine zoophilous pollination. Marine Ecology Progress Series, 469, pp. 1–6.
van Tussenbroek, B., Villamil, N., Márquez-Guzmán, J., Wong, R., Monroy-Velázquez, L.V. & Solis-Weiss, V. 2016. Experimental evidence of pollination in marine flowers by invertebrate fauna. Nature Communications, 7, 12980.
Vilks, G., Schafer, C.T. & Walker, D.A. 1975. The influence of a causeway on oceanography and foraminifera in the Strait of Canso, Nova Scotia. Canadian Journal of Earth Sciences, 1212, pp. 2086-2102.
Waldichuk, M. 1985. Roberts Bank coal port, British Columbia: How did the environmental impact assessment fare? Marine Pollution Bulletin, 16(5), pp. 179–185.
Walker, D.I., Kendrick, G.A. & McComb, A.J. 2006. Decline and Recovery of Seagrass Ecosystems: The Dynamics of Change. In: A.W.D. Larkum, R.J. Orth and C.M. Duarte (eds.) Seagrasses: Biology, Ecology and Conservation. Dordrecht: Springer.
Want, A., Matejusova, I., & Kakkonen, J. E. 2023. The establishment of the invasive non-native macroalga Sargassum muticum in the north of Scotland. Journal of the Marine Biological Association of the UK, 103, e69.
Waring, K. 2024. Islay’s Incredible Seagrass (Accessed on 25th September 2025).
Watson, H.C. 1837. The New Botanist's Guide to the Localities of the Rarer Plants of Britain, Vol. 2: Scotland and Adjacent Isles. London: Longman, Orme, Brown, Green, and Longmans.
Watson, K. 2013. The Flora of Renfrewshire. Glasgow: Glasgow Museums.
Waycott, M., Duarte, C.M., Carruthers, T.B.J., Orth, R.J., Dennison, W.C., Olyarnik, S., Calladine, A., Fourqurean, J.W., Heck, Jr K.L., Hughes, A.R., Kendrick, G.A., Kenworthy, W.J., Short, F.T. & Williams, S.L. 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. PNAS, 106(30), pp. 12377-12381.
WFD-UKTAG. 2014. UK Technical Advisory Group Guide to the Intertidal Seagrass Tool, Water Framework Directive: Transitional and Coastal Waters.
Whalen, M.A., Duffy, J.E. & Grace, J.B. 2013. Temporal shifts in top-down vs. bottom-up control of epiphytic algae in a seagrass ecosystem. Ecology, 94(2), pp. 510-520.
Whelan, P.M. & Cullinane, J.P. 1985. The algal flora of a subtidal Zostera bed in Ventry Bay, south-west Ireland. Aquatic Botany, 23, pp. 41-51.
Whelan, M.J., Linstead, C., Worrall, F., Ormerod, S.J., Durance, I., Johnson, A.C., Johnson D., Owen, M., Wiik, E., Howden, N.J.K., Burt, T.P., Boxall, A., Brown, C.D., Oliver D.M. & Tickner, D. 2022. Is water quality in British rivers “better than at any time since the end of the Industrial Revolution”? Science of The Total Environment, 843, 157014.
White, G. 1987. Report on survey of Poll Loisgann 15.069162 and Cùil Lochain 15.073169 Inverpolly National Nature Reserve, Wester Ross. Thursday 20 August 1987. The University of Technology, Loughborough. Unpublished report to Nature Conservancy Council. 6pp.
Wilkie, L. 2011. The Role of Intertidal Seagrass Zostera spp. in Sediment Deposition and Coastal Stability in the Tay Estuary, Scotland. PhD Thesis, University of St Andrews.
Williams, S.L. & Heck Jr., K.L. 2001. Seagrass community ecology. In: M.D. Bertness, S.D. Gaines & M.E. Hay (eds.) Marine community ecology. Sunderland, MA: Sinauer Associates.
Wilson, J.H. 1910. Nature study rambles, round St Andrews. St Andrews: W.C. Henderson & Son, University Press.
Wilson, D.P. 1949. The decline of Zostera marina L. at Salcombe and its effects on the shore. Journal of the Marine Biological Association U.K, 28, pp. 395-412.
Wood, H. 2006. Seasearch Survey Form: Skipness Seagrass Bed. Dive Date 09.06.2006
WoRMS. 2025. World Register of Marine Species at VLIZ. [Accessed 20 September 2025]
Xu, S., Zhou, Y., Wang, P., Wang, F., Zhang, X. & Gu, R. 2016. Salinity and temperature significantly influence seed germination, seedling establishment, and seedling growth of eelgrass Zostera marina L. PeerJ., 4, e2697.
Yates, C.C. 1913. Summary of survey of oyster bars in Maryland, 1906-1912. Washington, DC: United States Department of Commerce.
Yonge, C.M. 1949. The Seashore. London: Collins.
Yonge, C.M. 1960. Oysters. London: Collins.
Yu, L., Renton, J., Burian, A., Khachaturyan, M., Bayer, T., Kotta, J., Stachowicz, J.J., DuBois, K., Baums, I.B., Werner, B. & Reusch, T.B.H. 2024. A somatic genetic clock for clonal species. Nature Ecology & Evolution, 8, pp. 1327–1336.
Zhang, M., Dai, Z., Bouma, T. J., Bricker, J., Townend, I., Wen, J., Zhao, T. & Cai, H. 2021. Tidal-flat reclamation aggravates potential risk from storm impacts. Coastal Engineering, 166, 103868.
Zimmerman, R.C. & Alberte, R.S. 1996. Effect of light/dark transition on carbon translocation in eelgrass Zostera marina seedlings. Marine Ecology Progress Series, 136(1/3), pp. 305–309.
Zipperle, A.M., Coyer, J.A., Reise, K., Stam, W.T. & Olsen, J.L. 2009. Evidence for persistent seed banks in dwarf eelgrass Zostera noltii in the German Wadden Sea. Marine Ecology Progress Series, 380, pp. 73-80.
Zoutenbier, R. 2011. Mapping the Distribution of Seagrass in the Firth of Forth, Assessing the Sediment Composition on a Micro Habitat Scale and Analysing the Impact of Climate Change on Seagrass Distribution. MSc Thesis, Heriot-Watt University.
Zoutenbier, R., Barret, C. & Lyndon, A.R. 2016. Distribution and Status of seagrass Zostera spp. in the Estuary and Firth of Forth. Poster presentation at the 12th International Seagrass Biology Workshop. Nant Gwyrthern, North Wales.
zu Ermgassen, P.S.E., McCormick, H., Debney, A., Fariñas-Franco, J.M., Gamble, C., Gillies, C., Hancock, B., Laugen, A.T., Pouvreau, S., Preston, J., Sanderson, W.G., Strand, Å. and Thurstan, R.H. 2025. European native oyster reef ecosystems are universally collapsed. Conservation Letters, 18: e13068.
Annex 1 - Distribution maps
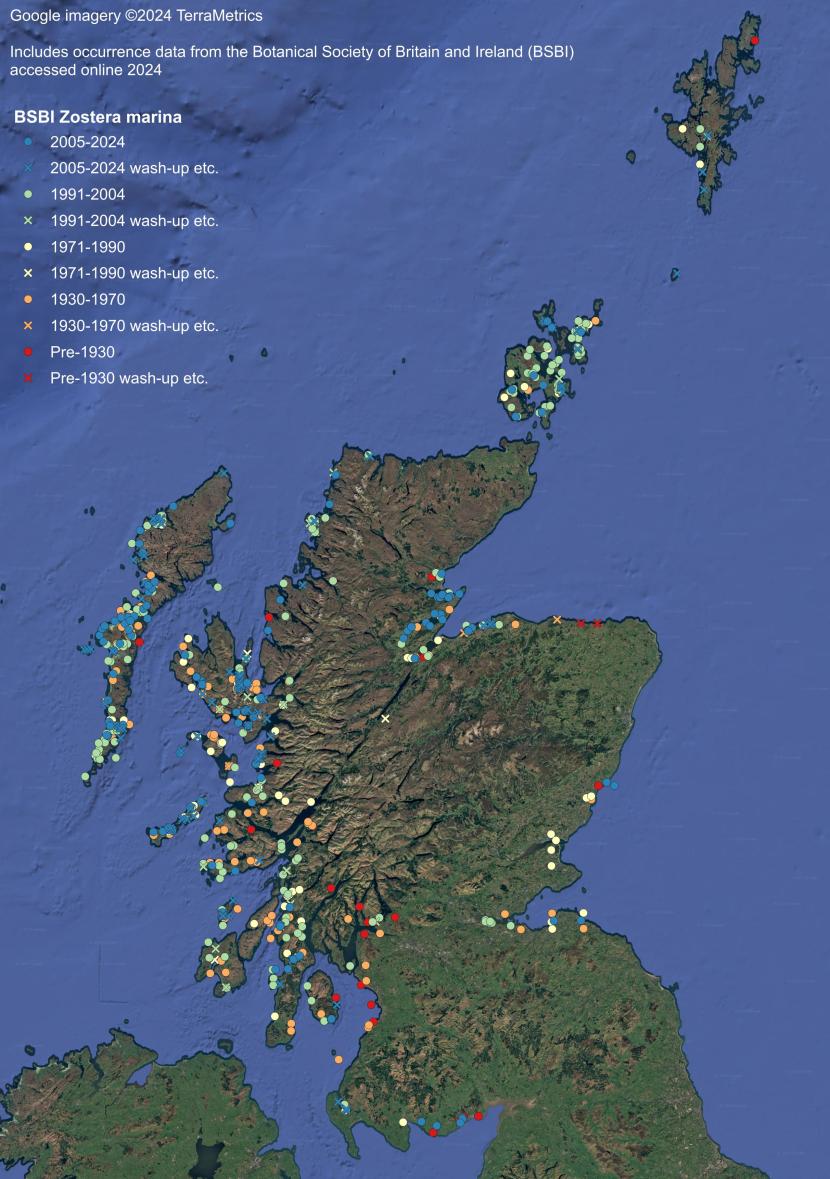
Figure 41. Zostera marina records in Scottish waters from the BSBI database. Data is typically recorded in squares ranging from 1-10 km2 though are presented here as centroids i.e. the central point of each recording square.
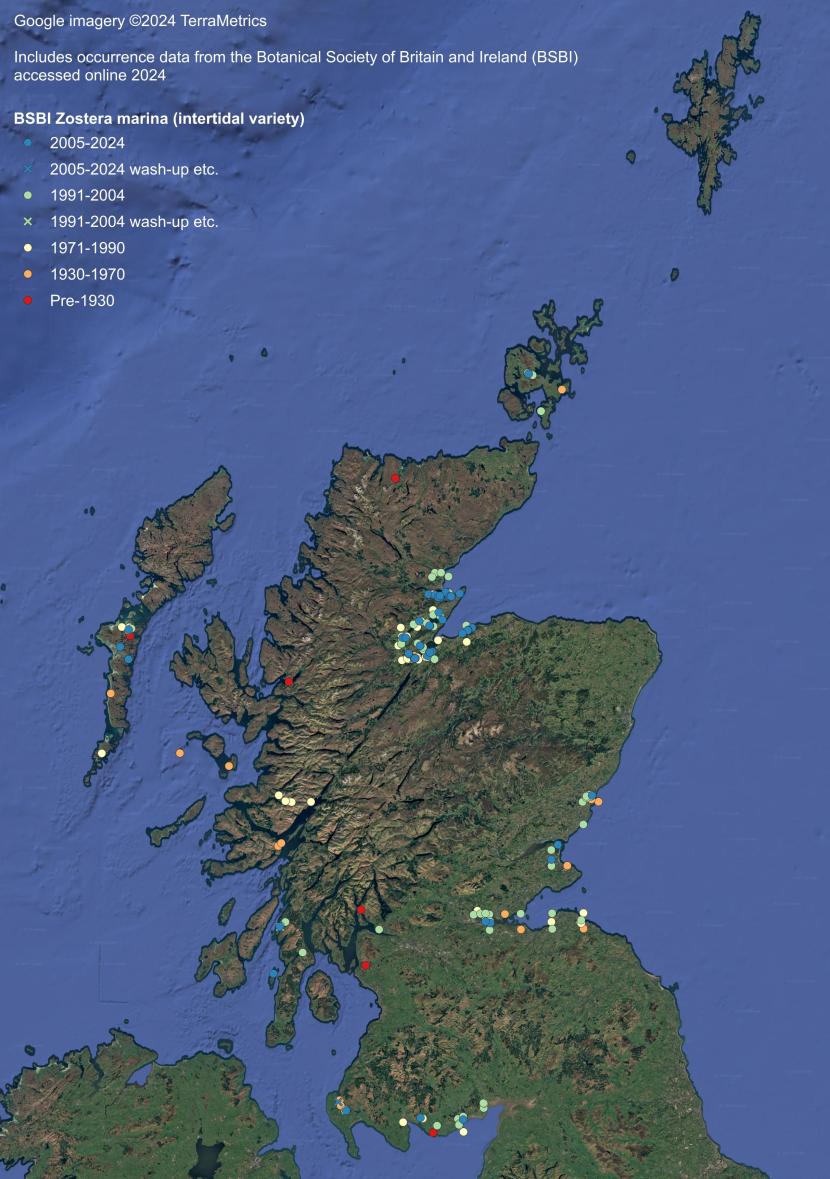
Figure 42. Zostera marina (intertidal variety) records in Scottish waters from the BSBI database. Data is typically recorded in squares ranging from 1-10 km2 though are presented here as centroids i.e. the central point of each recording square.
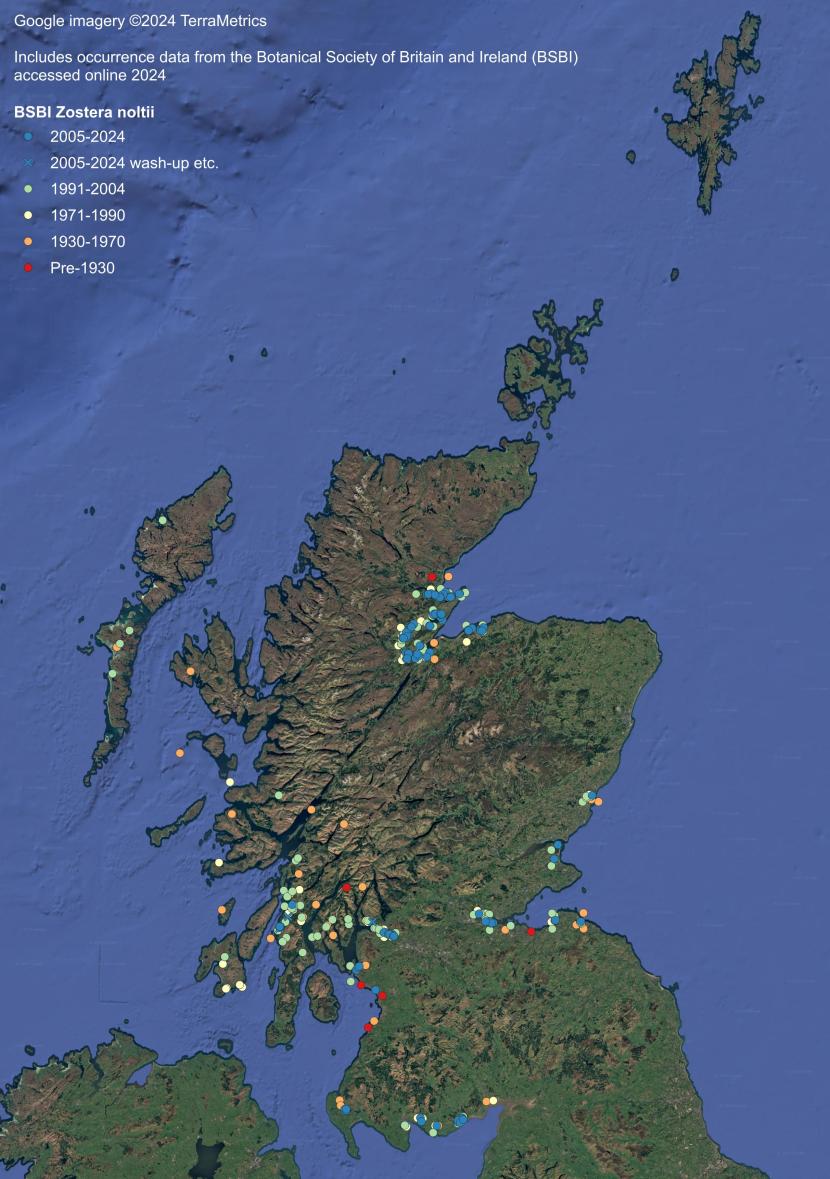
Figure 43. Zostera noltii records in Scottish waters from the BSBI database. Data is typically recorded in squares ranging from 1-10 km2 though are presented here as centroids i.e. the central point of each recording square.
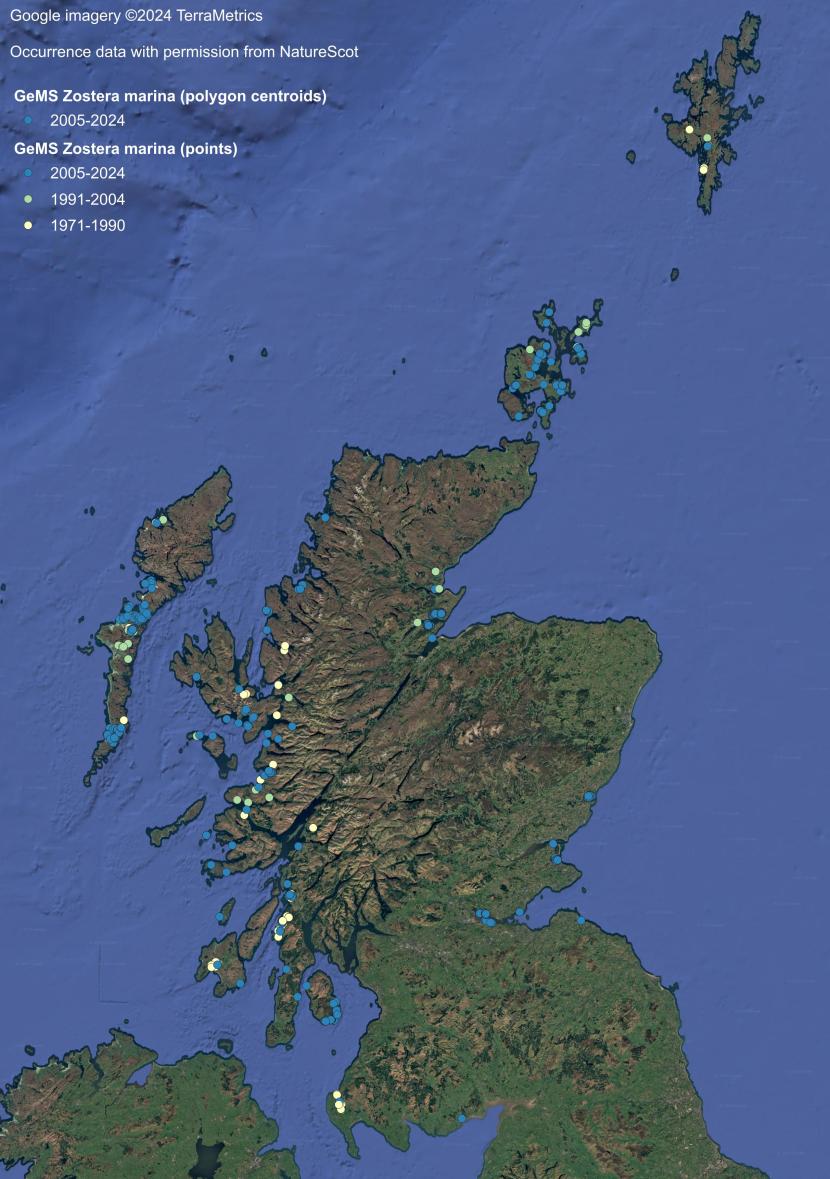
Figure 44. Zostera marina records in Scottish waters from the GeMS database.
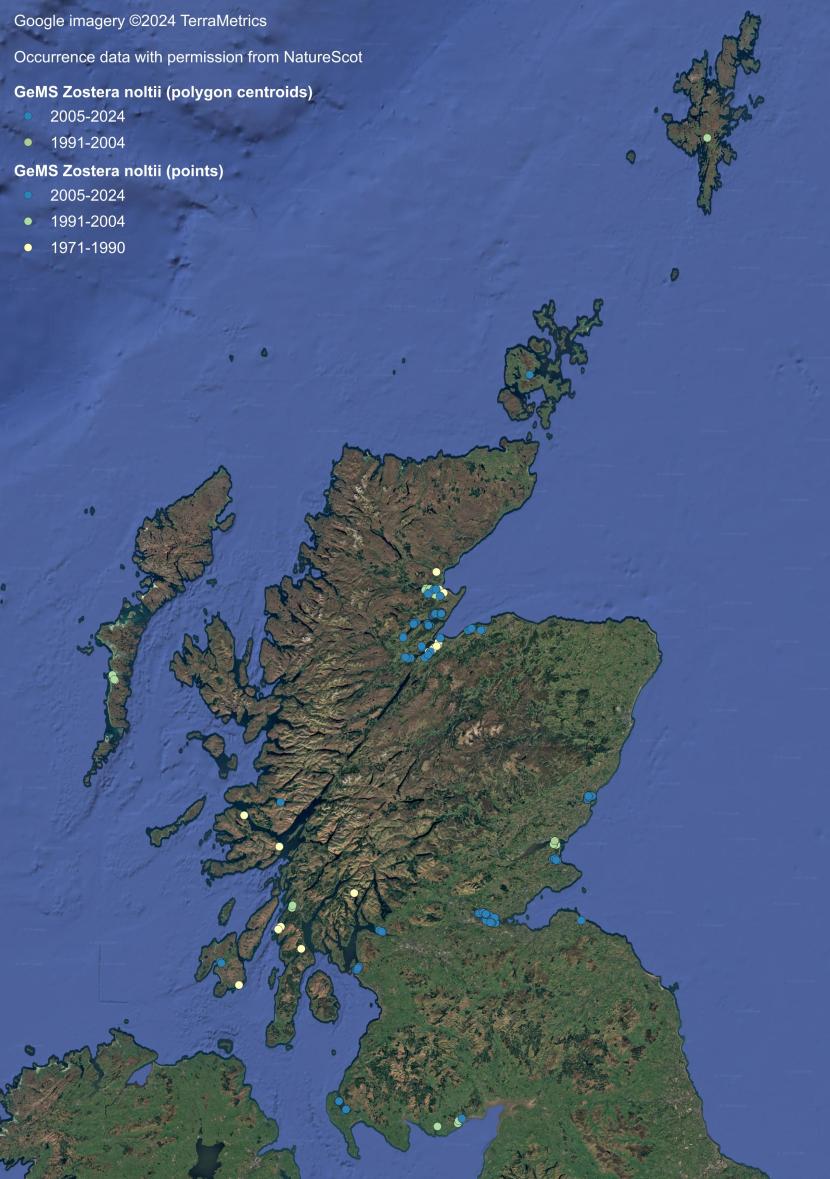
Figure 45. Zostera noltii records in Scottish waters from the GeMS database.
Annex 2: Historical vs contemporary Zostera beds
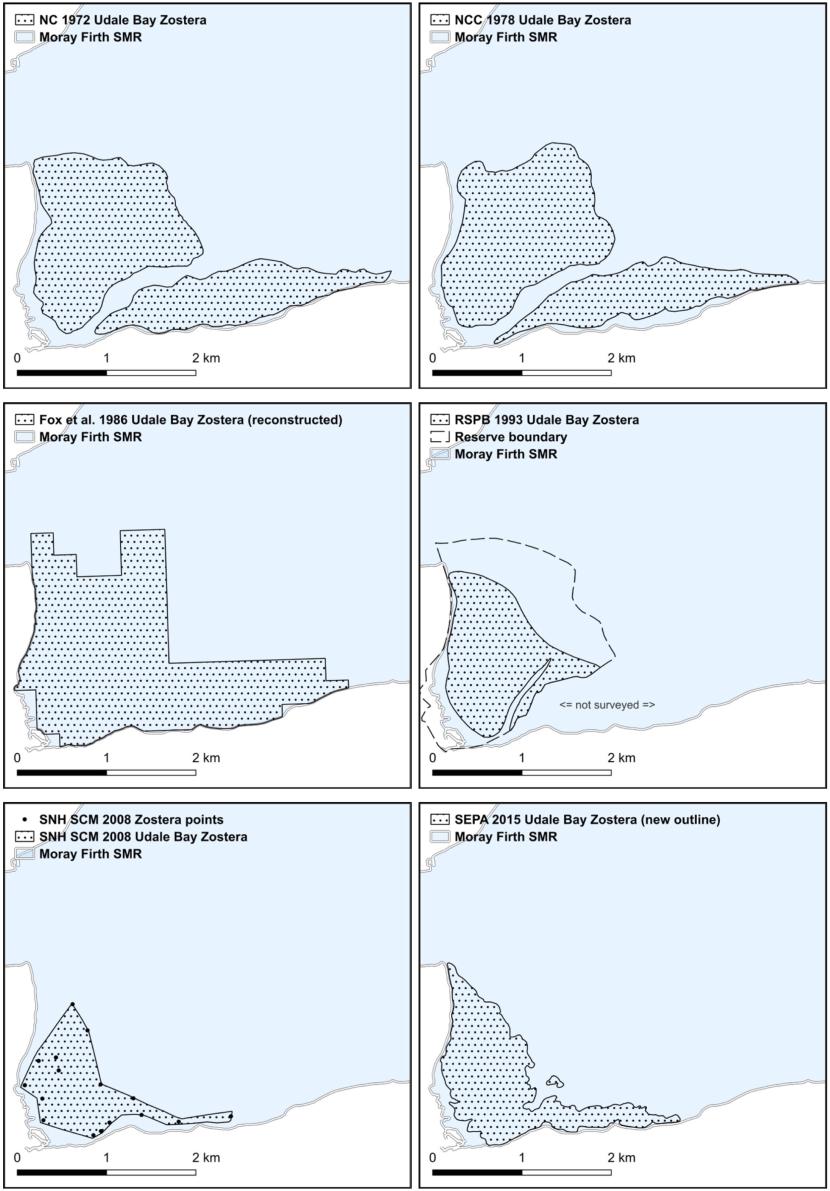
Figure 47. Historical vs. contemporary Zostera beds at Udale Bay, Cromarty Firth.
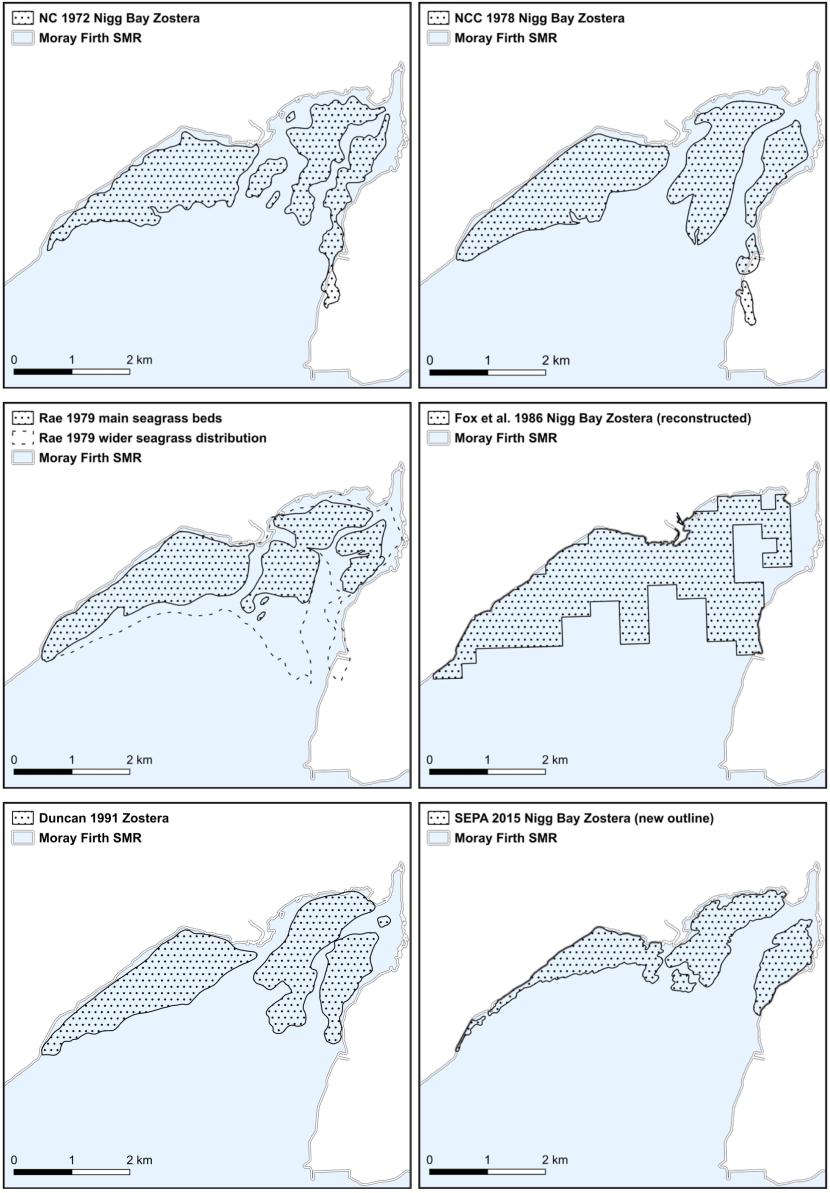
Figure 48: Historical vs. contemporary Zostera beds at Nigg Bay, Cromarty Firth.
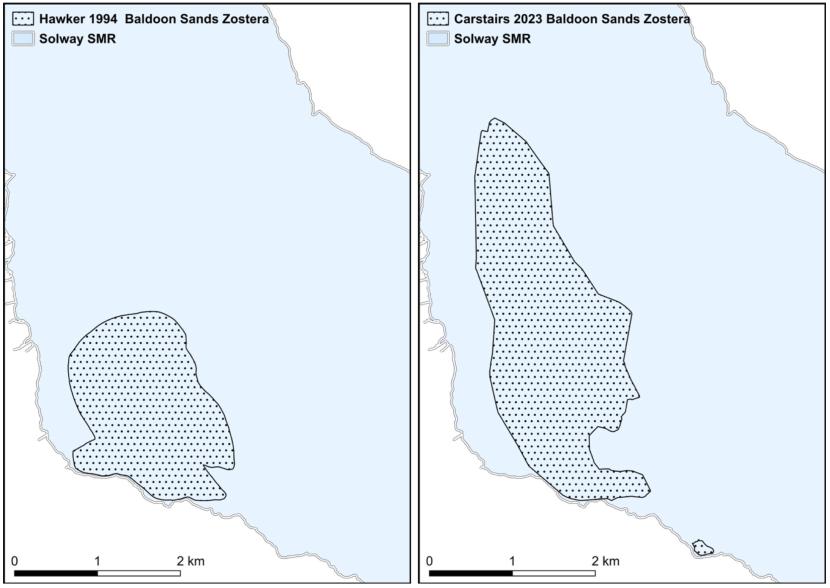
Figure 49. Historical vs. contemporary Zostera beds at Baldoon Sands, Solway Firth.
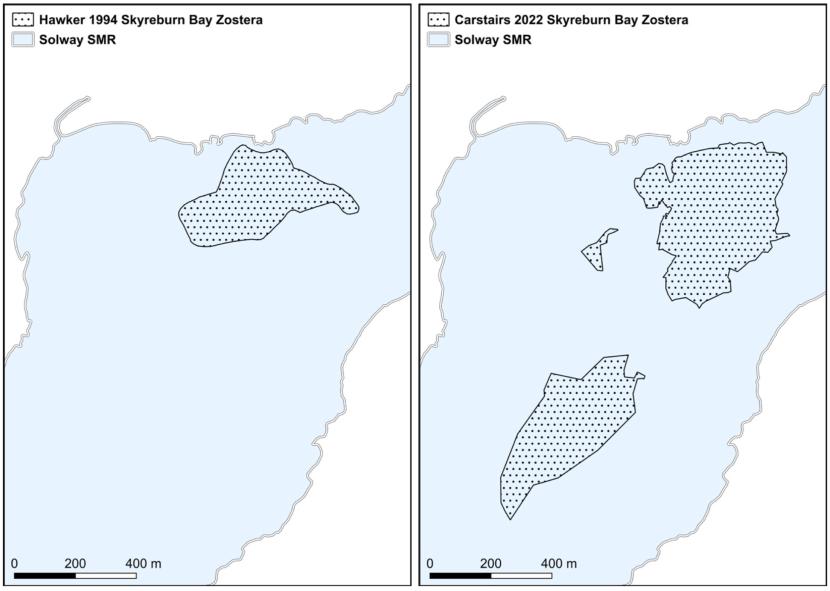
Figure 50. Historical vs. contemporary Zostera beds at Skyreburn Bay, Solway Firth.
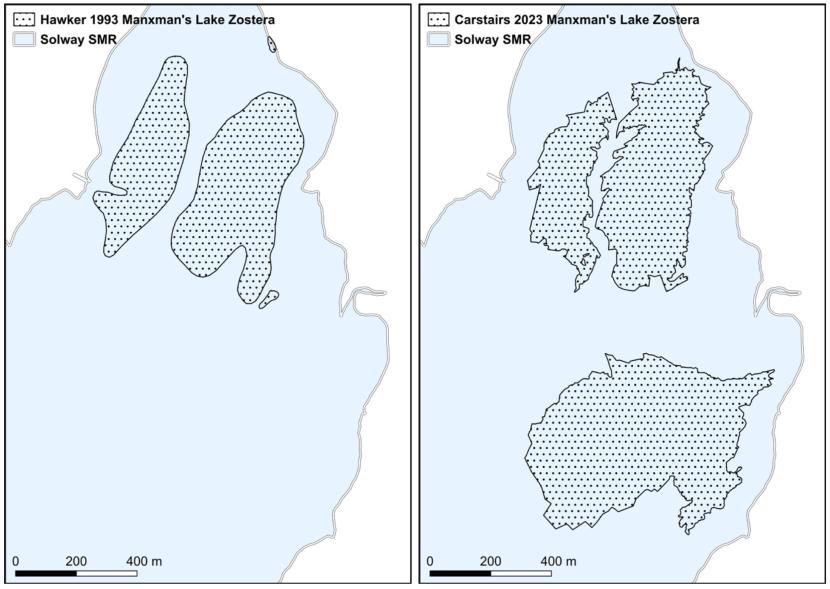
Figure 51. Historical vs. contemporary Zostera beds at Manxman’s Lake, Solway Firth.
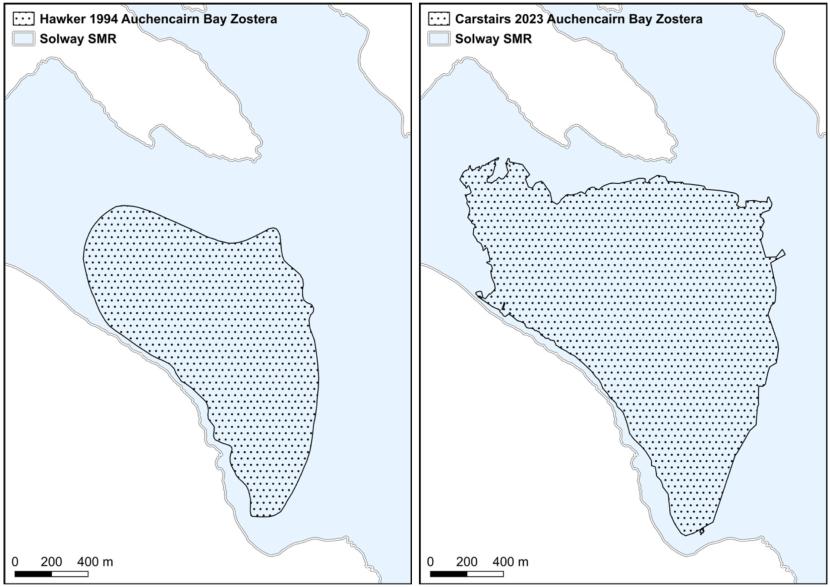
Figure 52. Historical vs. contemporary Zostera beds at Auchencairn Bay, Solway Firth.
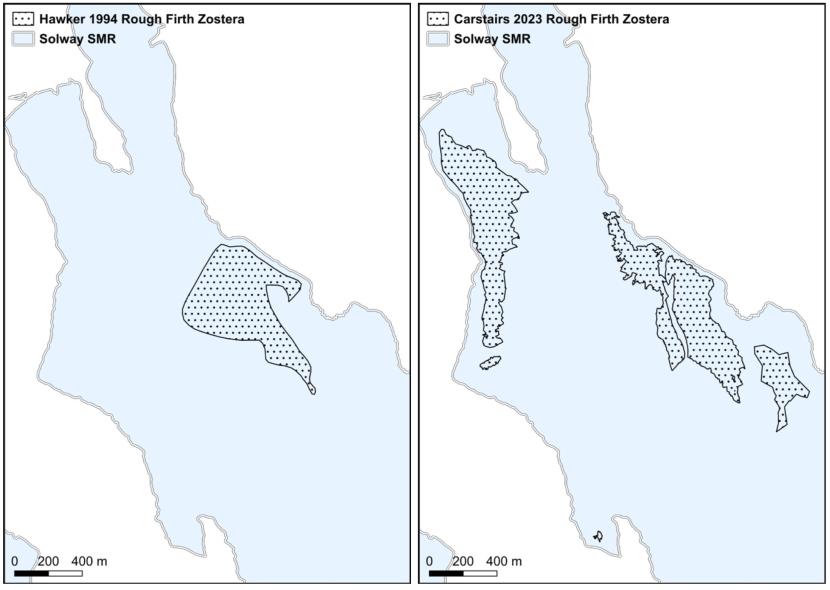
Figure 53. Historical vs. contemporary Zostera beds at Rough Firth, Solway Firth.






