NatureScot Research Report 1349 - Recruitment and reef development of Serpula vermicularis during a population decline of the species in Loch Creran, Argyll
Year of publication: 2026
Authors: Moore, C.G., Harries, D.B., Lyndon, A.R. and Tulbure, K.W. (Heriot-Watt University)
Cite as: Moore, C.G., Harries, D.B., Lyndon, A.R. and Tulbure, K.W. (Heriot-Watt University) Recruitment and reef development of Serpula vermicularis during a population decline of the species in Loch Creran, Argyll.
NatureScot Research Report 1349.
Keywords
Serpula vermicularis; organ-pipe worm; Loch Creran; serpulid reefs; Special Area of Conservation; decline; remediation; recruitment, fouling.
Background
Recognised as representing the most important site globally for the presence of aggregations or reefs of the polychaete worm Serpula vermicularis, condition monitoring of Loch Creran in 2015 revealed widespread degradation of the feature in the form of reef fragmentation and reduction in tube occupancy. The aim of this report is to describe the findings from recent studies in the loch on serpulid worm recruitment and reef development to further the understanding of the processes underlying the decline of the habitat.
Main findings
- The trial introduction of serpulid reef remediation substrates into Loch Creran led to their colonisation by serpulid reefs in areas where there was no indication of recovery of existing degraded reef material.
- Reef development on the remediation units in terms of percentage cover and reef height varied according to substrate type and location.
- While reefs developed at all locations, increase in coverage of the remediation units was only temporary and ceased or was replaced by a decline at the different study locations.
- Following an initial increase in height of reefs on the remediation units during the first three years, the height subsequently declined or exhibited no significant change at most of the locations studied. However, at one site reefs continued to increase in height throughout the 10 year study at a mean rate of c.2.9 cm a-1 on one substrate type. The similarity of this measure to pre-decline estimates of reef growth (3 cm a-1) and tube growth (3.3 cm a-1) is suggestive of established serpulids that are not experiencing physiological impairment.
- The results of a recruitment study involving the establishment of settlement substrates in Loch Creran and Loch Feochan were indicative of a marked decline in recruitment of juvenile Serpula vermicularis in Loch Creran from 2001 to 2023.
- No spatial patterns in recruitment density were identified within Loch Creran or between this loch and Loch Feochan.
- Taking into consideration the findings of the remediation and recruitment studies, a process of reef decline through fouling is proposed as a possible driver for the reduction in condition of the serpulid reef habitat in Loch Creran.
Introduction
The Loch Creran SAC was established in 2005 to afford protection for the marine feature reefs, which includes aggregations or reefs of the organ-pipe worm, Serpula vermicularis, which are rare on both national and global scales. Site condition monitoring of the feature in 2015 recorded widespread degradation in the form of reef fragmentation and reduction in tube occupancy. Extensive areas that in 2005 had consisted of dense reefs up to 60 cm in height, were observed in 2015 to have been reduced to fragmented reef rubble scattered on the sediment, with no indication of the cause (Moore et al., 2020). Coincidentally a PhD study of reef restoration techniques began in 2012 included an examination of the potential of a range of colonisation substrates in promoting reef development and the effect of location on development (Cook, 2016). The timescale of this work (2012 - 2014) was too short to monitor significant reef growth, but it provided the foundation for long-term monitoring of reef development, which is reported here. It was hoped that it would provide some indication of the continued viability of the loch to sustain reef development.
Even if the growth of reefs of Serpula vermicularis continues unabated in the loch, long-term survival of the habitat will be dependent upon larval settlement and subsequent recruitment at appropriate levels. A second aim of this report therefore is to examine evidence for a temporal decline in recruitment of S. vermicularis in Loch Creran. This involved the deployment of settlement substrates in 2023 to facilitate comparisons with historical measures of early recruitment in the loch, particularly those of Chapman (2004) and Chapman et al. (2007), which were carried out prior to the decline in reef condition. Intermediate assessment of recruitment success is provided by Cook (2016). Additional deployments in 2023 at two locations in Loch Feochan were employed to investigate spatial variation in recruitment.
One biotic factor that may exert a significant influence on reef development is predation. In Ardbear Lough, Ireland, Bosence (1979) observed several predatory species feeding on serpulid tubes or their contents including the sea urchins Psammechinus miliaris and Echinus esculentus, their stomachs being full of tube debris, and wrasse species Ctenolabrus rupestris and Crenilabrus melops, which were observed biting open tubes and extracting the worms. Very high densities of P. miliaris have been recorded on reef material in Loch Creran (e.g. 577 m-2 in Moore et al., 2020). A third aim of the current report is to provide preliminary observations from a PhD study, initiated in 2019, of the effect of predation on reef regeneration through the employment of predator exclusion cages.
Aims
- to assess temporal change in the extent of serpulid reefs established on experimental restoration units
- to examine evidence for a temporal decline in recruitment of S. vermicularis in the Loch Creran
- to make preliminary investigations on the effect of predation on reef regeneration through the employment of predator exclusion cages
2. Methods
2.1 Reef development
Reef restoration units were installed in Loch Creran in March 2012. At one site in the lower basin, (termed ‘Main Site’ and located in Port Columba Bay (Figure 1)), the effect of substrate type on reef development was assessed by deploying 6 replicates of 5 different substrate types (Figure 2) at a depth of 6 m:
- scallop shells in the form of a loose pile (LS)
- scallop shells in a small cylindrical mesh bag (radius 8 cm, length 25 cm, 1.6 cm mesh) (SSB)
- scallop shells in a large, cylindrical mesh bag (radius 16 cm, length, 30 cm, mesh 1.6 cm) (LSB)
- 6.4 - 25.6 cm diameter cobbles in a mesh bag with the same dimensions as the large scallop bag (CB)
- one 50 - 80 cm diameter boulder (B)
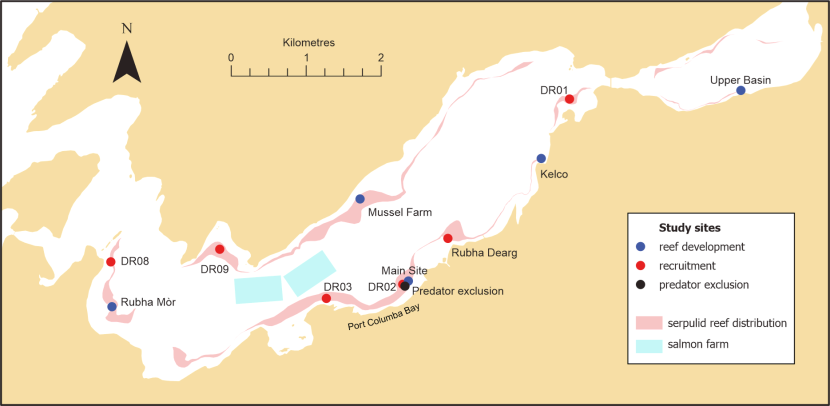
Figure 1. Locations utilised for the studies described in this report. Also shown is the pre-decline distribution of serpulid reefs as surveyed in 2005 and the licensed areas for salmon farming.
Click for a full description
The location of study sites is shown as blue dots for the reef development study, red dots for the recruitment study and a black dot for the predator exclusion experiment. The pre-decline distribution of serpulid reefs as determined in 2005 (Moore et al., 2006) is shown in pink and the location of the salmon farms in pale blue.
The units were deployed in six blocks, 10 m apart. Each block contained one replicate of each treatment. This permitted allocation of monitoring data to specific replicates.
The importance of deployment location was assessed by deployment of 6 replicates of one of the substrate types (scallops in large mesh bag) at an additional 4 locations (3 in the lower basin and one in the upper basin) at a depth of 6 m (Figure 1). At each location replicates had a minimum separation of 2 m and were tagged to permit the attribution of monitoring data to specific replicates.
Monitoring of the units every five months revealed the presence of serpulid settlement, but no well-defined reef structures had developed on the units by the end of the PhD study in 2014. However, for the purposes of the current study, subsequent monitoring of the units was carried out in December 2015, June 2017, October 2018 and August 2022.
All units were re-examined in situ by diving. Maximum serpulid reef height (defined as vertical distance from the substrate to the upper extremity of upwardly rising tubes) was measured by ruler to the nearest centimetre, and a visual estimate was made of percentage cover of the unit by upwardly rising tubes and the percentage cover by erect algae. Detailed GoPro video footage was taken of all units and still imagery of many of the units from the Main Site.

Figure 2. Substrate types deployed in the reef development study. A, cobbles in large bag; B, scallop shells in large bag; C, scallop shells in small bag; D, loose scallop shells; E, boulder.
2.2 Recruitment
Recruitment was investigated by deployment of settlement substrates at a depth of 6 m +/-1.4 m at six locations on 24-26 July 2023 in Loch Creran (Figure 1 and Annex 1) and two locations in Loch Feochan (Annex 1). Temporal and depth data are provided in Table 1.
| Location | Start date | End date | Duration (days) | Depth (m) |
|---|---|---|---|---|
| Rubha Dearg | 25/07/2023 | 26/11/2023 | 124 | 5.8 |
| DR01 | 24/07/2023 | 25/11/2023 | 124 | 5.8 |
| DR02 | 24/07/2023 | 25/11/2023 | 124 | 7.4 |
| DR03 | 24/07/2023 | 25/11/2023 | 124 | 7.4 |
| DR08 | 25/07/2023 | 26/11/2023 | 124 | 7.3 |
| DR09 | 24/07/2023 | 25/11/2023 | 124 | 5.8 |
| FLB | 26/07/2023 | 14/12/2023 | 141 | 6.1 |
| FUB | 26/07/2023 | 14/12/2023 | 141 | 4.7 |
The settlement substrates consisted of square section, black, plastic tubes of width 3 cm and length 20 cm (providing a settlement area of 240 cm2) attached to plastic canes inserted into sediment such that the tubes were raised around 50 cm from the seabed (Figure 3). At each location five replicate tubes were employed around 2 m apart along an isobath. To maximise comparisons with the 2001 settlement study at Rubha Dearg, Loch Creran by Chapman et al. (2007), at the same location in 2023, in addition to the deployment of five replicate tubes, five replicates of the same substrate units employed by Chapman et al. (2007) were used, each in the form of four vertically orientated, contiguous left valves of the scallop Pecten maximus attached to plastic canes and raised around 50 cm above the seabed. To examine recruitment levels to existing serpulid reefs, five replicates of a third substrate employed at Rubha Dearg each consisted of an uninhabited reef fragment previously collected from the loch, of the order of 20 cm long by 5 cm wide, attached to a plastic cane inserted into the sediment and raised around 50 cm above the seabed. At all locations except Rubha Dearg a second line of five replicate tube treatments was also deployed for long-term monitoring of reef development.
All settlement replicates were retrieved (except those for long-term monitoring) after a period of 124 days in Loch Creran and 141 days in Loch Feochan. The timing was chosen to be similar to that used by Chapman et al. (2007), to embrace most of the settlement period of Serpula vermicularis in the loch as determined by Chapman et al. (2007) and to allow sufficient post-settlement time for recruits to grow to a sufficient size to facilitate species identification. At the time of retrieval a visual indication of the scale of recruitment onto adjacent reef material was obtained at three of the sites (DR02, DR08, DR09) by close-up still photography using a Nikon D200 camera with 60 mm macro lens.
Upon retrieval replicate units were placed in seawater in separate plastic bags for transport to the laboratory. This prevented the chance of worms expelled from their tubes being excluded from the subsequent counts. All material was preserved in 5% seawater formalin. Prior to counting all settlement surfaces were photographed using a Nikon D7100 camera with 105 mm macro lens. All occupied serpulid tubes were counted and identified. The presence of heavy fouling by other taxa, such as the sea squirt Ascidiella aspersa, was also noted.
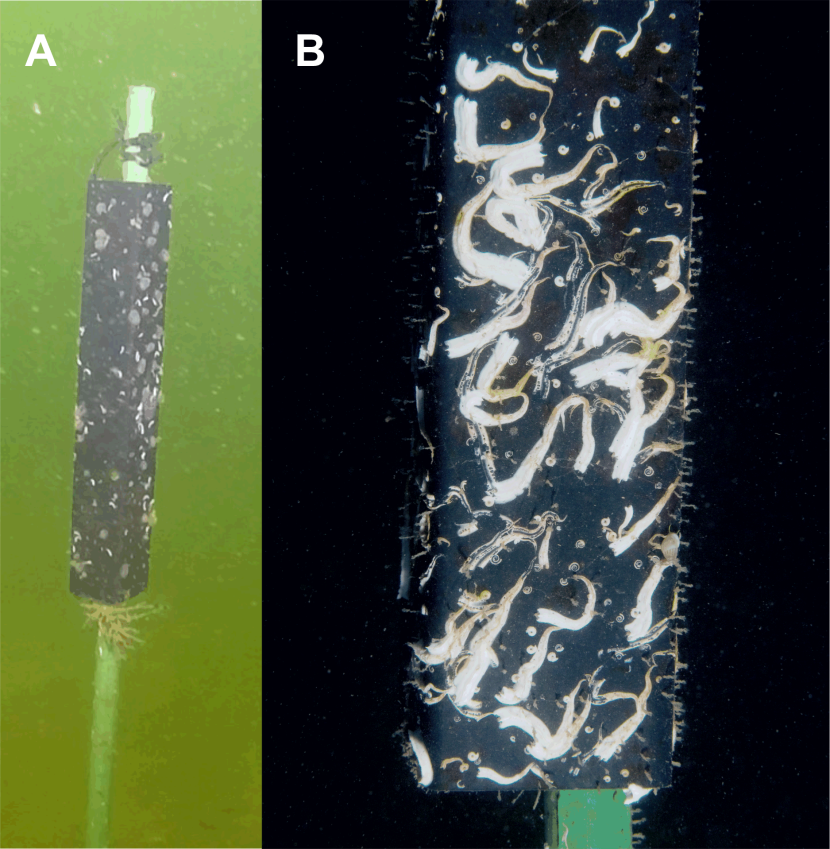
Figure 3. Settlement tube showing mode of deployment (A) and close-up of settlement surface with heavy recruitment of Spirobranchus spp. (B).
Click for a full description
Distant photo of settlement tube attached to upper end of plastic stake, and close-up of dense settlement of serpulid worms dominated by Spirobranchus spp. on the plastic tube.
Surface area of the scallop substrate was determined by importing scaled scans into Adobe Photoshop and employing the object measurement tool. Surface area of reef fragments was determined by building a 3D model by laser scanning using a Creality CR Scan Lizard scanner and importing the model to Blender software for area measurement. Some reef replicates required subdivision (achieved by cutting with a Dremel drill) to facilitate successful model production, and some accidental breakages occurred during this process. The resulting smaller reef fragments were sometimes difficult to laser scan successfully. For these small fragments surface area was estimated from a second order polynomial regression equation of surface area against dry weight for all reef fragments where 3D modelling was successful (n=25). Figure 4 shows a typical example of a reef fragment model in comparison to the source object. Although the claimed accuracy of the scanner was 0.05 mm, some loss in detail of the object is apparent. This was considered to be insufficient to invalidate the methodology.

Figure 4. Example of reef fragment employed in the recruitment study, showing the original object (left) and 3D model (right) rotated to display a similar orientation.
2.3 Predator exclusion
The predator exclusion study was initiated at a depth of c.6 m in Port Columba Bay, Loch Creran, during the period 15 - 20th August 2019 (Figure 1). Two different settlement substrates were deployed. One consisted of cylindrical bundles (approximately 21 x 10 x 6 cm) of fresh natural serpulid reef fragments (without live Serpula) bound together by cable ties. The other consisted of similarly sized bundles of scallop shells. The substrates were attached to the upper surface of a standard house brick to provide stability.
Exclusion cages were built from a plastic coated metal grid with a mesh size of 7 cm. This was assembled to form cuboids of side 45 cm and the top and sides of the cuboid covered with 4 mm plastic mesh.
Concrete paving slabs (45 x 45 cm) were deployed on the seabed and a brick with attached substrate was placed in the centre of each slab. Cages were placed over the slabs such that the base of the cage overlapped the edge of the slab and the cage was attached to the slab by cable ties.
For each substrate type there were 3 experimental treatments:
- substrate uncaged but placed on a paving slab
- substrate within an exclusion cage
- substrate within a control cage, which was identical to the exclusion cage except the lower 7 cm of the mesh was removed to allow access by predators
The experimental units were arranged in five distinct plots all at similar depths and with approximately 5 m between adjacent plots. Within each plot one replicate of each of the six experimental units was deployed haphazardly with 2 to 3 m separating adjacent experimental units. Overall, this provided five replicates of the six experimental treatments.
The site was revisited on 22nd - 23rd August 2022 in order to monitor reef development and to remove and clean the cages, as well as removing observed predators. Monitoring consisted of GoPro video footage, in situ observations and illustrative stills photography. This was repeated on 28th June 2023.
No well-defined reefs developed on any of the substrates during the experimental period. Recorded metrics included the number of tubes present and maximum tube height. It should be noted that in situ tube counts could not include small, newly settled juveniles, but only included the larger established adult tubes that could be readily distinguished by the diver. It should also be noted that the tubes included those of Serpula vermicularis and Protula tubularia. Both were present, but were not readily distinguishable by their tubes. The distinction is obvious when the radioles are extended, but in most instances, these were retracted due to disturbance caused by the presence of the diver. However, it should be noted that Protula has been recorded at considerably lower densities on reef material in the loch (Chapman et al., 2011), as well as on settlement plates (this study). Divers also made observations on other aspects such as differences in epibiota and on the degree of deterioration of the experimental substrate.
The coordinates for all experimental sites for all three studies described in this report are provided in Appendix 1.
3. Results
3.1 Reef development
3.1.1 Effect of substrate on reef development
Figure 5 shows the development of reef cover on the five substrate types at the main site over the ten year period of the investigation. Cover was markedly greater on the large cobble (overall mean 16.63 %) and large scallop bag (overall mean 10.96 %) units than the small scallop bag, loose scallops and boulder treatments, all of which had minimal reef development (overall mean covers <2 %). Reef height showed a broadly similar pattern (Figure 5) with mean heights attaining 26.33 cm and 18.67 cm for the large cobble and large scallop bag units in 2022 but <5 cm for the other treatments. The extent of reef development on one of the remediation units five years post deployment can be seen in Figure 6.
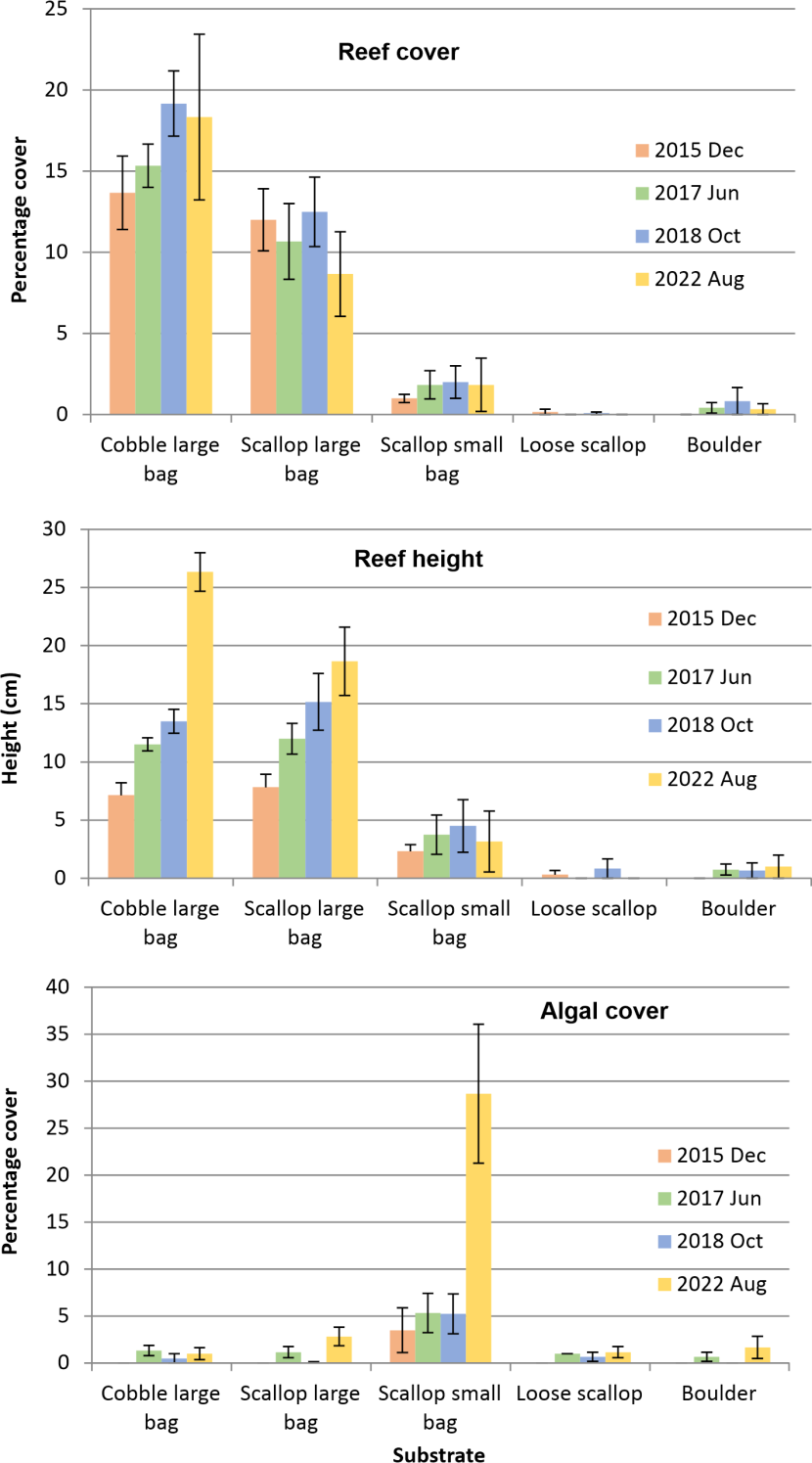
Figure 5. Effect of substrate type on the development of Serpula vermicularis reefs on experimental remediation units as percentage cover and maximum reef height, and the percentage cover of the units by algae.
Click for a full description
The bar graphs shows the effect of five substrate types (cobbles in a large mesh bag, scallop shells in a large mesh bag. scallop shells in a small mesh bag, scallops placed in a pile on the seabed, and a boulder) on serpulid reef development. Following deployment in March 2012 the graphs show the pattern of development in December 2015 (orange bars), June 2017 (green bars), October 2018 (blue bars) and August 2022 (yellow bars). The three bar graphs show the temporal pattern of mean percentage cover of the substrates by reefs, the mean maximum reef height and the mean percentage cover of the substrate by macroalgae. Error bars are standard error of the mean.

Figure 6. Reef development on a cobble replicate remediation unit five years post deployment.
The clearly minimal reef development on the smaller substrate units permitted statistical focus on the larger cobble and scallop bag treatments where repeated measure analyses could be employed in the form of two-way ANOVA testing for the effects of substrate type and year. A summary of statistical testing is provided in Annex 2. The Greenhouse-Geisser correction was applied to allow for a significant deviation from sphericity (Mauchly's test, p=0.004). The Shapiro-Wilks test revealed that data did not deviate from normal distributions, apart from the 2018 cobble replicate data (p=0.035), although a Q-Q plot indicated that deviation was minor. The ANOVA found no significant difference in reef cover between years (p=0.480) but mean reef cover on cobble was significantly greater than mean reef cover on scallop (p=0.042). In view of the slight deviation from normality, the results were confirmed by the non-parametric repeated measure Aligned Rank Transform ANOVA test (ART) (Wobbrock et al., 2011), which also showed no differences between years (p=0.439) but significantly higher reef coverage on the cobble units (p=0.028).
The reef height data for the cobble and scallop large bag treatments adhered to normality according to the Shapiro_Wilks test (p>0.102) and logged data passed Mauchly’s sphericity test, so this transformation was used in the repeated measure ANOVA, which revealed no difference between substrates (p=0.663) but significant variation in size between years (p<0.001). Post hoc comparisons revealed differences between all years (p<0.001) apart from 2017/2018 (p=0.350). The temporal pattern of reef growth on the cobble and scallop substrates is plotted in Figure 7. This shows a virtually linear relationship between reef height and time over the 10 year measurement period for the cobble units. The date of initiation of tube growth is unknown but the mean growth rate from 2015 to 2022 is 2.9 cm a-1. Growth is almost identical for the scallop units from 2012 to 2018 but slows from 2018 to 2022 attaining a mean of 1.6 cm a-1 from 2015 to 2022. There was a significant difference in mean reef height between the substrates in 2022 (t test, p=0.046).
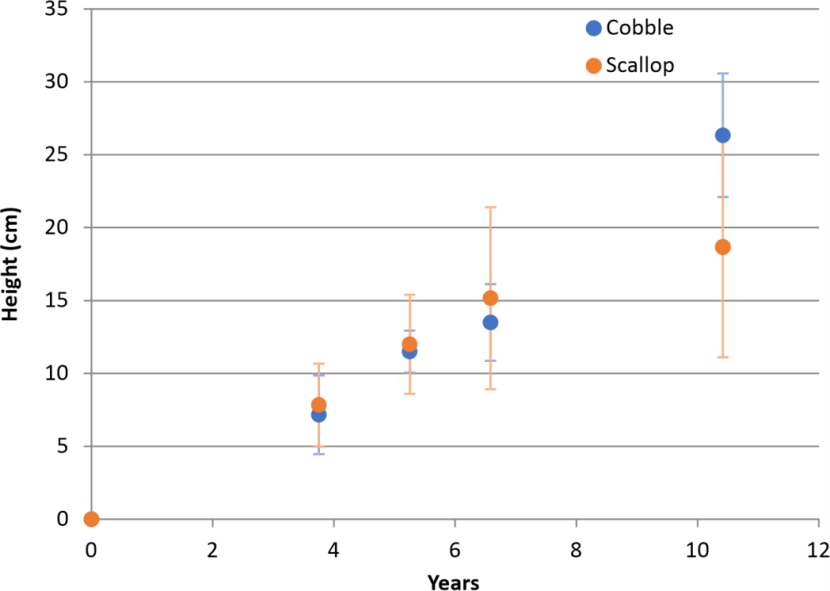
Figure 7. Temporal change in mean maximum height of reefs for replicate large bag remediation units containing scallop shells and cobbles.
Click for a full description
The scatter plot shows the increase in mean maximum size of reefs growing on large bag remediation units containing cobbles (orange dots) and scallop shells (blue dots) from the time of deployment up to around 10 years post-deployment. Error bars are standard error of the mean.
3.1.2 Effect of location on reef development
Figure 8 shows the development of reef cover on the large scallop bag units at the five locations in Loch Creran over the 10 year period of the investigation, although data for 2015 were not collected at the Mussel Farm and Upper Basin sites. Over the monitoring period as a whole it is clear that cover was markedly greater at the main site (overall mean 10.96 %) than all the other sites, where the mean did not exceed 4 % except at Rubha Mòr in 2017. Reef height (Figure 8) exhibited a similar pattern, with the mean maximum height of 18.67 cm at the main site more than double the greatest mean height recorded in any year at all the other locations. By contrast algal fouling of the experimental units was extensive at Rubha Mòr, reaching a mean of 86.67 % in 2022, moderately high at the Kelco and Mussel Farm sites (attaining around 30%), and minimal at the Main Site and Upper Basin locations (Figure 8).
Due to the absence of data at two locations in 2015, the loss of replicate identification tags at several sites from 2017 to 2022, and non-normal data, statistical analysis of the effect of location on reef development metrics was carried out using non-repeated measure ART ANOVA, which included data for all years except 2015. The results confirmed the patterns illustrated in Figure 8, revealing a significant difference in reef cover between locations (p<0.001), with post hoc testing (Elkin et al., 2021) showing higher cover at the Main Site than all the other sites (p <0.001 - p=0.026), and Rhu Mòr cover higher than at the Kelco, Mussel Farm and Upper Basin sites (p<0.001 - p=0.020). The ART ANOVA also revealed a significant overall temporal change in cover (p<0.001), but such patterns are considered below using, where possible, analyses employing repeated measures and 2015 data.
ART ANOVA also identified an overall location effect for reef height (p<0.001) with taller growth at the Main Site than elsewhere (p<0.001) and less growth at the Mussel Farm than either Rhuba Mòr (p<0.001) or Kelco (p=0.012).
Analysis of the algal cover data by ART ANOVA showed a locational difference (p<0.001) with higher fouling at Rubha Mòr than elsewhere (p<0.001). Higher algal cover was also found at Kelco and the Mussel Farm than at both the Main and Upper Basin sites (all p<0.001).
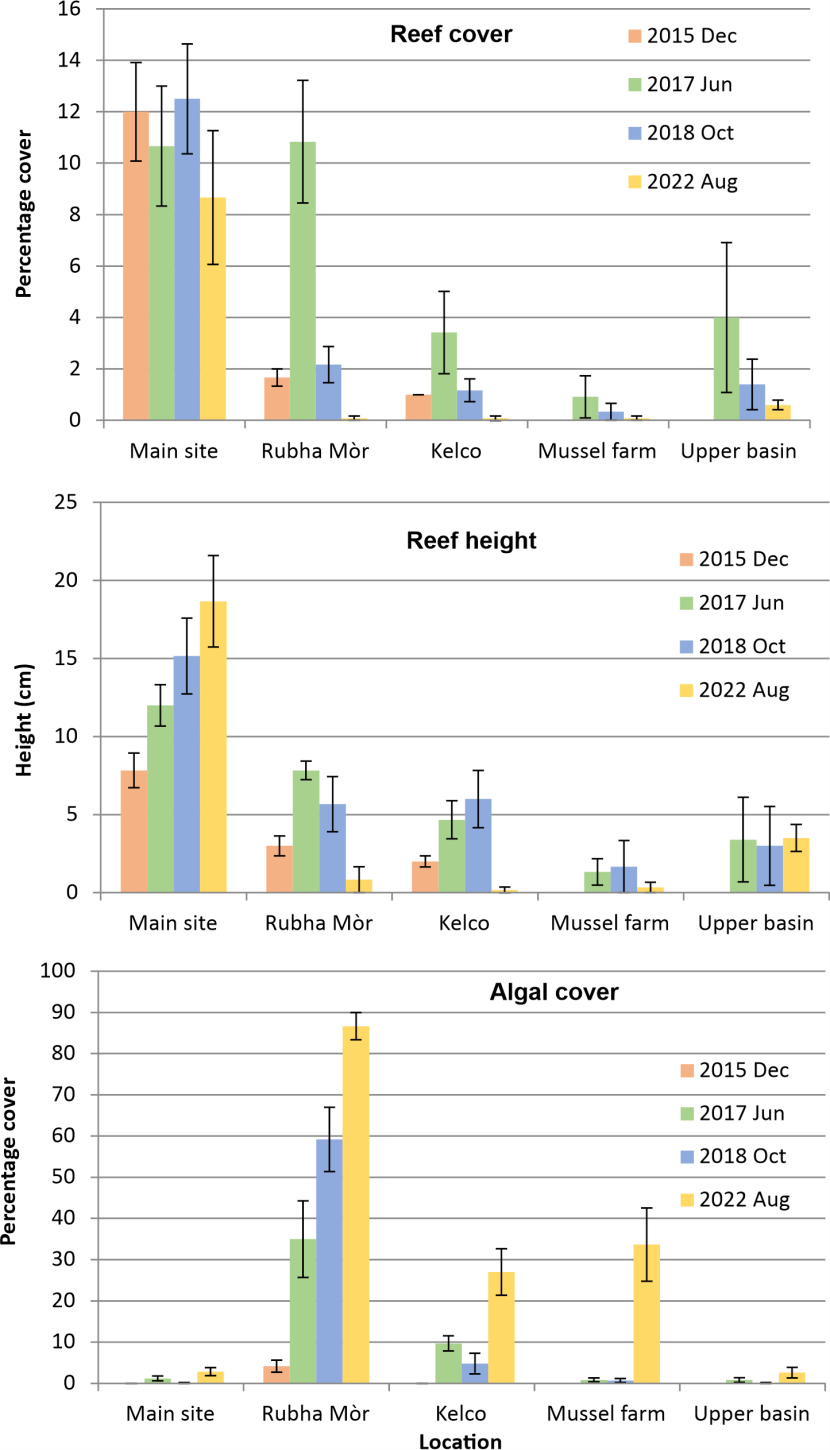
Figure 8. Effect of location on the development of Serpula vermicularis reefs on experimental remediation units as percentage cover and maximum reef height, and the percentage cover of the units by algae.
Click for a full description
The bar graphs shows the effect of location (i.e. the five sites in Loch Creran - Main Site, Rubha Mòr, Kelco, Mussel farm and Upper basin) on serpulid reef development. Following deployment of experimental remediation units in March 2012 the graphs show the pattern of development in December 2015 (orange bars), June 2017 (green bars), October 2018 (blue bars) and August 2022 (yellow bars). The three bar graphs show the temporal pattern of mean percentage cover of the units by reefs, the mean maximum reef height and the mean percentage cover of the substrate by macroalgae. Error bars are standard error of the mean.
3.1.3 Effect of location on temporal change in reef development
Temporal variation in reef development at the Main site has already been considered above. At Rubha Mòr the retention of replicate identity tags at the site throughout the study permitted the use of the repeated measure Friedman non-parametric test, followed by the Conover post hoc test, to assess change between years. The former was found to be significant (p=0.001). Although Figure 8 indicates a large peak in cover in 2017, similar to levels recorded at the Main site, the post hoc test revealed the only significant difference to be a large drop in the median from 10.0 in 2017 to 0.0 in 2022 (p=0.001). Reef height responded in a similar manner with the only significant difference between years being a drop from a median of 8.5 in 2017 to 0.0 in 2022 (p=0.023). Figure 8 shows a pattern of algal fouling increasing over the course of monitoring to reach a mean of 86.67 % in 2022. However, following a significant Friedman analysis result (p=<0.001), the only significant difference identified was between a median of 3.0 % in 2015 and 90.0 % in 2022 (p=0.023)
Replicate tags were lost at the remaining sites, so the less powerful Kruskal-Wallis test, followed by Dunn’s post hoc test with Bonferroni correction in the event of Kruskal-Wallis significance, was employed to test for temporal change at each location. At Kelco significant Kruskal-Wallis tests identified change in reef cover (p=0.003), reef height (p=0.008) and algal cover (p<0.001). Reef cover differed between 2017 (median 2.0 %) and 2022 (median 0.0 %) (p=0.002). Reef height differed between 2017 (median 5.0 cm) and 2022 (median 0.0 cm) (p=0.034) and between 2018 (median 6.5 cm ) and 2022 (p=0.012). Algal cover differed between 2015 (median 0.0 %) and 2022 (median 25.0 %) (p<0.001).
At the Mussel Farm site there was minimal reef development throughout the monitoring period and no significant temporal change in reef cover (p=0.700) nor height (p=0.724) according to the Kruskal-Wallis test. However, there was an overall increase in algal cover (p=0.003) with Dunn’s test showing increases from 2017 (median 0.5%) and 2018 (median 0.25%) to 2022 (median (25.0%) (respectively p=0.012 and 0.006).
In the Upper Basin no significant temporal change was recorded by Kruskal-Wallis analysis in reef cover (p=0.943), reef height (p=0.358) or algal cover (p=0.240).
3.2 Recruitment
Figure 9 shows a comparison between recruitment after a deployment of 124 days of serpulid species to the scallop, reef fragment and plastic tube settlement substrates at Rubha Dearg. Four taxa were recorded: Serpula vermicularis, Spirobranchus spp., Hydroides spp. and Protula tubularia. The serpulid fauna was strongly dominated by Spirobranchus spp., with Protula barely represented (mean 0.003 cm-2). Count data for Serpula and Hydroides were heteroscedastic and so Kruskal-Wallis tests were employed to identify any effects of substrate type on recruitment, both of which were not significant (p=0.087 and p=0.733 respectively). A oneway ANOVA test was used for the homoscedastic Spirobranchus data, which revealed a significant difference in mean density between substrates (p=0.005). A post hoc Tukey test showed higher recruitment to reef material than the tubes (p=0.004), but no significant differences between scallop shells and tubes (p=0.242), nor between scallop shells and reef fragments (p=0.074).
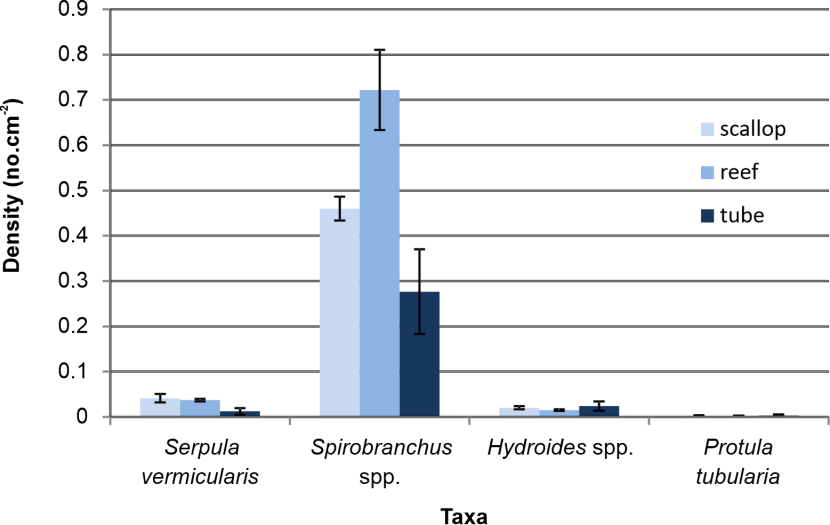
Figure 9. Recruitment of serpulid worms to scallop, reef and plastic tube settlement substrates at Rhuba Dearg, Loch Creran in 2023.
Click for a full description
The bar graph shows the recruitment of Serpula vermicularis, Spirobranchus spp., Hydroides spp. and Protula tubularia to three substrates: scallop shells (light blue), serpulid reef fragments (mid blue) and plastic tubes (dark blue). Bars represent mean density with attached standard error of the mean.
Figure 10 compares recruitment of the dominant serpulid taxa to similarly deployed scallop shell substrates at Rubha Dearg in 2023 and 2001 (Chapman et al., 2007). As all data sets were homoscedastic and showed no significant deviation from normality, T-tests were used to identify temporal differences in recruitment density for each taxon. All taxa showed significant temporal decreases in density (p<0.001), four-fold in the case of Spirobranchus, ten-fold for Hydroides and twenty-eight-fold for Serpula (from mean of 1.164 cm-2 in 2001 to 0.041 cm-2 in 2023).
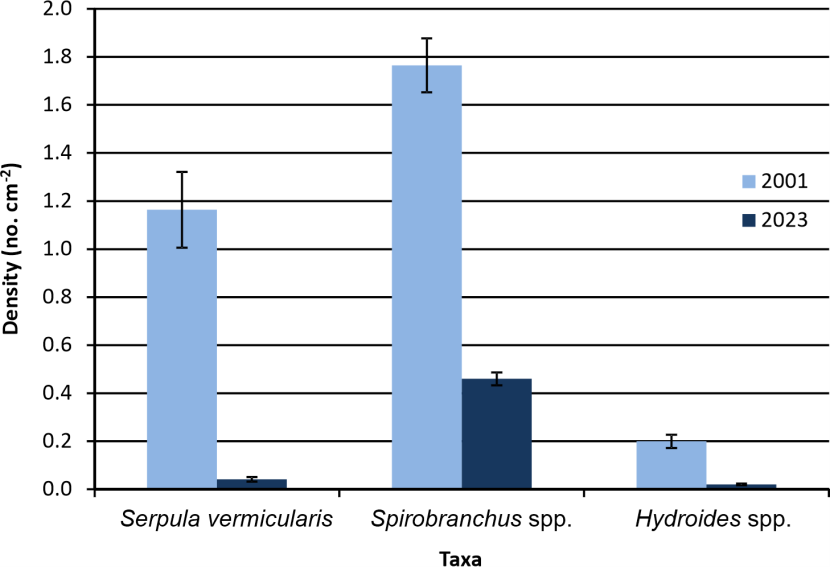
Figure 10. Recruitment of serpulid worms to a scallop shell settlement substrate at Rubha Dearg, Loch Creran in 2001 and 2023.
Click for a full description
The bar graph shows the recruitment of Serpula vermicularis, Spirobranchus spp. and Hydroides spp. to a scallop shell substrate in 2001 (light blue) and 2023 (dark blue). Bars represent mean density with attached standard error of the mean.
Spatial variation in serpulid density for the six sites in Loch Creran and two sites in Loch Feochan is illustrated in Figure 11. Due to data heteroscedasticity and non-normality locational differences between medians for each taxon were assessed with the Kruskal-Wallis test, which revealed no significant differences in the case of Serpula vermicularis (p = 0.164) and Spirobranchus spp. (p = 0.062). The median density of Hydroides spp. did vary significantly (p = 0.009), with Dunn’s post hoc test showing the only difference to be a higher density in the lower basin than the upper basin of Loch Feochan (p = 0.013).
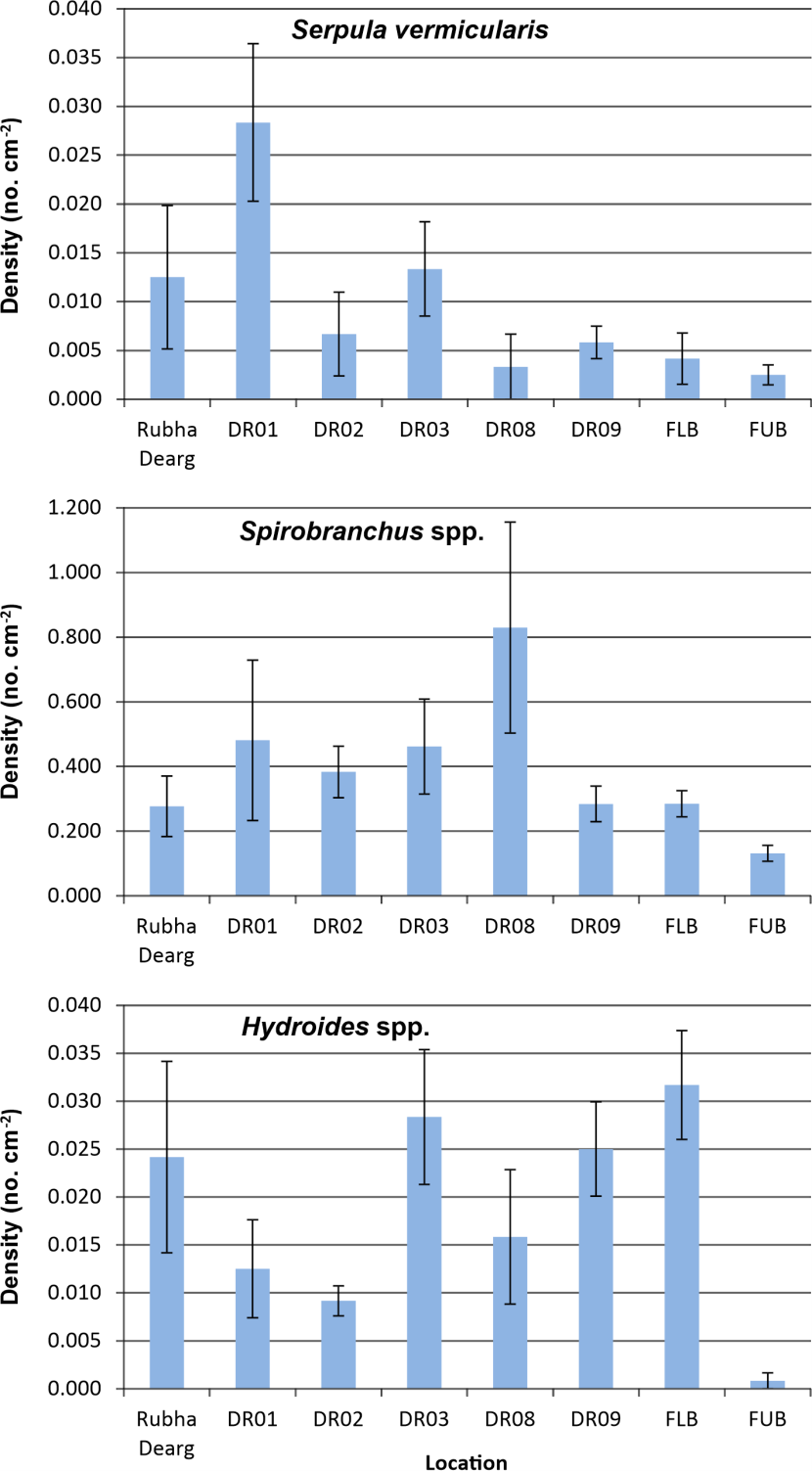
Figure 11. Recruitment of serpulid worms to a plastic tube settlement substrate at eight locations in Loch Creran and Loch Feochan in 2023.
Click for a full description
The bar graph shows the recruitment of Serpula vermicularis, Spirobranchus spp. and Hydroides spp. to a plastic tube substrate at six locations in Loch Creran (Rubha Dearg - DR09) and Loch Feochan (FLB, FUB) in 2023. Bars represent mean density with attached standard error of the mean.
3.3 Predator exclusion
Results from monitoring visits in 2022 and 2023 were broadly similar. The 2023 data are presented here.
The serpulid reef bundles underwent considerable fragmentation resulting in a loss of much of the originally available substrate (Figure 12A). For the open control treatment three replicates were entirely lost and a fourth reduced to about 10% of the original substrate volume. For the control cage treatment four replicates were entirely lost and only about a quarter of the original substrate remained on the fifth. The cage treatment was less degraded, with one replicate entirely lost and some loss of volume for two further replicates.
Due to the loss of replicates it is not possible to statistically test for differences in the degree of serpulid colonisation of serpulid reef substrate. The data have nevertheless been presented below (Figure 12). Weighting for calculation of mean values has been adjusted for reduction in the amount of substrate where appropriate.
None of the scallop substrate treatments showed any major deterioration. The substrate loss for the caged serpulid reef treatment was lower than the corresponding control treatments (Figure 12A), although not significantly so (Kruskal-Wallis test, p=0.095).
The number of live tubes growing on the caged scallop substrate was not significantly different from that on either of the corresponding control treatments (Kruskal-Wallis test, p=0.662) (Figure 12B). The number of tubes on caged serpulid substrates was slightly less than on caged scallop substrates, but the difference was not significant (T-test, p=0.372 - data homoscedastic and normal).
For the serpulid reef substrate there appears to be a greater mean maximum tube height on the caged treatment (Figure 12C), but this cannot be tested due to the loss of control replicates. For the scallop substrate the highest mean maximum tube height was recorded for the caged treatment, but the median was only significantly higher than the cage control (Kruskal-Wallis test followed by Bonferroni-corrected Dunn’s test, p=0.036). The maximum tube height on caged serpulid substrates was significantly less than on caged scallop substrates (T-test, p=0.026 - data homoscedastic and normal).
Algal turf species, solitary ascidians, Ophiothrix fragilis and Spirobranchus triqueter were prominent components of the epibiota developed on the experimental substrates. Of these, the algal turf was particularly profuse and was considerably more abundant on the open controls than on either of the caged treatments (Figure 12D).
While the results from this cage study might be suggestive of an effect of macrofaunal exclusion on reef decay and serpulid tube length, the level of statistical evidence precludes further consideration of these aspects in this report.
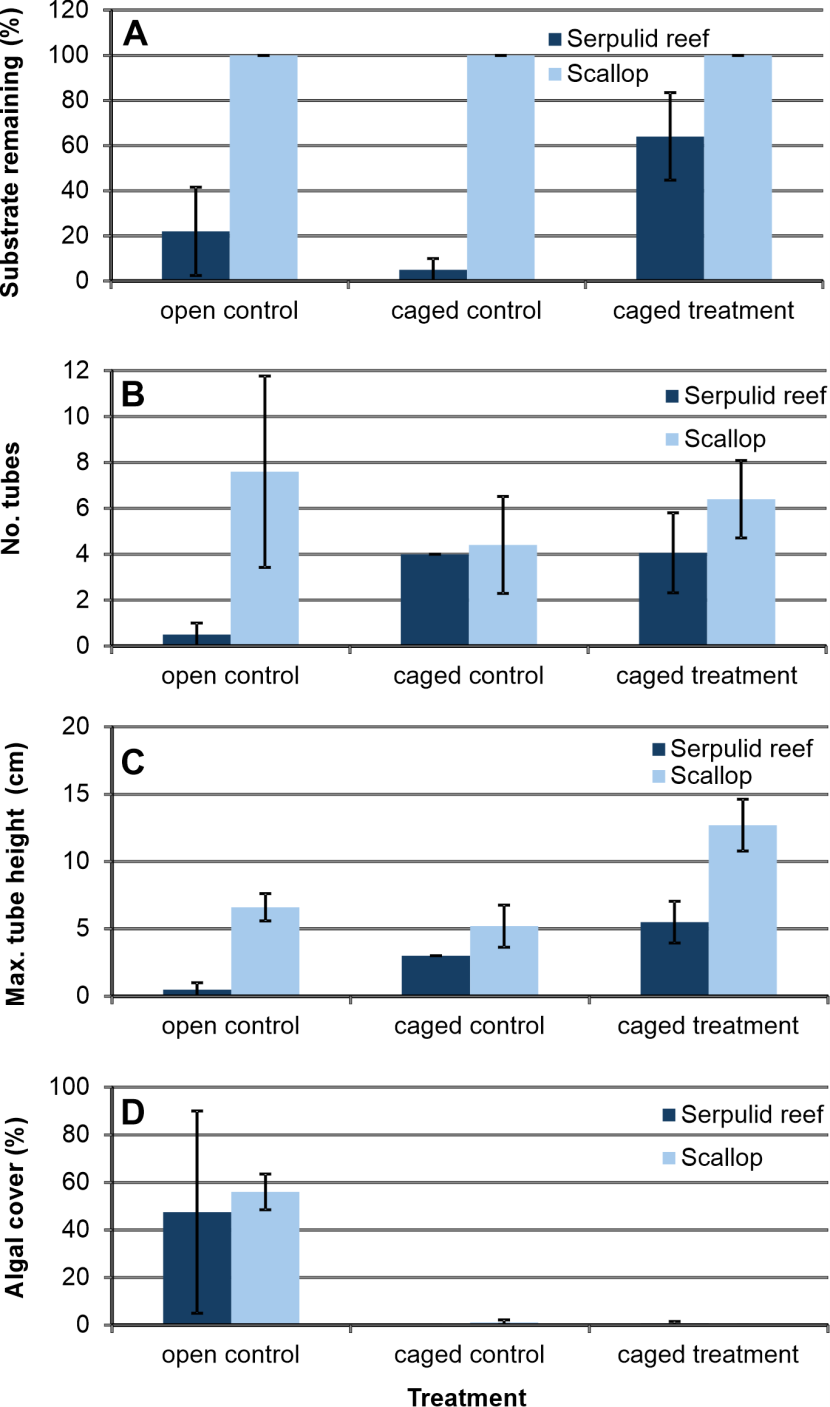
Figure 12. Effect of exclusion cage treatment on the condition of serpulid reef fragments and scallop shell substrates over the period August 2019 - June 2023.
Click for a full description
Bar graphs showing the effect of three types of exclusion cage treatment on the condition of serpulid reef (dark blue bars) and scallop shell substrates (light blue bars) over the period August 2019 - June 2023 in Loch Creran. The treatments were open control (no cage), caged control (cage with panel for predator access) and caged treatment (fully caged). Condition factors recorded were: A, percentage of substrate remaining at end; B, number of adult recruits (Serpula vermicularis and Protula tubularia); C, maximum tube height of recruits (S. vermicularis and P. tubularia); D, percentage cover by algal turf. Bars represent mean values, with attached standard error of the mean.
4. Discussion
4.1 Reef development
Although firm evidence for widespread serpulid reef degradation in Loch Creran was not revealed until 2013, there is some indication that a process of widespread reef decay was present several years before, possibly from around 2009 (Moore et al., 2020). It is therefore likely that the experimental remediation work (March 2012 to August 2022) fell within the period of decay. The five locations used for this work differed in terms of the pre-decline status of the serpulid reef habitat as revealed by monitoring work in 1994 (Moore, Saunders and Harries, 1998) and 2005 (Moore et al. 2006), as well as unpublished observations. The location of the Main and Rubha Mòr sites were in areas previously supporting extensive, dense populations of large reefs. Quantitative measures of reef cover in 2005 provided mean cover values of 8.9 % and 16.9 % respectively (locations termed respectively Sea Life Centre and South Shian in Moore et al. (2006)). In 1994 both locations were placed in the highest abundance category (>5 % cover) based on visual estimation by diver. Large reefs (defined as having a plan area >500 cm2) were recorded at the Mussel farm site in 1994 and 2005, but the maximum density recorded in the region in 1994 was <1 % cover. Reef development was even poorer in the region of the Kelco and Upper Basin sites, with narrow bands of sparse, relatively small reefs (Moore, Saunders and Harries, 1998; Moore et al., 2006). It might be expected that the environmental conditions responsible for the historical pattern of reef development at these locations, although not understood, might be reflected in the level of success of colonisation of the experimental units. This is partially true, with much higher reef cover values recorded for the Main Site and Rubha Mòr locations. However, this was only the case for 2017 at Rubha Mòr. At least from 2017 to 2022 the high levels of algal fouling recorded at Rubha Mòr (rising to a mean of 87%) could be a contributing causative factor. No correlation was apparent between reef development on the experimental units and the vicinity of point sources of pollution, such as aquaculture facilities.
Despite the widespread degradation of reefs at the Main Site and Rubha Mòr, with no visible evidence of recovery of fragmented reef material during the experimental period, the deployment of fresh substrates did lead to significant reef development at these locations, fuelling some optimism in the potential for habitat remediation. However, this development did not persist. At the Main Site following initial successful recruitment in the first three years resulting in 14 % and 12 % reef cover for the large cobble and scallop bag treatments respectively in 2015, no further significant temporal increase in cover occurred. At all the remaining sites, following lower initial levels of reef development, cover values decreased from 2017 to 2022, although not significantly so at the Mussel Farm and Upper Basin sites. Subsequent to the preparation of this report further visits to the restoration units were made in 2025. These data indicate a continued trend of degradation of serpulid reefs on the restoration units (Report in prep).
Hughes, Poloczanska and Dodd (2008) measured the tube extension rate of Serpula vermicularis in the loch in 2004 - 2005 at 3.3 cm a-1. Chapman (2004) measured the mean increase in length of recently settled worms over a two year period (2001 – 2003) as 5.7 cm a-1. Although higher than the estimate of Hughes, Poloczanska and Dodd (2008), the latter authors reported a significant negative correlation between body weight and tube growth, suggestive of a reduction in growth rate with age. Early tube growth is prostrate and subsequent upward growth is sinuous and so the rate of height increase of reefs will lag behind tube extension rate. In fact, the pre-decline reef growth rate in Loch Creran has been estimated at 3.0 cm a-1 based on measurement of the height of reefs growing on artificial substrates introduced to the loch (Moore et al., 2006).
In the current study while no persistent temporal increase in reef cover was recorded at Main Site, reef material continued to grow upwards throughout the monitoring period, attaining mean rates of height increase of approximately 2.9 cm a-1 and 1.6 cm a-1 for the cobble and scallop bag treatments respectively. The similarity of at least the growth on cobbles to the pre-decline reef growth estimate is suggestive that serpulid growth at a similar rate to pre-decline levels is still possible in the loch. This may reflect the absence of a temporal reduction in the physiological condition of adult serpulids, which might be expected in the presence of trophic or pollution pressures. At locations and on substrates where little or negative growth was recorded, this may reflect long-term historical differences in causative factors, such as current speed (and hence food supply), as well as grazing pressure.
4.2 Recruitment
4.2.1 Substrate preference
No significant difference in recruitment of Serpula vermicularis after 124 days immersion was found between the scallop, reef and plastic tube substrates. Mean S. vermicularis density on plastic tubes was around one third that recorded for the scallop and reef substrates (Figure 9) but the difference was not quite statistically significant (p=0.087). It should be noted, however, that recorded recruitment levels do not necessarily reflect settlement preferences, as grazing success by predators such as Psammechinus miliaris, which is present at high densities in Loch Creran(Moore et al., 2020) might be expected to be greater on a smooth plastic surface than on the more rugose scallop and reef substrates. For the much more abundant taxon, Spirobranchus spp., recruitment levels did indeed reflect the trend of increasing rugosity (plastic tube < scallop < reef) (Figure 9), although the difference between the reef and scallop substrates was not quite significant (p=0.074).
4.2.2 Spatial variation
No significant spatial variation in the density of Serpula vermicularis recruits was recorded, although mean density varied widely from 0.003 cm-2 to 0.028 cm-2. Possible linkages of spatial patterns to anthropogenic drivers are not evident. For example, Figure 13 shows the relationship between recruitment density and distance from the salmon farm installation (taken as the midpoint between the two cage sites employed, as revealed by satellite imagery). Clearly there is no correlation. Although the highest density was recorded at the most distant study site, the second highest value was obtained from the site closest to the cages (620 m).

Figure 13. Relationship between recruitment density of Serpula vermicularis and distance of study location from the salmon farm installation in Loch Creran.
Click for a full description
Scatter plot showing the relationship between mean recruitment density of Serpula vermicularis to plastic tube substrates at six locations in Loch Creran and distance of the location from the salmon farm installation in 2023.
4.2.3 Temporal change
Comparisons of the results of the current study with previous studies of Serpula vermicularis recruitment are complicated by variation in the methodologies employed, which are likely to strongly influence perceived differences in recruitment densities. Significant aspects of methodology include such factors as settlement substrate type and orientation (including artificial and natural substrates), season and duration of deployment. Notwithstanding these limitations, Table 2 presents a comparison of S. vermicularis recruitment densities from the literature, including studies from areas both known to support reefs, and from areas where they are believed to be absent.
Several studies have examined recruitment in Loch Creran through the use of settlement plates, with immersion periods ranging from one to at least 20 months. The best comparative data for analysis of temporal change in serpulid recruitment in the loch is derived from the use of scallop shell substrates in the current study and that of Chapman et al. (2007) in 2001. Chapman et al. (2007) employed fewer replicates but their experiments included the deployment of vertical scallop shells raised 50 cm above the sediment at the same depth of 6 m and in the same location, Rubha Dearg, as the current work. The duration of immersion was slightly shorter, commencing on 19th August and ending on 17th November (90 days). The current 2023 experiment extended from 25th July to 26th November (124 days). In another experiment Chapman et al. (2007) found settlement to occur predominantly from June to October and so the 2023 study embraced slightly more of this recorded settlement period. However, recorded densities represent a balance between settlement and mortality, with the latter also likely to increase with longer immersion times. Grazed and empty serpulid tubes were observed during the enumeration of recruits in 2023, but not at a level which could contribute significantly to an explanation of the twenty-eight-fold temporal decline in Serpula recruitment recorded between 2001 and 2023 (from a mean of 1.164 cm-2 to 0.041 cm-2). The level of mortality is unknown for the 2001 study.
Following the 2001 study, recruitment to vertical stone settlement plates in the loch was examined in 2013 - 2014 within the period of serpulid reef decline (Cook, 2016; Cook et al., 2021). A study of locational differences within the loch recorded median densities similar to mean values for 2023 at Main Site (0.078 cm-2), which was located close to Rubha Dearg, although higher densities were recorded elsewhere, with the maximum at the Mussel Farm site (0.366 cm-2). This locational study employed an immersion period of 12 months and so there was scope for both additional mortality and a prolonged period of recruitment. Cook’s seasonal study at Main Site perhaps provides a better comparison. This involved deploying plates every two months from January 2012 to November 2012, with retrieval in November 2013 (i.e. immersion periods varying from 12 - 22 months. Thus, although deployment was for longer than in 2001 and 2023, all plates will have experienced the recent summer recruitment peak, with some plates in the Cook study being immersed for summer peaks in two years. Depending upon the month of deployment median recruitment densities varied from a minimum of 0.083 cm-2 for November deployed plates (12 months immersion) to 0.235 cm-2 for July deployed plates (20 months immersion). These values are much lower than those recorded by Chapman’s 2001 90 day immersion study, but slightly higher than the 2023 study. Despite the methodological differences between the various recruitment studies, they are strongly suggestive that there has been a temporal decline in early recruitment between 2001 and 2023.
4.4.4 Larval retention
The probable cause(s) of a decline in recruitment of Serpula vermicularis in Loch Creran is unknown. It may be related to the reduction in size of the adult population in the loch, but this would be predicated on the existence of a process for the retention of competent larvae within the waters of the loch. Indeed, larval retention has been proposed as a contributory factor in the development of reefs in some serpulid species in poorly flushed waters (e.g. Bosence, 1979). A corollary of this thesis is that high densities of larvae will be generated with sufficiently short pelagic durations to avoid flushing from the water body. Indeed, several of the known instances of S. vermicularis reef development have been recorded in seemingly poorly flushed locations, such as Ardbear Lough and Linne Mhuirich (Bosence, 1979). However, serpulid reefs are also known to occur in well-flushed areas, such as in Blacksod Bay, which is open to the Atlantic Ocean (MERC Consultants, 2008). Here, S. vermicularis forms small reefs on a maerl and sand substrate. The related S. narconensis forms possibly the largest serpulid reef habitat in the world at a depth of 91 - 105 m in open Atlantic Ocean waters off South Georgia (Ramos and San Martin, 1999).
Serpula vermicularis produces planktotrophic larvae. There appears to have been no in situ studies of the duration of the planktonic stage. Holt et al (1998) state that comparisons with other serpulid species suggest that it may lie somewhere between six days and two months, but a subsequent comprehensive summary of the development of serpulid species (Kuprianova et al., 2001) reveals that the shorter development times (up to 10 days) all relate to studies carried out at temperatures >20 °C (where the temperature was recorded). They state that the development time of the pelagic feeding stage in serpulids is profoundly affected by temperature.
Based on observation of laboratory-reared individuals derived from Serpula vermicularis from Ardbear Lough, Leahy (1991 - cited in Chapman, 2004) estimated the length of the planktonic trochophore larval stage to be 42 days at 12°C. Young and Chia (1982) reared serpulids from Friday Harbor on the US Pacific coast at the same temperature and recorded metamorphosis to occur after 41 to 50 days, although some larvae adopted a benthic mode of life as early as 28 days. Their source material was identified as S. vermicularis but has subsequently been referred to the closely related S. columbiana (Kupriyanova, 1999; Kupriyanova et al., 2001). McGibney (2016) also reared serpulid larvae ascribed to S. vermicularis derived from material collected around 600 km farther south along the Pacific coast, recording a larval duration of 31 days, with the benthic phase commencing after 20 days.
Daily CTD readings at a depth of 6 m in the lower basin of Loch Creran in 2013 show the temperature rising from around 10 °C at the beginning of June to peak at around 13.5 °C during August and September (Cook, 2016). With the bulk of settlement of Serpula vermicularis occurring from mid-June to mid-October (Chapman et al., 2007), most pelagic larval development is likely to take place over this temperature range. Based on the larval development data from S. vermicularis and the closely related S. columbiana, some of which was obtained at a similar temperature to that experienced by Loch Creran larvae, it would be surprising if the pelagic larval phase in Loch Creran was significantly shorter than three weeks.
According to the mathematical modelling of flushing time of Scottish lochs by Edwards and Sharples (1986) the flushing time of Loch Creran is 3 days (the precise figure being 2.84 days). Flushing rate is defined as the time during which 60 % of water in the loch is removed by tidal exchange. The authors caution that flushing time may be longer for water held in deep basins. The flushing time assumes complete replacement of tidally discharged water with water of non-Creran origin. Studies of Loch Etive (Edwards and Edelsten, 1977) have indicated a 50 % exchange efficiency and if this is applied to Loch Creran, flushing time is extended to 6.5 days, which may be a more realistic estimate (CEFAS, 2008).
Larval density reduction can be modelled by the exponential decay formula:
Nt = N0.e-λt
where N0 and Nt are densities after 0 and t days and λ is the decay rate, which for a reduction to 40% over 6.5 days is given by:
λ = ln(1/0.4)/6.5
This results in larval retention estimates of 5 % after 21 days and 2 % after 28 days. If most of the discharged water during the ebb tide is not returned on the flood then a 3 day flushing time would provide larval retention estimates of just 0.2 % and 0.02 % after 21 and 28 days respectively. These figures assume a well-mixed water body, and so may significantly under- or over-estimate larval retention. Without better knowledge of the hydrodynamics of the loch and larval duration it is impossible to conclude whether recruitment levels in Loch Creran might be strongly influenced by adult population density or whether larval retention is a significant factor in promoting reef development. Flushing time represents only one possible factor contributing to reef development, as according to the ranking of Scottish lochs by Edwards and Sharples (1986), Creran is fairly typical of Scottish lochs, with 50 lochs having the same or shorter flushing time (0-3 days) and 59 lochs with longer flushing times (4-14 days). The other lochs on the list where serpulid reefs are known to occur include one with a longer flushing time (West Loch Tarbert - 5 days) and two with shorter flushing times (Loch Ailort - 2 days, and Loch Teacuis - 1 day). Long flushing times will increase the degree of larval retention, but will also increase the potential for localised factors, such as pollution, to influence larval and adult survival.
If recruitment levels and ultimately reef development are related to enhanced densities of competent larvae, which in turn are dependent upon larval retention, then one might expect Loch Creran to exhibit enhanced levels of larval density, settlement and early recruitment in comparison to coastal waters in general. Table 2 includes studies of recruitment of Serpula vermicularis from well-flushed locations not supporting reefs for comparison with pre-decline levels in Loch Creran. In Bantry Bay, Ireland, Cotter, O’Riordan and Myers (2003) recorded very high recruitment (up to 4.4 cm-2) on plates deployed for one month in July, although most of the recruits died before they were identifiable to species and so this figure assumed that the death rate for unidentified serpulid individuals during the month was similar for all serpulid species recorded. Beaumont (2006) recorded densities of S. vermicularis recruits from 0.02 - 0.14 cm-2 at several stations in Loch Linnhe over a 15 month panel deployment. Although this appears lower than several of the pre-decline Loch Creran estimates (up to 1.7 cm-2 - Chapman et al., 2007), the Loch Linnhe depths were significantly greater (15 m compared to mostly 6 m in Loch Creran). In Loch Creran Chapman et al. (2007) found recruitment density at 12 m depth to be markedly lower than that at shallower depths (0 - 6 m), with the density at 12 m very similar to that in Loch Linnhe (viz. 0.01 – 0.13 cm-2). Beaumont’s panel deployment period was, however, longer than that of Chapman’s, covering two probable peak settlement periods, although a study of mortality rate by Chapman (2004) recorded around 90 % post-settlement mortality during the first year. Bearing in mind the methodological differences and the paucity of comparative studies, no firm conclusions can be reached regarding spatial differences in early recruitment, although there are no strong indications of higher pre-decline levels in Loch Creran. Post-decline estimates (this study) found no significant difference between early recruitment densities in Loch Creran and Loch Feochan, despite the presence of the very large numbers of the species that manage to persist in Loch Creran.
| Author | Study | Period (months) | Date | Density (cm-2) | Notes | Location | Depth (m) | Reefs present |
|---|---|---|---|---|---|---|---|---|
| Bosence, 1979 | Depth study | 0.5 | Aug 1972 | 0.013 – 0.45 | lower surface of slates | Ardbear Lough | 4 - 17 | Yes |
| Cotter, O’Riordan and Myers, 2003 | Seasonal study | 1 | Feb 1999 - Dec 2000 | 0 - c.4.4 (July) | assuming Serpula and Spirobranchus have similar death rate on panels (most individuals died before firm ID possible) | Bantry Bay | 4 - 7 | No |
| Chapman et al., 2007 | Depth study | 3 | Aug - Nov 2001 | 0.872 -0.916 | vertical scallops | Loch Creran | 0 | Yes |
| Chapman et al., 2007 | Depth study | 3 | Aug - Nov 2001 | 1.164 -1.501 | vertical scallops | Loch Creran | 6 | Yes |
| Chapman et al., 2007 | Depth study | 3 | Aug - Nov 2001 | 0.012 - 0.133 | vertical scallops | Loch Creran | 12 | Yes |
| Chapman et al., 2007 | Seasonal study | 1 | Apr 2000 - Feb 2001 | 0.000 - 0.110 | horizontal scallops, lower surface | Loch Creran | 6 | Yes |
| Chapman et al., 2007 | Substrate study | 3 | Aug 2001 - Nov 2001 | 1.67 | vertical scallops | Loch Creran | 6 | Yes |
| Chapman et al., 2007 | Substrate study | 3 | Aug 2001 - Nov 2001 | c. 0.25 | live reef | Loch Creran | 6 | Yes |
| Chapman et al., 2007 | Substrate study | 3 | Aug 2001 - Nov 2001 | c. 0.7 | dead reef | Loch Creran | 6 | Yes |
| Beaumont, J. C., 2006 | Reef type and distance, predation | 15 | Aug 2003 - Oct 2004 | c. 0.016 - 0.136 | vertical plates | Loch Linnhe (12 sites) | 15 | No |
| Cook, 2016 | Seasonal study | 12-22 | Jan2012 - Nov 2013 | 0.083 - 0.235 | vertical stone tiles | Main site, Loch Creran | 6 | Yes |
| Cook, 2016 | Location study | 12 | Feb 2013 - Feb 2014 | 0.078 - 0.366 | vertical stone tiles | 5 Creran sites | 6 | Yes |
| Cook, 2016 | Gregariousness study | 20 | Apr 2013 - Sep 2014 | c.0 | vertical stone tiles | S. Shian, Loch Creran | 6 | Yes |
4.3 A mechanism for serpulid reef decay
The purpose of this report is to provide details of studies which may contribute to an understanding of the cause(s) of serpulid reef decline, particularly as it relates to Loch Creran. It is not intended to provide an exhaustive account of the many factors which may possibly have contributed to the decline (but for which there is no strong evidence), such as anthropogenic stressors (e.g. point source and diffuse contamination) and long-term changes in characteristics of pelagic waters (e.g. plankton, temperature and salinity), although such changes are known to have occurred (see for example Tett et al., 2008; Whyte, 2012). It is possible however, to postulate a possible mechanism for reef decay which is consistent with the findings of the studies described herein without invoking excessive speculation.
Serpulid reefs undergo a natural process of growth and fragmentation that has been described for Loch Creran (Moore et al., 2020) and Ardbear Lough (Bosence, 1979). This involves the development of structures so large (up to 1 m in height) that the strength of the calcareous framework can no longer support the weight. Bosence (1979) suggested that the base of the reef becomes weakened by biological erosion caused by agents including boring sponges and algae and tube biting predators such as fish and echinoids. Another factor is likely to be the heavy burden of epibionts that accrue on larger reefs.
The presence of serpulid reefs in Loch Creran remained unrecorded between the original report of their existence in 1882 (Smith, 1887) and their finding over 100 years later at five locations in the loch in 1990 by Connor (1990). At two of the sites (Port Columba Bay and off Rubha Garbh south) Connor (1990) noted that Serpula vermicularis formed unusually large ‘mini-reefs’ up to 20 cm across. By 2005 much larger reefs had become widespread including in Port Columba Bay and off Rubha Mòr (close to Rubha Garbh) (see Moore et al., 2020) and so 1990 - 2005 appears to represent a period of reef growth. In 2005, despite the widespread presence of pristine reefs, many of the larger reefs attaining heights of around 60 cm had started to collapse, forming petaloid, ring-shaped structures, clearly discernible on sonar imagery (Moore et al., 2009). Also as early as 2005 the basal region of many of the reefs were heavily fouled with epibionts, especially ascidians, such as Pyura microcosmus and Ascidia mentula, completely masking the calcareous substrate (Figure 14B). In some cases fouling extended to much of the reef surface including more distal regions (Figure 14C).
In settlement experiments in Loch Creran recruitment success of Serpula vermicularis has been linked to the time of year of settlement panel deployment (Cook et al., 2021). Panels deployed shortly before the summer settlement period were found to result in significantly higher recruitment density than panels deployed several months previously (all panels being retrieved at the same time). As panels with the longer immersion times were found to be not fouled by macroscopic taxa, it was proposed that the development of biofilms might represent the density limiting factor, as bacterial films have been found to adversely influence settlement success in other invertebrate species including serpulid worms (e.g. Huang et al., 2011 for Hydroides elegans). In any event these findings appear supportive of the contention that fouling in one form or another may represent a significant factor in recruitment success.
Additional evidence for deterioration in the attractiveness of aged substrate to serpulid recruits is provided by observations of minimal colonisation of fragmented reef material in the loch. Three metre wide tracks of reef rubble, possibly caused by dredging activity, were first observed off Rubha Mòr in 1998 and were still clearly evident in sidescan imagery obtained in 2005 and 2017 (Moore et al., 2020; Scottish Government, 2020) during a period of flourishing serpulid reefs and high recruitment to settlement panels (Chapman et al., 2007). The lack of significant recovery was still evident by the persistence of these tracks on sidescan imagery in 2015. Also, reanalysis of data from the 2001 recruitment study of Chapman et al. (2007), where, inter alia, recruitment to occupied reef fragments was compared to dead fragments, reveals that dead fragments supported triple the level of recruitment density (T test, p=0.011). The dead fragments had been dried prior to deployment, thereby degrading any adhering epibiota including biofilms. The results from the current study also indicate that recruitment to previously dried reef material is at a similar level to that of newly deployed scallop shells.
Reductions in the availability of suitable colonisation surfaces by fouling will be offset in the case of occupied tube material by the development of new surfaces, generated largely by adult tube growth and the development of juvenile recruits. The extent of long-term reef development will be governed by the balance between colonisation of these surfaces by subsequent recruits and by fouling biota. Where serpulid settlement is erratic (with significant reduction or absence in some years) suitable colonisation surface are likely to be largely restricted to recent adult tube growth. The rate of supply of such surfaces will decrease over time through death and probably as a result of a reduction in age-related growth rate. Such reductions have been indicated for Serpula vermicularis (Hughes, Poloczanska and Dodd, 2008) and reported in several other serpulid species (Reidi and Smith, 2015).
Fouling will not only adversely affect reef development through substrate competition but suspension feeding species may reduce food availability for the worms and consume serpulid larvae. The presence of dense epibionts, particularly if flexible such as algae, will almost certainly compromise the feeding activity of the worms.

Figure 14. Examples of fouling of serpulid reefs by epibiota in Loch Creran in 2005.
Click for a full description
Photos of serpulid reef fouling in Loch Creran in 2005. A, young, globular reef with little visual evidence of significant fouling; B, base of mature reef with heavy ascidian fouling; C, distal region of mature reef with heavy fouling by Amphilectus fucorum.
The population dynamics of S. vermicularis reefs in Loch Creran have not been examined. However, examination of reef imagery is suggestive of populations skewed heavily in favour of certain size classes (viz. larger tubes). This is also seen with the recently discovered Loch Ailort serpulid reefs, which appear to be made up of tubes of the same age class (Moore, 2019). Erratic spatial and temporal recruitment by serpulid species is known to occur. For example Medernach et al. (2000) recorded a total absence of recruitment by Ditrupa arietina in some years and Cook (2016) reported virtually no recruitment of S. vermicularis at South Shian from April 2013 to September 2014 in an area where once luxuriant reefs have now undergone considerable fragmentation. Erratic recruitment is likely to exacerbate reef decline.
In addition to ageing of the reef promoting an upward development of fouling intensity from the older basal regions to the younger distal region, the suspension feeding activity of the branchial crown of the worm may exert a negative impact on larval recruitment to the cleaner distal region of the tube. In the serpulid, Ficopomatus enigmaticus, Thorp (1994) found the presence of contiguous branchial crowns largely prevented the settlement of larvae. Thus there may be a Goldilocks zone for larval settlement in the subdistal region of the reef, which narrows with time if reef growth does not keep pace with the extension of fouling. If recruitment is impacted this could also lead to a reduction in larval supply if the density of competent larvae is related to larval retention within the waters of the loch.
While the rate at which tube surface area is lost to potential serpulid recruitment through fouling might be expected to be accelerated by a reduction in larval supply and subsequent settlement, larval density reduction may not necessarily be a driver of juvenile habitat loss, but could be a consequence.
In summary it is proposed that a possible mechanism for reef decline is one of progressive fouling of reef material by such agents as macroepibiota, biofilms and silt, which accompanies physical fragmentation of the reef structure through bioerosion and the weight of the epibiotic load. The rate at which tube area is lost to serpulid recruitment will experience temporal variation through annual variations in larval supply and the recruitment success and growth of the fouling community. Serpulid recolonization of fragmented material may also be adversely affected by enhanced predation levels. Intact reefs represent complex, highly-rugose, three-dimensional structures that afford refuges from predation for species capable of living within the mesh of intertwining tubes, such as juvenile serpulids. Creran reefs have also been reported to house extremely high densities of the grazing predator, Psammechinus miliaris (viz. 577 m-2 - Moore et al., 2020). Reef fragmentation is likely to increase grazing pressure on serpulid recruits through the provision of predator access to enhanced levels of grazing substrate.
Such a mechanism is broadly consistent with the results of the remediation study, where the introduction of fresh substrate brought about the development of new reef material in locations exhibiting marked reef decline. The temporary nature of this reef development may have been related to growth of the fouling community, whose macrobiotic elements were clearly markedly enhanced on at least some of the remediation units. The same process could explain the recent decline of the reef habitat at other widespread locations including Loch Teacuis (Kamphausen, 2015), Blacksod Bay (Scally, Pfeiffer and Hewitt, 2020) and Killary Fjord (Seasearch Ireland, 2012, 2018). Examination of video footage of the reefs of Killary Fjord reveal a transition from a habitat consisting of dense, large reefs with little evident fouling in 2012 (Seasearch, Ireland, 2012), to a mixture of collapsed and erect reef material bearing a dense cloak of epibionts in 2018 (Seasearch, Ireland, 2018). Following a survey of Loch Teacuis, Kamphausen (2015) concluded that the reef decline observed may be related to fouling by Saccharina latissima.
The theory also provides an explanation for regional variation in the timing of reef decay in Loch Creran. During the 2017 - 2019 condition monitoring of Loch Creran (Moore et al., 2020), while reef material was highly degraded throughout much of the loch, within Rubha Riabhach Bay on the northern side of the loch the reefs were dense, largely unbroken, attaining heights of up to 65 cm and pristine reefs were widely recorded. The overall visual impression was that the reef habitat in the bay resembled that occupying large areas at the principal reef locations elsewhere in the loch in 2005. Recent examination of the bay reveals a radical change in condition, with widespread reef collapse resembling that currently occurring throughout most of the loch (Harries et al. 2025). The temporal difference may simply reflect variation in the timing of the fouling process.
5. References
Beaumont, J. 2006. Quantifying biotic interactions with inshore subtidal structures: comparisons between artificial and natural reefs PhD thesis, SAMS/UHI.
Bosence, D. 1979. The factors leading to aggregation and reef formation in Serpula vermicularis. In: G. Larwood and B.R. Rosen, ed. 1979. Biology and systematics of colonial organisms. London: Academic Press, pp. 299-318.
Chapman, N. 2004. The ecology and conservation of the rare Serpula vermicularis (L.) reefs, Loch Creran, Argyllshire, Scotland. PhD thesis, Heriot-Watt University, Edinburgh.
Chapman, N.D., Moore, C.G., Harries, D.B. and Lyndon, A.R. 2007. Recruitment patterns of Serpula vermicularis L. (Polychaeta, Serpulidae) in Loch Creran, Scotland Estuarine Coastal and Shelf Science, 73, 598-606.
Chapman, N.D., Moore, C.G., Harries, D.B. and Lyndon, A.R. 2011. The community associated with biogenic reefs formed by the polychaete, Serpula vermicularis Journal of the Marine Biological Association of the United Kingdom, 92, 679-685.
Cook, R.L. 2016. Development of techniques for the restoration of temperate biogenic reefs PhD thesis, Heriot-Watt University, Edinburgh.
Cook, R.L, Sanderson, W.G., Moore, C.G. and Harries, D.B. 2021. The right place at the right time: Improving the odds of biogenic reef restoration Marine Pollution Bulletin, vol. 164, article number 112022.
Cotter, E., O’Riordan, R.M. and Myers, A.A. 2003. Recruitment patterns of serpulids (Annelida: Polychaeta) in Bantry Bay, Ireland Journal of the Marine Biological Association of the United Kingdom, 83, 41-48.
Edwards, A. and D.J.Edelsten, D.J. 1977. Deep Water Renewal of Loch Etive: A Three Basin Scottish Fjord Estuarine and Coastal Marine Science, 5, 575-595.
Edwards, A., and Sharples, F. 1986. Scottish Sea Lochs: A Catalogue (SAMS Internal Reports; No. 134). Scottish Association for Marine Science.
Elkin, L.A., Kay, M., Higgins, J.J. and Wobbrock, J.O. 2021. An aligned rank transform procedure for multifactor contrast tests Proceedings of the ACM Symposium on User Interface Software and Technology (UIST '21). Virtual Event (October 10-14, 2021). New York: ACM Press, pp. 754-768.
Harries, D.B., Moore, C.G., Lyndon, A.R., Tulbure, K., Schuler, M., Wilkes. J. and Crisp, E. (Unpublished). An assessment of temporal change in the biological condition of serpulid reefs within Loch Creran between 2017 and 2024. Report by Heriot Watt University on behalf of NatureScot.
Holt, T.J., Rees, E.I., Hawkins, S.J. and Seed, R. 1998. Biogenic Reefs (volume IX). An overview of dynamic and sensitivity characteristics for conservation management of marine SACs Scottish Association for Marine Science (UK Marine SACs Project).
Huang, Y.L., Li, M., Yu, Z.L. and Qian, P.Y. 2011. Correlation between pigmentation and larval settlement deterrence by Pseudoalteromonas sp Biofouling, 27, 287-293.
Hughes, D J , Poloczanska, E.S. and Dodd, J. 2008. Survivorship and tube growth of reef-building Serpula vermicularis (Polychaeta: Serpulidae) in two Scottish sea lochs Aquatic Conservation: Marine and Freshwater Ecosystems, 18, 117-129.
Kamphausen, L. 2015. Loch Teacuis serpulid survey March 2015. Report to Scottish Natural Heritage.
Kupriyanova, E.K. 1999. The taxonomic status of Serpula CF. Columbiana Johnson, 1901 from the American and Asian coasts of the North Pacific Ocean (Polychaeta: Serpulidae) Ophelia, 50, 21-34.
Kupriyanova, E.K. , Nishi, E. , Ten Hove, H.A. and Rzhavsk, A.V. 2001. Life-history patterns in serpulimorph polychaetes: ecological and evolutionary perspectives Oceanography and Marine Biology: an Annual Review, 39, 1-101.
Leahy, Y. (1991). Polychaete studies on the west coast of Ireland. Ph.D. thesis, National University of Ireland.
McGibney, M. 2016. Serpula vermicularis Scholars’ Bank. University of Oregon.
Medernach, L., Jordana, E, Gremare, A., Nozais, C, Charles, F. and Amouroux, J. M. 2000. Population dynamics, secondary production and calcification in a Mediterranean population of Ditrupa arietina (Annelida: Polychaeta) Marine Ecology Progress Series, 199, 171-184.
MERC Consultants 2008. Surveys of sensitive sublittoral benthic communities in Mullet/Blacksod Bay Complex SAC, Rutland Island and Sound SAC, Mulroy Bay SAC Report to National Parks and Wildlife Service, Galway.
Moore, C.G. 2019. Biological analyses of underwater video from monitoring and research cruises in Lochs Ailort and Fyne, the Sounds of Barra and Mull, inner Moray Firth, off Wester Ross, Noss Head and Rattray Head, and around the Southern Trench in outer Moray Firth Scottish Natural Heritage Research Report No. 1085.
Moore, C.G., Bates, C.R., Mair, J.M., Saunders G.R., Harries D.B. and Lyndon A.R. 2009. Mapping serpulid worm reefs (Polychaeta: Serpulidae) for conservation management. Aquatic Conservation: Marine and Freshwater Ecosystems, 19, 226-236.
Moore, C.G., Harries, D.B., Tulbure, K.W., Cook, R.L., Saunders, G.R., Lyndon, A.R., Kamphausen, L., and James, B. 2020. The current status of serpulid reefs, horse mussel beds and flame shell beds in Loch Creran SAC and MPA Scottish Natural Heritage Research Report No. 1156.
Moore, C.G., Saunders, G.R. and Harries, D.B. 1998. The status and ecology of reefs of Serpula vermicularis L. (Polychaeta: Serpulidae) in Scotland Aquatic Conservation: Marine Freshwater Ecosystems, 8, 645-656.
Moore, C.G., Saunders, G.R., Harries, D.B., Mair, J.M., Bates, C.R. and Lyndon, A.R. 2006. The establishment of site condition monitoring of the subtidal reefs of Loch Creran Special Area of Conservation Scottish Natural Heritage Commissioned Report No. 151.
Ramos, A. and San Martín, G. 1999. On the finding of a mass occurrence of Serpula narconensis Baird, 1885 (Polychaeta, Serpulidae) in South Georgia (Antarctica) Polar Biology, 22, 379–383.
Riedi, M.A. and Smith, A.M. 2015. Tube growth and calcification of two reef-building ecosystem engineers in southern New Zealand: Galeolaria hystrix and Spirobranchus cariniferus (Polychaeta: Serpulidae). Marine Geology, 367, 212-219.
Scally, L., Pfeiffer, N. and Hewitt, E. 2020. The monitoring and assessment of six EU Habitats. Directive Annex I Marine Habitats Irish Wildlife Manuals, No. 118. National Parks and Wildlife Service, Department of Culture, Heritage and the Gaeltacht, Ireland.
Scottish Government 2020. Case study: Persistent damage to the Loch Creran serpulid reefs Scotland’s Marine Assessment 2020.
Seasearch, Ireland 2012. Serpula vermicularis reefs at Rossroe, Killary Fjord - September 2012
Seasearch, Ireland 2018. Serpula vermicularis reefs Rossroe, Killary Fjord – June 2018
Tett, P., Carreira, C., Mills, D. K., van Leeuwen, S., Foden, J., Bresnan, E. and Gowen, R. J. 2008. Use of a Phytoplankton Community Index to assess the health of coastal waters. ICES Journal of Marine Science, 65, 1475-1482.
Thorp, C.H. 1994 Population variation in Ficopomatus enigmaticus (Fauvel) (Polychaeta: Serpulidae) in a brackish water millpond at Emsworth, West Sussex UK. Memoires de Museum National d'Histoire Naturelle, 162: 585-591.
Whyte, C. T. 2012. An Investigation into changes in the phytoplankton community in Loch Creran, a Scottish sea loch PhD thesis, Napier University, Edinburgh.
Wobbrock, J.O., Findlater, L., Gergle, D. and Higgins, J.J. 2011. The aligned rank transform for nonparametric factorial analyses using only ANOVA procedures. Proceedings of the ACM Conference on Human Factors in Computing Systems (CHI '11). Vancouver, British Columbia (May 7-12, 2011). New York: ACM Press, pp. 143-146.
Young, C.M. and Chia, F-S. 1982. Ontogeny of phototaxis during larval development of the sedentary polychaete, Serpula vermicularis (L.) The Biological Bulletin, 162, 457-468.
Annexes
| Site name | Study | Latitude | Longitude |
|---|---|---|---|
| Rubha Dearg | recruitment | 56.528117 | -5.324900 |
| DR01 | recruitment | 56.545300 | -5.299550 |
| DR02 | recruitment | 56.522433 | -5.334350 |
| DR03 | recruitment | 56.520433 | -5.350800 |
| DR08 | recruitment | 56.523917 | -5.397783 |
| DR09 | recruitment | 56.525883 | -5.374267 |
| FLB | recruitment | 56.351763 | -5.502424 |
| FUB | recruitment | 56.361252 | -5.454682 |
| Main Site | reef development | 56.522850 | -5.333150 |
| Rubha Mòr | reef development | 56.518550 | -5.397200 |
| Mussel Farm | reef development | 56.532483 | -5.344233 |
| Kelco | reef development | 56.538067 | -5.305250 |
| Upper Basin | reef development | 56.547017 | -5.262450 |
| Predator exclusion | predator exclusion | 56.522225 | -5.333859 |
| Analysis | Method | Variable | Factor (levels) | Post hoc | Probability | |
|---|---|---|---|---|---|---|
| Main Site substrate x year | 2 way repeated measure ANOVA | reef cover | year (2015, 2017, 2018, 2022) | N/A | 0.480 | |
| - | - | - | substrate (CB, LSB) | N/A | 0.042 | |
| - | - | - | year*substrate | N/A | 0.368 | |
| Main Site substrate x year | 2 way repeated measure ANOVA | reef height | year (2015, 2017, 2018, 2022) | N/A | <0.001 | |
| - | - | - | substrate (CB, LSB) | N/A | 0.663 | |
| - | - | - | year*substrate | N/A | 0.028 | |
| - | - | - | year | 2015 v. 2017 | <0.001 | |
| - | - | - | - | 2015 v. 2018 | <0.001 | |
| - | - | - | - | 2015 v. 2022 | <0.001 | |
| - | - | - | - | 2017 v. 2018 | 0.350 | |
| - | - | - | - | 2017 v. 2022 | <0.001 | |
| - | - | - | - | 2018 v. 2022 | <0.001 | |
| Main Site substrate x year | Aligned Rank Transform | reef cover | year (2015, 2017, 2018, 2022) | N/A | 0.439 | |
| - | - | - | substrate (CB, LSB) | N/A | 0.028 | |
| - | - | - | year*substrate | N/A | 0.460 | |
| Main Site substrate x year | Aligned Rank Transform | reef cover | year (2015, 2017, 2018, 2022) | N/A | 0.039 | |
| - | - | - | substrate (CB, LSB, SSB, LS, B) | N/A | <0.001 | |
| - | - | - | year*substrate | N/A | 0.444 | |
| - | - | - | year | 2015 v. 2017 | 0.709 | |
| - | - | - | - | 2015 v. 2018 | 0.024 | |
| - | - | - | - | 2015 v. 2022 | 0.675 | |
| - | - | - | - | 2017 v. 2018 | 0.260 | |
| - | - | - | - | 2017 v. 2022 | 1.000 | |
| - | - | - | - | 2018 v. 2022 | 0.285 | |
| - | - | - | substrate | CB v. LSB | 0.469 | |
| - | - | - | - | CB v. SSB | <0.001 | |
| - | - | - | - | CB v. LS | <0.001 | |
| - | - | - | - | CB v. B | <0.001 | |
| - | - | - | - | LSB v. SSB | <0.001 | |
| - | - | - | - | LSB v.LS | <0.001 | |
| - | - | - | - | LSB v. B | <0.001 | |
| - | - | - | - | SSB v. LS | 0.681 | |
| - | - | - | - | SSB v. B | 0.887 | |
| - | - | - | - | LS v. B | 0.994 | |
| Main Site substrate x year | Aligned Rank Transform | reef height | year (2015, 2017, 2018, 2022) | N/A | <0.001 | |
| - | - | - | substrate (CB, LSB, SSB, LS, B) | N/A | <0.001 | |
| - | - | - | year*substrate | N/A | <0.001 | |
| - | - | - | year | 2015 v. 2017 | <0.001 | |
| - | - | - | - | 2015 v. 2018 | <0.001 | |
| - | - | - | - | 2015 v. 2022 | <0.001 | |
| - | - | - | - | 2017 v. 2018 | 0.096 | |
| - | - | - | - | 2017 v. 2022 | <0.001 | |
| - | - | - | - | 2018 v. 2022 | <0.001 | |
| - | - | - | substrate | CB v. LSB | 0.963 | |
| - | - | - | - | CB v. SSB | <0.001 | |
| - | - | - | - | CB v. LS | <0.001 | |
| - | - | - | - | CB v. B | <0.001 | |
| - | - | - | - | LSB v. SSB | <0.001 | |
| - | - | - | - | LSB v.LS | <0.001 | |
| - | - | - | - | LSB v. B | <0.001 | |
| - | - | - | - | SSB v. LS | 0.345 | |
| - | - | - | - | SSB v. B | 0.349 | |
| - | - | - | - | LS v. B | 1.000 | |
| Rubha Mòr year | Friedman | reef cover | year (2015, 2017, 2018, 2022) | N/A | 0.001 | |
| - | - | - | year | 2015 v. 2017 | 0.203 | |
| - | - | - | - | 2015 v. 2018 | 1.000 | |
| - | - | - | - | 2015 v. 2022 | 0.841 | |
| - | - | - | - | 2017 v. 2018 | 0.572 | |
| - | - | - | - | 2017 v. 2022 | 0.001 | |
| - | - | - | - | 2018 v. 2022 | 0.310 | |
| Rubha Mòr year | Friedman | reef height | year (2015, 2017, 2018, 2022) | N/A | 0.003 | |
| - | - | - | year | 2015 v. 2017 | 0.396 | |
| - | - | - | - | 2015 v. 2018 | 1.000 | |
| - | - | - | - | 2015 v. 2022 | 1.000 | |
| - | - | - | - | 2017 v. 2018 | 1.000 | |
| - | - | - | - | 2017 v. 2022 | 0.023 | |
| - | - | - | - | 2018 v. 2022 | 0.110 | |
| Rubha Mòr year | Friedman | algal cover | year (2015, 2017, 2018, 2022) | N/A | <0.001 | |
| - | - | - | year | 2015 v. 2017 | 0.863 | |
| - | - | - | - | 2015 v. 2018 | 0.138 | |
| - | - | - | - | 2015 v. 2022 | 0.009 | |
| - | - | - | - | 2017 v. 2018 | 1.000 | |
| - | - | - | - | 2017 v. 2022 | 0.212 | |
| - | - | - | - | 2018 v. 2022 | 1.000 | |
| Kelco year | Kruskall-Wallis | reef cover | year (2015, 2017, 2018, 2022) | N/A | 0.003 | |
| - | - | - | year | 2015 v. 2017 | 0.302 | |
| - | - | - | - | 2015 v. 2018 | 1.000 | |
| - | - | - | - | 2015 v. 2022 | 1.000 | |
| - | - | - | - | 2017 v. 2018 | 0.332 | |
| - | - | - | - | 2017 v. 2022 | 0.042 | |
| - | - | - | - | 2018 v. 2022 | 1.000 | |
| Kelco year | Kruskall-Wallis | reef height | year (2015, 2017, 2018, 2022) | N/A | 0.008 | |
| - | - | - | year | 2015 v. 2017 | 1.000 | |
| - | - | - | - | 2015 v. 2018 | 0.965 | |
| - | - | - | - | 2015 v. 2022 | 0.731 | |
| - | - | - | - | 2017 v. 2018 | 1.000 | |
| - | - | - | - | 2017 v. 2022 | 0.034 | |
| - | - | - | - | 2018 v. 2022 | 0.012 | |
| Kelco year | Kruskall-Wallis | algal cover | year (2015, 2017, 2018, 2022) | N/A | <0.001 | |
| - | - | - | year | 2015 v. 2017 | 0.055 | |
| - | - | - | - | 2015 v. 2018 | 0.817 | |
| - | - | - | - | 2015 v. 2022 | <0.001 | |
| - | - | - | - | 2017 v. 2018 | 1.000 | |
| - | - | - | - | 2017 v. 2022 | 0.875 | |
| - | - | - | - | 2018 v. 2022 | 0.052 | |
| Mussel farm year | Kruskall-Wallis | reef cover | year (2017, 2018, 2022) | N/A | 0.700 | |
| Mussel farm year | Kruskall-Wallis | reef height | year (2017, 2018, 2022) | N/A | 0.724 | |
| Mussel farm year | Kruskall-Wallis | algal cover | year (2017, 2018, 2022) | N/A | 0.003 | |
| - | - | - | year | 2017 v. 2018 | 1.000 | |
| - | - | - | - | 2017 v. 2022 | 0.012 | |
| - | - | - | - | 2018 v. 2022 | 0.006 | |
| Upper basin year | Kruskall-Wallis | reef cover | year (2017, 2018, 2022) | N/A | 0.943 | |
| Upper basin year | Kruskall-Wallis | reef height | year (2017, 2018, 2022) | N/A | 0.358 | |
| Upper basin year | Kruskall-Wallis | algal cover | year (2017, 2018, 2022) | N/A | 0.240 | |
| All locations location x year | Aligned Rank Transform | reef cover | location (5 locations) | N/A | 0.001 | |
| - | - | - | year (2017, 2018, 2022) | N/A | <0.001 | |
| - | - | - | location*year | N/A | <0.001 | |
| - | - | - | location | Main v. RMòr | 0.026 | |
| - | - | - | - | Main v. Kelco | <0.001 | |
| - | - | - | - | Main v. Mussel | <0.001 | |
| - | - | - | - | Main v. Upper | <0.001 | |
| - | - | - | - | RMòr v. Kelco | 0.004 | |
| - | - | - | - | RMòr v. Mussel | <0.001 | |
| - | - | - | - | RMòr v. Upper | 0.020 | |
| - | - | - | - | Kelco v. Mussel | 0.189 | |
| - | - | - | - | Kelco v. Upper | 0.997 | |
| - | - | - | - | Mussel v. Upper | 0.114 | |
| - | - | - | year | 2017 v. 2018 | 0.173 | |
| - | - | - | - | 2017 v. 2022 | <0.001 | |
| - | - | - | - | 2018 v. 2022 | <0.001 | |
| Substrate | Kruskal-Wallis | Serpula density | substrates (scallop, reef, tube) | N/A | 0.087 | |
| Substrate | 1 way ANOVA | Spirobranchus density | substrates (scallop, reef, tube) | N/A | 0.005 | |
| - | - | - | substrate | scallop v. tube | 0.242 | |
| - | - | - | - | scallop v. reef | 0.074 | |
| - | - | - | - | tube v. reef | 0.004 | |
| Substrate | Kruskal-Wallis | Hydroides density | substrates (scallop, reef, tube) | N/A | 0.733 | |
| Year | T test | Serpula density | year (2001, 2023) | N/A | <0.001 | |
| Year | T test | Spirobranchus density | year (2001, 2023) | N/A | <0.001 | |
| Year | T test | Hydroides density | year (2001, 2023) | N/A | <0.001 | |
| Location | Kruskal-Wallis | Serpula density | location (RD, DR01, DR02, DR03, DR08, DR09, FLB, FUB) | N/A | 0.164 | |
| Location | Kruskal-Wallis | Spirobranchus density | location (RD, DR01, DR02, DR03, DR08, DR09, FLB, FUB) | N/A | 0.062 | |
| Location | Kruskal-Wallis | Hydroides density | location (RD, DR01, DR02, DR03, DR08, DR09, FLB, FUB) | N/A | 0.009 | |
| - | - | - | location | DR01 v. DR02 | 1.000 | |
| - | - | - | - | DR01 v. DR03 | 1.000 | |
| - | - | - | - | DR01 v. DR08 | 1.000 | |
| - | - | - | - | DR01 v. DR09 | 1.000 | |
| - | - | - | - | DR01 v. FLB | 1.000 | |
| - | - | - | - | DR01 v. FUB | 1.000 | |
| - | - | - | - | DR01 v. RD | 1.000 | |
| - | - | - | - | DR02 v. DR03 | 1.000 | |
| - | - | - | - | DR02 v. DR08 | 1.000 | |
| - | - | - | - | DR02 v. DR09 | 1.000 | |
| - | - | - | - | DR02 v. FLB | 0.591 | |
| - | - | - | - | DR02 v. FUB | 1.000 | |
| - | - | - | - | DR02 v. RD | 1.000 | |
| - | - | - | - | DR03 v. DR08 | 1.000 | |
| - | - | - | - | DR03 v. DR09 | 1.000 | |
| - | - | - | - | DR03 v. FLB | 1.000 | |
| - | - | - | - | DR03 v. FUB | 0.056 | |
| - | - | - | - | DR03 v. RD | 1.000 | |
| - | - | - | - | DR08 v. DR09 | 1.000 | |
| - | - | - | - | DR08 v. FLB | 1.000 | |
| - | - | - | - | DR08 v. FUB | 1.000 | |
| - | - | - | - | DR08 v. RD | 1.000 | |
| - | - | - | - | DR09 v. FLB | 1.000 | |
| - | - | - | - | DR09 v. FUB | 0.074 | |
| - | - | - | - | DR09 v. RD | 1.000 | |
| - | - | - | - | FLB v. FUB | 0.013 | |
| - | - | - | - | FLB v. RD | 1.000 | |
| - | - | - | - | FUB v. RD | 0.294 | |
| Exclusion cage treatment | Kruskal-Wallis | substrate loss - serpulid substrate | cage treatment (open control, cage control, cage) | N/A | 0.095 | |
| Exclusion cage treatment | Kruskal-Wallis | tube height - scallop substrate | cage treatment (open control, cage control, cage) | N/A | 0.035 | |
| - | - | - | - | open control v. cage control | 1.000 | |
| - | - | - | - | open control v. cage | 0.219 | |
| - | - | - | - | cage control v. cage | 0.036 | |
| Exclusion cage treatment | T-test | tube height - | cage treatment (serpulid and scallop cage) | N/A | 0.026 | |
| Exclusion cage treatment | Kruskal-Wallis | no. tubes - scallop substrate | cage treatment (open control, cage control, cage) | N/A | 0.662 | |
| Exclusion cage treatment | T-test | no. tubes | cage treatment (serpulid and scallop cage) | N/A | 0.372 | |





