
NatureScot Research Report 1145 - Distribution mapping and baseline monitoring survey of sea caves around Fair Isle
Fair Isle harbour photo © Graham Saunders
This research trip was undertaken 12 – 28 August 2022, with 18-24 August (inclusively) around Fair Isle.
Year of publication: 2024
Authors: Harries, D.B., Purcell, A., Moore, C.G., Lyndon A.R., Porter, J.S., Saunders, G.R. and Hume, C.L. (Heriot-Watt University)
Cite as: Harries, D.B., Purcell, A., Moore, C.G., Lyndon A.R., Porter, J.S., Saunders, G.R. and Hume, C.L. 2024. Distribution mapping and baseline monitoring survey of sea caves around Fair Isle. NatureScot Research Report 1145.
Keywords
Annex I habitat; benthos; condition; Demonstration and Research MPA; Fair Isle; Habitats Directive; marine; Marine Protected Area (MPA); MPA; monitoring; sea caves
Background
This research trip was undertaken 12 – 28 August 2022, with 18-24 August (inclusively) around Fair Isle. The aim of the current study was to create an inventory of the location, extent and nature of sea caves around Fair Isle. Additionally, the study set out to gather detailed biological data from a series of selected caves. The purpose of this was to gather information relevant to the distribution and quality of the Annex I habitat ‘submerged or partially submerged sea caves’ (as defined by the Habitats Directive) around Scotland. This information is also relevant to Fair Isle’s designation as a Demonstration and Research MPA.
Due to the extreme wave exposure of Fair Isle, it was necessary to consider a series of contingency survey targets in the event of weather conditions preventing access to the primary target. Weather conditions severely limited time available for surveying the primary target so some attention was diverted to the contingency targets of seagrass beds, flame shell beds and horse mussel beds around Orkney (not included in this report).
Main findings
A total of 175 potential cave sites (including arches and tunnels) have been identified on Fair Isle based on a combination of historical records and direct observation. All available information on caves has been collated into a cave inventory.
- It was estimated that approximately 60 of these sites are likely to have sufficiently reduced light levels to qualify as the Annex I habitat ‘submerged or partially submerged sea caves’.
- Some caves were extensive (~200 m in one case) and passages tended to be aligned with the rock bedding of the island.
- Four caves were subject to a detailed biological assessment. Three were on the east coast and one on the west coast.
- Cave biota tended to be heavily modified by the effects of scour which was concentrated at or near the floor of the caves. In shallower caves the effect of the scour tended to extend the entire wall height. At more extreme levels the biota was very impoverished with sparse spirorbins and mobile biota. At less extreme levels there was a community characterised by a turf of silty tubes constructed by sabellid polychaetes and / or tube dwelling amphipods.
- Where scour levels were reduced (deeper cave floor) a community characterised by Dendrodoa grossularia and Clathrina coriacea occurred in a number of caves.
- The upper walls of caves with a deep cave floor can support a more diverse and variable community characterised by crisid bryozoan turfs, aggregations of anemones, Tubularia indivisa and Alcyonium digitatum.
- Littoral and supralittoral zones appeared impoverished (relative to the open coast) within the caves.
- The biota recorded was broadly consistent with prior data recorded from Fair Isle caves and broadly similar to that recorded from comparable cave sites elsewhere in Scotland.
- No evidence of anthropogenic impacts or of anthropogenic activities with potential to impact the cave features was observed.
- Based on available data it should be concluded that the cave features were in good condition.
Acknowledgements
We would like to express our gratitude to Andy Cuthbertson and the crew of the Jean Elaine for ensuring we made the best use of the limited opportunities offered by the weather and for cheerfully accommodating changes of plan when required. We would also like to thank Martha Thomson, Neil Thomson & Stewart Thomson for their advice, hospitality and sharing their local knowledge of the Fair Isle sea caves. Thank you to Kieran Tulbure (HWU) for his efforts in photographically documenting the survey sites. Thank you to Owen Paisley for assisting with biological surveys and sample collection. We are also very grateful to Dr Andy Farrant (BGS) and Dr Iain Greig (University of Aberdeen) for advice and guidance on geological matters.
1. Introduction
The aim of this study was to create an inventory of the location, extent and nature of sea caves around Fair Isle. Additionally, the study set out to gather detailed biological data from a series of selected caves. The purpose of this was to gather information relevant to the designation of protected sites based on distribution and quality of the Annex I habitat ‘submerged or partially submerged sea caves’ (as defined by the Habitats Directive) around Scotland. This information is also relevant to Fair Isle’s designation as a Demonstration and Research MPA.
Due to the extreme wave exposure of Fair Isle, it was necessary to consider a series of contingency survey targets in the event of weather conditions preventing access to the primary target. Weather conditions severely limited time available for surveying the primary target so some attention was diverted to the contingency targets of seagrass beds south of Westray (which helped to create the Tuquoy bay polygon on the NMPi Seagrass beds datalayer), and flame shell bed and horse mussel bed Priority Marine Features (PMFs) in Scapa Flow, Orkney (videos to be analysed in 2024).
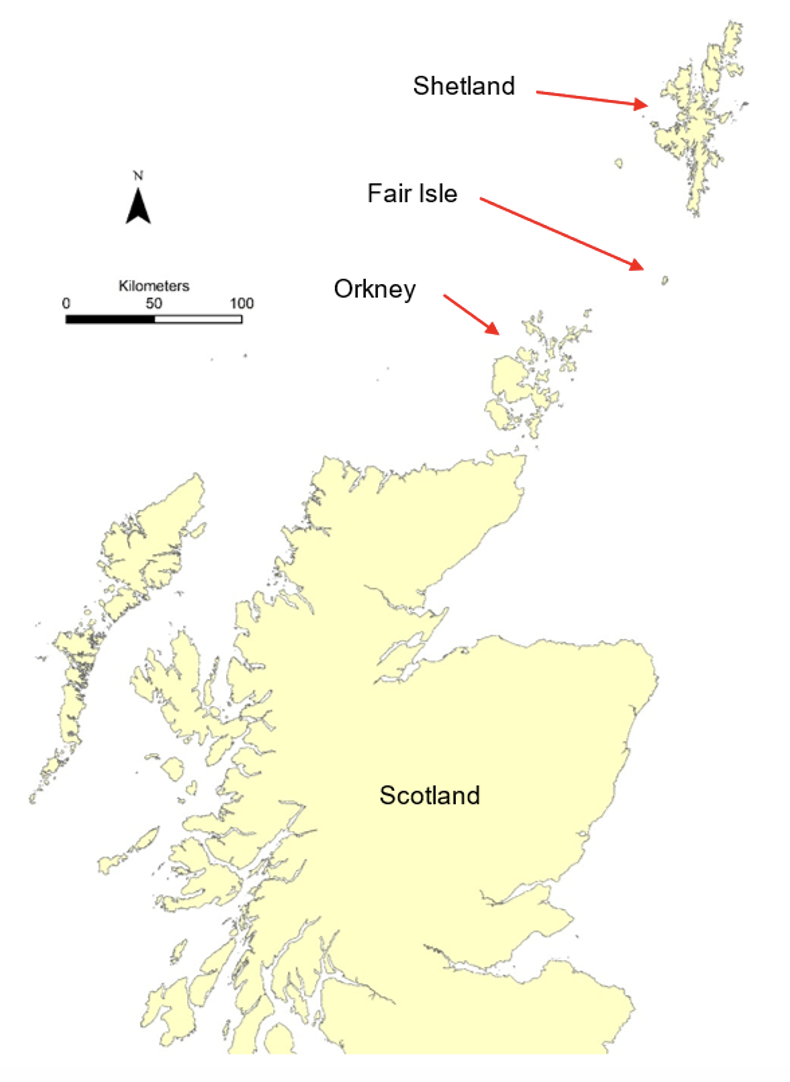
Figure 1. Map indicating location of Fair Isle. © Crown copyright and database rights [2024] Ordnance Survey 100017908.
1.1 The physical environment
Fair Isle is located about 40 km south of Shetland and about 50 km north-east of Orkney (Figure 1). The island is just under 5 km from north to south and roughly 2 km from west to east (Figure 2). It has a population of about 60 people. The main area of habitation and cultivation lies in the relatively low-lying southern part of the island. The northern part of the island consists of higher elevation moorland with Ward Hill (217 m) as the highest point.
The majority of the coastline consists of steep rocky cliffs. These are of significant height at many locations around the island and often exceed 100 m on the western and northern coasts. The only reliable harbour in regular use is North Haven on the eastern side of the island. Although the climate is relatively mild and maritime, the coasts are very exposed to wave action. Due to the small size of the island there is little protection from ocean swell and some degree of wave surge is to be expected on the shore for the majority of the time.
The rocks of Fair Isle are predominantly sedimentary and are Devonian in age. Most of the beds are formed from pebbly sandstones but there are beds of finer grained argillaceous sediments interspersed with the sandstone layers. In general terms the bedding tends to be steeply inclined and slopes downwards in an approximately south easterly direction at an angle of 55˚ to 75˚ from the horizontal. Broadly speaking, the alignment of the bedding tends to run along a south west to north east axis. The bedding is cut across by a number of faults which are mostly aligned west north west to east south east (Mykura 1972). See section 4.2 for further explanation.
1.2 Previous marine biological studies on Fair Isle
Marine Recorder contains a number of entries for Fair Isle and some of these relate to sea caves. The majority of the records derive from work conducted in 1987 as part of the “1986-87 OPRU/MNCR Shetland, Foula and Fair Isle survey”. Other records derive from Seasearch surveys in 2011, 2012 and 2016.
The 1987 records cover a total of 22 locations around Fair Isle. Of these, 9 relate to subtidal rocky habitats, 4 relate to subtidal rock with areas of coarse sediment, 2 relate to subtidal sediment, 5 relate to intertidal rock and 2 relate to sea caves. The 2011 Seasearch records cover 5 locations. Of these, 3 relate to subtidal rocky habitats and 2 relate to sea caves. The 2012 Seasearch record is of a single subtidal location with no useful biological data. The 2016 Seasearch records cover 2 locations. One of these relates to subtidal rocky habitats and the other to a sea cave.
It should be noted that a significant proportion (more than half) of the Fair Isle coordinates cited in Marine Recorder are clearly erroneous. This is apparent because coordinates placed a significant distance (50-100 m) offshore in deep water are often linked to biological records taken from rock surfaces at or just below the waterline. It is probable that the true location of these sites were considerably closer to the shore than is indicated by the coordinates. Other evidence of errors include a subtidal site which is plotted on land and a rocky intertidal site which is plotted on a sedimentary shore. GPS was not in routine public use in 1987 so it is likely these coordinates were determined by reference to coastal maps. However, errors also occur in the Seasearch records which were generated after 2010 presumably by means of the use of a GPS unit. On balance, all these Marine Recorder coordinates should be regarded as indicative only and it should be understood that they cannot be used for precise site relocation.
Figure 2. Map showing Fair Isle. © Crown copyright and database rights [2024] Ordnance Survey 100017908.
The intertidal sites covered in Marine Recorder were all rocky and typically tended to support biota indicative of exposed locations. Shores ranged from ‘exposed’ or ‘moderately exposed’ sites with communities characterised by Semibalanus balanoides to ‘very exposed’ shores supporting Chthamalus sp. and with Fucus distichus and Fucus spiralis f. nana present. However, there was considerable variation with some topographically complex sites supporting Ascophyllum nodosum or other fucoids in localised pockets of shelter.
The sublittoral fringe tended to be characterised by Alaria esculenta which corresponds to the general picture of a high degree of wave exposure. Forests of Laminaria hyperborea tended to predominate in the infralittoral. There was some variability in these forests with some supporting profuse growth of foliose red algae whereas others were subject to significant grazing by Echinus esculentus. It is probable that these differences arise from subtle differences in levels of wave exposure. The urchins are less able to sustain high population densities at more exposed locations and such sites are likely to sustain a more abundant growth of foliose red algae. The differences might be relatively localised with slightly steeper or shallower rock supporting fewer urchins and adjacent areas with a less steep profile or greater water depth supporting higher urchin densities.
In addition to urchin grazing, the effects of scour on infralittoral communities appeared significant at some locations. This is apparent in the occurrence of forests of mixed kelp species. This occurs where scour events clear rock surfaces periodically, favouring the establishment of faster growing species such as Saccorhiza polyschides, Saccharina latissima and Desmarestia sp. At one location, a ‘bed’ of Flustra foliacea was noted on the deeper rock. This species is also indicative of scour and supports the conclusion that scour is a significant factor in structuring many subtidal rock communities at Fair Isle. Scour levels are likely to be more intense on rock surfaces adjacent to coarse mobile sediments. So at some locations it is likely that Laminaria hyperborea forests might occur on the upper levels of rock slopes but mixed kelps or impoverished communities may occur at lower levels where they are subject to increased scour from neighbouring areas of coarse sediments.
Some reports also note more intense scour impacts on the floors and lower walls of subtidal gullies. This pattern is also characteristic of many sea cave habitats and will be discussed later. Vertical subtidal rock was noted at a number of sites and dense growth of Alcyonium digitatum was notable at many of these. Relatively few of the Marine Recorder records focussed on subtidal sediments. In most cases, the sediments were reported as coarse mobile sands and gravels. But in more sheltered embayments (e.g. North Haven) there were finer sands characterised by Arenicola marina casts.
Biological records relating to the 5 ‘cave’ sites included in Marine Recorder will be reviewed in the ‘Discussion’ section where they can be placed in context with the findings of this survey.
1.3 Conservation designations
Fair Isle has long standing recognition in terms of its importance for bird conservation. The island is important as a breeding site for a range of seabird species and also as a stop-over point for migratory species. The Fair Isle Bird Observatory has been established since 1948 and is an internationally renowned ornithological institution. The island is designated as a Site of Special Scientific Interest (SSSI) and a Special Protection Area (SPA) for a range of named seabird species. It is designated as a Special Area of Conservation (SAC) for the terrestrial habitats of ‘European dry heaths’ and ‘Vegetated sea cliffs of the Atlantic and Baltic coasts’. It is also designated as a Demonstration and Research Marine Protected Area (D&R MPA). This extends the potential for nature conservation into the subtidal zone and is intended to give the local community opportunities to develop new or improved forms of marine management to protect seabirds and promote ecotourism.
Numerous sea caves are known to occur around the Fair Isle coast and ‘submerged or partially submerged sea caves’ are listed under Annex I (Habitats) of the Habitats Directive. This study and findings will provide additional information for reporting and assessment of this Annex I feature at a Scottish and UK level. This is required due to amendments of the Habitats Regulations following the UK leaving the European Union. The findings are also of relevance to the site designation as a D&R MPA and to the local community more generally.
1.4 Human impacts
Fair Isle has been inhabited since at least the Neolithic. Population levels have probably been low for most of the time. Peak levels are thought to have occurred in the nineteenth century (with a population of over 300) and have subsequently declined to the current level of about 60 people (Butler 2016). The majority of human activity has doubtless revolved around some combination of crofting and fishing since the island was first inhabited, but tourism has also become increasingly important in recent times.
In the past, certain cave sites were used for the disposal of waste (including old vehicles). The gloup (geographic feature caused by the collapsed roof of a sea cave) at South Reeva is a notable example. This waste appears to have been largely removed and the practice has been discontinued. It is unlikely that present human activity has any significant potential to impact directly on the biota of the sea caves. Access to the caves is often difficult and dangerous and the caves harbour no resources likely to attract visits. The effects of any disturbance from human visits would be likely to be transitory in any event. These are extremely high energy environments subject to regular disturbance from storms and associated scour. Any disturbance arising from human visits is likely to be negligible in relation the intense and frequent disturbances that occur naturally due to waves driven by high winds.
Indirect impacts are potentially possible. Such impacts could occur due to climatic changes, ecological shifts due to fishing activities in the neighbouring waters or through pollution incidents such as oil spills. There are no reasons to suppose that Fair Isle is any more vulnerable to such events than is the case for other Scottish islands. Also, there are no reasons to suppose that the biota of sea caves would be any more susceptible to the impacts than the biota of the open coast. If such impacts were to materialise, they could well apply to sea caves but their broader effects on the local marine environment would be of greater concern.
1.5 Condition monitoring of sea caves
Although sea caves are not designated interest features on Fair Isle it is recognised that some do qualify as Annex I habitat. Therefore, it is appropriate that biological surveys are conducted in a manner to allow the potential for future monitoring and reporting of this feature, as required at a Scottish and UK level. Site Condition Monitoring (SCM) is undertaken to determine the biological condition of the interest feature (submerged or partially submerged sea caves) and to guide feature management action where appropriate.
The purpose of the current study was to assess the extent of the Annex I habitat of ‘submerged or partially submerged sea caves’ around Fair Isle and to establish a baseline biological data set that will facilitate the assessment of the favourable condition status of the habitat in the future.
To fulfil these objectives sea cave distribution was examined, and for selected caves, the topography was mapped and the biotopes surveyed by MNCR phase 2 methodology at relocatable points along the cave system.
Common Standards Monitoring (CSM) guidance has been drawn up in order to ensure a uniform approach to the monitoring of the condition of features (Anon, 1998). Thus, for the purposes of monitoring, each feature is represented by a series of attributes, which are measurable indicators of the condition of the feature at the site. For each attribute (e.g. extent of a habitat or presence of representative / notable biotopes), a target is set which is considered to correspond to the favourable condition of the feature.
The Common Standards Monitoring guidance produced for sea caves (JNCC 2004) lists attributes of the habitat together with the corresponding targets that should form the basis of the site condition monitoring (Table 1).
Table 1. Site attributes that should be utilised to define the condition of sea cave features in site condition monitoring (JNCC 2004). All attributes are discretionary.
Attribute | Target |
|---|---|
Extent of cave(s) | No change in dimensions of a cave, allowing for natural change that is part of a wider coastal geomorphological management regime. |
Number of caves in site | No reduction in the number of caves within a site allowing for natural change. |
Biotope composition of a cave | Maintain the variety of biotopes identified for the cave, allowing for natural succession or known cyclical change. |
Presence of representative/ notable biotopes | Maintain the presence of the specified biotope, allowing for natural succession/ known cyclical change. |
Species composition of representative or notable biotopes | No decline in biotope quality due to change in species composition or loss of notable species, allowing for natural succession/ known cyclical change. |
Presence and/or abundance of specified species | Maintain presence and/or abundance of the specified desirable species. Absence of the specified undesirable species (such as an invasive non-native species). |
In this project, the attribute ‘Extent of cave(s)’ is addressed by the topographic surveys conducted within selected caves. The attribute ‘Number of caves in site’ is addressed by the cave inventory. Remaining attributes are supported by the biological data collected within each of the surveyed caves.
It is necessary to consider what features characterise a ‘cave’. Although the term ‘cave’ is generally understood the distinction between rock arches, alcoves, rock overhangs and caves can be blurred. The Interpretation Manual of European Union Habitats (European Commission, 2013) does not attempt to define the term ‘cave’. It provides a definition of a sea cave that could be paraphrased as a cave that is wholly or partially within the sea (Other rocky habitats - 8330 - ‘Caves situated under the sea or opened to it, at least at high tide, including partially submerged sea caves. Their bottom and sides harbour communities of marine invertebrates and algae.’). Common standards monitoring guidance refers to a working definition of a cave proposed by Bunker and Holt (2003) who state that “a cave must be large enough to get a surveyor fully into the cave, turn round and exit without damaging the attached flora and fauna”. However, this definition is sufficiently broad to encompass large rock alcoves and overhangs where environmental conditions and biotic communities are likely to be indistinguishable from those of the open coast. The only biologically relevant parameter that consistently distinguishes caves from similar features is light. Surge gullies, rock arches, alcoves and rock overhangs are usually relatively well illuminated, whereas caves are typically dark. Chapman (1993) has proposed a useful biological definition of caves as “perpetually-dark voids, bounded by rock or similar inorganic materials and filled with gas and/or water”. Accordingly, we regard ‘caves’ which are fully illuminated due to large entrances to be a poor example of the feature and have attempted to concentrate efforts on sites where there is a significant reduction of light intensity within the cave.
2. Methods
2.1 Cave inventory
Prior to the survey, existing records of cave sites were gathered. These were derived from local knowledge imparted to NatureScot staff by Fair Isle residents, from ordnance survey maps and from the Grampian Speleological Group database. An additional major source of records was derived from a photographic survey of cave locations conducted in 2019 by Carol Hume (NatureScot) and Fair Isle residents. The Marine Recorder database was also scrutinised for cave records.
During the 2021 survey, the coast was systematically examined at close quarters from an inflatable boat whenever sea conditions were suitable. To maximise coverage in the limited time available, two inflatables operated independently with a team of 3 or 4 surveyors in each. Cave entrances were photographed and the coordinates recorded by GPS. If sea conditions were safe, snorkellers entered the caves and made notes on the configuration, extent, water depth and light levels. Wherever possible, a sketch was made of the cave layout.
2.2 Biological surveys
Detailed biological and physical surveys were conducted at four cave sites. The choice of available sites was severely constrained by weather conditions. Preferred cave sites were those that were extensive and had significant water depth. The essential criterion for site selection was that the cave should be sufficiently dark to require the use of a torch. Although the selected sites met this essential criterion they were not all ideal in terms of depth and extent. Although some potentially more suitable sites were identified they could not be safely accessed.
Monitoring surveys of caves were initiated by conducting a physical survey and deploying a relocatable fixed datum line for precise spatial mapping of biotopes. Methods are adapted from Ellis (1988) and have formerly been used in Berwickshire (ERT, 2004), on Papa Stour (ERT, 2005), on Mousa (Harries et al., 2009, Harries et al., 2019) and on St Kilda, North Rona and Loch Eriboll (Harries et al., 2018). The physical survey allows cave extent and depth range to be assessed. It also provides a means of defining the location of points within the cave from which biological data was recorded and enables those points to be relocated by subsequent monitoring surveys.
A point on the upper shore/supralittoral just outside the cave entrance was marked with a galvanized metal piton (‘relocation piton’) hammered into a rock crevice. The position of the marker was fixed by GPS and several photographs taken of it from different viewpoints. The proximity of sea cliffs can reduce GPS accuracy, so several waypoints were taken and subsequently plotted and cross referenced with photographs to ensure coordinates were reliable. The piton provided the relocation point for a fixed line that was set up within the cave to act as a datum line to define the position of points within the cave. The line was set up to run in straight line sections with additional attachment points positioned between each section as required to prevent the line being forced into a curve by obstructions or changes in the direction of the cave passage. The compass bearing and length of each section of line was recorded and the height/depth of the individual attachment points was noted at the beginning and end of the section. An approximate plan view sketch was produced of the cave floor with notes on the composition of the substrate. At selected points along the line the cross sectional shape of the passage was sketched and the cross sectional dimensions (e.g. distance to floor, ceiling and both walls) were directly measured at floor level and estimated in relation to the fixed datum line for the upper parts of the cross section. The arrangement of the datum line is typically a near vertical descent from the relocation piton to a shotline located below the water surface. The line then continues along a single wall when possible, following the main axis of the cave. Attachment points were photographed or videoed to aid future surveyors in repositioning the datum line.
Following the physical survey, an assessment was made of changes in biological communities along the length of the cave and a series of locations were identified for subsequent collection of biological data from cross sections of the cave passage. The number of biological cross section surveys were determined by the time available and the extent of the cave. They varied from one to three on the caves examined. The biological surveys were conducted within a 2 m wide band up one wall of the passage at specified distances along the datum line. Where appropriate, the wall was subdivided into biological zones and the depth of the zone boundaries recorded. Within each biological zone, the abundance of each component species was estimated using the MNCR SACFOR scale and notes made on the nature and inclination of the substrate. Where appropriate, samples were collected to establish the identity of difficult, cryptic or morphologically variable biota.
Video footage was obtained along the length of the cave using the datum line for orientation and detailed footage was obtained of the biota at each of the biological cross sectional survey locations. Additional underwater wide angle and macro digital still images were taken to illustrate the biota at the biological cross sectional survey locations.
Where the terms ‘left’ and ‘right’ are used in the context of cave descriptions the orientation is as viewed into the cave.
All coordinates were generated by differential GPS receivers (Garmin Montana 600) set to the WGS84 datum. Depths and heights were related to chart datum by determination of the tidal rise using TotalTide software (Hydrographic Office, Taunton). The secondary port of ‘Fair Isle’ was used. In the text of this report, the heights / depths of locations are cited as metres ‘ACD’ (above chart datum) or metres ‘BCD’ (below chart datum) as appropriate.
Surface stills photographs were taken using different cameras depending on the circumstances and the personnel collecting the records (Olympus TG-5, Olympus TG-6, Nikon Coolpix S32 & Panasonic DMC-FZ62). Wide angle underwater stills photographs were taken with a Nikon D200 and 14 mm lens. Macro underwater stills were taken with a Nikon D200 and 60 mm lens with additional macro images taken using an Olympus TG-5. Underwater video footage was taken with a GoPro Hero 4 attached to frame bearing video lights.
2.3 Data processing
2.3.1 Cave inventory
Historical records from cave sites were plotted using ArcGIS 10.6.1 and cross-referenced with information noted in 2021. Erroneous coordinates and duplicate records of the same cave were identified and corrected where possible by scrutiny of descriptive accounts and published maps of cave locations. All accessible records were collated to produce a detailed geo-referenced cave inventory. Caves were categorised based on the degree of certainty that a cave is present and based on the availability of information on extent and light levels.
2.3.2 Physical data
An orthographic projection-based approach was taken to process the physical survey data and Adobe Illustrator was used to generate the final cave surveys. Initially, once the physical survey data was collected an initial centreline was constructed in plan view - each leg was individually generated to the correct length (scale used 1 cm : 1 m) and then rotated to the correct bearing as described in the survey data. Each leg was then joined to form a single line and the final centreline realised. Width components (in this case circles of the desired diameter as per the scale 1 cm : 1 m) were then added to both sides of the centreline which facilitated the realisation of the left and right hand walls as seen in plan view. Wall and floor information was then added - with line detailing being derived from both the original sketches and photographs gathered on the date of the respective survey.
2.3.3 Voucher specimens
Voucher specimens were preserved in ethanol. Approximately 150 voucher specimens were collected for validation of in-situ records. The collection includes ~27 sponge specimens. Spicules were extracted using bleach and slides examined under a compound microscope and eyepiece graticule measurements made of a range of spicules where appropriate. Identifications utilised Ackers et al. (2007). Cnidarians were represented by ~10 specimens with hydroids composing a significant component of the biota at some sites. Identifications were made using Schuchert (2012) for athecate hydroids and Cornelius (1995) for thecate hydroids. All specimens were examined using a dissection microscope and in the case of certain hydroids temporary slides prepared for examination with compound microscope. Within the polychaetes (~33 specimens), small sabellids and spirorbins were a prominent component at many sites. These were examined using a combination of dissection and compound microscopy and identified using Hayward & Ryland (1990). Within the Crustacea (~40 specimens), tube dwelling amphipods were a significant component of the biota at certain sites. Identifications were made using a dissection microscope and literature including Lincoln (1979) for the amphipods. Molluscs were represented by ~14 specimens identified using Tebble (1976) and Hayward & Ryland (1990). Bryozoans were represented by ~10 specimens. Crisid turf was a significant component of the cave biota at one site. Crisids were identified using Hayward & Ryland (1990). Other bryozoan taxa were identified using Ryland & Hayward (1977), Hayward & Ryland (1979) and Hayward (1985). Echinoderms were represented by 4 specimens and were identified with Southward & Campbell (2006). Ascidians were represented by ~10 specimens identification literature was primarily Millar (1970). Taxon names were synonymised to correspond to WoRMS (2022).
2.3.4 Biological data
Wherever possible, in-situ species identifications were validated (and corrected where appropriate) by detailed cross-referencing with the identifications of the voucher specimens and with macro photographs. In-situ abundance records were also validated (and adjusted where appropriate) by detailed cross-referencing with video footage and stills images. Abundance scales were assigned using the MNCR SACFOR scale. Similarly, site descriptions were cross-referenced with video and stills imagery for consistency and validation. Biotopes were assigned based on the classification scheme of Connor et al. (2004).
2.3.5 Site coding
Surveyed sites have been allocated a formal site code and a site name relating to neighbouring geographic features (where possible). The format of the site code includes an initial pair of characters to indicate location (i.e. ‘FI’ to denote Fair Isle), a second pair of characters denotes the year of survey (i.e. ‘21’ denotes 2021) and a third pair of characters denotes the feature surveyed (i.e. ‘CV’ denotes cave). The final pair of characters identifies the specific site such that ‘FI21CV02’ indicates the second cave site surveyed at Fair Isle in 2021.
Individual zones and cross sections are identified by numerals following the site code. For example, ‘FI21CV01.3.2’ denotes the second zone (surveying vertically from cave floor to ceiling) on the third cross section (surveying further into the cave from the entrance) of the first cave surveyed at Fair Isle.
During the survey fieldwork, cave sites were allocated a ‘cave inventory code’ (CI1, CI2 etc) when initially examined. Those selected for survey were referred to by a ‘field reference’ name or code which was used on the written labels of the voucher collection. The ‘field reference’ and ‘cave inventory code’ are cited in the Results section and in appropriate appendices to ensure specimens and field records can be linked to the relevant survey location.
3. Results
3.1 Introduction
The intention of the 2021 survey team was to conduct a systematic search of the entire Fair Isle coastline and document all cave sites present. However, during the 7-day visit to Fair Isle a combination of wave surge and sea fog limited the area accessible for a systematic search to the area around North Haven and two southward facing stretches of the east coast (Figure 3).
The selection of caves for detailed biological survey was similarly constrained to sites that could be safely accessed in the prevailing conditions. Nevertheless, the cave sites were sufficiently different in their structure to represent a range of different cave habitats. The cave floor of FI21CV03 lay in the lower intertidal and sublittoral fringe so this was essentially an intertidal site. The floors of the remaining sites were subtidal but of differing depths with FI21CV01 at 2 m BCD, FI21CV02 at 4 m BCD and FI21CV04 at 7.5 m BCD (at the entrances). The range of water depth is significant because it relates to the level of scour and hence the diversity and composition of the biota. The locations of the four surveyed sites are indicated on Figure 3.
3.2 Cave inventory
A total of 175 cave or cave-like sites have been documented around the coast of Fair Isle (see Figures 4 & 5). This figure includes arches and tunnels in addition to closed caves. This was done because differentiating between an ‘arch’ and a ‘tunnel’ is subjective and because ‘tunnels’ can produce the same environmental conditions as ‘caves’ and hence contain typical cave communities. Of the 175 sites, we have a high level of confidence that an entrance is present in 145 cases. This confidence is variously based on having visited the sites, access to photographs of cave entrances taken from cliff tops or from local knowledge provided by the Fair Isle inhabitants. The remaining 30 sites are noted from maps or other sources, but their presence is not confirmed. The cave inventory contains more than 175 records. This is because two entrances are recorded for some tunnels but only one has been considered for these estimates to avoid ‘double counting’ of sites.
Of the 145 confirmed sites there are 82 where there is sufficient information to form a judgement on whether they qualify as an Annex I ‘submerged or partially submerged sea caves’ feature. This judgement is based on light levels within the cave. Reduced light is the primary environmental factor distinguishing caves from sites with similar topography such as overhangs, rock alcoves and surge gullies. Direct measurement of light levels would have been impractical within the time constraints, so a pragmatic approach was taken based on noting if a torch was required. Of these 82 sites, there were 20 where light levels were judged to be sufficiently reduced to qualify as a cave feature (represented in Figures 4 & 5 and in Annex I). The remaining 62 sites were noted as partially or fully illuminated.
A large proportion of the recorded caves were of limited extent. For example, 65% of the recorded caves were less than 20 m in length. Nevertheless, there remains a large number of extensive caves including a number of over 50 m in length and one tunnel that exceeds 200 m. The topography of the caves reflects the local geology, and the general pattern is of cave passages orientated along a SW to NE axis and with a rift-shaped cross section inclined towards the NW.
Cave locations are shown in figures 4 & 5. The full detailed list of cave inventory records is presented in Annex 1.
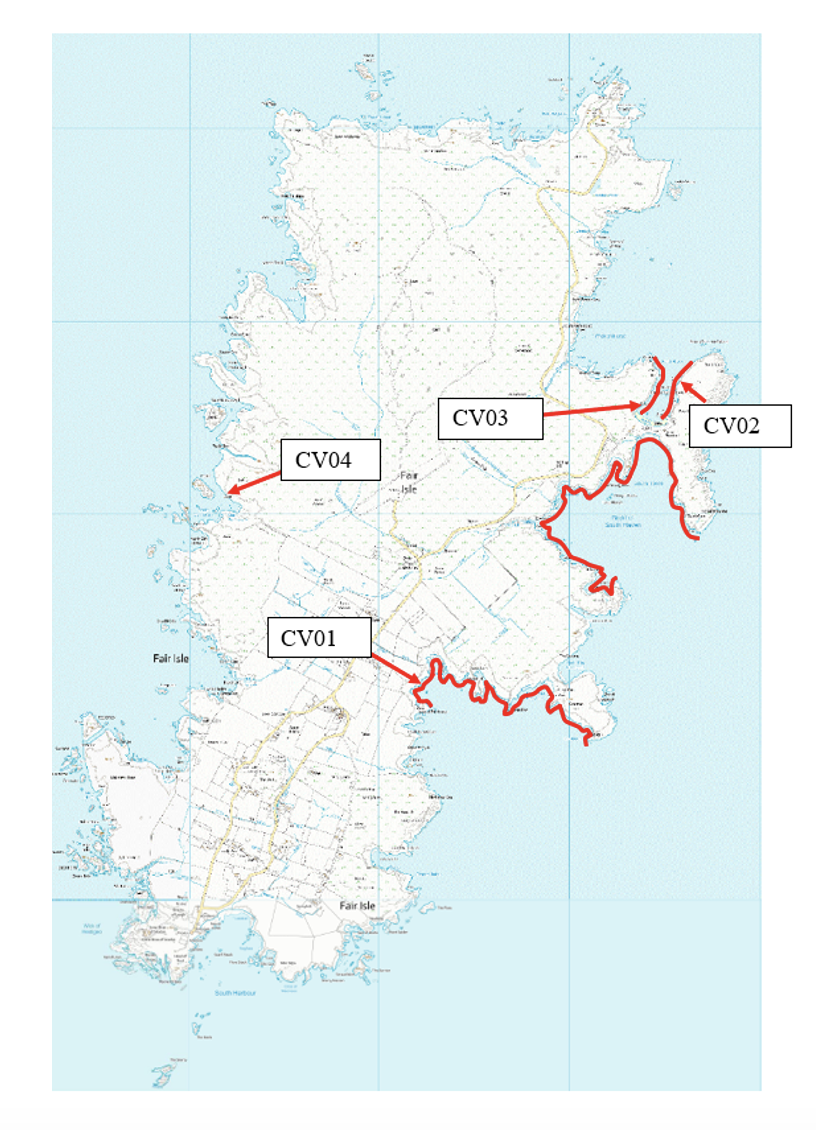
Figure 3. Map of Fair Isle showing areas of coast where a systematic search was made for caves in 2021 (red lines). The positions of the four biological survey sites are also indicated © Crown copyright and database rights [2024] Ordnance Survey 100017908
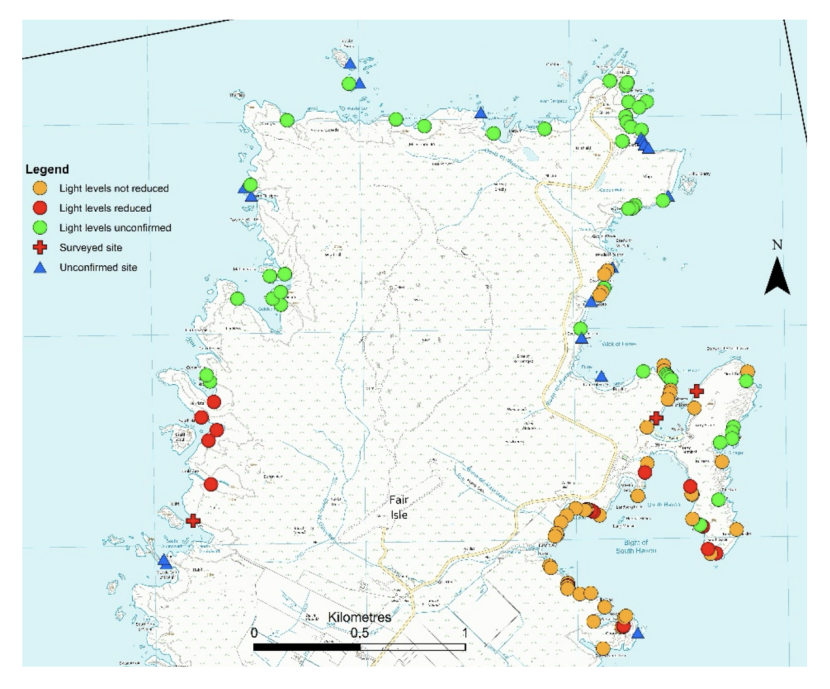
Figure 4. Northern part of Fair Isle showing location of cave sites. © Crown copyright and database rights [2024] Ordnance Survey 100017908
Click for a full description
Symbols indicate level of confidence in evidence of cave presence and availability of information on light levels. Reduced light levels are indicated by red symbols (both circles and crosses). Higher resolution maps are presented in Annex 1B.
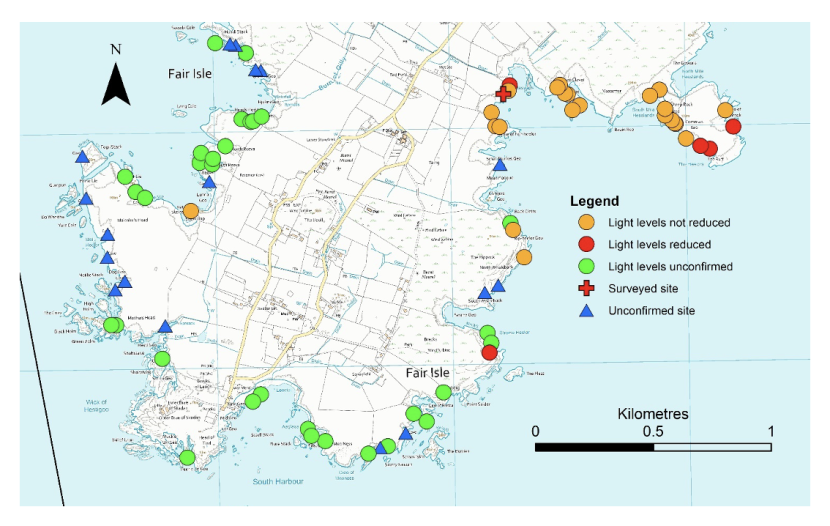
Figure 5. Southern part of Fair Isle showing location of cave sites. © Crown copyright and database rights [2024] Ordnance Survey 100017908
Click for a full description
Symbols indicate level of confidence in evidence of cave presence and availability of information on light levels. Reduced light levels are indicated by red symbols (both circles and crosses). Higher resolution maps are presented in Annex 1B.
3.3 FI21CV01 – Heeds o Sheeldegeo (Hesswells) cave 3
(field ref. ‘A3’; cave inv. Code ‘CI45’; – see section 2.3.5 for explanation of site coding)
Physical Description (Figure 6)
Cave entrance was ~3.5 m in width at waterline but rapidly narrowed above to <1 m in width with a ceiling at ~7 m ACD. A main passage trended NW and extended for ~47 m from the entrance. About 17 m within the entrance a side passage branched off the main passage on the right and ran north for a further ~39 m. Just within the entrance the main passage abruptly narrowed to ~1 m in width. But it rapidly broadened once more beyond this point reaching ~4 m in width near the junction. However, the majority of the inner part of the main passage was considerably narrower with the last ~25 m of this passage being less than ~2 m in width. The northern side passage was rather broader, exceeding ~4 m in width for much of its length. The passage cross section was predominantly ‘teardrop shaped’ throughout. It was broader at the base due to erosive scouring by the floor boulders and narrowed above into constricted inclined rift-like fissures in the ceiling. Cave floor at the entrance was at ~2 m BCD and progressively shallowed within the cave with the last ~3 m of the main passage occupied by an intertidal (>1 m ACD) boulder beach. A corresponding boulder beach was also present in the last ~6 m of the northern side passage. Cave floor was predominantly composed of rounded boulder slabs throughout. The northern side passage appeared relatively silty with sand infill between the floor boulders. The rear of the main passage appeared more scoured with polished floor boulders and considerable quantities of drift algae.
Biological description (Tables 2, 3 & 4)
The subtidal zone of the entrance area was a dense forest of Laminaria digitata (superabundant). There was an understory of foliose red algae (common) including Odontothalia dentata. Rock surfaces were well covered by coralline crusts (abundant) and patchy Halichondria panicea (occasional). Echinus esculentus (present).
The kelp forest and algal cover did not extend far within the entrance. By 5 m within the cave the kelp was absent and foliose red algae was relatively sparse (occasional). There was also less cover of coralline crusts, but they remained abundant. On larger stable boulders there was increased coverage of Fabricia stellaris tube turf (locally common) and Spirorbis tridentatus (locally occasional). Similar biota occurred on the cave walls with increased coverage of sponge crusts (locally common on upper walls) with increasing height on the walls.
By 10 m within the cave there were no foliose reds or coralline crusts evident. From this point onward the biota remained similar for the rest of the cave. The rounded boulders and cobbles of the cave floor showed evidence of scour and supported a relatively impoverished community. This was characterised by sparse Fabricia stellaris tube turf (rare to frequent) and Spirorbis tridentatus tubes (rare). The abundance of the Fabricia stellaris was variable with denser populations present on larger more stable boulders. In some areas there were significant numbers of Actinia equina amongst the boulders. The level of scour increased as the floor became shallower within the cave. In the northern side passage the rock surfaces of the floor were relatively bare 30 m into the cave. Beyond 40 m into the cave, they were scoured almost entirely bare, with a clean pebble beach at 45 m into the cave. A similar pattern could be seen in the main passage with rock surfaces almost entirely bare at the inner end.
The biota at the base of the cave walls tended to be similar to that of the floor but became increasingly abundant with height above the floor due to reduced scour. Cave wall biota was characterised by Fabricia stellaris tube turf which was generally superabundant on the upper part of the cave walls. Grantia compressa was also a major component of the biota and was generally common or abundant on the mid and upper walls. Spirorbis tridentatus and sponge crusts were present throughout but were generally very sparse (rare). The sponge crusts (predominantly Leuconia sp.) were more prominent (frequent) on the upper walls in the outer part of the cave. This cave wall community was present throughout the cave but became more impoverished in the inner parts of the cave where scour from the shallow cave floor extended over more of the wall area.
The area near the waterline (~1.0 – 2.0 m ACD) at the entrance was characterised by sparse Semibalanus balanoides (rare) and Patella vulgata (common). The upper walls and ceiling were characterised by Hildenbrandia rubra and Verrucaria sp. (both common). Due to time constraints, the area above the waterline further within the cave was not closely assessed but appeared to be predominantly bare rock.
In terms of biotopes, the subtidal zone of the entrance area corresponded to the environmental conditions and biota described for IR.FIR.SG.FoSwCC. The intertidal part of the entrance walls corresponded to an impoverished version of LR.HLR.MusB.Sem. Upper walls and ceiling corresponded to LR.FLR.CvOv.VmucHil. Two main biotopes characterised the inner part of the cave beyond about 10 m from the entrance. The floor and lower walls supported a sparse scoured biota corresponding to IR.FIR.SG.CC.Mo. This extended over a greater proportion of the walls further into the caves where the floor was shallower. Where there was sufficient water depth, the upper parts of the cave walls supported a somewhat richer and more abundant biota indicative of a reduction in the level of scour. This was ascribed to IR.FIR.SG.CCBalPom. The match to the characterising species is poor but the situation and environmental conditions correspond well.
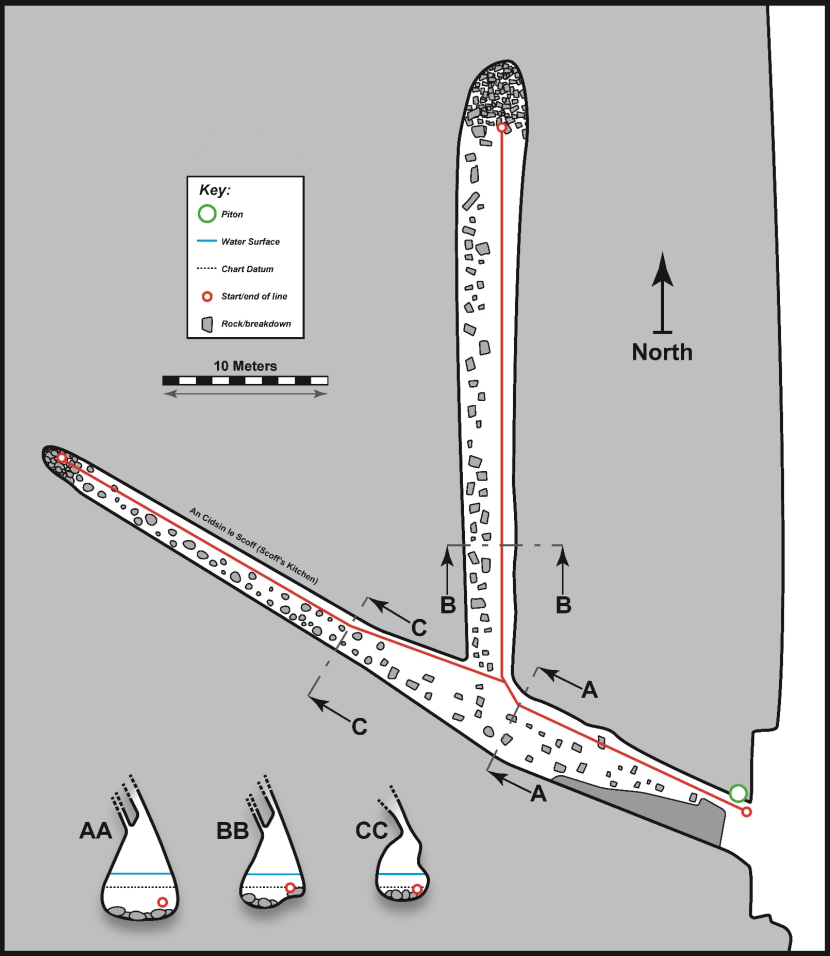
Figure 6. FI21CV01 – Heeds o Sheeldegeo (Hesswells) cave 3 (‘A3’; ‘CI45’) .
Click for a full description
Physical survey (scale bar applies to both floor plan and passage cross sections, associated biological data is provided in tables 2, 3 & 4). ‘AA’ is cross section 1, ‘BB’ is cross section 2 and ‘CC’ is cross section 3.
Table 2. FI21CV01 – Heeds o Sheeldegeo (Hesswells) cave 3 (‘A3’; ‘CI45’) – Biological summary – cross section 1 (right wall, ~15 m from entrance, 15 m on main tape)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor of rounded boulders and cobbles. | -1.9 | -2.2 | Sparse biota with occasional Fabricia stellaris tube turf, rare Spirorbis tridentatus and sponge crusts. | IR.FIR.SG.CC.Mo |
2 | Vertical to slightly overhanging cave wall. | +1.2 | -2.0 | Biota sparse near base of wall, becoming increasingly abundant with height above floor. Fabricia stellaris tube turf common becoming abundant on upper wall. Grantia compressa common on upper wall. Leuconia(?) crusts occasional becoming frequent on upper wall. Spirorbis tridentatus occasional. | IR.FIR.SG.CC.BalPom |
Table 3. FI21CV01 – Heeds o Sheeldegeo (Hesswells) cave 3 (‘A3’; ‘CI45’) – Biological summary – cross section 2 (left wall, ~25 m from entrance, 25 m on main tape – northern side passage)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor of rounded boulders and cobbles and base of rock wall to ~0.5 m above floor. | -0.7 | -1.2 | Sparse biota. Fabricia stellaris tube turf frequent, Spirorbis tridentatus rare and Actinia equina frequent amongst boulders. | IR.FIR.SG.CC.Mo |
2 | Vertical to slightly overhanging cave wall. | +1.2 | -0.7 | Fabricia stellaris tube turf abundant becoming superabundant on upper wall. Grantia common, Halichondria and Spirorbis tridentatus rare. | IR.FIR.SG.CC.BalPom |
Table 4. FI21CV01 – Heeds o Sheeldegeo (Hesswells) cave 3 (‘A3’; ‘CI45’) – Biological summary – cross section 3 (left wall, ~27 m from entrance, 10 m on left branch tape – main passage)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor of rounded boulders and cobbles. Drift algae present... | -0.7 | -0.7 | Sparse biota, rock surfaces appear smooth & bare. Fabricia stellaris tube turf and Spirorbis tridentatus rare. Actinia equina locally abundant between boulders. | IR.FIR.SG.CC.Mo |
2 | Vertical to slightly overhanging cave wall. | 0 | -0.7 | Scoured lower wall. Fabricia stellaris tube turf frequent but restricted to crevices, Spirorbis tridentatus rare. | IR.FIR.SG.CC.Mo |
3 | Vertical to slightly overhanging cave wall. | +1.2 | 0 | Fabricia stellaris tube turf superabundant, Grantia abundant, Leucosolenia sp.and Spirorbis tridentatus rare. | IR.FIR.SG.CC.BalPom |
3.4 FI21CV02 – Buness, Nort gabel
(field ref. & cave inv. Code ‘CI04’)
Physical Description (Figure 7)
Cave entrance was ~2.5 m in width at waterline with a ceiling at ~5 m ACD. The cave consisted of a single linear passage trending NE and extending for ~82 m from the entrance. Passage dimensions remained similar for most of the length of the cave. Passage shape was in the form of a rift inclined to the NW. Passage width was broader at the cave floor due to erosive scouring by the floor boulders. Cave floor at the entrance was at ~4 m BCD and progressively shallowed within the cave to a small intertidal (>1.2 m ACD) boulder beach at the back. Cave floor was predominantly composed of rounded boulders throughout. Because of excessive wave surge and reduced visibility on the day of the survey, only the first 30 m of the cave was subject to a detailed physical survey. But snorkellers had accessed the back of the cave and measured overall length during a visit on an earlier date.
Biological description (Table 5)
The subtidal zone of the entrance area was a forest of Laminaria digitata (abundant). There was an understory of foliose red algae (common) including Odontothalia dentata. Coralline crusts (frequent) and patchy Halichondria panicea (rare) occurred on rock surfaces. The kelp forest and foliose algal cover did not extend far within the entrance. By 5 m within the cave both were absent. Patches of coralline crusts continued within the cave as far as 10 m but were absent by 15 m.
Most rock surfaces within the cave were dominated by tube turf. Collected samples indicated the composition of the tube turf was mixed and variable. Samples from the floor and lower walls contained large numbers of Jassa falcata and very low numbers of Fabricia stellaris. Samples from higher on the walls contained large numbers of Jassa falcata, Fabricia stellaris and cf. Pseudoparatanais batei. On the upper cave walls and the larger floor boulders the tube turf was superabundant but was less dense (common or abundant) in less stable floor areas which appeared to be subject to some scour. Dendrodoa grossularia were also notable. They were common on most surfaces and abundant locally. Clathrina coriacea was present but rather sparse (rare). The abundances of both Dendrodoa grossularia and Clathrina coriacea were insufficient to justify the designation of the IR.FIR.SG.DenCcor biotope but it is probable that the biotope would occur if scour levels were lower. Sponges were prominent. Grantia compressa were abundant on the upper parts of the walls and sponge crusts (predominantly Halichondria panicea) were frequent in the same areas. Spirorbis tridentatus were present but very sparse (rare) throughout although more abundant (occasional) near the base of the walls.
Sea conditions prevented biological surveys beyond 20 m from the entrance. However, when the cave was examined by snorkellers on an earlier date it was noted that the community throughout the cave was dominated by tube turf with abundant Grantia compressa on the upper parts of the walls. It is thought that the community described above is representative of that found throughout the cave although it is likely to be impoverished towards the back of the cave as scour increases due to the shallower cave floor.
In terms of biotopes, the subtidal zone of the entrance area corresponded to the environmental conditions and biota described for IR.FIR.SG.FoSwCC. Within the cave, the floor and subtidal walls were scoured but not to the extent of IR.FIR.SG.CC.Mo. These areas were ascribed to IR.FIR.SG.CCBalPom. The match to the characterising species is poor but the situation and environmental conditions correspond well. The area could be regarded as a very impoverished version of IR.FIR.SG.DenCcor but overly scoured for the appropriate designation of this biotope. The biota of the lower part of the intertidal walls was similar to that of the subtidal area. This was ascribed to LR.FLR.CvOv.SpR.Den as the closest potential match of the intertidal cave biotopes. With increasing height on the wall, the biota became more impoverished and did not clearly match any biotope so was ascribed to the biotope complex LR.FLR.CvOv. Above this area was a band of sparse barnacles which represented an impoverished version of LR.HLR.MusB.Sem. Upper walls and ceiling corresponded to LR.FLR.CvOv.VmucHil. Both of these two biotopes are likely to diminish further into the caves but both were recognisable at 20 m from the entrance.
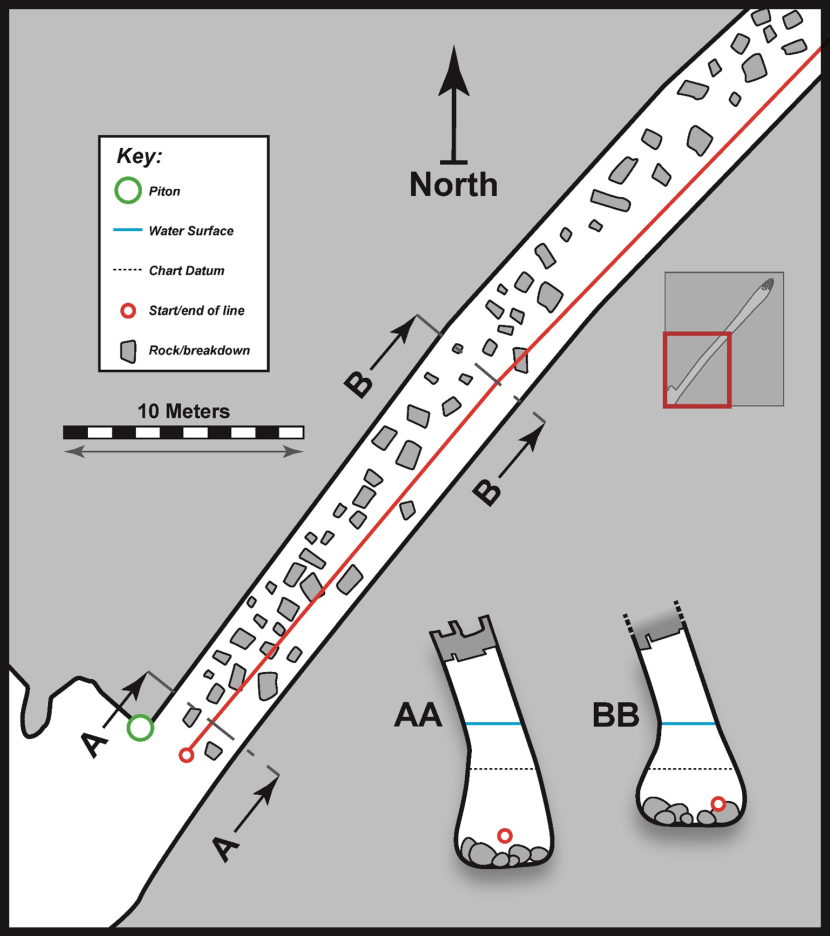
Figure 7. FI21CV02 – Buness, Nort gabel (‘CI04’).
Click for a full description
Physical survey (scale bar applies to both floor plan and passage cross sections, associated biological data is provided in table 5). ‘BB’ is cross section 1. Insert shows whole cave based on snorkeller records.
Table 5. FI21CV02 – Buness, Nort gabel (‘CI04’) – Biological summary – cross section 1 (right wall, ~20 m from entrance, 20 m on tape)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor of rounded boulders and cobbles. Including some very large stable boulders. | -1.9 | -2.4 | Tube turf (predominantly Jassa falcata based on samples) abundant and locally superabundant on larger boulders. Dendrodoa grossularia frequent and locally abundant on larger boulders. Sparse Spirorbis tridentatus (rare) and Halisarca dujardinii (rare). | IR.FIR.SG.CC.BalPom |
2 | Vertical & slightly overhanging rock walls. | -0.4 | -2.4 | Tube turf (predominantly Jassa falcata based on samples) abundant. Dendrodoa grossularia common (locally abundant in some areas). Sparse Spirorbis tridentatus (rare) but locally occasional on lower wall. Halichondria (rare). | IR.FIR.SG.CC.BalPom |
3 | Vertical & slightly overhanging rock walls. | +1.1 | -0.4 | Tube turf (mixed composition based on samples) superabundant. Grantia compressa abundant, particularly near the top of the zone. Dendrodoa grossularia common. Halichondria rare overall but frequent in upper part of zone. Sparse Spirorbis tridentatus (rare). | LR.FLR.CvOv.SpR.Den |
4 | Vertical & slightly overhanging rock walls. | +1.6 | +1.1 | Tube turf (mixed composition assumed) tube turf superabundant. | LR.FLR.CvOv |
5 | Vertical & slightly overhanging rock walls. | +2.1 | +1.6 | Sparse Semibalanus (rare). | LR.HLR.MusB.Sem |
6 | Vertical & slightly overhanging rock walls. | >+2.1 | +2.1 | Algal stains. Hildenbrandia (common) and green algal stains (common). | LR.FLR.CvOv.VmucHil |
3.5 FI21CV03 – North Haven west cave 3
(field ref. & cave inv. Code ‘CI01’)
Physical Description (Figure 8)
Cave entrance was ~3 m in width at the waterline with a ceiling at ~5 m ACD. A main passage trended W and extended for ~22 m from the entrance. Just within the entrance a side passage branched off the main passage on the right and ran northwest for a further ~16 m. About 6 m from the entrance, the main passage abruptly narrowed to ~2 m in width. Beyond this point it broadened once more to ~5 m in width. The northwest side passage was ~3.5 m in width for much of its length. The passage cross section was somewhat variable with sloping slabs forming the ceiling and stepped blocky walls. Cave floor at the entrance was at ~0 m BCD and progressively shallowed within the cave to a small intertidal (>1 m ACD) cobble beach at the back of the main passage. A corresponding intertidal (>1 m ACD) area lay at the back of the northwest side passage. The cave floor at the entrance consisted of boulder slabs. Within the cave, the floor consisted of bedrock slabs in some areas and boulders with pebbles in others.
Biological description (Table 6)
The entrance area had a dense forest of Laminaria digitata (superabundant) with coralline crusts, (abundant), foliose red algae (common) and Patella vulgata (frequent). This extended for ~5 m into the cave along the main passage. From 5 to 8.5 m along the main passage there was abundant coralline crusts, common foliose red algae and patches of Halichondria panicea (occasional). From 8.5 to 12 m along the main passage the floor was composed of scoured bedrock with sparse biota of coralline crusts (frequent), Halichondria panicea (occasional), Actinia equina, Carcinus maenas and Caryophyllia smithii (all rare). The cave walls in this same area were dominated by common Fabricia stellaris tube turf and frequent Leuconia johnstoni. Biota otherwise sparse including Grantia compressa, Spirorbis tridentatus and Halichondria panicea (all rare). From 12 m to the back of the cave there was very little biota and the substrate consisted of clean scoured cobbles and boulders.
The Laminaria digitata forest extended only for the first 3 m into the northwest side passage with coralline crusts and foliose red algae extending a further 1.5 m into the cave. From 4.5 to 11 m along this passage the biota was characterised by Fabricia stellaris tube turf which was common on the floor and becoming abundant on parts of the walls. Samples of tube turf indicated it was of mixed composition but composed primarily of Fabricia stellaris but with a significant proportion of Amphicorina sp. Sponge crusts were also notable with Halichondria panicea more prominent on the floor (frequent) and Leuconia johnstoni more prominent on the walls (frequent). Spirorbis tridentatus were sparse (rare) throughout. From 11 m to the back of the passage the substrate consisted of clean scoured bedrock and boulders.
In terms of biotopes, the subtidal zone of the entrance area corresponded to the environmental conditions and biota described for IR.FIR.SG.FoSwCC. Within the cave, the floor and cave walls supported a community that was not a clear match to any intertidal cave biotope. It was not severely scoured and was tentatively ascribed to LR.FLR.CvOv.FaCr. This biotope is somewhat of a ‘catch-all category and is noted to be variable in composition.
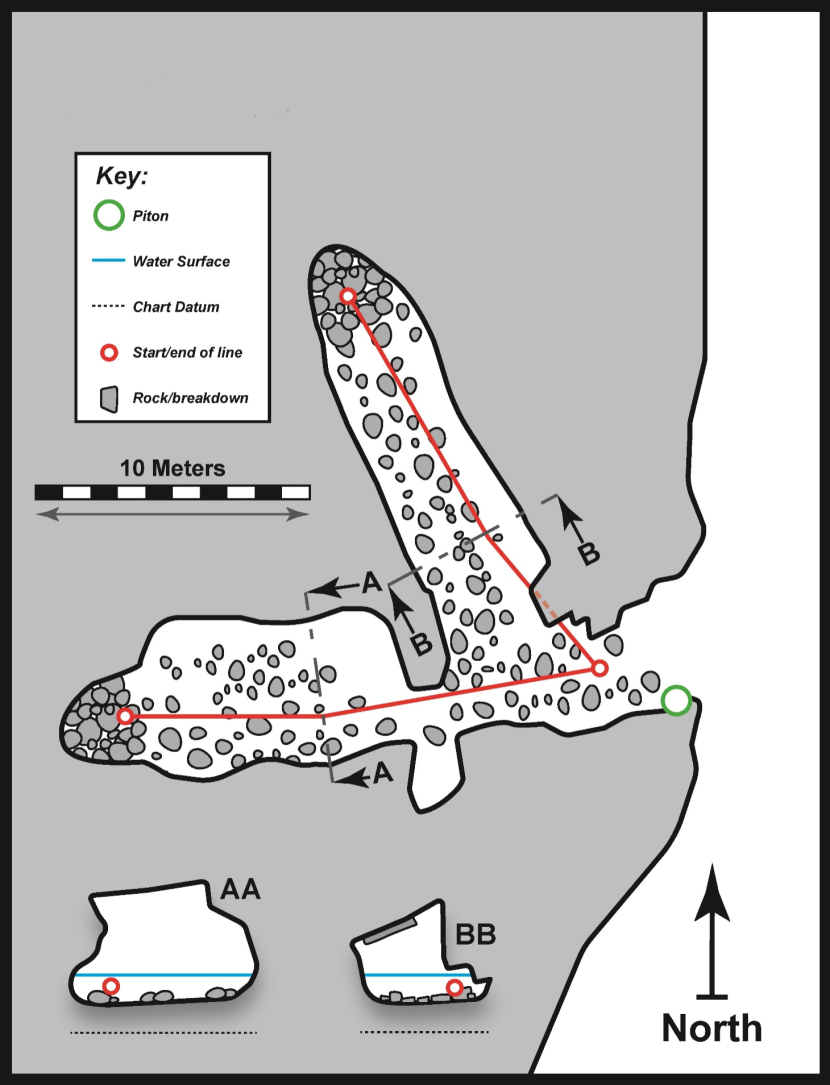
Figure 8. FI21CV03 – North Haven west cave 3 (‘CI01’).
Click for a full description
Physical survey (scale bar applies to both floor plan and passage cross sections, associated biological data is provided in table 6). ‘BB’ is cross section 1.
Table 6. FI21CV03 – North Haven west cave 3 (‘CI01’) – Biological summary – cross section 1 (right wall, ~7 m from entrance, 6 m on right hand tape)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor of boulders & cobbles. | +1.0 | +1.0 | Sparse Fabricia stellaris tube turf (common) and patches of Halichondria panicea (frequent). | LR.FLR.CvOv |
2 | Vertical rock wall. | +2.0 | +1.0 | Locally dense Fabricia stellaris tube turf (abundant) and patches of Leuconia johnstoni (frequent) in lower part of the zone. Actinia equina locally common in upper part of the zone. | LR.FLR.CvOv |
3 | Vertical rock wall & ceiling. | >+2.0 | +2.0 | Sparse patches of cave mould (rare). | LR.FLR.CvOv |
3.6 FI21CV04 – The Diamond cave (Colsta) – South entrance
(field ref. & cave inv. Code ‘CI66’)
Physical Description (Figure 9)
Southern cave entrance was ~14 m in width at the waterline with a ceiling at ~9 m ACD. The cave consisted of a single linear passage trending NE and extending for ~200 m from the southern entrance and connecting into Troila Geo at a northern entrance. Passage dimensions remained considerable throughout the cave but became narrower towards the north. Width was reduced to ~6.5 m at 85 m from the southern entrance and narrowed further towards the northern entrance. Passage shape was in the form of a rift inclined to the NW. Cave floor at the southern entrance was at ~7.5 m BCD and progressively shallowed within the cave. Floor depth at 85 m from the southern entrance was ~5 m BCD and the floor of the northern entrance was an intertidal (~0.7 m ACD) cobble beach. The cave floor was predominantly composed of boulders with cobbles, pebbles, and sand infill. Some extensive areas of rippled sand were present deeper within the cave. For example, the majority of the cave floor was composed of rippled sand from about 75 to 90 m from the southern entrance. Time constraints prevented the completion of a full physical survey but snorkellers swam through to inspect the northern entrance.
Biological description (Tables 7 & 8)
The cave floor boulders at the entrance were dominated by a forest of Laminaria hyperborea (abundant) with coralline crusts (abundant), foliose reds (frequent) and Echinus esculentus (common). The algal dominance diminished with increasing distance into the cave. Laminaria hyperborea was reduced to common by 5 m and absent by 10 m. At 10 m the foliose reds were reduced to rare and the coralline crusts were reduced to occasional by 20 m. Foliose reds were absent beyond this point but sparse patches of coralline crusts (rare) continued to ~30m.
The cave floor biota was generally sparse. Beyond 25 m it was characterised by sparse Jassa falcata tube turf (frequent), Caryophyllia smithii (frequent, locally common on larger boulders) and Dendrodoa grossularia (locally occasional on larger boulders). From 65 m the biota of the floor boulders began to appear more scoured and was relatively sparse. From ~75 m to 90 m there were extensive areas of the cave floor occupied by rippled sand. In this area and beyond to the end of the surveyed area at 100 m the boulders were distinctly scoured with many surfaces of clean bare rock and significant numbers of Actinia equina (frequent to common).
The Dendrodoa grossularia and Clathrina coriacea community (IR.FIR.SG.DenCcor) was a significant component of the cave wall biota throughout the surveyed area. The biota near the base of the wall was generally more impoverished due to the impacts of scour. This was particularly prominent at distances beyond 70 m due to the sand on the cave floor. Spirorbis tridentatus and Jassa falcata tube turfs tended to predominate on the scoured wall bases with other biota also present but in relatively low abundance. Above this scoured zone, Dendrodoa grossularia was normally abundant. The Clathrina coriacea component of the community was rather sparse on the cave walls in the outer part of the cave but beyond ~30 m from the entrance it was more prominent and was common over extensive areas of the walls. The biota on the upper parts of the cave walls appeared more diverse and profuse. Prominent taxa included Phellia gausapata (locally abundant), Cylista elegans (common to abundant), Crisia eburnea bryozoan turf (locally abundant), Tubularia indivisa (frequent) and Alcyonium digitatum (locally frequent).
The intertidal zone at the waterline was dominated by a narrow band of dense Semibalanus balanoides (abundant to superabundant). In the partially illuminated outer parts of the cave the lower part of the supralittoral cave walls were dominated by Hildenbrandia rubra (abundant) and the upper walls and ceiling by green algal stains. In the darker areas further within the cave the supralittoral zone appeared to consist of bare rock devoid of obvious biota.
In terms of biotopes, the subtidal zone of the well-illuminated entrance area corresponded to the environmental conditions and biota described for IR.FIR.SG.FoSwCC. Within the cave, the floor showed biota typical of scoured conditions. In the deeper water of the outer part of the cave the level of scour was not severe, and these areas were ascribed to IR.FIR.SG.CCBalPom. The match to the characterising species is poor but the situation and environmental conditions correspond well. As the floor became shallower further into the cave (e.g. cross section 2), the evidence of scour was more pronounced and justified the allocation of the IR.FIR.SG.CC.Mo biotope. Throughout the cave the scour extended up the base of the walls to about 1 m above the floor. This relatively impoverished community was also ascribed to IR.FIR.SG.CCBalPom. The majority of the cave wall area within the caves was dominated by a well-developed IR.FIR.SG.DenCcor biotope. However, the community on the upper reaches of the walls shifted to one more dominated by anemones and other taxa. This provided a reasonable match to the description of the IR.FIR.SG.CrSpAsAn biotope. At and above the waterline, there was a band of barnacles corresponding to an impoverished version of LR.HLR.MusB.Sem. Upper walls and ceiling in the outer part of the cave corresponded to LR.FLR.CvOv.VmucHil. But with decreased light levels in the inner part of the cave the supralittoral rock surfaces appeared barren of obvious life.
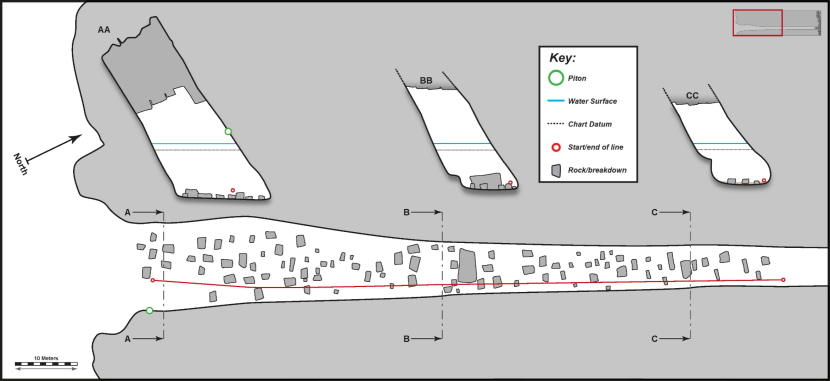
Figure 9. FI21CV04 – The Diamond cave (Colsta) – South entrance (‘CI66’).
Click for a full description
Physical survey (scale bar applies to both floor plan and passage cross sections, associated biological data is provided in tables 7 & 8). ‘BB’ is cross section 1 and ‘CC’ is cross section 2. Insert shows whole cave based on snorkeller records.
Table 7. FI21CV04 – The Diamond cave (Colsta) – South entrance (‘CI66’) – Biological summary – cross section 1 (right wall, ~45 m from entrance, 45 m on tape)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor predominantly composed of boulders with patches (~10%) of sand and gravel. | -6.1 | -6.1 | IR.FIR.SG.CCBalPom | |
2 | Vertical & slightly overhanging rock walls. | -5.1 | -6.1 | Scoured base of rock wall. Spirorbis tridentatus (abundant) & Dendrodoa grossularia (common). | IR.FIR.SG.CCBalPom |
3 | Vertical & slightly overhanging rock walls. | -2.1 | -5.1 | Lower rock wall. Dendrodoa grossularia (abundant) and Clathrina coriacea (common). | IR.FIR.SG.DenCcor |
4 | Vertical & slightly overhanging rock walls. | +0.9 | -2.1 | Upper rock wall. Mixed faunal turfs. Dendrodoa grossularia (occasional, locally common) and Clathrina coriacea (frequent). Crisia eburnea bryozoan turf (abundant). Other taxa becoming more prominent near the top of this zone. Phellia gausapata (occasional, locally abundant upper zone), Tubularia indivisa (frequent, locally abundant upper zone), Alcyonium digitatum (occasional, locally frequent upper zone). | IR.FIR.SG.CrSpAsAn |
5 | Vertical & slightly overhanging rock walls. | +1.4 | +0.9 | Intertidal rock wall. Semibalanus balanoides (abundant) and Cylista elegans (abundant). | LR.HLR.MusB.Sem |
6 | Vertical & slightly overhanging rock walls. | +2.9 | +1.4 | Intertidal rock wall. Semibalanus balanoides (abundant). | LR.HLR.MusB.Sem |
7 | Vertical & slightly overhanging rock walls. | +4.9 | +2.9 | Supralittoral rock wall. Hildenbrandia rubra (abundant). | LR.FLR.CvOv.VmucHil |
8 | Vertical & slightly overhanging rock walls. | >+4.9 | +4.9 | Supralittoral rock wall. Green algal stains (abundant). | LR.FLR.CvOv.VmucHil |
Table 8. FI21CV04 – The Diamond cave (Colsta) – South entrance (‘CI66’) – Biological summary – cross section 2 (right wall, ~85 m from entrance, 85 m on tape)
Zone | Physical notes | Upper limit | Lower limit | Biological notes | Biotope |
|---|---|---|---|---|---|
1 | Cave floor predominantly composed of rippled sand with scattered boulders (~10%). | -5.1 | -5.1 | Bare sand and scoured boulders. Actinia equina (occasional, locally frequent). Jassa falcata tube turf (rare). | IR.FIR.SG.CC.Mo |
2 | Vertical & slightly overhanging rock walls. | -4.6 | -5.1 | Scoured base of rock wall. Dendrodoa grossularia (common). Cylista elegans (frequent). Jassa falcata tube turf (occasional). Bryozoan crusts (occasional). Spirorbis tridentatus (rare). | IR.FIR.SG.CCBalPom |
3 | Vertical & slightly overhanging rock walls. | -4.1 | -4.6 | Scoured base of rock wall. Dendrodoa grossularia (common). Jassa falcata tube turf (common, locally abundant in upper zone). Spirorbis tridentatus (frequent). Bryozoan crusts (occasional). Hydroid turf (occasional). | IR.FIR.SG.CCBalPom |
4 | Vertical & slightly overhanging rock walls. | -1.1 | -4.1 | Lower rock wall. Dendrodoa grossularia (superabundant) and Clathrina coriacea (common). Tubularia indivisa (rare, locally frequent in upper zone). | IR.FIR.SG.DenCcor |
5 | Vertical & slightly overhanging rock walls. | +0.9 | -1.1 | Upper rock wall. Mixed faunal turfs. Dendrodoa grossularia (common) and Clathrina coriacea (occasional). Cylista elegans (common). Phellia gausapata (rare, locally abundant). Tubularia indivisa (frequent). | IR.FIR.SG.CrSpAsAn |
6 | Vertical & slightly overhanging rock walls. | +1.7 | +0.9 | Intertidal rock wall. Semibalanus balanoides (superabundant) and Cylista elegans (common). | LR.HLR.MusB.Sem |
7 | Vertical & slightly overhanging rock walls. | +2.9 | +1.7 | Intertidal rock wall. Semibalanus balanoides (abundant). Actinia equina (frequent). | LR.HLR.MusB.Sem |
8 | Vertical & slightly overhanging rock walls. | >+2.9 | +2.9 | Supralittoral rock wall. Bare rock. | Bare rock. |
4. Discussion
4.1 Extent & number of caves
The collation of a comprehensive inventory of the sea caves of Fair Isle is a significant challenge. The primary obstacle is wave action. The island is at an extremely exposed location with no nearby islands to dissipate waves. Its eastern shores are open to the full width of the North Sea and its western shores are exposed to the full fetch of the Atlantic Ocean. The issue is compounded by the small size of the island which can allow ocean swells to wrap around the coastline as they are refracted by the shallow coastal waters. This has the effect that significant wave surge can still be experienced on a lee shore. A further issue that we experienced on the site was abrupt but short-lived increases in swell height that occurred with apparently unpredictable but long (30 minutes or more) intervals. This has the effect that a site that could be safely accessed in existing background swell levels could suddenly become unsafe due to an unexpected significant increase in swell height. The increased swell levels were generally of short duration but were sufficient to create hazardous levels of surge.
The rough seas create one problem in that access to the caves requires the use of a boat. The coastline is generally characterised by high and rugged cliffs so access from the land is not a practical option. But a greater issue is that sea caves have a funnelling effect on waves and the wave surge becomes amplified within the confines of the caves. A small amount of wave surge may present no difficulties on the open coast but its amplification within caves can create significant working difficulties and is potentially hazardous.
The prevailing sea conditions during the 2021 survey restricted close examination of the coast to some protected areas on the east coast (Figure 3) and one cave on the west. The highly exposed rock cliffs on the north and the west of the island were inaccessible and potentially contain many significant sites. Even within the examined areas it was not always possible to enter the caves. In these areas the sea was sufficiently calm to safely approach cave entrances in an inflatable boat but amplified wave surge within the entrances sometimes prevented snorkellers from making a full assessment of the site. The importance of entering the cave entrance for close examination was demonstrated by the case of FI21CV01. When viewed from the boat at the entrance this appeared little more than an alcove leading to an impassable fissure. When entered by a snorkeller it was established that the unpromising looking entrance opened out into a relatively extensive cave after a short, constricted section.
Despite the limited coverage of the 2021 survey there is a considerable amount of additional information on cave locations provided by local knowledge, the NatureScot 2019 survey and other sources. In the majority of these cases there is limited information available on the extent of these sites. Although cave entrances might be visited by a boat it would be rare that conditions would be suitably calm for entering the cave and rare that the occupants of the boat would be suitably equipped (or inclined) to swim into the cave. However, it is clear that a large number of cave entrances exist and that in some cases there is good evidence that extensive caves or tunnels lie within the entrances. A total of 175 sites have been documented. These probably encompass the vast majority of the larger and more conspicuous entrances but may overlook some less conspicuous entrances requiring examination by snorkellers or swimmers. Another category of cave subject to under-reporting would be sites that are entirely subtidal. No such caves were documented in 2021 but local accounts indicate that they do occur. Finding unknown submerged cave sites would be extremely difficult and it would be near impossible to conduct a systematic and comprehensive search for submerged caves around the island.
A major part of the remit of this study was to assess the number and extent of caves that can be regarded as qualifying as the Annex I habitat ‘submerged or partially submerged sea caves’. Based on the arguments presented in the Introduction (Section 1.5) we regard reduced light levels as the main factor determining if sites can be regarded as Annex I cave habitats. Although we lack data on the majority of caves, it is possible to estimate the number of qualifying caves on Fair Isle. This estimate is based on the assumptions that the assessed caves are representative of the site as a whole and that the records of the total number of caves is roughly correct.
There were 96 sites where passage length estimates are available. It should be appreciated that a high value for passage length does not necessarily imply light levels are sufficiently reduced for the site to qualify as a cave feature. A relatively long cave with a large entrance may admit a considerable amount of light whereas a short cave with a constricted entrance may be relatively dark. But it is certainly probable that longer caves are likely to be dark and the information can be used to estimate the probable number of sites at Fair Isle that qualify as Annex I sea cave features.
Table 9 shows the number of caves recorded in four categories of passage length. If we assume that this ‘sample’ is representative of the size distribution of all Fair Isle caves, we can estimate the total number of caves in each length category based on the estimated total number of 175 sites.
Table 9. A breakdown of the numbers of cave records in terms of passage length and estimation of overall number of caves present on Fair Isle in each size category.
Passage length category | Number of caves recorded in each length category | Percentage of recorded caves in each length category | Estimated number of caves in each length category based on a total of 175 sites |
|---|---|---|---|
<10 m | 40 | 42% | 73 |
10-20 m | 22 | 23% | 40 |
20-50 m | 20 | 21% | 36 |
>50 m | 14 | 15% | 26 |
Table 10 shows the number of caves in each length category where both passage length and light levels were estimated and the number of these caves where light levels were significantly reduced. This is used to calculate the percentage of caves in each category that qualify as an Annex I sea cave feature. The expected relationship between passage length and darkness is clear. Only 5% of caves less than 10 m in length extend into darkness whereas 100% of caves greater than 50 m in length are dark.
Table 10. A breakdown of the numbers of cave records in terms of passage length and presence of reduced light levels. This is used to estimate the percentage of caves with reduced light for each length category.
Passage length category | Number of caves where both length & light levels were noted | Number of dark caves | Percentage of dark caves in each length category |
|---|---|---|---|
<10 m | 40 | 2 | 5% |
10-20 m | 20 | 4 | 20% |
20-50 m | 11 | 7 | 64% |
>50 m | 7 | 7 | 100% |
Table 11 shows the estimated total number of caves of each length category and the expected percentage that will have reduced light levels. This is used to estimate the total number of caves with reduced light levels.
Table 11. A breakdown of the estimated numbers of caves in each length category, the percentage of these that are likely to show reduced light levels and hence an estimate of the total number of caves on Fair Isle with reduced light levels.
Passage length category | Estimated number of caves in each length category based on a total of 175 sites | Percentage of dark caves in each length category | Estimate of number of total dark caves |
|---|---|---|---|
<10 m | 73 | 5% | 4 |
10-20 m | 40 | 20% | 8 |
20-50 m | 36 | 64% | 23 |
>50 m | 26 | 100% | 26 |
Totals | 175 | - | 60 |
So based on the existing information we estimate approximately 60 caves on Fair Isle are likely to qualify as Annex I habitat ‘submerged or partially submerged sea caves’ with reduced light levels as the defining factor.
4.2 Cave configuration
In general, many of the Fair Isle caves tend to have a passage direction aligned on an axis running roughly south west to north east. Additionally, the cave walls tend to be inclined, sloping downward to the south east. Not all caves correspond to this generalisation, but it holds true for many of the caves for which data is available. Of the four caves that were examined in detail, the two largest caves (FI21CV02 & FI21CV04) correspond to the description. In the cave inventory there are 59 caves where there is sufficient information to assess the configuration of the cave. Of these, 45 (76%) correspond to this generalised configuration. Those which differ were often of limited extent. Additionally, many of the images of cave entrances suggest the walls of the entrance are inclined as described above.
The configuration of the caves corresponds well to the configuration of the beds of sedimentary rocks which form Fair Isle. In general, the alignment of the bedding (‘strike’) runs approximately south west to north east. The beds are steeply inclined and sloping downward (‘dip’) in an approximately south easterly direction (Mykura 1972). The caves can be envisaged as voids created by partial removal of areas of specific beds of rock.
The rocks of Fair Isle are predominantly sandstone but there is some variation in the composition of the beds. The beds are therefore likely to differ in terms of how susceptible they are to wave erosion with caves potentially developing along weaker bedding.
A closer examination of the area of ‘Diamond Cave’ (FI21CV04) in relation to the geological map of Mykura (1972) and satellite imagery provides some additional support for the interpretation outlined above (Figures 10 & 11). The predominant rock type of the area is reported as ‘pebbly sandstone' The map also shows three areas defined as ‘predominantly fine grained sediments’. The eastern most two of the areas appears to be a single bed which has been cut across by the fault responsible for the formation of the imposing gorge of Troila Geo. The third structure is to the north of the Troila Geo fault and some 200 m north west of the first bed. Both of these ‘fine grained’ sediment beds are aligned approximately on a south west to north east axis and dip towards the south east. The position and orientation of Diamond cave corresponds to the expected position of the westernmost fine grained bed if it is assumed to continue to the south of the Troila Geo fault in the same manner as the easternmost bed. The satellite image (Figure 11) shows that all three areas of fine grained rock shown on the map are visible as shallow gullies. This implies that this rock type is less resistant to erosion than the surrounding rock and supports the interpretation that Diamond Cave has been formed by the sea level erosion of the same layer of rock.
Another factor influencing cave topography is change in relative sea level due to either post glacial rebound or to subsidence of the crust. Such changes can have consequences for the depth of the cave floor. This in turn has consequences for cave biota as increased floor depth provides a greater area for the establishment of diverse communities above the influence of scour associated with mobile substrates on the cave floor.
Sea caves are generally created by erosion of areas of relatively weak rock by the forces exerted by ocean waves. These erosive forces are only well-developed at or near the sea surface because the wave energy is less focused and less powerful below the surface. Consequently, the active formation of sea caves tends to occur intertidally. In areas where the crust is rising due to glacial rebound the caves tend to be in the mid to upper intertidal zone (e.g. Berwickshire coast). Where rebound is pronounced the caves may become elevated features at the rear of ‘raised beaches’ (e.g. some parts of the Argyll coast). In subsiding areas (e.g. St Kilda) the converse occurs, and caves may have a deep cave floor or become entirely submerged ‘fossil’ features comparable to the ‘fossil’ caves of raised beaches on the west coast of Scotland.
Subsidence of the crust also occurs on Shetland and is apparent in sea caves around Papa Stour and Mousa. At these sites there are a number of sea caves where the cave floor is relatively deep. It is probable that the erosion forming the floors of these caves occurred when sea levels were relatively low and hence the rocks of the cave floor were at or near sea level where erosive forces were strong. Subsequently the crust has subsided resulting in the cave floor now lying in depths where erosion is less intense than at the water surface. The same appears to be true to some extent of the water depth in the Fair Isle caves. But of the caves documented so far the water depths are not as great as those recorded in the Shetland caves. This may indicate less subsidence in Fair Isle and correspondingly implies that the existence of a significant number of well-developed subtidal caves is unlikely. However, this comes with the caveat that only a very small proportion of the caves have been documented so such conclusions are based only on informed speculation.
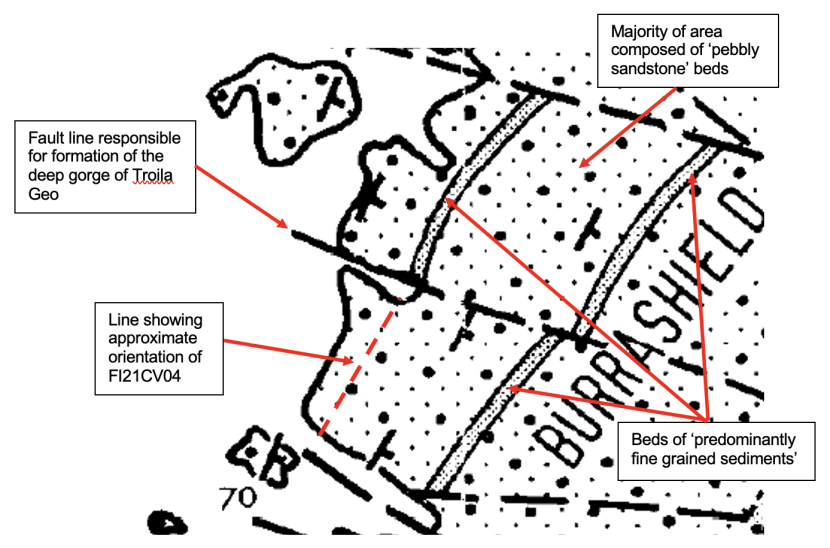
Figure 10. Detail from geological map of Mykura 1972.
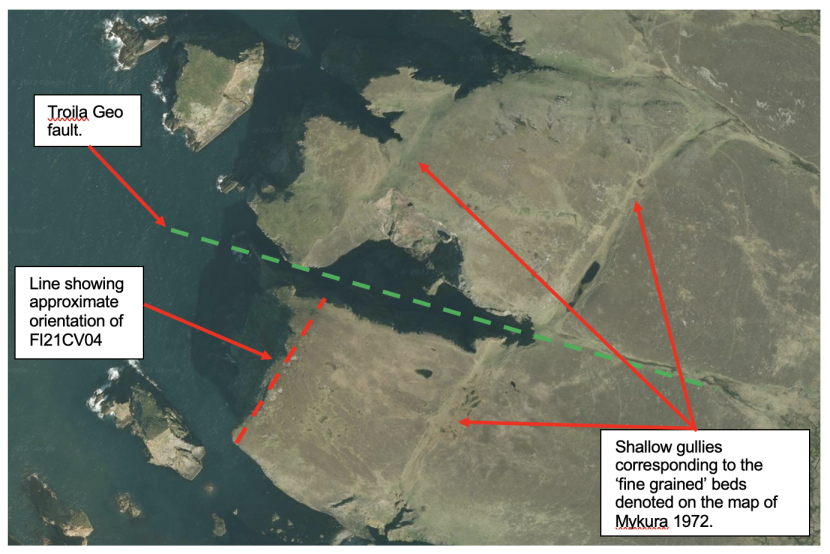
Figure 11. Satellite imagery of Fair Isle showing some surface features. Google Earth Image ©2024 Airbus (Data SIO, NOAA, U.S. Navy, NGA, GEBCO).
4.3 Review of historical records on Fair Isle cave biota
Marine Recorder includes 5 records relating to ‘cave’ habitats around Fair Isle. Two are derived from a 1987 survey conducted as part of the “1986-87 OPRU/MNCR Shetland, Foula and Fair Isle survey”. Two relate to Seasearch records made in 2011 and one to Seasearch records made in 2016.
The first of the two 1987 records was in North Haven (MR sample codes 261.194.001 to 261.194.005 and Annex 1a site code MR02). The coordinates plot in open water ~65 m north east of the Yessness headland which is on the west side of North Haven. These coordinates are therefore only indicative and do not identify a specific cave entrance. Also, the descriptions associated with the record clearly relate to two different caves.
The first of these descriptions (MR sample codes 261.194.001 to 261.194.004) is almost certainly one of the five cave entrances on the tip of the Yessness headland (CI81 to CI85). Due to wave surge we were unable to examine these sites fully in 2021 so cannot confidently link the 1987 description to a specific cave. However, based on the entrance description, it is probable that it relates to CI81. The 1987 description highlights a cave wall community dominated by Dendrodoa grossularia and ascribes the IR.FIR.SG.DenCcor biotope. Further within the cave they note the cave walls are subject to a greater degree of scour with sparse spirobins, Verruca stromia and white sponge crusts. They describe floor depth as rising from ~7 m at the entrance to ~1.5 m near the end of the cave. This account can be related to the 2021 findings at other sites which also found Dendrodoa grossularia was prominent on cave walls where floor depth was sufficient to provide cave wall space above the scour zone (e.g. FI21CV04). The scoured community that they note in the shallower areas at the back of this cave is also comparable to scoured communities noted at shallower sites examined in 2021.
The second description provided in this record (MR sample code 261.194.005) almost certainly relates to FI21CV02 (CI04). The physical description that they provide of the cave is a close match to our observations made in 2021. The 1987 surveyors only examined the biota in the first 20 m of the cave. This is exactly the same as was done in 2021 and the coverage was presumably limited in this way for similar reasons in both surveys (wave surge). The biota described in 1987 was similar to that noted in 2021. The presence of Grantia compressa was highlighted and they also describe a ‘slimy cover’ of undefined biota. This was undoubtedly the ‘tube turf’ which was noted as a significant component of the cave biota in 2021.
The second of the two 1987 records (MR sample code 261.188.003) corresponds to the 2021 site CI52. This is the south western end of the tunnel complex (Da Heelers) that runs through Sheep Rock. In 2021 it was not possible to enter the tunnels due to wave surge. It is not clear how far the 1987 surveyors penetrated into the tunnels but it does appear they did go a reasonable distance beyond the entrance. Parts of the description refer to the “entrance area” and mention the presence of algae. But other parts of the description indicate a biota more typical of a cave environment. They highlight sponge crusts, anemones and Tubularia. They also mention ‘domiculous (tube dwelling) gammeridae’ and note the scoured floor area with mobile substrates. This is in broad accordance with the biota found in the 2021 site FI21CV04 where anemones and Tubularia were notable on the upper cave walls and where tubes of the amphipod Jassa falcata were prominent. The scoured cave floor noted in 1987 is a general characteristic of caves visited in 2021 but the ‘diverse sponge fauna’ that they note on the walls is less recognisable.
The first of the two 2011 Seasearch records was in North Haven (MR sample code MRMCS0070000079D.01). The coordinates correspond to the 2021 site CI03. However, the 2011 description does not correspond to CI03, particularly in regard to water depth at the entrance. It is highly probable that the 2011 description actually relates to CI02 which is nearby and is a more prominent entrance. The 2011 description of the biota provides limited information but does mention a “Clathrina/Dendrodoa mix” on the walls. Wave surge prevented survey of this cave in 2021 but the presence of IR.FIR.SG.DenCcor is plausible given the water depth. However, the extent of this biotope is likely to be relatively limited due to the small dimensions of the cave.
The second of the two 2011 Seasearch records (MR sample code MRMCS0070000079F.01 and Annex 1a site code MR01) is unlikely to constitute a genuine cave record. The coordinates are placed ~100 m offshore in the vicinity of Stroms Heelor. The description refers to a variety of rocky substrates including rock outcrops, boulder areas, gullies and rock walls. Biota of mixed kelps and Alcyonium are mentioned. A “small cave” is mentioned in relation to a vertical rock wall but no further details are provided. It is likely that this was a rock alcove of limited extent. If a significant cave had been present it would surely have merited more emphasis in the description provided by the surveyor.
The 2016 Seasearch record (MR sample code MRMCS020000000AD.001) coincides with the 2021 site GSG03. The description gives an account of a heavily scoured gully with mobile boulders and cobbles on the floor and leading to a “small cave”. The sea state prevented an examination of this area in 2021 so the entrance could not be confirmed. However, local knowledge indicates a cave is present at this location and extends back into full darkness. This is at odds with the 2016 description of a “small cave”. Given the known imprecision of the Marine Recorder coordinates it is possible that the 2016 surveyors were describing a neighbouring site distinct from that of the dark cave described by the Fair Isle inhabitants.
4.4 General overview of Fair Isle cave biota
This section needs to begin with a clear proviso that our knowledge of the sea cave biota remains too limited to draw general conclusions with a high degree of confidence. It is based on a detailed assessment of only four sites supplemented with passing observations made in the course of compiling the cave inventory and with the handful of historical records outlined above.
Scour is normally a significant influence on the biota of sea caves and this is certainly the case in the Fair Isle caves. The degree of scour is linked to the depth and the composition of the cave floor. Cave floors entirely of bedrock or large stable boulders are associated with relatively little scour. If loose mobile substrates (e.g. cobbles, pebbles, gravel or sand) are present then the scour levels are more pronounced. Scour impacts are typically concentrated at or just above the cave floor. Where the cave floor is at a shallow depth the scour impact can cover the entire cave wall. Where the floor is deeper the scoured zone might be restricted to the base of the wall with a more profuse and diverse biota developed on the upper wall beyond the reach of the most intense scour.
At the more intense levels of scour the rock surfaces are almost entirely bare, with attached biota restricted to protective crevices. Mobile biota may be present and at shallow sites the anemone Actinia equina may be common. Communities corresponding to this description were encountered in Fair Isle (e.g. the shallow inner areas of FI21CV01 & FI21CV04). Where scour levels are less pronounced more attached biota can establish although the communities tend to be sparse and impoverished. Scour resistant biota such as Spirorbis tridentatus often predominate at such locations (e.g. the bases of the cave walls in FI21CV04).
Another community indicative of scour is ‘tube turf’. The ‘turf’ is a velvety layer of small (<2 mm) silty tubes that can form a dense cover on cave walls and floor boulders. The species which form the tube turf can differ from site to site and sometimes a mix of species contribute to the formation of the turf at a given site. The sabellid fan worm Fabricia stellaris is often a major component of the tube turf. On first impressions, the fragile silty tube inhabited by these worms would seem poorly adapted to the high energy environments of sea caves. However, these worms are unusual amongst sabellids in that they can reattach to rock surface and readily rebuild their tubes if displaced (Lewis 1968). It is probable that this ability to reattach is a key factor allowing them to maintain dense populations in sea cave environments where wave action regularly displaces biota from the rock surfaces. Another major contributor to tube turf is the tube-building amphipod Jassa falcata. At the sites surveyed in Fair Isle, the tube turfs in shallower more scoured sites (FI21CV01 & FI21CV03) tended to be dominated by Fabricia stellaris and at deeper sites (FI21CV02 & FI21CV04) the tube turf was dominated by Jassa falcata. Clearly, far too few sites have been assessed to determine if this distributional difference applies more generally but it would be interesting to investigate further. This account focusses only on the most abundant of the tube building species and perhaps over-simplifies matters. In reality, the tube turf can contain a diverse range of tube builders including other species of amphipods & sabellids, small terebellid polychaetes and taneids.
Many of the caves visited on Fair Isle in 2021 tended to have a relatively shallow floor and the associated scour influenced most of the cave wall area. The biota of such caves was commonly characterised by dense cover of tube turf, sometimes profuse growths of the purse sponge Grantia compressa and sparse sponge crusts formed by Leuconia sp. and or Halichondria panicea. This assemblage was characteristic of the shallower surveyed sites (FI21CV01 & FI21CV03) but was also frequently observed in the many shallow caves visited when gathering data for the cave inventory. Similar assemblages have been previously reported as occurring frequently in the sea caves of Mousa (Harries et al., 2009, Harries et al., 2019). These caves are formed in sandstones of a similar age to those on Fair Isle and similarly, the majority of the cave floors are relatively shallow.
In the Fair Isle caves with greater water depth other assemblages become established on the cave walls. One that may be common is characterised by the sea squirt Dendrodoa grossularia which is sometimes associated with significant cover of the lace sponge Clathrina coriacea. This assemblage was an obvious characterising feature of the cave walls in FI21CV04 but was also present in an impoverished form at the shallower site of FI21CV02. These were the two deepest caves examined in 2021 and the probability of the assemblage being widespread in suitable sea cave sites on Fair Isle is increased because it was also noted in two of the historical records of sea caves with relatively deep floors. The Dendrodoa / Clathrina assemblage is a recognised sea cave community noted at many sites. For example, impoverished versions of this have been reported from Mousa sea caves (Harries et al., 2009, Harries et al., 2019) and more well-established versions from the sea caves of Loch Eriboll (Harries et al., 2018).
FI21CV04 was the deepest site examined in 2021 and the upper cave walls supported a diverse mosaic of biota. Prominent components of this mosaic included dense aggregations of anemones such as Phellia gausapata and Cylista elegans and turfs of crisid bryozoans. In the shallower areas just below the waterline there were profuse growths of the hydroid Tubularia indivisa and the soft coral Alcyonium digitatum. It is probable that similar communities occur in other Fair Isle sea caves where floor depth is sufficent to minimise the impact of scour. The 1987 record from the Da Heelers tunnels in Sheep Rock (CI52) certainly highlights the presence of anemones and Tubularia as characteristic. An example of another site where such assemblages may occur is Muckle Rip-heelor (CI47) with water depth of 12 m BCD at the entrance. Wave surge excluded the possibility of a full survey but the cave was inspected by snorkeller and profuse growth of Tubularia was noted on the upper walls.
Variations of this diverse upper wall assemblage have been noted in sea caves with deep floors at a number of other locations. For example, dense anemones and Tubularia were notable on upper cave walls in St Kilda and North Rona (Harries et al., 2018) and also at one of the deeper caves on Mousa (Harries et al., 2009, Harries et al., 2019). Crisid turfs were also noted at this same cave on Mousa but were best developed in the St Kilda caves where they were better developed than at either Mousa or Fair Isle.
The account provided above highlights some similarities between the Fair Isles caves and those of other sites such as St Kilda, North Rona, Loch Eriboll and Mousa. There are also parallels that could be drawn with the biota of Papa Stour sea caves (ERT, 2005). North Rona caves did differ to some extent in terms of the prominence of colonial ascidians (mainly didemnids) on the cave walls which have not been observed in such high abundances in Fair Isle or the other sites. Berwickshire coast is another significant Scottish sea cave site but at Berwickshire all the more extensive caves are intertidal so are not comparable with the essentially subtidal areas surveyed at Fair Isle and the other sites.
Although weather conditions limited the detailed surveys of the Fair Isle caves to only four sites, those sites are a good reflection of the gradient of scour associated with cave floor depth and the consequences of the scour on the composition and diversity of communities within the caves. FI21CV01 & FI21CV03 were shallow and tended to support the assemblage of tube turf, Grantia compressa and sparse sponge crusts. FI21CV02 was somewhat deeper and supported a similar assemblage but with the addition of significant numbers of Dendrodoa grossularia which resembled an impoverished version of the Dendrodoa / Clathrina assemblage. The increased water depth at FI21CV04 was associated with a well-developed Dendrodoa / Clathrina assemblage and also the diverse mosaic of anemones, crisid bryozoan turf and Tubularia.
4.5 Anthropogenic impacts
It is appropriate to include an assessment of the potential for anthropogenic impacts on the sea cave habitats, but the subject does not warrant a lengthy assessment and has been addressed in the Introduction (section 1.4). No anthropogenic impacts were noted during the 2021 survey and direct impacts are thought to be unlikely because the caves are inaccessible and offer no resources likely to be exploited so direct disturbance is unlikely. There are no significant local sources of pollution. The caves could be impacted in the event of an oil spill, but they are such high energy sites that it is probable the oil would be rapidly dispersed and the site recolonised in a short space of time. Broad scale impacts such as climate change or ecological shifts linked to overfishing have potential to impact the caves but such impacts would have much broader consequences and there is no evidence to suggest the cave biota would be any more vulnerable to such impacts than other marine habitats.
4.6 Recommendations
It would be desirable to gather more information on the Fair Isle caves both in terms of the number and extent of caves and in terms of the biota. The main obstacle to this is that suitable sea conditions for access are relatively rare and are unpredictable. The infrequent windows of opportunity for accessing caves can only be readily exploited by those who are based on the island for lengthy periods. It would also of course require access to a suitable boat, an interest in documenting sea caves and ideally the capacity to swim into the caves (i.e. health and safety precautions, wetsuit, snorkel gear etc). The gathering of information by such means would also benefit from a straightforward but robust protocol on recording information. Imagery and reliable recording of coordinates would be an important component of such information. Also, the noting of factors such as light levels and water depth should be highlighted due to their importance in predicting the nature and diversity of biological communities that might occur at a site. NatureScot have developed a recording form in collaboration with other organisations that may help with this.
A further recommendation for future surveys of this sort is that it might be advantageous to seek the advice of a structural geologist. Ideally, they would participate in both the planning phase and in the fieldwork. A suitably experienced geologist might be of value in predicting where geological weaknesses might favour cave formation and hence help prioritise areas to be searched for sea caves. But it should also be noted that the areas examined during the 2021 survey were determined entirely by the prevailing sea conditions and this limitation would remain regardless of predictions of priority areas for sea caves.
4.7 Assessment of site condition
There are no robust baseline data to enable the condition of the caves to be assessed by direct comparison with a previous state. However, there are some historical data dating back to 1987 and comparisons have been drawn with this in the account above. Another approach to condition assessment is to draw comparisons with other sites deemed to be in good condition. Again, such comparisons have been drawn with suitable sites in the account above. A further approach to condition assessment is through an assessment of the evidence that past, present or future activities have potential to damage the feature. This has also been addressed in the account above.
The overall conclusion is that the Fair Isle sea caves should be regarded as being in good condition. This is based on the following points:
- There is no evidence of current anthropogenic impacts on the feature.
- No existing activities in the area have a realistic potential to impact on the feature.
- The potential for future anthropogenic impacts on the feature is extremely unlikely (assuming the former practice of using certain gloups for waste disposal does not resume).
- The biota recorded within the caves visited in 2021 was broadly comparable to that reported in 1987.
- The biota recorded within the caves visited in 2021 was broadly comparable to that recorded at similar sites around Scotland (St Kilda, North Rona, Loch Eriboll, Mousa & Papa Stour).
5. References
Ackers, R.G., Moss, D., Picton, B.E., Stone, S.M.K. & Morrow, C.C. 2007. Sponges of the British Isles (“Sponge V”). Marine Conservation Society.
Anon, 1998. A Statement on Common Standards Monitoring. Peterborough: Joint Nature Conservation Committee.
Bunker, F. & Holt, R. 2003. Surveys of Sea Caves in Welsh Special Areas of Conservation. CCW Marine Monitoring Report No. 6.
Butler, R.W. 2016. Changing politics, economics and relations on the small remote island of Fair Isle. Island Studies Journal, 11(2), 687-700.
Chapman, P. 1993. Caves & Cave Life. Harper Collins Publishers. ISBN: 0 00 219908 4.
Connor, D.W., Allen, J.H., Golding, N., Howell, K.L., Lieberknecht, L.M., Northern, K.O. & Reker, J.B. 2004. The National Marine Habitat Classification for Britain and Ireland. Version 04.05. Peterborough: Joint Nature Conservation Committee. ISBN: 1 861 07561 8 (internet version).
Cornelius, P.F.S. 1995. North-West European thecate hydroids and their medusae: Keys and notes for identification of the species. Volume 50 of Synopses of the British fauna, Linnean Society of London. (Field Studies Council).
Ellis, B. 1988. An introduction to cave surveying. A handbook of techniques for the preparation and interpretation of conventional cave surveys. BCRA Cave studies series no.2. ISBN 0 900265 04 3.
ERT (Scotland) Ltd, 2004. Site condition monitoring: Berwickshire and North Northumberland Coast (BNNC) littoral and sublittoral caves, June 2003. Scottish Natural Heritage Commissioned Report No. 069 (ROAME No. F02AA409).
ERT (Scotland) Ltd, 2005. Site Condition Monitoring: surveys of marine rocky environments in the Papa Stour cSAC July-August 2003. Scottish Natural Heritage Commissioned Report No. 102 (ROAME No. F02AA409b).
European Commission, 2013. EUR 28 – Interpretation Manual of European Union Habitats. European Commission, DG Environment, Nature ENV B.3, 146pp.
Harries, D.B., Moore, C.G., Cook, R.L., Kamphausen, L., Porter, J.S., Lyndon, A.R. & Pye, S.E. 2019. 2016 site condition monitoring of the rocky reefs and sea caves of Mousa SAC and survey of sedimentary habitats of the Mousa to Boddam MPA. Scottish Natural Heritage Research Report No. 1080.
Harries, D.B., Moore, C.G., Lyndon, A.R. & Mair, J.M. 2009. The establishment of site condition monitoring of the rocky reefs and sea caves of Mousa Special Area of Conservation. Scottish Natural Heritage Commissioned Report No. 326 (ROAME No. R07AC705).
Harries, D.B., Moore, C.G., Porter, J.S., Sanderson, W.G., Ware, F.J. & Kamphausen, L. 2018. The establishment of site condition monitoring of the sea caves of the St Kilda and North Rona Special Areas of Conservation with supplementary data from Loch Eriboll. Scottish Natural Heritage Research Report No. 1044.
Hayward, P.J. 1985. Ctenostome Bryozoans: Keys and Notes for the Identification of the Species. Issue 33 of Synopses of the British fauna, Linnean Society of London.
Hayward, P.J. & Ryland J.S. 1979. British Ascophoran Bryozoans: Keys and Notes for the Identification of the Species. Issue 14 of Synopses of the British fauna, Linnean Society of London.
Hayward, P.J. & Ryland J.S. 1990. The Marine Fauna of the British Isles and North-West Europe. Oxford University Publications.
JNCC, 2004. Common Standards Monitoring Guidance for Marine, Version August 2004, ISSN 1743-8160.
Lewis, D.B. 1968. Some aspects of the ecology of Fabricia sabella (Ehr.) (Annelida, Polychaeta). J. Linn. Soc. (Zool.), 47(313) 515-526.
Lincoln, R.J. 1979. British Marine Amphipoda: Gammaridea. British Museum (Natural History).
Millar, R.H. 1970. British Ascidians: Keys and Notes for the Identification of the Species. Issue 1 of Synopses of the British fauna, Linnean Society of London.
Mykura, W. 1972. The Old Red Sandstone sediments of Fair Isle, Shetland Islands. Bulletin Geol. Surv. Great Britain, 41, 1-31.
Ryland, J.S. & Hayward, P.J. 1977. British Anascan Bryozoans: Keys and Notes for the Identification of the Species. Issue 10 of Synopses of the British fauna, Linnean Society of London.
Schuchert, P. 2012. North-West European Athecate Hydroids and Their Medusae: Keys and Notes for the Identification of the Species. Issue 59 of Synopses of the British fauna, Linnean Society of London.
Southward, E.C. & Campbell, A.C. 2006. Echinoderms: Keys and Notes for the Identification of the Species. Issue 56 of Synopses of the British fauna, Linnean Society of London.
Tebble, N. 1976. British Bivalve Seashells. A Handbook for Identification, 2nd ed. Edinburgh: British Museum (Natural History), Her Majesty's Stationary Office.
WoRMS Editorial Board, 2022. World Register of Marine Species. World Register of Marine Species at VLIZ. Accessed 2022-06-14. doi:10.14284/170.
ANNEX 1 – Fair Isle Cave Inventory
This annex provides summary information on all known Fair Isle caves. Summary information is tabulated in Annex 1A. Cave locations are shown in the maps presented in Annex 1B. Photographs and sketch surveys (where available) are shown in Annex 1C. Four caves were subject to detailed survey in 2021 and detailed information on these is provided in Annex 2.
- Annex 1A – Fair Isle cave inventory (CSV)
The sites are ordered on the table running clockwise around the coast of Fair Isle starting from the jetty at North Haven. The ‘Source’ column indicates the main sources of information on the sites and this is also reflected in the site code. Site codes prefixed by ‘CI’ were visited in 2021 (with ‘HWU 2021’ noted in the ‘source’ column). Site codes prefixed by ‘NS’ represent information gathered by Carol Hume (NatureScot) in 2019 based on site visits & local knowledge (with ‘SNH 2019’ noted in the ‘source’ column). Site codes prefixed by ‘GSG’ are based on information derived from the Grampian Speleological Group cave database (with ‘GSG’ noted in the ‘source’ column). Site codes prefixed by ‘MR’ are derived from pre-existing Marine Recorder records indicating cave presence (with ‘MR data’ noted in the ‘source’ column). Site codes prefixed by ‘XC’ relate to reported cave locations which were inspected, and no cave was found to be present. The ‘Source’ column may indicate other information sources. For example, ‘OS Map’ denotes cave indicated on Ordnance Survey maps and ‘N & S Thompson’ indicates information provided by Neil & Stewart Thompson (residents of Fair Isle). Sites confirmed as Annex I qualifying sea caves are indicated by an additional column on the table.
- Annex 1B – Cave location maps (PDF)
Maps showing locations of caves listed in Annex 1A
- Annex 1C – Cave photographs and survey sketches (PDF)
Photographs and survey sketches (where available) of caves listed in Annex 1A and mapped in Annex 1B
ANNEX 2 – Fair Isle cave site relocation details with physical and biological data
This Annex contains sheets intended for field use. They provide information to allow for the relocation of the entrance, repositioning of the survey line and recognition of the biological zones.
ANNEX 3 – Fair Isle cave species abundance data (SACFOR)
This Annex contains the semi-quantitative biological abundance data from the zones of the 4 surveyed caves.
- Annex 3A – FI21CV01 (CSV)
- Annex 3B – FI21CV02 (CSV)
- Annex 3C – FI21CV03 (CSV)
- Annex 3D – FI21CV04 (CSV)
ANNEX 4 – Survey log
Outline of activities during the survey.
- Annex 4 – Survey log (DOCX)
ANNEX 5 – Video log
File names for video files segmented into biotopes, with time details for video files of sample events (cave code with cross section and zone suffix).
- Annex 5 – Video log (CSV)
Find out more
- Blog coming soon!
- Fair Isle Management Research Organisation







