NatureScot Research Report 1370 - Potential actions to support recovery of seabird populations in Scotland
Published: 2024
Authors: Furness, R.W., Goodship, N., Raveh, S. (MacArthur Green) and Philip, E. (NatureScot)
Cite as: Furness, R.W., Goodship, N., Raveh, S. and Philip, E. 2024. Potential actions to support recovery of seabird populations in Scotland. NatureScot Research Report 1370
Keywords
seabird; conservation; pressures; mitigation; compensation; action; Scotland
Background
There are multiple seabird work initiatives that have recently concluded, or are currently underway, that have been developed to identify some of the threats and pressures to which seabirds are vulnerable in Scottish waters and/or at breeding colonies. For the main, they focus on specific threats/pressures and/or certain species and therefore there can overlap in proposed actions suggested to reduce these pressures. For instance, actions identified to address climate change resilience are similar to proposed compensatory actions recommended for marine energy development derogations. A holistic approach that considers the multitude of actions that could be implemented across Scotland, what these are likely to consist of, and how these would be most appropriately delivered will provide clarity on collaborative opportunities, increase efficiencies in funding, resourcing and delivery and most significantly, maximise benefits for seabird recovery.
The aim of this project is therefore to develop proposals for potential actions to aid delivery of seabird conservation in Scotland. This project seeks to draw together existing knowledge and outputs from different seabird work programmes to provide a series of potential seabird conservation actions for Scotland. These will provide the basis for a Scotland-wide approach to tackling seabird population declines.
Key unpublished references were supplied by NatureScot and the Scottish Government Marine Directorate.
Main findings
- There is strong evidence that climate change, mammal predators (especially introduced mammal species), and depletion of forage fish stocks by industrial fisheries are major pressures on many seabird species breeding in Scotland.
- There are many other smaller pressures affecting particular species of seabirds.
- Most seabird populations in Scotland have declined in recent years, so there is a need for potential actions that improve the conservation status of seabirds.
- The potential actions that this report has identified include:
- Ecosystem-based management of forage fish fisheries.
- Supplementary feeding.
- Monitoring and reduction of fisheries bycatch of seabirds.
- Monitoring and reduction of aquacultural exclusion net entanglement of seabirds.
- Eradication of invasive mammals from seabird islands.
- Biosecurity for seabird islands.
- Predator-exclusion fences.
- Nest rafts, tern terraces and chick shelters, seabird nest boxes, artificial colony structures.
- Education programmes to raise public support for seabird conservation.
- Strategic selection of areas for marine renewable development.
- Reduction of human disturbance of seabirds.
- Reduction of marine litter.
- Vegetation management at seabird colonies.
- Create islets for terns or gulls.
- Monitoring of disease.
- Reducing harvest and lethal control of seabirds.
- Enhanced monitoring of seabird demography.
Acknowledgements
We thank NatureScot and Marine Directorate, especially Erica Knott, Jen Graham, Lucy Quinn, Sarah Lawrence, Elaine Tait and Jayne Burns, for their inputs to this review throughout the process, and thank Lucy Quinn and Elaine Tait for helpful, detailed and timely comments on the draft report.
- Part 1 Summary of pressures threats and appropriate adaptive responses
- Part 2. Potential seabird conservation actions
- Part 3. Recommendations on potential actions and priorities
- References
- Annex 1
- Abbreviations
Introduction
There are multiple seabird work initiatives that have recently concluded, or are currently underway, that have been developed to identify some of the threats and pressures to which seabirds are vulnerable in Scottish waters and/or at breeding colonies. For the main, they focus on specific threats/pressures and/or certain species and therefore there can overlap in proposed actions. For instance, actions identified to develop climate change resilience are similar to proposed compensatory actions recommended for marine energy development derogations. A holistic approach that considers the multitude of actions that could be implemented across Scotland, what these are likely to consist of, and how these would be most appropriately delivered will provide clarity on collaborative opportunities, increase efficiencies in funding, resourcing and delivery and most significantly, maximise benefits for seabird recovery.
The aim of this project is therefore to develop a list of potential actions that could be taken to aid delivery of seabird conservation in Scotland. This project seeks to draw together existing knowledge and outputs from different seabird work programmes to provide a comprehensive series of detailed potential seabird conservation actions for Scotland. These will provide a Scotland-wide approach to tackling seabird population declines.
There are three elements to this project:
- A review, carried out in February-March 2023 based on works published before that date plus selected unpublished reports provided by NatureScot, to identify a comprehensive 'long-list' of potential seabird conservation actions.
- A series of descriptions of potential seabird conservation actions.
- Discussion and recommendations on potential actions and priorities, including discussion of actions/options likely to have the greatest benefit.
The report provides a short narrative on the potential feasibility and/or long-term practicalities of the potential conservation actions – it is not however a detailed feasibility assessment.
Methods
Part 1: Summary of pressures, threats and appropriate adaptive responses
Part one of this review summarises pressures and threats facing Scottish seabirds and the work completed to date to identify a comprehensive ‘long-list’ of potential seabird conservation actions.
This review uses reference material detailed in Annex 1 which was listed by NatureScot for review, as well as information in Mitchell et al. (2004), Forrester et al. (2007) and recently published literature.
The review assessed 20 threats/pressures that are known to currently impact seabird populations in Scotland; for each threat/pressure, a section on appropriate adaptive responses is summarised.
Threats/pressures assessed include:
- Climate change;
- Depletion of forage fish stocks;
- Fisheries bycatch;
- Predation;
- Marine renewables collision with turbine blades;
- Disturbance and displacement;
- Pollution;
- Fishery discards;
- Nest site interspecific competition;
- Changes to nesting habitat;
- Refuse disposal and agricultural practices;
- Disease epidemics; and
- Lethal control, sport shooting and harvesting.
The review collates all potential recommended actions across all of the key reference material to be clear on which species will benefit (directly or indirectly) and the different threats and pressures addressed.
Two types of potential conservation actions are provided:
- Appropriate direct actions: Conservation action is aimed at reducing or eliminating the specific threat/pressure for the particular seabird species; and
- Appropriate indirect actions: Conservation action is aimed at the species but because it is not possible to address the specific threat/pressure, other threats/pressures are identified which could make the species more resilient to the specific threat/pressure.
This review is focused on providing conservation actions for the following seabird species breeding in Scotland:
- Kittiwake, Rissa tridactyla;
- Herring gull, Larus argentatus;
- Great black-backed gull, Larus marinus;
- Lesser black-backed gull, Larus fuscus;
- Sandwich tern, Sterna sandvicensis;
- Roseate tern, Sterna dougallii;
- Common tern, Sterna hirundo;
- Arctic tern, Sterna paradisaea;
- Little tern, Sternula albifrons;
- Great skua, Stercorarius skua;
- Arctic skua, Stercorarius parasiticus;
- Common guillemot, Uria aalge;
- Razorbill, Alca torda;
- Black guillemot, Cepphus grylle;
- Puffin, Fratercula arctica;
- European storm-petrel, Hydrobates pelagicus;
- Leach’s storm-petrel, Oceanodroma leucorhoa;
- Fulmar, Fulmarus glacialis;
- Manx shearwater, Puffinus puffinus;
- Gannet, Morus bassanus;
- Cormorant, Phalacrocorax carbo; and
- Shag, Gulosus aristotelis.
This review does not consider in detail the pressures that might be managed for these same species in overseas wintering areas or on their migration routes.
Part 2: Potential seabird conservation actions
Part two presents a series of individual descriptions of potential seabird conservation actions that can be cross-referenced by species, threat(s)/pressure(s) and activity.
Each potential action includes:
- Summary description – of the potential action;
- Scope of potential action – which threat(s), pressure(s) will be managed, which species will benefit and geographic scope, including where this would benefit from international collaboration;
- Options - for each potential action, where these exist, highlighting ‘stacked benefits’ (multiple options for some actions and opportunity to target multiple species with a single action) and collaborative opportunities;
- Delivery involvement - who needs to be involved i.e. skills required, organisational input;
- Delivery mechanism(s) - i.e. strategic, site-based, new or existing working groups/projects, funding streams;
- Timescales for delivery; and
- Dependencies/constraints - including on action outwith Scotland.
Part 3: Recommendations on potential actions and priorities
Part three provides discussion and recommendations on potential actions and priorities, including discussion of actions/options likely to have the greatest benefit and where it might be best to target actions. This takes account of necessary timescales for implementation (i.e. actions that can be readily implemented), the constraints that need to be addressed as a matter of priority, potential actions that focus on the most vulnerable species and potential actions that could achieve multiple species benefits.
Part 1. Summary of pressures, threats and appropriate adaptive responses
Climate change
Impacts
Mitchell et al. (2004) identified a number of seabird species in Britain and Ireland that are subject to pressures and threats from climate change. They suggested that species at greatest risk were fulmar, Manx shearwater, shag, Arctic skua, great skua, kittiwake, common guillemot and puffin. Forrester et al. (2007) and Burthe et al. (2014), suggested that fulmar, Manx shearwater, shag, Arctic skua, great skua and common guillemot were particularly vulnerable to climate change in a Scottish context.
From analysis of climate envelope models and local climatic suitability indices in relation to seabird population sizes in Britain and Ireland, Russell et al. (2014) concluded that climate influences the population sizes of multiple seabird species in Britain and Ireland, and that climate change has already altered seabird abundances and distributions. There is widespread empirical evidence from seabird studies around the world for both indirect (bottom-up food web effects) and direct effects of climate change on seabird demography (Furness, 2016).
Climate envelope modelling at the scale of Europe (Huntley et al., 2007; Russell et al., 2015) predicted declines in the breeding range across Europe of most of the seabird species that breed in Britain and Ireland, but with some inconsistent predictions between the two studies despite similar methodology and overlap of authors involved in these two studies. For the species included in this review, Huntley et al. (2007) predicted decreases in breeding range for 20 and an increase for two species. Russell et al. (2015) predicted decreases in breeding range for 15 and increase for seven species.
The Marine Protected Area Management and Monitoring (MarPAMM) project resulted in several reports on climate change impacts on seabird populations. These reports included: a review of the literature on this topic (Johnston et al., 2021b), predictions of change in seabird numbers resulting from climate change modelled from 2000 to 2050 at a Great Britain and Ireland scale and for a defined area west of Scotland (Davies, Humphreys and Pearce-Higgins, 2021), modelling of at-sea distributions predicted from the combination of the predicted changes in numbers and pre-existing at-sea species distribution models (Cleasby, Wilson and Davies, 2021) and an assessment of adaptation responses that could reduce impacts of climate change on these populations (Pearce-Higgins, Davies and Humphreys, 2021).
Searle et al. (2022a,b) examined the potential impacts of climate change on seabirds off the east coast of Scotland. Their models found little or no effect of climate metrics on the breeding success of shag, herring gull or razorbill. The best model for gannet predicted a small increase in breeding success as a result of climate change. For kittiwake, great black-backed gull, common guillemot and puffin models predicted large decreases in breeding success with climate change. Breeding success does not necessarily determine population trend, as seabird populations are likely to be especially strongly influenced by change in adult survival, so direct comparison between Searle et al. (2022a,b) and climate envelope predictions or the MarPAMM predictions is not straightforward.
Davies, Humphreys and Pearce-Higgins (2021) predicted decreases in breeding numbers for 14 of the species included in this review and increases for three species, but the authors were unable to derive models for five species. However, the models they developed in most cases explained low percentages of the variance and so were considered by the authors to have poor or very poor predictive power. Their models also tended to emphasise terrestrial metrics as having more influence than marine ones (such as sea surface temperature), although the review by Johnston et al. (2021b) and the analysis by Searle et al. (2022a,b) emphasised the evidence for climate change impacts on seabirds being mostly related to bottom-up effects of climate change on marine food webs with terrestrial climate change impacts being relatively minor (see also Carroll et al., 2015, but see Olin et al., 2023 for an example of direct impacts of solar heating on breeding seabirds). Searle et al. (2022a,b) chose to classify models of climate change impact on seabird breeding success as not being better than null models unless the model explained more than 10% of the variance. On that criterion, 12 of the models derived by Davies, Humphreys and Pearce-Higgins (2021) would fail.
While some predictions are strongly consistent for a few seabird species (for example, all studies that considered Arctic skua, Arctic tern or black guillemot predicted a decrease in the numbers of these species), some predictions are strongly contradictory. Russell et al. (2015) predicted an increase in fulmar numbers whereas Huntley et al. (2007) and Davies, Humphreys and Pearce-Higgins (2021) predicted a decrease. Davies, Humphreys and Pearce-Higgins (2021) predicted a large decrease (-82%) in European storm-petrel numbers in Britain and Ireland, yet Mitchell et al. (2004) and Forrester et al. (2007) did not identify climate change as an important threat to storm-petrels in Britain and Ireland, and both Huntley et al. (2007) and Russell et al. (2015) predicted increases in European storm-petrel breeding range as a consequence of predicted climate change. Searle et al. (2022a,b) predicted an increase in gannet breeding success and improved habitat availability for foraging (warmer waters) whereas Mitchell et al. (2004) and Forrester et al. (2007) suggested little or no influence of climate change on gannets and both Huntley et al. (2007) and Russell et al. (2015) predicted reduced breeding range of gannet in Europe as a result of predicted climate change.
In summary, there is increasing evidence of climate change influencing seabirds in Scotland and it is likely that this will be one of the strongest influences on seabird populations in coming years. However, it seems that there is little confidence to be placed in predictions regarding exactly what those impacts will be in terms of seabird population change. The most confident predictions relate to the likely losses of northern seabird species from Scotland which have their southern limit of breeding range currently in or close to Scotland, including Arctic skua, Arctic tern, black guillemot and possibly also kittiwake and puffin.
Appropriate adaptive responses
Pearce-Higgins, Davies and Humphreys (2021, p. 5) state “There are no potential counteracting adaptation responses [direct conservation actions] to directly address the impacts of climate change on the marine environment. Instead, we suggest that marine responses should seek to reduce the impacts of other pressures on seabirds, making those populations more resilient to potentially negative climate change impacts”.
The adaptive response would be to achieve net zero globally, and possibly to reduce atmospheric carbon dioxide levels back towards historic levels. The discussions on how to achieve this are continuing elsewhere and are likely to be ongoing for some time. We also know that we are locked into future climate change impacts as a result of current emissions. Given this, and that climate change is potentially one of the most severe impacts on seabird populations, the objective must be to make seabird populations more resilient by addressing other pressures. The options set out in this report will help achieve this.
Depletion of forage fish stocks
Definition and distribution of forage fish
Forage fish are short-lived, small oily fish that feed on plankton and grow quickly. Most forage fish are shoaling pelagic species. Forage fish are a key food source for many large predatory fish as well as many marine mammals and seabirds. They are also targeted by ‘industrial’ fisheries that harvest these abundant fish to provide the raw material for conversion into fish meal and fish oil.
Species composition of forage fish differs between ecosystems, but in each environment, they play an equivalent ecological role of energy transfer from plankton to top predator. In Scottish waters, the key forage fish are sandeel, juvenile herring and sprat (part of the herring family). In the Arctic, the role is played by capelin and juvenile herring, but in warm-temperate seas, sardines and anchovies are present (especially in upwelling regions).
These forage fish species differ in their habitat requirements and spatial distribution. Sandeels are an unusual kind of forage fish in that they spend the winter buried in the seabed sand, but come the spring and summer, they migrate up through the water column to the zooplankton layer to feed. Sandeels therefore require sandy sediment with appropriate grain size in which to bury themselves and are patchily distributed in the North Sea (Figures 1 and 2) and off the west coast of Scotland (Figure 3).
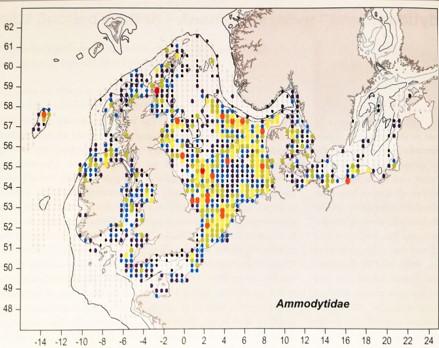
Figure 1. Distribution of sandeels around the UK. From Heessen, Daan and Ellis (2015). Warmer colours represent higher abundance within a particular sampling square.
Click for a full description
Map illustrating distribution of sandeel around the UK. Red dots represent sampling squares in the sea with the highest sandeel abundance, abundance in sample squares decreases in the order of orange, yellow, green and light blue dots, dark purple dots represent sampling squares in the sea with the lowest sandeel abundance. The highest sandeel abundances are located within the North Sea but distribution is patchy.
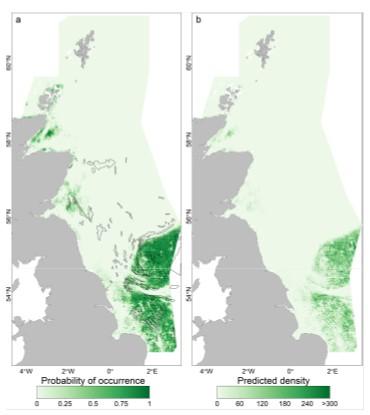
Figure 2. Predicted probability of presence a) and predicted density b) of lesser sandeels based on habitat modelling, and locations of major fishing grounds for lesser sandeel (inside black lines). From Langton et al. (2021).
Click for a full description
A figure showing two maps of the North Sea around the UK. The first map a) illustrates the lesser sandeel probability of presence ranging from 0 light green to 1 dark green. The second map b) illustrates lesser sandeel predicted density ranging from 0 light green to >300 dark green. The ranges are based on habitat modelling and locations of major fishing grounds for lesser sandeel - the latter are represented by black polygons on the maps. Sandeels are patchily distributed in the North Sea, the highest probability of presence and predicted density (dark green) are located on sand banks.
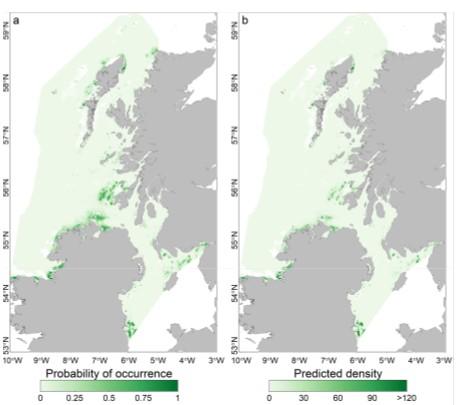
Figure 3. Predicted probability of presence a) and predicted density b) of lesser sandeels based on habitat modelling. From Langton, Boulcott and Wright (2021).
Click for a full description
A figure showing two maps of the Atlantic Ocean off the west coast of Scotland. The first map a) illustrates the probability of lesser sandeel presence ranging from 0 light green to 1 dark green. The second map b) illustrates lesser sandeel predicted density ranging from 0 light green to >300 dark green. The ranges are based on habitat modelling for lesser sandeel and are patchily distributed.
Sprats tend to occur in estuarine habitats; Scotland is located on the northern edge of their distribution and therefore they are rarely found in northern Scotland (Figures 4 and 5). Highest concentrations of sprats are found in the south-eastern North Sea in German and Dutch waters, and in the Irish Sea.
Figure 4. Distribution of sprat around the UK. From Heessen et al. (2015). Warmer colours represent higher abundance within a particular sampling square.
Click for a full description
Map illustrating distribution of sprat around the UK. Red dots represent sampling squares in the sea with the highest sprat abundance, abundance in sample squares decreases in the order of orange, yellow, green and light blue dots, dark purple dots represent sampling squares in the sea with the lowest sprat abundance. The highest sprat abundances are located in the south-eastern North Sea in German and Dutch waters, and in the Irish Sea.
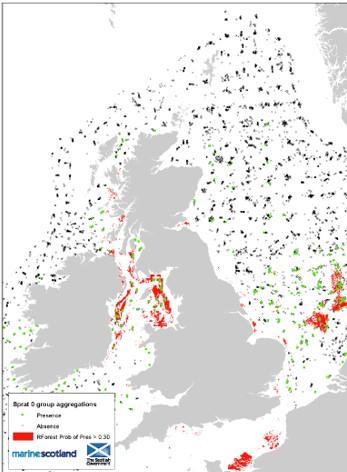
Figure 5 Modelled distribution of sprat around the UK. From Aires, González-Irusta and Watret (2014)
Click for a full description
Map illustrating modelled distribution of sprat in seas around the UK. Green dots represent sprat presence and black crosses represent sprat absence. Red polygons illustrate areas with a probability occurrence of more than 30% sprats. The highest sprat abundances are located in the south-eastern North Sea in German and Dutch waters, and in the Irish Sea.
Herring are widely distributed throughout UK waters (Figure 6), but juvenile herring (the only size class that most seabirds can swallow) have a more restricted distribution. In the west of Scotland, herring tend to be predominantly older fish (Heessen, Daan and Ellis, 2015). In the North Sea, adult herring tend to predominate in the northern North Sea while the highest abundance of juvenile herring is found in the south-eastern North Sea in German and Dutch waters (Aires, González-Irusta and Watret, 2014; Heessen, Daan and Ellis, 2015).
Distribution of herring around the UK. From Heessen, Daan and Ellis (2015). Warmer colours represent higher abundance within a particular sampling square.
Click for a full description
Map illustrating distribution of herring around the UK. Red dots represent sampling squares in the sea with the highest herring abundance, abundance in sample squares decreases in the order of orange, yellow, green and light blue dots, dark purple dots represent sampling squares in the sea with the lowest herring abundance. Herring are widely distributed throughout UK waters.
Depletion of sandeel
Since commissioning this report, the Scottish Government (2023) have published a full review of the evidence on the potential effects of sandeel fisheries management on the marine environment, including seabirds. This review supported a consultation on a proposal to close fishing for sandeel in Scottish waters. In 2024, a full closure prohibiting fishing for sandeel Scottish waters was implemented. In this case, the most beneficial appropriate adaptive response has been made and sandeel fisheries management is not considered further by this report.
Depletion of sprat or juvenile herring
Common terns breeding at Imperial Dock lock SPA in Leith, Firth of Forth, feed almost entirely on sprats while breeding (Jennings et al., 2012). The total number of breeding common terns in the Firth of Forth were reduced by about 40% when the Firth of Forth sprat stock collapsed in the early 1980s after heavy fishing pressure removed 88,000 tonnes from this small area (Fernandez et al., 2005; Jennings et al., 2012). When the fishery closed, the sprat stock rebuilt (Fernandez et al., 2005), as did common tern numbers (Jennings et al., 2012). Sprats are believed to represent a significant part of breeding kittiwake diet and to support high breeding success at small colonies of kittiwakes in the upper Firth of Forth (Lothian Ringing Group, pers. comm.).
During summer, sprats tend to be in shallow marine habitats influenced by freshwater inflows into the sea. Their distribution in the North Sea is predominantly southern, mainly south and east of the Dogger Bank, but spreads over much larger areas when the sprat stock increases in biomass (Heessen, Daan and Ellis, 2015; ICES, 2020). Similarly, sprat catches tend to focus on the highest sprat density areas in the southern North Sea, but catches are taken as far north as Shetland in years of high stock biomass, as in 2019 (ICES, 2020). Sprat is a species predicted to increase considerably in range and abundance as a result of warming sea temperature in UK waters/North Sea (Kjesbu et al., 2022) so may become a more important food for seabirds in future.
Appropriate adaptive responses
The evidence that depletion of sprat stocks affects seabirds in Scottish waters is less clear than for sandeels, but the example of common tern in the Firth of Forth suggests that reducing fishing pressure on coastal aggregations of sprats/juvenile herring would also benefit those seabirds that breed in these inshore areas where they feed more on sprats than on sandeels. Potential future dependency by seabirds on sprat/juvenile herring as their range and abundance increase should be accounted for in ecosystem-based management of these fisheries.
Appropriate actions could be closure of forage fish fisheries in Scottish waters, or the ecosystem-based management of those fisheries to set thresholds for the biomass of forage fish that must be maintained at levels appropriate for dependent predators (for example the ‘one-third for the birds’ threshold advocated by Cury et al., 2011) rather than allowing depletion to the much lower threshold of ‘Blim’, the biomass required to maintain future recruitment of the forage fish.
Another potential action relating to food shortage having an adverse impact on seabird breeding success is supplementary feeding of seabirds (chicks and/or adults). This action has been shown to be effective in several species such as Arctic skua, kittiwake and puffin.
In Shetland, supplementary feeding of Arctic skuas on Foula was introduced when their breeding success decreased considerably. Supplementary feeding resulted in higher attendance at breeding territories by adults (i.e. less time spent away searching for food), improved breeding success and higher return rates of adults the following breeding season (Davis, Nager and Furness, 2005). Subsequent supplementary feeding increased breeding success of supplemented Arctic skua pairs and resulted in a halt to the decline in breeding numbers (Gear, 2022).
Supplementary feeding of kittiwake chicks at a colony in Alaska where breeding success is food-limited has also been demonstrated to increase chick survival and fledging success (Gill, Hatch and Lanctot, 2002; White et al., 2010). However, whereas supplementary feeding at the Alaskan colony is from inside a building, with sliding hatches that allow food to be given to chicks without disturbance (Gill, Hatch and Lanctot, 2002), this mechanism would not be possible at a natural colony or at an artificial colony where such feeding hatches are not built into the structure. Furthermore, supplementary feeding of kittiwakes at a natural colony may result in scavenging birds such as corvids and large gulls being attracted into the colony which could reduce kittiwake breeding success. Supplementary feeding of kittiwake chicks would also be unlikely to be sustainable at the scale that may be required to achieve recovery of the Scottish population of this species.
Supplementary feeding of puffin chicks at St Kilda, the Isle of May, and at a colony in north Norway, showed that the work rate required of the parents was reduced when the chick was given food, which is likely to increase adult survival as well as benefit breeding success (Harris, 1978; Cook and Hamer, 1997; Wernham and Bryant, 1998; Dahl, Barrett and Ims, 2005; Harris and Wanless, 2011).
Bycatch in fisheries
Longline fishery
Impacts
Seabird bycatch on longlines occurs when scavenging seabirds attempt to steal bait off hooks that are being deployed on longlines (either pelagic or demersal) behind fishing vessels (Dunn and Steel, 2001). Once set, hooks are too deep in the water to catch scavenging seabirds. Very few birds are caught when longlines are being hauled back into the vessel as most baits have gone by then. Bycatch is therefore predominantly limited to seabirds that scavenge baits as the lines are being set.
Northridge, Kingston and Coram (2020) present estimates of seabird bycatch taken by UK fishing vessels in UK waters. Using these estimates, Miles, Parsons and O’Brien (2020) carried out an assessment of the likely impact of this bycatch on UK seabird populations. Almost all the sampling for seabird bycatch on longlines was in the northernmost UK waters, where there is demersal longline fishery that targets large and valuable deep-water fish such as hake, ling, torsk, large adult haddock and cod. Miles, Parsons and O’Brien (2020) estimated that the fishery may kill about 5,000 fulmar, 200 gannet, 56 great black-backed gull and 14 kittiwake per year. Although the sampling did not record any bycatch of great skua, it has previously been reported that some great skua mortality occurs on longline hooks (Furness, 1987; Mitchell et al., 2004), while Mitchell et al. (2004) also noted cases of Manx shearwater being taken on longlines, although that is probably mostly during the nonbreeding season outside UK waters.
Due to uncertainties associated with the estimates provided in the Northridge, Kingston and Coram (2020) paper (including limited to 103 longline hauls), a further analysis was presented by Kingston, Northridge, Paxton and Buratti (2023) to update these estimates. This paper estimated a mortality of approximately 1,000 to 2,000 fulmar, 50 to 150 gannet, and 10 to 20 great shearwater and great skua per year over the last decade. These latest figures provide more robust estimates for UK waters.
Bycatch does vary from vessel to vessel, seasonally, between years and differences in fishing effort (usually measured as numbers of hooks set), so there continues to be uncertainty around predicted impacts. It is clear however, that longline bycatch presents an important threat to some species of seabirds, particularly fulmar. Løkkeborg and Robertson (2002) reported a high bycatch of fulmars (but no other seabird species) in longline fishing for torsk in Norwegian waters in late summer (32 fulmars caught on longlines set over 12 days of fishing without deployment of any bycatch reduction actions, giving a bycatch rate of 0.55 fulmars per 1000 hooks set).
There is very little pelagic longline fishing in UK waters.
Appropriate adaptive responses
Globally, the most severe impact of longline bycatch on seabird populations results from bycatch of albatrosses (Pardo et al., 2017). To minimise that impact, numerous studies have investigated a range of potential mitigation actions to reduce fishery bycatch rates. Efficacy of these actions is well established globally, although predominantly in the context of bycatch of albatrosses and large petrels (ACAP 2019, Melvin et al., 2019). Deployment of bycatch reduction actions in the longline fishery has been demonstrated to be successful, not only in reducing the rate of bycatch of seabirds (Dunn and Steel 2001) but also in allowing recovery of populations that had been depleted by high levels of bycatch mortality (Dasnon et al., 2022).
Several studies with a focus on bycatch reduction actions of fulmars on demersal longlines have been carried out. Fulmars are attracted to fishing vessels to scavenge on offal and other discarded fishing waste, which means that the foraging distribution of fulmars is influenced by commercial fishing vessel presence and activity (Darby et al., 2021). Preventing discharge of offal and other fishery waste at sea would help to reduce attraction of seabirds such as fulmars, gannets and gulls to fishing vessels, as has been shown for gannets (Clark et al., 2020) and could help to reduce longline bycatch. Deployment of a bird-scaring streamer line as a single action in Alaska was also showed to significantly reduced non-albatross seabird (mostly fulmar) bycatch by 77-90% (Melvin et al., 2019). However, setting only at night reduced bycatch of some species, but increased bycatch of fulmars (Melvin et al., 2019), so would be unsuitable in a fishery where fulmar bycatch is significant. Best practice is considered to be use of a combination of reduction actions that are most appropriate and practical for the particular fishery and bycatch seabird species concerned (Løkkeborg, 2011).
There are several effective methods to reduce seabird bycatch. There are no requirements for longline vessels fishing in UK waters to reduce bycatch of seabirds, although voluntary management actions are practiced by some UK vessels (Kingston et al., 2023). Results of a questionnaire circulated to skippers in the Scottish longline fleet indicated that approaches that were favourable to industry included the use of bird-scaring lines; offal management approaches; night setting; and, the deterrence of birds from the line hauling area (Kingston et al., 2023).
Bycatch continues to be monitored in Scottish waters in this fleet under the UK Bycatch Monitoring Programme (BMP). Total observer sampling effort in the fishery over the last decade equates to about 0.5% of the annual UK fishing effort in a typical year, although there has been a significant increase in observer monitoring in recent years. The evidence suggests that there would be scope to reduce bycatch of fulmars, gannets, great black-backed gulls and great skuas in longline fisheries in northern Scottish waters, although numbers may be much smaller than in some overseas longline fisheries to which Scottish seabirds may be exposed, especially during the nonbreeding season.
Midwater trawl bycatch
Impacts
Midwater trawls (pelagic net fishing) in UK waters are thought to take around 500 common guillemots, 60 razorbills and 50 cormorants per year based on an extrapolation of data from sampling 2,239 hauls by UK vessels in UK waters (Northridge, Kingston and Coram, 2020). While much of these observed hauls were from Scottish waters, all of the seabird bycatch occurred off the south coast of south-west England with no seabird bycatch in the hauls sampled from Scottish waters. This suggests that the impact of this fishery in Scottish waters is probably very small, and possibly negligible, although these species may be at risk of bycatch in wider UK and overseas waters during the nonbreeding season.
Christensen-Dalsgaard et al. (2022) found several clusters of tens of drowned gulls on beaches in Norway which led to observing seabird bycatch in purse seine nets used by fisheries targeting herring along the Norwegian coast. They found that 10% of 91 coastal purse-seine fishing events led to bycatch of seabirds, with a total of 32 bycaught seabirds (0.36 birds per haul). These included 13 common gulls, nine herring gulls, six great black-backed gulls and four gulls not identified to species. There is very limited purse seine fishery in Scottish waters (approximately two to three weeks a year with a handful of vessels) and observations to date through the BMP have recorded no seabird bycatch.
Appropriate adaptive responses
Based on the available evidence in Northridge, Kingston and Coram (2020) and Miles, Parsons and O’Brien (2020) it would appear adaptive responses in relation to reducing seabird bycatch from midwater trawl fisheries in Scottish waters is not required.
Set net bycatch
Impacts
Based on sampling from 18,916 hauls of set nets by UK fishing vessels, scaled up to the entire fishery, set nets are estimated to take a bycatch each year of 1,500 common guillemots, 215 cormorants, 120 gannets, 70 fulmars, 70 razorbills, 25 shags and 20 herring gulls (Northridge, Kingston and Coram, 2020). In relation to the total UK population of each species, these numbers represent an additional 9% to normal natural mortality of adult cormorants, 1.3% for common guillemot, 0.5% for shag, 0.3% for fulmar, gannet and razorbill (Miles, Parsons and O’Brien, 2020). However, almost all of this bycatch occurred in English waters (Bradbury et al., 2017; Northridge, Kingston and Coram, 2020); with a small hot-spot for bycatch in set nets occurring north of Shetland where gannets, fulmars and gulls were taken (Northridge, Kingston and Coram, 2020). There was no recorded bycatch of guillemots, razorbills, cormorants or shags in set nets in Scottish waters, but data from inshore fisheries is largely unavailable. Cleasby, Wilson and Davies (2022) also investigated bycatch of seabirds in static nets (gillnet fisheries) around the UK. They concluded (p. 157) that “lack of fishing effort data for smaller vessels made it difficult to assess seabird-fishery overlap in Scottish waters”. Whilst there is uncertainty at the level of mortality risk presented by set nets in Scottish waters, evidence from elsewhere indicates that set net bycatch can present an additional pressure on populations. Dierschke, Christensen-Dalsgaard and Koschinski (2022) listed guillemot as a species involved in bycatch in nets set in the Celtic Seas, which includes waters off the west of Scotland. Bærum et al. (2019) estimated an annual bycatch in the Norwegian coastal gillnet fishery of 1,580 to 11,500 seabirds per year, with fulmar being the most numerous seabird in the bycatch sample and common guillemot the most numerous of the diving seabirds.
Set nets are also used by the aquaculture industry in Scottish waters, generally as predator exclusion netting around and over marine salmon cages, but were not sampled by Northridge, Kingston and Coram (2020) or Miles, Parsons and O’Brien (2020). There do not appear to be detailed studies of bycatch of seabirds in aquaculture set nets. However, there have been individual cases of small numbers of gannets (tens of individuals) becoming entangled in netting covering salmon cages in different parts of Scotland and different seasons, especially where top nets use a mesh of >200 mm suspended from a central pole (NatureScot, 2020). Both the top and side nets can also trap or entangle small or moderate numbers of gulls and possibly a few great skuas, cormorants, and shags. In the sub-sea nets auks have also been recorded as bycatch.
Appropriate adaptive responses
Monitoring is required in Scottish waters to improve understanding on the potential for impacts of bycatch on seabirds. However, based on evidence from elsewhere, implementing site-based set net bycatch reduction measures in seabird foraging areas particularly during the breeding season may considerably reduce bycatch mortality.
At Filey Bay off the east coast of Yorkshire, there was a hotspot of common guillemot and razorbill bycatch in set nets for salmon and sea trout, with estimated bycatch mortality of 200 guillemots and 323 razorbills in 2008 as well as 186 guillemots and 277 razorbills in 2009 (Quayle, 2015). After bycatch reduction actions were introduced in 2010, the bycatch mortality was reduced to an average of 11 guillemots and 43 razorbills per year in 2010-2014 (Quayle, 2015). This was achieved by establishing local by-law regulations requiring fishers to only set nets during the day, to have high visibility corline netting in the leader/tailpiece of the net, restricting use of monofilament to 70 m in length or less, to remain at their nets in order to remove any birds that became entangled and release these alive wherever possible and to report bycatch mortality. It is uncertain which of these different individual actions was most effective in reducing bycatch. Cleasby, Wilson and Davies (2022) found that guillemots and razorbills rarely foraged at night during the breeding season, but that these species showed peak foraging activity around sunrise and sunset. This suggests that the restriction of gillnet fishing to between 5am to 9pm in June used at Filey Bay may not have been especially effective in reducing the risk of bycatch – it is more likely that the reduced bycatch in this location was achieved through the use of high visibility corline and the attendance of fishers at nets releasing any birds that became entangled. Results presented by Quayle (2015) demonstrate that bycatch of common guillemots and razorbills can be reduced considerably by such methods.
Based on their experience in Norway, Bærum et al. (2019) suggested that seasonal closures of the gillnet fishery could be effective as a means of reducing bycatch, as the current rates of bycatch show strong seasonal variation. O’Keefe et al. (2023) reviewed evidence about actions that can reduce bycatch, advocating gear switching to avoid use of set net designs that cause high levels of seabird bycatch, but also advocating using seasonal closures to avoid periods when seabirds are most at risk at particular set net fishing locations, and the possible use of visual and acoustic deterrents. Rouxel et al. (2021) showed that long-tailed ducks may avoid buoys with large ‘looming’ eyes painted on them, which could reduce bycatch. Ørsted (2022) has recently investigated this method for reducing bycatch of auks and reported on the results of the first year of its bycatch reduction technology selection phase, undertaken during the 2021/2022 nonbreeding season. This has provided evidence that the use of looming eye buoys can reduce auk bycatch in active fisheries (trials undertaken along the south coast of the UK). Based on the first year of data, Ørsted (2022, p. 22/25) is “confident that the LEB [looming eye buoys] can be implemented as a compensation measure within active gillnet fisheries to compensate for impacts to guillemot and razorbill”. A second season of testing this approach, funded by Ørsted, has been underway off the south coast of England from September 2022 to March 2023 and should report on those results during 2024.
Field et al. (2019) tested flashing white Light-Emitting Diode (LED) lights, green LED lights and high contrast monochrome net panels as deterrents, but found no reduction in bycatch of sea ducks when these were used and an increase in bycatch in nets with added flashing white LED lights. These studies suggest that there are no visual or acoustic deterrents that can currently be identified as likely to give strong reductions in seabird bycatch in set nets, so further research on conservation actions for seabirds in this area would be desirable. Cleasby, Wilson and Davies (2022) suggest that bycatch reduction might be achieved by closing areas of high auk abundance to gillnet fisheries as well as areas where water depth is shallow (>100 m), and by deployment of above-water deterrents (e.g. Rouxel et al., 2021). Above-water deterrents are typically more visible under low light conditions such as around peak foraging periods of sunrise and sunset than are underwater deterrents which may be relatively ineffective (e.g. Field et al., 2019).
Possibly the most effective action to reduce guillemot and razorbill bycatch mortality may be the training of fishers to release birds that become tangled in their nets (Quayle, 2015).
Fishery discards
Impacts
There is broad agreement that seabird species which benefit most from scavenging on discards and offal behind fishing vessels in the waters around the British Isles are fulmars, gannets, great skuas, great black-backed gulls, herring gulls, lesser black-backed gulls and kittiwakes (Hudson and Furness, 1988; Furness, Ensor and Hudson, 1992; Garthe, Camphuysen and Furness, 1996; Mitchell et al., 2004; Forrester et al., 2007; Sherley et al., 2020). Although there is evidence that discards form an important part of the diet of great skuas during the breeding season, most of these scavenging seabirds feed more on fishery waste in the nonbreeding season than they do while breeding. In the case of great skua, reductions in the amount of fishery waste being discarded has caused a diet change in this species (Votier et al., 2004; Church et al., 2018), which has had impacts on other seabird species, as great skua has become more predatory (Votier et al., 2004). Coulson (2015) suggested that fishery discards were likely to have been much more important in supporting the increase of herring gull numbers in Britain and Ireland than were opportunities to scavenge at landfill sites.
Reduced amounts being discarded by fisheries as a consequence of the European Union (EU) Common Fisheries Policy landings obligation have reduced food supply to scavenging seabirds. Sherley et al. (2020) estimated that discards could have supported about three million scavenging seabirds in the North Sea in 2010, which is two million fewer than in 1990. The decrease is also geographically variable. Discard amounts have decreased more in Scottish waters than in the south-eastern North Sea, because beam trawls, found predominantly in the south-eastern North Sea, still produce large amounts of discards, whereas technical actions have reduced bycatch of undersized fish in Scottish demersal trawls for whitefish and in the Nephrops fishery.
Reduced availability of discards may have reduced breeding success of great skua, and possibly of great black-backed gull at some colonies, and has increased predation rates by great skuas, and possibly by great black-backed gulls on smaller seabirds. However, the largest impact may be on scavenging seabirds at sea, with decreases in numbers in the North Sea in winter.
Appropriate adaptive responses
Reducing discards, preferably to zero, is a sensible fishery policy. The current policy in Scottish waters is to reduce unnecessary discards as part of the Future Catching Policy to reduce waste and improve the sustainability of fish stocks. However, although decreased from levels seen in previous decades, discards throughout the North Sea still support very large numbers of scavenging seabirds (Sherley et al., 2020). Discards also attract scavenging seabirds to fishing vessels thereby increasing the risk of seabird bycatch in fisheries.
Management should aim to develop actions that minimise impacts of the withdrawal of this artificial food supply from scavenging seabirds.
Mammalian predators
Impacts
Many seabirds breed predominantly on offshore islands that lack terrestrial mammalian predators. Seabirds nest in a variety of habitats including on the ground in the open, under boulders or in burrows. These seabirds are very vulnerable to mammalian predation if invasive non-native predators manage to establish a population on a remote seabird island (Jones et al., 2008). Invasive non-native predators include brown (Norway) rat, black (ship) rat, house mouse, feral cat, feral ferret, and American mink. In the context of seabird islands that have been free from terrestrial mammals, mammalian predators can also include native predators such as hedgehog, fox, stoat, weasel and mice. These predators may consume eggs and chicks of ground-nesting and burrow-nesting seabirds, and in some cases, they may take adult seabirds too. For example, mice have been found to have a small impact on Manx shearwater breeding success at Rum (Lambert et al., 2021) and have famously attacked and eaten albatross chicks on some sub-Antarctic islands. Herbivores such as deer, sheep and rabbits can also be problematic in seabird colonies on islands. There are records of deer and sheep killing seabird chicks in order to eat their bones for their mineral content and records of rabbits altering the soil stability and ground vegetation which damages areas used by burrow-nesting seabirds.
Mitchell et al. (2004) identified mammalian predators as having a strong influence on breeding distribution and populations in Britain and Ireland of Manx shearwater, European storm-petrel, Leach’s storm-petrel, Sandwich tern, common tern, little tern, black guillemot and puffin, a moderate influence for shag, lesser black-backed gull, herring gull, roseate tern, Arctic tern and razorbill, and a slight influence on great black-backed gull. Forrester et al. (2007) came to a similar conclusion in the context of populations in Scotland, but listed fulmar, cormorant and common guillemot as vulnerable at some Scottish colonies where they nest on the ground rather than just on sea cliffs or isolated sea stacks. Many Scottish islands have populations of invasive non-native mammalian predators, especially rats. Some of these have been present for over 100 years, and there is a tendency to take this for granted because the mammal-free condition was so long ago that there is little or no understanding of the ecological situation before colonisation by invasive mammals. For example, an estimated 92% of the land area of the archipelagos of Orkney and Shetland comprises islands with rats present and European storm-petrels now breed almost exclusively on the small area of rat-free islands (de Leon et al., 2006). Historically, European storm-petrels did breed much more widely on islands in these archipelagos before they were eradicated by rats and other mammalian predators, but that was so long ago that the much wider distribution is long forgotten. De Leon et al. (2006, p. 64) concluded that “the presence or absence of rats is the single most important influence on storm-petrel breeding distribution in Orkney and Shetland”.
In a few situations, seabirds have apparently managed to coexist with invasive non-native mammalian predators. These cases might suggest that the impact of invasive non-native mammalian predators can be exaggerated, but the exceptions are interesting. On Rum, the largest colony of Manx shearwaters coexists with brown rats and mice. The shearwaters nest only at high altitude on the slopes of the Rum mountains, whereas rats mainly occur at the coast. Although rats do visit the shearwater colonies and will scavenge on abandoned eggs, their predation impact seems to be slight – though localised events probably occur. The numbers of rats at the shearwater colonies appear to be very small, and probably the cold winter conditions prevent rats from maintaining a population in the colony area. This contrasts with numerous islands, including adjacent island of Canna where Manx shearwater populations have been eliminated or reduced to very small numbers by invasive non-native mammalian predators, where the shearwater colony is not spatially separated from rat habitat (e.g. Booker et al., 2019). Further work would be required to gain a better understanding of rat distribution in relation to current and potential Manx shearwater breeding sites and enable a whole-island assessment of the risk from rat predation.
Appropriate adaptive responses
Eradication of invasive non-native mammalian predators on seabird islands.
The eradication of invasive non-native mammalian predators is well established as a conservation tool. Appropriate methods have been described in detail, and the successes (and occasional failures) have been reviewed in the literature many times (Courchamp, Chapuis and Pascal, 2003; Jones et al., 2016). In Europe, 90 islands were successfully cleared of one or more invasive non-native mammalian predators in the 25 years up to 2016 (DIISE, 2016). However, until recently there were still many islands in the UK where invasive non-native mammalian predators (especially brown rats) have strong negative effects on seabirds. Brooke et al. (2007), Ratcliffe et al. (2009) and Stanbury et al. (2017) developed lists of top priorities for eradication of invasive non-native mammalian predators from islands to conserve vulnerable seabirds. Setting priorities is sensible in terms of obtaining the greatest gain from available finite resources and also to prioritise sites where recolonisation by invasive non-native mammalian predators is relatively low risk. Some successful eradications have been nullified by swift recolonisation by the invasive non-native mammalian predator from nearby populations. For example, brown rats were eradicated from Handa, northwest Scotland, but reinvaded about ten years later (Stanbury et al., 2016). Brooke et al. (2007) considered global seabirds with an emphasis on rare endemic species at risk from invasive non-native mammalian predators. Ratcliffe et al. (2009) considered petrels and shearwaters in the UK, Channel Islands and Isle of Man. Stanbury et al. (2017) considered seabirds in the UK and crown dependencies.
The ranking of colonies by Stanbury et al. (2017) identified 22 colonies in Scotland within the 25 top priority sites in the UK and crown dependencies. Although invasive non-native mammalian predator eradications have been carried out already at some of those sites, there is clearly much scope for further removal of invasive non-native mammalian predators from Scottish islands, and there may be a case for control of mammalian predator impacts on seabirds on the Scottish mainland, for example by protecting vulnerable ground-nesting seabirds with predator-proof fences (Young et al., 2012, Short, 2020). Mitchell et al. (2018) provide an online database of UK island seabird colonies where mammalian predators are present.
In the UK, some examples of using electric fences to exclude foxes from colonies have been successful, but electric fences are not fully effective in excluding predators and require regular maintenance (White and Hirons, 2019; Short, 2020). Use of an electric fence greatly improved breeding success and numbers of Sandwich terns at Forvie National Nature Reserve (NNR), Aberdeenshire, but required daily checks because foxes managed to cross the fence line on average several times per year so needed to be cleared out of the protected area (Short, 2020).
A more expensive, but more effective, alternative is the use of predator-proof fences, as deployed in Hawaii at Ka’ ena Point Natural Area Reserve (Young et al., 2012). These 2m tall fences were set up in November 2010 to February 2011 around 20ha of coastal habitat within Ka’ ena Point to prevent predators (including dogs, cats, mongooses, rats and mice) from entering the protected area. Predators were eradicated within the enclosed 20ha – it took three months to complete for all predators except mice, which were eradicated within an additional six months. This was the first predator proof fence constructed in the United States at the time of its completion (Young et al., 2012). Such completely predator-proof fencing may be unnecessary to protect colonies just from foxes, but it might be especially appropriate for colonies subject to predation by rats or mink as well as by foxes. Cooper (2013) lists a further ten examples of successful deployment of predator-proof fencing around seabird colonies in New Zealand, Hawaii (USA) and Azores (Portugal), and these are also reviewed in detail by White and Hirons (2019).
Eradication of invasive non-native mammalian predators has been proposed as a compensation action for impacts of offshore wind farm developments (e.g. Skeate, 2022). Ratcliffe et al. (2009) assumed a cost of eradication of rats at £440 per hectare, based on the cost of eradication at Canna, but these costs will vary considerably depending on the site and a variety of other factors. The cost of eradicating rats from the Isles of Scilly was estimated at £5 million over 5 years and the subsequent biosecurity programme for this and other sites in England at £250,000 (Lock et al., 2022). Deployment of predator-proof fence to exclude mammalian predators from a six hectare area of lesser black-backed gull nesting habitat at Orfordness has recently been implemented as a compensation action for impacts of offshore wind farms Norfolk Vanguard, Norfolk Boreas, East Anglia ONE North, and East Anglia TWO (MacArthur Green and Royal Haskoning DHV, 2022).
An essential part of invasive non-native mammalian predator eradication from seabird islands is the development of effective biosecurity actions to minimise the risk of the mammalian predator re-establishing its population on the island by recolonisation from elsewhere (Bell et al., 2019; Bambini and Varnham, 2020). Biosecurity planning should include awareness raising, prevention measures on key pathways, on-island surveillance, and rapid incursion response plans.
Avian predators
Impacts
The impact of avian predators on seabirds is generally considered to be less than the impact of mammals and is thought often to be a sustainable impact. However, some avian predation impacts can be exacerbated particularly where the predator population has been affected by human activities. Large gull populations for instance increased considerably due to provision of refuse and fishery discards, and in some circumstances, are recognised as having impacts on smaller seabirds (Scopel and Diamond, 2017). For example, great black-backed gulls can feed on adult storm-petrels, Manx shearwaters or puffins. Large gulls can steal fish from terns and take tern eggs and chicks. Large gulls can take eggs and chicks from kittiwake nests. Great skuas deprived of forage fish and discards switch to killing adult seabirds and taking seabird eggs and chicks. These impacts have the potential to cause the decline of colonies of smaller seabirds. Corvids and birds of prey can also have impacts on seabird populations.
Mitchell et al. (2004) provided evidence that strong effects occurred on populations of Leach’s storm-petrel, Arctic skua and little tern, moderate effects on European storm-petrel, kittiwake, Sandwich tern, roseate tern, common tern and puffin and slight effects on fulmar, Manx shearwater, shag, great skua, lesser black-backed gull, great black-backed gull, herring gull, Arctic tern, common guillemot, razorbill and black guillemot. Forrester et al. (2007) provided a shorter but predominantly similar list in a Scottish context, and listed Arctic tern as exposed to moderate rather than slight effects. Black guillemots seem to be particularly vulnerable to both mammal and avian predators at their nests and seem to be limited in their breeding distribution and numbers by a lack of safe nest sites. Nests have been predated by herring gulls at some colonies (BTO, 2023) and by hooded crows, gulls and great skuas at others (Johnston et al., 2019).
Impact levels are difficult to classify for this particular pressure because they tend to be site-specific. Particular avian predators may be present at some colonies, but not at others, and the impact may vary depending on particular features of the colony. Topography may allow avian predators to access nests at some colonies, but not at others. For example, as kittiwake numbers declined in Shetland after the collapse of the Shetland sandeel stock, rates of predation by great skuas increased, but some of the remaining kittiwakes moved into sea caves to nest where they are much less at risk from great skuas. Many kittiwake colonies on open cliff faces were completely deserted.
The recent increase in the white-tailed eagle population in Scotland may represent a threat to some seabird colonies. White-tailed eagles in Norway have caused major disruption to some seabird colonies, even being considered responsible for the decline in breeding numbers of kittiwakes and gannets at some sites in Norway (Barrett, Strøm and Melnikov, 2017; Anker-Nilssen, Fayet and Aarvak, 2023). White-tailed eagles take adult auks, gulls and fulmars and cause panic flights at seabird colonies resulting in eggs and chicks being exposed to other potential predators such as crows and large gulls. It remains to be seen how seabird populations in Scotland will respond to further recovery of the white-tailed eagle population around Scotland, but this may involve some redistribution of seabird colonies into locations less vulnerable to eagle attack and disturbance.
Appropriate adaptive responses
Appropriate adaptive responses are likely to be species-specific and also site-specific because impacts are strongly site-specific.
Removal, or diversionary feeding, of avian predators
In many cases, avian predators are themselves protected species of conservation concern. Killing avian predators to reduce impacts on vulnerable seabirds may be undesirable but can be an effective strategy in some cases (Scopel and Diamond, 2017). Killing avian predators may often also be ineffective if the removed predator is replaced by another. However, many avian predators are predation specialists, taking a particular prey type but ignoring other potential prey. Where a seabird colony is within the foraging range of a particular predator, it may be ‘protected’ by the presence of that predator if the predator does not specialise on the seabird as prey. For example, great skuas are highly territorial and exclude other great skuas from their territory. A great skua that feeds only on fish may hold a territory adjacent to a black guillemot colony and represent no threat to the black guillemots. However, a great skua that specialises in killing black guillemots as they return to their nest can wipe out an entire black guillemot colony (Furness, 1987). Similarly, an individual great black-backed gull may specialise in killing storm-petrels, or puffins. Removal of specialist avian predators that have impacts on particular seabird species could lead to the territory being taken by a new pair of avian predators with a different feeding habit, so removing that predation impact on a particular colony of seabirds. Removal of a small number of specialist individuals has been demonstrated to greatly reduce impacts of large gulls on storm-petrels (Sanz-Aguilar et al., 2009). Similarly, Westerberg et al. (2019) found that only a few great black-backed gulls at Skokholm were responsible for most of the predation on Manx shearwaters at that colony - many of the other great black-backed gulls did not have any impact on the shearwaters.
Diversionary feeding of predators to reduce predation impact on puffins, guillemots, razorbills and kittiwakes was evaluated as a possibility by Skeate (2022). However, diversionary feeding of avian predators is not considered further in this review because the scientific evidence for this being a successful adaptive response is inadequate and there would need to be a large body of evidence collected on the efficacy and sustainability of that approach before it could be recommended. For further review see Skeate (2022).
Supplementary feeding of Arctic skuas
Great skuas kill and eat large numbers of Arctic skua chicks at some colonies where the two species nest side by side (Furness, 1987). Arctic skua chicks are vulnerable at all times, but newly fledged Arctic skua chicks are especially vulnerable to attack. Chicks are vigorously protected by their parents. When food is scarce, parents spend a lot of time away from the territory searching for food for their large chicks. At such times, the risk of fledglings being killed by great skuas increases because only one parent (or neither) will be on territory to defend their chick. Supplementary feeding of Arctic skuas allows adults to spend more time on the territory and as a result, the rate of killing of Arctic skua chicks by great skuas is reduced. Supplementary feeding leads to an increase in Arctic skua chick survival, and also improves the body condition of adult Arctic skuas so increases their return rate the following season (Davis, Nager and Furness, 2005). Supplementary feeding therefore potentially reduces impacts of great skuas on colonies of Arctic skuas and can help reduce impacts on breeding numbers (Gear, 2022). Similar supplementary feeding of great skuas would be possible if extreme pressures reduced their numbers drastically but would be inappropriate if the population remains moderately large.
Tern terraces; nest boxes/chick shelters
Nest boxes/chick shelters provide protection for nesting terns from adverse weather conditions as well as from avian predator attack (Steel and Outram, 2020). First developed for roseate terns, these nest boxes are also used by breeding common terns and Arctic terns and are used as shelters by Sandwich terns. Chicks of all tern species apart from little tern use chick shelters. These are considered by the RSPB to represent best practice for protecting nesting terns from avian predators (Babcock and Booth, 2020a).
Bamboo canes around tern nests
Bamboo canes placed vertically in the ground around tern nests reduce the frequency of attack by large gulls and therefore can provide partial protection for terns against avian predation (Boothby, Redfern and Schroeder, 2019). These are considered by RSPB to represent best practice for protecting nesting terns from avian predators (Babcock and Booth, 2020b).
Nest boxes
Black guillemots readily use artificial nest sites (ANS) which can be purpose-designed nest boxes (as deployed at various locations such as Carrickfergus waterfront) or holes or drain-pipes in walls (as in the seafront wall at Oban). Provision of nest boxes can allow black guillemot populations to increase, as seen at Copeland Bird Observatory, Strangford Lough, and Bangor Marina, because the main limiting factor for these birds seems to be safe nesting places (BTO, 2023). Nest boxes provide sites that are safe from most potential predators.
Nest boxes are used by European storm-petrels and Leach’s petrel and may increase breeding success by making nest sites more secure from predators.
Marine renewables collision with turbine blades
Impacts
The marine energy industry has grown in recent years, with more, larger-scale offshore wind farms being deployed as well as tidal turbines and wave energy converters. Consequently, studies assessing impacts to seabird populations are emerging but still relatively new. Recognising the lack of empirical evidence, Furness et al. (2013) computed a theoretical index of collision risk of seabird species based on the overlap of their at-sea flight height distribution with the rotor-swept area of turbines, their flight agility, the proportion of time spent flying at sea, the amount of nocturnal flight activity, and the conservation importance of the species. Classifying species with risk score >500 identified gannet, lesser black-backed gull, herring gull, great black-backed gull and kittiwake as high risk, scores of 101-499 as moderate risk included cormorant, shag, Arctic skua, great skua, Sandwich tern, roseate tern, common tern, Arctic tern and little tern, scores <100 as low risk included fulmar, Manx shearwater, European storm-petrel, Leach’s storm-petrel, common guillemot, razorbill, black guillemot and puffin.
Empirical evidence gathered since has broadly supported Furness et al. (2013) classifications (McGregor, 2022a). Cumulative impact assessments have suggested that the highest population-level impacts of offshore wind farm collision mortality in Scottish North Sea waters would be likely to occur with kittiwake, herring gull, lesser black-backed gull and great black-backed gull (Tjørnløv et al., 2022). Collision risk for gannets has also been assessed as potentially high, particularly in Scotland where research has showed that the greatest risk is to females during the chick rearing period (Lane et al., 2020). There is recent evidence of high macro-avoidance of offshore wind farms by non-breeding gannets (Natural England, 2022, Tjørnløv et al., 2022) which has suggested that collision risk might be less for that species than previously thought (and conversely that displacement risk could be higher). Risk for coastally distributed seabirds is now thought to be relatively low, as most offshore wind development has moved to deeper waters, especially with the development of floating turbines. However, there remains potential for impacts as a result of wet storage requirements. This reduces perceived risk for coastal species including herring gull, terns, cormorant and shag which spend relatively little time far away from land. However, new developments of offshore wind in northern and western Scotland may bring a greater focus on seabird species that are scarce in English waters and off east Scotland, where offshore wind has mostly been developed up until now. That could include Arctic skua, great skua, and possibly risk to shearwaters and storm-petrels if attracted towards hazard warning lights at offshore wind farms (Deakin et al., 2022).
Wave energy converters seem to represent little threat to seabirds, although this type of development could cause some displacement of seabirds and could potentially alter habitat through reducing wave action (Grecian et al., 2010). Tidal turbines have the potential not only to displace seabirds and to alter habitat, but also to cause mortality through collision (Furness et al., 2012). However, seabird activity in areas of high tidal flow tends to be low, so that the overlap between seabird foraging and the very few locations where tidal flow is strong enough to power tidal flow turbines is likely to be small (Johnston et al., 2021a).
Appropriate adaptive responses
Offshore wind farm developers already include much mitigation in their assessments to reduce potential impacts on seabird populations, including increasing the gap between the sea surface and the lowest point of the rotor-swept area because most seabird flight activity is relatively close to the sea surface. However, some adaptive responses may be possible.
It would be pragmatic to strategically select sites for offshore wind farm development away from foraging hot-spots used by seabirds such as frontal systems where prey tend to be aggregated, or over sandbanks that support high densities of sandeels, and as distant as practicable from major seabird colonies accommodating species considered to be highest collision risk.
Recent research suggests that painting turbine blades with high contrast bands (black and white) is likely to make these more visible to seabirds so may reduce collision risk (May et al., 2020, Martin and Banks, 2023). This possibility is now being tested on offshore turbines, but results are not yet available.
Where impacts from offshore wind farms have an adverse effect on the integrity of SPA features (i.e. seabirds), there is a requirement for compensation to be delivered if the development is approved. Several offshore wind farm developers are now delivering compensation on this basis for predicted impacts of offshore wind farms in English waters; the amount of compensation is likely to increase in the future since it is the contribution of each development to the in-combination total impact that requires compensation. Compensation measures which are currently in place include predator-proof fencing to exclude mammal predators from lesser black-backed gull nesting habitat and construction of artificial nesting ledges for kittiwakes. Current development plans are considering compensation measures such as eradication of mammal predators from islands and restoring historic colonies. Such compensation would be more effective if coordinated and strategic across multiple projects.
Disturbance and displacement
Impacts
Disturbance at colonies
Terns, especially Sandwich terns and little terns, are highly sensitive to impacts of human disturbance near to their nest sites (e.g. Short, 2020). Many breeding sites of Sandwich tern and little tern have been lost as a consequence of human disturbance (Mitchell et al., 2004, Forrester et al., 2007).
There can be particular cases of human disturbance having a negative impact on seabird species that appear to be highly tolerant of human activity. For example, kittiwakes nest on buildings in many coastal towns in Norway, Faroes, Scotland, England and France. In most cases there seem to be no adverse effects of human disturbance and breeding success in urban colonies tends to be higher than in natural colonies (Christensen-Dalsgaard et al., 2019). However, there is some evidence that human disturbance has reduced breeding numbers of kittiwakes in part of the colony at Dunbar Harbour (Skeate, 2022).
Offshore wind farm displacement
Dierschke, Furness and Garthe (2016) reviewed evidence for displacement of seabirds by offshore wind farm structures (avoidance of offshore wind farms). They listed gannet as showing strong avoidance/displacement, fulmar, Manx shearwater, Sandwich tern, common guillemot and razorbill as showing moderate avoidance/displacement, and cormorant, shag, lesser black-backed gull, herring gull and great black-backed gull showing some evidence of attraction to offshore wind farms. This attraction was strongest for cormorant, with offshore wind farms providing new opportunities for cormorants to extend their foraging activity further offshore by using wind farm structures to roost and dry plumage between foraging bouts. There was too little data to allow assessment of possible displacement or attraction of storm-petrels, skuas, roseate tern, little tern, black guillemot or puffin.
Displacement of auks from offshore wind farms seems to vary among sites (Dierschke et al., 2016). Some report no change in auk densities at an offshore wind farm between baseline data and operational data (Dierschke et al., 2016; Trinder, 2023) or even an increase within a wind farm area (Degraer et al., 2023). At other offshore wind farms displacement may result in auk density reduced by 40 to 70% (Dierschke et al., 2016, Peschko et al., 2020). The cost of displacement to auk populations is also uncertain. There may be no cost if feeding conditions for auks are good, or displacement might reduce fitness if auks are already struggling to find food. It seems highly likely that impacts of displacement would be greater when auks are constrained by food shortage. Analysis of guillemot energy budgets through the nonbreeding season in the west of Scotland and in the east of Scotland (Buckingham et al., 2023) suggests that energy costs peak in late winter/early spring (late February/early March). Energy costs were lower than for the same species wintering off eastern Canada, suggesting some scope for increasing effort if necessary. However, birds in Canada may be able to sustain high work rates because there is high availability of food. It is unclear whether Scottish guillemots have adequate forage fish availability to allow high energy expenditure.
Disturbance/displacement by shipping
Disturbance to seabirds at sea can include disturbance by vessels (Schwemmer et al., 2011; Fliessbach et al., 2019) and resulting habitat loss through displacement away from shipping lanes (Dierschke and Mercker, 2022). Diving seabirds may dive or take off, non-diving seabirds may take off as a ship approaches, adding to their energy expenditure and potentially reducing foraging activity. However, the costs of disturbance by ships appears to be small for most seabird species in UK waters, with most seabirds only showing avoidance behaviour at short distances and for a short period. Fliessbach et al. (2019) derived a disturbance vulnerability index in relation to ship approach and ranked black guillemot, razorbill, common guillemot, cormorant and gannet as among the seabirds more vulnerable to this kind of disturbance. In contrast, some seabirds, such as terns and gulls, may benefit from the turbulence caused by ship propellers, bringing prey to the sea surface and allowing them to feed close behind passing ships.
Appropriate adaptive responses
Disturbance by people can be countered by signage, by re-routing of paths, and by seasonal funding wardens/rangers to meet people who are about to disturb nesting seabirds.
Strategic siting of new offshore wind farm developments away from foraging hot-spots used by seabirds such as frontal systems where prey tend to be aggregated, or over sandbanks that support high densities of sandeels, and as distant as practicable from major seabird colonies accommodating species considered to be at highest risk of displacement.
Compensation measures as outlined in the appropriate adaptive responses for marine renewables collision section above will also be relevant where disturbance and displacement impacts from offshore wind farms have an adverse effect on the integrity of SPA features (i.e. seabirds).
Artificial light pollution
Impacts
In their literature review, Deakin et al. (2022) highlight that there is abundant evidence for light-induced disorientation in shearwaters and petrels with observers reporting that once attracted to the vicinity (i.e. within several tens of metres) of a powerful light source, birds seem unable to escape and may end up flying around for several hours. Potential effects are described as “Sub-lethal affects that may influence survival in the longer term, or the ability to rear young, could accrue from the wasteful expenditure of energy in circling flight for protracted periods. This may lead to loss of body condition resulting in birds becoming more vulnerable to starvation or predation. Flight costs of European storm petrels have been estimated at 3.9 times basal metabolic rate (Bolton, 1996), close to the maximum sustainable work rate (Drent and Daan, 1980). Prolonged periods of flight, without opportunity to feed or rest, may lead to dehydration or exhaustion of birds that escape collision”.
Attraction to lights was noted by Mitchell et al. (2004) as a slight threat to Manx shearwater. Fledglings can be grounded by artificial lights, and this is known to occur at Kinloch on Rum and at Mallaig, the nearest town to Rum (Syposz et al., 2018), although the numbers of fledglings grounded are relatively small in relation to the very large size of this population. Manx shearwater, European storm-petrel, Leach’s storm-petrel and puffin fledglings are known to be grounded by artificial lights in Village Bay, St Kilda, but the numbers grounded represent considerably less than 1% of the local population fledging from burrows (Miles et al., 2010), and recent improvements to the lighting infrastructure in Village Bay have almost eliminated routine fledgling groundings, now generally limited to nights when there is thick fog in the bay. The two storm-petrel species can be grounded after being attracted to lights on oil production platforms and on ships at sea (Deakin et al., 2022). Attraction of adults to artificial lights are more likely to occur during foggy, misty and light rain conditions (Guilford et al., 2019; Deakin et al., 2022). Attracted birds may die from collision with objects at artificial lights, but most often, they are simply grounded and depart at first light in the morning. However, risk of predation is likely to be increased substantially for birds grounded on land.
Overnight anchoring/mooring of cruise ships or other brightly illuminated vessels in vicinity of petrel or shearwater colonies is also a growing concern. While systematic evidence for storm petrels is more limited than that relating to shearwaters, a number of potentially damaging impact pathways exist:
- Potential for direct illumination of colony areas leading to depression in number of visits to the colony by breeding or prospecting adults during breeding season. This could result in reduced provisioning of dependent young, with consequences for breeding success and/or fledgling survival or disruption to colony recruitment.
- Potential for fledglings to be attracted to artificial light sources, either directly from the colony or from sea following initial fledging flight with associated risk of grounding and mortality.
- Potential for adults to be attracted to artificial light sources with associated risk of grounding and mortality.
- Potential for adults to become disorientated by presence of artificial light sources and to waste energy in circling such sources over prolonged periods.
Appropriate adaptive responses
Miles et al.(2010) showed that the number of grounded birds can be limited by reducing the amount and/or intensity of artificial lighting, although that adaptive response was more effective in the case of storm-petrels than for Manx shearwaters.
Deakin et al. (2022) list potential management options for artificial lights:
- Alter pattern of illumination of offshore wind farm hazard warning lights to flashing rather than steady.
- Alter wavelength of lights (but it is uncertain which colour would be least attracting).
- Shield lights to avoid upwards radiation of light.
- Reduce intensity of lights.
- Reduce numbers of lights.
- Cover windows with blinds.
- Turn off lighting at key times in the season (e.g. chick fledging period).
- Use radar-activated lighting for aircraft warning lights at sea.
- Train people in safe handling and release of stranded birds at lights.
Oil pollution
Impacts
Impacts of oil pollution on seabirds are well known and are still of concern (King, Elliott and Williams, 2021). Seabirds are vulnerable to major oil spills, but most mortality is caused by chronic oil pollution. Seabirds that spend more time on the sea surface are at greater risk than seabirds that spend most time flying. Mitchell et al. (2004) listed oil pollution as a strong effect on common guillemot populations, a moderate effect on razorbill, black guillemot and puffin, and a slight effect on gannet, cormorant, shag, great skua, kittiwake. Forrester et al. (2007) broadly matched that list, but suggest the effect of oil pollution is moderate for common guillemot and slight for razorbill and puffin. There is considerable evidence of seabird mortality related to oil pollution from major oil spills, but also from routine beached bird surveys that also detect oiling caused by chronic pollution. Levels of oiling of seabirds found on beached bird surveys have decreased considerably over the decades all around the coasts of Europe. Highest levels have historically been in the southern North Sea, with much lower levels of seabird oiling around Scotland. Long-term monitoring of beached birds in the southern North Sea led Stienen et al. (2017, p. 194) to conclude that “oil pollution in the southern North Sea has almost halted”. They attributed this to legislation and monitoring to prevent chronic discharges from shipping traffic, which has been the predominant source of oil pollution in European (including UK) waters. Younger seabirds tend to be at higher risk than adults (Votier et al., 2008).
Although major oil spills can kill large numbers of seabirds, especially common guillemots and razorbills, the breeding populations of these bird species at colonies in the general area of such events tend to show surprisingly little impact. This appears to be at least in part because oil mortality particularly affects the youngest age classes more than adults, but also because recruitment rates of immature guillemots increase considerably to recompense for losses of breeding adults, buffering impacts of oil spills on breeding populations (Votier et al., 2008). This is only possible for a population that has a pool of immature birds available to fill spaces created in the adult population, so might differ for a population subject to poorer environmental conditions.
Appropriate adaptive responses
It is unlikely that further efforts to reduce risk of oil pollution would represent significant gains for seabird conservation in Scotland. Nevertheless, continued awareness of risks from oil pollution and continued monitoring remain desirable.
Chemical pollution
Impacts
There are numerous studies investigating chemical contaminants in seabirds. These show high concentrations in certain species, especially species high in the food web such as great skua, and species that can feed on larger (and therefore older) fish that tend to accumulate higher concentrations of contaminants, such as gannet (Power et al., 2021) and cormorant (Mason, Ekins and Ratford, 1997). There have been instances where contaminants have had severe impacts on seabird populations, but these are rare and localised instances, such as mass mortality of Sandwich terns in the Netherlands due to pesticide pollution in the 1960s (Mitchell et al., 2004).
Contaminants in seabirds may have interactive effects with other pressures, for example, concentrations of lipid-soluble contaminants will increase in the blood as lipid stores are used up particularly during periods of food shortage. High concentrations of contaminants could kill birds that are already starving. However, the concentrations of most legacy contaminants such as mercury, Dichlorodiphenyltrichloroethane (DDT), dieldrin and Polychlorinated biphenyls (PCBs) have decreased in seabirds around the North Atlantic in recent decades (Pereira et al., 2009, Braune et al., 2015, Harris and Wanless 2011). Unexpected emerging pollutants may still be a concern, but seem unlikely to have detectable impacts on seabird populations in Britain and Ireland (Leat et al., 2019). In cases where contaminants are atmospherically transported and condense in cold air, higher incorporation into food chains may occur in Arctic rather than temperate systems (Leat et al., 2019).
Appropriate adaptive responses
Since the evidence strongly indicates that contaminants such as heavy metals and halogenated organic chemicals have little impact at the population level on seabirds in Britain and Ireland, there is probably no scope for adaptive responses relating to contaminants. This situation may contrast with the situation in marine mammals, where contaminant levels seem not to have decreased to the same extent as in seabirds. Any adaptive responses to reduce contaminant levels in marine mammals would likely reduce levels in seabirds too, but there would probably be very little gain to seabird populations.
Plastic pollution
Impacts
Mitchell et al. (2004) identified plastic pollution as a slight threat to gannet, cormorant and puffin. Forrester et al. (2007) mentioned plastic pollution affecting fulmar and gannet in Scotland. Plastic pollution affecting seabirds includes ingestion of small fragments of plastic and entanglement in plastic waste. These are problems for different seabird species (O’Hanlon et al., 2017).
Ingested plastic
Ingestion of plastic by seabirds is a global concern (Wilcox et al., 2015), with evidence of damage to internal organs by ingested plastic (Lavers, Hutton and Bond, 2019; Rivers-Auty et al., 2023).
A monitoring programme has been established to collect dead fulmars found washed-up on beaches in order to quantify the plastic in their stomachs as part of OSPAR Ecological Quality Objectives (EcoQO) (van Franeker et al., 2021). Fulmar was chosen as an indicator species as it is widespread across the OSPAR region, and as a petrel it has a surface feeding habit that lends itself to plastic monitoring in surface waters. The anatomy of petrels - a narrowing between the proventriculus and gizzard - prevents pellets containing indigestible material in the gizzard being regurgitated. This means that ingested plastic remains and accumulates in the gizzard reducing the amount of food that petrels can digest. Plastic particles can also cause ulceration of the digestive tract. The same anatomical arrangement occurs in storm-petrels, which also accumulate plastic, but much less is known about how much plastic storm-petrels consume because this aspect is not monitored.
As a result, plastic consumption in fulmars is better understood than in other species. The amount of plastic in fulmar stomachs sampled from beaches around the North Sea has declined significantly between 2002 and 2018 (van Franeker et al., 2021), suggesting that this problem is probably declining in European waters as a result of increased concern about plastic particles in the marine environment.
In other seabird taxa, such as gulls and skuas, plastic is ingested, but indigestible material is regurgitated as a pellet (Hammer et al., 2016), which is possible because there is a wide opening between the proventriculus and gizzard, so plastic does not accumulate. Harris and Wanless (2011) note that a few puffins have been found dead with rubber bands in the stomach, particularly in east Scotland. Small numbers have also been found with ingested plastic, more often in east Scotland than in west Scotland. This was found more often in juvenile puffins than in adults.
Studies worldwide on accumulation of plastic in the stomachs of petrels suggest that this might have a detrimental effect on seabirds accumulating large amounts of plastic (Wang et al., 2021; Rivers-Auty et al., 2023). However, the impact, if any, at the population level is highly uncertain.
Entanglement
Entanglement of seabirds occurs in abandoned, lost or otherwise discarded fishing gear (ALDFG), in recreational fishing line, in other types of plastic litter at sea, and in plastic waste incorporated into seabird nest material (O’Hanlon et al. 2017). Camphuysen (2008) reviewed entanglement of seabirds found during beached bird surveys on the coast of the Netherlands from 1970 to 2007. He reported that 513 complete carcasses of seabirds out of 215,347 (0.24%) had been entangled in plastic waste. Most of that was nylon fishing line or fragments of fishing net/rope. He noted that the frequency of entanglement, although low, had increased since 2004. About 50% of entangled seabirds were gannets or herring gulls, the next most frequent victims being cormorants and great black-backed gulls. There appear to be no published data of this kind for Scotland. However, the low incidence of entanglement in the Netherlands suggests that this is a relatively minor cause of seabird mortality.
Grant et al. (2021) describe a standardised method for estimating the level of plastic waste incorporated into seabird nests. Plastic is a nest material used by many seabird species, especially gannets, cormorants and shags, and less frequently by kittiwakes. It is most likely gathered at sea along with more appropriate materials such as seaweed. In most cases the presence of plastic in seabird nests is benign, but occasionally a chick (or adult) may become entangled in the plastic at the nest and die there. Amounts of plastic (mostly from fishing gear) in gannet nests vary among colonies, with a tendency for there to be more in regions with more fishing activity (O’Hanlon et al., 2019).
Appropriate adaptive responses
The level of seabird mortality caused by plastic waste is relatively small and is very unlikely to have any detectable effect on population size or trend. However, it represents a welfare concern and additional pressure.
Probably the most effective solution to the problem is education. However, other actions that target regulation to further reduce the amount of fishing waste being discarded at sea and reduce the discard of fishing line at freshwater loch and river fisheries would also be effective.
There has been success, in certain locations, in removal of ALDFG gear from the seabed as an action to reduce mortality of seabirds and other marine animals. Stolte et al. (2022) describe a successful project of this type in Germany, using the combined efforts of the World Wild Fund (WWF) for nature, divers, fishers and public authorities. There may be merit in considering removal of ALDFG in targeted locations, although adopting similar methods (i.e. use of divers) is not likely to be feasible in Scotland due to seabed conditions and accessibility.
Removal of plastic from nests at seabird colonies has been attempted, but it is not clear that such removal is worthwhile, even if carried out at a time of year when the birds are absent in order to ensure no disturbance to the birds. Nests of some seabirds, such as gannets, cormorants, shags and kittiwakes can persist from year to year, depending on their exposure to winter rain and sea spray. Where nests persist, birds add material the next season, but do not need to spend so much time and effort finding nest material in order to build a whole new nest. Where plastic has been removed from nests, the breakdown of the rest of the nest material, and loss of that material through erosion and washing off into the sea, seems likely to be greater. That would require birds to find more material when they renovate nests the following spring. Finding nest material at sea is competitive, and birds may be more likely to use suboptimal nest material, such as plastic, as a result of competition to find nest material in the sea around large seabird colonies. Providing natural nest material, such as fragments of seaweed placed on the sea near to a colony during the nest-building period in spring might reduce the amount of plastic that birds collect, but the most appropriate adaptive response would probably be to encourage fishers to avoid ALDFG arising.
Neurotoxin poisoning
Impacts
Gulls scavenging on refuse may contract botulism (Ortiz and Smith, 1994). This tends to develop in anaerobic conditions, such as with decaying domestic waste in polythene sacks and was widespread across landfill sites in the UK in the 1980s-1990s (Ortiz and Smith, 1994). Large mortality incidents among gulls from botulism were seen throughout the UK, including in Scotland (MacDonald and Standring, 1978) although much of the botulism-related mortality occurred in southern England. Mitchell et al. (2004) note that outbreaks of botulism had moderate local impacts on herring gull, lesser black-backed gull and possibly great black-backed gull populations. Coulson (2015) suggests that botulism may have resulted in landfill being a negative influence on gull populations rather than a cause of population increase. Changes in the management of refuse disposal have greatly reduced this as a source of food for large gulls, and this has also reduced the problem of botulism in gulls in the UK.
Algal blooms can produce neurotoxins that are taken up into the food chain. Algal blooms were considered responsible for large numbers of shags (and some terns) being killed off the north-east coast of England, significantly reducing the local breeding population of shags, but only for a few years before numbers recovered (Armstrong et al., 1978; Coulson et al., 1968; Mitchell et al., 2004). A similar algal bloom production of neurotoxin was considered to be responsible for the death of thousands of kittiwakes (Coulson, 2011; Coulson and Strowger, 1999).
Appropriate adaptive responses
It is thought that toxic algal blooms are more likely in eutrophic waters, so reducing amounts of sewage and agricultural run-off should help to reduce risk of toxic algal blooms. However, blooms may also be more likely in warmer conditions, so climate warming could increase this risk.
Impacts on seabirds at the population level are probably negligible from botulism now and are probably extremely local in effect from algal toxins in shag and kittiwake. Reducing neurotoxin poisoning would not be a high priority for seabird conservation but may be of greater priority for public health.
Nest site interspecific competition
Impacts
Mitchell et al. (2004) listed interspecific competition for nesting habitat as having a moderate impact on European storm-petrel, Arctic skua, Sandwich tern, roseate tern, common tern, Arctic tern and a slight effect on Manx shearwater, Leach’s storm-petrel, little tern and razorbill. Forrester et al. (2007) did not consider this to be a major influence on any seabirds in Scotland, which is consistent with the classification by Mitchell et al. (2004). Impacts on terns come mainly from expanding populations of large gulls moving into tern nesting habitat and displacing terns. Impacts on Arctic skua come from expanding populations of great skuas moving into Arctic skua nesting habitat (Phillips, Furness and Stewart,1998). Impacts on razorbills come from common guillemots moving into boulderfield habitat where the cliff-nesting habitat has become full. Impacts on petrels and shearwaters come from expanding populations of puffins where these take over nesting habitat. More recently, there has been evidence of the expansion of some gannet colonies displacing other nesting seabirds, including displacement of large gulls from the top of the Bass Rock and common guillemots from broad ledges and platforms at Foula, Fair Isle and St Abbs.
Appropriate adaptive responses
The impacts of nest site competition are considered to be mostly local and colony-specific, therefore, there may be little need to consider management actions to limit these effects. However, where such management actions are required (perhaps mostly in relation to tern sites), the deployment of nest boxes in conjunction with bamboo canes may be effective actions that can reduce impacts.
Past management regimes at some tern colonies has been to remove adult Larus gulls and their eggs and nests (e.g. herring gull, lesser black-backed gull and great black-backed gull) from the immediate vicinity of tern colonies. This management, although effectively reducing nest site competition was primarily to reduce gull predation. Gull species were removed from the General Licence in 2020 due to the declining population statuses of gulls. From 2020, there has been a presumption against issuing licences for lethal control for the purpose of wild bird conservation, so this is no longer a potential conservation action.
Changes to nesting habitat
Impacts
Nesting habitat change is a highly species-specific issue as each seabird species has particular nesting habitat/nest site requirements. Mitchell et al. (2004) and Forrester et al. (2007) identify nesting habitat change as a factor having a slight effect on Arctic skua populations in parts of Scotland. At some Arctic skua colonies, agricultural improvement has reduced the availability of nesting habitat, elsewhere, the growth of conifer plantations has done the same. Onshore wind and local development may have further reduced Arctic skua nesting habitat in Shetland. These losses are from potentially large areas of nesting habitat for Arctic skuas, but these effects combine with the spread of great skuas to further reduce the availability of habitat at some sites.
Lesser black-backed gulls have also lost traditional colony sites for similar reasons, including tree growth at Flanders Moss, Stirling, which has seen the species abandon that once-large colony. Terns require suitable open ground for nesting and colonies may be lost through vegetation growth onto colony areas (which can be facilitated by the nutrient input onto that area from tern excrement). Sandwich terns in particular will abandon traditional colonies if vegetation grows onto the nesting area. Tern colonies may also be subject to erosion losses and flooding, and development of large tern colonies on the mainland creates a ‘honeypot’ attracting mammal predators, so may be unsustainable. Puffin colonies can quickly become damaged by burrowing activity as puffin numbers increase at a colony, resulting in soil erosion and instability so that the colony can collapse or wash away. Puffin colonies can also become overgrown with nitrophilous vegetation if not grazed. This last problem can include rampant growth of invasive plants such as tree mallow, making the area much less suitable for puffin nesting. Change can also come about as a result of the increase in numbers of another species of seabird. Nesting habitat of large gulls on top of the Bass Rock was colonised by the expanding numbers of gannets spreading onto the top of the island from the surrounding cliffs. Gannets tear up the vegetation, resulting in erosion of the soil, making the habitat unsuitable for gull nests, but also covering the former gull nesting area with gannet nests. Leach’s storm-petrels nest at high density on Dun, St Kilda, on the areas not occupied by puffin burrows, but appear to be excluded from nesting within the area of high density puffin burrows.
Appropriate adaptive responses
Changes to planning policy.
There are only a small number of Arctic skua colonies in Scotland, mostly in Shetland, Orkney, and the Western Isles. There would be an opportunity to ensure that no further loss of nesting habitat occurs around these colonies, including those lying outwith protected areas, by implementing planning policy that would address avoiding habitat deterioration on the land in and around Arctic skua colonies. This policy could be prescribed to a range of seabird species.
Tern nest boxes
Common terns, Arctic terns and roseate terns will use nest boxes and this can increase breeding success by protecting the incubating birds from weather and avian predators (Babcock and Booth, 2020a). Sandwich terns seem less interested in nesting inside tern nest boxes but will nest against the nest box for protection. Chicks will use these boxes as shelter and use chick shelters, which is likely to increase chick survival (Babcock and Booth, 2020a). Steel and Outram (2020) report on the gains to tern breeding numbers at the Isle of May as a result of deploying tern nest boxes.
Predator-proof fences
Predator-proof fences allow increased breeding success by terns or gulls because the colonies are protected from mammal predator attacks. It is possible to protect tern colonies with an electric fence to reduce risk of fox depredations (Short, 2020), but a more secure approach is to protect colonies with predator-proof permanent fences.
Tern rafts
Common terns readily use rafts provided as nesting sites (Furness et al., 2013; Coccon et al., 2018; Manikowska-Slepowronska, Slepowronski and Jakubas, 2022). However, other tern species are less readily attracted to rafts for nesting.
Artificial islets for terns and gulls
Creating artificial islets within ponds or wetlands can attract nesting terns and gulls and can provide nesting habitat that allows high breeding success where natural habitat has been lost. A particularly good example of this is along the south coast of France, where 154 artificial islets were created for terns and gulls with high success in terms of use and increases in tern and gull numbers (Schwartz et al., 2022). A good example in Scotland is St John’s Pool, Caithness (Hughes, O’Hanlon and Smith, 2021). There may be opportunities to use dredge sediment waste material more creatively to form such artificial islands for terns and gulls (Ausden et al., 2018; Manning, Scott and Leegwater, 2021). Habitat enhancement to make coastal seabird colonies safe from mammal predation and human disturbance was a major component of the potential actions suggested for the recovery of England’s breeding seabirds at a cost estimated around £5.5 million over 5 years (Lock et al., 2022).
Grazing and vegetation management at puffin colonies
Encouraging development of a short sward of grass over puffin burrows will help to ensure minimal erosion of soil and also prevent development of rank vegetation that might inhibit puffin access to burrows. Grazing can help to maintain a short sward, providing the grazing animals are light enough to avoid trampling of burrows. Grazing by rabbits or sheep may be appropriate.
Refuse disposal and agricultural practices
Impacts
Mitchell et al. (2004) identify refuse disposal as having a moderate effect on populations of herring gulls and lesser black-backed gulls in the UK. Forrester et al. (2007) suggest a moderate effect on herring gull populations in Scotland, but only a small effect on lesser black-backed gulls in Scotland. Neither suggest that great black-backed gull populations were significantly affected by refuse disposal. Small numbers of great black-backed gulls do occur on landfill sites, predominantly in winter, but these numbers are probably a very small part of the population. Coulson (2015) considered the likely roles of fishery discards and landfill as determinants of population change in the herring gull and suggested that whereas there is strong evidence that fishery discards contributed to population growth, landfill probably had a minimal influence. Firstly, landfill only became available as a major feeding opportunity long after the population growth had been well established. Secondly, mortality from botulism acquired from feeding on landfill sites may have killed as many or more gulls than the feeding opportunity helped to fuel population increase.
Changes in the management of refuse disposal to reduce amounts of edible material available to scavengers at landfill have greatly reduced this as a source of food for large gulls, so that this is probably no longer an important influence on distribution or population trend of either herring gull or lesser black-backed gull. Further management actions to exclude gulls from being able to exploit landfill for foraging are likely to continue the process of causing gulls to increase foraging effort and switch habitat use to recompense for the loss of landfill opportunities (Langley et al., 2021).
Lesser black-backed gulls can sustain a breeding population in inland locations where they can feed on earthworms, so management of pasture can influence the local population of this species. Great black-backed gulls will feed inland in spring on dead lambs and afterbirths in crofting areas and sheep farming areas. Herring gulls and lesser black-backed gulls will feed in urban areas, especially coastal towns where they can scavenge on refuse in bins and on streets. They will also feed in spring on feed in troughs of livestock such as sheep and will feed in chicken and pig farms where feed for those animals is open to exploitation by gulls. Whether these feeding opportunities influence demography and population growth of gulls in uncertain, but it seems likely that, at least in Scotland, such opportunities are relatively minor influences on gull distributions and demography.
Appropriate adaptive responses
None.
Disease epidemics
Impacts
Rather little is known about diseases of seabirds. Numerous diseases have been detected in seabirds worldwide, including blood parasites, herpesviruses, avian cholera, tick-borne viruses, avian influenza, Newcastle disease, and puffinosis, but impacts of diseases on seabird populations are little understood. There is widespread prevalence of antibodies against Borrelia, the spirochete that causes Lyme disease, among seabirds in the North Atlantic (Staszewski, McCoy and Boulinier, 2008). Many seabirds have ticks in the colony and there are cases of tick infections causing breeding failure of seabirds. Most seabirds carry feather lice, often of more than one species but usually of species endemic to particular seabird host species. Some seabirds carry fleas, including one flea species that is endemic to the Manx shearwater population at Rum (Kwak, Heath and Palma, 2019). However, feather lice and fleas apparently have little or no impact on their hosts.
Mitchell et al. (2004) noted that Manx shearwater populations may be slightly affected by puffinosis. Forrester et al. (2007) did not mention diseases of seabirds as a significant influence on populations. Puffinosis can affect Manx shearwater adults and chicks and can kill a significant number of chicks at some colonies. For example, 4% of chicks at Skokholm died of puffinosis in 1964 (Harris, 1965). The disease was thought to be caused by a coronavirus, but recent research suggests that this was a misinterpretation of laboratory results due to contamination, and that puffinosis is caused by infection of abraded skin by soil bacteria (Esmonde et al., 2022). Puffinosis can affect fulmars and gulls, as noted by MacDonald et al. (1967) in Orkney and Shetland, and can occasionally affect other seabirds. There does not seem to be a significant amount of puffinosis in Manx shearwaters at Rum, which might perhaps relate to the difference in burrow substrate in Rum compared to the fertile soil-based burrows at Welsh colonies of the species.
Until 2021, it seemed that avian influenza had little or no impact on seabirds in Britain and Ireland because the experience in Scotland was of low pathogenicity, but an outbreak of Highly Pathogenic Avian Influenza (HPAI) occurred among great skuas at several Scottish colonies in summer 2021 (Banyard et al., 2022). In 2022, HPAI severely affected great skua colonies. It killed over 1,400 adults from a population of about 1,500 pairs at Foula, the largest colony of this species (Camphuysen, Gear and Furness, 2022), and killed large numbers in the Faroe Islands, Iceland and Norway as well as throughout Scotland. This outbreak also affected many gannet colonies from Wales to Canada to Norway (Falchieri et al., 2022), killed over 8,000 adult Sandwich terns at many colonies in the Netherlands (Rijks et al., 2022) and at some colonies in the UK, and to a smaller extent killed large gulls, kittiwakes, roseate, common and Arctic terns, fulmars, cormorants, Manx shearwater, common guillemots and razorbills at some UK colonies, but especially in Scotland (Falchieri et al., 2022). Impacts on breeding seabirds were very patchy, with some colonies escaping infection, but others being badly hit. The virus has persisted into summer 2023, particularly in kittiwakes and other gulls, terns and auks. Great skua and gannet however do not appear to have succumbed to the outbreak in 2023, suggesting perhaps some resilience/immunity. It seems highly likely that further outbreaks will occur in future years. At this stage, the dynamics of the virus make it impossible to predict the extent to which this will develop, how quickly seabirds will develop immunity to the virus and if there will be longer-term impacts as a result of reduced fertility and/or absence of breeding adults in future generations.
Appropriate adaptive responses
A critical component of the response to diseases in wild birds is understanding how the disease is moving through a population and whether birds have developed a level of immunity.
Monitoring and reporting of dead or sick wild birds in combination with testing for example, for HPAI, allows the distribution, spread and any seasonality of infection and mortality to be identified. Serological testing of apparently healthy birds could also allow estimates of recovery rates from infection and focus management at colonies to aid in species recovery.
There has been debate over the effectiveness of carcass clearance of HPAI infected birds. Kuiken and Cromie (2022) make the case that Government departments responsible for wildlife protection must develop policies that prevent spillover of diseases of livestock into wild animals. This would include examples such as HPAI from poultry farms to seabirds, and Newcastle disease from chicken farms to seabirds. Preventive actions they recommend include restricting contacts between farmed birds and wild birds. Specifically in the context of HPAI, Kuiken and Cromie (2022, p.5) argue that adaptive short-term responses include “accurate monitoring of the virus and associated mortality of wild birds, and, where appropriate, coordinated removal of infected wild bird carcasses from affected sites to limit virus spread. Long-term recommendations include enhanced protection of seabird and waterbird sites, vaccination of poultry against HPAI, reduction of poultry farm size and density, and avoidance of waterbird-rich areas as a location for poultry farms”.
Biosecurity plans to limit transmission of disease at and between seabird colonies, as well as ensuring safe working environments will be a useful tool for responding to outbreaks and minimising transmission risk to and from colonies.
Lethal control and harvesting
Impacts
Adult herring gulls, lesser black-backed gulls and great black-backed gulls have historically been culled in very large numbers at many of the largest gull colonies in the UK. For example, about 45,000 adult herring and lesser black-backed gulls were culled over a 15-year period in the 1970s, early 1980s on the Isle of May (Mitchell et al., 2004). In the same period, destruction of nests, eggs and young also occurred. Coulson (2015) concluded that culling of herring gulls and lesser black-backed gulls was probably one of the most important drivers of population decline of those species in parts of the UK in the 1970s-2000s. Culling will not only have reduced breeding numbers in the culled colonies, but will probably have removed many of the young birds produced at colonies not subject to lethal control when those young birds recruited to breed. For example, the University of Glasgow and Clyde Ringing Group ringed several thousand lesser black-backed gull chicks at Flanders Moss near Stirling in the early 1980s. Of those ringed birds recovered at breeding age during the summer, most were recovered at colonies where culling operations continued, especially Horse Island in the Clyde. Those sites appear to have acted as sinks for breeding age birds such that culling will have impacted all colonies and not just those where the culls occurred.
Although some culls reported the numbers of birds killed, many did not. Continued legal control of large gulls under General Licences until 2020 also was without reporting of the numbers killed, so that it is impossible to be certain how important gull control has been in driving population change. Lethal control of large gulls, especially of great black-backed gulls, has occurred in crofting and sheep farming areas of Scotland, as these birds are seen as threats to weak lambs and ewes. Deliberate persecution is the main factor leading to ring recoveries of herring gull, lesser black-backed gull and great black-backed gull (Wernham et al., 2002). Removal of gulls from the General Licences has resulted in a reduction of the numbers of adults being lethally controlled, although it remains possible to obtain a specific licence from NatureScot to control gulls, primarily for the purpose to protect human health or safety or to prevent serious damage to agriculture.
Shooting of seabirds for sport is not permitted in the UK. It is now rare that licences are provided to permit the harvest of seabird eggs or chicks in the UK. The best known example of this is the controlled harvest of gannet chicks at Sula Sgeir under licence from NatureScot.
Appropriate adaptive responses
There may be scope to reduce the annual harvest of gannet chicks at Sula Sgeir. At present, the licence allows a take of up to 2000 birds per year, but a smaller harvest would be an opportunity to reduce other impacts on gannets without necessarily having to end the tradition of harvesting. During the 2022, 2023 and 2024 HPAI outbreaks, the local community cancelled their annual harvest in recognition of wider potential population impacts as a result of HPAI.
Continued analysis of the numbers of gulls controlled each year under each licence type is required. Considering the population trends of gulls, both nationally and regionally where available, in conjunction with the licensing data would help to determine how much scope there may be to improve the conservation status of these species by reducing the amount of licences being permitted. For a reduction in permitted licences to be successful and to mitigate against illegal persecution occurring, stakeholder engagement with local councils and other groups of interest would be key. Should further outbreaks of HPAI occur, this should also be taken into consideration when determining the number of licences being permitted. However, where gulls pose a genuine risk to public health and safety, NatureScot will continue to issue licences.
Generic actions for all pressures
Some impacts (such as the impact of climate change) would be difficult to address with direct actions, at least in the short term, so would best be addressed by actions aimed at building resilience by reducing pressures on seabird populations caused by other threats/pressures. Direct actions are summarised in Table 1.
Table 1
| Pressure | Potential seabird conservation actions | Key seabird species that benefit | Strength of evidence | Strength of effect |
|---|---|---|---|---|
| Climate change | Achieve Net zero globally. Conservation actions should aim to reduce the impacts of other pressures on seabirds making populations more resilient to potentially negative climate change impacts. | All | moderate | moderate |
| Depletion of forage fish stocks | Ecosystem-based management of sprat/juvenile herring fisheries | Kittiwake, common tern, common guillemot | moderate | strong |
| Depletion of forage fish stocks | Supplementary feeding of Arctic skuas | Arctic skua | good | strong |
| Depletion of forage fish stocks | Supplementary feeding of great skuas | Great skua | good | strong |
| Fishery discards | None recommended | - | - | - |
| Longline fishery bycatch | Monitor and reduce longline bycatch of seabirds | Fulmar, gannet, great skua, great black-backed gull | deficient | strong |
| Midwater trawl bycatch | None recommended | - | - | - |
| Static net bycatch | Monitor and reduce set net fishery bycatch of seabirds | Fulmar, gannet, cormorant, shag, common guillemot, razorbill | deficient | slight |
| Static net bycatch | Monitor and reduce aquaculture exclusion net entanglement of seabirds | Gannet, cormorant, shag, large gulls, auks | deficient | moderate |
| Mammalian predators | Eradicate invasive mammals from islands | Manx shearwater, European storm-petrel, Leach’s storm-petrel, shag, lesser black-backed gull, herring gull, great black-backed gull, Sandwich tern, roseate tern, common tern, Arctic tern, common guillemot, razorbill, black guillemot, puffin | good | strong |
| Mammalian predators | Biosecurity actions for seabird islands | Manx shearwater, European storm-petrel, Leach’s storm-petrel, shag, lesser black-backed gull, herring gull, great black-backed gull, Sandwich tern, roseate tern, common tern, Arctic tern, common guillemot, razorbill, black guillemot, puffin | good | strong |
| Mammalian predators | Predator-exclusion fences around mainland seabird colonies - Predator-proof fencing | Gulls, terns | moderate | strong |
| Mammalian predators | Predator-exclusion fences around mainland seabird colonies- Electric fencing (with daily wardening) | Gulls, terns | good | strong |
| Mammalian predators | Nest rafts for common terns | Common tern | good | moderate |
| Avian predators | Tern terraces, nest boxes and chick shelters | terns | moderate | strong |
| Avian predators | Tern terraces, nest boxes and chick shelters - Bamboo canes | terns | moderate | moderate |
| Avian predators | Nest boxes for black guillemots | Black guillemot | good | strong |
| Avian predators | Nest boxes for storm-petrels | European storm-petrel | moderate | moderate |
| Avian predators | Nest boxes for storm-petrels | Leach’s storm-petrel | deficient | moderate |
| Avian predators | Supplementary feeding of Arctic skuas | Arctic skua | good | strong |
Marine renewables collision with turbine blades
| Strategic selection of areas for marine renewable development | Gannet, gulls, skuas | deficient | slight |
Marine renewables collision with turbine blades
| New artificial nest sites for kittiwakes | Kittiwake | good | slight |
| Disturbance and displacement | Education programmes to gain public support for seabird conservation | all | good | moderate |
| Disturbance and displacement | Reduce human disturbance of seabirds | All, but mainly terns | good | moderate |
| Disturbance and displacement | Manage shipping disturbance of seabirds | Gannet, cormorant, auks | deficient | slight |
| Disturbance and displacement | Manage airborne vehicle disturbance of seabirds | Gannet, cormorant, auks | deficient | slight |
| Artificial light pollution | Reduce artificial light at sites near storm-petrel, shearwater and puffin colonies | Manx shearwater, European storm-petrel, Leach’s storm-petrel, puffin | deficient | slight |
| Oil pollution | None recommended | - | - | - |
| Chemical pollution | None recommended | - | - | - |
| Plastic pollution | Manage marine litter | Gannet, cormorant, shag, fulmar, large gulls | deficient | slight |
| Neurotoxin poisoning | None recommended | - | - | - |
| Nest site interspecific competition | None recommended | - | - | - |
| Changes to nesting habitat | Vegetation management at puffin colonies | Puffin | deficient | slight |
| Changes to nesting habitat | Create islets for terns and gulls | Terns, large gulls | good | strong |
| Refuse disposal and agricultural practices | None recommended | - | - | - |
| Disease epidemics | Monitoring of diseases | all | deficient | slight |
| Disease epidemics | Supplementary feeding of great skuas | Great skua | good | slight |
| Lethal control and harvesting | Reduce harvest of gannet chicks | Gannet | good | moderate |
| Lethal control and harvesting | Reduce lethal control of large gulls | Herring gull, lesser black-backed gull, great black-backed gull | moderate | moderate |
| Generic actions for all pressures | Education programmes to gain public support for seabird conservation | all | good | moderate |
| Generic actions for all pressures | Demographic monitoring of seabirds | all | moderate | moderate |
Part 2. Potential seabird conservation actions
The identification of funding opportunities for conservation action was not in the scope for this report.
Ecosystem-based management of sprat/juvenile herring fisheries
Summary description
Sprat and juvenile herring tend to be caught together, in fisheries that target sprat. In Scotland, sprat fisheries have been carried out predominantly in estuarine waters, such as in the Moray Firth and Firth of Forth (Fernandez et al., 2005; ICES, no date). Sprat fisheries risk depletion of the local stock of sprats, as occurred in the Firth of Forth in the early 1980s (Jennings et al., 2012). Recovery of fish stocks may lead to calls from fishers to reopen a local fishery, as in the Clyde Sea area where a large increase in sprat stock has occurred after the closure. High abundance of sprats has generated a pressure to reopen sprat fishing in the Clyde Sea area (Lawrence and Fernandes, 2021).
Scope of potential action
Depletion of forage fish is a pressure affecting many seabird species in Scotland. Some of these species, especially kittiwake, terns and common guillemots can forage primarily on sprats where these are available - and if abundant sandeels are not available. This action could reduce the impacts of sprat stock depletion by sprat fisheries, although those fisheries tend to be short-lived and opportunistic. Geographic scope for this measure would be areas where sprats are sufficiently abundant to potentially support a sprat fishery, such as Firth of Forth, Moray Firth, Clyde Sea area.
Options
- Reducing fishing mortality on sprat to maintain stocks above the ‘one-third-for-the-birds’ threshold (Cury et al., 2011) could improve the prey base available for seabirds, but would require detailed monitoring of sprat stock biomass by acoustic survey.
- Closure of Scottish coastal waters (or parts of them) to sprat fishing would ensure that sprat stocks would not be depleted by fishing, without the need to invest in an expensive acoustic survey programme.
Delivery involvement
This would need to involve fishers, fishing organisations and Scottish Government. Either option would require statutory instruments to change the position with regard to sprat fishing in the coastal waters of Scotland so would require Scottish Government to lead. This would also require consultation with fishers, fishing organisations and NGOs.
Delivery mechanism(s)
Strategic across wider seas.
Scottish Government could potentially implement wider seas fisheries restrictions through statutory instruments further to formal consultation on any proposed measures.
Measures could be focused on areas where sprats are sufficiently abundant such as Firth of Forth, Moray Firth and Clyde Sea area.
Timescales for delivery
Implementation would require a consultation period and development of appropriate measures, which could require two to five years to bring to effect. As sprat is a short-lived fish, stock recovery following constraint on fishing would be expected within two or three years of implementation. Recovery of seabird breeding numbers would be much slower, with a time lag of about 5 years due to the period taken for increased breeding success to feed through to breeding numbers and a constraint on seabird population growth being limited to no more than about 3% increase per annum for most seabird populations.
Dependencies/constraints
There may be opposition from fishing interests to a sprat fishery closure, or to constraints on sprat catches.
Supplementary feeding of Arctic skuas
Summary description
Arctic skua respond well to supplementary feeding. Territories are defended by an adult pair which means that supplementary food provided within a territory makes it likely that only the target birds will get the food. Providing the food in late evening means that adult attendance is likely to be high as all breeding adults are on territory overnight. Arctic skuas can be given tinned wet cat food and hen’s eggs (Davis, Nager and Furness, 2005). They can also be given forage fish or small strips cut from larger fish such as mackerel or herring (Gear, 2019). Pairs given supplementary food quickly learn to take this as soon as it is presented.
In 2001 and 2002, Davis, Nager and Furness (2005) gave supplementary food to 28 pairs of Arctic skua at Foula which were monitored alongside 25 control pairs. Supplemented pairs fledged 0.9 chicks per pair whereas control pairs fledged 0.5 chicks per pair and attended their territory more of the time than control pairs, so perhaps gained from not having to work so hard searching for food. Return rates of adults given supplementary food were higher than for control pairs, and the gain in adult survival from supplementary feeding may have a stronger impact on population change than the gain in terms of chick production.
Subsequent supplementary feeding at Foula started in 2019 (Gear, 2019) because the population had declined to just 19 pairs. The eight fed pairs fledged 12 chicks (1.5 chicks per pair) whereas eight matched control pairs fledged only four chicks (0.5 chicks per pair). Based on this, all pairs were given supplementary food in 2020-22, resulting in the decline in breeding numbers being halted. In the four years of supplementary feeding by Sheila Gear Arctic skuas at Foula averaged 0.93 chicks per pair, whereas in the four years before supplementary feeding the breeding success averaged 0.21 chicks per pair. Numbers of pairs declined from ca. 250 pairs in the mid-1970s to 63 pairs in 2009 to 19 pairs in 2019 but increased to 21 pairs in 2022 (Gear, 2022). The increased number of pairs cannot be attributed to increased production of fledglings because Arctic skuas do not start to breed until four years old (Horswill and Robinson, 2015) so suggests an effect on adult survival combined with recruitment from elsewhere. Similar supplementary feeding would be practical at other Arctic skua colonies if birds are food-limited and so are experiencing poor breeding success.
Scope of potential action
Supplementary feeding would be practical at Arctic skua colonies where local people or a reserve manager would be available to carry out the feeding daily throughout much of the breeding season (at least throughout chick-rearing and preferably from egg laying to post-fledging of chicks). Given the small size of Arctic skua populations the amount of supplementary food needed is modest, but the time input to provide this to pairs daily is considerable. Potential colonies where supplementary feeding could be practical include Foula, Fair Isle, Papa Stour, Fetlar, Unst, Papa Westray, Westray, Rousay, Eday, Flotta, St Kilda, and Handa.
Options
- Supplementary feeding of Arctic skua pairs on a daily basis through chick-rearing period.
- Supplementary feeding of Arctic skua pairs on a daily basis from laying to chick-fledging.
- Supplementary feeding of Arctic skua pairs on a daily basis from laying to chick dispersal from colony.
Delivery involvement
Input from NatureScot, NGOs and local communities would be required. This action would require work by reserve staff where they are already present at colonies, or work by local people willing to carry this out for several hours per day every day for at least the chick-rearing period (about 30 days in June-July) or for longer (up to 80 days if from laying (end of May) to chick-fledging (early August). Providing supplementary food is a fairly simple activity, but requires reasonable understanding of the need to do this in a way that avoids disturbance to the birds themselves and avoids risk of attracting other species to feed on the supplementary food.
Funders and/or funding partners may be required to implement action.
Delivery mechanism(s)
Site-based with a focus on one or a small number of selected colonies where the birds are readily accessible and provide as many pairs to feed as is practical.
It would be inappropriate to select an area, such as mainland Caithness, where there are very few pairs that are located far apart in relatively inaccessible terrain. Colonies such as Foula, Fair Isle, Papa Westray would seem particularly suitable. It would be appropriate to monitor breeding success achieved, and in the longer term any changes in breeding numbers at the colony. Where a colony recovers in breeding numbers there would be a case for reducing the amount of supplementary feeding, halting supplementary feeding if the evidence indicates that the local population has become self-sustaining once more.
Timescales for delivery
This could be implemented within one to two years, following the model already in place at Foula, Shetland. Supplementary feeding should not be seen as a permanent solution, but rather as a means of avoiding the extinction of particular colonies to maintain a population until conditions improve to allow the birds to be self-sustaining without supplementation. This might take many years, but restoring breeding populations that have been lost usually proves to be much more difficult than maintaining declining populations.
As seen at Foula, immediate benefits should be apparent within the first season with increased chick survival and the following year, with adult return rates. It will take four to five years however before fledged chicks are ready to breed and therefore, recruited into the breeding population.
Dependencies/constraints
It is uncertain where recruits into Scottish Arctic skua colonies come from, but the fact that this species is at the southern edge of its breeding range in Scotland suggests that either immigrants come from further north or that the population is not supported by immigration. Most Arctic skuas in Scotland are dark phase whereas in the Arctic almost all Arctic skuas are light phase. This implies that very few immigrants to Scottish colonies come from northern populations of Arctic skuas. The Scottish population may therefore be especially vulnerable to local extinction, and may be one of the first species to disappear from our breeding avifauna if the recent trend is not halted or reversed.
Supplementary feeding of great skuas
Summary description
Great skuas are opportunists that will scavenge within their breeding territory when able to do so. Although no studies have tested the possibility of supplementary feeding great skuas to improve breeding success or adult survival, the evidence from supplementary feeding of Arctic skuas indicates that this is highly likely to work for great skua. As with Arctic skua, there might be a need to train great skuas to take supplementary food, but once the habit has established it should be possible to feed pairs on their territory in the late evening (when all adults are present on territory).
Scope of potential action
Given the relatively large size of some great skua colonies (hundreds of pairs) and the considerably larger body size compared to Arctic skua (so a larger food need), it would be impractical to provide supplementary food to large numbers of great skua pairs. However, the huge impact of HPAI on great skuas in Scotland in 2021 and 2022 raises the possibility that the population of this species might be considerably reduced in future. If numbers were to be reduced to levels where local extinction appears to be a risk, supplementary feeding could help to maintain remaining numbers to prevent colony extinctions and support numbers until the population could recover or stabilise.
Options
Provide short-term supplementary food to pairs of great skuas at colonies that have been hit the hardest by HPAI and at risk of local extinction. Providing supplementary food at colonies also supporting seabirds that are vulnerable to great skua predation however, should be avoided.
Delivery involvement
Input from NGOs and local communities would be required. This would require work by reserve staff at colonies where staff are already present, or work by local people willing to carry this out for several hours per day every day for at least the chick-rearing period (about 30 days in June-July) or for longer (up to 80 days if from laying (end of May) to chick-fledging (early August). Providing supplementary food is a fairly simple activity, but requires reasonable understanding of the need to do this in a way that avoids disturbance to the birds themselves and avoids risk of attracting other species to feed on the supplementary food. Since the efficacy of this has not been tested with great skuas the breeding success of supplemented pairs should be monitored and compared with that of control pairs at the same colony. If it proves to be successful in boosting breeding success then it could be applied to all pairs rather than requiring a control sample.
Funders and/or funding partners may be required to implement action.
Delivery mechanism(s)
Site-based with a focus on one or a small number of selected colonies where the birds are readily accessible and provide as many pairs to feed as is practical.
Until very recently this would be considered unnecessary for a species that has been increasing in numbers for many decades, but the mass mortality of great skuas from HPAI in 2021 and 2022 put that in some doubt. The impact of HPAI on great skua is uncertain and it is unclear how many colonies will remain in future. Possible locations where it might be appropriate for supplementary feeding would be Hermaness, Noss, Hoy, Foula and Handa. It would be appropriate to monitor breeding success achieved, and in the longer term any changes in breeding numbers at the colony. Where a colony recovers in breeding numbers there would be a case for reducing the amount of supplementary feeding, halting supplementary feeding if the evidence indicates that the local population has become self-sustaining once more.
Timescales for delivery
This could be implemented within one to two years. Supplementary feeding should not be seen as a permanent solution, but rather as a means of avoiding the extinction of particular colonies to maintain a population until conditions improve to allow the birds to be self-sustaining without supplementation. This might take many years, but restoring breeding populations that have been lost usually proves to be much more difficult than maintaining declining populations.
If successful, there is the potential for immediate benefits being apparent within the first season with increased chick survival. It will take four to five years however before fledged chicks are ready to breed and therefore, recruited into the breeding population.
Dependencies/constraints
Where colonies have seabird species that are vulnerable to great skua predation, and/or other pressures it may not be appropriate to enhance the rate of great skua population recovery.
At present, great skuas are sometimes considered to be problem birds because they kill smaller seabirds, displace Arctic skuas, and occasionally attack and kill lambs and ewes, usually but not always when those animals are sick or dying. The public attitude to great skuas might change if the species becomes rare as a result of HPAI (or any other pressure), but unless the public attitude did change, there could be resistance to supplementary feeding of a great skua. Scotland is home to a high proportion of the world population of this species (approximately 60%), so it would be important to take action to avoid the extinction of the Scottish great skua population if that were to be a risk.
Monitor and reduce longline bycatch of seabirds
Summary description
Deployment of bycatch management actions in the longline fishery has been demonstrated to be successful, not only in reducing the rate of bycatch of seabirds (Dunn and Steel 2001) but also in allowing recovery of populations that had been depleted by high levels of bycatch mortality (Dasnon et al., 2022).
Potential bycatch reduction actions include:
- Deployment of bird-scaring plastic streamer lines over the longline.
- Setting lines through an underwater tube so that hooks are deployed too deep for most scavenging seabirds to reach.
- Use of a line shooter.
- Use of water cannon to deter seabirds from approaching.
- Setting lines only at night to avoid attention from diurnally-active seabirds,
- Avoiding discharge of offal so that scavenging seabirds are not attracted in the first place.
- Adding weights to lines to increase sinking speed.
- Use of hook designs that reduce risk of snagging seabirds.
- Deployment of baited hooks in a capsule that only opens once the line has reached a specific depth (Løkkeborg and Robertson, 2002, Løkkeborg, 2003; 2011, Melvin et al., 2019; Gilman et al., 2020; Dasnon et al., 2022; Gilman et al., 2022).
Many of these actions can be very effective. However, best practice is to use a combination of reduction actions that are most appropriate and practical for the particular fishery and bycatch concern (Løkkeborg, 2011).
Currently there are no legal requirements for UK longline fishing vessels to reduce bycatch of seabirds. Voluntary management actions are practiced by some members of the fleet (Kingston et al., 2023). Bycatch is monitored in Scottish waters in this fleet under the UK Bycatch Monitoring Programme. Total observer sampling effort in the fishery over the last decade equates to about 5% of the annual UK fishing effort in a typical year. The evidence suggests that there would be scope to reduce bycatch of fulmars, gannets, great black-backed gulls and great skuas in longline fisheries in northern Scottish waters, although numbers may be much smaller than in some overseas longline fisheries to which Scottish seabirds may be exposed, especially during the nonbreeding season.
Scope of potential action
The main impact of longline fishery bycatch of seabirds by UK vessels in Scottish waters is on fulmar and gannet, with rare instances of great black-backed gull and great skua also recorded. All of these species could benefit if actions were introduced to reduce seabird bycatch in the longline fishery although the small numbers of gulls and skuas caught on longlines in Scottish waters suggest that the gain for those species may be small. Several tried and tested methods are possible, but which of these would be most effective in the Scottish fishery is uncertain. Setting lines only at night has been shown to increase bycatch of fulmars, so is likely to be unsuitable.
Continued detailed monitoring of seabird bycatch as part of the UK Bycatch Monitoring Programme and experimental study of the efficacy of mitigation measures in the Scottish waters is required.
Delivery involvement
This would need to involve fishers, fishing organisations and Scottish Government. Either option would require statutory instruments to change the position with regard to long-line fishing in the coastal waters of Scotland so would require Scottish Government to lead. This would also require consultation with fishers, fishing organisations and NGOs.
Delivery mechanism(s)
Strategic across wider seas.
Scottish Government could potentially implement wider seas fisheries restrictions through statutory instruments further to formal consultation on any proposed measures.
Trials on the use of streamers and on altered buoyancy regimes are ongoing through the European LIFE funded project Coordinated Development and Implementation of Best Practice in Bycatch Reduction in the North Atlantic, Baltic and Mediterranion regions (‘the CIBBRiNA project’) in collaboration with the UK Bycatch Monitoring Programme. The Future Catching Policy will provide the opportunity for the Scottish Government to deliver on obligations to minimise and, where possible, eliminate bycatch of sensitive marine species.
The Scottish Government will be consulting on fisheries management measures in marine and colony extension Special Protection Areas (SPAs) which may include measures for the longline fleet. Further research and experimentation is needed to determine what options would be most effective.
Timescales for delivery
Two to five years of research would be necessary before effective management actions could be implemented to provide sufficient time to determine which actions are most appropriate to implement in a Scottish context.
Effective techniques however could be implemented voluntarily within one or two years on confirmation of appropriate methods. Changes in legislation to make bycatch reduction methods mandatory would however take longer, potentially up to five years to implement.
Dependencies/constraints
Cooperation of fishers would be essential. Although actions in the Scottish longline fleet may be possible, numbers of Scottish gannets caught on longlines are likely much higher in overseas fisheries in southern Europe and west Africa, so developing management actions in the wintering areas of those species might be more effective but more difficult to achieve.
Monitor and reduce set net fishery bycatch of seabirds
Summary description
Monitor the scale of mortality of seabirds in set nets in Scottish waters and where appropriate, implement actions to reduce bycatch mortality of seabirds in set nets.
Scope of potential action
Reducing bycatch of seabirds in set nets, where it occurs, could benefit common guillemot, razorbill, gannet, fulmar and large gulls, but extending current monitoring data on bycatch would help identify the detail on this.
Options
- Full or partial closure of Marine Protected Areas (MPAs) with high seabird foraging abundance to set net fishing during seasons when diving seabird numbers are highest.
Require attendance of fishers at nets while nets are operational and
train fishers to release birds that become entangled in nets before they drown.
- Deploy above water deterrents to reduce seabird numbers close to set nets.
- Use high visibility corline in nets to increase visibility to seabirds.
Delivery involvement
To ensure any of the options above are targeted at key pressure hot-spots, discussions with fishers to establish locations, scale, frequency, gear type and methods is a priority within MPAs where this activity is carried out.
Seabird researchers and fisheries observers would be required to establish level of bycatch and test mitigation measures where bycatch is reported.
NatureScot are responsible for providing advice to Marine Directorate on seabird impacts and potential options to avoid these. Marine Directorate are responsible for implementing fisheries measures.
Delivery mechanism(s)
Scottish Government could potentially implement site-based fisheries restrictions through statutory instruments further to formal consultation on any proposed measures.
Extending current monitoring bycatch of seabirds and testing management actions would be strategic based on the evidence. Implementing bycatch reduction actions should be site-based in areas with bycatch hot-spots.
Timescales for delivery
One year to complete assessments and consultation on proposed measures with implementation of MPA measures within two years.
Dependencies/constraints
Cooperation of fishers would be essential.
Monitor and reduce aquaculture exclusion net entanglement of seabirds
Summary description
The finfish farming sector in Scotland uses protective nets to safeguard stock from seals, otters and birds. All sites have top nets over the tops of stocked cages and some may also deploy sub-surface nets around the cages (increasingly with double sub-surface netting for seal management). Marine birds attracted to the stock or to wild fish congregating around cages risk entanglement in nets or entrapment in cages but the extent of this issue is not fully understood.
A particular concern has arisen from the recent and increasing adoption of pole-mounted top nets for marine cages. This system lifts nets much higher above the water surface than systems using central floating supports, which typically used nets with 50 mm mesh to reduce risk of bird entanglement. Given much greater net area, and hence weight to be supported, larger mesh sizes have been used in pole-mounted systems. In some locations mesh sizes as large as 300 mm or more have been used for the ceiling nets and there have been instances of gannets diving through such nets into fish cages. They can then be trapped under the net or may become entangled in the top net when trying to dive through (NatureScot, 2020).
Since late 2020 NatureScot has worked closely with the aquaculture sector and local planning authorities (LPAs) to address these risks. Standard conditions are attached to planning permissions for new /extended fish farm sites wishing to adopt pole-mounted top nets as well as to all permissions under permitted development processes for amendments to top nets at existing farms. NatureScot have also provided standing advice for LPAs to ensure that NatureScot are consulted where such amendments involve proposed use of pole-mounted top nets with mesh sizes of greater than 100 mm and/or are located in sensitive locations with respect to proximity to SPAs for large gulls, shags or cormorants. The consent conditions require monitoring and standardised data recording using an agreed proforma, adapted from that previously used at sites in Orkney to monitor auk and diver entanglement in hamster wheel top nets and sub-surface nets. This also records relevant context data, including details of nets deployed and stocking, to assist in understanding circumstances under which entanglement/entrapment of birds may occur
In addition to these routine returns, the conditions stipulate that any exceptional incidences of entanglement, defined on basis of frequency, persistence and numbers of birds (details vary with site) require immediate notification to NatureScot and LPA such that adaptive management actions could be applied if required.
Assessment of mortality in pole-mounted top nets further to the implementation of these consent conditions is on-going and the first two years of data are currently being analysed to inform any amendments to NatureScot advice.
Scope of potential action
Reducing mortality of seabirds in aquaculture cage nets would likely benefit gannet, large gulls, cormorants, shags and auks. Geographic scope is likely to be a concern mostly in the west from Argyll to the Western Isles to Shetland where almost all marine finfish aquaculture in Scotland is located.
Options
- All new aquaculture applications and proposed extensions to existing aquaculture developments should continue to be subject to consent conditions in accordance with NatureScot advice. Voluntary returns from existing sites not subject to formal consenting conditions would further assist in understanding bird interactions with finfish farms in Scotland.
- There is a prerequisite for monitoring of seabird mortality associated with aquaculture cage design, so that appropriate adaptation actions can be determined based on evidence.
Delivery involvement
Continuing discussions with the aquaculture industry and LPAs is a priority. NatureScot has already established a pole-mounted top nets working group with the industry.
Further monitoring of seabird mortality at marine cages with pole-mounted top nets beyond the self-reporting mechanism now in place could be carried out by camera surveillance. This should involve NatureScot and LPAs based on the planning consents issued.
Delivery mechanism(s)
Mitigation and entanglement monitoring can be implemented through site-based planning consent conditions.
Monitoring of entanglement/entrapment can be used to inform development of Best Practice Guidelines for the industry. At a site level, adaptation plans should be developed and implemented in any instances where entanglement monitoring highlights concerning levels or frequency of bird mortality. Mandatory action is required where there could be risk of cumulative or in-combination population level impacts on SPA populations.
Timescales for delivery
Currently ongoing.
Dependencies/constraints
Cooperation of aquaculture industry is essential.
Eradicate invasive mammals from seabird islands
Summary description
Invasive mammalian predator eradication from islands is a well-established procedure that has now been carried out at hundreds of sites world-wide and at a small number of islands in the UK. Those include eradication of black rats from the Shiant Isles, eradication of brown rats from Ailsa Craig, Canna, and Handa (reviewed in Part 1). However, although eradication was successful at Handa, rats re-colonised the island subsequently, indicating the importance also of biosecurity for islands that have been cleared or never had invasive mammalian predators. Brooke et al. (2007), Ratcliffe et al. (2009) and Stanbury et al. (2017) developed lists of top priorities for eradication of invasive mammalian predators from islands to conserve vulnerable seabirds. Twenty-two of the 25 top priority sites listed by Stanbury et al. (2017) are in Scotland. Although eradications have now been carried out or are underway at a few of these sites, many have still got invasive mammalian predators impacting seabird populations and preventing recolonisation by seabirds that have been extirpated.
In some cases, eradication of other non-predatory invasive mammals could show conservation benefits for seabirds, for example eradication of rabbits where severe erosion has reduced seabird nesting opportunities and damaged soil stability (Spatz et al. 2023).
Scope of potential action
Which seabird species would benefit from eradication of invasive mammalian predators would depend not only on the predator species, but also on the habitat on the island, and therefore the extent of overlap between seabird and predator. In broad terms, eradication of invasive mammalian predators from islands is an action that would benefit storm-petrels, shearwaters, terns, ground-nesting gulls, shag, cormorant, auks, and possibly fulmar to a small extent at a small number of sites. Given the considerable gains for seabirds that could result from eradication of invasive mammalian predators and the proven effectiveness for several seabird species, this would also be a very high priority action. For common guillemot, invasive predator eradication may have no benefit for those birds nesting on cliff ledges but may improve breeding success of those pairs nesting in coastal boulderfields and may allow increasing numbers to breed in coastal boulderfields. Around Scotland, islands important as seabird colonies, or where seabirds used to breed before being wiped out by invasive mammalian predators, and where invasive predators continue to represent a strong pressure on seabirds, are predominantly in west or north Scotland (especially Orkney and Shetland).
Eradication of non-predatory invasive mammals such as rabbits could be worth consideration at seabird colonies such as Eilean Mor in the Flannan Isles, where the dense rabbit population has eroded the west slopes of the island, exposing areas of bare rock topped with a honeycomb soil cap that is at risk of being irreparably lost in winter storms. The potential benefits of rabbit eradication, and the extent of the future erosion risk to burrow nesting species on Eilean Mor have not been explored. The Flannan Isles are one of the few SPA puffin colonies that recorded a population increase during Seabirds Count, and it is unknown whether the removal of rabbits and associated competition for limited burrow space could increase the resilience of this population.
Options
- Update of the review by Stanbury et al. (2017) to include sites where particular seabird species are now locally extinct as well as sites where populations have been depleted.
- Strategic selection of target islands followed by site-based eradication projects island by island.
Delivery involvement
Landowners, managers and local communities.
A partnership approach including Scottish Government, NatureScot and NGOs is required to develop a prioritised programme of eradication.
Funders and/or funding partners would be required to implement action.
Invasive mammalian predator eradication is often carried out by internationally-renowned specialist professional teams. Some training has been done in the UK, and there would be possibilities for UK teams to do this work, or to work with an international team.
Delivery mechanism(s)
Site-based with overview from a strategic ‘Seabird Recovery Partnership’. Delivery would need to develop a long term (decadal) programme of work.
Timescales for delivery
Planning could require one to two years.
Rat eradications typically take one winter to complete, followed by intensive monitoring which should record no signs of rats for two years in order to meet the internationally agreed criterion for rat-free status, and pre-eradication site-based planning may require a year or two in advance of implementation. Timescales to eradication in other species may be considerably longer; stoats for example may take several years to remove, and require a longer period of intensive monitoring post-eradication. Such a programme should probably be delivered over a period of ten to twenty years.
Recovery of seabird populations can be surprisingly quick where there are birds able to immigrate into newly cleared habitat – for example, the first recorded storm-petrel chick was found in the third season post-eradication. Growth rates of seabird populations however rarely exceed 3% growth per annum, so recovery may take decades to complete.
Dependencies/constraints
Some seabird islands are privately owned so the landowner would need to be supportive of the programme. Some seabird islands have a resident human population which would have views about eradications, especially in the case of feral cats. For example, it is important to appreciate that on some islands, cats go ‘feral’ in the summer to feed on seabirds but may return to croft houses for the winter. Some landowners would not agree to eradication of feral cats where their island is also affected by introduced rats or rabbits.
Biosecurity is obviously much more difficult at an island with a resident human population than at remote islands with no human visits. Eradication would not normally be worthwhile unless a biosecurity programme was established as well.
Biosecurity for seabird islands
Summary description
Biosecurity identifies risks of invasive mammalian predators getting onto seabird islands and promotes actions to reduce that risk as much as possible (Bambini and Varnham, 2020). This is an integral part of eradication of invasive mammalian predators from seabird islands (Bell et al., 2019) but is also important where there is a risk of novel introductions to islands free from invasive mammalian predators.
Prevention is far more cost-effective than eradication, but requires a long-term commitment, so short-term biosecurity programmes need to be carried forward into permanent programmes.
Scope of potential action
Scope for potential action includes SPA colonies, non-SPA colonies and sites where seabird populations used to be present but have been extirpated by invasive non-native mammalian predators. The islands included in the Biosecurity for Scotland project are Ailsa Craig, Auskerry, Calf of Eday, Canna & Sanday, Copinsay, Fair Isle, Fetlar, Flannan Isles, Forth Islands, Foula, Glas Eileanan, Hoy, Mousa, Mingulay, Berneray & Pabbay, Monach Isles, North Rona & Sula Sgeir, Noss, Papa Stour, Papa Westray, Pentland Firth Islands, Priest Island, Ramna Stack & Gruney, Rousay, Rum, Shiant Isles, Staffa, St Kilda and Treshnish Isles.
Options
Biosecurity for Scotland (2023 – 2026) is a continuation of ‘Biosecurity for LIFE’ (2018 – 2023). The Biosecurity for Scotland project ends in March 2026, and one of the project’s actions is to develop a Future Biosecurity Partnership for the long-term continuation of biosecurity at the project’s 38 sites.
A long-term biosecurity programme should include the following elements:
- Education and awareness raising
- Prevention measures (maintaining barriers on invasion pathways such as boats and harbours)
- Routine on-island surveillance (early detection)
- Rapid incursion response plans
Delivery involvement
Land managers, local communities and commercial vessel owners.
Biosecurity for Scotland is funded through the Nature Restoration Fund, and its partners are RSPB, NatureScot, and National Trust for Scotland. An ongoing programme of biosecurity should have a similar team of partners.
Additional funders and/or funding partners would be required to implement this action beyond 31 March 2026.
Delivery mechanism(s)
Site-based, supported by the Biosecurity for Scotland, Nature Recovery Fund project with overview from a strategic ‘Seabird Recovery Partnership’.
Timescales for delivery
Further action to ensure long-term biosecurity should be developed as the Biosecurity for Scotland project continues. There is a need for a permanent seabird island biosecurity programme for Scotland, in order to embed and maintain the measures set up by Biosecurity for LIFE and Biosecurity for Scotland.
Dependencies/constraints
Biosecurity depends on the behaviour of a large number of different stakeholders/groups. In 2022, the Biosecurity for LIFE project commissioned a survey to gather information on what support the stakeholders affiliated with biosecurity of Scottish islands would need to continue with biosecurity implementation after the end of the project (Pearson, 2022).
When asked for the one thing someone could do today which would improve prevention, the most popular theme was to increase awareness locally for communities/visitors/public through more signs and leaflets at harbours with website address to find out more, followed by controlling rats on boats/harbours more.
When asked what they considered as the main challenges to regular surveillance continuing after the Biosecurity for LIFE project, the most popular themes were funding people and boats for surveillance visits and lack of available skilled people (contractors, volunteers) to verify sightings/give advice.
When asked what they considered as the main challenges to carrying out an effective incursion response on your island(s) after the Biosecurity for LIFE project, the most popular theme was access to enough skilled staff/volunteers, who would have the requisite knowledge and time to help to carry out the work.
The top three themes identified by stakeholders when asked what resources will be required to meet any/all these challenges were “funding (staff/equipment/boating/people)”, “a dedicated paid staff member to coordinate surveillance and incursion”, and “sustained awareness raising”. The stakeholder responses captured the value of having a biosecurity personnel dedicated to providing ongoing support / training / advice to stakeholders as they seek to maintain long-term biosecurity measures. There are various examples of biosecurity losing its effectiveness over time post-eradication, when biosecurity knowledge is lost during staff/volunteer turnover, so maintaining a point of contact to ensure that continuity is maintained will be important for a successful long-term biosecurity programme in Scotland.
Predator-exclusion fences
Summary description
Electric fences can be set up seasonally around tern nesting areas as a temporary action to exclude foxes, although these require daily maintenance and clearing out of foxes that manage to cross the fence line (Short, 2020). This approach could be applied to any ground-nesting seabird colony where foxes have access to the area. Permanent predator-proof fences can be constructed that require little maintenance and are effective in excluding all mammal predators.
Key aspects of mammal-proof fences are a height of about 2m, wire mesh of a gauge of at least 1 mm to prevent foxes from chewing through it, and mesh size no more than 50 mm, with at least 500 mm buried horizontally at a depth of 100-150 mm below ground, water crossings with mesh to the base of drainage channels to prevent access by aquatic species such as otter, floppy overhanging top of 350-400 mm angled at approximately 45o to the outside to inhibit climbing (MacArthur Green and Royal Haskoning DHV, 2022). See also White and Hirons (2019).
Scope of potential action
This action would be effective to protect ground-nesting seabirds against mammalian predators. Bird species that would benefit with predator-proof fencing around their nesting areas include: terns and large gulls in particular as well as puffins, Manx shearwaters, storm-petrels, but potentially also cormorants, shags, black guillemots, razorbills and common guillemots that nest in boulderfields.
Geographic scope would most appropriately target mainland colonies of terns and large gulls where mammalian predation is limiting breeding success or breeding numbers, or where colonies have been lost as a result of predation pressure from mammals. It would be possible to establish mammal-free areas on islands too, potentially as an alternative to predator eradication in cases where eradication would not be possible or sustainable. There are examples of this overseas, for example predator-proof fence excluding cats from important areas of a petrel colony on the island of Corvo in the Azores.
Options
- Use of electric fencing to exclude foxes from seabird colonies subject to predation by foxes.
- Use of permanent fencing to create mammalian-predator-free areas for ground-nesting seabirds.
Delivery involvement
Land managers, particularly at bird reserves.
Funders and/or funding partners would be required to implement action.
Delivery mechanism(s)
Site based with overview from a strategic ‘Seabird Recovery Partnership’.
Timescales for delivery
Requirement for planning permission would set a limit on how soon a permanent fence could be established, but electric fencing can be set in place in a matter of hours or days depending on the scale of operation. Permanent fences can be constructed in a period of weeks (outside the breeding seasons of birds that are to be protected).
Dependencies/constraints
White and Hirons (2019) note that vandalism can be a significant issue with fences, especially where a few people consider the use of a fence to be an inappropriate limitation on their activities. There would be value in appropriate signage in such situations and monitoring of the integrity of fences. There would also be a need to consider other features of protected sites that might be affected by construction of a permanent fence. Some sites may have physical features making it impossible to place a permanent fence in the most appropriate position if anywhere.
Nest rafts for common terns
Summary description
Evidence indicates that common terns readily take to nesting on purpose-built rafts, and that productivity on these artificial colonies is usually higher than at natural colonies. Provision of predator-proof nesting rafts for common terns appears to be an effective way to increase productivity in this species in the UK. Such rafts are not inaccessible to avian predators, so provision of nest boxes and chick shelters on rafts would enhance these. Successful rafts for common terns have been deployed at Avoch and at various sites in Argyll.
Scope of potential action
Rafts could provide relatively safe nesting opportunities for common terns in locations where there are no natural sites or natural sites have been lost to erosion, habitat change or invasive predators. This action would work well for common tern, but tern rafts may be ignored by other tern species. Locations around Scotland can be indicated by changes in common tern breeding numbers in different areas based on Seabird Count (ref). However, local knowledge would most likely refine that.
Options
Provision of tern rafts at sites around Scotland where high quality natural nesting habitat is unavailable.
Delivery involvement
Land managers, particularly at bird reserves and local communities. Suitable locations might best be identified by local ornithologists. Crown Estate Scotland would have a role in licencing moorings of rafts in marine areas (which would require a marine licence for sea-bed lease and a Marine Works Consent), but there may be scope for establishing some rafts in freshwater lochs adjacent to the sea.
Funders and/or funding partners may be required to implement action.
Delivery mechanism(s)
Site based with overview from a strategic ‘Seabird Recovery Partnership’.
Timescales for delivery
Initially, suitable sites would need to be short-listed and ranked. Planning arrangements and licences would need to be arranged before rafts could be installed.
Dependencies/constraints
Access and maintenance issues would need to be resolved, as would licensing. Sites that appear to be suitable may not necessarily be adopted by terns, so there would be a risk that some rafts would not be used. Colonisation probability may be increased by use of dummy terns and playback of tern colony sounds. Sites that are successful may attract increased interest from predators, including protected species such as peregrine and kestrel, which can cause terns to desert, either temporarily or in some cases permanently. So the success of this action is not guaranteed in all cases. Success may be enhanced by combining actions such as tern nest boxes and chick shelters on rafts.
Tern terraces, nest boxes and chick shelters
Summary description
Tern terraces are areas of habitat managed to optimise an existing breeding site for tern nesting. Tern nest boxes were initially developed for roseate terns (Babcock and Booth, 2020a) but are useful for Arctic and common terns too. Sandwich terns prefer to nest in the open, but often use the nest box as a protection beside the nest (Steel and Outram, 2020). Tern terraces have been used successfully at the Isle of May. Provision of tern terraces on the Isle of May in 2015 not only increased numbers of common terns and Arctic terns nesting there, but also attracted roseate tern and Sandwich tern to recolonise the island (Steel and Outram, 2020).
Tern terraces provide the combination of suitable substrate (gravel and sand), nest box protection for birds that prefer that, chick shelters to provide protection against adverse weather and security for chicks from avian predators, bamboo canes to deter predatory attacks by large gulls, chicken wire fences on the boundary to prevent predators such as gulls and oystercatchers from walking into the tern colony and to stop chicks from running out of the colony (Steel and Outram, 2020). The high density of nesting terns also increases the ability of the tern community to defend itself from predators and that may be a reason for Sandwich tern being attracted to the tern terrace even though it may not choose to nest inside tern nest boxes.
Scope of potential action
The Isle of May could be used as a model of best practice for this action that could be implemented at other sites around Scotland. Tern terraces probably work best for common tern and Arctic tern, but may also attract Sandwich tern in some locations. They could attract roseate tern but this species is now extremely scarce in Scotland. This action might best be combined with creation of new artificial islets for terns at locations where there are no predator-free natural sites.
Options
- Creation of tern terraces on islands that are free from mammal predators and likely to support terns or held tern colonies in the past.
- Creation of tern terraces on newly constructed artificial islets for terns.
Delivery involvement
Land managers, particularly at bird reserves and local communities. Local bird clubs or branches of the Scottish Ornithologists’ Club would be well placed to identify likely sites for creation of tern terraces.
Funders and/or funding partners may be required to implement action.
Delivery mechanism(s)
Site based with overview from a strategic ‘Seabird Recovery Partnership’.
Site selection would be an important first step, as the locations where terns have nested around Scotland in the past are well known, and many of those local populations have been lost.
Timescales for delivery
The Isle of May example indicates that terns may start to use newly constructed tern terraces immediately, although the development of a larger colony at such sites may occur over the period of a few successive summers.
Dependencies/constraints
A key constraint would be locating suitable sites. Construction of tern terraces would need to be negotiated with the landowner, and sites would need to be ones where other potential impacts on terns (such as human disturbance, mammal and avian predators) would be minimised to reduce risk of the action being ineffective. It may be possible to combine tern terraces with use of islets or rafts to create new nesting habitat, and with use of predator-exclusion fences at sites with mammalian predators.
Nest boxes for black guillemots
Summary description
Black guillemot is vulnerable to mammalian and avian predators taking eggs, chicks, and breeding adults. Nest boxes provide black guillemots with better protection against predators than natural nests , and allow black guillemots to nest in locations where natural nest sites are lacking but other ecological conditions (such as food) are suitable for the species (Greenwood, 2015, Leonard et al., 2015). Black guillemots that nest on North Ronaldsay commute to the coast of Sanday to forage, suggesting that provision of nest box colonies at Sanday might allow a new local breeding population to develop. There may be many other cases similar to that elsewhere around the Scottish coastline.
Many designs of nest box have been tried, but there has not been a detailed assessment of the optimal design. Wooden boxes work well. These require a wooden ‘stop’ two-thirds of the way to the back of the box to stop eggs from rolling out. The box should also have a shelf at the front as black guillemots like a safe landing area and will sit outside the box (Leonard et al., 2015). British Trust for Ornithology (BTO) and Ards Borough Council have been deploying 80 new black guillemot nest boxes in the local authority areas of North Down and Ards, which has been a successful project that could be used as a good model for any future deployments of nest boxes.
Scope of potential action
This action reduces direct threats to black guillemots from mammalian and avian predators. Nest boxes can also be used to reduce pressures on black guillemots from climate change, human disturbance and disturbance by shipping. No other seabird species would be expected to benefit from deployment of black guillemot nest boxes as they are unlikely to be used by anything other than black guillemots.
Geographic scope for this action in Scotland could be anywhere from the west coast of Dumfries and Galloway to the coasts of the outer Moray Firth and the coast of Banff and Buchan. Sites such as the waterfront at Oban might be ideal as they would provide nest sites in an area without natural sites and allow black guillemot numbers to increase in a location where they can be enjoyed by local people and tourists. Similarly, there are many piers and jetties all around the coasts of west and north Scotland and the islands where black guillemot nest boxes could be incorporated. Sites close to human activity may be particularly suitable for black guillemot nest boxes because predators are less likely to approach nests when these are in areas (Johnston et al., 2019). Black guillemot nested for the first time on Inchmarnock in the Clyde after a nest box was provided there (Middleton, 1981), indicating the potential to extend the number of locations where this species can breed.
Options
Nest box deployment with monitoring of breeding success and any predation (the latter by trail-cam deployment) at sites selected for lack of nearby natural nesting habitat for black guillemot.
Delivery involvement
Land managers and local communities.
Skills required include experience in positioning nest boxes for black guillemots and box design to optimise black guillemot nest success. This may best be obtained from practitioners such as BTO and local ornithologists who have deployed black guillemot nest boxes in locations such as Northern Ireland (BTO, 2023) and Arran. Monitoring would best be done by local ornithologists, and recording of predation attempts could be done by deployment of trail cameras at nests (see Johnston et al., 2019).
Funders and/or funding partners may be required to implement action.
Delivery mechanism(s)
Site based with overview from a strategic ‘Seabird Recovery Partnership’.
Site selection could be influenced by locations with interested local ornithologists who could manage a small programme of nest box deployment and monitoring. Black guillemots nest in small, loose, colonies and are social, especially near the breeding area in late winter and spring. Natural colonies in Scotland are often of a few tens of pairs. Provision of sets of 20 nest boxes might therefore be an appropriate start, with the possibility of adding more boxes if uptake is high.
Site selection could be based largely on opportunity, but there is some evidence in relation to potential sites. The national seabird censuses indicate where the main natural aggregations occur. Gaps in that distribution might indicate areas lacking natural sites.
Providing nest sites for black guillemots could form a useful part of the ecotourism of a location and an educational opportunity.
Timescales for delivery
Nest boxes could be deployed at short notice during the early winter (because black guillemots return to establish nest sites very early in the year). It is likely that nest boxes would be taken up by black guillemots very soon after their deployment, although their breeding success tends to be poor in their first year of breeding (Leonard et al., 2015). Building up larger numbers may take several years as black guillemots only fledge about 1 chick per nest and birds do not start to breed until about five years old (Horswill and Robinson 2015).
Dependencies/constraints
Nest boxes would need to be placed in a way that did not interfere with the normal functioning of the structure (such as a pier or jetty) on which it was placed. There could also be issues if structures are listed and so cannot be modified without planning permission.
Nest boxes for petrels
Summary description
Nest boxes for petrels have been used successfully at several colonies including Mousa (Shetland), St Agnes and Gugh (Isles of Scilly), Skokholm (Wales), and Benidorm Island (Spain) (Bolton 1996, de Leon and Minguez 2003, Milborrow and Murphy 2008, Libois et al., 2012, British Birds 2019). Breeding success in nest boxes at Mousa was the same as in natural sites on Mousa (Bolton, 1996). However, at Benidorm Island breeding success and adult survival were both higher in nest boxes compared to natural sites (de Leon and Minguez, 2003; Libois et al., 2012), and provision of nest boxes there resulted in a large increase in breeding numbers on the island which had been subject to high predation rates by gulls before the nest boxes were installed. Nest boxes also allowed higher breeding success by Madeiran storm-petrels than achieved in natural sites (Bolton et al., 2004) so is advocated as a conservation action for petrels in general.
At Skokholm, a wall has been constructed with 100 nest chambers with inspection lids built into the wall, allowing development of a storm-petrel colony that can easily be monitored (British Birds, 2019).
Scope of potential action
This action directly reduces threats to European storm-petrel and Leach’s petrel from mammalian and avian predators. Nest boxes could also allow petrels to breed successfully in locations where natural nest sites are unavailable. They could therefore enhance resilience in relation to other pressures on European storm-petrel and Leach’s petrel. Both species could benefit at sites where natural nest sites are limited or unavailable.
Geographic scope would be throughout Shetland, Orkney, the Western Isles and islands off the west coast of Scotland, especially in locations where European storm-petrels and Leach’s petrel are unable to breed in natural sites due to presence of mammalian predators.
Options
- Nest boxes built into existing coastal boulderfields.
- Nest boxes built into existing coastal walls.
- New walls with built-in nesting chambers, as created at Skokholm (British Birds 2019).
Delivery involvement
Land managers, local communities and academia.
Requires understanding of storm-petrel nest structure that provides safety from predators (based on experience at locations such as Skokholm and Benidorm Island). Monitoring of use and breeding success achieved would be a high priority so the projects should be linked with where such services could be established. One of the key opportunities these would provide is to be able to monitor adult survival and breeding success of storm-petrels, which are currently very poorly known and for which there are no adequate long-term time series.
Funders and/or funding partners would be required to implement action.
Delivery mechanism(s)
Site based with overview from a strategic ‘Seabird Recovery Partnership’, with a strong presumption that selected sites would be monitored on a long-term basis.
Timescales for delivery
Construction may be possible within one year, but it may require some years for numbers of storm-petrels using nest boxes to build up. First breeding attempts of seabirds are usually less successful than attempts by experienced/established pairs, so data on breeding success would be affected for the first few years by the slow process of colonisation of new sites. Monitoring of long-term breeding success and adult survival may need to discard data for the first few years of a project.
Dependencies/constraints
There are very few sites where Leach’s storm-petrels breed in Scotland, but there might be scope for restoring breeding at sites where this species has been lost, apparently affected by mammal predation (e.g. Foula, Shetland).
New artificial nest sites for kittiwakes
Summary description
Kittiwakes readily nest on artificial sites, including on window ledges of warehouses, window ledges of offices and houses, on church towers, on piers and jetties, on harbour walls, ruined castle walls, on stone bridges, metal girders, and on offshore structures such as oil and gas platforms. Whilst appropriate natural sites are not limited in Scotland breeding success on artificial sites tends to be higher than at nearby natural colonies (Christensen-Dalsgaard, Langset and Anker-Nilssen, 2019), possibly because predation impacts are lower at ‘urban’ sites and possibly because colony size is generally smaller at artificial sites so there may be less intra-specific competition and there is little or no inter-specific competition. Highest success is seen on offshore platforms, which have the additional benefit of being close to foraging grounds so reduce flight time of breeding kittiwakes (Christensen-Dalsgaard et al., 2019). Most artificial structures used by kittiwakes however are ones they have adopted in areas lacking natural nesting habitat (sea cliffs with narrow ledges) which is not the case in Scotland.
Scope of potential action
The use of artificial nest sites is an action that could be used to enhance the resilience of kittiwakes in Scotland. It would not benefit any other seabird species.
Geographical scope could potentially be any part of the Scottish coast where the coast is open to the sea, or in offshore waters. However, the fact that there is a large amount of natural habitat in Scotland for kittiwake nesting suggests that artificial nest sites in Scotland would be much less worthwhile than in England. There could be merit in considering offshore artificial nest sites for kittiwake in Scotland since the (limited) evidence available indicates that kittiwakes nesting on offshore artificial nest sites achieve higher breeding success. Where kittiwake populations have declined due to pressures from climate change, depletion of sandeel stocks and other factors, offshore artificial nest sites may be an action that could improve demography of kittiwakes to stop population decline. The success of the Hornsea offshore wind farms offshore artificial nest sites (which would be monitored as a requirement of the planning consent of that project) could inform whether the gain in productivity achieved from offshore artificial nest sites could justify such an action being considered in Scotland.
Options
- Artificial nest sites for kittiwake at sites on the Scottish coast.
- Offshore artificial nest sites for kittiwake in Scottish waters.
Delivery involvement
Local Planning Authorities, land managers, Crown Estates, consultancies and academia.
Artificial nest sites would require specialist design and build with an appropriate monitoring scheme in place to determine success.
Artificial nest sites at a coastal site would require planning permission and land ownership or landowner lease consent.
An offshore artificial nest sites would require Crown Estate Scotland licencing.
Funders and/or funding partners would be required to implement action.
Delivery mechanism(s)
Strategic with an overview from a ‘Seabird Recovery Partnership’ and a strong presumption that artificial nest sites would be monitored on a long-term basis.
Timescales for delivery
Kittiwakes may not adopt a structure that is created for them. Alternatively, colonisation may take some years, especially in regions where kittiwake populations have low breeding success so are not producing large numbers of young birds that will be seeking potential nest sites. Productivity gains from artificial nest site will only start to increase breeding numbers of the regional population about five years later, as kittiwakes do not start to breed until about 5 years old.
Dependencies/constraints
One option could, in theory, be to use existing offshore structures that are due to be decommissioned in order to add suitable ledges for kittiwakes. However, there are issues with legal ownership of offshore structures and decommissioning requirements that may make adaptation of existing structures impractical.
Education programmes to raise public support for seabird conservation
Summary description
Education would improve public attitudes to seabird conservation, both in terms of reducing impacts such as caused by fishing waste and ingested plastic, but also appreciating the need for actions to reduce impacts on less popular seabirds such as large gulls.
Scope of potential action
There are already some excellent examples of good practice in this regard, including the Scottish Seabird Centre in North Berwick, the Biosecurity for LIFE project, and the BBC series with Sir David Attenborough looking at wildlife in Britain. Further examples could help to improve public understanding which could reduce impacts of human disturbance and persecution of seabirds and improve support for seabird conservation actions. Increased public support would be important to achieve many of the options listed in this report, so should be seen as an important interacting action that facilitates many of the conservation actions being suggested. All seabird species would benefit from an improved public understanding of their conservation challenges, with the geographic scope being throughout Scotland. The lack of understanding is remarkable and can be difficult for those regularly immersed in seabird ecology to understand. For example, Channel Four’s Gogglebox recently showed several families to be amazed to learn from the Attenborough series that there are puffins nesting in Britain. Most people do not realise that kittiwakes are different from herring gulls and are not an urban nuisance or scavenge on urban waste.
Options
- Educational displays (e.g. panels) and apps explaining the action being put into practice at a particular location and why it is important for conservation.
- Education through school and university teaching programmes.
- Education through popular science journalism.
- Education through co-design using a youth work approach (Boyle & Copley, 2022).
- Education through volunteering participation in citizen science monitoring of seabirds.
- Education through volunteering to carry out actions that reduce impacts on seabirds.
- Targeted training on biosecurity and safe release of seabirds from nets and boat/land strandings.
Delivery involvement
Scottish Government, NatureScot, NGOs, education, communications and media professionals, tourism, local communities and academics.
Education communication skills would be crucial. The programme would require close coordination between those with communication skills and the conservation practitioners involved in each of the actions being taken forward.
Delivery mechanism(s)
Strategic with potential overview from a strategic ‘Seabird Recovery Partnership’. Communications plans and campaigns should be developed. Television programmes might be one of the most powerful ways to get across messages about seabird conservation and actions to achieve this. However, there would also be scope for site-based options, especially through display panels at key sites, and through community-based action.
Timescales for delivery
This programme should be carried out alongside conservation actions, or in advance of actions that may meet public resistance to ensure there is good public understanding of the actions and why they need to be undertaken.
Dependencies/constraints
Many of the actions would depend for their success on public support, which may be lacking in particular cases or from particular stakeholder groups, so the overall success of actions would be likely to be strongly influenced by the quality of education programme accompanying these.
Strategic selection of areas for marine renewable development
Summary description
Areas made available for developers to propose construction of offshore wind farms are already subject to strategic spatial planning through the ‘Sectoral marine plan for offshore wind energy’, with numerous interests needing to be taken into account. These interests include likely impacts on seabird populations and ensuring future development avoids areas where vulnerable seabird densities are high.
Scope of potential action
Future updates to the Sectoral plan should continue to select areas as distant as practicable from major seabird colonies for species that are considered to at high risk of collision. Foraging hot-spots used by seabirds such as frontal systems where prey tend to be aggregated, or over sandbanks that support high densities of sandeels should also be avoided.
Delivery involvement
Scottish Government, Crown Estate Scotland, Offshore wind industry, NatureScot and NGOs.
Delivery mechanism(s)
Strategic, based on the known spatial distribution of the largest seabird colonies around Scotland.
Timescales for delivery
Development of the next Sectoral marine plan for offshore wind energy is in progress.
Dependencies/constraints
To deliver net zero energy targets, Sectoral Marine Planning should consider and minimise impacts on seabirds through use of mitigation and where required compensation. Similarly for individual projects where adverse impacts cannot be avoided, they should be mitigated for and where necessary compensation measures implemented.
Reduce human disturbance of seabirds
Summary description
Disturbance of breeding seabirds can occur when people visit areas where seabirds nest. Often such disturbance is unintended and results from people enjoying the countryside without realising their effect on nesting birds. Tern colonies on mainland sites are especially vulnerable to human disturbance. Little terns nesting on beaches are often not noticed by people. Island seabird colonies can also be vulnerable. Disturbance can be reduced by signage, by education programmes, and by actions including fences that redirect paths away from vulnerable seabirds. Visitor management during the breeding season of vulnerable seabirds provides a more powerful means of reducing risk of human disturbance. Short (2020, p.16) notes that human disturbance of nesting terns at Forvie remains an ongoing problem, despite signage and the electric fence across the peninsula and states “In most cases, incursions were due to people not noticing the signs, or not understanding the signs (in the case of international visitors), and only in a very few cases were the access restrictions deliberately ignored. Light disturbance was noted on occasion from walkers, anglers and kayakers. Staff and volunteer presence remains an important factor in protection of the colonies from predation and disturbance.”
Scope of potential action
Reducing human disturbance represents a potentially valuable action to help several seabird species. Species that could particularly benefit from this action include terns, but there are specific cases where many seabird species could benefit from reduced human disturbance at specific sites. For example, fulmars can be very susceptible to disturbance at the nest. Even seabirds thought to be rather resilient to human disturbance, such as urban-nesting kittiwakes, can be affected by high levels of human disturbance at parts of colonies such as Dunbar Harbour and St Abb’s Head. Geographic scope is throughout Scotland, but particularly at a small number of specific seabird colonies. This action could be combined with other actions, such as “education”, “nest rafts” and “create islets for terns and gulls”.
Options
- Provide information at sites (e.g. signs) to encourage people to avoid walking into areas used by nesting terns.
- Encourage responsible access by e.g. re-routing footpaths and placing fences to discourage people from walking into areas used by nesting terns.
- Provision of hides for viewing birds without disturbance, potentially also using cameras that give close-up live images of the colony.
- Promote the SMWWC.
- Provide seasonal warden/ranger positions to encourage responsible behaviour and avoid disturbance.
- Developing best practice guidance for recreational user groups.
Delivery involvement
Land managers, particularly at bird reserves, local communities, communications professionals and targeted engagement with site-specific recreational users.
Delivery mechanism(s)
Site-based with potential overview from a strategic ‘Seabird Recovery Partnership’.
Timescales for delivery
Site-based actions could be implemented immediately but a programme of action would benefit from a longer-term strategic approach.
Dependencies/constraints
Tern colonies vulnerable to human disturbance will be at risk of being lost, so actions would need to be put in place at sites that are considered to be sustainable providing human disturbance can be managed.
Manage shipping disturbance of seabirds
Summary description
Disturbance by ships/boats is unlikely to be a major impact on seabird populations in most parts of Scottish waters but may have some local impacts that could be countered by regulating vessel activity at sensitive sites and during sensitive times of year. This might be especially relevant where vessel traffic is increasing considerably.
Scope of potential action
Fliessbach et al. (2019) ranked black guillemot, common guillemot, razorbill, cormorant and gannet as particularly vulnerable to impacts of disturbance by vessels. Reducing vessel activity would therefore be an action to directly benefit those species. Geographic scope would cover high at-sea concentrations of those species where they are affected by high or increasing levels of vessel activity. This may be especially in the vicinity of the largest colonies of these species during the breeding season.
Options
- Constraints on vessel movements in areas of high at-sea concentrations of seabirds associated with major seabird colonies during the breeding season.
- Promote the Scottish Marine Wildlife Watching Code to vessel owners.
Delivery involvement
Scottish Government, Local planning and Harbour authorities and NatureScot.
Delivery mechanism(s)
Requirement for approved vessel movement management plans included as conditions for planning and licence consent.
Site-based, at selected major colonies of black guillemot, common guillemot, razorbill, cormorant and gannet.
Timescales for delivery
Not limited.
Dependencies/constraints
Acceptance of such an action (with its implied costs for vessel operators) would require a more secure evidence base that quantifies gains to seabird conservation that could be obtained by reducing disturbance by vessels. This may be a limited gain that is not easy to quantify.
Manage airborne vehicle disturbance of seabirds
Summary description
Disturbance to breeding seabirds may occur when unmanned aerial vehicles (UAVs), which are increasingly being used for filming and photographing wildlife, or aircraft and helicopters are flown too close to seabird colonies causing birds to leave suddenly, known as “flushing” events. Eggs and chicks may be left exposed, damaged, or displaced if adult birds at the colony flush due to disturbance, which can increase the chance of predation. This source of disturbance is unlikely to have major impact on seabird populations in most parts of Scottish waters, although there may be some local impacts that could be countered by regulating the activity of these airborne vehicles at sensitive sites and during sensitive times of year.
Scope of potential action
The action of reducing airborne vehicle disturbance is most likely to benefit cliff nesting and ground nesting seabirds that are generally vulnerable to human disturbance including cormorants, auks, gulls, gannets, and terns. Even for potentially sensitive species, the impact of airborne vehicle disturbance on breeding birds may depend on the intensity and direction of the disturbance and can vary widely between different colonies. NatureScot recommend that drone flights take off at least 100 meters away from seabirds (and any cliffs containing seabirds), as well as keeping a minimal flight height of 50 meters (up to 100 meters for larger types of UAVs), and establishing a minimal distance of 50 meters away from seabirds and 200 meters from large seabird flocks.
Reactions of birds to helicopter disturbance and noise can also be highly variable between species and also within the same species; actively breeding birds with eggs and young chicks are less likely to flush than non-breeding birds (Anderson, 2007 and Brisson-Curadeau et al., 2017). Relatively few studies have investigated helicopter disturbance at seabird colonies, further research is required. A safe working distance for aircraft in Scotland over protected breeding raptor nests is considered to be at least 500 meters in altitude above the sensitive location; Scottish Natural Heritage (2015). NatureScot (2017) guidance recommends that a minimum of 300 meters altitude is maintained when close to colonies or large groups of birds.
Options
- Guidance on the use of UAVs at seabird colonies to minimise disturbance to seabirds.
- Provide information (e.g. through using signs) to highlight the potential dangers of disturbance in areas where people use UAVs near seabird colonies. Work with local bird clubs and NGOs to identify priority sites (such as popular recreational areas near important seabird colonies) where signs should be installed first.
- Education about the impacts of human disturbance to wildlife through school and university teaching programmes.
- Education through volunteering to carry out actions that reduce impacts on seabirds.
- Greater enforcement of no-fly zones for helicopters flying over seabird colonies.
Delivery involvement
NatureScot, NGOs, bird clubs, academic’s and general public.
Control of UAV use would best be done by local groups (NGOs and bird clubs) taking responsibility for local sites. Restriction of helicopter movement over seabird colonies would probably require use of bylaws to regulate aircraft activity in specific zones close to seabird colonies.
Delivery mechanism(s)
Strategic, implemented at all designated seabird colonies.
Timescales for delivery
Not limited.
Dependencies/constraints
For UAV use, seabird colonies that are vulnerable to human disturbance potentially could be at risk of being lost if disturbance is great enough, so actions would need to be put in place at sites that are considered to be sustainable providing human disturbance can be managed.
For aircraft restriction, acceptance of such an action (with its implied costs for aircraft operators) would require a more secure evidence base that quantifies gains to seabird conservation that could be obtained by reducing disturbance by aircraft. This may be a limited gain that is not easy to quantify.
Reduce artificial light at sites near storm-petrel, shearwater and puffin colonies
Summary description
Actions to reduce the extent to which artificial lights attract or disorientate seabirds. Disorientation by lights during foggy weather could affect a wide range of seabird species, but is a rare event, and seems most often to affect procellariiforms. Attraction to artificial lights is especially an issue with storm-petrel, shearwater, and puffin fledglings.
Scope of potential action
Reducing artificial light in the marine environment is an action that could directly benefit procellariiforms. Geographic scope of this would mostly be in the areas close to major colonies of storm-petrel, Manx shearwater and puffin. The main sites in Scotland of concern would be St Kilda (storm-petrels, Manx shearwater and puffin), Rum (Manx shearwater), Shetland and Orkney, North Rona, Flannan Isles, Priest Island, Treshnish Isles, Sanda (storm-petrels and puffin) and Isle of May (puffin).
Options
The risk to seabirds from attraction to lights or disorientation could be reduced using the following options:
- Shielding lights to minimise light spill upwards.
- Using selected wavelengths of light that are less likely to attract birds (but at present it is uncertain which wavelengths cause greatest or least effect).
- Using flashing lights rather than continuous lights where that would be appropriate.
- Requiring ships in the area to cover windows with blinds.
- Training boat crew and local communities where strandings are an issue to safely capture and release stranded birds.
- Switching lights off at times of year of particular concern.
Delivery involvement
Where lighting is a legal requirement for safely, discussions with regulators may be required to investigate whether there would be options to permit use of selected wavelengths, or flashing lights or fewer lights. To allow the development of effective management actions, there is a need for research by seabird ecologists to assess the extent of the problem in Scottish waters and the effectiveness of alternative solutions.
Delivery mechanism(s)
Probably site-based rather than strategic, with emphasis on the surroundings of the largest colonies of storm-petrels, Manx shearwaters and puffins.
Timescales for delivery
Because of high uncertainty about impacts and efficacy of different potential methods to reduce artificial light, delivery of effective management actions may require some years of research.
Dependencies/constraints
Some lights are necessary for safety. The extent of the problem of attraction to artificial lights in Scottish waters is uncertain, so an assessment of the extent of this would help to allow better targeting of management actions. The behavioural responses of storm-petrels, Manx shearwaters and puffins to different wavelengths of light are not well understood. Better understanding of this could help determine whether choice of wavelengths of light could help reduce the impacts.
Reduce marine litter
Summary description
Reducing amounts of fishing gear ending up as marine litter and removing ALDFG gear would reduce impacts of plastic waste on seabirds.
Scope of potential action
This action would reduce the minor impact of plastic entanglement on seabirds, principally on gannets, large gulls, procellariiforms, shags and cormorants. Geographic scope would be throughout Scottish waters.
Options
- Education programme to identify the welfare issue caused by entanglement and the way to reduce this.
- Levy on the fishing industry to cover the costs of removal of ALDFG from the seabed and fragments of fishing gear from the shoreline.
- Projects to recover ALDFG from the seabed and fragments of fishing gear from the sea surface and shoreline, or to prevent plastic from getting from rivers into the sea.
Delivery involvement
Education programmes should target schoolchildren, the general public, commercial and sport/recreational fishers. Political action would be required to establish a levy on the fishing industry to cover the costs of removing ALDFG and fishing gear litter (following the polluter pays principle). Projects to recover ALDFG gear would require involvement of conservation NGOs such as Wildfowl & Wetlands Trust Limited (WWT), appropriate method of recovery, and relevant public authorities (following the example of Stolte et al. (2022)). Projects to recover beach-cast fishing debris could build on existing beach-cleaning volunteer programmes.
Delivery mechanism(s)
Strategic.
Timescales for delivery
The educational programme could be started at any time and would likely need to be run for at least ten years. Recovering fishery debris would need to await development of a levy to raise funds to carry out the clean-up work.
Dependencies/constraints
Recovery of ALDFG from the waters around Scotland would be more difficult than the work recently done in the Baltic Sea (Stolte et al., 2022) as Scottish waters tend to be deeper than the coastal Baltic waters, with rougher sea and stronger currents. Use of Remotely Operated Vehicles might be necessary rather than deployment of divers.
Vegetation management at seabird colonies
Summary description
Management of grazing intensity above puffin colonies to maintain a sward of short grass with strong root development to minimise erosion and undermining.
Scope of potential action
This action could directly be applied to target grazing pressures at puffin colonies. Geographic scope would be limited to any puffin colony where growth of vegetation over the colony was having an impact on puffins, or where erosion was causing loss of nesting habitat. This may apply to few colonies, but there have been problems of vegetation growth affecting puffins (for example on some of the Forth Islands), and some puffin slopes have eroded after heavy burrowing activity by increasing breeding numbers of puffins at particular colonies, as at Grassholm, Wales, and at part of Dun, St Kilda (Mitchell et al., 2004). The west slopes of Eilean Mor in the Flannan Isles ae subject to heavy erosion and loss of soil cap due to burrowing by the island’s dense rabbit population.
Delivery involvement
Getting grazing levels correct would require skills of agricultural experts in grazing who would have to work with the owners and managers of sites where this action was to be implemented. Monitoring and adaptive management would be an important part of this.
Delivery mechanism(s)
Site-based with potential overview from a strategic ‘Seabird Recovery Partnership’. However, likely to be limited to very few sites.
Timescales for delivery
Not strongly constrained in terms of timescale.
Dependencies/constraints
Agreement between landowners, individuals with grazing rights (if any), and conservation managers would be required. Managing livestock could be a constraint if animals need to be moved to/from sites and it would be necessary to consider any possible impacts of grazing animals on other protected features of sites (especially including vegetation features).
Create islets for terns and gulls
Summary description
Creating a pool with sand and gravel islets in the pool will provide new nesting habitat suitable for nesting by terns or gulls. A good example of this in Scotland is St John’s Pool, Caithness (Hughes, O’Hanlon and Smith, 2021). Another good example elsewhere is the creation of islets for terns and gulls in the south of France, where 154 such sites were created (Schwartz et al., 2022). The French example selected sites based on presence of suitable foraging habitat and resources, presence of water resource allowing maintenance of water around the created islet to secure colonies from terrestrial predation, absence of uncontrolled human disturbance, past presence of breeding gulls or terns in the general area, absence of major predator populations in the immediate area, absence of an airport in the vicinity. The created islets averaged 63 m2 (range 2 to 1,400 m2), distance to shore from 9 m to 270 m (median 45 m), water level between islet and shoreline from 0.1 to >1 m, median 0.3 m, slope of islet surface of about 10% (Schwartz et al., 2022 online supplementary material). There may be scope for creative use of dredge sediment waste in such a programme.
Scope of potential action
This action would reduce impacts of nesting habitat loss (whether through habitat change, competition with other species, or unsustainable predation pressure) on tern and gull species. Common tern, Arctic tern and Sandwich tern would be the species most likely to benefit. Large gulls could benefit from large developments, but large gull species would be less likely to colonise small ponds and islands. This action would be most successful if the aim was to specifically benefit either terns or large gulls, but not both together. Islets in ponds are likely to be colonised by black-headed gulls and possibly by common gulls (species not considered in this review because they are predominantly terrestrial rather than marine) but colonisation of islets by terns, especially by Sandwich terns, may be more likely if black-headed gulls have already colonised. Geographic scope could be throughout coastal Scotland, with a particular suitability of low-lying land in sea lochs or in coastal bays of mainland or low-lying islands. Selection criteria similar to those designed in France (Schwartz et al., 2022 online supplementary material) would be appropriate to short-list potential sites. The Sheringham Shoal and Dudgeon Extension offshore wind farm planning application includes plans to create a pool and islets at Loch Ryan, near Stranraer, to provide nesting habitat for Sandwich tern and possibly other species (Equinor, 2022).
Options
- Creation of small coastal pools with islets for tern and small gull nesting.
- Creation of large coastal pools with larger islets designed for large gull nesting.
Delivery involvement
Scoping potential sites could be done with local ornithologists and community groups. There would most likely be more potential sites than funding could develop, so some selection should follow principles similar to those used in the south of France.
Delivery mechanism(s)
Strategic selection of sites, but then a site-based approach with national coordination of the project. It might be appropriate to have a pilot project to demonstrate the effectiveness and to learn lessons in relation to design and implementation in Scotland.
Timescales for delivery
Probably requires one to two years for short-listing potential sites, a further two or three years for a pilot trial at a few sites but further sites could be rolled out over time as appropriate.
Dependencies/constraints
In addition to issues of land ownership or lease, planning permission would be required in order to construct and maintain an inland pool as well as a controlled activity regulations licence from SEPA for abstraction and/or impounding of freshwater. Any works below mean high water springs which might be necessary for a pool fed by seawater on a tidal basis would require a marine licence from Marine Scotland and possibly a Marine Works Consent and sea-bed lease from Crown Estate Scotland.
This action could be combined with the use of predator-proof fence around pools holding newly created islets to further reduce risks from mammal predators and human disturbance.
Reduce harvest of gannet chicks
Summary description
Gannet chicks are harvested as food at colonies in Iceland, the Faroes and Scotland. Sula Sgeir is the only colony in Scotland at which an annual harvest of gannet chicks is allowed.
Scope of potential action
This action would involve reducing the number of gannet chicks harvested under licence. Harvests of gannet chicks are cultural traditions, but an agreement to restrict the scale of this harvest could be used as a mechanism to reduce impacts on gannets.
Geographic scope in Scotland is limited to Sula Sgeir, but because immature gannet prospect possible nesting opportunities at many colonies throughout the species’ range in the eastern North Atlantic, increased production of chicks at Sula Sgeir would be expected to benefit gannet colonies over a wide geographic area but probably especially those closest to Sula Sgeir.
Delivery involvement
NatureScot and the local community.
Delivery mechanism(s)
Site-based through existing licencing process.
Timescales for delivery
A voluntary suspension by the community took place between 2022-2024 due to HPAI. Should the local community wish to reinstate the harvest, reductions in the total allowable harvest further to what is considered sustainable would require stakeholder engagement.
Dependencies/constraints
A gannet chick that survives to be large enough to be harvested is almost certain to survive to fledge, as chick mortality is low in gannets and mostly occurs among very young chicks. However, mortality of fledglings is high. Horswill and Robinson (2015) provide best estimates of demographic parameters of UK gannets as juvenile survival 0.424, and immature survival to breeding age 0.61. Based on these estimates, 25.8% of chicks i.e. one in four chicks will reach breeding age and therefore are available to recruit into a colony.
Reduce lethal control of large gulls
Summary description
Licensed control of adult gulls (or prevention of breeding which can also impact on population dynamics) was permitted under the General Licences, but since April 2020 now requires a specific licence and subsequent reporting of numbers lethally controlled. Numbers licensed for lethal control of adults and chicks from 2020 onwards are now published online, however the taking of nests and eggs are not currently publicly published. The impact of licenced control on large gull populations remains unclear.
Scope of potential action
This action reduces the impact of licenced control on populations of large gulls. Populations of herring gull, lesser black-backed gull and great black-backed gull would benefit (as would common gull and black-headed gull populations which are also subject to licenced control, but which were not considered as part of this review). Gains could be throughout Scotland, as these gull populations are meta-populations with extensive movement of young birds reared in one colony to breed in other colonies, generally nearby but potentially throughout the country. This action could also be used to alleviate other pressures affecting large gull populations, most especially in their more natural-nesting populations. There would also be an advantage to establishing several protected sites for large gulls to breed so that those new sites could take up a significant part of the recruitment of young gulls to reduce the pressure on urban colonies in particular.
Options
- The licensing regime can transition to a presumption against issuing licences except under specific and well-defined circumstances.
- Population Viability Analysis (PVA) of large gull populations (individual colonies, regional, and national populations) with comparison of counterfactuals where licenced control is or is not occurring in the model population (at levels indicated by licence data) to assess the role of licenced control in large gull population trends.
- Increasing emphasis on preventative actions (such as non-lethal methods of deterrence) and ensuring that in the planning process for new buildings the prevention of unwanted birds nesting is taken into consideration at the design stage.
- Increased public and stakeholder engagement around gulls, especially in those areas where gull conflict is perceived, is required alongside the implementation of a reduced lethal control of gulls.
- Regular monitoring of gull populations, especially in non-traditional colonies such as urban environments.
Delivery involvement
NatureScot as the licencing authority. PVA modelling should be carried out by researchers with PVA modelling expertise and preferably with experience of PVA models for colonial seabirds. The modelling would need to be done in close collaboration with NatureScot licensing team and ornithologists to ensure that the best available data on control levels can be incorporated into the modelling. For consideration of gull population surveys in non-traditional colonies, working with organisations with experience of robust bird survey designs would be required. Stakeholder engagement would be required to enable a transition to a presumption against issuing licences except under specific circumstances.
Delivery mechanism(s)
Strategic, considering the whole of Scotland, but with a focus on regional populations of gulls most likely to be impacted by current and recent control levels.
Timescales for delivery
NatureScot commenced a transition to a presumption against issuing licences in 2022 with the aim for full implementation by 2024, further to discussions with stakeholders. PVA modelling should be carried out immediately based on recent population data and current licensing numbers. That could report within one year, to inform the future strategy in terms of licensing. Designing and implementing a survey protocol for non-natural nesting populations of gulls should be a priority, particularly in those areas where higher levels of conflict are perceived.
Dependencies/constraints
Reducing the numbers of birds licensed for lethal control may be challenging for some stakeholders, and therefore early stakeholder engagement to explain any changes to the licensing regime is necessary.
Demographic monitoring of seabirds
Summary description
Impacts on seabirds, or the management of impacts through direct or indirect conservation actions, will be seen in terms of changes to demographic rates. For this reason, monitoring of demographic rates likely to be affected by impacts or conservation actions should be a high priority. These include breeding success and adult survival rates as a minimum, in addition to monitoring of breeding numbers. There is already a Seabird Monitoring Programme and defined Key Sites for Demographic Monitoring, but that programme could be expanded to include more species and a more representative set of sites around Scotland.
Scope of potential action
The aim of this action should be to monitor breeding success and adult survival of all species of breeding seabird, preferably at several colonies distributed around Scotland. However, for some species this would be impractical so a pragmatic approach is needed. Analysis of time series should help to evaluate impacts of particular pressures and the success or otherwise of conservation actions. More detailed monitoring (including measurement of age of first breeding, frequency of nonbreeding by adults, and immature survival) would be possible for a few species at selected colonies (such as the Isle of May). There should be particular emphasis on monitoring that informs policy in terms of conservation actions. Existing monitoring is very valuable in that context but is not sufficient at present.
Options
- Maintain the long-term monitoring at the Isle of May to ensure that this time series is continued.
- Develop long-term monitoring to extend to species not adequately monitored at present.
Delivery involvement
This requires a coordinated approach with inputs from BTO, RSPB, JNCC, NatureScot and Scottish Government. Long-established local ringing groups and bird observatories also have a valuable role to play in maintaining long-term monitoring of adult survival.
Funders and/or funding partners would be required to implement action.
Delivery mechanism(s)
Strategic with overview from the Seabird Monitoring Programme.
Timescales for delivery
This is a high priority as the evidence should underpin policy and at present the evidence is not adequate to do so for most seabird species in Scotland.
Dependencies/constraints
Funding would need to be provided, and to ensure continued data collection to maintain time series, funding would need to be assured into the future.
Part 3. Recommendations on potential actions and priorities
Many very different possible actions are listed in Part 2. All of these could have a place in actions to improve the conservation status of seabirds in Scotland, but only a few of these actions have potential to have a particularly large effect. These include eradication of invasive mammals from seabird islands and habitat management to create safer nesting areas.
Eradication of invasive mammalian predators from seabird islands also has scope for large gains for several species of seabird in Scotland. The methodology is well developed and effective, and gains for seabird populations have been demonstrated clearly, especially for burrow-nesting seabirds such as storm-petrels, Manx shearwater, puffin, black guillemot and for ground-nesting seabirds such as terns, large gulls, shags. There may be some particular sites where vulnerable seabirds can coexist with invasive mammalian predators under some ecological conditions (possibly this is the case at Rum for Manx shearwaters and rats under the current conditions but that might not be the case in future if ecological conditions alter for example through climate change, or epidemics such as avian influenza), and there may be some specific colonies where invasive mammals have impacts on seabirds that are generally thought to be at low risk. For example, negative impacts on kittiwakes nesting close to the ground on structures that rats can easily climb, as on part of the castle walls at Dunbar Harbour (Skeate, 2022). This requires careful consideration of site-specific issues when evaluating the relative merits of eradication programmes at different sites. This action would be most effective if carried out as a strategic programme with oversight from Government (for example through a Seabird Recovery Partnership) with costs of individual site-specific actions being provided by a variety of stakeholders contributing to the overall programme. That would ensure that action was focused on top priority sites to give the best value outcome. Putting biosecurity actions into place for sites that have been cleared of invasive mammalian predators would also need to be an integral part of that programme.
Habitat management to create safer nesting areas for terns and gulls seems also to offer considerable potential gain. Creating islets for terns and gulls has been demonstrated to be effective in attracting these birds to breed in areas where they were otherwise constrained by the presence of mammalian predators or human disturbance. A combination of habitat creation (coastal ponds/lagoons with islets for seabird nesting), with predator-exclusion fence surrounding to protect the colony from mammal predators and human disturbance, could also have a viewing hide integrated into the site for public education/ecotourism. Such sites could also be made more resilient to avian predators by including tern terraces where the aim is to restore tern breeding. Again, this programme would be most effective if done as a strategic programme with oversight by Government, but with sites suggested for short-listing by local stakeholder and community groups.
Nest boxes have been demonstrated to be effective for some particular seabird species. This is especially the case for black guillemot, where nest boxes allow breeding birds better protection against predators (mammalian and avian),and allow nesting in areas that lack natural nest habitat. Nest boxes probably also improve European storm-petrel and Leach’s petrel safety from predators. Nest boxes also open opportunities for monitoring demography of species that currently lack demographic monitoring because birds in natural nest sites are not readily monitored. This might also allow better monitoring of Leach’s storm-petrel, a species now of global conservation concern and in serious decline in Scotland.
There is a shortage of evidence on the impacts of fisheries set nets and long-line fisheries on seabirds in Scotland. Research is needed urgently to quantify the impact and to develop implementation of appropriate conservation actions. Actions to reduce long-line bycatch of seabirds globally are already well developed but are currently not deployed across the whole long-line fleet in Scottish waters. Conservation actions also need further development for set-net bycatch.
All of these actions require a parallel programme of education (in the broadest sense) to gain better societal support for seabird conservation, and of seabird demographic monitoring. The latter is important not only to measure the success of actions, but also to gain a better understanding of the impacts of pressures on seabird populations and the relative importance of different pressures and different conservation actions.
Ultimately, no single action will trigger seabird recovery. A combination of actions will be necessary to facilitate recovery, maintain recovering populations and increase seabird resilience to increasing pressures associated with our changing environment. Implementing these actions will require sustained funding and collaborative approaches in Scotland and internationally.
References
ACAP 2019. ACAP review of seabird bycatch mitigation measures and summary advice for reducing the impact of fishing on seabirds. ACAP.
Aires, C., González-Irusta, J.M. and Watret, R. 2014. Updating fisheries sensitivity maps in British waters. Scottish Marine and Freshwater Science Report 5. No. 10. Marine Scotland Science.
Anderson, B.A. 2007. A Literature Review of the Effects of Helicopter Disturbance and Noise on Selected Wildlife Species. ABR, Inc.—Environmental Research & Services: Anchorage, Alaska
Anderson, O.R.J., Thompson, D. and Parsons, M. 2022. Seabird Bycatch Mitigation: Evidence base for possible UK application and further research. JNCC Report No. 717. JNCC, Peterborough.
Anker-Nilssen, T., Fayet, A.L. and Aarvak, T. 2023. Top-down control of a marine mesopredator: increase in native white-tailed eagles accelerates the extinction of an endangered seabird population. Journal of Applied Ecology 10.1111/1365-2664.14343
Armstrong, I.H., Coulson, J.C., Hawkey, P. and Hudson, M.J. 1978. Further mass seabird deaths from paralytic shellfish poisoning. British Birds 71: 58-68.
Ausden, M., Dixon, M., Lock, L., Miles, R., Richardson, N. and Scott, C. 2018. Sea change in the beneficial use of dredged sediment. RSPB, Sandy.
Babcock, M. and Booth, V. 2020a. Roseate tern terraces and nest boxes. Tern conservation best practice. LIFE14 NAT/UK/000394.
Babcock, M. and Booth, V. 2020b. Tern conservation best practice: using canes to deter avian predators. RSPB.
Bærum, K.M., Anker-Nilssen, T., Christensen-Dalsgaard, S., Fangel, K., Williams, T. and Volstad, J.H. 2019. Spatial and temporal variations in seabird bycatch: incidental bycatch in the Norwegian coastal gillnet-fishery. PLoS ONE 14: e0212786.
Bambini, L. and Varnham, K. 2020. Technical assessment of the biosecurity measures required on the 42 UK seabird island SPAs. Biosecurity for LIFE [LIFE17/GIE/000572]. RSPB, Sandy.
Banyard, A.C., Lean, F.Z.X., Robinson, C., Howie, F., Tyler, G., Nisbet, C., Seekings, J., Meyer, S., Whittard, E., Ashpitel, H.F., Bas, M., Byrne, A.M.P., Lewis, T., James, J., Stephan, L., Lewis, N.S., Brown, I.H., Hansen, R.D.E. and Reid, S.M. 2022. Detection of highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b in great skuas: a species of conservation concern in Great Britain. Viruses 14: 212.
Barrett, R.T., Strøm, H. and Melnikov, M. 2017. On the polar edge: the status of the northern gannet (Morus bassanus) in the Barents Sea in 2015-16. Polar Research 36: 1390384.
Bell, E., Floyd, K., Boyle, D., Pearson, J., St Pierre, P., Lock, L., Buckley, P., Mason, S., McCarthy, R., Garratt, W., Sugar, K. and Pearce, J. 2019. The Isles of Scilly seabird restoration project: the eradication of brown rats (Rattus norvegicus) from the inhabited islands of St Agnes and Gugh, Isles of Scilly. IUCN Species Survival Commission Occasional Papers 62: 88–94.
Bolton, M. 1996. Energy expenditure, body-weight and foraging performance of storm petrels Hydrobates pelagicus breeding in artificial nesting chambers. Ibis 138: 405-409.
Bolton, M., Medeiros, R., Hothersall, B. and Campos, A. 2004. The use of artificial breeding chambers as a conservation measure for cavity-nesting procellariform seabirds: a case study of the Madeiran storm petrel (Oceanodroma castro). Biological Conservation 116: 73-80.
Booker, H., Price, D., Slader, R., Frayling, T., Williams, T. and Bolton, M. 2019. Seabird recovery on Lundy: population change in Manx shearwaters and other seabirds in response to eradication of rats. British Birds 112: 217-230.
Boothby, C., Redfern, C. and Schroeder, J. 2019. An evaluation of canes as a management technique to reduce predation by gulls of ground-nesting seabirds. Ibis 161: 453-458.
Boyle, J. and Copley, L. 2022. Co-Design and Conservation: A Case-Study from RSPB Biosecurity for LIFE in Coastal and Island Primary Schools and Youth Groups Across Scotland, Concept, 13(2), pp. 1-13
Bradbury, G., Shackshaft, M., Scott-Hayward, L., Rexstad, E., Miller, D. and Edwards, D. 2017. Risk assessment of seabird bycatch in UK waters. Report to Defra. Project No MB0126.
Braune, B., Gaston, A.J., Hobson, K.A., Gilchrist, H.G. and Mallory, M.L. 2015. Changes in trophic position affect rates of contaminant decline at two seabird colonies in the Canadian Arctic. Ecotoxicology and Environmental Safety 115: 7-13.
Brisson-Curadeau, É., Bird, D., Burke, C., Fifield, D.A., Pace, P., Sherley, R.B. and Elliott, K.H., 2017. Seabird species vary in behavioural response to drone census. Scientific reports 7: pp.1-9
British Birds 2019. Storm petrel nest boxes. British Birds e-newsletter 53.
Brooke, M.de.L., Hilton, G.M., Martins, T.L.F., 2007. Prioritizing the world's islands for vertebrate-eradication programmes. Animal Conservation 10, 380-390.
BTO 2023. Black guillemot nest boxes. BTO
Buckingham, L., Daunt, F., Bogdanova, M.I., Furness, R.W., Bennett, S., Duckworth, J., Dunn, R.E., Wanless, S., Harris, M.P., Jardine, D.C., Newell, M.A., Ward, R.M., Weston, E.D. and Green, J.A. 2023. Energetic synchrony throughout the non-breeding season in common guillemots from four colonies. Journal of Avian Biology 2023: e03018.
Burthe, S., Wanless, S., Newell, M.A., Butler, A. and Daunt, F. 2014. Assessing the vulnerability of the marine bird community in the western North Sea to climate change and other anthropogenic impacts. Marine Ecology Progress Series 507: 277-295.
Camphuysen, K. 2008. Entanglement of seabirds in marine litter and fishing gear, 1970-2007. Sula 21: 88-92.
Camphuysen, C.J., Gear, S.C. and Furness, R.W. 2022. Avian influenza leads to mass mortality of adult great skuas in Foula in summer 2022. Scottish Birds 42: 312-323.
Carroll, M.J., Butler, A., Owen, E., Ewing, S.R., Cole, T. et al. 2015. Effects of sea temperature and stratification changes on seabird breeding success. Climate Research 66: 75-89.
Christensen-Dalsgaard, S., Langset, M. and Anker-Nilssen, T. 2019. Offshore oil rigs – a breeding refuge for Norwegian black-legged kittiwakes Rissa tridactyla? Seabird 32: 20-32.
Christensen-Dalsgaard, S., Ytrehus, B., Langset, M., Wiig, J.R. and Bærum, K.M. 2022. Seabird beachcast events associated with bycatch in the Norwegian purse seine fishery. Marine Environmental Research 177: 105625.
Church, G.E., Furness, R.W., Tyler, G., Gilbert, L. and Votier, S.C. 2018. Change in the North Sea ecosystem from the 1970s to the 2010s: great skua diets reflect changing forage fish, seabirds, and fisheries. ICES Journal of Marine Science 76: 925-937.
Clark, B.L., Vigfusdottir, F., Jessopp, M.J., Burgos, J.M., Bodey, T.W. and Votier, S.C. 2020. Gannets are not attracted to fishing vessels in Iceland – potential influence of a discard ban and food availability. ICES Journal of Marine Science 77: 692-700.
Cleasby, I.R., Wilson, L.J., Crawford, R., Owen, E., Rouxel, Y. and Bolton, M. 2022. Assessing bycatch risk from gillnet fisheries for three species of diving seabird in the UK. Marine Ecology Progress Series 684: 157-179.
Cleasby, I.R., Wilson, L.J. and Davies, J.G. 2021. Predicting seabird distributions in response to climate change using habitat modelling. Report to AFBI and Marine Scotland.
Coccon, F., Borella, S., Simeoni, N. and Malavasi, S. 2018. Floating rafts as breeding habitats for the Common tern, Sterna hirundo. Colonization patterns, abundance and reproductive success in Venice Lagoon. Rivista Italiana di Ornitologia 88: 23–32.
Cook, M.I. and Hamer, K.C. 1997. Effects of supplementary feeding on provisioning and growth rates of nestling puffins Fratercula arctica: evidence for regulation of growth. Journal of Avian Biology 28: 56-62.
Cooper, J. 2013. Predator-proof fences are helping to protect procellariiform seabirds, including ACAP-listed albatrosses and petrels
Coulson, J.C. 2011. The Kittiwake. T & AD Poyser, London.
Coulson, J.C. 2015. Re-evaluation of the role of landfills and culling in the historic changes in the herring gull (Larus argentatus) population in Great Britain. Waterbirds 38: 339-354.
Coulson, J.C., Potts, G.R., Deans, I.R. and Frazer, S.M. 1968. Dinoflagellate crop in the North Sea: mortality of shags and other sea-birds caused by paralytic shellfish poison. Nature 220: 21-27.
Coulson, J.C. and Strowger, J. 1999. The annual mortality rate of black-legged kittiwakes in NE England from 1954 to 1998 and a recent exceptionally high mortality. Waterbirds 22: 3-13.
Courchamp, F., Chapuis, J-L. and Pascal, M. 2003. Mammal invaders on islands: impact, control and control impact. Biological Reviews 78: 347-383.
Cury, P.M., Boyd, I.L., Bonhommeau, S., Anker-Nilssen, T., Crawford, R.J.M., Furness, R.W., Mills, J.A., Murphy, E.J., Österblom, H., Paleczny, M., Piatt, J.F., Roux, J-P., Shannon, L. and Sydeman, W.J. 2011. Global seabird response to forage fish depletion – one-third for the birds. Science 334: 1703-1706.
Dahl, H.K., Barrett, R.T. and Ims, R.A. 2005. Effects of supplementary feeding on provisioning and growth rates of Atlantic puffin Fratercula arctica chicks in north Norway. Atlantic Seabirds 7: 133-143.
Darby, J.H., de Grissac, S., Arneill, G.E., Pirotta, E., Waggitt, J.J., Borger, L., Shepard, E., Cabot, D., Owen, E., Bolton, M., Edwards, E.W.J., Thompson, P.M., Quinn, J.L. and Jessopp, M. 2021. Foraging distribution of breeding northern fulmars is predicted by commercial fisheries. Marine Ecology Progress Series 679: 181-194.
Dasnon, A., Delord, K., Chaigne, A. and Barbraud, C. 2022. Fisheries bycatch mitigation measures as an efficient tool for the conservation of seabird populations. Journal of Applied Ecology 59: 1674-1685.
Davies, J.G., Humphreys, E.M. and Pearce-Higgins, J.W. 2021. Projected future vulnerability of seabirds within the INTERREG VA area to climate change. Report to AFBI and Marine Scotland.
Davis, S.E., Nager, R.G. and Furness, R.W. 2005. Food availability affects adult survival as well as breeding success of parasitic jaegers. Ecology 86: 1047-1056.
Deakin, Z., Cook, A., Daunt, F., McCluskie, A., Morley, N., Witcutt, E., Wright, L. and Bolton, M. 2022. A review to inform the assessment of the risk of collision and displacement in petrels and shearwaters from offshore wind developments in Scotland. Report to Scottish Government.
Degraer, S., Brabant, R., Rumes, B. and Vigin, L. 2023. Environmental impacts of offshore wind farms in the Belgian part of the North Sea: Progressive insights in changing species distribution patterns informing marine management. Memoirs on the Marine Environment, Royal Belgian Institute of Natural Sciences, Brussels.
Dierschke, V., Furness, R.W. and Garthe, S. 2016. Seabirds and offshore wind farms in European waters: avoidance and attraction. Biological Conservation 202: 59-68.
Dierschke, V., Christensen-Dalsgaard, S. and Koschinski, S. 2022. Pilot assessment of marine bird bycatch. OSPAR 2023 Quality Status Report for the North-East Atlantic. OSPAR Commission, London.
Dierschke, V. and Mercker, M. 2022. Pilot assessment of marine bird habitat quality. OSPAR 2023 Quality Status Report for the North-East Atlantic. OSPAR Commission, London.
DIISE 2016. The database of island invasive species eradications, developed by island conservation, coastal conservation action laboratory UCSC, IUCN SSC invasive species specialist group. University of Auckland
Drent, R.H. and Daan, S., 1980. The prudent parent: energetic adjustments in avian breeding 1. Ardea, 55(1–2), pp.225-252.
Dunn, E.K. and Steel, C., 2001. The impact of long-line fishing on seabirds in the north-east Atlantic: recommendations for reducing mortality. RSPB/JNCC, Sandy.
Esmonde, N.P.G., Hanna, R.E.B., Patel, J.G., Smyth, V.J., Caplat, P., Smyth, W., Jaggers, P., Padget, O., Guilford, T., Perrins, C. and Reid, N. 2022. Case report of puffinosis in a Manx shearwater (Puffinus puffinus) suggesting environmental aetiology. Animals 12: 3457.
Equinor 2022. Sheringham Shoal and Dudgeon offshore wind farm extension projects DCO application Appendix 2: Sandwich tern compensation document. PINS 5.5.2 DOC C282-RH-2-GA-00187.
Falchieri, M., Reid, S.M., Ross, C.S., James. J., Byrne, A.M.P., Zamfir, M., Brown, I.H., Banyard, A.C., Tyler, G., Philip, E. and Miles, W. 2022. Shift in HPAI infection dynamics causes significant losses in seabird populations across Great Britain. Vet Record 8/15 October 2022: 294-296.
Fernandez, T.F., Hollywood, C., Huxham, M. and Kinross, J. 2005. Sprat survey of the Firth of Forth, December 2004. Commissioned by Fife Council. Napier University, Edinburgh.
Field, R., Crawford, R., Enever, R., Linkowski, T., Martin, G., Morkunas, J., Morkune, R., Rouxel, Y. and Oppel, S. 2019. High contrast panels and lights do not reduce bird bycatch in Baltic Sea gillnet fisheries. Global Ecology and Conservation 18: e00602.
Fliessbach, K.L., Borkenhagen, K., Guse, N., Markones, N., Schwemmer, P. and Garthe, S. 2019. A ship traffic disturbance vulnerability index for northwestern European seabirds as a tool for marine spatial planning. Frontiers in Marine Science 6: 1-15.
Forrester, R.W., Andrews, I.J., McInerny, C.J., Murray, R.D., McGowan, R.Y., Zonfrillo, B., Betts, M.W., Jardine, D.C. and Grundy, D.S. 2007. The Birds of Scotland. The Scottish Ornithologists’ Club, Aberlady.
van Franeker, J.A., Kuhn, S., Anker-Nilssen, T., Edwards, E.W.J., Gallien, F., Guse, N., Kakkonen, J.E., Mallory, M.L., Miles, W., Olsen, K.O., Pedersen, J., Provencher, J., Roos, M., Stienen, E., Turner, D.M. and van Loon, W.M.G.M. 2021. New tools to evaluate plastic ingestion by northern fulmars applied to North Sea monitoring data 2002-2018. Marine Pollution Bulletin 166: 112246.
Furness, R.W. 1987. The Skuas. T & AD Poyser, Calton.
Furness, R.W., Ensor, K. and Hudson, A.V. 1992. The use of fishery waste by gull populations around the British Isles. Ardea 80: 105-113.
Furness, R.W., Wade, H.M., Robbins, A.M.C. and Masden, E.A. 2012. Assessing the sensitivity of seabird populations to adverse effects from tidal stream turbines and wave energy devices. ICES Journal of Marine Science 69: 1466-1479.
Furness, R.W., MacArthur, D., Trinder, M. and MacArthur, K. 2013. Evidence review to support the identification of potential conservation measures for selected species of seabirds. Report to Defra.
Furness, R.W., Wade, H.M. and Masden, E.A. 2013. Assessing vulnerability of marine bird populations to offshore wind farms. Journal of Environmental Management 119: 56-66.
Furness, R.W. 2016. Impacts and effects of ocean warming on seabirds. In: D. Laffoley and J.M. Baxter, ed. 2016. Explaining ocean warming: causes, scale, effects and consequences. IUCN, Gland.PP, pp 271-288.
Garthe, S., Camphuysen, C.J. and Furness, R.W. 1996. Amounts of discards by commercial fisheries and their significance as food for seabirds in the North Sea. Marine Ecology Progress Series 136: 1-11.
Gear, S.C. 2019. Seabird monitoring Foula 2019. Report to SOTEAG.
Gear, S.C. 2022. Seabird monitoring Foula 2022. Report to SOTEAG.
Gill, V.A., Hatch, S.A. and Lanctot, R.B. 2002. Sensitivity of breeding parameters to food supply in black-legged kittiwakes Rissa tridactyla. Ibis 144: 268-283.
Gilman, E., Beverly, S., Musyl, M. and Chaloupka, M. 2020. Commercial viability of locating pelagic longline branchline weights at the hook to reduce seabird bycatch. Endangered Species Research 43: 223-233.
Gilman, E., Musyl, M., Wild, M., Rong, H. and Chaloupka, M. 2022. Investigating weighted fishing hooks for seabird bycatch mitigation. Scientific Reports 12: 2833.
Grant, M.L., O’Hanlon, N.J., Lavers, J.L., Masden, E.A., James, N.A. and Bond, A.L. 2021. A standardised method for estimating the level of visible debris in bird nests. Marine Pollution Bulletin 172: 112889.
Grecian, W.J., Inger, R., Atrill, M.J., Bearhop, S., Godley, B.J., Witt, M.J. and Votier, S.C. 2010. Potential impacts of wave-powered marine renewable energy installations on marine birds. Ibis 152: 683-697.
Greenwood, J.G. 2015. Breeding sites and breeding success in 2014 of black guillemots at Bangor Marina, Co. Down. Northern Ireland Seabird Report 2014.
Guilford, T., Padget, O., Bond, S. and Syposz, M. 2019. Light pollution causes object collisions during local nocturnal manoeuvring flight by adult Manx shearwaters Puffinus puffinus. Seabird 31: 48-55.
Hammer, S., Nager, R.G., Johnson, P.C.D., Furness, R.W. and Provencher, J.F. 2016. Plastic debris in great skua (Stercorarius skua) pellets corresponds to seabird prey species. Marine Pollution Bulletin 103: 206-210.
Harris, M.P. 1965. Puffinosis among Manx shearwaters on Skokholm. British Birds 58: 426-433.
Harris, M.P. 1978. Supplementary feeding of young puffins Fratercula arctica. Journal of Animal Ecology 47: 15-23.
Harris, M.P. and Wanless, S. 2011. The Puffin. T & AD Poyser, London.
Heessen, H.J.L., Daan, N. and Ellis, J.R. eds., 2015. Fish atlas of the Celtic Sea, North Sea, and Baltic Sea based on international research-vessel surveys. Wageningen Academic Publishers.
Horswill, C. and Robinson, R.A. 2015. Review of seabird demographic rates and density dependence. JNCC Report No. 552. JNCC, Peterborough.
Hudson, A.V. and Furness, R.W. 1988. Utilization of discarded fish by scavenging seabirds behind whitefish trawlers in Shetland.Journal of Zoology, London 215: 151-166.
Hughes, R.D., O’Hanlon, N. and Smith, J. 2021. Colonisation of St John’s Pool, Caithness by terns and gulls. Scottish Birds 41: 205-212.
Huntley, B., Green, R., Collingham, Y.C. and Willis, S.G., 2007. A climatic atlas of European breeding birds (p. 521). Barcelona: Lynx Edicions.
ICES no date. Sprat. ICES FishMap species fact sheet.
ICES. 2020. Sandeel in Division 3.a and Subarea 4. Section 9 in Herring Assessment Working Group for the Area South of 62°N (HAWG).ICES
Jones, H.P., Tershy, B.R., Zavaleta, E.S., Croll, D.A., Keitt, B.S., Finkelstein, M.E. and Howald, G.R. 2008. Severity of the effects of invasive rats on seabirds: a global review. Conservation Biology 22: 16-26.
King, M.D., Elliott, J.E. and Williams, T.D. 2021. Effects of petroleum exposure on birds: a review. Science of the Total Environment 775: 142834.
Kuiken, T. and Cromie, R. 2022. Protect wildlife from livestock diseases. Science 378: 5.
Jennings, G., McGlashan, D.J. and Furness, R.W., 2012. Responses to changes in sprat abundance of common tern breeding numbers at 12 colonies in the Firth of Forth, east Scotland. ICES journal of marine science, 69(4), pp.572-577.
Johnston, D.T., Furness, R.W., Robbins, A.M.C., Tyler, G.A. and Masden, E.A. 2019. Camera traps reveal predators of breeding black guillemots Cepphus grylle. Seabird 32: 72-83.
Johnston, D.T., Furness, R.W., Robbins, A.M.C., Tyler, G.A., McIlvenny, J. and Masden, E.A. 2021a. Tidal stream use by black guillemots Cepphus grylle in relation to a marine renewable energy development. Marine Ecology Progress Series 669: 201-212.
Johnston, D.T., Humphreys, E.M., Davies, J.G. and Pearce-Higgins, J.W. 2021b. Review of climate change mechanisms affecting seabirds within the INTERREG VA area. Report to AFBI and Marine Scotland.
Jones, H.P., Holmes, N.D., Butchart, S.H.M., Tershy, B.R., Kappes, P.J., Corkery, I., Aguirre-Muñoz, A., Armstrong, D.P., Bonnaud, E., Burbidge, A.A. and Campbell, K., 2016. Invasive mammal eradication on islands results in substantial conservation gains. Proceedings of the National Academy of Sciences of the United States of America 113: 4033-4038.
Kingston, A., Northridge, S., Paxton, C. G. m., & Forti Buratti, J. P. 2023. Improving understanding of seabird bycatch in Scottish longline fisheries and exploring potential solutions. Marine Scotland.
Kjesbu, O.S., Sundby, S., Sandø, A.B., Alix, M., Hjøllo, S.S., Tiedemann, M., Skern‐Mauritzen, M., Junge, C., Fossheim, M., Thorsen Broms, C. and Søvik, G., 2022. Highly mixed impacts of near-future climate change on stock productivity proxies in the North East Atlantic. Fish and Fisheries 23: 601-615.
Kwak, M.L., Heath, A.C.G. and Palma, R.L. 2019. Saving the Manx shearwater flea Ceratophyllus (Emmareus) fionnus (Insecta: Siphonaptera): the road to developing a recovery plan for a threatened ectoparasite. Acta Parasitologica 64: 903-910.
Lambert, M., Carlisle, S., Cain, I., Douse, A. and Watt, L. 2021. Unexpected involvement of a second rodent species makes impacts of introduced rats more difficult to detect. Scientific Reports 11: 19805.
Lane, J.V., Jeavons, R., Deakin, Z., Sherley, R.B., Pollock, C.J., Wanless, R.J. and Hamer, K.C., 2020. Vulnerability of northern gannets to offshore wind farms; seasonal and sex-specific collision risk and demographic consequences. Marine Environmental Research, 162, p.105196.
Langley, L.P., Bearhop, S., Burton, N.H.K., Banks, A.N., Frayling, T., Thaxter, C.B., Clewley, G.D., Scragg, E. and Votier, S.C. 2021. GPS tracking reveals landfill closures induce higher foraging effort and habitat switching in gulls. Movement Ecology 9: 56.
Langton, R., Boulcott, P. and Wright, P.J. 2021. A verified distribution model for the lesser sandeel Ammodytes marinus. Marine Ecology Progress Series 667: 145-159.
Lavers, J.L., Hutton, I. and Bond, A.L. 2019. Clinical pathology of plastic ingestion in marine birds and relationships with blood chemistry. Environmental Science and Technology 53: 9224-9231.
Lawrence, J.M. and Fernandes, P.G. 2021. A switch in species dominance of a recovering pelagic ecosystem. Current Biology 31: 1-7.
Leat, E.H.K., Bourgeon, S., Hanssen, S.A., Petersen, A., Strøm, H., Bjørn, T.H., Gabrielsen, G.W., Bustnes, J.O., Furness, R.W., Haarr, A. and Borgå, K. 2019. The effect of long-range transport, trophic position and diet specialization on legacy contaminant occurrence in great skuas, Stercorarius skua, breeding across the Northeast Atlantic. Environmental Pollution 244: 55-65.
de Leon, A. and Minguez, E. 2003. Occupancy rates and nesting success of European storm-petrels breeding inside artificial nest-boxes. Scientia Marina 67: 109-112.
de Leon, A., Minguez, E., Harvey, P., Meek, E., Crane, J.E. and Furness, R.W. 2006. Factors affecting breeding distribution of storm-petrels Hydrobates pelagicus in Orkney and Shetland. Bird Study 53: 64-72.
Leonard, K., Greenwood, J.G. and Wolsey, S. 2015. Nest boxes for black guillemots. Northern Ireland Seabird Report 2014.
Libois, E., Gimenez, O., Oro, D., Minguez, E., Pradel, R. and Sanz-Aguilar, A. 2012. Nest boxes: a successful management tool for the conservation of an endangered seabird. Biological Conservation 155: 39-43.
Lock, L., Donato, B., Jones, R. and MacLeod-Nolan, C. 2022. England’s breeding seabirds: a review of the status of their breeding sites and suggested measures for their recovery. Natural England and RSPB.
Løkkeborg, S. 2003. Review and evaluation of three mitigation measures – bird-scaring line, underwater setting and line shooter – to reduce seabird bycatch in the North Atlantic longline fishery. Fisheries Research 60: 11-16.
Løkkeborg, S. 2011. Best practices to mitigate seabird bycatch in longline, trawl and gillnet fisheries – efficiency and practical applicability. Marine Ecology Progress Series 435: 285-303.
Løkkeborg, S. and Robertson, G. 2002. Seabird and longline interactions: effects of a bird-scaring streamer line and line shooter on the incidental capture of northern fulmars Fulmarus glacialis. Biological Conservation 106: 359-364.
MacArthur Green and Royal HaskoningDHV, 2022. Norfolk Projects Offshore Wind Farms Lesser black-backed gull Implementation and Monitoring Plan. Ref PB5640.009.0005.
MacDonald, J.W., McMartin, D.A., Walker, K.G., Carins, M. and Dennis, R.H. 1967. Puffinosis in fulmars in Orkney and Shetland. British Birds 60: 356-360.
MacDonald, J.W. and Standring, K.T. 1978. Outbreak of botulism in gulls on Firth of Forth, Scotland. Biological Conservation 14: 149-155.
Manikowska-Slepowronska, B., Slepowronski, K. and Jakubas, D. 2022. The use of artificial floating nest platforms as conservation measure for the common tern Sterna hirundo: a case study in the RAMSAR site Druzno Lake in Northern Poland. European Zoological Journal 89: 229–240.
Manning, W.D., Scott, C.R. and Leegwater, E. eds, 2021. Restoring estuarine and coastal habitats with dredged sediment: a handbook. Environment Agency, Bristol.
Martin, G.R. and Banks, A.N. 2023. Marine birds: vision-based wind turbine collision mitigation. Global Ecology and Conservation 42: e02386.
Mason, C.F., Ekins, G. and Ratford, J.R. 1997. PCB congeners, DDE, dieldrin and mercury in eggs from an expanding colony of cormorants (Phalacrocorax carbo). Chemosphere 34: 1845-1849.
May, R., Nygard, T., Falkdalen, U., Astrom, J., Hamre, O. and Stokke, B.G. 2020. Paint it black: efficacy of increased wind turbine rotor blade visibility to reduce avian fatalities. Ecology and Evolution 10: 8927-8935.
McGregor, R. 2022a. Assessment of compensatory measures for impacts of offshore windfarms on seabirds. Natural England Commissioned Report NECR431.
Melvin, E.F., Dietrich, K.S., Suryan, R.M. and Fitzgerald, S.M. 2019. Lessons from seabird conservation in Alaskan longline fisheries. Conservation Biology 33: 842-852.
Middleton, J.R. 1981. Breeding of the black guillemot on Inchmarnock. Western Naturalist 10: 26-27.
Milborrow, J. and Murphy, J. 2008. Improvement of nest boxes and trial of new techniques for storm petrel monitoring. CCW Contract Science Report No. 832.
Miles, W., Money, S., Luxmoore, R. and Furness, R.W. 2010. Effects of artificial lights and moonlight on petrels at St Kilda. Bird Study 57: 244-251.
Miles, J., Parsons, M. and O’Brien, S. 2020. Preliminary assessment of seabird population response to potential bycatch mitigation in the UK-registered fishing fleet. JNCC Report, Defra Project code ME6024.
Mitchell, I., Thomas, S., Bambini, L., Varnham, K., Phillips, R., Singleton, G., Douse, A., Foster, S., Kershaw, M., McCulloch, N., Murphy, M. and Hawkridge, J. 2018. Invasive mammal presence on island seabird colonies. UK Marine Online Assessment Tool.
Mitchell, P.I., Newton, S.F., Ratcliffe, N. and Dunn, T.E. 2004. Seabird Populations of Britain and Ireland. T & AD Poyser, London.
Natural England 2022. Natural England interim advice on updated collision risk modelling parameters (July 2022).
NatureScot 2017. A guide to best practice for watching marine wildlife. Report.
NatureScot 2020. Pole-mounted top nets and birds at finfish farms. NatureScot Interim Technical Briefing Note.
Northridge, S., Kingston, A. and Coram, A. 2020. Preliminary estimates of seabird bycatch by UK vessels in UK and adjacent waters. Report to JNCC, Defra.
O’Hanlon, N.J., James, N.A., Masden, E.A. and Bond, A.L. 2017. Seabirds and marine plastic debris in the north eastern Atlantic: a synthesis and recommendations for monitoring and research. Environmental Pollution 231: 1291-1301.
O’Hanlon, N.J., Bond, A.L., Lavers, J.L., Masden, E.A. and James, N.A. 2019. Monitoring nest incorporation of anthropogenic debris by northern gannets across their range. Environmental Pollution 255: 113152.
O’Keefe, C.E., Cadrin, S.X., Glemarec, G. and Rouxel, Y. 2023. Efficacy of Time-Area Fishing Restrictions and Gear-Switching as Solutions for Reducing Seabird Bycatch in Gillnet Fisheries. Reviews in Fisheries Science & Aquaculture 31: 29-46.
Olin, A.B., Dück, L., Berglund, P-A., Karlsson, E., Bohm, M., Olsson, O. and Hentati-Sundberg, J. 2023. Breeding failures and reduced nest attendance in response to heat stress in a high-latitude seabird. Marine Ecology Progress Series
Ørsted 2022. Hornsea Project Four Bycatch Reduction Technology Selection Phase Summary. Deadline: 5, Date: 20 June 2022. Document Reference: G5.13. Revision: 01.
Ortiz, N.E. and Smith, G.R. 1994. Landfill sites, botulism and gulls. Epidemiology and Infection 112: 385-391.
Pardo, D., Forcada, J., Wood, A.G., Tuck, G.N., Ireland, L., Pradel, R., Croxall, J.P. and Phillips, R.A. 2017. Additive effects of climate and fisheries drive ongoing declines in multiple albatross species. Proceedings of the National Academy of Sciences of the United States of America 114: E10829-E10837.
Pearce-Higgins, J.W., Davies, J.G. and Humphreys, E.M. 2021. Species and habitat climate change adaptation options for seabirds within the INTERREG VA area. Report to AFBI and Marine Scotland.
Pearson, J. 2022. Summary – Biosecurity legacy survey for Scottish island stakeholders. Unpublished.
Pereira, M.G., Walker, L.A., Best, J. and Shore, R.F. 2009. Long-term trends in mercury and PCB congener concentrations in gannet (Morus bassanus) eggs in Britain. Environmental Pollution 157: 155-163.
Peschko, V., Mercker, M. and Garthe, S. 2020. Telemetry reveals strong effects of offshore wind farms on behaviour and habitat use of common guillemots (Uria aalge) during the breeding season. Marine Biology 167: 118.
Phillips, R.A., Furness, R.W. and Stewart, F.M. 1998. The influence of territory density on the vulnerability of Arctic skuas Stercorarius parasiticus to predation. Biological Conservation 86: 21-31.
Power, A., White, P., McHugh, B., Berrow, S., Schlingermann, M., Tannian, M., Newton, S., McGovern, E., Murphy, S., Crowley, D., O’Hea, L., Boyle, B. and O’Connor, I. 2021. Persistent pollutants in northern gannet Morus bassanus eggs in Ireland: levels and colony differences. Environmental Pollution 268: 115723.
Quayle, H. 2015. Filey Bay: Safe Seas for Seabirds. RSPB report.
Ratcliffe, N., Mitchell, I., Varnham, K., Verboven, N. and Higson, P. 2009. How to prioritize rat management for the benefit of petrels: a case study of the UK, Channel Islands and Isle of Man. Ibis 151: 699-708.
Rijks, J.M., Leopold, M.F., Kühn, S., in ‘t Veld, R., Schenk, F., Brenninkmeijer, A., Lilipaly, S.J., Ballmann, M.Z., Kelder, L., de Jong, J.W., Courtens, W., Slaterus, R., Kleyheeg, E., Vreman, S., Kik, M.J.L., Gröne, A., Fouchier, R.A.M., Englesma, M., de Jong, M.C.M., Kuiken, T. and Beerens, N. 2022. Mass mortality caused by Highly Pathogenic Influenza A(H5N1) virus in Sandwich terns, the Netherlands, 2022. Emerging Infectious Diseases 28: 2538-2542.
Rivers-Auty, J., Bond, A.L., Grant, M.L. and Lavers, J.L. 2023. The one-two punch of plastic exposure: macro- and micro-plastics induce multi-organ damage in seabirds. Journal of Hazardous Materials 442: 130117.
Rouxel, Y., Crawford, R., Cleasby, I.R., Kibel, P., Owen, E., Volke, V., Schnell, A.K. and Oppel, S. 2021. Buoys with looming eyes deter seaducks and could potentially reduce seabird bycatch in gill nets. Royal Society Open Science 8: 210225.
Russell, D.J.F., Wanless, S., Collingham, Y.C., Anderson, B.J., Beale, C., Reid, J.B., Huntley, B. and Hamer, K.C. 2014. Beyond climate envelopes: bio-climate modelling accords with observed 25-year changes in seabird populations of the British Isles. Diversity and Distributions 21: 211-222.
Russell, D., Wanless, S., Collingham, Y., Huntley, B. and Hamer, K. 2015. Predicting future European breeding distributions of British seabird species under climate change and unlimited/no dispersal scenarios. Diversity 7: 342-359.
Sanz-Aguilar, A., Martinez-Abrain, A., Tavecchia, G., Minguez, E. and Oro, D. 2009. Evidence-based culling of a facultative predator: efficacy and efficiency components. Biological Conservation 142: 424-431.
Schwartz, T., Besnard, A., Pin, C., Scher, O., Blanchon, T., Béchet, A. and Sadoul, N. 2022. Efficacy of created and restored nesting sites for the conservation of colonial Laridae in the South of France. Conservation Biology 2022: e14005.
Scopel, L.C. and Diamond, A.W. 2017. The case for lethal control of gulls on seabird colonies. Journal of Wildlife Management 81: 572-580.
Scottish Natural Heritage (SNH). 2015. The use of helicopters and aircraft in relation to disturbance risks to Schedule 1 & 1A raptors and wider Schedule 1species
Schwemmer, P., Mendel, B., Sonntag, N., Dierschke, V. and Garthe, S. 2011. Effects of ship traffic on seabirds in offshore waters: implications for marine conservation and spatial planning. Ecological Applications 21: 1851-1860.
Searle, K.R., Waggitt, J., Evans, P., Bogdanova, M., Daunt, F. and Butler, A. 2022a. Study to examine the impact of climate change on seabird species off the east coast of Scotland and potential implications for environmental assessments. Marine Scotland Science Report.
Searle, K.R., Butler, A., Waggitt, J.J., Evans, P.G.H., Quinn, L.R., Bogdanova, M.I., Evans, T.J., Braithwaite, J.E. and Daunt, F. 2022b. Potential climate-driven changes to seabird demography: implications for assessments of marine renewable energy development. Marine Ecology Progress Series 690: 185-200.
Sherley, R.B., Ladd-Jones, H., Garthe, S., Stevenson, O. and Votier, S.C. 2020. Scavenger communities and fishery waste: North Sea discards support 3 million seabirds, 2 million fewer than in 1990. Fish and Fisheries 21: 12422.
Short, D. 2020. Breeding of four species of tern and black-headed gull at Forvie National Nature Reserve, 2020. Report to NatureScot.
Skeate, E. 2022. Berwick Bank Wind Farm Derogation Case Colony Compensatory Measures Evidence Report. SSE Renewables.
Spaans, B., Leopold, M,. and Plomp, M. 2018. Using a drone to determine the number of breeding pairs and breeding success of Sandwich terns Sterna sandvicensis. Limosa 91:30-37
Spatz, D.R., Jones, H.P., Bonnaud, E., Kappes, P., Holmes, N.D. and Guzmán, Y.B., 2023. Invasive species threats to seabirds. In Conservation of marine birds (pp. 97-130). Academic Press.
Stanbury, A., Thomas, S., Aegerter, J., Brown, A., Bullock, D., Eaton, M., Lock, L., Luxmoore, R., Roy, S., Whitaker, S. and Oppel, S. 2017. Prioritising islands in the United Kingdom and crown dependencies for the eradication of invasive alien vertebrates and rodent biosecurity. European Journal of Wildlife Research 63: 31.
Staszewski, V., McCoy, K.D. and Boulinier, T. 2008. Variable exposure and immunological response to Lyme disease Borrelia among North Atlantic seabird species. Proceedings of the Royal Society B Biological Sciences 275: 2101-2109.
Steel, D. and Outram, B. 2020. Terns – restoring biodiversity to the Isle of May’s breeding seabirds. Scottish Birds 40: 206-211.
Stienen, E.W.M., Courtens, W., van der Walle, M., Vanermen, N. and Verstraete, H. 2017. Long-term monitoring study of beached birds shows that chronic oil pollution in the southern North Sea has almost halted. Marine Pollution Bulletin 115: 194-200.
Stolte, A., Dederer, G., Lamp, J., Fenn, C., Lee, M., Frank, W., Howe, C., Günther, M., Vesper, H. and Werner, S. 2022. The quest for ghost gear in the German Baltic Sea: a team effort between WWF, divers, fisherfolk, and public authorities. Frontiers in Marine Science 9: 981840.
Syposz, M., Gonçalves, F., Carty, M., Hoppitt, W. and Manco, F. 2018. Factors influencing Manx shearwater grounding on the west coast of Scotland. Ibis 160: 846-854.
Tjørnløv, R.S., Skov, H., Armitage, M., Barker, M., Jørgensen, J.B., Mortensen, L.O., Thomas, K. and Uhrenholdt, T. 2022. Resolving key uncertainties of seabird flight and avoidance behaviours at offshore wind farms. Final Report for the study period 2020-2021. AOWFL, Aberdeen.
Trinder, M. 2023. Beatrice Offshore Wind Farm Year 2 Post-construction Ornithological Monitoring Report. MacArthur Green, Glasgow. bowl_2021_post_construction_ornithology_monitoring_report_25_07_2023.pdf (marine.gov.scot)
Votier, S.C., Furness, R.W., Bearhop, S., Crane, J.E., Caldow, R.W.G., Catry, P., Ensor, K., Hamer, K.C., Hudson, A.V., Kalmbach, E., Klomp, N.I., Pfeiffer, S., Phillips, R.A., Prieto, I. and Thompson, D.R. 2004. Changes in fisheries discard rates and seabird communities. Nature 427: 727-730.
Votier, S.C., Birkhead, T.R., Oro, D., Trinder, M., Grantham, M.J., Clark, J.A., McCleery, R.H. and Hatchwell, B.J. 2008. Recruitment and survival of immature seabirds in relation to oil spills and climate variability. Journal of Animal Ecology 77: 974-983.
Wang, L.M., Nabi, G., Yin, L.Y., Wang, Y.Q., Li, S.X., Hao, Z. and Li, D.M. 2021. Birds and plastic pollution: recent advances. Avian Research 12: 59.
Wernham, C.V. and Bryant, D.M. 1998. An experimental study of reduced parental effort and future reproductive success in the puffin, Fratercula arctica. Journal of Animal Ecology 67: 25-40.
Wernham, C., Toms, M., Marchant, J., Clark, J., Siriwardena, G. and Baillie, S. 2002. The migration atlas: movements of the birds of Britain and Ireland. T & AD Poyser, London.
Westerberg, K., Brown, R., Eagle, G. and Votier, S.C. 2019. Intra-population variation in the diet of an avian top predator: generalist and specialist foraging in great black-backed gulls Larus marinus. Bird Study 66: 390-397.
White, G. and Hirons, G. 2019. The predator exclusion fence manual: Guidance on the use of predator exclusion fences to reduce mammalian predation on ground-nesting birds on RSPB reserves. Version 3, October 2019. RSPB.
White, J., Leclaire, S., Kriloff, M., Mulard, H., Hatch, S.A. and Danchin, E. 2010. Sustained increase in food supplies reduces broodmate aggression in black-legged kittiwakes. Animal Behaviour 79: 1095-1100.
Wilcox, C., van Sebille, E. and Hardesty, B.D. 2015. Threat of plastic pollution to seabirds is global, pervasive, and increasing. Proceedings of the National Academy of Sciences of the United States of America 112: 11899-11904.
Young, L.C., Vanderwerf, E.A., Mitchell, C., Yeun, E., Miller, C.J., Smith, D.G. and Swenson, C. 2012. The use of predator proof fencing as a management tool in the Hawaiian Islands: a case study of Ke’ena Point Natural Area Reserve. University of Hawaii Pacific Cooperative Studies Unit Technical Report 180: 1-87.
Annex 1
| Reference | Status | Overview | Source |
|---|---|---|---|
Scottish Seabird Conservation Action Plan (SSCAP)
| Draft in confidence | The draft SSCAP is led by Marine Scotland and has used an evidence approach to identifying key threats/pressures that seabirds in Scotland are vulnerable to. Draft action plans to address these key threats are in an early stage of development. Draft species accounts are also available. | Marine Directorate |
| Conservation and Management Advice documents (CMA) | Draft in confidence | Draft CMA documents are available for most seabird marine protected areas (Special Protection Areas and Nature Conservation Marine Protected Areas). The CMAs identify current feature (designated species) condition, existing management and the conservation objectives for the MPA. | NatureScot |
| Marine Protected Area Management and Monitoring (MarPAMM) | Seabirds and climate change
| There are four relevant reports that consider how climate change affects seabirds and how vulnerable different species are to predicted climatic changes. They also advise how the new understanding could inform management plans and present species and habitat climate change adaptation options for seabirds in the MarPAMM region. Species Factsheets are also available for 19 seabird species. | Marine Scotland
|
| UK Centre for Ecology and Hydrology, Bangor University and Biomathematics and Statistics Scotland (BioSS) joint project. | Published Report “Impact of climate change on seabird species and potential implications for environmental assessments” and associated Searle et al., 2022 | This study investigated the potential impacts of climate change on seabird distribution, abundance and demography off the east coast of Scotland, and examined integration of these climate models into standard population forecast models used in assessments for offshore wind developments. | Marine Scotland
|
| Scottish Offshore Wind Energy Council (SOWEC) | Published report Report to Crown Estate Scotland and SOWEC: HRA Derogation Scope B - Review of seabird strategic compensation options | This report reviewed possible options for compensation under The Birds and Habitats Directives for seabird populations, with emphasis on Scotland. There is strong evidence that seabird species are highly dependent for breeding success and for survival on healthy stocks of forage fish, especially sandeels Ammodytes marinus. Therefore, several seabird species would benefit from actions that allow stocks of forage fish (sandeels and sprats Sprattus sprattus) to recover from depletion caused by fishing on those stocks. Modelling sandeel stock dynamics indicates that reducing fishing mortality on those stocks would lead to some recovery of stock biomass. | Marine Scotland |
| UK Bycatch Mitigation Initiative | Published Report Marine wildlife bycatch mitigation initiative and in draft material | There are three publications including a recently published study of methods for seabird bycatch mitigation that have or might be applied to UK fisheries (Anderson, Thompson and Parsons, 2022). There are projects underway looking at bycatch “hotspots” and possible reasons for these elevated bycatch rates (Northridge et al., in prep) and identifying improvements to seabird bycatch monitoring (JNCC, in prep). Bycatch “toolkits” are also in preparation, which will be practical guides for the fishing industry and regulators to further understand and reduce seabird bycatch in UK fisheries. There is also work underway funded by the Scottish Government to improve understanding of seabird bycatch in longline fisheries and potential solutions. | JNCC Marine Scotland |
| Avian influenza Task Force | Draft in confidence | Draft HPAI Response Plan – including proposals for mitigation | NatureScot |
| Biosecurity for Life | Draft in confidence | Draft Island biosecurity action plans – these are available for some seabird island SPAs. Draft Scotland Action Programme | NatureScot RSPB |
| Species on the Edge project | Draft in confidence | Draft site-based actions proposed for tern species | NatureScot |
| Draft PMF marine birds | Draft in confidence | Draft assessment against the PMF selection criteria to establish a (pre-consultation) ‘long-list’ of potential marine birds to consider for PMF status. | NatureScot |
| Natural England modelling on sandeel closures | Natural England (2021). A Natural Capital Account for the Industrial Sandeel Fisheries. Final Report by effect and ABPmer for Natural England. Unpublished.
|
|---|---|
| Draft MPA and Priority Marine Features (PMFs) Fisheries actions | Phase 1 fisheries measures -->actions Draft in confidence - ‘Phase 2’ fisheries actions |
| Collaboration on Offshore Wind Strategic Compensation (COWSC) Expert groups | Terms of Reference |
| ECOWINGS | ECOWINGS |
| Draft Oslo and Paris Conventions (OSPAR) Marine Bird Recovery Plan | OSPAR Task and Finish Group set up to take forward the recovery plan and develop brief actions for consultation Spring 2024. |
| Case Report for the OSPAR List of Threatened and/or Declining Species and Habitats: Kittiwake | Published Care Report and Recommendations |
Abbreviations
Artificial nest sites (ANS)
Biomathematics and Statistics Scotland (BioSS)
British Trust for Ornithology (BTO)
Collaboration on Offshore Wind Strategic Compensation (COWSC)
Conservation and Management Advice documents (CMA)
Department of Agriculture, Environment and Rural Affairs (DAERA)
Dichlorodiphenyltrichloroethane (DDT)
Ecological quality Objective (EcoQO)
Habitats Regulations Assessment (HRA)
Highly Pathogenic Avian Influenza (HPAI)
International Council for the Exploration of the Sea (ICES)
Invasive Alien Vertebrates (IAVs)
Invasive Non-Native Species (INNS)
Joint Nature Conservation Committee (JNCC)
Light-Emitting Diode (LED)
Marine Protected Area (MPA)Marine Protected Area Management and Monitoring Project (MarPAMM)
Nature Restoration Fund (NRF)
Non-governmental organizations (NGOs)
Priority Marine Features (PMFs)
Polychlorinated biphenyls (PCBs)
Population Viability Analysis (PVA)
Scottish Marine Environmental Enhancement Fund (SMEEF)
Scottish Offshore Wind Energy Council (SOWEC)
Scottish Ornithologists’ Club (SOC)
Scottish Seabird Conservation Action Plan (SSCAP)
Scottish and Southern Electricity Networks (SSE)
Site of Special Scientific Interest (SSSI)
Spawning Stock Biomass (SSB)
Special Protection Area (SPA)
The Royal Society for the Protection of Birds (RSPB)
Total stock biomass (TSB)
UK Centre for Ecology and Hydrology (UKCEH)
Wildfowl & Wetlands Trust Limited (WWT)
World Wild Fund for Nature (WWF)





