NatureScot Research Report 1302 - Reviewing, assessing and critiquing the evidence base on the impacts of muirburn on wildfire prevention, carbon storage and biodiversity
Year of publication: 2022
Authors: Holland, J.P., Pollock, M., Buckingham, S., Glendinning, J. & McCracken, D.
Cite as: Holland, J.P., Pollock, M., Buckingham, S., Glendinning, J. & McCracken, D. 2022. Reviewing, assessing and critiquing the evidence base on the impacts of muirburn on wildfire prevention, carbon storage and biodiversity. NatureScot Research Report 1302.
Keywords
muirburn; moorland; carbon, peat, biodiversity, fire
1. Background
Muirburn involves the controlled burning of old heather and grass to promote new growth. It is a tool used traditionally in Scotland by land managers, including gamekeepers, farmers, crofters and conservation managers to improve grazing, provide food and shelter for red grouse (Lagopus lagopus scotica) and other gamebirds, reduce the risk of wildfires and for selected conservation objectives for habitats and species. However, the benefits and disadvantages of muirburn are contested.
In 2019 the Grouse Moor Management Review Group (an independent group set up by Scottish Government) produced a report (often referred to as the Werritty Report) on grouse moor management and muirburn in Scotland. This set out a number of recommendations including a proposal to introduce legislation to require a year-round muirburn licence. Other recommendations, supported by the Scottish Government in their response, included ‘…to ban muirburn on peatland (except in very limited cases as part of an approved habitat restoration programme) and to ‘…undertake a review of the current definition of peatland, taking expert advice on whether it should be revised and a stricter definition imposed’. It was made clear that the licensing regime should apply to all muirburn and not just that carried out on grouse moors. This means that licensing will apply to all land users and land managers practicing muirburn whatever the intended purpose.
NatureScot has been tasked by Scottish Government to develop a year-round licensing approach to muirburn. The Muirburn Code sets out statutory restrictions and good practice for practitioners who burn or cut vegetation. Currently muirburn can take place between 1st October and the 15th April (or 30th April with the permission of the land owner) without a licence. Out of season licences can be granted, provided certain criteria and conditions are met.
Developing an effective licensing approach for application throughout the year requires knowledge of the key processes and factors controlling the impacts of muirburn practices on biodiversity and soil conditions, carbon storage and risks to peatland and greenhouse gas (GHG) emissions. It also requires an understanding of muirburn’s relationship with the prevention and management of wildfires.
There is wide-ranging research on fire management practices worldwide, as well as research on elements of muirburn in the UK. Some of this research remains contested. To develop an effective, informed and practical licensing approach to muirburn NatureScot needs to base the guidance and licensing scheme it develops on relevant research and evidence. This review aims to provide an unbiased analysis of this existing evidence base.
The review focuses on the key questions set by NatureScot in their Statement of Requirements:
- What is muirburn?
- What is the current understanding of the relationship between muirburn and wildfire prevention? Can muirburn play a role in controlling the onset, intensity, severity and extent of wildfires?
- What is the current understanding of the impact of muirburn on carbon emissions and storage? What is the impact of muirburn on carbon functions (sequestration, GHG emission and other loss of carbon (C)) from soils and above ground vegetation? What are the timescales for recovery and resilience of carbon sequestration potential and GHG emissions following a muirburn event over immediate, short and long timescales on soils and habitats? What are the indicators to evaluate the dynamics of carbon losses and recovery following muirburn events?
- What is the current understanding of the impact of muirburn on habitats and species?
- What are the ranges and types of practical approaches that can be used to identify peat and peatlands relevant to inform muirburn management decisions? Knowing that the depth of peat layer is a commonly used criteria for identifying peat soils, have there been studies looking at the impacts of muirburn on different type/depth of peat soils? Have there been other factors used to identify peat and peatlands in studies of the assessment of the impacts of muirburn?
The review does not specifically look at the impact of muirburn as an inhibitor of natural vegetation succession (i.e. its role in preventing the establishment of woodland) or its impact on wider land use, as these areas are too far removed from the direct and immediate impacts that would be used to inform decisions relating to licensing.
This review gives no commentary on whether muirburn is an acceptable or necessary management practice for the management of moorland, just what the impact of muirburn is likely to be in practice. This is a review of the evidence; it does not provide recommendations as to how the evidence could be used by NatureScot and Scottish Government in terms of the licensing process.
2. Main findings
This review focuses on a set of key questions on the impact of muirburn set by NatureScot, concisely expressed as:
- What is muirburn?
- What is the relationship between muirburn and wildfire?
- What is the impact of muirburn on carbon emissions and storage?
- What is the impact of muirburn on habitats and species?
- What are the practical approaches that can be used to identify peat and peatlands?
This review, like many of the previous reviews that have been referenced, has found that the evidence base surrounding the impacts of muirburn on wildfire, soil carbon and habitats and species is somewhat limited and sometimes contested. The evidence base is most limited for the relationship of muirburn with wildfire, then carbon, with the most evidence on the impacts of muirburn on habitats and species.
A quick scoping review of the peer reviewed literature was carried out using the Web of Science online database using a set of search terms relevant to each of the three main questions (questions 2, 3 and 4 above). A limited number of selected grey literature sources were also reviewed.
- What is muirburn?
- Muirburn can be concisely defined as “the intentional and controlled burning of moorland vegetation”.
- What is the relationship between muirburn and wildfire?
- What we know
- There is evidence that muirburn causes a proportion of wildfires that occur on moorland, however, there remains uncertainty regarding this proportion.
- There is evidence from field studies that the immediate effect of burning on Calluna vulgaris-dominated habitats is to reduce above ground biomass (fuel load).
- Studies suggest that fire intensity in Calluna is controlled by fuel structure, windspeed and fuel moisture content.
- What we know we don’t know
- There is a lack of research into the question of whether variation in fuel loads resulting from muirburn influence the subsequent occurrence or likelihood of wildfire in moorland in the UK.
- This review found that the evidence base relating to the role of muirburn in controlling fire intensity and fire severity was too limited to draw firm conclusions.
- There was a lack of evidence relating to muirburn controlling the extent of wildfire in moorland.
- There was a lack of research into the relationship between muirburn and wildfire on non-Calluna dominated habitats.
- What we know
- What is the impact of muirburn on carbon emissions and storage?
- What we know
- There is evidence of burning of above-ground biomass on peat during muirburn with potential impact on carbon sequestration.*
- The data available appears to be focused on dissolved organic carbon (DOC) fluxes and aboveground (vegetation) carbon, with information on greenhouse gas emissions and soil carbon stocks being more limited.
- What we know we don’t know
- There is limited empirical evidence describing the effects of muirburn on carbon fluxes including greenhouse gas (GHG) emissions from UK moorlands.
- There is no overall consensus as to the net impacts of muirburn on carbon budgets, with evidence supporting gains, losses and no difference in carbon stores/fluxes following muirburn.
- The majority of data available is for stream or soil water dissolved organic carbon (DOC) concentrations. Few studies directly measured or calculated the effects of muirburn events on greenhouse gas emissions or carbon stocks prior and post fire events.
- There are few complete carbon budgets from UK moorland sites subject to managed burning. In particular, there is a lack of information on soil carbon stocks.
- There are few long-term studies from which the impacts of burning on carbon budgets can be determined, with limited geographical coverage. Those studies that do exist do not cover the range of habitats and habitat condition, and often have to make assumptions where data is lacking.
- There is little or no carbon budget data for dry heathlands or other important moorland habitats.
- What we know
- What is the impact of muirburn on habitats and species?
- What we know
- The impact and influence of muirburn on moorland habitats and species is complex.
- Moorland management (which includes managed burning) affects the abundance and diversity of bird species. Some species benefit from moorland management while others do not. It is difficult to disentangle the impact of muirburn on birds from the impact of other moorland management practices.
- Burning results in a change in plant species composition, but this change is not always consistent. Moorland vegetation responds to managed burning in a complex manner, with many factors including habitat type and condition, soil moisture content, geographical location, scale of muirburn management, burn rotation length, fire severity and intensity, air pollution and other management such as grazing, all having an effect.
- There is consistent evidence that Calluna cover increases over time following burning.
- There is mixed evidence as to the response of Sphagnum to prescribed burning, however there is evidence that Sphagnum capillifolium can recover from managed burning.
- What we know we don’t know
- In general, there is limited evidence describing the effects of muirburn on the full range of moorland habitats and species across the UK. For some moorland vegetation types, species groups and species, there is no evidence base, and for some others there is a contested evidence base. The review concludes that the evidence from the primary literature is focussed on blanket bog and upland heath, and no studies were identified from the Web of Science search that looked at the impact of muirburn on other important moorland habitats such as grassland (e.g. Molinia caerulea dominated grassland or Nardus stricta dominated grassland), flushes or alpine heaths.
- There is a lack of evidence on the impact of muirburn on small mammals, reptiles, or amphibians.
- The review concludes that the evidence base is too limited to draw firm conclusions regarding the impact of muirburn on terrestrial invertebrates.
- What we know
- What are the practical approaches that can be used to identify peat and peatlands?
- There are a number of approaches that can be used to identify peat and peatlands, however the only practical, reliable and cost-effective approach, is the use of some form of probe to measure either the actual peat depth or to ascertain whether the peat depth is beyond a particular threshold.
- There is a lack of evidence relating to the impact of muirburn on different depths of peat.
* This amended line replaces the previous line in the report to clarify the emphasis on impact. (There is some evidence of significant burning of soil surface peat biomass during muirburn rotation with likely impact on carbon sequestration.)
3. What is Muirburn?
3.1. Definition
For the purpose of this review the following concise definition of muirburn has been used:
“The intentional and controlled burning of moorland vegetation”
Muirburn is not specifically defined in the Muirburn Code (2021), however there is a definition in the “Supplement to the Muirburn Code: A guide to best practice” which was published to support a previous edition of the Code. In the supplement (page 1), muirburn is defined as:
“The knowledgeable and controlled application of fire to a predetermined area, at a specified time of day and season, and under specified weather and fuel conditions, so as to ensure that the intensity, rate of spread, and limits of spread of the fire meet planned resource management objectives. Prescribed burning of moorland in Scotland is usually referred to as muirburn, and there are four main land management objectives which apply:
- To produce a continuous supply of vigorous and nutritious new growth, by removing accumulated dead and woody plant material which makes the vegetation unpalatable and indigestible for grazing animals;
- To maintain moorland vegetation which is varied in composition and height, allowing greater access by livestock, and which provides increased foraging and nesting opportunities for moorland game and wildlife;
- To maintain the cover of heather, blaeberry and other characteristic moorland plants, in the long-term, so as to provide year-round forage and cover for livestock, moorland game and wildlife, and to maintain internationally renowned moorland landscapes;
- To reduce the accumulation of potential fuel and so reduce the risk of damaging, high intensity wildfires
3.2. Legal Context
The primary legislation governing muirburn in Scotland is the Hill Farming Act 1946, as amended by more recent legislation (the Climate Change (Scotland) Act 2009 and the Wildlife and Natural Environment (Scotland) Act 2011). There is no clear definition of muirburn in the Hill Farming Act 1946, the Climate Change (Scotland) Act 2009 or the Wildlife and Natural Environment (Scotland) Act 2011. Muirburn is also subject to legislation that applies to specific sites including areas designated for their natural or cultural heritage, or to protected species and their nests. Further legislation applies to muirburn in relation to burning close to public roads, and in certain cases, smoke from muirburn fires may be subject to legislation if it is a public nuisance (Scotland’s Moorland Forum, 2017b; RSPB, 2021). The legislation covers the burning of all moorland vegetation, including species such as gorse.
The Muirburn Code produced by Scotland’s Moorland Forum (updated in May 2021) provides guidance on good practice for burning moorland vegetation and sets out the statutory regulations that must be complied with under the legislation governing muirburn.
3.3. What is Moorland?
Moorland or moor is a broad habitat mosaic found mainly in the uplands, which is characterised by low-growing vegetation, usually on acidic soils that are often low in nutrients and sometimes water-logged. Moorland habitats are comprised of a range of uncultivated upland vegetation types including wet and dry heath, blanket bog, wet and dry grasslands and bracken. Moorland habitat can extend down to sea-level, particularly in the north and west of Scotland, and as there is no defined upper altitudinal limit to moorland, it can include a range of alpine communities at high altitude. Vegetation types that would not be included under the term moorland include improved and semi-improved grassland, arable land, lowland swamps and fens, woodland, scrub (on agriculturally improved ground, including Ulex europaeus on inbye ground), salt marsh and other coastal communities, as well as parks and other urban green spaces. Although not usually described as moorland, the Muirburn Code applies to lowland heath and coastal heath. The Muirburn Code does not apply to other lowland habitats such as semi-natural grasslands, lowland fens or scrub.
3.4. Moorland Habitats
The main moorland habitats are upland dry heaths (dominated by Calluna vulgaris (henceforth referred to as Calluna), Vaccinium myrtillus and Erica cinerea), wet heaths (dominated by Erica tetralix and Molinia caerulea), blanket bogs on peat soils (with more than 50cm of peat, as defined in Scotland), dry and wet grasslands and bracken (Figure 1; see Annex 1)
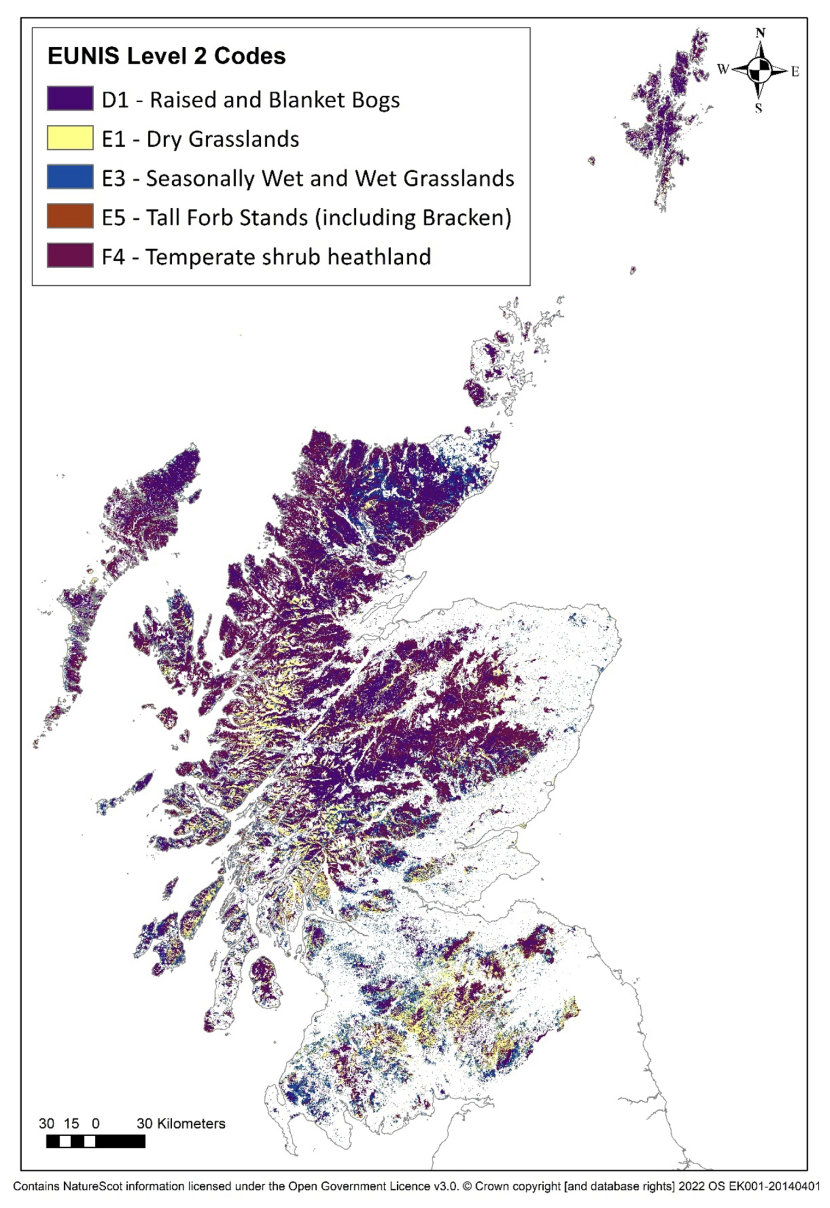
Figure 1 - Main moorland habitat types. (This is a EUNIS level 2 habitat map and may include some level 4 habitat types that are not moorland (Strachan, 2017)).
There are a range of other vegetation types that often occur within a moorland mosaic. These include certain scrub communities (Salix aurita, Ulex europaeus and Juniperus communis), as well as a range of upland springs, flushes, fens and mires. In addition, there are a range of other alpine and sub-alpine habitat types that occur mainly at high altitude. These include alpine and subalpine grasslands; subalpine moist or wet tall-herb and fern stands; arctic, alpine and subalpine scrub; and evergreen alpine and subalpine heath and scrub (Figure 2; see Annex 2 for the other moorland EUNIS habitats and their corresponding NVC types). These vegetation types would not be expected to form the primary focus of muirburn but as they may occur within the moorland mosaic they may be impacted by it to differing extents.
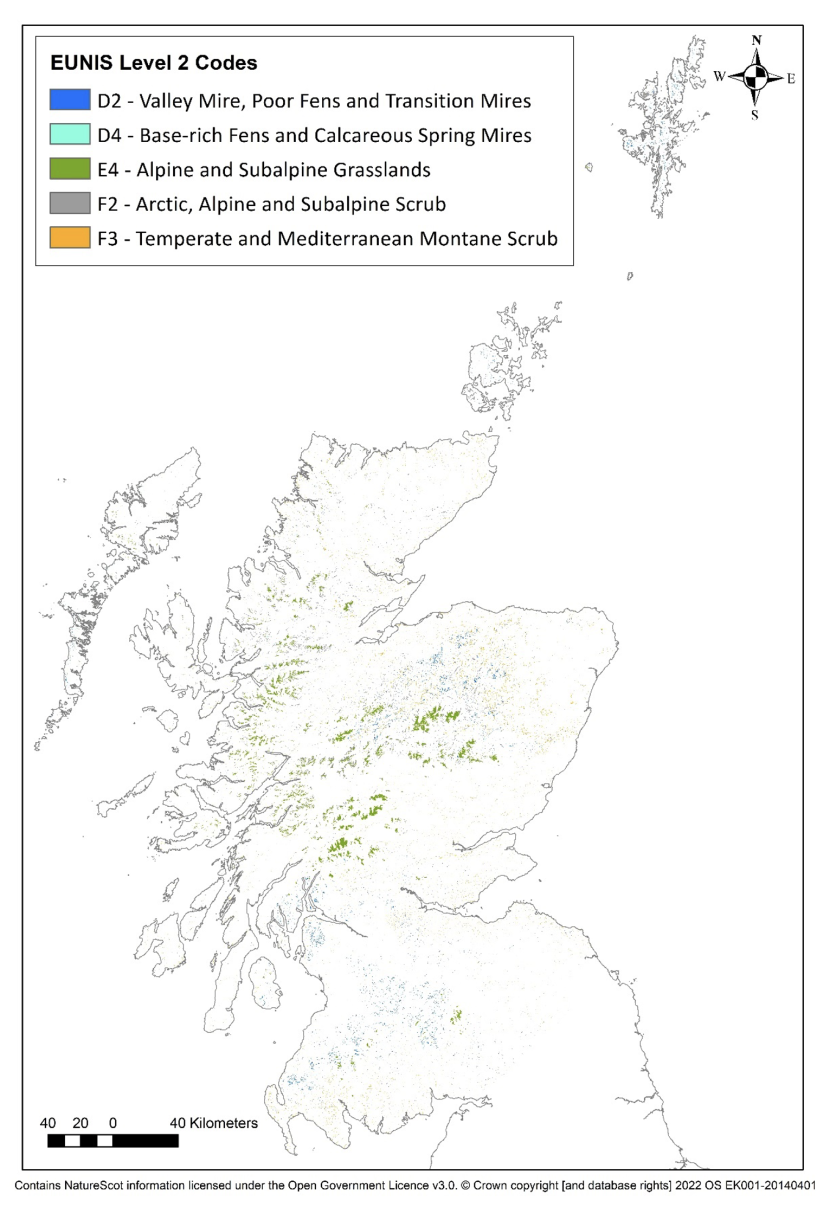
Figure 2 - Other moorland, alpine and sub-alpine habitat types. (This is a EUNIS level 2 habitat map and may include some level 4 habitat types that are not moorland (Strachan, 2017)).
4. Review Methodology and Approach
A quick scoping review (Collins et al., 2015) of the peer reviewed literature was carried out. While seeking to maintain rigour and transparency, it should be noted that due to the limited time and resources available this was not an exhaustive systematic review.
The team carrying out this review are not directly involved in muirburn research, have no vested interest in muirburn or moorland management, and have no pre-conceived opinions on muirburn management.
For each of the three topic areas (i.e. the relationship between muirburn and wildfire; the impact of muirburn on soils and soil carbon; the impact of muirburn on habitats and species) the review team agreed upon a set of search terms to locate the relevant literature (see Table 1). These search terms were split into three categories: terms relating to moorland and peatland (population); terms relating to muirburn and prescribed burning (intervention); and terms relating to the particular topic area (outcome). The search terms for the biodiversity outcome were more extensive than for the other two outcomes due to the much wider scope of this topic area.
Due to the limited time available, only the Web of Science online database was used to search for relevant peer-reviewed scientific literature.
| CATEGORY | Search terms |
|---|---|
| Population: Moor | heath* OR moor* OR bog OR peat* OR heather OR Calluna OR Molinia OR mire OR fen* OR flush* |
| Intervention: Muirburn | "prescribed burn*" OR muirburn or "manag* burn*" OR "manag* fire" OR "control* burn*" OR "rotation* burn*" OR "pastoral fire" OR "fuel reduction" OR swaling |
| Outcome: Soil / Carbon | soil OR carbon |
| Outcome: Biodiversity | habitat OR biodiversity OR conservation OR "species diversity" OR ecosystem* OR flor* OR fauna OR animal* OR bird* OR insect* OR invertebrate* OR communit* OR species OR composition OR frequency OR abundance OR diversity OR structure OR NVC OR cover OR richness OR sphagnum moss* OR sphagn* OR Erica tetralix OR Eriophor* OR Molinia OR Calluna OR Trichophorum OR population* OR “breeding success” OR restor* OR revegetat* OR “vegetation management” OR remediation OR regenerat* OR livestock OR stocking OR grazing OR designated site* OR SSSI OR “site* of special scientific interest” OR water catchment* OR “special area* of conservation” OR SAC* OR “special protection area*” OR SPA |
| Outcome: Wildfire | wildfire OR fire OR "wild land fire" OR "fuel load" OR "fire prevention" OR "fire risk" OR "fire suppression" |
In addition, the impacts of muirburn have been looked at previously in a number of reports and reviews. It was not possible to do a full review of the grey literature, so a number of key publications, identified by the review team in consultation with NatureScot, that were commissioned by the Scottish Government, ClimateXChange, and Natural England (which include comprehensive systematic reviews) were used as a basis for this current review (see below). Following comments from an independent reviewer, the main findings from an additional DEFRA funded study by Heinemeyer et al. (2019) were also included in the habitats and species section. Although these reviews and reports do not specifically focus on the questions being asked in this current review, they do contain information that is relevant and appropriate.
- Werritty, A., Pakeman, R.J., Shedden, C., Smith, A. and Wilson, J.D. 2015. A Review of Sustainable Moorland Management. Report to the Scientific Advisory Committee of Scottish Natural Heritage.
- Werritty, A. 2019. Grouse Moor Management Review Group. Report to the Scottish Government.
- Brooker, R., Hester, A., Newey, S. and Pakeman, R. 2018. Socio-economic and biodiversity impacts of driven grouse moors in Scotland: Part 2 Biodiversity impacts of driven grouse moors in Scotland. Report to the Scottish Government.
- Newey, S., Fielding, D., Miller, D.G., Matthews, K.B. and Thomson, S. 2020. Biodiversity considerations on grouse moors. Part 4 of Research to assess socioeconomic and biodiversity impacts of driven grouse moors and to understand the rights of gamekeepers. Report to the Scottish Government.
- Chapman, S., Hester, A., Irvine, J. and Pakeman, R. 2017. Muirburn, peatland and peat soils - an evidence assessment of impact. ClimateXChange Report.
- Glaves, D.J., Morecroft, M., Fitzgibbon, C., Leppitt, P., Owen, M. and Phillips, S. 2013. The effects of managed burning on upland peatland biodiversity, carbon and water. Natural England Evidence Review, NEER004.
- Glaves, D.J., Crowle, A.J.W., Bruemmer, C. and Lenaghan, S.A. 2020. The causes and prevention of wildfire on heathlands and peatlands in England. Natural England Evidence Review NEER014.
- Baggaley, N.J., Britton, A.J., Barnes, A., Buckingham, S., Holland, J.P., Lilly, A., Pakeman, R.J., Rees, R.M., Taylor, A. and Yeluripati, J. 2021. Understanding carbon sequestration in upland habitats. ClimateXChange Report.
- Heinemeyer, A., Vallack, H.W., Morton, P.A., Pateman, R., Dytham, C., Ineson, P., McClean, C., Bristow, C. and Pearce-Higgins, J.W. 2019. Restoration of heather-dominated blanket bog vegetation on grouse moors for biodiversity, carbon storage, greenhouse gas emissions and water regulation: comparing burning to alternative mowing and uncut management. Final Report DEFRA Project BD5104. DEFRA.
The peer reviewed articles were screened for relevance to the specific questions that had been set by NatureScot. Papers that did not directly relate to the questions were removed; as were papers that did not relate to moorland habitats, species or soils (for example forest environments), or were from locations with non-temperate climates that were not relevant to Scottish moorlands (for example the Mediterranean, Arctic, California, Australia etc.). The screened references were reviewed, and an assessment was made of their scientific robustness and relevance to the research questions using a set of criteria (see Table 2 and Annex 6, Annex 9 and Annex 12). The criteria involved an assessment of the study design (i.e. whether the study was replicated and had controls); the geographical coverage (i.e. the number of contrasting study sites); the system diversity coverage (i.e. the number of vegetation or soil types studied); the number of burn events assessed; and whether sampling was carried out pre, during and post burning. For the carbon question an additional criterion was included related to the number of key areas of a carbon budget included in the study (i.e. soil carbon, vegetation carbon, water (dissolved organic carbon (DOC)) and/or gas measurements). For each criterion, a high, medium and low rating was given where possible. Information on the geographical location, habitat/soil type, type of study (field study, laboratory based, modelling) and key findings from each of the papers were extracted.
| Criteria | Explaination |
|---|---|
| Experimental design - High | The study includes both control sites and is replicated |
| Experimental design - Medium | The study includes control sites or has replication |
| Experimental design - Low | The study does not include control sites or replication |
| Geographical coverage - High | The study includes 3 or more contrasting experimental/study sites |
| Geographical coverage - Medium | The study includes 2 contrasting experimental/study sites |
| Geographical coverage - Low | The study includes 1 experimental/study site |
| System diversity coverage - High | The study includes 3 or more contrasting soil or vegetation types |
| System diversity coverage - Medium | The study includes 2 contrasting soil or vegetation types |
| System diversity coverage - Low | The study includes 1 soil or vegetation type |
| Number of burn events - High | The study includes the monitoring of 3 or more burn events (or 3 ages of burned vegetation) or includes the monitoring of short rotation and long rotation burns |
| Number of burn events - Medium | The study includes the monitoring of 2 burn events (or 2 ages of burned vegetation) |
| Number of burn events - Low | The study includes the monitoring of 1 burn event (or 1 age of burned vegetation) |
| Burn event coverage - High | The study includes samples covering pre, during (or immediately after burning), and post burn events |
| Burn event coverage - Medium | The study includes samples covering pre and post burn events or uses a space-for-time substitution study |
| Burn event coverage - Low | The study includes samples covering only post burn events |
| Not applicable | The criterion is not applicable to this study or not enough information is provided, or it is unclear |
The main findings and conclusions that have been reached following the review process were based on a combination of the number of studies/sources of evidence, the scientific robustness of those studies, the consistency of the evidence, and the relevance of the evidence to the questions that had been set.
The previous reviews and other grey literature were reviewed separately from the peer reviewed articles. Some of the previous reviews used their own robustness criteria and gave assessments of the level of evidence available. Where this is the case, this is indicated in this report.
5. The Relationship Between Muirburn and Wildfire
5.1. Summary of Main Evidence
- The primary literature search generated 259 publications of these 22 were included in the review.
- The review process identified six primary sources which estimated the proportion of wildfires that result from managed burning, giving a range of 15%-60%, or 24%-68 if lowland statistics are excluded. Due to the limited evidence base and caveats acknowledged by the source authors the review team note no basis to offer a degree of confidence in this range. The review concludes that there is evidence that muirburn directly causes a proportion of wildfires that occur, however there remains uncertainty regarding this proportion.
- The search process identified no primary studies which directly studied whether variation in fuel loads resulting from muirburn influence the subsequent occurrence or likelihood of wildfire on moorland. This is in agreement with what has been reported by previous reviews, and authors in the field; in that there has been a lack of research into this question in the Scottish/UK context (Worrall et al., 2010b; Glaves et al., 2013; Davies et al., 2016b; Luxmoore, 2018; Glaves et al., 2020).
- With respect to fuel management, there is evidence from field studies that the immediate effect of burning on Calluna moorland is to reduce above ground biomass. Modelling studies suggest that shorter burning rotations maintain lower average biomass.
- The review process identified a small number of studies which have modelled fire behaviour in Calluna. Overall, this evidence base is too limited to draw firm conclusions however these studies suggest that:
- Fireline intensity;and rate of spread are influenced by vegetation structure, making fires more difficult to predict and control. Insofar as muirburn does influence fuel structure, this indicates a plausible mechanism through which muirburn may influence the intensity of wildfires.
- Calluna fuel load and structure may influence the severity of burns, however as soil moisture conditions are also found to control fire severity, the marginal effect (as distinct from other factors) of Calluna fuel load and structure on fire severity across these studies is unclear.
5.2. Research Questions - Muirburn and Wildfires
The research question set up for this review was:
a) What is the role of muirburn in controlling the onset, intensity, severity and extent of wildfires?
Subsequently this research question has been considered as three separate but related sub-questions:
- What is the role of muirburn in controlling the onset of wildfires?
- What is the role of muirburn in controlling the intensity and severity of wildfires?
- What is the role of muirburn in controlling the extent of wildfires?
In its approach to the research questions, the review considered:
- The extent to which the practice of muirburn may be a direct cause of wildfire, due to the potential for managed burns to escape control; and
- The potential for muirburn to control the occurrence, intensity, severity and extent of wildfires by controlling the build-up of fuel.
5.3. Search for Primary Literature
A search for primary literature was conducted in the Web of Science database. Search terms and syntax are detailed in Table 3.
The search generated 259 results, of these 22 were included in the review and 237 excluded on the basis that they did not provide relevant evidence to the review questions.
Screening for relevance followed an iterative process. Sources were assessed first on the basis of their title, then abstract, then on a scan of their full text. At each stage, sources were excluded if it was clear that they did not include information relevant to the research questions. Sources were removed where the habitat of the study clearly differed to the moorland habitat as defined for the review, for instance those reporting results for land uses (e.g. forest systems) and climates (Mediterranean, California, Australia) that are outside of the moorland habitat as defined for the review. Sources were removed relating to outcomes other than wildfire occurrence, intensity, severity, extent, or fire behaviour and characteristics following managed burning (e.g. wildfire emissions). Sources were removed relating to interventions other than muirburn or managed burning (e.g. fire-fighting chemicals).
| Search terms | heath* OR moor* OR bog OR peat* OR heather OR calluna OR molinia OR mire OR fen* OR flush* AND "prescribed burn*" OR muirburn or "manag* burn*" OR "manag* fire" OR "control* burn*" OR "rotation* burn*" OR "pastoral fire" OR "fuel reduction" OR swaling AND wildfire OR fire OR "wild land fire" OR "fuel load" OR "fire prevention" OR "fire risk" OR "fire suppression" OR licensing |
|---|---|
| Tool: | Web of Science |
| No. hits | 259 |
| Sources excluded from the review | Sources removed on screening of title = 116 Sources removed on screening of abstract = 74 Sources removed on screening of text = 9 Sources subsequently removed at further reading = 38 |
| Sources Included | Sources included = 22 |
In addition to the 22 sources included from the Web of Science search, a further 7 sources were included from searches of the grey literature, a further 12 sources were included from cross-citation, and one further source was included following notification by an independent reviewer. Five review studies were pre-known to the review team. See Annex 3 and Annex 4 for more details of the papers included in the review.
Primary studies: 29
Position/discussion papers: 10
Methodological criticism: 2
Review studies: 5
An assessment of the robustness and relevance to the research questions of the papers was carried out (see Annex 5) using a set of assessment criteria (see Annex 6). The modelling papers and papers using wildfire occurrence statistics were not included in the assessment of robustness and relevance as they did not fit the assessment criteria.
5.4. Research Question 1: What is the Role of Muirburn in Controlling the Onset of Wildfires?
In approaching this question, the review team considered:
- What is the evidence that muirburn directly causes a proportion of wildfires?
- What is the evidence that muirburn reduces fuel load?
- What is the evidence that muirburn influences wildfire occurrence by reducing fuel load?
5.4.1. What is the evidence that muirburn causes a proportion of wildfires in moorland?
The Web of Science search identified no primary sources which had studied the relationship between wildfire occurrence and muirburn in moorland habitats. Five sources were subsequently identified from an existing review of the causes and prevention of wildfire on heathlands and peatland in England, carried out by Glaves et al. (2020). These were assessed independently by the review team as part of the review process and included in the review. One further relevant source was identified by the review team during a search of the grey literature.
The review process identified six primary sources which estimated the proportion of wildfire that results from managed burning (Table 4), giving a range of 15% to 60%, or 24% to 68% if lowland statistics are excluded. Due to the limited evidence base and caveats acknowledged by the source authors the review team note no basis to offer a degree of confidence in this range.
The review concludes that there is evidence that muirburn directly causes a proportion of wildfires that occur, however there remains uncertainty regarding this proportion.
| Source | Description | Data | Temporality | Finding |
|---|---|---|---|---|
| Cosgrove (2004) | Summary statistics | Data reported to Badenoch and Strathspey Fire Protection Group | 2003 | The supposed cause of 4 out of 14 (29%) were muirburn activities |
| Legg et al. (2006) | Questionnaire administered to 42 Scottish Estates | Questionnaire administered by the Game Conservancy Trust and Scottish Gamekeepers Association | 2003 | 9 of 17 (53%) from escaped managed fires |
| Worrall et al. (2010b) | Ranger reports in the Peak District National Park | Ranger reports | 1976- 2004 | 10 of 41 (24%) attributed to escaped managed burns |
| Martin (2018) | Case study of Darwen and Tufton Moors | Mapping and description of 22 wildfires over a period of 22 years | 1995- 2017 | 8 of 22 (36%) likely due to escaped managed burns (7 from burns on Molinia grassland for livestock grazing, and one from an out-of-control grouse moor strip burn getting out-of-control) |
| Luxmoore (2018) | Scotland wide analysis of FRS records | FRS | 2009/10- 2014/15 | 140 of 233 (60%) potentially caused by muirburn |
| Glaves et al. (2020) | Glaves et al. (2020) Analysis of amalgamated English wildfire data | Data submitted to Natural England by; Dorset County Council, Lancashire Fire & Rescue Services, Moors for the Future, and Peak District National Park | 2011- 2018 | Uplands and lowlands (15%) due to escaped managed burns Uplands only (68%) due to escaped managed burns. |
5.4.2. What is the evidence that muirburn reduces fuel load?
Fuel Load: the total amount of combustible material (fuel) expressed quantitatively in terms of mass per unit area.
The search process identified five primary studies which have studied the effect of burning on above ground biomass in Calluna moorland. Results from a further four modelling studies are introduced.
There is evidence that the immediate effect of managed burning on Calluna heath is to reduce above ground biomass.
From after the fact sampling of recent burns at two sites in Northern England, Farage et al. (2009) found that burning reduced above ground biomass by 16 ± 4% in their first burn year and 24 ± 5% in their second burn year.
Subsequent commentors have questioned the findings of Farage et al. (2009), commenting that the reported biomass remaining following burning (1262 g m-2) is unusually high and that an underestimate could arise from their measurement approach based on paired plots (Legg et al., 2010).
From before and after assessment at six managed burns at the same site in the Peak District, Worrall et al. (2013b) estimated the average biomass loss during burns to be 75 +/- 9%.
From after the fact assessment of five peatland wildfires (Davies et al., 2016a) and 27 controlled burns reported in Legg and Davies (2007), Davies et al. (2016a) found that the consumption of surface fuels (heather and graminoids) is a roughly constant proportion of pre-fire fuel load. Modelling the relationship, they found a positive linear relationship between pre-fire fuel load and mean fuel consumption for surface fuels. Controlling for Fire Weather Information variables (the dryness of ground conditions as assessed using the Canadian Fire Weather Information System indices) did not improve model fit. No such relationship was found for ground fuels (mosses, litter, tussock-bases, and woody stems).
Taylor et al. (2021) conducted two studies of flame spread, one comprising sixteen laboratory scale flame spread experiments in Calluna dwarf-shrub, recreated under controlled laboratory conditions (no wind), another comprising ten controlled burns on Calluna heath near Glensaugh Farm in North-East Scotland. In their laboratory experiment they measured total biomass pre- and post-burn. For fires that successfully spread they reported the percentage of biomass consumed ranged from 51.29% - 98.17%. In their field experiment they similarly measured total biomass pre- and post-burn, reporting average biomass consumed ranging from 49.0% to 93.3%. In neither study did they note whether the proportion of surface and ground fuels consumed differed.
From assessment of four replicate blocks at Moor House in the North Pennines of Northern England, Milligan et al. (2018) found that vegetation structure (height) varied observably between three burn rotations, 10-years, 20-years, and no-burn since 1954. For the 10-year rotation vegetation height was clustered in the 0-20 cm category, with a marked reduction above. For the 20-year and no-burn rotations, vegetation height was more variable, with a peak in the 20-30 cm category.
The following modelling studies drawing on chronosequence data predict that average biomass will be reduced where burning rotations are shorter.
From controlled burning experiments of five moorlands in the Peak District National Park in Northern England along a chronosequence, Allen et al. (2013) developed a matrix model of above ground biomass. From this they predicted that above ground biomass would be greater where the rotation interval of prescribed burning is longer. Modelling the effect of an increased frequency of prescribed burning was found to reduce above ground biomass.
Following Allen et al. (2013), Santana et al. (2016) developed matrix models of above ground biomass at four UK sites along a North-South gradient. Comparing across models they found that above ground biomass and litter accumulation patterns differed between sites, although the differences were not ordered along a North-South gradient. From modelling simulations, they predicted that above ground biomass will be reduced with shorter burning rotations.
Clay et al. (2015) measured biomass across two sites in Northern England along a 10-year chronosequence. Comparing burned plots to no-burn control plots they found that the average biomass for burned plots was less than half that of no-burn control plots. Fitting a regression on years since last burn they estimated that around 80% of biomass was lost during burns.
In a study assessing the combined influence of burning and grazing Alday et al. (2015) compared plots at Moor House, in the North Pennines of Northern England, burned on 10-year and 20-year rotations to no-burn control plots. They found that managed burning reduced total biomass relative to the control group significantly and that this effect was greater for the shorter rotation group. There was no effect of grazing, either individually or through interaction. Through non-linear mixed effects regression, they estimated that Calluna biomass and height reached asymptotes at 20 years and 15 years, respectively.
Whitehead et al. (2021) studied vegetation response following management burns conducted over a ten-year period at Langholm Moor in South-West Scotland. From forty-eight plots, eight in each of six burn years, their analysis found that years since burning is a significant predictor of heather cover, with heather cover increasing with the number of years since burning. They further found that biomass, height and heather coverage were strongly positively correlated.
With respect to fuel management, the review concludes that there is evidence from field studies that the immediate effect of burning on Calluna moorland is to reduce above ground biomass. Modelling studies suggest that shorter burning rotations maintain lower average biomass among burned plots.
5.4.3. What is the evidence that muirburn influences wildfire occurrence by reducing fuel load?
The search process identified no primary studies which have directly studied whether variation in fuel loads resulting from muirburn influence the occurrence of wildfire in moorland. This is in agreement with what has been reported by previous reviews, and authors in the field; in that there has been a lack of research into this question in the Scottish / UK context (Worrall et al., 2010b; Glaves et al., 2013; Davies et al., 2016b; Luxmoore, 2018; Glaves et al., 2020).
5.4.3.1. Contextual accounts which suggest that by managing fuel load, muirburn may influence wildfire occurrence
Three contextual accounts are introduced here which suggest that fuel management may have an influence on wildfire occurrence. The Web of Science search identified one source, Log et al. (2017) which provides a contextual account of two large wildfires in Calluna-dominated Norwegian coastal heath. Two further contextual accounts are reported from preceding reviews.
Glaves et al. (2013, 2020) and Werritty et al. (2015) cited McMorrow et al. (2009) who found that ‘heather moorland’ in the Peak District of Northern England (which is mostly managed by rotational burning) experienced fewer wildfires than other moorland habitats, relative to its size. This has been taken as an indication that fuel management through managed burning may have a role in reducing wildfire occurrence in the Peak District.
Glaves et al. (2020) noted Luxmoore’s (2018) account based on an assessment of the National Trust for Scotland Estate, where no managed burning is carried out, that over a period of eighteen years there were 12 wildfires affecting 1,463 ha (less than 2% of the area), indicating that while a policy of no-burning has not caused wildfire to cease at the National Trust for Scotland Estate, comparatively it has not resulted in widespread wildfire.
Giving an after the fact assessment of the conditions leading up to two large wildfires in Norway, Log et al. (2017) argue that a lack of controlled burning during the fifty years prior had led to the build-up of a high fuel load, in the form of late developmental phase Calluna and increased abundance of fire prone tree species. The occurrence of particular climatic conditions; below zero temperatures in the three-weeks preceding, low relative humidity and cold air from the mountains had provided the conditions for adiabatic heating, in effect freeze drying the Calluna and resulting in winter desiccation. They claim that these conditions made the area susceptible to ignition and sustained burning and that a lack of fuel load management had been an enabling condition for the fires, which on the basis of property damage were the most severe in Norway since World War II.
The review concludes that there is a lack of evidence from field studies that variation in fuel loads resulting from muirburn influence the occurrence of wildfire in moorland. There has been a lack of research into this question in the Scottish / UK context.
5.5. Research Question 2: What is the Role of Muirburn in Controlling the Intensity and Severity of Wildfires?
In approaching this question, the review team considered the potential influence of fuel management through muirburn on the intensity and severity of wildfires in the Scottish moorland context defined for the review.
Intensity and severity are introduced together as the evidence base that has been identified overlaps significantly.
Intensity and severity are two key aspects of fire behaviour, they are defined in the Supplementary Information to The Muirburn Code (Scotland’s Moorland Forum, 2017c, pages 4.1 and 4.2) as follows:
“Fire Intensity: The intensity of a fire is a measure of the rate that energy is released by a fire and is a combination of the amount of fuel that burns and the rate of spread of the fire. A good guide in the field to the intensity of a fire is the flame length. Fire intensity is used to judge how difficult a fire is to put out”.
“Fire Severity: A general term most commonly used to describe the combined effects of both flaming and smouldering combustion on a fire site that creates impacts on plants, soils, and wildlife”.
“Burn Severity: A qualitative assessment of the heat pulse directed toward the ground during a fire. Burn severity relates to soil heating, large fuel and duff consumption, consumption of the litter and organic layer beneath trees and isolated shrubs, and mortality of buried plant parts”.
See Keeley (2009) for more detailed definitions of the terms fire intensity, fire severity and burn severity.
A previous review (Glaves et al., 2020) noted a lack of research investigating the effect of fuel load management through muirburn on wildfire intensity/severity in the UK context.
The present review similarly notes a lack of studies that have directly investigated whether variation in fuel load due to muirburn results in changes to wildfire intensity or severity. Reflecting on this, the review team considered that to design a field experiment addressing this question at the spatial scale on which wildfires occur would be infeasible as it would involve subjecting large areas of land to elevated risk. In this context feasible study designs are effectively limited to smaller scale experimental studies or statistical modelling.
The search process identified seven controlled burning studies in Calluna moorland which have investigated the factors influencing fire behaviour. While these have been carried out on a limited spatial scale, and in a more limited range of conditions than occur more generally in the overall Scottish moorland context, the results from these studies are considered relevant to the review questions.
5.5.1. What is the evidence that fuel load and structure control the intensity of wildfire?
Fuel Structure
Within the research cited in this review, the term fuel structure is used variously to refer to:
- the proportion of different fuel types; e.g. fine versus coarse fuel;
- the height and density of the Calluna canopy; and
- the continuity and uniformity of fuel load across the landscape.
For a general discussion on fuel characteristics and their influence on wildfire behaviour see Section 8, Part B, 1. “The Effects of Fuel” in, Scottish Fire and Rescue Services (2011) Wildfire Operational Guidance.
Calluna Development Phase
Davies et al. (2009; 2019) use Calluna development phase as a measure of fuel structure.
Gimingham (1988) described four characteristic phases in Calluna morphology:
- “Pioneer. Early stages of establishment and growth. Regular branching from the axis of a single leading shoot, at first. Height up to ca 6 cm, shape pyramidal, cover incomplete. Up to 6 years of age.
- Building. Becoming bushy, branches radiating from the centre, productivity of peripheral shoots high and flowering vigorous. Height up to ca 0.5 m (or more), shape hemispherical, canopy dense, cover approaching 100%. Up to about 15 years of age (depending on habitat).
- Mature. Extension growth declining, though green shoot production and flowering still high. Shape still hemispherical, but central branches inclined to spread sideways, initiating gap formation; cover slightly reduced. Up to 20 years of age (or more).
- Degenerate. Central branches dying, creating gap. Some other branches which have become partially buried in litter etc. may remain alive (because of adventitious rooting), sustaining a ring of foliage bearing twigs. Cover much reduced. Eventually whole plant may die. Death at 30-40 years of age.”
As described by Gimingham (1988) Calluna development phases are distinguished by changes in branch structure as well as the height, shape, density and continuity of the canopy. Davies et al. (2009) further note characteristic changes in the proportion of fine fuels and coarse stems that occur across development phases.
Empirically, and further distinguishing Late Building from Building phases, Davies et al. (2009) find significant differences in fuel load, height and bulk density (their CDI measure) across three development phases in their study – Building, Late Building and Mature.
From controlled burning experiments (twenty ignition attempts) in Calluna at two sites in Scotland, Davies and Legg (2011) found that moisture content of Calluna in the lower canopy, and the moss/litter layer controlled both ignition and rate of spread. Davis and Legg (2011) found a critical threshold of 60-70% fuel moisture content in the lower canopy for sustaining fires; above 70% both spot and line ignitions failed to spread, while below 60% fires developed rapidly.
Taylor et al. (2021) conducted two studies of flame spread, one comprising sixteen laboratory scale flame spread experiments in Calluna dwarf-shrub recreated under controlled laboratory conditions (no wind), another comprising ten controlled burns on Calluna heath near Glensaugh Farm in North-East Scotland.
To conduct their laboratory experiments sixteen 1.8 m by 1.0 m fuel beds were formed using fuel harvested from plots in the Pentland Hills Regional Park in South-East Scotland. In doing so they sought to recreate the fuel loadings observed where the fuel was harvested, distinguishing four fuel types: fine green, fine dead, coarse, and moss. From these they observed qualitatively that the fuel moisture content of the fine dead fuel was a dominant factor in determining rate of spread, and length of flames. However this alone was not sufficient to support successful flame spread (whether or not the flame traversed the entire length of the fuel bed) under all conditions. They found that successful flame spread must be supported by sufficiently low fuel moisture content in the other fuels, particularly the coarser heather and the fine green.
In a separate study of ten controlled burns in Calluna heath, Taylor et al. (2021) observed the variation in rate of spread and length of flames across burns. Examining the pairwise correlation between rate of spread and environmental variables; relative humidity; wind speed; shrub fuel load; and fuel moisture content they found no discernible trends (that might explain variation in rate of spread). They further noted that the fuel moisture content values measured in the field for which fire successfully spread were higher than the thresholds determined in the laboratory. They reason that this can be explained due to the influence of wind which they consider would support the flame spread in the field but was absent in the laboratory experiments.
From 27 controlled burns at two sites in Scotland, Davies et al. (2009) found that rate of spread was controlled by wind speed, vegetation height, and live fuel moisture content, with the relationship primarily determined by windspeed. They further found an interaction between windspeed and vegetation structure, with a stronger effect of windspeed on rate of spread observed in plots of mature Calluna.
From 27 controlled burns at two sites in Scotland, Davies et al. (2019) found that fireline intensity (the rate of heat output per metre kW m-1) and flame height increased significantly across three developmental phases of Calluna. From best subsets regression they concluded that fireline intensity could be adequately modelled from fuel height and wind speed, although model predictions were improved by further accounting for live fuel moisture content.
Overall, this evidence base is too limited to draw firm conclusions, however these studies suggest that fire intensity in Calluna is controlled by fuel structure, windspeed and fuel moisture content. Insofar as muirburn does influence fuel structure, this indicates a plausible mechanism through which muirburn may influence the intensity of wildfires.
Some caveats should be noted:
This is a relatively small evidence base drawn from a limited number of sites in Scotland.
These controlled burning studies were only carried out in Calluna, and therefore do not fully reflect the variety of moorland fuel types, notably grasses and sedges as well as other dwarf shrubs and gorse, which experience different patterns of flammability over the course of a year.
These studies were only carried out during the legal burning season and therefore do not reflect potential differences in fire behaviour in summer.
They were carried out on a relatively small scale. The plot size in Davies and Legg (2011) ignition experiments was 2 m by 2 m, while two designs 15 m by 20 m and 20 m by 20 m were used in Davies et al. (2009; 2019). Taylor et al. (2021) used 1.8 m by 1.0 m fuel beds in their laboratory experiment, and 10 m by 10 m plots in their field experiment. Accordingly, potential changes in fireline intensity that occur over greater scales will not be reflected.
5.5.2. What is the evidence that fuel load and structure control the severity of wildfire?
From fifteen experimental burns at Crubenmore, near Dalwhinnie, Davies (2005) applied a generalised linear model to assess the influence of fuel group (high, medium, low) on various measures of fire severity. The study found that fuel group could explain variation in fuel consumption (a measure of fire severity). No statistically significant relationship was observed for fuel group with respect to other measures of severity; consumption of the moss-litter layer or ground surface heating. Separately, the study observed significant variation in ground surface heating between burn days, but found this could not be explained by variation in fuel load, rate of spread or fireline intensity.
From after the fact assessment of five UK wildfires Davies et al. (2016a) found results which suggest that the severity of wildfires in Calluna dominated moorland is influenced by fuel structure and fuel load (Davies et al., 2016a). This finding is based firstly; on the observation that severity (as measured by a composite burn index, which reflects damage across multiple fuel layers) varied substantially within particular wildfires, sometimes more than the average variation that was observed between wildfires; and secondly on the basis of statistical analysis in which fixed factors determined by sub plot location were found to explain variation in fuel consumption (a further measure of fire severity). The authors reason that this is due to variation in fuel load and structure present at the stand level and that these explain the variation in severity (as measured by fuel consumption). However as these were not directly observed before the fact, this is in effect an assumption. Fuel consumption is estimated after the fact through comparison of paired burned and unburned plots. The authors maintain that plots can be matched sufficiently well by measuring the diameter of remaining stems, yet it must be noted that this approach overall will entail a degree of measurement imprecision.
In a separate analysis of the same five wildfires, Davies et al. (2016a) found that variation in severity (as measured by a composite burn index, which reflects damage across different fuel layers) could be explained by the dryness of ground conditions on the day, as measured by indices from the Canadian Fire Weather Information System.
A further controlled burning study similarly found a significant role for soil moisture in controlling severity, yet reported no result for Calluna fuel load or structure. From 19 experimental burns at two sites in Scotland, Grau-Andrés et al. (2018) found that the moisture content of the moss-litter layer, controlled variation in fire severity, and that their dry heath site was more sensitive to this change than the wet bog site. At both sites, a simulated drought led to increased consumption of the moss-litter layer (one measure of severity) when burned, relative to control plots where no drought treatment was applied. The effect on soil heating (a further measure of severity) however differed between sites. In burns following their simulated drought treatment, mean time above the ecologically significant threshold of 50oC measured at the soil surface increased from 34 seconds to 10 minutes at the dry heath site, with peak temperature reaching 158oC, while at the raised bog site, average maximum temperature never exceeded 15oC. While they control for plot level variation in vegetation structure, they do not report a result. Sites were selected to have similar ground fuel cover, > 85% mature Calluna, and >63% cover of pleurocarpous mosses, but contrasting edaphic characteristics.
A further study found that Calluna cover influences temperature during burns and can explain damage to moss. From 16 experimental burns at Moor House in the North Pennines, Noble et al. (2019a) found that higher temperatures were associated with a greater proportion of cell damage in Sphagnum capillifolium. Maximum surface temperature during burns varied considerably from 33oC to 538oC and was higher in plots with greater Calluna cover. They found dwarf shrub cover to be the best predictor of maximum fire temperature. Further laboratory testing of five common species of Sphagnum indicated that all five species of Sphagnum experienced greater cell damage at higher temperatures.
The results of Davies et al. (2016a) suggest that variation in pre-fire fuel load and structure of Calluna influence fuel consumption during fires. Noble et al. (2019a) found that Calluna cover explained variation in maximum burn temperature and therefore damage to Sphagnum moss (a contextually relevant measure of severity). Grau-Andrés et al. (2018) controlled for variation in fuel structure in their study, yet did not report whether it was found to influence severity, finding that soil moisture plays a leading role in explaining variation in severity of burns across two measures of severity, consumption of the moss-litter layer and soil surface temperature response.
For further context, the conceptual model developed by Davies, Legg and Grau-Andrés identified the moss-litter layer as being key to understanding fire behaviour in this system. Where the moss-litter layer is dry it adds considerably to the available fuel load in the system, the effect of moisture being to control the total available fuel load.
Overall, the findings from four modelling studies suggest that Calluna fuel load and structure may influence fire severity, however as soil moisture conditions are also found to control fire severity the marginal effect (as distinct from other factors) of Calluna fuel load and structure across these studies is unclear.
A further limitation of this evidence base with respect to the research question is that measures of severity relate to the plot level and are centred on consumption of Calluna biomass and damage to moss. There is no clear link from this evidence base to wider measures of severity, based on area affected, economic damage or the level of resources required to be employed in firefighting.
5.6. Research Question 3: What is the Role of Muirburn in Controlling the Extent of Wildfires?
The search process identified no primary studies which have directly studied whether variation in fuel loads resulting from muirburn influence the extent of wildfire in moorland.
The review concludes that there is a lack of evidence that variation in fuel loads resulting from muirburn influences the extent of wildfire in moorland. This has been identified as a research gap.
5.7. Evidence from Previous Reviews and Reports
A series of previous reviews have sought to assess the effects of muirburn/moorland management practices, of these only Glaves et al. (2013) and Glaves et al. (2020) have specifically addressed a research question with respect to wildfire. Four further reviews are additionally introduced here, on the basis that they have highlighted further relevant evidence in the process of investigating muirburn impacts with respect to other outcomes (Table 5).
| Year and Authors | Title | Main Focus | Wildfire Research Question |
|---|---|---|---|
| Glaves et al. (2013) | The effect of managed burning on upland peatland biodiversity, carbon and water (NEER004) | Managed burning effects | “Is there a relationship between managed burning of upland peatlands and ‘wildfire’ (risk, hazard, occurrence, severity, extent and damage)?” |
| Werritty et al. (2015) | A Review of Sustainable Moorland Management. Report to the Scientific Advisory Committee of Scottish Natural Heritage | Moorland management | No research question stated with respect to wildfire |
| Chapman et al. (2017) | Muirburn, Peatland and Peat Soils – An Evidence Assessment of Impact | Effect of muirburn on carbon and peat forming vegetation | No research question stated with respect to wildfire |
| Brooker et al. (2018) | Socio- economic and biodiversity impacts of driven grouse moors in Scotland. | Assessment of moorland management practices | No research question stated with respect to wildfire |
| Baggaley et al. (2021) | Understanding carbon sequestration in upland habitats | Carbon stocks and the relationship between carbon and biodiversity | No research question stated with respect to wildfire |
| Glaves et al. (2020) | The causes and prevention of wildfire on heathlands and peatlands in England | Wildfire | 1. What are the main factors that contribute to the risk and occurrence of wildfire? 2. What are the main wildfire ignition sources? 3. What factors influence fire behaviour and severity? 4. What are the most effective measures for preventing wildfire? 5. What are the characteristics of effective firebreaks? 6. How effective is the Met Office Fire Severity Index in predicting potential fire severity? 7. How effective is ‘fire watching’ in preventing and reducing the impact of wildfire? 8. What are the most effective measures for reducing the negative impacts of wildfire? |
a) Glaves, D.J., Morecroft, M., Fitzgibbon, C., Leppitt, P., Owen, M. and Phillips, S. 2013. The effects of managed burning on upland peatland biodiversity, carbon and water. Natural England Evidence Review, NEER004. Peterborough: Natural England
The review found no studies that specifically provided evidence on the direct relationship between managed burning and occurrence and severity of wildfire in the UK.
They found moderate evidence (as assessed by Glaves et al., 2013) that fuel load and fuel structure are critical factors in fire behaviour, and that managed burning can be used to reduce fuel load and therefore fire hazard (citing; Albertson et al., 2009; 2010; Davies, 2005; Davies et al., 2010; Davies and Legg, 2011; Aylen et al., 2007; McMorrow et al., 2009; Davies et al., 2008 and Legg and Davies, 2009).
They found moderate evidence (as assessed by Glaves et al., 2013), citing McMorrow et al. (2009) that heather moorland in the Peak District which is mostly managed by rotational burning is less prone to the occurrence of wildfires than other moorland habitats.
They further found moderate evidence (as assessed by Glaves et al., 2013) that there may be an increased need for risk management in the future due to climate change, and that this is likely to include fuel management by cutting or burning and the creation of a network of fire breaks and fire control zones (citing; Albertson et al., 2009; 2010; Aylen et al., 2007; McMorrow et al., 2009; and Davies et al., 2008).
b) Werritty, A., Pakeman, R.J., Shedden, C., Smith, A. and Wilson, J.D. 2015. A Review of Sustainable Moorland Management. Report to the Scientific Advisory Committee of Scottish Natural Heritage
With respect to the relationship between muirburn and wildfire occurrence, Werritty et al. (2015, p.15) state that “evidence suggests that over 50% of wildfires with known causes may themselves be caused by loss of control of prescribed burns” (citing; Legg et al., 2006; Worrall et al., 2010b; and personal communication with the National Trust for Scotland).
Citing McMorrow et al. (2009) they note that Peak District grouse moor management is associated with lower frequency of wildfire, which has been taken to imply that the practice of prescribed burning reduces or at least is associated with reduced wildfire frequency in that region.
With respect to wildfire severity, Werritty et al. (2015, p.15) state that, “whilst large, intense fires can be destructive, many may have no greater impact than prescribed burns” (citing; Maltby et al., 1990 and Clay et al., 2010a).
Overall, Werritty et al. (2015, p.15) describe a contested evidence base, “the relationship between the use of prescribed fire and the frequency and extent of wildfires on moorland remains contested, and this is an area where the evidence base needs to be developed.”
c) Chapman, S., Hester, A., Irvine, J. and Pakeman, R. 2017. Muirburn, peatland and peat soils – an evidence assessment of impact. ClimateXChange Report
Chapman et al. (2017) note that wildfire results from a variety of causes, including accident, lightning strikes, arson and indeed muirburn which has gone out of control, and that it is usually prevalent in dry weather and in areas where there has been an accumulation of readily combustible plant material.
Citing Legg and Davies (2009) they note that wildfires will often burn hotter and can be more harmful than moorland properly managed by muirburn.
Chapman et al. (2017, p.3) further note that “it has been suggested that climate change will increase the risk of wildfire in areas where warmer and drier summers are predicted which will lower moisture levels, increasing the risk of ignition (Albertson et al., 2010)”.
d) Brooker, R., Hester, A., Newey, S. and Pakeman, R. 2018. Socio-economic and biodiversity impacts of driven grouse moors in Scotland: Part 2 Biodiversity impacts of driven grouse moors in Scotland. Report to the Scottish Government
In a discussion on muirburn impacts in context, the authors argue that fire intensity/severity is critical to understanding muirburn impacts. They note that very few fire studies have included detailed measurements of fire severity linked through to impacts, severely limiting the ability to link reported fire impacts to the actual characteristics of the fire causing those impacts and that there has been a strong reliance on modelling work to predict likely impacts from different types of fires.
In exception to this, Brooker et al. (2018) note the work of Davies, Legg and colleagues whose studies have identified key factors affecting fire intensity/severity; particularly (a) fuel load and (b) soil/vegetation moisture and weather conditions. They further note:
- There are strong links between severity of burn and fuel load (Davies et al., 2016a)
- Longer rotation burning generally leads to greater fuel loads (Milligan et al., 2018)
- Soil/vegetation moisture and weather conditions – these factors are critical for fire severity and correlate with time of year (Grau-Andrés et al., 2018).
e) Baggaley, N.J., Britton, A.J., Barnes, A., Buckingham, S., Holland, J.P., Lilly, A., Pakeman, R.J., Rees, R.M., Taylor, A. and Yeluripati, J. 2021. Understanding carbon sequestration in upland habitats. ClimateXChange Report
In a section titled “burning” in relation to dry heath, Baggaley et al. (2021) note that evidence for the effects of management burning on above and particularly below-ground carbon stocks in dry heathland is surprisingly limited.
- Studies of management burns on heathlands in the UK uplands suggest that <30-100% of the above-ground carbon stock may be lost during a management burn, while soil carbon stocks should not be impacted if the burn is properly managed (citing: Kayll et al., 1966; Farage et al., 2009; Legg et al., 2010; Worrall et al., 2013b).
- The size of the above-ground biomass carbon stock is positively related to time since fire, and reaches a maximum at around 20 years (citing, Alday et al., 2015).
- The amount of carbon lost during fires is impacted by variations in climate across the UK; carbon losses are greater, under warmer, drier conditions (citing; Davies et al., 2016a; Santana et al., 2016).
- Wildfires may be more intense than management burns and may remove both above-ground carbon stocks and a portion of the soil carbon store (citing; Maltby et al., 1990).
f) Glaves, D.J., Crowle, A.J.W., Bruemmer, C. and Lenaghan, S.A. 2020. The causes and prevention of wildfire on heathlands and peatlands in England. Natural England Evidence Review NEER014
The main conclusions from Glaves et al. (2020) regarding wildfire in general are:
- Wildfires occur across the country, on all main habitats but particularly (by area) on heathlands and peatlands.
- In the uplands, wildfires are most common during Spring, unless it has been a particularly dry year in which case the less common Summer wildfires may be more frequent.
- In the lowlands, there is greater frequency overall, while a Spring peak is still evident, wildfires are comparatively more common through Summer, Autumn and Winter.
- Data from the Fire and Rescue Service Incident Reporting System for eight years 2008-2016/2017, indicates that by area wildfires are most common in open semi-natural habitats, 59% of all wildfires, with 48% occurring on mountain heath and bog.
- The majority are due to human causes, either accidental or deliberate.
- Arson is more frequent in the lowlands
- The proportion of wildfire due to accident greater is in the uplands
- There is an association with recreation
- Escaped managed burns are also significant
- Risk and occurrence of wildfire is associated with:
- hot dry conditions, especially drought
- vegetation characteristics
- fuel load
- moisture
- human related characteristics
- public access
- managed burning
- arson
- Wildfire incidence is episodic, coinciding with dry spells, resulting in variation between years which makes determination of temporal trends difficult.
Evidence on wildfire occurrence attributable to managed burning
Glaves et al. (2020) note that while data limitations had previously been cited as limiting information on the frequency of managed burns initiating wildfires in the UK, recently more data has become available. Sources of data now include statistics on wildfire occurrence compiled nationally, regionally and locally by the Fire and Rescue Service, and for some National Parks and Areas of Outstanding Natural Beauty. They conclude that collectively this provides strong evidence that managed burns escaping control cause a proportion of wildfires, citing the following:
- Their own analysis of (2002–18) English wildfire data which includes 57 cases where managed burns escaped control were identified as the cause of ignition, comprising 15% of the 382 ignitions where a specific cause was identified.
- Luxmoore (2018), a Scotland-wide analysis of Fire and Rescue Service records which reported that 60% of ‘primary’ wildfires (140 out of a total of 233, 2009/10 to 2014/15) were “potentially caused by muirburn”, ranging between 48% and 67% per year and varying geographically by region.
- Worrall et al. (2010b), a study utilising data from ranger reports in the Peak District National Park spanning 1976 to 2004 found that 10 out of 41 (24%) of fires for which the cause was identified could be attributed to escaped managed burns, although they tended to be larger resulting in 51% of the area affected by wildfires being due to this cause.
- Legg et al. (2006), a study utilising data from a questionnaire administered to 41 Scottish Estates recorded 17 wildfires in total in 2003, of which nine had been caused by muirburn and two further by vegetation burning for livestock, yielding a total of 65% due to escaped managed fires.
- Martin (2018), a case study of Darwen and Turton Moors, recorded 22 “main wildfires” between 1995 and 2017, of which one related to grouse moor strip burning, and a further seven were considered likely to have been due to managed burns for livestock grazing getting out of control, yielding a total of 36% resulting from escaped managed burns.
- Luxmoore (2018), a case study of National Trust for Scotland properties covering an upland area of 63,316 ha where no managed burning takes place. Over a period of eighteen years, twelve large wildfires occurred affecting a total area of 1,463 ha or 2% of the upland area. Glaves et al. (2020) note that while a policy of not burning did not cause wildfires to cease, it did not result in widespread wildfire at this site either.
While noting the need for more complete data on escaped managed burns and other causes of wildfire, Glaves et al. (2020) suggest this gives a range for the proportion of wildfires resulting from escaped managed burns of between 15% and 60%, or 24% to 65% if data from the lowlands is excluded.
Evidence on managed burning reducing fuel load in the UK
Glaves et al. (2020) note that whilst monitoring and managing fuel load is often advocated in the UK, especially for upland heathland and peatland there appears to be limited evidence of its direct effect on wildfire incidence, behaviour, severity and extent. They surmise that this may in part be due to data limitations; the relatively short Incident Reporting System wildfire data set timescale; and limited availability of mapped data on managed/prescribed burning especially at a local scale. As this becomes available in the future it might make quantitative analysis of the relationship between burning and wildfires more practical in future.
Evidence from elsewhere in the world on managed burning reducing fuel load
Glaves et al. (2020) further considered international evidence relating to the effectiveness of managed burning in reducing wildfire hazard, noting that managing fuel load through prescribed burning is widely practiced elsewhere in the world, particularly in shrub and forest habitats in southern Europe, North America and Australia.
From this evidence base Glaves et al. (2020, p.70) note “strong, but in some specific respects contradictory, evidence that [managed burning] can be beneficial in reducing hazard and hence the incidence, intensity, severity and extent of wildfires, and in facilitating fire suppression efforts”.
In assessing the validity of this evidence base Glaves et al. (2020, p.70) note that evidence tends to be from modelling or theoretical studies, rather than empirical, and that within this literature there remain “considerable apparently unresolved questions over the effects of fuel load management, in particular in relation to the spatial arrangement, size, extent and type of fuel treatments [limiting] the conclusions that can be drawn from these approaches [and] highlighting the need for more, properly designed experiments addressing [the] question [of fuel treatment effects]”.
Summarising this international evidence base Glaves et al. (2020) found moderate evidence that the intensity and severity of wildfires is influenced by the following factors:
- The speed of subsequent fuel accumulation – rapid fuel accumulation can limit effectiveness to a relatively short post treatment period.
- The size, shape, pattern and degree of continuity of managed burns – simulations suggest that long linear, especially gridded treatment areas are more likely to be effective than patches.
- The proportion of area subjected to fuel reduction – higher proportions are likely to be more effective although this may result in as much or more of the area being treated than is expected to be spared from wildfire. Comparatively, the proportion of total area treated per annum has a greater influence than the treatment unit size.
Additionally, they note that the interaction of other factors notably fire weather and ignition prevention may be as or more important in controlling wildfire than fuel management approach and effort, and that the best results of managed burning are likely to be obtained in heterogenous landscapes and in climates where the likelihood of extreme weather conditions is low.
Commenting on the applicability of this evidence base to the UK, Glaves et al. (2020) note that such effects are likely to vary geographically and between sites and habitats. In applying this evidence base to the UK context they note that typical managed burning of UK moorland involves the creation of an extensive patchwork of small blocks (median 0.25-0.28 ha.) managed on 10-20 year rotations, whereas the described findings suggest that “such burning might not necessarily provide the most effective spatial pattern, frequency or approach specifically for reducing wildfire risk, occurrence and impact. Where this is an objective, a more strategic approach targeted at high risk locations such as access hotspots/routes, probably with more frequent and varied treatments, might be more effective and efficient, and potentially also result in a smaller total area being burnt” (Glaves et al. 2020, p.71).
5.8. Conclusions
5.8.1. What we know
- The review concludes that there is evidence that muirburn directly causes a proportion of wildfires that occur on moorland, however, there remains uncertainty regarding this proportion.
- The review concludes that there is evidence from field studies that the immediate effect of burning on Calluna moorland is to reduce above ground biomass.
- Studies suggest that fire intensity in heather is controlled by fuel structure, windspeed and fuel moisture content.
5.8.2. What we know we don’t know
- There is a lack of research into the question of whether variation in fuel loads resulting from muirburn influence the subsequent occurrence or likelihood of wildfire in moorland in the UK.
- This review found that the evidence base relating to the role of muirburn in controlling fire intensity and fire severity was too limited to draw firm conclusions.
- There was a lack of evidence relating to muirburn controlling the spatial extent of wildfire in moorland.
- There was a lack of research into the relationship between muirburn and wildfire on non-heather dominated habitats.
6. The Impact of Muirburn on Carbon Emissions and Storage
6.1. Summary of Main Evidence
- The primary literature search generated 117 publications. From this, carbon data relevant to Scottish systems experiencing muirburn was available from 17 papers.
- The quick scoping review showed there to be limited empirical data describing the effects of muirburn on carbon sequestration or greenhouse gas (GHG) emissions.
- The carbon cycle is complex and dependent upon biogeochemical processes as well as anthropogenic (management) and climatic factors resulting in significant variation spatially and temporally making full budget analyses difficult. The majority of studies available considered individual components of the carbon (or greenhouse gas) budget, with most data available for stream or soil water dissolved organic carbon (DOC) concentrations. Few studies have directly measured or calculated the effects of muirburn events on GHG emissions or carbon stocks (i.e. soil, water and gas carbon measurements) prior and post fire events. Therefore, comparing results from single site experiments with wider catchment areas could lead to contradictory interpretation or lack the evaluation of cumulative impacts occurring spatially and temporally.
- There is poor understanding of the interactions between burning and other disturbances (such as, grazing, drainage, and nutrient deposition) on carbon cycling and vegetation dynamics (Davies et al., 2016c).
- There are few complete carbon budgets from UK peatland sites subject to management burning (Davies et al., 2016c). In particular changes in soil carbon stocks do not seem to be reported often, in response to muirburn events. There is at present little evidence of how fire affects the emission of carbon dioxide (CO2) and methane (CH4) via perturbation to aerobic and anaerobic respiration (Gray et al., 2021), although Heinemeyer et al. (2019) reported that the burning of Calluna-dominated blanket bog, at a study site in Northern England, lead to a switch from a net C sink to a net C source.
- The reporting of key parameters would help in calculating/estimating carbon stocks and budgets, for example it is recommended to accurately record bulk density and total peat depth measurements as part of peat carbon stock assessments (Morton and Heinemeyer, 2019).
- There is still limited carbon budget data for dry heathlands or other moorland habitats with the majority of research being conducted on a few UK peatland sites.
- There are few long-term studies from which the impacts of managed burning on carbon budgets can be determined, with limited geographical coverage. Moor House in the North Pennines provides a unique insight into the long-term effects of rotational burning; however, this is only one site, and it is not clear how representative it is of other muirburn sites.
- There is limited evidence referring to optimal fire rotation intervals in relation to post-fire recovery and net carbon budgets.
6.2. Research Questions – Muirburn and Carbon
The research questions set up for this review were:
- What is the impact of muirburn on carbon functions (sequestration, GHG emissions and other loss of carbon) from soils and above ground vegetation?
- What is the timescale for recovery and resilience of carbon sequestration potential and GHG emissions following a muirburn event over immediate, short and long timescales on soils and habitats?
- What are the indicators to evaluate the dynamics of carbon losses and recovery following muirburn events?
6.3. Search for Primary Literature
A search for primary literature was conducted in the Web of Science database. Search terms and syntax are detailed in Table 6.
The search generated 117 results, of these 17 were included in the review as they contained carbon data relevant to Scottish systems experiencing muirburn.
Screening for relevance followed an iterative process. Sources were assessed first on the basis of their title, then abstract, then on a scan of their full text. At each stage, sources were excluded if it was clear that they did not include information relevant to the research questions. Sources were removed where the habitat of the study clearly differed to the moorland habitat as defined for the review, for instance those reporting results for land uses (e.g. forest systems) and climates (Mediterranean, California, Australia etc.) that are outside of the moorland habitat as defined for the review. In addition, papers that were not focused on prescribed burns/wildfires were also removed. A more rigorous interrogation of the papers was carried out to identify papers reporting on the effects of fire on carbon balance (i.e. soil carbon, vegetation carbon, greenhouse gases, carbon loss in leachates etc.). Papers were excluded if they did not include carbon data or discussion related to carbon.
| Search terms | heath* OR moor* OR bog OR peat* OR heather OR Calluna OR Molinia OR mire OR fen* OR flush* AND "prescribed burn*" OR muirburn or "manag* burn*" OR "manag* fire" OR "control* burn*" OR "rotation* burn*" OR "pastoral fire" OR "fuel reduction" OR swaling AND soil OR carbon |
|---|---|
| Tool: | Web of Science |
| No. hits | 117 |
| Initial screening | Removed papers reporting results for land uses (E.g. forest systems) and climates (Mediterranean, Arctic, California, Australia etc) that are not relevant or Scottish/UK moorlands. In addition, papers that were not relevant i.e were not focused on prescribed burns/wildfires were also removed. |
| Second Screening | A more rigorous interrogation of the papers to identify papers reporting on the effects of fire on carbon balance (i.e. soil carbon, vegetation carbon, greenhouse gases, C loss in leachates etc). Papers were excluded if they did not include carbon data or discussion, were not relevant to UK soils or climates (e.g. Mediterranean, tropical etc.) or were not relevant to muirburn (e.g. forest systems). Sources removed = 100 |
| Sources included in the review | Sources included = 17 |
Information on the 17 papers that include empirical evidence relating to carbon is given in Annex 7. An assessment of each of the paper’s robustness and relevance to the carbon research questions was carried out (see Annex 8); using a set of criteria (see Annex 9).
6.4. Research Question 1: What is the Impact of Muirburn on Carbon Functions (Sequestration, GHG Emissions and Other Loss of Carbon) from Soils and Above Ground Vegetation?
In approaching this question, the review team considered:
- What is the evidence of the impact of muirburn on gaseous carbon losses?
- What is the evidence of the impact of muirburn on aqueous carbon losses?
- What effects does muirburn have on vegetation cover and potential soil/peat erosion?
- What is the evidence of the impact of muirburn on carbon storage and sequestration?
Evidence from both primary literature and previous reviews/reports are discussed together in the sections below.
6.4.1. What is the evidence of the impact of muirburn on gaseous carbon losses?
During the process of combustion carbon is released to the atmosphere in the form of various gases and particulates with most of the carbon in the form of CO2 (Clay and Worrall, 2011). The activity of rotational heather burning can affect gaseous pathways because alterations in the vegetation community alter the gross carbon dioxide (CO2) fluxes of respiration and photosynthesis, as well as reducing above-ground carbon stocks (Ramchunder et al., 2009, Ward et al., 2007). Prescribed burning on peatland vegetation alters the soil thermal regime (Brown et al., 2015), which may therefore affect biologically regulated processes such as organic matter decomposition and respiration rates. However, Morton and Heinemeyer (2019) outlined surface peat carbon losses, apparently resulting from enhanced decomposition under rising temperatures, might also be explained by apparent changes due to peat shrinkage and expansion, demonstrating the complexity in temperature and moisture driving gaseous losses.
In a fire severity manipulation experiment on two Scottish heathland sites, Grau-Andrés et al. (2019) showed burning increased CH4 flux at a raised bog and Ward et al. (2007) found rotational burning and grazing (at Moor House in the North Pennines of Northern England) had significant effects on net and gross CO2 fluxes, with the greatest effects observed in the burning treatment where rates of respiration and photosynthesis were greater relative to unburned treatments.
Rotational burning reduced net CH4 fluxes on 12 out of the 15 sampling dates at Moor House (North Pennines) with a mean reduction of 12% relative to unburned plots with a significant interaction between burning and grazing with the lowest CH4 fluxes occurring in the no burn, ungrazed plots (Ward et al., 2007). Chen et al. (2008) showed soils from both frequent burning treatments and unburned controls (at Moor House) had no clear differences in methane oxidation. This indicates that burning potentially causes a relatively small impact on CH4 compared to variability of vegetation biomass and spatial heterogeneity.
Ward et al. (2012) compared 10-year interval burned plots (last burned in winter 2007, sampled 18 months after burning) with areas unburned since 1954 (at Moor House) and found no significant differences in gross and net CO2 fluxes due to either burning or grazing treatments and no significant differences in CH4 flux due to burning within sites. There was, however, a trend for higher CH4 flux in areas that had been burned relative to unburned areas (Ward et al., 2012). Grau-Andrés et al. (2019) claimed that methane emission fluxes have also been observed to decline one year after wildfires.
6.4.2. What is the evidence of the impact of muirburn on aqueous carbon losses?
Studies suggest dissolved organic carbon leaching may increase following prescribed burning and then decrease over the initial few years after the burn as the vegetation canopy becomes more established and bryophytes re-appear (Holden et al., 2013). Glaves et al. (2013) outlines that there is strong evidence that managed burning leads to increased water colouration and/or dissolved organic carbon in peatland watercourses. However, Glaves et al. (2013) also noted that the relatively small number of small plot- or stand-scale studies of soil water colouration and/or DOC in relation to burning have shown inconsistent evidence, which may reflect differences in time since burning (as effects have been shown to be greatest soon after burning) and sampling too deep in the peat (as effects have been shown to tend to occur in the upper layer).
Davies et al. (2016c) highlights that increased colouration of water from DOC occurs in areas without moorland burning (as DOC is strongly associated with the dominance of Calluna rather than burning per se), and Jaffé et al. (2013) also questioned whether increased DOC transport offsite leads to net carbon loss or simply serves as a conveyer for some of it to be accumulated elsewhere. Davies et al. (2016c) also suggests prescribed burning is associated with changes in DOC quality and associated water colouration, rather than the quantity of DOC.
Despite alterations to soil, Ramchunder et al. (2009) states that Ward et al. (2007), Worrall et al. (2007) and Clay et al. (2009) did not find any relationship between DOC release and burning. Clay et al. (2009) showed burning does not significantly affect DOC concentration in either soil water or runoff with peaks in DOC concentration and water colour seen in the weeks following the managed burn but that these effects are short-lived and no significant effect was observed between the year before and year after burning at Moor House.
In another study at Moor House, in the North Pennines, Worrall et al. (2007) found that the DOC content showed no significant difference between grazing treatments but showed a significant decrease with the presence of burning, though no direct relationship with the depth to water table could be found. A potential explanation for differences in water colouration is that there may be a relatively continuous effect of burns within a rotationally burnt catchment (when a proportion is burnt most years) compared to plots that are burnt less frequently with moderate to long intervals where any effect is perhaps more temporary. In addition, Worrall et al. (2007) showed that, statistically, burn management explains only a small proportion of the variance in the composition of the DOC, with the variation being dominated by the differences between days of sampling and seasonal variation. It is worth noting that DOC fluxes are naturally variable due to soil heterogeneity and therefore not evenly distributed across a moorland and so when attempting to identify burn impacts, there is a need to consider variations in burn severity, above ground vegetation, distance from watercourses and landscape topography.
Worrall et al. (2013a) showed DOC concentrations of surface runoff water at sites in the Peak District were not significantly different (P< 0.05) between any of the managed burn treatments and the control. However, Worrall et al. (2013a) did find that DOC concentration in soil water significantly (P< 0.05) decreased with both burning and cutting, but these differences could be explained by differences in water table and changes in flowpath through the soil profile. The study by Worrall et al. (2013a) suggests that declines in soil water DOC concentration are brought about as different levels in the peat profile become the dominant source of water due to changes in the depth to water table, with these changes in the depth to water table being brought about by changes in evapotranspiration resulting from a loss of vegetation. In a fire severity manipulation experiment on a Calluna raised bog by Grau-Andrés et al. (2019) they showed that burning had no significant effect on soil water DOC. However, it is worth noting that a lack of effect seen on DOC fluxes may relate to sampling depth as Glaves et al. (2013) outline that the effects on DOC are likely to occur in the upper layers of peat and so when reporting outputs for DOC effects following burns it is important to ensure that water table fluctuations are accounted for.
In a study by Clay et al. (2012) results showed that there was an elevated water colour in the few years immediately following prescribed burning but that this was not matched by a rise in DOC concentration. Clay et al. (2012) therefore propose that burning appears to affect the composition of the DOC rather than the absolute DOC concentration, and therefore highlights that the use of water colour as a proxy for DOC concentration should be treated with caution. Furthermore, as has been noted elsewhere there is a disconnect between the direction and magnitude of DOC changes between some plot-scale studies and catchment-level monitoring (Davies et al., 2016c).
6.4.3. What effects does muirburn have on vegetation cover and potential soil/peat erosion?
As outlined by Ramchunder et al. (2009), peat as a material has very low density and so it can be eroded very easily by the removal from bare peat surfaces through the action of running water, wind and chemical oxidation. A loss in vegetation through burning or by other means, exposes the soil to erosion and desiccation (making it vulnerable to shrinkage). In an assessment of stream DOC in the North York Moors, Yallop and Clutterbuck (2009) found the proportion of exposed peat surface resulting from new heather burning was consistently identified as the most significant predictor of variation in DOC concentration (estimated from stream water colour). Worral et al. (2007) showed carbon loss from eroding peat within a rotational heather burning system occurred primarily through increased gaseous fluxes (CO2 and CH4), particulate and dissolved organic carbon losses to watercourses and excess dissolved CO2 pathways. Alday et al. (2015) suggest that any carbon losses could be minimized by using “cool burn” or “pressurized fuel-assisted” burning as some vegetation remains after the fire and so the peat should be left relatively unaffected. Alday et al. (2015) state that the overall carbon balance within moorland systems will depend on the biomass consumed by a fire and the time it takes for the ecosystem to recover via plant growth and biomass accumulation during the inter-fire interval. Blundell and Holden (2014) implied that the vegetation type influences burn effects and so it is important to consider vegetation type, as woody material can potentially provide a source for black carbon. Due to the high water content of moss, it may be more resistant to the effects of fire compared to other vegetation types, however the water content of mosses is highly variable.
Therefore, restoration of peatland plant community composition could be essential to the post-fire recovery of peatlands, with an overall aim of the preservation of peatland carbon stores (Shepherd et al., 2021). It is also suggested that after burning there is often relatively rapid vegetation recovery and hence carbon accumulation during the post-fire succession (Alday et al., 2015).
6.4.4. What is the evidence of the impact of muirburn on carbon storage and sequestration?
Glaves et al. (2013) report that there is moderate evidence suggesting that managed burning results in reduced peat accumulation; reduced above and below ground carbon storage compared to no burning; erosion and reduction in the level of the soil surface; increases in gross CO2 fluxes of respiration and photosynthesis; and carbon losses through fuel consumption during burning and in conversion to char. Worrall et al. (2010a) carried out a meta-analysis of existing managed burn data and showed that there was only a 7% probability of improving the carbon budget (i.e. retaining C inputs and minimising system-level C losses) and a 40% probability of improving the greenhouse gas budget by introducing prescribed burning onto a peatland. Davies et al. (2016c) noted that rates of peat accumulation are lower in areas burnt by management fires, suggesting that in terms of carbon sequestration burning may not be beneficial to the system carbon budget.
A key characteristic of peatlands is a high water table, which leads to the slow decomposition of organic matter leading to its accumulation (and carbon within the organic matter complexes). As discussed, increased temperatures and/or exposed peat (due to a loss of vegetation cover) can leave the peat vulnerable to desiccation and shrinkage. Such a change in soil structure may be important for hydrology, water quality and biota in peatlands. If such changes result in changes to water flowpaths, by, for example, creating connected macropore channels for water flow this may increase infiltration rates and saturated hydraulic conductivity (Holden et al., 2013).
Holden et al. (2013) found that rotational heather burning leads to changes in upper layer water movement in blanket peat, which reduces the role of macropores and increases the role of micropores. This may increase the potential for leaching of dissolved organic carbon from the peat system because there would be increased contact time between infiltrating water and the peat matrix.
Water table depth is very important for maintaining the stability and function of peatlands as carbon stores (Brown et al., 2014). Brown et al. (2014) found water table depth to be significantly deeper for burned catchments than for unburned ones, which may reduce C sequestration potential and C storage capacity.
However, in a study based on a long-term experiment (at Moor House in the North Pennines), Worrall et al. (2007) showed that the depth to water table in soil differed between different heather burning rotations and grazing intensities, with the greatest depth to water table on plots where burning did not occur or for longer burning cycles where livestock had been excluded. Worrall et al (2007) found burning on 20-year and 10-year cycles decreased the depth to water table by 8% and 26% respectively (post hoc analysis on normalised data for grazed and ungrazed plots). Worrall et al. (2013a) found the depth to water table decreased at burnt and cut sites relative to controls in line with a change in evapotranspiration due to loss of vegetation.
Brown et al. (2014) showed that prescribed burns on UK moorlands had clear effects on peat hydrology, peat chemistry and physical properties with a reduction in organic matter content of the upper peat layers. Brown et al. (2015) state that prescribed peatland vegetation burning alters soil thermal regimes (increases soil temperature), which can impact soil respiration rates and carbon loss. A reduced capacity for a peatland to hold and maintain water will affect leached carbon losses, peatland exposure to decomposition, soil mineralisation rates and therefore the overall soil carbon storage capacity.
Rosenburgh et al. (2013) showed that the soil carbon to nitrogen ratio (C:N) of prescribed burn sites (in the North Peak) declined since burning, with the lowest C:N ratio in the oldest stands, which are likely to be the most N saturated and produce the greatest leaching losses of N.
Clay et al. (2010c) showed that managed burn sites were a mean source of carbon at a rate of approximately 117.8 gCm-2year-1 compared to unburnt sites with a mean source of 156.7 gCm-2year-1 (during the study period 2006-2007 at Moor House in the North Pennines), which if extrapolated then the catchment would be a net source of carbon of between 62 and 206 gCm-2year-1 over a 3-year period. However, after 30 years at the same site, Garnett et al. (2000) found that there was significantly less carbon stored in the blanket peat in plots which had been burned every ten years compared to plots that had been unburnt since 1954. Carbon budgets for prescribed burn plots in Northumberland were all sources of carbon and ranged from near neutral carbon balance (3-yr-old plots: 4 gCm-2yr-1) to large net sources (8-yr-old plots: 269 gCm-2yr-1) suggesting that burning of Calluna-dominated landscapes leads to an ‘avoided loss’ of carbon, as shown by Clay et al. (2015).
In a fire severity manipulation experiment on two heathland sites in Scotland, Grau-Andrés et al. (2019) showed that the response of carbon fluxes to increased fire severity in drought plots was similar to plots burnt under ambient conditions associated with traditional managed burning. Grau-Andrés et al. (2019) showed that averaged across all burnt plots, burning altered mean net ecosystem exchange from a net carbon sink in the heathland (−0.33 μmol CO2 m-2s-1 in unburnt plots) to a carbon source (0.50 μmol CO2 m-2s-1 in burnt plots) and at the raised bog (−0.38 and 0.16 μmol CO2 m-2s-1, respectively).
Ward et al. (2007) found that neither burning nor grazing affected total ecosystem carbon storage when sampled to a depth of 1m due to the greatest stocks (over 99%) being contained within the soil ‘O‘ horizon, which was unaffected by either burning or grazing. Ward et al. (2007) showed differences in carbon stocks (between burned and unburned sites) in the aboveground vegetation and upper peat horizons only, with the greatest change in carbon seen as a result of burning, with a 56% reduction relative to the unburned control.
Worrall et al. (2013b) showed pre-burn and post-burn above-ground carbon stocks were 445 ± 141 gCm-2 and 97 ± 35 gCm-2 respectively (it was noted that material collected post-fire contained charred vegetation material with a substantial input of unburnt litter material when sampling). This suggests that the biomass consumed by a fire as well as time for post-fire recovery during fire interval could significantly impact C inputs and subsequent C balances as outlined by Alday et al. (2015).
In addition, some studies note that muirburn can stimulate vegetation regrowth and subsequent carbon input to the system. In terms of C budgets, this may also increase C losses, if steady state is assumed, or increase the fuel load of subsequent fires. Burning may also change composition, structure and hence function especially of peatland habitats. However, as noted previously there is a lack of knowledge on whether this would be similar for dry heath or other habitats. Burning biomass of any kind will inevitably release above-ground carbon to the atmosphere through combustion, but may also transform some to a more recalcitrant form, i.e. charcoal (Allen et al., 2016). Black carbon produced from burns can contribute to the carbon sink due to its long mean residence time, often on the millennial time scale and its high degree of resistance to chemical agents i.e. black carbon may have the potential to remove significant amounts of carbon from the short term bio-atmospheric system and transfer it to the longer geological carbon cycle (Clay and Worrall, 2011). Production of refractory black carbon could be an important addition to carbon accumulation. Worrall et al. (2103b) showed that the production of char during some prescribed fires can lead to greater carbon storage than if no char was produced. This occurs even when allowing for the greater biomass loss in order to achieve a greater proportion of char in the burn products (Worrall et al., 2013b).
6.4.5. Overview on carbon functions
Overall there is limited empirical data available in the literature to characterise carbon functions. In addition, where data exists most studies have considered individual components of the carbon (or greenhouse gas) budget of a peatland under prescribed burning, especially dissolved organic carbon as outlined by Clay et al. (2015). The carbon cycle is complex and dependent upon biogeochemical processes as well as anthropogenic (management) and climatic factors resulting in significant variation spatially and temporally making full budget analyses difficult. Alday et al. (2015) state that the overall carbon balance within moorland systems will depend on the biomass consumed by a fire and the time it takes for the ecosystem to recover via plant growth and biomass accumulation during the inter-fire interval.
6.5. Research Question 2: What is the Timescale for Recovery and Resilience of Carbon Sequestration Potential and GHG Emissions Following a Muirburn Event Over Immediate, Short and Long Timescales on Soils and Habitats?
A key factor to consider when quantifying C budgets in systems where burning occurs is the frequency, intensity and duration of burn events as these will determine the time for recovery and restoration (this may vary depending on the severity of the burn, soil conditions and growth rate of vegetation). Prescribed burning on peatlands usually takes place on rotations of 15 to >25 years (Ashby and Heinemeyer, 2021), but few studies of the impact of prescribed burning on carbon fluxes have long-term records; one of the longest records available only considered 33 months of data – 21 months prior to a burn and 12 months afterwards (Clay et al., 2010c).
There is limited evidence in relation to the recovery and resilience of moorland carbon, however studies suggest that any impacts seen from the action of burns may be recovered over decadal time periods. For example, Allen et al. (2016) suggested that rotations in the range 8–18 years (12.5–5.5% burned annually) would minimise carbon loss from above-ground vegetation. Clay et al. (2009) suggest that longer burning rotations may be beneficial in order to reduce water colour in upland peat, but that burning in itself does not lead to dramatic increases in DOC in soil water or runoff water.
Holden et al. (2013) suggest fire influences the near-surface hydrological functioning of peatlands but that recovery in terms of saturated hydraulic conductivity and macropore flow may be possible within two decades if there are no further fires. Based on evidence gathered at Moor House (North Pennines), Clay et al. (2010c) suggest that if total combustion of vegetation occurs, provided burning occurs at cycles longer than 32 years, then less carbon is predicted to be lost than in a no‐burn scenario. However, for the same site, Garnett et al. (2000) showed that after 30 years there was significantly less carbon stored in the blanket peat in plots which had been burned every ten years compared to unburnt plots which indicates that recovery (in terms of carbon stores) may not always occur.
In a study focusing on wildfires, Grau-Andrés et al. (2019) reported that post-fire effects such as reduced primary productivity (and decreased aboveground carbon source) may be transient with reports showing peatland carbon sequestration to be greater than carbon loss after fire (13 years in the study highlighted), as ground vegetation regenerated. If poorly managed, prescribed burns may periodically run out of control, causing damage to deep peat: recovery from deep burns takes longer, and if the seedbank is destroyed is also dependent on adequate seed dispersal (Sanderson et al., 2020).
6.6. Research Question 3: What are the Indicators to Evaluate the Dynamics of Carbon Losses and Recovery Following Muirburn Events?
In order to determine whether there is a net loss, gain or no difference in the overall carbon budget, measurements of carbon input, carbon stored and carbon outputs are needed. In addition, to establish the effect of muirburn on a carbon budget, then budget estimates should be made pre- and post- fire events. Attempts have been made to estimate complete carbon budgets that consider the overall impacts of burning (Glaves et al., 2013). So far, these have produced inconsistent evidence, with predictions of both positive and negative overall effects of burning, although the estimates provide strong evidence that burning affects the processes controlling carbon budgets of upland peatlands (Glaves et al., 2013).
The development of an effective monitoring approach will likely need to address a range of issues including:
- Complexity of natural systems
When using existing datasets and outputs from studies to assess the direction of change and causality between muirburn and carbon balance, it is important to consider that there are quite often confounding effects from multiple management practices (such as grazing) occurring at a given site. In addition to management, there are other factors influencing carbon fluxes in these systems such as naturally occurring spatial heterogeneity and seasonal variations in carbon fluxes (particularly DOC) due to moisture and temperature fluctuations.
- Influence of historical management on baseline data
There is limited information reported on the historical management and condition of field sites used in these studies, which is particularly important in terms of estimating the recovery and resilience to any impacts of burn events. For example, Albertson et al. (2010) highlight that Moor House in the North Pennines, is less degraded/modified than a field site in the Peak District, where carbon loss is potentially more severe. A baseline carbon budget is also needed to evaluate long-term effects of burning. Although unburnt sites are sometimes used as controls, there may still be significant differences in the initial carbon stocks due to differences in historical land use and condition.
- Change in methodology and reporting
Another consideration is the range of methodologies used to estimate or measure the different components of the carbon cycle in these systems. As outlined by Yallop and Clutterbuck (2009) numerous previous studies have used water colour as a proxy for DOC concentration, however Clay et al. (2012) highlight that the use of water colour as a proxy for DOC concentration should be treated with caution as it is proposed that burning appears to affect the composition of the DOC rather than just the absolute DOC concentration.
To fully capture changes in soil carbon stocks, measurements or estimates of soil carbon concentration, soil depth/dating and bulk density are needed. It is worth noting that studies discussing changes of carbon may use different approaches to express carbon concentration and flux based on analytical results; for example using standard conversion rates such as assuming the carbon concentration to be 50% of the soil dry mass or using specific measurements or estimates of peat depth and bulk density. However, it is more difficult to estimate peat depth and bulk densities and so Morton and Heinemeyer (2019) recommend accurately recording bulk density and total peat depth measurements as part of peat carbon stock assessments to more accurately determine losses and gains.
There does not appear to be a common consensus over what is the most appropriate indicator to represent effects of burning on the carbon balance, as there are limited studies reporting muirburn effects on carbon stocks and GHG emissions. In order to investigate the temporal changes in DOC production it would ideally require monitoring sites across the duration of a burn cycle. However, given that these typically range from 8 to 25 years, using a linear approach to monitoring would pose significant financial and logistical problems (Clay et al., 2012).
6.7. Conclusions
There is poor understanding of the interactions between burning and other disturbances, such as fire, grazing, drainage, and nutrient deposition, on carbon cycling and vegetation dynamics (Davies et al., 2016c). Moorland systems are valuable landscapes in terms of soil carbon stores, and peatlands in particular are unique in relation to their complex hydrology and carbon storage capacity. However, there is limited data describing changes in carbon budgets due to the effects of muirburn, which is essential for the conservation of these sensitive systems.
6.7.1. What we know
- There is evidence of burning of above-ground biomass on peat during muirburn with potential impact on carbon sequestration.*
- The data available appears to be focused on dissolved organic carbon (DOC) fluxes and aboveground (vegetation) carbon, with information on greenhouse gas emissions and soil carbon stocks being more limited.
6.7.2. What we know we don’t know
- The review concludes that there is limited empirical evidence describing the effects of muirburn on carbon fluxes including greenhouse gas (GHG) emissions from UK moorlands.
- There is no overall consensus as to the net impacts of muirburn on carbon budgets, with evidence supporting gains, losses and no difference in carbon stores/fluxes following muirburn.
- The majority of data available is for stream or soil water dissolved organic carbon (DOC) concentrations. Few studies directly measured or calculated the effects of muirburn events on greenhouse gas emissions or carbon stocks prior and post fire events.
- There are few complete carbon budgets from UK moorland sites subject to managed burning. In particular, there is a lack of information on soil carbon stocks.
- There are few long-term studies from which the impacts of managed burning on carbon budgets can be determined, with limited geographical coverage. Those studies that do exist do not cover the range of habitats and habitat condition, and often have to make assumptions where data is lacking.
- There is little or no carbon budget data for dry heathlands or other important moorland habitats.
* This amended line replaces the previous line in the report to clarify the emphasis on impact. (There is some evidence of significant burning of soil surface peat biomass during muirburn rotation with likely impact on carbon sequestration.)
7. The Impact of Muirburn on Habitats and Species
7.1. Summary of Main Evidence
- The primary literature search generated 312 results, of these 31 were included in the review.
- The impact and role of muirburn on moorland habitats and species is complex, and there is often a lack of consensus and a contested evidence base in the literature. Factors including burn rotation length, fire severity and intensity, the presence of other factors such as management (e.g. grazing, drainage or predator control), the habitat type, geographical location, climatic conditions and scale of muirburn management all have an effect. There is limited definitive evidence of positive or negative impacts, although defining what are considered positive and negative impacts is often difficult. Some species have been found to benefit from muirburn while others do not, and whether this is positive or negative will depend on the perceived value of the species concerned (e.g. its rarity, or contribution to favourable condition of the habitat or its provision of ecosystem services) and the management objectives of the site.
- The evidence from the primary literature indicates that moorland management (which includes managed burning) affects the abundance and diversity of bird species (4 studies – Smith et al., 2001; Tharme et al., 2001; Hancock et al., 2011; Robertson et al., 2017). Some species benefit from moorland management while others do not.
- The evidence from the primary literature indicates that burning increased carabid species richness at a costal heath in Norway (1 study – Bargmann et al., 2015; 2016). One study found that Lepidoptera species rapidly recolonised dwarf shrub heath after burning (Haysom and Coulson, 1998). Burning did not result in substantial declines in the abundance of Orthoptera on four German peat bogs (1 study – Hochkirch and Adorf, 2007). The burning of peatland was found to have affected the macroinvertebrate assemblage of moorland catchment streams (2 studies – Johnston and Robson, 2018; Ramchunder et al., 2013). Burning was found to have an impact on the soil macrofauna, with earthworms initially predominating after burning followed by an increase in enchytraeid worms, but both diminish over time (1 study – Mallik and FitzPatrick, 1996).
- There is mixed evidence as to the response of Sphagnum to prescribed burning. Evidence from Moor House, in the North Pennines (Lee et al., 2013a; Milligan et al., 2018) and two other sites (Whitehead and Baines, 2018; Whitehead et al., 2021) indicate that short-burn rotations lead to an increase in Sphagnum. However, Noble et al. (2018a) also from Moor House found that the long-unburned reference plots had more Sphagnum than the ‘intermediate’ treatments and were similar to the short rotation plots. Noble et al. (2018b) found that burned sites had less Sphagnum cover on a national scale than unburned sites.
- The evidence from the primary literature indicates that hotter fires are likely to have a greater impact on Sphagnum growth and survival (2 studies – Taylor et al., 2017; Noble et al., 2019a). One study suggests that prescribed burning has only a limited and transitory effect on the abundance of Sphagnum capillifolium (Grau-Andrés et al., 2017), and another study suggests that Sphagnum capillifolium has the ability to recover following exposure to temperatures that are likely to occur in prescribed burns (Taylor et al., 2017), however the study by Noble et al. (2018a) suggests that burning can have a negative impact on Sphagnum and this can persist for several decades.
- The evidence from the primary literature indicates that burning results in a change in plant species composition and biomass. Blanket-bog vegetation responds to managed burning in a complex manner. There is consistent evidence that Calluna increases following burning (Lee et al., 2013a; Milligan et al., 2018; Alday et al., 2015; Whitehead et al., 2018; 2021; Noble et al., 2018b). Evidence from Moor House, in the North Pennines, indicates that short-burn rotations lead to an increase in Eriophorum vaginatum (Lee et al., 2013a; Milligan et al., 2018), however in a UK wide study by Noble et al. (2018b) plots burned 2-10 years previously had less Eriophorum vaginatum cover than unburned sites.
- There is little evidence on the impact of prescribed burning on moorland reptiles (one study, not peer reviewed, by Newey et al., 2020, found small and inconsistent effects of burning on adders), and no evidence was found on the impact of prescribed burning on moorland amphibians, small mammals or gastropods.
7.2. Research Question – Muirburn and Biodiversity
The research question set up for this review was:
a) What is the current understanding of the impact of muirburn on habitats and species?
Subsequently this research question has been considered as five separate sub-questions:
- What is the impact of muirburn on vegetation?
- What is the impact of muirburn on birds?
- What is the impact of muirburn on small mammals, reptiles and amphibians?
- What is the impact of muirburn on invertebrates?
- What is the impact of muirburn on soil microbes?
7.3. Search for Primary Literature
A search for primary literature was conducted in the Web of Science database. Search terms and syntax are detailed in Table 7.
The search generated 312 results, of these 31 were included in the review and 281 were excluded on the basis that they did not provide relevant evidence to the review question or were not original studies.
Screening for relevance followed an iterative process. Sources were assessed first on the basis of their title, then abstract, then on a scan of their full text. At each stage, sources were excluded if it was clear that they did not include information relevant to the research question. Sources were removed where the habitat of the study clearly differed to the moorland habitat as defined for the review, for instance those reporting results for land uses (e.g. forest systems) and climates (Mediterranean, California, Australia etc.) that are outside of the moorland habitat as defined for the review.
| Search terms | heath* OR moor* OR bog OR peat* OR heather OR calluna OR molinia OR mire OR fen* OR flush* AND "prescribed burn*" OR muirburn or "manag* burn*" OR "manag* fire" OR "control* burn*" OR "rotation* burn*" OR "pastoral fire" OR "fuel reduction" OR swaling AND habitat OR biodiversity OR conservation OR "species diversity" OR ecosystem* OR flor* OR fauna OR animal* OR bird* OR insect* OR invertebrate* OR communit* OR species OR composition OR frequency OR abundance OR diversity OR structure OR NVC OR cover OR richness OR sphagnum moss* OR sphagn* OR Erica tetralix OR Eriophor* OR Molinia OR Calluna OR Trichophorum OR population* OR “breeding success” OR restor* OR revegetat* OR “vegetation management” OR remediation OR regenerat* OR livestock OR stocking OR grazing OR designated site* OR SSSI OR “site* of special scientific interest” OR water catchment* OR “special area* of conservation” OR SAC* OR “special protection area*” OR SPA |
|---|---|
| Tool: | Web of Science |
| No. hits | 312 |
| Sources excluded from the review | Sources excluded on initial screening = 243 Sources excluded on second screening = 36 |
| Sources included | Sources included = 33 |
In addition to the 31 original studies (see Annex 10); there were 6 other papers that were considered relevant but were not reviewed. These included three reviews (Davies et al., 2016b; Harper et al., 2018; Reed et al., 2009), two critiques of other papers or reports (Ashby and Heinemeyer, 2020; 2021), and one rebuttal paper (Brown and Holden, 2020). Most of the original papers were from blanket bog (intact and modified) or Calluna dominated wet or dry heath (see Annex 10). There were no papers identified that studied the impact of prescribed burning on alpine heath communities, grasslands, flushes, tall herb communities or montane scrub communities.
Twenty-one papers focussed on the impact of prescribed burning on vegetation, four on birds and eight on invertebrates (see Annex 10).
Twenty-seven papers were from studies in the UK (mainly from England) (see Annex 10). Eight of the papers used data from Moor House in the North Pennines.
Thirty papers reported on field studies, and two on glasshouse experiments (see Annex 10).
An assessment of each of the paper’s robustness and relevance to the habitat and species research question was carried out (see Annex 11); using a set of criteria (see Annex 12).
7.4. Research Question 1: What is the Impact of Muirburn on Vegetation?
7.4.1. What is the impact of muirburn on vegetation composition, above-ground biomass and vegetation height?
The search process identified eleven primary sources that looked at the impact of muirburn on vegetation composition/cover; one paper that looked at the impact of muirburn on above-ground biomass; and one paper that looked at both vegetation cover and above-ground biomass. The search process identified nine primary sources that looked at the impact of burning on Sphagnum, and two that looked at the impact on propagules.
7.4.1.1. Moor House (Hard Hill) long-term, replicated plot experiment
Eight of the papers used data collected from the Moor House (Hard Hill) long-term burning and grazing replicated plot experiment. The Hard Hill experiment is located on an area of Calluna vulgaris-Eriophorum vaginatum blanket mire (National Vegetation Classification type M19b (Rodwell, 1991)) at Moor House National Nature Reserve in the North Pennines of Northern England (Ward et al., 2007). The study site extends across an area of 1km2 at an altitude of 590 to 630m and consists of four replicated treatment blocks that have been subjected to a range of burning and grazing treatments since 1954 (Ward et al., 2007). Each block is composed of six 10 m by 30 m treatment plots. Within each block, the plots were subjected to different burn cycles: (a) a short-rotation burn cycle (approximately every 10 years), (b) a long-rotation burn cycle (approximately every 20 years), and (c) unburned since 1954 (Ward et al., 2007; Milligan et al., 2018). Half of the treatment plots were fenced off to prevent sheep grazing, while the other half were open to sheep grazing (Ward et al., 2007). In addition to the formal experimental plots each block had an associated unburned (and unfenced) reference plot that was considered to have been unburned since at least the mid-1920s (Lee et al., 2013a). There was no pre-treatment vegetation assessment, and the reference plots (which were outside the formal experimental design) were only sampled occasionally, and hence not all the papers include data from the reference plots (Lee et al., 2013). Moor House is not typical of many managed moorland sites and the results from the Hard Hill experimental plots are probably only applicable to less-modified, high altitude blanket bog with low levels of grazing.
In a study carried out at the Moor House replicated long-term plots, nine years after burning (in the 10-year burn treatment), Ward et al. (2007) measured vegetation community composition, soil and vegetation carbon stocks and fluxes of CO2, CH4 and DOC. They found that both burning and grazing reduced aboveground carbon stocks, and that burning reduced carbon stocks in the surface peat. They also found that burning and grazing strongly affected the vegetation composition, causing an increase in graminoids and a decrease in dwarf shrubs and bryophytes compared to the unburned and un-grazed controls. Soil microbial properties showed minor responses to burning, with the C:N ratio of the microbial biomass higher in the burnt plots relative to the unburnt plots. There were increases in the CO2 fluxes of respiration and photosynthesis in the burnt and grazed plots relative to the unburnt and un-grazed controls.
In another study carried out at Moor House, Ward et al. (2012) looked at how prescribed burning and grazing influenced the short-term uptake and cycling of carbon in the vegetation and soil in the Moor House long-term plots 18 months after burning (in the 10-year burn treatment). They found that burning affected the vegetation composition and structure reducing the abundance of mature dwarf-shrubs and increasing graminoids.
Lee et al. (2013a) investigated the impact of managed burning on the vegetation composition and diversity in the replicated long-term plots at Moor House. They found that the cover of Calluna and Hypnum jutlandicum increased in the long-unburned ‘reference’ plots, as did bryophyte species richness, but bryophyte cover did not, and lichen diversity declined. Neottia cordata, the locally rare Lesser Twayblade, which is an indicator species of the M19 Calluna vulgaris-Eriophorum vaginatum mire community (Rodwell, 1991), increased in the long-unburned ‘reference’ plots. They found that there was some divergence between the burning rotation treatments with the no-burn treatment changing towards a Calluna and Hypnum jutlandicum community, while the most-disturbed 10-year rotation plots had a much greater abundance of both Eriophorum and Sphagnum spp. They conclude that blanket-bog vegetation responds to managed burning in a complex manner. At the Moor House site where the experiment burn rotation interval is long (20 years), Calluna became dominant and the preferred peat-forming species showed no increase, however, where the burn rotation interval is short (10 years), the abundance of the peat forming species Eriophorum vaginatum and Sphagnum increased.
In a study by Lee et al. (2013b) they looked at the impact of managed burning on the propagule banks of two contrasting blanket bog sites differing in climate, productivity and pollution history. They used the long-term replicated plots at Moor House as well as a chronosequence from more modified bog in the Peak District of Northern England. The propagule banks at both sites were very species-poor and were composed mainly of common species. The chronosequences showed that there were few species that showed significant effects with time since burning. The Calluna seed bank increased in the above-ground litter fraction which acted as a barrier to seed transfer to the underlying peat. Where there is an extensive and deep litter layer a large proportion of the regeneration potential is susceptible to loss by burning. As the burn rotation-interval increased the Calluna seed bank and the frequency of occurrence of Sphagnum species propagules in the peat at Moor House increased. In other words, burning depleted the Sphagnum propagule bank in the peat in the plots at Moor House. They suggest that prescribed burning rotations at two temporal scales may be required to conserve Sphagnum species, short-rotation burns (every 10-years) to enhance Sphagnum abundance in the vegetation (Lee et al., 2013a) and long-rotations (>55 years) to maintain propagules in the surface peat.
Alday et al. (2015) used the Moor House replicated long term plots to look at the cumulative effects of multiple fires and low-grazing levels on the above ground biomass and height of blanket bog vegetation. They found that there was no significant effect of sheep-grazing or its interaction with burning on above-ground biomass or vegetation height, and significant reductions in above-ground biomass and vegetation height were only produced by repeated burning. They found no significant differences in above-ground biomass or vegetation height between the plots that hadn’t been burnt since the 1950s and the unburnt reference plots (no burn since the 1920s). Graminoid vegetation was more abundant in the short rotation treatments (10-year) (independent of grazing), while other vascular plant biomass was only abundant in short rotation un-grazed plots. The biomass of Calluna was greater in the longer rotation interval plots (20-year), while the biomass of bryophytes was greater in the short rotation plots. Both the burning rotation treatments (10-year and 20-year) produced significant reductions in total above-ground biomass compared to unburnt reference plots. No burning for more than 50 years produced a stand dominated mainly by Calluna and litter with a low biomass of bryophytes and other vascular plant species, and a reduction in peat-forming species. They concluded that in order to maximize the carbon fixation in similar moorlands, the fire return-interval should be around 20 years. However, they suggest that this return-interval could reduce the amount of some important peat-forming species such as Sphagnum and Eriophorum, which favour rotation intervals of 10-years. They also suggest that vegetation height might be a useful tool for guiding land managers on when to implement prescribed burning.
Milligan et al. (2018) also used the Moor House long-term replicated plot data to look at changes in vegetation composition and plant functional types. They found that the least disturbed plots were dominated by Calluna and pleurocarpous mosses, whereas the most disturbed plots (burned every 10 years) had greater Eriophorum vaginatum, Sphagnum spp., acrocarpous mosses, liverworts, and lichens. They found that at Moor House increasing the burning frequency increased the species richness of vascular plants with time. Repeated burning at the 10-year rotation period increased the species abundance-weighted Ellenberg values for moisture, reaction and light through time, indicating that the burning treatments were producing a vegetation composed of species that preferred moister and less acidic conditions.
Noble et al. (2018a) looked at the effects of burning and grazing treatments on Sphagnum using the Moor House long-term replicated plots. They found that the unburned reference plots and the plots with the most frequently burned rotation treatment (every 10-years) had greater Sphagnum abundance and hummock height than the plots with the intermediate treatments (20-year rotation and no-burn since 1954). The abundance of the three most common species (S. capillifolium, S. subnitens and S. papillosum) showed similar patterns. At Moor House the low level of grazing had no impact on Sphagnum abundance, and there was no interaction between grazing and burning treatments. They suggest that fire can have a negative impact on Sphagnum, and this can persist for several decades (as appears to be the case in the no-burn since 1954 plots compared to reference plots not subject to the 1954 burns), but the burning rotation interval together with other factors including the level of atmospheric pollution may alter the effects, and in some cases Sphagnum abundance may recover.
Noble et al. (2019a) monitored fire temperatures and their impact on Sphagnum capillifolium in the field at Moor House, as well as cell damage to five different Sphagnum species in response to high temperature in the laboratory. They found that higher temperatures were associated with more cell damage in Sphagnum capillifolium, and all five species of Sphagnum tested in the laboratory showed greater damage at higher temperatures, with the damage occurring immediately after heat exposure. Hotter fires are likely to have a greater impact on the survival and growth of Sphagnum.
7.4.1.2. Other sites
Hancock et al. (2005) looked at whether fire would be a useful management tool to improve the rate of Scots pine regeneration on Calluna dominated moorland. The investigation was carried out at Abernethy Forest in the Cairngorms National Park. The experiment used 10 plots on open moorland approximately 300m from the forest edge. Within each plot there were 10 small exclosures, two for each of four burning treatments, and two unburnt controls. Within each of the exclosures, 100 Scots pine seeds were sown. The numbers of Scots pine seedlings were counted after 2, 11 and 30 months. They found that two months after sowing the Scots pine seed, the burnt plots had 3.5 times the number of seedlings compared to the unburnt controls. They found no significant additional effect of either double burning or year of burn on the number of seedlings established. They suggest that slower moving fires and shallower moss and litter depths post-fire, were associated with greater seedling establishment.
Davies and Legg (2008) looked at lichen diversity across a burning chronosequence at Mar lodge in the Scottish Highlands. They found that the immediate effect of burning was to significantly reduce the diversity of lichens, however it largely recovered within 20 years. The lichen population dynamics were significantly different between areas of wet and dry blanket bog, with terricolous lichens (species growing on the ground) in the wet sites being replaced by pleurocarpous mosses. In general, areas that had not been burnt for 25 years or more, had lower lichen diversity than areas were the heath was 10 to 15 years old. They conclude that whilst fire allows the formation of areas of young vegetation with a high diversity of terricolous lichens (species growing on the ground), areas of older Calluna provide better habitat for corticolous lichens (species growing on bark), and that burn return intervals of less than 15–20 years are likely to lead to a decline in lichen diversity.
Davies et al. (2010) recorded pre-and post-fire vegetation structure and composition, fire behaviour characteristics, and proxy measures of fire severity, at 15 experimental burns on Calluna-dominated upland dry heath (NVC community H12; Rodwell, 1991) in the Cairngorms. They found that regeneration after burning was strongly linked to the age of the stand and the post-fire substrate type. They also found that fire behaviour and severity had little effect, although burning-induced heating of the ground-surface may promote the establishment of Calluna seedlings. Vegetative regeneration was found to be extremely poor in stands of older Calluna, as was seedling establishment where the post-fire substrate was dominated by mats of live or dead pleurocarpous moss.
Harris et al. (2011) looked at the effect of managed burning on vegetation composition at the regional scale from a chronosequence from five severely modified and degraded moorland sites in the Peak District of Northern England. They found that the only species to show an increasing response following burning was Calluna, while all other species showed an increase immediately following burning, but then either declined or showed a unimodal/skewed response. Their results suggest that managed burning maintains plant species diversity in the immediate post-burn phase, but as the vegetation ages and increases in height, most species disappear, and the vegetation becomes dominated by Calluna. They recommend that moorland vegetation should be burned before it reaches 25cm in height to maintain the pre-burn plant species diversity.
Hancock et al. (2011) carried out an experiment at Abernethy Forest to test whether prescribed burning would be a valuable technique for capercaillie (Tetrao urogallus) habitat management. They monitored the vegetation, arthropods and capercaillie dung over a 7-year period (1 year pre-treatment and 6 years post treatment) in 700 metre square plots, that were randomly assigned to control, mowing, or burning treatments. Six years after the treatments, Vaccinium myrtillus cover was significantly higher in the burnt and mown plots compared to the control plots.
In a glasshouse germination trial, Bargmann et al. (2014) compared the seed banks from heath that had not been burnt for 28 years with seed banks from newly burnt Norwegian coastal heath (burned the previous year). They found that both the Ericaceae; and Gramineae species responded to fire cues, whereas herbaceous species did not. The germination responses were stronger in seed banks from the heath that had not been burnt for 28 years compared to seed banks that had already been exposed to recent fire cues. An aqueous smoke solution treatment was found to be a more effective cue than ash across all the plant functional groups.
Taylor et al. (2017) carried out a glasshouse experiment that looked at the ability of Sphagnum capillifolium to recover from exposure to high temperatures, similar to those produced in a managed peatland fire. They found that the level of recovery of Sphagnum capillifolium was related to three factors, the temperature, the post-treatment environmental conditions and the pre-treatment stem moisture content. Recovery was slowest when the Sphagnum was heated to 400 Co for 30 seconds. They concluded that Sphagnum capillifolium has the ability to recover following exposure to temperatures that are likely to occur in prescribed burns, as long as some living material remains.
Grau-Andrés et al. (2017) carried out a study at Braehead Moss, a raised bog in Southern Scotland, to quantify the recovery of Sphagnum following prescribed burns with a range of fire severities. They measured Sphagnum cover and chlorophyll fluorescence F-v/F-m ratio (an estimate of photosynthetic capacity) up to 36 months post-fire. They found that the cover of Sphagnum capillifolium was similar in unburnt and burnt plots. They also found that five months after burning the chlorophyll fluorescence (F-v/F-m) of Sphagnum capillifolium was 0.44 in low fire severity plots and 0.24 in higher fire severity plots compared to 0.67 in unburnt plots. After 22 months this had risen to 0.76 in burnt plots and there were no differences between severity treatments. Similar post-fire recovery was observed for other Sphagnum species, however their low abundance prevented statistical analysis of the data. The results from this study show that Sphagnum capillifolium is resilient to low–moderate fire severities and that there is only a limited and transitory effect of prescribed burning on the abundance and photosynthetic capacity of Sphagnum capillifolium.
Whitehead and Baines (2018) looked at vegetation change across a time-series of burns at an actively managed grouse moor in the North Pennines. The cover of Sphagnum and Eriophorum vaginatum was highest on areas that were last burned within 3-10 years. The cover of Sphagnum was low immediately after burning suggesting an initial burn effect, but then increased, peaking in age-categories 3–6 and 7–10 years before levelling off between 11–17 years and then declining to virtual absence. Sphagnum cover varied between sample blocks, being greatest where peat was deeper. The number of Sphagnum spp. Was lowest in the longer unburned age class (greater than 17 years). The cover of Calluna and other bryophytes showed a linear increase over time since burning, as did the overall vegetation height. The results from this actively managed grouse moor showed that the cover and species richness of Sphagnum, correlated with the reduced dominance of tall Calluna. They suggest that Sphagnum can benefit from a burn return-period of up to 10 years.
Noble et al. (2018b) looked at how managed burning, atmospheric pollution and grazing are related to vegetation community composition and the cover of four taxa (Sphagnum spp., Calluna vulgaris, Eriophorum vaginatum and Campylopus introflexus) using two datasets from a total of 2,013 plots across 95 blanket bog sites in England. Analysis of the data showed differences in vegetation community composition between burned and unburned plots at both the regional (Pennine) and national scales. Burned sites had less Sphagnum and more Calluna cover on a national scale, while on a regional scale, plots burned between 2 and 10 years ago had less Eriophorum vaginatum and more Campylopus introflexus cover than unburned sites. The authors suggest that to promote peat forming vegetation, the routine burning or heavy grazing of peatlands should not be used, and that atmospheric pollution may hinder peat-forming species, particularly on sites that are burned.
Noble et al. (2019b) monitored vegetation composition in different burn-age categories at three blanket bog sites in the north of England. They found that the most recently burned plots were likely to have more bare peat, a thinner moss layer and lower vascular plant height. Following burning, graminoid cover initially increased but after more than ten years graminoid cover was low. The cover of dwarf shrubs increased after burning and remained high after more than ten years. At the site with the most Sphagnum a large proportion of the Sphagnum cover was bleached one year after burning, but then recovered. There was no significant difference in Sphagnum re-introduction success according to burn-age, though success decreased over the study period in the most recent and intermediate burn-age categories at the most Sphagnum-poor site. They found no evidence to support the use of burning as a tool to increase Sphagnum or promote Sphagnum re-establishment.
Whitehead et al. (2021) looked at the post-burn vegetation succession at a moor in south-west Scotland, comparing areas burned between 10 and 5 years prior to sampling (in 2019) with longer-unburnt control plots (unburned since at least 2008). They found Calluna cover, vegetation height and total above-ground biomass increased linearly over time since burning. They also found that Eriophorum cover was lower during the first eight years after burning. The cover of Sphagnum in plots that had been burnt eight to ten years prior to sampling averaged five times higher than that in the no-burn control plots. Sphagnum cover was also positively correlated with peat depth. They conclude that this study supports earlier studies in Northern England by Milligan et al. (2018) and Whitehead and Baines (2018), showing that prescribed burning at regular intervals can increase Sphagnum cover by reducing the cover of Calluna and the biomass of the canopy vegetation.
The review concludes that the evidence from the primary literature indicates that burning results in a change in plant species composition and structure, but that this change is not always consistent. Moorland vegetation responds to managed burning in a complex manner, with habitat type and burn rotation period being important factors, but other potentially confounding factors such as air pollution and grazing may also be important.
There is consistent evidence that Calluna increases over time following burning (Lee et al., 2013a; Milligan et al., 2018; Alday et al., 2015; Whitehead et al., 2018; 2021; Noble et al., 2018b).
Evidence from Moor House in the North Pennines indicates that short-burn rotations lead to an initial increase in Eriophorum vaginatum (Lee et al., 2013a; Milligan et al., 2018), however in an England-wide blanket bog study by Noble et al. (2018b), plots burned 2-10 years ago had less Eriophorum vaginatum cover than unburned sites. One Scottish study showed an increase in Vaccinium myrtillus following burning (Hancock et al., 2011).
The review concludes that the evidence from the primary literature is focussed on blanket bog, and that no studies were identified that looked at the impact of muirburn on other important moorland habitats such as grassland (e.g. Molinia caerulea dominated grassland/mire or Nardus stricta dominated grassland), flushes or alpine heaths. However, it should be noted that Glaves et al. (2013) did review several studies on the burning of Molinia caerulea dominated grassland (Miles, 1971; Ross et al., 2003; Marrs et al., 2004).
The review concludes that there is evidence of mixed responses of Sphagnum to prescribed burning on blanket bog, though most of the evidence relates to one species, Sphagnum capillifolium.
Evidence from Moor House in the North Pennines (Lee et al., 2013a; Milligan et al., 2018) and two other sites (Whitehead and Baines, 2018; Whitehead et al., 2021) indicate that short-burn rotations can lead to an increase in Sphagnum. However, Lee et al. (2013b) found that occurrence of Sphagnum species propagules in the peat at Moor House decreased with increasing frequency of burning and Noble et al. (2018a) found high Sphagnum frequency/cover in the longest-unburned ‘reference’ plots at Moor House and (2018b) that burned sites had less Sphagnum cover on a national scale in English blanket bog than unburned sites.
The review concludes that there is some limited evidence indicating that Sphagnum capillifolium can recover from managed burning.
The review also concludes that there is evidence that hotter fires have a greater impact on Sphagnum growth and survival.
There is very little evidence of the impact of muirburn on other Sphagnum species other than Sphagnum capillifolium.
The review concludes that the evidence base is too limited to draw firm conclusions regarding the impact of muirburn on propagule banks. However, evidence from the only long-term replicated burn experiment in the UK at Moor House in the North Pennines, indicates that burning depletes the Sphagnum propagule bank in the peat.
The review concludes that there is very limited evidence from the primary literature on the impact of muirburn on lichens, however the study by Davies and Legg (2008) showed that burning significantly reduced the diversity of lichens in the short term, but that they largely recovered within 20 years.
7.5. Research Question 2: What is the Impact of Muirburn on Birds?
The search process identified four primary sources that looked at the impact of prescribed burning on birds. It is likely that the search process missed some studies because their titles and abstracts referred to moorland management rather than directly to burning or muirburn. Glaves et al. (2013) evaluated a number of additional bird studies (e.g. Picozzi, 1968; Haworth and Thompson, 1990; Moss et al., 2005; Daplyn and Ewald, 2006; Amar et al., 2009). The main findings from Glaves et al. (2013) relating to the impacts of muirburn on birds is given in Section 7.9. In addition to the papers identified from the Web of Science search, two recent papers highlighted by an independent reviewer have also been reviewed (Douglas et al., 2017; Littlewood et al., 2019).
Smith et al. (2001) looked at the abundance of red grouse and meadow pipits (Anthus pratensis) at sites across Northern England and Scotland. They found that meadow pipit abundance declined with increasing muirburn and increasing Calluna but increased with increasing grass cover. They found that bird species diversity declined with increasing Calluna and Sphagnum cover and with habitat patchiness, and that bird species diversity increased from west to east and on moors with more muirburn.
Tharme et al. (2001) examined data from 320 1km moorland squares across eastern Scotland and Northern England. They looked at whether the population densities of 11 species of moorland breeding birds differed between moorland managed for red grouse and similar moorland vegetation that was not managed for red grouse. Management included rotational burning and predator control. They found that densities of breeding European golden plover (Pluvialis apicaria), northern lapwing (Vanellus vanellus), red grouse and Eurasian curlew (Numenius arquata) were higher on grouse moors compared to the other moors, while meadow pipit, Eurasian skylark (Alauda arvensis), whinchat (Saxicola rubetra) and carrion/hooded crow (Corvus corone/Corvus cornix) were less abundant on grouse moors. They found evidence of a positive effect of burning on the density of red grouse and European golden plover and a negative effect on meadow pipit. They concluded that moorland management benefits some breeding bird species but not others.
Hancock et al. (2011) carried out an experiment to test whether prescribed burning would be a valuable technique for capercaillie habitat management. They monitored the vegetation, arthropods and capercaillie dung over a 7-year period (1 year pre-treatment and 6 years post treatment) in 700 metre square plots, that were randomly assigned to control, mowing, or burning treatments. Capercaillie dung counts suggested that areas that had been burnt or mown, had more summer capercaillie usage than control areas.
Robertson et al. (2017) examined the impact of burning on Calluna height and structure, and red grouse density and breeding success, at 36 moorland sites across Northern England. They found that variation in Calluna height was positively associated with the extent of burning and the relationship between Calluna height and burning was similar on blanket bog and heath (on shallow peat). Red grouse breeding success and post-breeding density were found to be positively associated with the extent of burning, though pre-breeding density was not, and the relationships between red grouse and burning were similar on heath and blanket bog. They suggest that moors which were burned more frequently had higher red grouse breeding success but that this may only apply to moors with burning rotations of more than 8 years.
Douglas et al. (2017) looked at changes in bird abundances in relation to sheep and cattle grazing, vegetation burning and cutting at Geltsdale nature reserve in Cumbria, Northern England. The study used six experimental plots established in 1999, that ranged in size from 1.9 to 2.1 km2, and in altitude from 230-620 m above sea-level. The plots comprised a mosaic of dry and wet heath, blanket bog and rough grassland. The plots had a range of grazing levels (with both sheep and cattle grazing) but the grazing levels were not consistent within plots. The burning and cutting regimes were also not implemented experimentally but followed management needs. Bird surveys were conducted in all plots in 1999, 2002, 2004, 2006, 2009, 2011 and 2013. Douglas et al. (2017) found that where a greater percentage area of a plot had been burned, European golden plover increased in the immediate post-burn years while red grouse declined.
Littlewood et al. (2019) carried out bird surveys across 18 estates in Northern England and South East Scotland. The survey sites ranged from intensively managed grouse moors to moorland sites with no grouse management. Littlewood et al (2019) found positive associations between predator control and the abundance of three ground-nesting wader species, with strong effects for European golden plover and Eurasian curlew, and less strong effects for common snipe (Gallinago gallinago). The only evidence for an effect of burning were a very weak positive relationship between burning and European golden plover numbers and a weak negative effect of burning on the abundance of Eurasian wren (Troglodytes troglodytes).
The review concludes that the evidence from the primary literature indicates that moorland management (which includes managed burning) affects the abundance and diversity of bird species. Some species benefit from moorland management while others do not.
Moorland management often includes more than just prescribed burning; it generally includes predator control, which will affect the abundance and diversity of bird species.
7.6. Research Question 3: What is the Impact of Muirburn on Small Mammals, Reptiles and Amphibians?
The search process identified no primary studies which have directly studied the impact of muirburn on small mammals, reptiles or amphibians. The burning of moorland patches is likely to eliminate most of these species in the short term if they are unable to escape from the fire. Those protected underground in burrows may survive, but no evidence was found to confirm this.
The review concludes that there is a lack of evidence on the impact of muirburn on small mammals, reptiles, or amphibians. This has been identified as a research gap.
7.7. Research Question 4: What is the Impact of Muirburn on Invertebrates?
The search process identified eight sources that looked at the impact of prescribed burning on invertebrates. Two sources (one study) looked at Carabidae (ground beetles), one at Lepidoptera (butterflies and moths), one at Orthoptera (grasshoppers and crickets), one at Arachnida (spiders), two at stream macroinvertebrates, and one at soil macrofauna.
Bargmann et al. (2015; 2016) looked at species diversity and composition of carabid beetles in a coastal heathland in western Norway. The first paper used a 22-year chronosequence of time since last fire. They found that burning increases alpha species richness and is especially important for the richness of open habitat species. They also showed that there were compositional differences between carabid assemblages along the chronosequence. They found a higher species turnover between consecutive years in patches that have been burnt recently compared to patches that have not been burnt for a long time. In the second study they looked at the characteristic species of the successional stages within the coastal heathlands and the traits of species in burnt areas versus areas of older heath. They identified ten carabid species that were indicative of the pioneer stage (0-5 years old), one species that was associated with the building stage (6-14 years old), and one species indicative of the mature stage (15-25 years). Six of the ten species that were identified as characteristic of recently burnt heath were heathland or open habitat specialists. Diet and moisture preference were identified as significant traits determining the abundance of carabids following fire. Collembolan specialists and species that had no moisture preference were more abundant in recently burnt heathland.
Haysom and Coulson (1998) looked at the effects of vegetation structure on the Lepidoptera assemblage associated with Calluna vulgaris-Vaccinium myrtillus heaths under rotational burning at four locations in Northern England and southern Scotland. They found that there was a significant progressive increase in larval diversity with increasing dwarf shrub height, due to the occurrence of uncommon moth species in the taller dwarf shrubs, and a change in the contribution of common species in different height classes. They suggest that recolonization of the growing dwarf shrubs by Lepidoptera occurs rapidly after a burn, and although the density of recolonising migrants may be low, most common species come back within a few years. Haysom and Coulson (1998) suggest that in the short term, the burning of moorland patches is likely to eliminate most invertebrates, particularly those living above-ground. They suggest that recolonization by local immigration is therefore critical in the longer term. Small patches of muirburn are unlikely to create a problem for most invertebrates, particularly flying insects, because they are only required to move short distances to recolonize burned areas (Haysom and Coulson, 1998). For species that are less mobile, or that have small or dispersed populations, recolonisation in the short to medium term is likely to be more difficult.
In a study looking at Orthoptera in four German peat bogs, Hochkirch and Adorf (2007) found that all species persisted on the burned plots, and none experienced substantial declines in abundance compared to the unburned plots. Overall, differences in the composition and abundance of Orthoptera species were more evident between the four bog sites than between the fire treatments or fire season. They suggest that small-scale fires between late winter and early spring do not appear to represent a threat to Orthoptera species.
Hancock et al. (2011) carried out an experiment to test whether prescribed burning would be a valuable technique for capercaillie habitat management. They monitored the vegetation, arthropods and capercaillie dung over a 7-year period (1 year pre-treatment and 6 years post treatment) in 700 metre square plots, that were randomly assigned to control, mowing, or burning treatments. They found that the biomass of spiders was significantly higher in burnt and mown plots than controls, however, the biomass of caterpillars did not show clear differences between treatments.
Johnston and Robson (2018) investigated the effects of ash input on four headwater streams in the UK. They found that the deposition of ash onto streambeds has an effect on the macroinvertebrate assemblage, however the effect appears to be relatively small compared to the high between-stream variation.
In a study examining the benthic macroinvertebrates from 16 peatland sites across Northern England that were either burnt or had no recent history of burning, Ramchunder et al. (2013), found that there were significant differences in species-richness, diversity, dominance, community composition and functional feeding groups between burned and unburnt catchments. They found that in burned catchments, higher levels of suspended sediment concentration and fine benthic particulate organic matter were associated with a lower abundance of some mayflies, stoneflies and caddis-flies, and increased abundance of some Diptera Chironomidae and Simuliidae larvae. Their study suggests that rotational burning has an impact on peatland stream ecosystems.
Mallik and FitzPatrick (1996) looked at biological activity in heathland soils from two sites subject to periodic burning. They found that there were more enchytraeid worms in recently burned soils compared to soils that had burned ten years before. The population of enchytraeid worms in the upper 8 cm of the soil declined with time since burning. They concluded that there appears to be a succession in the soil macrofauna following burning in which earthworms initially predominate after burning followed by an increase in enchytraeids but both diminish over time.
There is some evidence that burning benefits open-ground species such as ground beetles and spiders (2 studies – Bargmann et al., 2015; 2016; Hancock et al., 2011). In the short-term the burning of moorland patches is likely to eliminate most invertebrates, particularly those living above-ground, however the limited evidence indicates that recolonization by local immigration is likely to result in no long-term detrimental impacts to common, mobile species (Haysom and Coulson, 1998). There is a lack of information on the impact of muirburn on less mobile or rare species. There is also no information on the relationship between the size of burn patch and recolonisation, or the effect of repeated burns on invertebrate assemblages.
The review concludes that the evidence base is too limited to draw firm conclusions regarding the impact of muirburn on terrestrial invertebrates.
There is limited evidence from one study that streams within burned catchments had a lower abundance of some mayflies, stoneflies and caddis-flies, and increased abundance of some Diptera larvae (Ramchunder et al., 2013), Another study (Johnston and Robson 2018) showed an effect of adding ash to watercourses, though this was relatively small compared to differences between watercourses.
7.8. Research Question 5: What is the Impact of Muirburn on Soil Microbes?
The search process identified only two sources from the primary literature that looked at the impact of muirburn on soil microbes.
There was one paper that looked at the effect of a single low intensity burn on the soil microbial respiratory activity and fungal community structure of a moorland site adjacent to a native pine woodland. Curlevski et al. (2011) found that a single prescribed burn had no lasting effect on either fungal taxonomic richness, fungal community composition or microbial activity.
In a study carried out at Moor House in the North Pennines, Ward et al. (2012) looked at how prescribed burning and grazing influenced the short-term uptake and cycling of carbon in the vegetation and soil. They found that burning affected the soil microbial community by reducing the total abundance of fungi but it had no effect on the total abundance of soil bacteria.
The review concludes that the evidence base is too limited to draw firm conclusions regarding the impact of muirburn on soil microbes.
7.9. Evidence from Previous Reviews and Reports
A series of previous reviews and reports have sought to assess the effects of muirburn/moorland management practices. Some of these reviews included information on the impact of muirburn on habitats and species.
a) Glaves, D.J., Morecroft, M., Fitzgibbon, C., Leppitt, P., Owen, M. and Phillips, S. 2013. The effects of managed burning on upland peatland biodiversity, carbon and water. Natural England Evidence Review, NEER004. Peterborough: Natural England
The review caried out by Glaves et al. (2013) identified a number of impacts of muirburn on upland peatland habitats and species from the literature. Glaves et al. (2013) cite specific references in their review not all of which have been read for this review due to time constraints. The main findings from Glaves et al. (2013) are summarized below:
Impacts on flora:
There is strong evidence (as interpreted by Glaves et al. (2013)) for:
- Change in species composition (change in relative abundance).
- Initial period of graminoid dominance (10-20 years) and decline in dwarf shrub cover and diversity.
- Molinia caerulea and Trichophorum germanicum initial dominance especially in the west or Eriophorum vaginatum particularly in the Pennines.
- Calluna declines initially but then increases.
- Bryophytes decline initially, but some early colonists may increase relatively quickly. Sphagnum showed mixed responses (increases and decreases).
- Creation of bare ground.
There is moderate evidence (as interpreted by Glaves et al. (2013)) for:
- Increase in Rubus chamaemorus.
- Composition of blanket bog vegetation can continue to show change more than 80 years after the last burn.
- Relatively flat, un-patterned bog surfaces resulting from fire.
- At Moor House in the North Pennines, more frequent burning has promoted the dominance of Eriophorum vaginatum with increased Calluna cover under the longer rotation.
There is weak evidence (as interpreted by Glaves et al. (2013)) for:
- Empetrum nigrum may decline following burning.
Impacts on fauna:
There is strong evidence (as interpreted by Glaves et al. (2013)) for:
- Burning indirectly affects the invertebrate community composition of upland peatland habitats, typically benefiting open-ground species such as ground beetles and surface-active spiders.
- Increase in overall invertebrate species-richness or diversity, through increases in the structural diversity of the vegetation and the presence of open patches.
- Correlations between habitat type, vegetation composition and structure, and densities of some moorland breeding birds, particularly waders.
- Certain species are associated with particular moorland vegetation characteristics.
- Red grouse and European stonechat (Saxicola rubicola) are associated with increasing Calluna.
- Common snipe and Eurasian curlew with structural heterogeneity.
- European golden plover and Eurasian skylark with short vegetation.
- Waders with wet conditions.
- Whinchat with dense vegetation.
- European stonechat with tall vegetation.
- Meadow pipit with grass-heather mixes.
- There are correlations between burning and/or predator control intensity and densities of some moorland breeding birds.
- Higher densities of red grouse, European golden plover, Eurasian curlew, northern lapwing, common redshank (Tringa totanus) and ring ouzel (Turdus torquatus) with increased burning/predator control.
- Lower densities of meadow pipit, Eurasian skylark, northern wheatear (Oenanthe oenanthe) and twite (Linaria flavirostris) with increased burning/predator control.
- Dates of first egg-laying of some moorland bird species and the legal burning season overlap in the first half of April, indicating a potential vulnerability for ground nesting species.
There is moderate evidence (as interpreted by Glaves et al. (2013)) for:
- Too frequent burning is likely to make peatland sites less suitable or unsuitable for the large heath butterfly, but that occasional burning may be beneficial.
- Burning is correlated with changes in the diversity and composition of aquatic invertebrate assemblages in watercourses draining peatland catchments, with declines in certain groups, (mayflies and stoneflies), and increases in flies.
- Earlier nesting over time for eight moorland bird species, which may in the future increase the proportion of first nest attempts by mid-April.
- Greater declines in European golden plover under more intensive burning management.
There is weak evidence (as interpreted by Glaves et al. (2013)) for:
- Correlation between burning and/or predator control intensity and overall diversity of moorland breeding birds.
b) Werritty, A., Pakeman, R.J., Shedden, C., Smith, A. and Wilson, J.D. 2015. A Review of Sustainable Moorland Management. Report to the Scientific Advisory Committee of Scottish Natural Heritage
Werritty et al. (2015) were tasked with providing “an overview of the main moorland management practices, in terms of what is known about their extent, practices and impacts on the natural heritage” (Werritty et al., 2015, p.8); muirburn was one on the moorland practices they reported on. The authors cite specific references in the text, not all of which have been read for this review due to time constraints.
The authors note that moorlands have been created by grazing, tree-felling and fire. They state that dwarf-shrub heaths are maintained by management and have considerable international conservation significance, though species that do not tolerate fire are absent, as are woodland and scrub species. A mosaic of patches of vegetation resulting from managed muirburn can lead to the development of plant, invertebrate and bird communities of conservation significance.
They note that “Conservation outcomes from muirburn depend heavily on the interactions between burning rotation length, patch size, edaphic conditions, other anthropogenic pressures (grazing, drainage and atmospheric deposition), and the timescales over which these interactions are measured” (Werritty et al., 2015, p.15).
On blanket bogs and on wet heaths on “deeper peat soils” (Werritty et al., 2015, p.15) there is evidence of damage to habitat condition, stream invertebrate diversity and problems with re-establishment of Sphagnum.
The authors also highlight the importance of sheep grazing, and to a lesser extent, deer grazing, to upland habitat composition.
c) Werritty, A. 2019. Grouse Moor Management Review Group. Report to the Scottish Government
Werritty (2019) was tasked with reporting on managing grouse moors sustainably and within the law; muirburn was one of the topics covered. A list of references was provided but specific papers were not cited in the text. The reference list includes several papers that have not been reviewed here as they did not appear in our Web of Science search and time constrains meant we were unable to look beyond this.
Werritty (2019) state that three-quarters of Europe’s Calluna-dominated moorlands, a priority habitat for the EU, are within the UK, and that this is largely not a natural landscape but a cultural one created by deforestation, fire and grazing. Muirburn limits succession of the vegetation towards the woodlands that would be the natural habitat in a proportion of the ‘cultural’ moorlands currently present. Establishment of young trees is reliant on nearby seed sources, so even without muirburn, conversion of moorland to woodland would only occur over a limited area in the short and medium term. Regeneration of woodland results in biodiversity losses for the species associated with open moorland and biodiversity gains for the species associated with woodland.
Werritty (2019) asks “is burning necessary to retain heather-dominance?” (Werritty, 2019, p.36). While moorland managers have assumed that burning was needed to keep Calluna dominant, this is known to be incorrect for some unburned areas where Calluna plants have stayed dominant for 40-60 years by stem layering or rooting. Werritty (2019) highlights the lack of information on the proportion of Scottish moorland that has never been burned, and therefore the inability to predict the proportion of Calluna dominated moorland that would convert to woodland if burning stopped. The debate about the ‘need’ for burning to maintain Calluna dominance (rather than to provide young nutritious shoots) would be resolved by this information.
Werritty (2019) highlights fire characteristics, and especially fire intensity as a driver of biodiversity and wider ecosystem outcomes. Some of the debate about the opposing biodiversity impacts of muirburn is attributed to a lack of enquiry into fire characteristics. Few studies have recorded fire intensity, so understanding of the impact of fire intensity on the impact of the fire is limited. Future studies should record fire characteristics, including fire intensity and fire impacts. Fire intensity is affected by fuel load, weather, moisture content and other factors. The deeper the fire burns down into the moss layer, the litter layer, and the soil/peat below, the greater the impact of the fire and the likelihood of damage.
The impacts of muirburn depend on the habitat being burned. Werritty (2019) states that in dry heaths, muirburn carried out following the Muirburn Code can benefit plant, bird and invertebrate biodiversity compared with unburned moorland, by creating a mosaic of habitat structures. There is inconclusive evidence that muirburn is damaging to blanket bog and wet heath habitats and peat-forming plants but note that one long term study from the Pennines (the Hard Hill experiment at Moor House) has found burning to be beneficial. There is strong evidence that muirburn can be damaging to biodiversity (and hydrology, soil stability and other system components) “in some situations” (Werritty, 2019, p.35).
There is evidence of both positive and negative impacts of muirburn on biodiversity (and soil stability and hydrology), with most positive examples coming from dry heath and most negative examples coming from wet heaths and peatlands. There is a high level of disagreement in the literature and a large number of knowledge gaps.
d) Newey, S., Fielding, D., Miller, D.G., Matthews, K.B. and Thomson, S. 2020. Biodiversity considerations on grouse moors. Part 4 of Research to assess socioeconomic and biodiversity impacts of driven grouse moors and to understand the rights of gamekeepers. Report to the Scottish Government
Newey et al. (2020) did a wide-scale correlative desk-study looking at relationships between the intensity of grouse moor management and the presence/absence of ten moorland biodiversity indicator species: birch (Betula spp.), blaeberry (Vaccinium myrtillus), green hairstreak butterfly (Callophrys rubi), adder (Vipera berus), Eurasian curlew, European golden plover, merlin (Falco columbarius), common kestrel (Falco tinnunculus), lesser redpoll (Acanthis cabaret) and whinchat. ‘Intensity of grouse moor management’ was defined as the percentage area of one kilometre Ordnance Survey grid squares with signs of muirburn, assessed by aerial photography or satellite imagery. Over 3,600 one km squares were used, with the study focussed on areas where driven grouse moor management was already known to be an important land use (mainly selected upland areas in the Eastern half of Scotland). As the study utilised existing species distribution data which had been collected for a different purpose, the results should be interpreted with caution.
The indicator species chosen were those thought likely to be positively or negatively affected by grouse moor management intensity and that had adequate occurrence data: birch, green hairstreak butterfly, adder, blaeberry (at the one km square resolution) and Eurasian curlew, merlin, lesser redpoll, European golden plover, kestrel and whinchat (at the 10 km square resolution). The burning categories were:
- ‘no burning / very small area burned’ (0-20 %),
- ‘small area burned’ (21-40 %),
- ‘moderate area burned’ (41-60 %),
- ‘large area burned’ (61-80 %) and
- ‘very large area burned’ (81-100 %)
The majority of squares were in the lowest burn area category, with decreasing numbers of squares as the burn area increased. At the 10 km square level, less than ten squares were in the ‘large area burned’ category and only one square was in the ‘very large area burned’ category, meaning that the results should be interpreted very cautiously. Species distribution data was acquired from various existing sources.
The authors state that it is difficult to draw any firm conclusions, but the tentative findings are:
- Birch presence was negatively associated with increasing burn area.
- Blaeberry presence was slightly higher in the ‘small area burned’ category and in the ‘large area burned’ category than in other categories.
- Green hairstreak butterfly is a relatively rare species, apparently present in a greater proportion of squares in the ‘small area burned’, very large area burned’ and ‘no burning / very small area burned’ categories than in the other two categories. However, this species is hard to detect, the results may stem from the fact that it is easier to find it in short vegetation that has been burned.
- Adder, another relatively rare species, had relatively high presence in the ‘no burning / very small area burned’ category, then apparently showed a trend of increasing presence with increasing area burned. Again, this may stem from the fact that it is easier to find it in short vegetation that has been burned.
- Eurasian curlew presence was positively associated with area burned.
- European golden plover abundance increased with area burned up to the category ‘moderate area burned’ then decreased.
- Merlin presence increased with area burned up to the category ‘moderate area burned’ then decreased.
- Common kestrel presence apparently increased slightly with increasing burn area.
- Lesser redpoll was present in a greater proportion of squares in the ‘large area burned category’ than in other categories.
- Whinchat was present in a greater proportion of squares in the ‘large area burned category’ than in other categories with lower burning.
The authors suggest that birch presence having a negative relationship with area burned is unsurprising, as one of the purposes of muirburn is to maintain open habitats. They note that muirburn prevents the succession of vegetation towards scrub and woodland and the establishment of the species and communities that are associated with these habitats. They also note that “species may be responding to aspects of moorland management other than muirburn” (Newey et al., 2020, p.2) and that for the bird species studied, occurrence is probably also influenced by other characteristics of the landscape.
e) Heinemeyer, A., Vallack, H.W., Morton, P.A., Pateman, R., Dytham, C., Ineson, P., McClean, C., Bristow, C. and Pearce-Higgins, J.W. 2019. Restoration of heather-dominated blanket bog vegetation on grouse moors for biodiversity, carbon storage, greenhouse gas emissions and water regulation: comparing burning to alternative mowing and uncut management. Final Report DEFRA Project BD5104. DEFRA
In a DEFRA funded project, Heinemeyer et al. (2019) looked at the initial management impacts of burning and mowing on Calluna-dominated blanket bog, in a replicated, multi-year study at three sites in Northern England under grouse moor management (Mossdale and Nidderdale in the Yorkshire Dales National Park, and Whitendale in the Forest of Bowland Area of Outstanding Natural Beauty, in Lancashire). The study was designed as a paired catchment scale study together with plot level replication. At each site there were two similar adjacent sub-catchments that were randomly allocated either a burning or mowing management, with various plot-level managements, including an additional uncut plot-level management within each mown sub-catchment. The project covered an initial 5-year period.
The main findings from the study relating to biodiversity included:
- The re-growth of Calluna was initially slower on burnt plots compared to mown plots, although after 4 years, Calluna cover and height was similar on burnt and mown plots.
- Cotton-grass (Eriophorum spp.) cover increased on both burnt and mown plots. Eriophorum vaginatum cover was significantly greater on mown than burnt plots, however this was the case to some extent in the pre-management period.
- The cover of non-Sphagnum mosses, particularly Hypnum jutlandicum and Campylopus introflexus, was greater on burnt than on mown plots, but this difference was also partly the case in the pre-management period.
- The cover of Sphagnum was not significantly affected by management and remained relatively constant across time under all management regimes, however there was an increase in total cover of Sphagnum on mown plots in the final year (year 4) at the wetter sites. This together with differences in cotton-grass and non-Sphagnum moss cover as well as a more species-specific ecological assessment, indicated a possible different long-term trajectory between burnt and mown plots.
- Overall plant species diversity was low and decreased from the wettest to the driest site. The driest site had the highest number of Sphagnum species, but the lowest Sphagnum cover, with the wettest site having the highest cover.
- Of the management treatments burning appeared to be the least beneficial towards supporting characteristic bog vegetation, particularly at the driest site. The uncut option showed few drawbacks except for the limited recovery of a peat-forming bryophyte layer at the driest site. Mowing appeared to encourage the growth of key species and the re-establishment of a peat forming bog community, particularly at the wettest site, together with an increase in Sphagnum capillifolium.
- There was higher cranefly emergence in the mown plots compared to the burnt plots in the dry year after management (2014). However, cranefly emergence was reduced in the following two wet years in the mown plots.
- Cranefly emergence was consistently lower on the wettest site, particularly in the wetter mown plots.
- Cranefly abundance on transects was overall higher in mown than burnt catchments.
- The modelling of European golden plover fledging production indicated that numbers would be higher in mown than burnt areas, with greatest effect in the relatively dry year of 2014 when cranefly abundance was lowest.
- Modelling of the effects of drier summers, based on cranefly emergence and soil moisture data, predicted a greater resilience to future drier summers of upland bird numbers (i.e. dunlin (Calidris alpina), European golden plover and red grouse) under mowing, particularly when brash was left, compared to burning.
7.10. Conclusions
7.10.1. What we know
- The review concludes that the impact and influence of muirburn on moorland habitats and species is complex.
- The review concludes that moorland management (which includes prescribed burning) affects the abundance and diversity of bird species. Some species benefit from moorland management while others do not.
- The review concludes that burning results in a change in plant species composition, but that this change is not always consistent. Moorland vegetation responds to managed burning in a complex manner, with many factors including habitat type and condition, soil moisture content, geographical location, scale of muirburn management, burn rotation length, fire severity and intensity, air pollution and other management such as grazing, all having an effect.
- There is consistent evidence that Calluna increases over time following burning.
- The review concludes that there is mixed evidence as to the response of Sphagnum to prescribed burning, however there is some evidence indicating that Sphagnum capillifolium can recover from managed burning.
7.10.2. What we know we don’t know
- In general, there is limited evidence describing the effects of muirburn on the full range of moorland habitats and species across the UK. For some moorland vegetation types, species groups and species, there is no evidence base, and for some others there is a contested evidence base. The review concludes that the evidence from the primary literature is focussed on blanket bog and upland heath, and no studies were identified from the Web of Science search that looked at the impact of muirburn on other important moorland habitats such as grassland (e.g. Molinia caerulea dominated grassland or Nardus stricta dominated grassland), flushes or alpine heaths.
- The review concludes that there is a lack of evidence on the impact of muirburn on small mammals, reptiles, or amphibians.
- The review concludes that the evidence base is too limited to draw firm conclusions regarding the impact of muirburn on terrestrial invertebrates.
8. Identifying Practical Approaches for Defining Peat and Peatlands for Muirburn Licensing Purposes
8.1. Setting the Research and Development Questions
The research and development questions set up for this review were:
- What are the ranges and types of practical approaches that can be used to identify peat and peatlands relevant to inform muirburn management decisions?
- Knowing that the depth of peat layer is a commonly used criteria for identifying peat soils, have there been studies looking at the impacts of muirburn on different type/depth of peat soils?
- Have there been other factors used to identify peat and peatlands in studies of the assessment of the impacts of muirburn?
8.2. Research and Development Question 1: What are the Ranges and Types of Practical Approaches that can be Used to Identify Peat and Peatlands Relevant to Inform Muirburn Management Decisions?
8.2.1. Introduction
There is no mention of the words peat or peatland in the primary legislation governing muirburn in Scotland (the Hill Farming Act 1946, as amended by more the Climate Change (Scotland) Act 2009 and the Wildlife and Natural Environment (Scotland) Act 2011).
In the Muirburn Code, peat is defined as “an organic soil, which contains more than 60 per cent organic matter and exceeds 50cm in thickness” (Scotland’s Moorland Forum, 2017, p.8; see Bruneau and Johnson (2014) for more information on peat). The Muirburn Code states that “burning should not take place on peatland, except as part of a habitat restoration plan, approved by NatureScot….Areas with peat hags, bare peat or erosion should not be burnt” (Scotland’s Moorland Forum, 2017a, p.8).
None of the papers reviewed specifically focused on practical approaches for identifying peat and peatlands that were relevant to inform muirburn management decisions.
8.2.2. Mapping and measuring peat depth
A number of methods have been used to measure peat depth, but most of these are not practical or appropriate for land managers. Some of the methods are used to map at scale and some are only appropriate for research purposes or to aid large scale management decision making. Some of these methods are outlined below:
The use of manual methods (e.g. peat depth probes, core samples and ground penetrating radar) have been the most widely used methods for measuring peat depth (Parry et al., 2014), however they are not practical over large areas (Fyfe et al., 2014; Gatis et al., 2019). More recent studies have focussed on the use of spatial modelling using field sampling and digital elevation models (Parry et al., 2012; Poggio and Gimona, 2014; Young et al., 2018), elevation, slope and disturbance covariates (Holden and Connolly, 2011), topographical (elevation, slope, aspect) and superficial geology covariates (Finlayson et al., 2021), remote sensing and site characteristics (Aitkenhead, 2016) and digital soil mapping (Aitkenhead and Coull, 2019), to map peat depth.
Airborne gamma radiometric data has also been used to estimate peat depth, as peat attenuates naturally occurring radiation that is emitted from underlying bedrock (Keaney et al., 2013; Gatis et al., 2019). Radiometric surveys have limited ability to infer peat depth where the organic horizon exceeds 50cm, and the radiometric signal varies with bedrock type (Minasny et al., 2019). Marchant (2021) developed a framework for optimizing the locations and number of manual peat depth measurements required for surveys that incorporate radiometric data measurements from airborne surveys.
8.2.3. Practical approaches
- Peat probes. The use of peat probes is probably the most practical method for measuring peat depth. Peat probes are narrow rods usually made from metal or fibreglass. They are simple to use and are relatively low cost (in the region of £100). The peat depth can be measured relatively easily by pushing the rod into the peat until the mineral base is reached. The depth to which the probe sinks indicates the peat depth in that local area. Details on carrying out a peat depth survey are given in a NatureScot guidance document Peat Depth and Peatland Condition Survey. This guide is designed for applicants wishing to restore peatlands under the Peatland Action Programme. The survey is completed on an approximate 100m by 100m grid which provides information on the variability of the peat depth. At each intersection of the grid a location is taken using a GPS (Peat depth and peatland condition survey). Other peatland survey guidance is available in relation to developments on peatland (Scottish Government, Scottish Natural Heritage, SEPA, 2017). The detailed methodology used in these two guidance documents may not be appropriate for muirburn licensing purposes. The NatureScot guidance document advises that bamboo canes should not be used as an alternative to a peat probe as they will snap and could cause injury. If all that is required is to determine whether the peat depth is above a particular threshold, then a simple low cost 1 metre fibreglass or metal rod marked with the threshold depth may be sufficient, with measurements taken across the proposed area to be burnt using a 100 m by 100 m grid. This will be much quicker than measuring the full peat depth using interconnecting rods, i.e. less than a minute once the sampling point is located. As such, it could be reasonable to increase sampling intensity if this is desirable, especially if the proposed area to be burned is small, as there can be considerable variation in peat depth on sites, even at nearby points.
- Vegetation assessment. The presence of key blanket bog indicator species could be used as a proxy for identifying peatlands. This however provides no information on peat depth. NatureScot have produced an information sheet on Blanket Bog (UK BAP Priority Habitat) which provides advice on how to recognise blanket bog. Many upland land managers carry out herbivore impact assessments on their moorlands, including areas of blanket bog, using the Best Practice Guidance. This guidance provides information on key indicator species of blanket bog. The presence of indicator species such as Sphagnum spp., Cowberry (Vaccinium vitis-idaea), Hare’s-tail Cotton-grass (Eriophorum vaginatum), Common Cotton-grass (Eriophorum angustifolium), Crowberry (Empetrum nigrum), Deer grass (Trichophorum sp.) and Cross-leaved heath (Erica tetralix) could be used as a proxy to identify the presence of peat. However, the absence of any or all of these indicator species does not necessarily mean that the substrate is not peat; neither does the presence of these species necessarily mean that the substrate is peat.
- Habitat and soil maps. The use of habitat and soil maps to identify peatland areas could be used, however the existing national data sets are not at a fine enough resolution to be used with any degree of confidence. They also give no indication of peat depth. They could however be used to inform land managers of where peat depth assessments should be carried out.
- Airborne gamma radiometric data and spatial modelling. Although these techniques are neither available nor practical for land managers, improved national coverage of airborne gamma radiometric data and/or spatial modelling may help inform NatureScot as to the distribution of peat on particular sites. The development of digital soil mapping could provide high resolution maps with information on carbon stocks that could be useful for regulatory bodies and policymakers (Aitkenhead and Coull, 2019).
The review concludes that there are a number of approaches that can be used to identify peat and peatlands, however the only practical, reliable and cost-effective approach, is the use of some form of probe to measure either the actual peat depth or to ascertain whether the peat depth is beyond a particular threshold. There is however a constraint with this method in terms of the time required to carry out a survey, which will depend on the scale involved and the level of detail required.
8.3. Research and Development Question 2: What are the Impacts of Muirburn on Different Depths of Peat?
Detailed information on the depth of peat or variation in peat depth was rarely given in the primary papers that were reviewed. Very few studies were found in the literature that specifically looked at the impact of muirburn on different depths of peat. Whitehead and Baines (2018) measured vegetation and peat depth in their study on vegetation responses to prescribed burning on an actively managed grouse moor in Upper Teesdale in the North Pennines. They found that Sphagnum cover was greatest where peat was deeper, while Calluna cover was lower where peat depth was greater. Whitehead et al. (2021) measured vegetation and peat depth from moorland blocks of different ages since last burned, on a managed grouse moor in south-west Scotland. They found that the cover of Sphagnum was positively correlated with peat depth.
The review concludes that there is a lack of evidence to determine the impacts of muirburn on different depths of peat.
8.4. Research and Development Question 3: Have there been Other Factors Used to Identify Peat and Peatlands in Studies of the Assessment of the Impacts of Muirburn?
No studies looking at the impacts of muirburn have been found in the primary literature that have used other factors to identify peat and peatlands.
9. Identifying the Knowledge Gaps
The questions relating to the impacts of muirburn, around which this review has focussed, have not been fully addressed and a number of research gaps have been identified. Since a lack of evidence does not imply an absence of impact, more research is required to fill these knowledge gaps in order to provide robust and definitive answers to these questions.
9.1. Why are there Knowledge Gaps and What are Some of the Problems with the Existing Evidence Base?
- There is a relatively limited geographic distribution of study sites in the UK. UK moorlands occupy a wide geographic, altitudinal and climatic range. They also differ widely in terms of species composition, previous and current management, levels of pollution, and degree of damage.
- The impacts of muirburn are complex and there are a limited number of studies assessing these impacts and interactions. There is a need for more replicated, controlled, randomised studies, with before and after measurements.
- There has been a particular research focus on blanket bog vegetation and to a lesser extent upland Calluna-Vaccinium dry heath, with some studies looking at coastal heath and raised bog.
- Much of the research focusses on muirburn as a management tool for grouse moor management, with only a few studies including sites burnt for livestock grazing.
- Many of the studies identified in the review have used the long-term replicated plots on blanket bog at Moor House NNR in the North Pennines of Northern England. This is the only long-term replicated study site in the UK. Moor House is not representative of the range of moorland habitats and muirburn practices found across Scotland. Priority should be given to setting up long term monitoring programs that cover more than one burn rotation on upland dry and wet heath.
- Many of the studies identified in the review use chronosequences (space-for-time substitution studies) where an assumption is made that no other variable besides age has changed between the sites being studied.
- There is limited information on the intensity (or the variables associated with intensity: fuel load, fuel moisture content, litter layer moisture content, wind speed) or severity of controlled burns.
- Experimental burns tend to be small scale (i.e. they may not be representative of actual management practice). More evidence from studies where the burning is carried out in the same manner as would be done by moorland managers would improve the knowledge base.
- Detailed information on the depth of peat or variation in peat depth was rarely given in the primary papers that were reviewed. This information should be recorded.
- Much of the research work has been carried out by a small number of research teams, and some of these research teams appear to have reached conflicting conclusions. Scientifically rigorous and unbiased research is essential to enable well-informed and unbiased policy decision making.
- Many studies are confounded by other factors such as grazing management, drainage, air pollution and predator control. Developing studies that remove or fully account for these factors would improve the knowledge base.
- Although there have been a number of reviews on the impact of muirburn, including this current review, a full systematic review of all the evidence to date would help inform future muirburn management and muirburn policy.
9.2. Research Gaps Relating to the Relationship Between Muirburn and Wildfire
- There is a limited evidence base with respect to the frequency that muirburn becomes a direct cause of wildfire, further research is required to improve confidence in the range of estimates.
- Research is needed to look at whether muirburn controls wildfire occurrence by managing fuel load, as there is currently no direct evidence in a UK context.
- Research is required to investigate the marginal influence of variation in Calluna fuel load on the severity of burns, across different seasons and fire weather conditions.
- Research is required to investigate the influence of variation in Calluna fuel load on the spatial extent of wildfires in a UK context, as there is currently no direct evidence in a UK context.
- Research is required to consider what data would be required to enable a robust approach to these questions.
9.3. Research Gaps Relating to the Impact of Muirburn on Carbon Emissions and Storage
- Further research is needed to disentangle the effect of different fire rotation-intervals on above-ground biomass, as such knowledge will assist in determining the fire rotation-interval that optimizes carbon fixation by means of vegetation growth (Alday et al., 2015).
- Further research is required to better understand the interactions between burning and other disturbances (such as, grazing, drainage, and nutrient deposition) on carbon cycling and vegetation dynamics.
- There are few complete carbon budgets from UK peatland sites subject to management burning and further research would improve the knowledge base. There is at present little evidence of how burning affects the emission of carbon dioxide (CO2) and methane (CH4).
- The accurate recording of bulk density and total peat depth as part of peat carbon stock assessments would help in calculating/estimating carbon stocks and budgets.
- There is little or no carbon budget data for dry heathlands or other moorland habitats.
9.4. Research Gaps Relating to the Impact of Muirburn on Habitats and Species
- Research is required on the impact of prescribed burning on moorland reptiles, amphibians, small mammals and gastropods.
- Further research is required to look at the impact of muirburn on invertebrates particularly less-mobile species and species with restricted populations. Research is also required on the relationship between the size of burn patch and recolonisation by invertebrates, and the effect of repeated burns and burn interval period on invertebrate assemblages.
- Further research is required on the interaction of grazing and muirburn and its impacts on moorland vegetation.
- Research is required on the impact of prescribed burning on other moorland habitat types other than blanket bog and upland dry and wet heath, such as alpine heaths, flushes and montane grasslands. Although these habitats are not the key focus for muirburn management they do form part of the moorland mosaic and are often of high conservation value.
- More research is required on the impact of muirburn on Sphagnum in particular the impact on species other than Sphagnum capillifolium.
10. Overall Conclusion
This review, like many of the previous reviews that have been referenced, has found that the evidence base surrounding the impacts of muirburn on wildfire, soil carbon and habitats and species is limited and sometimes contested.
The questions relating to the impacts of muirburn, around which this review has focussed, have not been fully addressed. In particular, there are a number of knowledge gaps that need to be filled in order to provide robust and definitive answers to these questions and to determine the pros and cons of muirburn in regard to the suite of upland ecosystem services that the UK moorlands provide.
11. Glossary
Acrocarpous Moss: Acrocarpous mosses produce their sporophytes (spore containing capsules) at the ends of main stems, grow in an obviously vertical orientation, and are usually sparsely branched.
Arachnida: Spiders, ticks, mites, harvestmen and other related invertebrates.
Biomass: Is the mass of organic material that comes from living organisms in a given area. Above-ground biomass relates to the organic material above the ground surface (i.e. it does not include live plant roots, soil micro- and macro-organisms, or soil organic matter from decomposing plants and animals). Biomass can include both live and dead material.
Burn Rotation: The period of time (or interval) between repeated managed burns.
“Burn Severity: A qualitative assessment of the heat pulse directed toward the ground during a fire. Burn severity relates to soil heating, large fuel and duff consumption, consumption of the litter and organic layer beneath trees and isolated shrubs, and mortality of buried plant parts” (Scotland’s Moorland Forum, 2017c, page 4.1). See Keeley (2009) for a more detailed definition of the term burn severity.
Carabid (Carabidae): Ground beetles
Carbon sequestration: The process of capturing and storing atmospheric carbon dioxide.
Chironomidae: Non-biting midges
Chronosequence (and associated space-for-time substitution): A set of sites that share similar attributes (e.g. soil, geology, altitude), but represent different ages since they were formed. Space-for-time substitution: space-for-time substitution is a method for studying slow ecological processes (such as vegetation change following burning), where the relationships between ecological variables are studied at sites that are at different stages of development (for example time since last burned) (Walker et al., 2010).
Collembolan specialists: Feed on Collembola (springtails).
Corticolous Lichens: Lichens that grow on the surface of bark.
DOC: Dissolved Organic Carbon. Dissolved Organic Carbon or DOC is a measurement of the amount of organic matter in water that can pass through a filter (normally 0.45 µm).
Ephemeroptera: Mayflies.
Ericacaea: The heath or heather family.
“Fire Behaviour: The manner in which a fire reacts to the influences of fuel, weather, and topography and relates to the way fuels ignite, flame develops and fire spreads” (Scotland’s Moorland Forum, 2017c, page 4.1).
“Fire Intensity: The intensity of a fire is a measure of the rate that energy is released by a fire and is a combination of the amount of fuel that burns and the rate of spread of the fire. A good guide in the field to the intensity of a fire is the flame length. Fire intensity is used to judge how difficult a fire is to put out” (Scotland’s Moorland Forum, 2017c, page 4.1). See Keeley (2009) for a more detailed definition of the term fire intensity.
“Fire Severity: A general term most commonly used to describe the combined effects of both flaming and smouldering combustion on a fire site that creates impacts on plants, soils, and wildlife” (Scotland’s Moorland Forum, 2017c, page 4.2). See Keeley (2009) for a more detailed definition of the term fire severity.
Fireline Intensity: The rate of heat release per unit length of fire front (the line or strip along which there is continuous flames) per unit of time.
Fuel Load: The total amount of combustible material (fuel) expressed quantitatively in terms of mass per unit area.
Gramineae: Grasses.
Graminoid: Herbaceous plants with a grass-like morphology. Refers to grasses, sedges, cotton-grasses and rushes.
Lepidoptera: Butterflies and moths.
Orthoptera: Grasshoppers, crickets and related insects.
Plecoptera: Stoneflies
Pleurocarpous Moss: Pleurocarpous mosses produce sporophytes (spore containing capsules) laterally from the sides of stems, and tend to grow in a horizontal spreading manner forming carpets, and are usually much branched.
“Prescribed Burning: Is the planned application of fire under prescribed environmental conditions, and within defined boundaries, to achieve required objectives” (Scotland’s Moorland Forum, 2017c, page 4.2). Prescribed burning is an alternative name for controlled or managed burning.
Simuliidae: Blackflies.
Succession: The process of change in the species composition of an ecological community over time.
Terricolous Lichens: Lichens that grow on the soil as a substrate.
Trichoptera: Caddisflies.
Wildfire: An unplanned, unmanaged, and unpredictable fire in an area of combustible vegetation. In the UK the term wildfire is defined as “any uncontrolled vegetation fire which requires a decision, or action, regarding suppression” (Scottish Government, 2013)
12. References
Aitkenhead, M.J. 2016. Mapping peat in Scotland with remote sensing and site characteristics. European Journal of Soil Science, 68(1), 28–38.
Aitkenhead, M. and Coull, M. 2019. Mapping soil profile depth, bulk density and carbon stock in Scotland using remote sensing and spatial covariates. European Journal of Soil Science, 71(4), 553–567.
Albertson, K., Aylen, J., Cavan, G. and McMorrow, J. 2009. Forecasting the outbreak of moorland wildfires in the English Peak District. Journal of Environmental Management, 90, 2642–2651.
Albertson, K., Aylen, J.M., Cavan, G. and McMorrow, J. 2010. Climate change and the future occurrence of moorland wildfires in the Peak District of the UK. Climate Research, 45, 105–118.
Alday, J.G., Santana, V.M., Lee, H., Allen, K.A., and Marrs, R.H. 2015. Above-ground biomass accumulation patterns in moorlands after prescribed burning and low-intensity grazing. Perspectives in Plant Ecology, Evolution and Systematics, 17, 388–396.
Allen, K.A., Harris, M.P.K. and Marrs, R.H. 2013. Matrix modelling of prescribed burning in Calluna vulgaris-dominated moorland: short burning rotations minimize carbon loss at increased wildfire frequencies. Journal of Applied Ecology, 50(3), 614–624.
Allen, K.A., Denelle, P., Sánchez Ruiz, F.M., Santana, V.M. and Marrs, R.H. 2016. Prescribed moorland burning meets good practice guidelines: A monitoring case study using aerial photography in the Peak District, UK. Ecological Indicators, 62, 76–85.
Ashby, M.A. and Heinemeyer, A. 2020. Prescribed burning impacts on ecosystem services in the British uplands: A methodological critique of the EMBER project. Journal of Applied Ecology, 57(11), 2112–2120.
Ashby, M.A. and Heinemeyer, A. 2021. A Critical Review of the IUCN UK Peatland Programme’s Burning and Peatlands Position Statement. Wetlands, 41, 56.
Aylen, J., Cavan, G. and Albertson, K. 2007. The best strategy for mitigating moorland wildfire risk. A Report to Moors for the Future.
Baggaley, N.J., Britton, A.J., Barnes, A., Buckingham, S., Holland, J.P., Lilly, A., Pakeman, R.J., Rees, R.M., Taylor, A. and Yeluripati, J. 2021. Understanding carbon sequestration in upland habitats. ClimateXChange Report.
Bargmann, T., Hatteland, B.A. and Grytnes, J.A. 2015. Effects of prescribed burning on carabid beetle diversity in coastal anthropogenic heathlands. Biodiversity Conservation, 24, 2565–2581.
Bargmann, T., Heegaard, E., Hatteland, B.A., Chipperfield, J.D. and Grytnes, J.A. 2016. Species trait selection along a prescribed fire chronosequence. Insect Conservation Diversity, 9, 446–455.
Bargmann, T., Maren, I.E. and Vandvik, V. 2014. Life after fire: smoke and ash as germination cues in ericads, herbs and graminoids of northern heathlands. Applied Vegetation Science, 17, 670–679.
Blundell, A. and Holden, J. 2014. Using palaeoecology to support blanket peatland management. Ecological Indicators, 49, 110–120.
Brooker, R., Hester, A., Newey, S. and Pakeman, R. 2018. Socio-economic and biodiversity impacts of driven grouse moors in Scotland: Part 2 Biodiversity impacts of driven grouse moors in Scotland. Report to the Scottish Government.
Brown, L.E., Holden, J. and Palmer, S.M. 2014. Effects of Moorland Burning on the Ecohydrology of River Basins. Key findings from the EMBER project. University of Leeds.
Brown, L.E., Palmer, S.M., Johnston, K. and Holden, J. 2015. Vegetation management with fire modifies peatland soil thermal regime. Journal of Environmental Management, 154, 166–176.
Bruneau, P.M.C. and Johnson, S.M. 2014. Scotland’s peatland – definitions & information resources. Scottish Natural Heritage Commissioned Report No. 701.
Chapman, D.S., Bonn, A., Kunin, W.E. and Cornell, S.J. 2010. Random Forest characterization of upland vegetation and management burning from aerial imagery. Journal of Biogeography, 37, 37–46.
Chapman, S., Hester, A., Irvine, J. and Pakeman, R. 2017. Muirburn, peatland and peat soils – an evidence assessment of impact. ClimateXChange Report.
Chen, Y., McNamara, N.P., Dumont, M.G., Bodrossy, L., Stralis-Pavese, N. and Murrell, C. 2008. The impact of burning and Calluna removal on below-ground methanotroph diversity and activity in a peatland soil. Applied Soil Ecology, 40(2), 291–298.
Clay, G.D. and Worrall, F. 2011. Charcoal production in a UK moorland wildfire – How important is it? Journal of Environmental Management, 92(3), 676–682.
Clay, G.D, Worrall, F. and Aebischer, N.J. 2012. Does prescribed burning on peat soils influence DOC concentrations in soil and runoff waters? Results from a 10-year chronosequence. Journal of Hydrology, 448–449, 139–148.
Clay, G.D., Worrall, F. and Aebischer, N.J. 2015. Carbon stocks and carbon fluxes from a 10-year prescribed burning chronosequence on a UK blanket peat. Soil Use and Management, 31(1), 39–51.
Clay, G.D., Worrall, F. and Fraser, E.D.G. 2009. Effects of managed burning upon dissolved organic carbon (DOC) in soil water and runoff water following a managed burn of a UK blanket bog. Journal of Hydrology, 367, 41–51.
Clay, G.D., Bonn, A., Evans, M.G., Hewson, W., Parnell, M., Wilkinson, R. and Worrall, F. 2010a. Grindsbrook Wildfire 2008 - a case study. Moors for the Future Report. Edale: Moors for the Future Partnership.
Clay, G.D., Worrall, F. and Fraser, E.D.G. 2010b. Compositional changes in soil water and runoff water following managed burning on a UK upland blanket bog. Journal of Hydrology, 380, 135–145.
Clay, G.D., Worrall, F. and Rose, R. 2010c. Carbon budgets of an upland blanket bog managed by prescribed fire. Journal of Geographical Research: Biogeosciences, 115, G04037.
Collins, A.M., Coughlin, D., Miller, J., Kirk, S. 2015. The Production of Quick Scoping Reviews and Rapid Evidence Assessments: A How to Guide, NERC/DEFRA.
Cosgrove, P. 2004. The Burning Issue: a briefing paper on fire and the natural heritage. CNPA Briefing Paper No. 3.
Curlevski, N.J.A., Artz, R.R.E., Anderson, I.C. and Cairney, J.W.G. 2011. Response of soil microbial communities to management strategies for enhancing Scots pine (Pinus sylvestris) establishment on heather (Calluna vulgaris) moorland. Plant and Soil, 339, 413–424.
Davies, M.G. 2005. Fire Behaviour and Impact on Heather Moorland. PhD Thesis. University of Edinburgh.
Davies, M.G., Gray, A., Hamilton, A. and Legg, C.J. 2008. The future of fire management in the British uplands. The International Journal of Biodiversity Science and Management, 4(3), 127–147.
Davies, G.M. and Legg, C.J. 2008. The effect of traditional management burning on lichen diversity. Applied Vegetation Science, 11, 529–538.
Davies, G.M., Legg, C.J., Smith, A.A. and MacDonald, A.J. 2009. Rate of spread of fires in Calluna vulgaris-dominated moorlands. Journal of Applied Ecology, 46(5) 1054–1063.
Davies, G.M., Smith, A.A., MacDonald, A.J., Bakker, J.D. and Legg, C.J. 2010. Fire intensity, fire severity and ecosystem response in heathlands: factors affecting the regeneration of Calluna vulgaris. Journal of Applied Ecology, 47, 356–365.
Davies, M.G. and Legg, C.J. 2011. Fuel moisture thresholds in the flammability of Calluna vulgaris. Fire Technology, 47, 421–436.
Davies, G.M., Gray, A., Rein, G. and Legg, C.J. 2013. Peat consumption and carbon loss due to smouldering wildfire in a temperate peatland. Forest Ecology and Management, 308, 169–177.
Davies, G.M. and Legg, C.J. 2016. Regional variation in fire weather controls the reported occurrence of Scottish wildfires. PeerJ, 4, e2649.
Davies, G.M., Domènech, R., Gray, A. and Johnson, P.C.D. 2016a. Vegetation structure and fire weather influence variation in burn severity and fuel consumption during peatland wildfires. Biogeosciences, 13, 389–398.
Davies, G.M., Kettridge, N., Stoof, C.R., Gray, A., Ascoli, D., Fernandes, P.M., Marrs, R., Allen, K.A., Doerr, S.H., Clay, G.D., McMorrow, J. and Vandvik, V. 2016b. The role of fire in UK peatland and moorland management: the need for informed, unbiased debate. Philosophical Transactions of the Royal Society B, 371, 20150342.
Davies, G.M., Kettridge, N., Stoof, C.R., Gray, A., Marrs, R., Ascoli, D., Fernandes, P.M., Allen, K.A., Doerr, S.H., Clay, G.D., McMorrow, J. and Vandvik, V. 2016c. Informed debate on the use of fire for peatland management means acknowledging the complexity of socio-ecological systems. Nature Conservation, 16, 59–77.
Davies, G.M., Legg, C.J., Smith, A.A. and MacDonald, A. 2019. Development and participatory evaluation of fireline intensity and flame property models for managed burns on Calluna-dominated heathlands. Fire Ecology, 15, 30.
Douglas, D.J.T., Beresford, A., Selvidge, J., Garnett, S., Buchanan, G.M., Gullett, P. and Grant, M.C. 2017. Changes in upland bird abundances show associations with moorland management. Bird Study, 64, 242–254.
Evans, C.D., Malcolm, I.C., Shilland, E.M., Rose, N.L., Turner, S.D., Crilly, A., Norris, D., Granath, G. and Monteith, D.T. 2017. Sustained biogeochemical impacts of wildfire in a mountain lake catchment. Ecosystems, 20, 813–829.
Farage, P., Ball, A., McGenity, T.J., Whitby, C. and Pretty, J. 2009. Burning management and carbon sequestration of upland heather moorland in the UK. Australian Journal of Soil Research, 47(4), 351–361.
Finlayson, A., Marchant, B.P., Whitbread, K., Hughes, L. and Barron, H.F. 2021. Estimating organic surface horizon depth for peat and peaty soils across a Scottish upland catchment using linear mixed models with topographic and geological covariates. Soil Use and Management, 37, 628–639.
Fyfe, R.M., Coombe, R., Davies, H. and Parry, L. 2014. The importance of sub-peat carbon storage as shown by data from Dartmoor, UK. Soil Use and Management, 30, 23–31.
Gagkas, Z., Campbell, G., Owen, J. and Davies, M. 2022. Provision of analyses of Scottish Fire and Rescue Service (SFRS) Incident Reporting System (IRS) data in relation to wildfire incident. Report Prepared for Scottish Government, March 2022.
Garnett, M.H., Ineson, P. and Stevenson, A.C. 2000. Effects of burning and grazing on carbon sequestration in a Pennine blanket bog, UK. The Holocene, 10(6), 729–736.
Gatis, N., Luscombe, D.J., Carless, D., Parry, L.E., Fyfe, R.M., Harrod, T.R., Brazier, R.E. and Anderson, K. 2019. Mapping upland peat depth using airborne radiometric and lidar survey data. Geoderma, 335, 78–87.
Gimingham, C.H. 1988. A reappraisal of cyclical processes in Calluna heath. Vegetatio, 77, 61– 64.
Glaves, D.J., Crowle, A.J.W., Bruemmer, C. and Lenaghan, S.A. 2020. The causes and prevention of wildfire on heathlands and peatlands in England. Natural England Evidence Review NEER014.
Glaves, D.J., Morecroft, M., Fitzgibbon, C., Leppitt, P., Owen, M. and Phillips, S. 2013. The effects of managed burning on upland peatland biodiversity, carbon and water. Natural England Evidence Review, NEER004.
Grau-Andrés, R., Gray, A. and Davies, G.M. 2017. Sphagnum abundance and photosynthetic capacity show rapid short-term recovery following managed burning. Plant Ecology and Diversity, 10, 353–359.
Grau-Andrés, R., Davies, M.G., Gray, A., Scott, E.M. and Waldron, S. 2018. Fire severity is more sensitive to low fuel moisture content on Calluna heathlands than on peat bogs. Science of the Total Environment, 616–617, 1261–1269.
Grau-Andrés, R., Gray, A., Davies, M.G., Scott, M.E. and Waldron, S. 2019. Burning increases post-fire carbon emissions in a heathland and a raised bog, but experimental manipulation of fire severity has no effect. Journal of Environmental Management, 233, 321–328.
Gray, A., Davies, G.M., Domenech, R., Taylor, E. and Levy, P.E. 2021. Peatland wildfire severity and postfire gaseous carbon fluxes. Ecosystems, 24, 713–725.
Hancock, M., Egan, S., Summers, R., Cowie, N., Amphlett, A., Rao, S. and Hamilton, A. 2005. The effect of experimental prescribed fire on the establishment of Scots pine Pinus sylvestris seedlings on heather Calluna vulgaris moorland. Forest Ecology and Management, 212, 199–213.
Hancock, M.H., Amphlett, A., Proctor, R., Dugan, D., Willi, J., Harvey, P. and Summers, R.W. 2011. Burning and mowing as habitat management for capercaillie Tetrao urogallus: An experimental test. Forest Ecology and Management, 262, 509–521.
Harper, A.R., Doerr, S.H., Santin, C., Froyd, C.A. and Sinnadurai, P. 2018. Prescribed fire and its impacts on ecosystem services in the UK. Science of the Total Environment, 624, 691–703.
Harris, M.P.K., Allen, K.A., McAllister, H.A., Eyre, G., Le Duc, M.G. and Marrs, R.H. 2011. Factors affecting moorland plant communities and component species in relation to prescribed burning. Journal of Applied Ecology, 48, 1411–1421.
Haysom, K.A. and Coulson, J.C. 1998. The Lepidoptera fauna associated with Calluna vulgaris: effects of plant architecture on abundance and diversity. Ecological Entomology, 23, 377–385.
Heinemeyer, A., Vallack, H.W., Morton, P.A., Pateman, R., Dytham, C., Ineson, P., McClean, C., Bristow, C. and Pearce-Higgins, J.W. 2019. Restoration of heather-dominated blanket bog vegetation on grouse moors for biodiversity, carbon storage, greenhouse gas emissions and water regulation: comparing burning to alternative mowing and uncut management. Final Report DEFRA Project BD5104. DEFRA.
Hochkirch, A. and Adorf, F. 2007. Effects of prescribed burning and wildfires on Orthoptera in Central European peat bogs. Environmental Conservation, 34, 225–235.
Holden, J., Wearing, C., Palmer, S., Jackson, B., Johnston, K. and Brown, L.E. 2013. Fire decreases near-surface hydraulic conductivity and macropore flow in blanket peat. Hydrological Processes, 28(5), 2868–2876.
Holden, N.M. and Connolly, J. 2011. Estimating the carbon stock of a blanket peat region using a peat depth inference model. Catena, 86(2), 75–85.
Jaffé, R., Ding, Y., Niggemann, J., Vähätalo, A.V., Stubbins, A., Spencer, R.G.M., Campbell, J. and Dittmar, T. 2013. Global charcoal mobilization from soils via dissolution and riverine transport to the oceans. Science, 340, 345–347.
Johnston, K. and Robson, B.J. 2015. Experimental effects of ash deposition on macroinvertebrate assemblages in peatland streams. Marine and Freshwater Research, 69, 1681–1691.
Kayll, A.J., 1966. Some characteristics of heath fires in North-East Scotland. Journal of Applied Ecology, 3(1), 29- 40.
Keaney, A., McKinley, J., Robinson, M. and Ruffell, A. 2013. Spatial statistics to estimate peat thickness using airborne radiometric data. Spatial Statistics, 5, 3–24.
Keeley, J. (2009). Fire intensity, fire severity and burn severity: A brief review and suggested usage. International Journal of Wildland Fire. 18. 116–126.
Kelly, R., Montgomery, I.W. and Reid, N. 2018. Differences in soil chemistry remain following wildfires on temperate heath and blanket bog sites of conservation concern. Geoderma, 315, 20–26.
Legg, C., Bruce, M. and Davies, G.M. 2006. Country Report for the United Kingdom. International Forest Fire News, 34, 94–98.
Legg, C., Davies, G.M. and Gray, A. 2010. Comment on: ‘Burning management and carbon sequestration of upland heather moorland in the UK’. Australian Journal of Soil Research, 48(1), 100–103.
Legg, C. and Davies, G.M. 2009. What determines fire occurrence, fire behaviour and fire effects in heathlands? Proceeding of the 10th National Heathlands Conference – Managing Heathlands in the Face of Climate Change. Natural England Commissioned Report, NECR014. Pages 45–55.
Legg, C and Davies, M. 2007. A Fire Danger Rating System for Vegetation Fires in the UK. The FireBeaters Project Phase 1 Final Report.
Lee, H., Alday, J.G., Rose, R.J., O’Reilly, J. and Marrs, R.H. 2013a. Long-term effects of rotational prescribed burning and low-intensity sheep grazing on blanket-bog plant communities. Journal of Applied Ecology, 50, 625–635.
Lee, H., Alday, J.G., Rosenburgh, A., Harris, M., McAllister, H. and Marrs, R.H. 2013b. Change in propagule banks during prescribed burning: A tale of two contrasting moorlands. Biological Conservation, 165, 187–197.
Li, P., Irvine, B., Holden, J. and Mu, X. 2017. Spatial variability of fluvial blanket peat erosion rates for the 21st Century modelled using PESERA-PEAT. Catena, 150, 302–316.
Littlewood, N.A., Mason, T.H.E., Hughes, M., Jaques, R., Whittingham, M.J. and Willis, G. 2019. The influence of different aspects of grouse moorland management on non-target bird assemblages. Ecology and Evolution, 9, 11089–11101.
Log, T., Thuestad, G., Velle, L.G., Khattri, S.K. and Kleppe, G. 2017. Unmanaged heathland – A fire risk in subzero temperatures? Fire Safety Journal, 90, 62–71.
Luxmoore, R. 2018. The relationship between prescribed burning and wildfires: an analysis of wildfire occurrence in the Scottish uplands. National Trust for Scotland.
Mallik, A.U. and FitzPatrick, E.A. 1996. Thin section studies of Calluna heathland soils subject to prescribed burning. Soil Use and Management, 12, 143–149.
Maltby, E., Legg, C. and Proctor, M.C.F. 1990. The ecology of severe moorland fire on the North York Moors: Effects of the 1976 fires and subsequent vegetation and surface development. Journal of Ecology, 78(2), 490–518.
Marchant, B.P. 2021. Using remote sensors to predict soil properties: Radiometry and peat depth in Dartmoor, UK. Geoderma, 403, 115232.
Martin, S. 2018. An assessment of wildfires on Darwen and Turton Moors, Lancashire 1995–2017. Consultancy report to D Livesey, Belmont, Lancashire.
McMorrow, J.M, Lindley, S.J., Aylen, J., Cavan, G., Albertson, K. and Boys, D. 2009. Moorland wildfire risk, visitors and climate change: patterns, prevention and policy. In: Bonn, A., Allot, T., Hubacek, K. and Stewart, J. eds. 2009. Drivers of Environmental Change in Uplands. Routledge, Chapter 23, pp. 404–431.
Milligan, G., Rose, R.J., O’Reilly, J. and Marrs, R.H. 2018. Effects of rotational prescribed burning and sheep grazing on moorland plant communities: Results from a 60-year intervention experiment. Land Degradation and Development, 29, 1397–1412.
Minasny, B., Berglund, O., Connolly, J., Hedley, C., de Vries, F., Gimona, A., Kempen, B., Kidd, D., Lilja, H., Malone, B., McBratney, A., Roudier, P., O’Rourke, S., Rudiyanto., Padarian, J., Poggio, L., ten Caten, A., Thompson, D., Tuve, C. and Widyatmanti, W. 2019. Digital mapping of peatlands – A critical review. Earth-Science Reviews, 196, 102870.
Morton, P.A. and Heinemeyer, A. 2019. Bog breathing: the extent of peat shrinkage and expansion on blanket bogs in relation to water table, heather management and dominant vegetation and its implications for carbon stock assessments. Wetlands Ecology and Management, 27, 467–482.
NatureScot. 2021. Peat depth and peatland condition survey guidance. Peatland Action, NatureScot.
Newey, S., Fielding, D., Miller, D.G., Matthews, K.B. and Thomson, S. 2020. Biodiversity considerations on grouse moors. Part 4 of Research to assess socioeconomic and biodiversity impacts of driven grouse moors and to understand the rights of gamekeepers. Report to the Scottish Government.
Noble, A., O’Reilly, J., Glaves, D.J., Crowle, A., Palmer, S.M. and Holden, J. 2018a. Impacts of prescribed burning on Sphagnum mosses in a long-term peatland field experiment. PloS ONE, 13(11), e0206320.
Noble, A., Palmer, S.M., Glaves, D.J., Crowle, A., Brown, L.E. and Holden, J. 2018b. Prescribed burning, atmospheric pollution and grazing effects on peatland vegetation composition. Journal of Applied Ecology, 55, 559–569.
Noble, A., Crowle, A., Glaves, D.J., Palmer, S.M. and Holden, J. 2019a. Fire temperatures and Sphagnum damage during prescribed burning on peatlands. Ecological Indicators, 103, 471–478.
Noble, A., Palmer, S.M., Glaves, D.J., Crowle, A., Holden, J. 2019b. Peatland vegetation change and establishment of re-introduced Sphagnum moss after prescribed burning. Biodiversity Conservation, 28, 939–952.
Parry, L.E., Charman, D.J. and Noades, J.P.W. 2012. A method for modelling peat depth in blanket peatlands. Soil Use and Management, 28, 614–624.
Parry, L.E., West, L.J., Holden, J. and Chapman, P.J. 2014. Evaluating approaches for estimating peat depth. Journal of Geophysical Research: Biogeosciences, 119, 567–576.
Poggio, L. and Gimona, A. 2014. National scale 3D modelling of soil organic carbon stocks with uncertainty propagation – An example from Scotland. Geoderma, 232–234, 284–299.
Ramchunder, S.J., Brown, L.E. and Holden, J. 2009. Environmental effects of drainage, drain-blocking and prescribed vegetation burning in UK upland peatlands. Progress in Physical Geography: Earth and Environment, 33(1), 49–79.
Ramchunder, S.J., Brown, L.E. and Holden, J. 2013. Rotational vegetation burning effects on peatland stream ecosystems. Journal of Applied Ecology, 50(3), 636–648.
Reed, M.S., Bonn, A., Slee, W., Beharry-Borg, N., Birch, J., Brown, I., Burt, T.P., Chapman, D., Chapman, P.J., Clay, G.D., Cornell, S.J., Fraser, E.D.G., Glass, J.H., Holden, J., Hodgson, J.A., Hubacek, K., Irvine, B., Jin, N., Kirkby, M.J., Kunin, W.E., Moore, O., Moseley, D., Prell, C., Price, M.F., Quinn, C.H., Redpath, S., Reid, C., Stagl, S., Stringer, L.C., Termansen, M., Thorp, S., Towers, W. and Worrall, F. 2009. The future of the uplands. Land Use Policy, 26(1), S204–S216.
Robertson, G.S., Newborn, D., Richardson, M. and Baines, D. 2017. Does rotational heather burning increase red grouse abundance and breeding success on moors in Northern England? Wildlife Biology, 2017(SP1).
Rodwell, J.S. (ed.) 1991. British Plant Communities. Volume 2: Mires and Heaths. Cambridge: Cambridge University Press.
Rosenburgh, A., Alday, J.G., Harris, M.P.K., Allen, K.A., Connor, L., Blackbird, S.J., Eyre, G. and Marrs, R.H. 2013. Changes in peat chemical properties during post-fire succession on blanket bog moorland. Geoderma, 211–212, 98–106.
RSPB. 2021. How to prevent nature and carbon going up in smoke: Licensing Muirburn. RSPB
Sanderson, R., Newton, S. and Selvidge, J. 2020. Effects of vegetation cutting on invertebrate communities of high conservation value Calluna upland peatlands. Insect Conservation and Diversity, 13, 239–249.
Santana, V.M., Alday, J.G., Lee, H., Allen, K.A. and Marrs, R.H. 2016. Modelling carbon emissions in Calluna vulgaris-dominated ecosystems when prescribed burning and wildfires Interact. PloS ONE, 11(11), e0167137.
Scotland’s Moorland Forum. 2017a. The Muirburn Code – Management of moorland by burning and cutting. NatureScot and Scottish Government.
Scotland’s Moorland Forum. 2017b. The Muirburn Code 2017 – Supplementary Information 2 – Offences & Legislation. NatureScot and Scottish Government.
Scotland’s Moorland Forum. 2017c. The Muirburn Code 2017 – Supplementary Information 4 – Fire Behaviour and Effects, Wildfire and Contingency Planning. NatureScot and Scottish Government.
Scottish Executive. No Date. Prescribed Burning on Moorland – Supplement to the Muirburn Code: A Guide to Best Practice. Scottish Executive.
Scottish Government. 2013. Fire and Rescue Service Wildfire Operational Guidance. ISBN 9781782564980.
Scottish Government, Scottish Natural Heritage, SEPA. 2017. Peatland Survey. Guidance on Developments on Peatland, on-line version only.
Shepherd, H.E.R., Catford, J.A., Steele, M.N., Dumont, M.G., Mills, R.T.E., Hughes, P.D.M. and Robroek, J.M. 2021. Propagule availability drives post-wildfire recovery of peatland plant communities. Applied Vegetation Science, 243(3), e12608.
Smith, A.A., Redpath, S.M., Campbell, S.T. and Thirgood, S.J. 2001. Meadow pipits, red grouse and the habitat characteristics of managed grouse moors. Journal of Applied Ecology, 38(2), 390–400.
Stewart, G.B., Glendell, M., McMorran, R., Troldborg, M., Gagkas, Z., Ovando, P., Roberts, M., Maynard, C., Williams, A., Clay, G. and Reed, M.S. 2021. Uplandia: making better policy in complex upland systems – Final Report. Defra and Natural England.
Strachan, I.M. 2017. Manual of terrestrial EUNIS habitats in Scotland. Version 2. Scottish Natural Heritage Commissioned Report No. 766.
Taylor, E.S., Levy, P.E. and Gray, A. 2017. The recovery of Sphagnum capillifolium following exposure to temperatures of simulated moorland fires: a glasshouse experiment. Plant Ecology and Diversity, 10, 77–88.
Taylor, A.F.S., Bruce, M., Britton, A.J., Owen, I.J., Gagkas, Z., Pohle, I., Fielding, D. and Hadden, R. 2021. Fire Danger Rating System (FDRS) Report. James Hutton Institute and University of Edinburgh.
Tharme, A.P., Green, R.E., Baines, D., Bainbridge, I.P. and O'Brien, M. 2001. The effect of management for red grouse shooting on the population density of breeding birds on heather-dominated moorland. Journal of Applied Ecology, 38(2), 439–457.
Walker, L.R., Wardle, D.A., Bardgett, R.D. and Clarkson, B.D. 2010. The use of chronosequences in studies of ecological succession and soil development. Journal of Ecology, 98(4), 725–736.
Ward, S.E., Bardgett, R.D., McNamara, N.P., Adamson, J.K. and Ostle, N.J. 2007. Long-term consequences of grazing and burning on northern peatland carbon dynamics. Ecosystems, 10, 1069–1083.
Ward, S.E., Ostle, N.J., Oakley, S., Quirk, H., Stott, A., Henrys, P.A., Scott, A. and Bardgett, R.D. 2012. Fire accelerates assimilation and transfer of photosynthetic carbon from plants to soil microbes in a northern peatland. Ecosystems, 15, 1245–1257.
Werritty, A. 2019. Grouse Moor Management Review Group. Report to the Scottish Government.
Werritty, A., Pakeman, R.J., Shedden, C., Smith, A. and Wilson, J.D. 2015. A Review of Sustainable Moorland Management. Report to the Scientific Advisory Committee of Scottish Natural Heritage.
Whitehead, S., Weald, H. and Baines, D. 2021. Post-burning responses by vegetation on blanket bog peatland sites on a Scottish grouse moor. Ecological Indicators, 123, 107336.
Whitehead, S.C. and Baines, D. 2018. Moorland vegetation responses following prescribed burning on blanket peat. International Journal of Wildland Fire, 27(10), 658–664.
Worrall, F., Armstrong, A. and Adamson, J.K. 2007. The effects of burning and sheep-grazing on water table depth and soil water quality in a upland peat. Journal of Hydrology, 339, 1–14.
Worrall, F., Bell, M.J. and Bhogal, A. 2010a. Assessing the probability of carbon and greenhouse gas benefit from the management of peat soils. Science of the Total Environment, 13, 2657–2666.
Worrall, F., Clay, G., Marrs, R. and Reed, M.S. 2010b. Impacts of Burning Management on Peatlands. Commissioned Report for the IUCN UK Peatland Programme’s Commission of Inquiry on Peatlands, IUCN.
Worrall, F., Rowson, J. and Dixon, S. 2013a. Effects of managed burning in comparison with vegetation cutting on dissolved organic carbon concentrations in peat soils. Hydrological Processes, 27(26), 3994–4003.
Worrall, F., Clay, G.D. and May, R. 2013b. Controls upon biomass losses and char production from prescribed burning on UK moorland. Journal of environmental management, 120, 27–36.
Yallop, A.R. and Clutterbuck, B. 2009. Land management as a factor controlling dissolved organic carbon release from upland peat soils 1: Spatial variation in DOC productivity. Science of the Total Environment, 407(12), 3803–3813.
Young, D.M., Parry, L.E., Lee, D. and Ray, S. 2018. Spatial models with covariates improve estimates of peat depth in blanket peatlands. PLoS ONE, 13(9), e0202691.
13. Acknowledgements
The authors would like to thank two independent quality assurance reviewers for comments on the draft manuscript.
Annexes
| EUNIS code | Habitat | EUNIS name | EUNIS level | Annex I name | NVC (* part only) | BAP Priority Habitat(s) | Phase 1 |
|---|---|---|---|---|---|---|---|
| F4.11 | Wet Heath | Northern wet heaths | 4 | Northern Atlantic wet heaths with Erica tetralix | M15* M16* | UH, LH | D2 |
| F4.13 | Wet Heath | [Molinia caerulea] wet heaths | 4 | * | M25* | * | B5 |
| F4.21 | Dry Heath | Submontane [Vaccinium] - [Calluna] heaths | 4 | European dry heaths | H12* H18* H21* H22* | UH, LH | D1 |
| F4.22 | Dry Heath | Sub-Atlantic [Calluna] - [Genista] heaths | 4 | European dry heaths | H9 H16* | UH, LH | D1 |
| F4.25 | Dry Heath | Boreo-Atlantic [Erica cinerea] heaths | 4 | European dry heaths | H7* H10* | UH, LH, MC | D1 |
| D1.21 | Blanket bogs | Hyperoceanic low-altitude blanket bogs, typically with dominant [Trichophorum] | 4 | Blanket bogs | M1* M2* M3* M15* M17 M18* M25* | BB | E1.6.1, E1.7, E1.8 |
| D1.22 | Blanket bogs | Montane blanket bogs, [Calluna] and [Eriophorum vaginatum] often dominant | 4 | Blanket bogs | M1* M2* M3* M15* M19* M20* | BB | E1.6.1, E1.7, E1.8 |
| D1.24 | Blanket bogs | Wet bare peat and peat haggs on blanket bogs | 4 | Blanket bogs | BB | E4 | |
| E1.71 | Dry Grassland | [Nardus stricta] swards | 4 | * | U5* | * | B1.1 |
| E1.72x | Dry Grassland | Other Agrostis-Festuca grassland | 5 | * | U4* CG10* | LC LD | B1.1, B1.2 |
| E1.72# | Dry Grassland | Species-rich Nardus grassland, on siliceous substrates in mountain areas | 5 | Species-rich Nardus grassland, on siliceous substrates in mountain areas (and submountain areas in continental Europe) | CG10* CG11 U4* U5c | LC, UC. LD | B3.1 |
| E1.73 | Dry Grassland | [Deschampsia flexuosa] grassland | 4 | * | U2 | * | B1.1 |
| E3.42 | Seasonally wet and wet grasslands | [Juncus acutiflorus] meadows | 4 | * | M23a | PM, UF | B5 |
| E3.52 | Seasonally wet and wet grasslands | Heath [Juncus] meadows and humid [Nardus stricta] swards | 4 | * | U6 U5b | * | B1.1 |
| E5.31 | Bracken | Sub-Atlantic [Pteridium aquilinum] fields | 4 | * | U20 W25* | * | C1 |
(information drawn from Strachan (2017))
| EUNIS code | Habitat | EUNIS name | EUNIS level | Annex I name | NVC (* part only) | BAP Priority Habitat(s) | Phase 1 |
|---|---|---|---|---|---|---|---|
| F2.323 | Subalpine deciduous scrub | Northern British willow brush | 5 | * | Salix aurita scrub non-NVC | WW | A2 |
| F3.15 | Temperate thickets and scrub | [Ulex europaeus] thickets | 4 | * | W23* | * | A2 |
| F3.16#1 | Temperate thickets and scrub | [Juniperus communis] scrub | 5 | Juniperus communis formations on heaths or calcareous grasslands | W19* | UH | A2 |
| D2.22 | Poor fens and soft-water spring mires | [Carex nigra], [Carex canescens], [Carex echinata] fens | 4 | * | M6 M7 | LF, UF | E2.1 |
| D2.2C | Poor fens and soft-water spring mires | Soft water spring mires | 4 | * | M31-33 M35-36 | LF, UF | E2.3 |
| D2.31 | Transition mires and quaking bogs | [Carex lasiocarpa] swards | 4 | Transition mires and quaking bogs | M4* M5* M9* | LF, UF | E3 |
| D2.32 | Transition mires and quaking bogs | [Carex diandra] quaking mires | 4 | Transition mires and quaking bogs | M9* | LF, UF | E3 |
| D2.33 | Transition mires and quaking bogs | [Carex rostrata] quaking mires | 4 | Transition mires and quaking bogs | M4* M5* M8 M9* | LF, UF | E3 |
| D2.33€ | Transition mires and quaking bogs | Transition mires and quaking bogs (Annex I) | 4 | Transition mires and quaking bogs | M4* M5* M8 M9* M29* S27 | LF, UF | E3 |
| D2.37 | Transition mires and quaking bogs | Rhynchospora alba quaking bogs | 4 | Depressions on peat substrates of the Rhynchosporion | M1* M2* | LRB, BB, LF | E1.6.1, E1.6.2 |
| D4.12 | Rich fens, including eutrophic tall-herb fens and calcareous flushes and soaks | [Schoenus ferrugineus] fens | 4 | Alkaline fens | M10* | LF, UF | E2.2 |
| D4.15 | Rich fens, including eutrophic tall-herb fens and calcareous flushes and soaks | [Carex dioica], [Carex pulicaris] and [Carex flava] fens | 4 | Alkaline fens | M10* | LF, UF | E2.2 |
| D4.17 | Rich fens, including eutrophic tall-herb fens and calcareous flushes and soaks | [Carex saxatilis] fens | 4 | Alpine pioneer formations of the Caricion bicoloris-atrofuscae | M12 | UF | E2.2 |
| D4.19 | Rich fens, including eutrophic tall-herb fens and calcareous flushes and soaks | British [Carex demissa] - [Saxifraga aizoides] flushes | 4 | Alkaline fens | M11* | UF | E2.2 |
| D4.1C | Rich fens, including eutrophic tall-herb fens and calcareous flushes and soaks | [Carex rostrata] alkaline fens | 4 | Alkaline fens | M9* | LF, UF | E2.2 |
| D4.1N | Rich fens, including eutrophic tall-herb fens and calcareous flushes and soaks | Hard water spring mires | 4 | Petrifying springs with tufa formation (Cratoneurion) | M37 M38 | LF, UF | E2.3 |
| D4.24 | Basic mountain flushes and streamsides, with a rich arctic-montane flora | British mica flushes | 4 | Alpine pioneer formations of the Caricion bicoloris-atrofuscae | M10* M11* M34 | UF | E2.2 |
| D4.24€ | Basic mountain flushes and streamsides, with a rich arctic-montane flora | Alpine pioneer formations of the Caricion bicoloris-atrofuscae | 4 | Alpine pioneer formations of the Caricion bicoloris-atrofuscae | M10* M11* M12 M34 | UF | E2.2 |
| E4.115# | Alpine and subalpine grasslands | Polytrichastrum-Kiaeria snowbed | 6 | Siliceous alpine and boreal grasslands | U11 | MH | D4 |
| E4.115x | Alpine and subalpine grasslands | Rhytidiadelphus-Deschampsia snowbed | 6 | * | U13b | MH | D4 |
| E4.116 | Alpine and subalpine grasslands | Boreo-alpine [Deschampsia]-[Anthoxanthum] communities | 5 | * | U13a | MH | D4 |
| E4.117 | Alpine and subalpine grasslands | Boreo-alpine herb-rich acid snowbed communities | 5 | Siliceous alpine and boreal grasslands | U14 | MH | D4 |
| E4.12 | Alpine and subalpine grasslands | Boreo-alpine calcicline snow-patch grassland and herb habitats | 4 | Alpine and subalpine calcareous grasslands | CG12 | UC | B3.1 |
| E4.12€ | Alpine and subalpine grasslands | Alpine and subalpine calcareous grasslands | 4 | Alpine and subalpine calcareous grasslands | CG12 CG13* CG14 | UC | B3.1 |
| E4.21 | Alpine and subalpine grasslands | Oroboreal [Carex bigelowii]-[Racomitrium] moss-heaths | 4 | Siliceous alpine and boreal grasslands | U9 U10 | MH | D4 |
| E4.22 | Alpine and subalpine grasslands | Rock pavement lichen communities | 4 | * | non-NVC lichen communities | IR | D3 |
| E4.23 | Alpine and subalpine grasslands | Rock pavement, plateau and summital moss heaths | 4 | * | non-NVC bryophyte communities | IR | D3 |
| E4.32 | Alpine and subalpine grasslands | Oroboreal acidocline grassland | 4 | Siliceous alpine and boreal grasslands | U7 U8 | MH | D4 |
| E4.32€ | Alpine and subalpine grasslands | Siliceous alpine and boreal grasslands | 4 | Siliceous alpine and boreal grasslands | U7 U8 U9 U10 U11 U12 U14 | MH | D4 |
| E5.59 | Subalpine moist or wet tall-herb and fern stands | Oro-boreal tall-herb communities | 4 | Hydrophilous tall herb fringe communities of plains and of the montane to alpine levels | U17 | IR | C2 |
| E5.5B | Subalpine moist or wet tall-herb and fern stands | Alpine and subalpine fern stands | 4 | * | U19, Dryopteris affinis stands | IR | C3.2 |
| E5.5x | Subalpine moist or wet tall-herb and fern stands | Luzula sylvatica-Vaccinium myrtillus tall-herb community | 4 | * | U16 | IR | C3.2 |
| F2.1# | Arctic, alpine and subalpine scrub | Sub-Arctic Salix spp. scrub | 4 | Sub-Arctic Salix spp. scrub | W20 | MH | A2 |
| F2.11 | Arctic, alpine and subalpine scrub | Boreo-alpine acidocline snow-patch [Salix herbacea] scrub | 4 | Siliceous alpine and boreal grasslands | U12 | MH | D4 |
| F2.29#1 | Evergreen alpine and subalpine heath and scrub | [Dryas] mats not on limestone pavement | 5 | Alpine and subalpine calcareous grasslands | CG13* CG14 | UC | B3.1 |
| F2.25 | Evergreen alpine and subalpine heath and scrub | Boreo-alpine and arctic heaths | 4 | Alpine and Boreal heaths | H10* H12* H13-15 H16* H17 H18* H19-20 H21* H22* | MH | D1 |
| Authors | Country | Location | Habitat | Methodology | Main Findings |
|---|---|---|---|---|---|
| Albertson et al. (2010) | England | Peak District, Northern England | Peak District National Park in entirety (habitat mosaic) | Probit model. Wildfire data from 1 June 1976 to 31 December 2008 from reports by rangers in the PDNP. Met office weather data. | Does not estimate wildfire occurrence due to muirburn. Does not model muirburn or fuel load. They find that fire weather and recreation control wildfire risk Peak recreation days having the strongest influence. They predict that wildfire risk will increase due to climate change, and peak hazard may switch from Spring to Summer, although changes will be slow to occur. |
| Alday et al. (2015) | England | Moor House NNR, North Pennines, Northern England | Eastern side of Hard Hill. Blanket bog > 50cm deep peat. The vegetation at Moor House can be described as Calluna vulgaris–Eriophorum vaginatum blanket mire (M19) and E. vaginatum blanket and raised mire (M20) communities. | GLM mixed effects. Field data from biomass monitoring in July 2011. 28 subplots. | Managed burning reduces biomass, shorter rotation greater effect. No effect of grazing, either individually or through interaction. Calluna biomass reached an asymptote of 795.87 +/- 80gm-2 approximately 20 years after fire, and vegetation height reaches an asymptote 36.31 +/-0.84 cm approx. 15 years after last fire. |
| Allen et al. (2013) | England | Peak District, Northern England | Moorland
| Age structured matrix model. Data on above ground biomass was obtained from Harris et al. (2011). Chronosequence. | They find that above ground biomass is greater where the rotation interval of prescribed burning is longer. Increasing the frequency of prescribed burning reduces above ground biomass. |
| Arnell et al. (2021) | UK | UK | Modelling Study. Applies Met Office Fire Severity Model (MOFSI) to UKCP18 climate projections to forecast wildfire danger days under potential climate change scenarios at six representative UK sample sites. MOFSI is based on indices from the Canadian Fire Weather Information System. | Predicts increasing risk of summer wildfires. Greatest in the South and East of England but increasing in the North also. Through comparison of heat maps they suggest that the distribution of actual wildfire relates more to land use than fire weather. Fire weather conditions greatest in the South- East of England. Observed wildfires cluster in locations throughout the UK. Significant hotspots in Scotland. | |
| Cosgrove (2004) | Scotland | Badenoch and Strathspey | Diverse | Summary statistics. Data reported to Badenoch and Strathspey Fire Protection Group | "Formal monitoring of wildfires across the Cairngorms is patchy, with only large fires and ‘call-outs’ usually recorded. The attending Fire Service tries to determine the cause of each fire with the evidence available, but this is not always possible. Consequently, the reason for a fire starting is known as the ‘supposed cause’. The supposed cause of 13 out of 14 (93%) wildfires tackled in Badenoch and Strathspey in 2003 was human action. 29% were started by muirburn activities that got out of control, destroying c1135ha of habitat. The supposed cause of an additional 29% of fires was by discarded cigarettes and camp/bonfires destroying c433ha of habitat" |
| Clay et al. (2015) | England | Emblehope moor and Ray Demesne moor, Northumberland
| Emblehope moor. Ray Demesne moor. Both sites peat, 50cm or greater. Dominant vegetation types across both sites include Calluna vulgaris, Eriophorum vaginatum, Eriophorum angustifolium, Sphagnum spp., Polytrichum spp., Molinia caerulea and Rubus chamaemorus. | Linear regression. Data from field survey at two sites along 10 year chronosequence.
| They find that burning reduces biomass. The average biomass of burned plots less than half of that for unburned. Their regression estimates that 80% of above ground biomass is lost during burns. |
| Davies (2005) | Scotland | Crubenmore | H12, species poor Calluna. Glacial till. Peaty deposits. | GLM | Variously fuel group, burn day, rate of spread and fireline intensity were not found to have significant effects on; moss - litter consumption; time above 100oC; the slope of the temperature regression; or height change of plant temperature measure. There were significant differences in total consumption between fuel loadings (low, med, high). There was no significant difference in fuel consumption with burn day. There was a significant difference in ground surface heating (above 50oC) between burn days. The duration of temperature above 50oC did not differ significantly between fuel groups, or with changes in rate of spread or fireline intensity. |
| Davies et al. (2009) | Scotland | Crubenmore Estate near Dalwhinnie and Black Hill, Pentlands | H12, Calluna vulgaris dominated upland heath | GLM regression used to investigate differences in fuel structure between growth phases of Calluna (Building, Late Building, Mature. Redundancy Analysis used to explore the difference within and between control variables. Experimental data from 27 controlled burns. | They find considerable variation in median height and fuel load across the three development classes of Calluna, building, late building and mature. Modelling found that rate of spread determined primarily by wind speed vegetation height, and live fuel moisture content. Non-linear relationship between wind speed and rate of spread, the effect greater in mature plots. |
| Davies et al. (2010) | Scotland | Carn Dhomhnuill Bhan, near Cairngorms | Glacial till. Substantial peat deposits. H12. Calluna vulgaris dominated upland heath. | Principal components analysis. Data from 15 experimental fires, burned October to April in plots selected across three developmental phases of Calluna. | Fireline intensity varied considerably, ranging from 75 to 530 kW m-1 in Building, 548 to 1433 kW m-1 in Late-building and 347 to 3389 kW m-1 in Mature stand Intensity varied significantly between building and mature classes but not for others. |
| Davies and Legg (2011) | Scotland | Crubenmore Estate near Dalwhinnie and Castlelaw Hill, Pentlands | Calluna vulgaris dominated, underlain by deep mat of pleurocarpous mosses. | GLM model in Minitab used to determine whether significant differences existed between, fires that were sustaining and fires that failed to spread. Experimental data from twenty ignition attempts. | They find that moisture content of Calluna in the lower canopy, and the moss/litter layer controls both ignition and rate of spread. The find a critical threshold of 60- 70% FMC in the lower canopy for sustaining fires. Above 70%, both spot and line ignitions failed to spread. Below 60% fires developed rapidly. |
| Davies et al. (2016a) | UK | Five sites following wildfire. Three England, two Scotland. | 1) Anglezarke N. England. Deep peat. Calluna vulgaris, blanket mire M19. 2) Mardsen N England. Deep peat. Calluna vulgaris, blanket mire M19. 3) Loch Doon SW Scotland. Shallow peat. Molinia. M25a. 4) Wainstalls N England. Deep peat. Calluna vulgaris. M19. 5) Finzean NE Scotland. Calluna vulgaris H12. | Generalized Linear Modelling (GLM). Data from post hoc assessment of five different wildfire sites, burnt during spring 2011 and spring 2012. Burn severity was assessed using an adapted Composite Burn Index (CBI). CBI uses visual estimate of impact on five fuel strata. Fuel consumption was estimated from comparison of burnt and unburnt paired plots. | The find that the severity of burns, can be explained by reduced moisture content in the ground fuel layer (moss-litter). They find substantial variation in burn severity both within and between fires. On average, mean pCBI (composite burn index) varied 1.6 fold between wildfires but up to 1.7 fold within fires. |
| Davies and Legg (2016) | Scotland | Modelling of four Scottish Fire and Rescue Service regions. | Diverse habitats. Noted that SFRS data frequently lacks vegetation information. | Logistic regression. Wildfire data obtained from four Scottish Fire and Rescue Services, covering 1st Jan 2003 to 15th March 2007. Fire Weather Index (FWI) codes were calculated for the date and location using Met Office data. | Does not estimate wildfire occurrence due to muirburn. Does not model muirburn or fuel load. They find regional variation in Scottish wildfire. Greatest in Highland in total count by region and per capita basis. On per area basis most common in Lothian and Borders. They find that two FWI indices are able to predict wildfire "tolerably well". |
| Davies et al. (2019) | Scotland | Crubenmore Estate near Dalwhinnie and Castlelaw Hill, Pentlands | H12 Calluna vulgaris. Most plots unerlain by deep carpet pleurocarpous moss. | Best subsets regression. Data from 27 controlled burns. Intensity calculated as product of heat of combustion, fuel consumption and fire rate of spread per Byram (1959). | Fireline intensity and flame height increased significantly across age related Calluna phases • fireline intensity could be adequately modelled from fuel height and wind speed • model improved by accounting for live fuel moisture content |
| Farage et al. (2009) | England | Mossdale Moor, upper Wensleydale North Yorkshire | Not specified | Comparison of means. Biomass data from field surveys in 2003 and 2004. Burn consumption estimated by comparison of burnt and unburnt plots. N = 8. | • 16 +/- 4% is estimated to have been consumed in the burn in 2003 • and 24 +/-5% in 2004 |
| Gagkas et al. (2021) | Scotland | Nationwide study of Scotland | Diverse habitats. Analysis by Level 3 property type categories. | Comparative statistics at National and Local Authority level assessing: Temporality, Fuel Types, Ignition motives, and Burnt area size. Cluster Analysis at LA level. Wildfire data obtained from FRS 1st April 2009 to 31st December 2020. Filtered based on FRS codes and visual inspection of satellite imagery. | They identify 9,725 wildfires, of which motive of ignition is recorded for a subset of 919 incidents (and source of ignition for 927). The categories summarised in the report do not detail the proportion of such that were caused by muirburn. Other intentional burn out of control: 100 counts (10.8% total wildfires for which cause of ignition recorded) Other: 129 counts (14%). |
| Glaves et al. (2020) | National Study of England | England | Not stated | Summary Statistics. Data amalgamated from data submitted to Natural England from; Dorset county Council; Lancashire Fire & Rescue Services; Moors for the Future; Peak District National Park. | Cause of ignition is available for 12 % of overall incidents recorded. Within this subset, the main causes were ‘camp fires’ (49%), land management burns (15%), barbeques (10%), and ‘reignited’ fires and military training (both 5%) with no other individual causes greater than 3%. They note differences between lowlands and uplands. On the basis of N = 62 upland wildfires for which cause is recorded, the majority were assigned to land manager burns (68%), followed by camp fires (9%) and barbeques (8%). They note "Care is needed in interpreting these findings given the small proportion of overall fires where a specific cause was assigned and potential bias and subjectivity in these assessments (para. 5.3), and the relatively small number in the uplands (62, 10% of all upland fires) which emphasises the need for better recording of information about the causes of wildfire incidents." |
| Grau-Andrés et al. (2018) | Scotland | Glen Tanar and Braehead Moss | Glen Tanar, dry heath peaty podzols. Braehead Moss, raised bog with deep > 1.5m peat. Both sites > 85% mature Calluna vulgaris and > 63% cover of pleurocarpous mosses. | GLM mixed effects. Data from 19 experimental fires. Fuel load and structure assessed by fuel rule and visual inspection prior to burn. Fire severity assessed by two metrics: consumption of moss- litter layer, and fire induced soil heating. | They found that the moisture content of the moss- litter layer controls fire severity, across both measures of fire severity. Measured as consumption of the moss-litter layer during fire, they found no significant difference between the dry heath site and the raised bog site. Measured as soil heating however, they found a substantial difference in the effect of low FMC on soil heating. Following drought treatment, mean time above 50oC at the soil surface increased from 34s to 10min at the dry heath site, with peak temperature reaching 158oC. While at the raised bog site, average maximum temperature never exceeded 15oC. |
| Legg et al. (2006) | Scotland | Scotland | Not stated | Summary Statistics. Questionnaire data from survey administered by the Game Conservancy Trust and Scottish Gamekeepers Association. Responses from 42 Estates in 2003. | Seventeen wildfires were reported from thirteen estates, averaging 150ha. Cause was recorded as: 9 due to escaped managed burn, two were recorded as having been started by crofters, three as arson, two as accidental, and one unknown. For context they note that "Twenty of the 42 respondents in the survey reported active management by burning during 2003 with an estimated 4300 fires; one estate estimated that they had burned 900-1000 fires during the season. It is likely that this rather small sample of estates is biased towards those who experienced wildfires as they are perhaps more likely to respond to the questionnaire. The survey also targeted moorland estates managed for game and does not include information about the many wildfires that occurred on public land or land managed for other purposes" |
| Log et al. (2017) | Norway | Flatanger and Froya | Calluna vulgaris dominated heath | After the fact assessment of two severe wildfires, account of wildfires and discussion of pre-fire conditions. | Giving an after the fact assessment of the conditions leading up to two large wildfires in Norway, Log et al. (2017) argue that a lack of controlled burning during the fifty years prior had led to the build up of high fuel load, in the form of late developmental phase mature - degenerate Calluna and increased abundance of fire prone tree species. Combined with particular climatic conditions, below zero temperatures in the three-weeks preceding, low relative humidity and cold air from the mountains had given the conditions for adiabatic heating leading, freeze drying Calluna and resulting in winter desiccation. They conclude these conditions made the area susceptible to ignition and sustained burning and that a lack of fuel load management had been an enabling condition for the fires, which on the basis of property damage were the most severe in Norway since the World War II. |
| Maltby et al. (1990) | England | North York Moors Northern England | Calluna dominated. Variable sandy stony thin peaty hummus to peat greater than 1m deep. | After the fact assessment of wildfire outcome. Ongoing vegetation and soil survey. | In late summer of a drought year in 1976, a significant wildfire affected an area of approx 600ha. The fire burned deeply into blanket peats and destroyed thinner peats, reducing them in extreme cases to ash. the fire resulted in substantial physical and chemical alterations of the soil. Vegetation has been slow to develop. |
| Milligan et al. (2017) | England | Moor House NNR, North Pennines, Northern England | Calluna dominated. M19. 1.5 - 2.5 m peat | Field study. | They find that vegetation structure (height) varied observably between the three burn rotations. For the short (10 year) rotation vegetation height was clustered in the 0- 20cm category, with a marked reduction above. For the 20 year and no burn rotations, vegetation height was more variable, with a peak in the 20 - 30 cm category. |
| Noble et al. (2019) | England | Moor House NNR, North Pennines, Northern England | Hard Hill - not specified in paper | Generalized Linear Modelling (GLM) mixed effects. Data from 16 experimental burns and laboratory testing. | Max temperature at moss surface varied from 33oC to 538oC. Max temp higher in plots with greater Calluna cover. Dwarf shrub cover best predictor of maximum fire temperature R squared = 0.46, p = 0.008. Analysis of cell damage to moss samples found considerable variation. Measured in June, 2- 38% for temperatures under 100oC and consistently high (93 – 100%) for temps above 400oC. Positive linear relationship with temperature. Further laboratory testing of five common species of sphagnum indicated that all five species of sphagnum experienced greater cell damage at higher temperatures. |
| Santana et al. (2016) | UK | Four sites, UK | (1) Kerloch - poorly drained peaty podzols; (2) Moor House - blanket bog > 50cm peat; (3) Howden - blanket bog > 50cm peat; (4) Hartland Moor, Studland Heath and Morden Bog - podzols of low fertility derived from Eocene deposits | Age structured matrix model. Chronosequence data obtained from prior publications.
| • Above ground biomass and litter accumulation patterns differed between sites • Differences not ordered on north-South gradient • Modelling simulations showed predicted above ground biomass increasing in the rotation interval of prescribed burning |
| Taylor et al. (2021) | Scotland | Laboratory study; fuel sampled from Pentland Hills Regional Park Field study; Glensaugh Farm, Scotland | Not detailed | Laboratory study. Field study. | Laboratory study: qualitatively they observe that the fuel moisture content of the fine dead fuel was a dominant factor in determining flame spread rate, and length of flames (energy release). However low fuel moisture content of fine fuel alone was not sufficient to support flame spread (whether or not the flame traversed the entire length of the fuel bed) under all conditions, they find this must be supported by sufficiently low fuel moisture content in the other fuels, particularly the coarser heather and the fine green. They identify fuel moisture thresholds for fire to spread under laboratory conditions. Fine green 47- 65%; fine dead 26- 33%; coarse 54- 60%; moss 84- 135%. Field study: •No discernible trends were observed between the environmental variables of relative humidity, wind speed and temperature, or fuel moisture content and either the mass lost or flame spread rate in these experiments. The FMC values measured in the field for which fire spread was higher than the thresholds determined in the laboratory. |
Whitehead et al. (2021)
| Scotland | Langholm Moor, SW Scotland | Mix of mire communities M18 and M19, peat > 0.5m, average 1.55 m | GLM Regression. | They find that years since burning is a significant predictor of heather cover, with cover being significantly lower in the first seven years relative to the no burn controls. There were significant positive correlations between each pairwise combination of vegetation height, biomass and heather cover (vegetation height and biomass r = 0.79, heather cover and biomass r = 0.80, vegetation height and heather cover r = 0.77, all n = 64, P < 0.001). |
| Worrall et al. (2010a) | England | Peak District | Not stated | Summary Statistics. Methodology not published. | “The Peak District National Park ranger reports between 1976 and 2004 record 341 wildfires. Of the 341 reported wildfires, 41 have an attributed cause and of those 41, ten have been attributed to prescribed burns, i.e. a little under 25% of wildfires with a known cause are due to prescribed burning. However, when the area of the wildfires is considered of the 41 fires with an assigned cause, those due to prescribed burns represented 51% of the burnt area, i.e. fires from prescribed burns appear to have been bigger when they did occur". They note that it has been suggested that there may be a lack of consistency with FRS data recording. "There are data quality issues surrounding the recording of vegetation fires in the UK which makes any analysis difficult. Before the introduction of the Incident Recording System (IRS) in the UK, Fire and Rescue Services often had a “favourite” attributed cause which varied over time (Walker et al., 2009)" |
| Worrall et al. (2013b) | England | Peak District | Calluna. Not specified | Field study. Biomass assessed pre and post management burn. | They estimate average biomass loss during burns to be 75 +/- 9%. |
(information drawn from Strachan (2017))
Annex 4 – Additional critique and review papers relevant to the relationship between muirburn and wildfire
Ashby and Heinemeyer (2020) Prescribed burning impacts on ecosystem services in the British uplands: A methodological critique of the EMBER project
EMBER project does not study wildfire/ burn behaviour/ factors influencing severity/ above ground carbon. Criticisms:
- Chronosequence, assumes similar pre-disturbance conditions.
- Lack of control for variation in climate / elevation between sites both in study design and statistical modelling.
- Suggests peat temperature finding of one study may be due to temp probe exposure to sun.
Davies et al. (2016c) Informed debate on the use of fire for peatland management means acknowledging the complexity of socio-ecological systems
Responds to George Monbiot article which raised three criticisms of a prior paper.
- that they framed their research question too narrowly
- that they made the implicit assumption that moorlands were the right ecosystem for the UK countryside
- that they failed to adequately engage with arguments for a cessation of managed burning.
Ashby and Heinemeyer (2021) A Critical Review of the IUCN UK Peatland Programme's Burning and Peatlands Position Statement
Contests the IUCN position that wildfire risk does not generally apply to wet blanket bog habitat. Questions the evidence base for rewetting as a management strategy to reduce fire risk, arguing that since water tables drop naturally in summer, rewetting cannot be relied upon to eliminate wildfire risk on peat bog. Agrees with IUCN position that wildfire incidence is primarily due to human causes.
Legg et al. (2010) Comment on: 'Burning management and carbon sequestration of upland heather moorland in the UK'
Farage et al. (2009) report biomass remaining after fire as 1262 gm-2. Legg and Davies (2010) note that this is unusually high. In their own study of 26 experimental fires they recorded mean biomass remaining as 277gm-2 with a maximum of 940 gm-2 which occurred in one late mature stand which was observably atypical, and had stems which collapsed close to the ground, un-aerated and remained too damp to burn. Their description of their experimental design suggests that they only collected samples from a single burned and unburned stand in each of the two years. If this is the case then it would be pseudo-replication and would invalidate any statistics to estimate confidence intervals or the estimates of carbon lost due to burning.
Davies et al. (2016b) The role of fire in UK peatland and moorland management: the need for informed, unbiased debate
Argues for the need to distinguish burn severity, not to conflate the outcomes of severe wildfire with managed burning. Notes knowledge gap with respect to true fire regime across UK Moors often assumed 15-25 years on management basis, but likely much longer in many cases. Lack of effective method to determine. Details five factoids which are often repeated but insufficiently evidenced, and agues they should be avoided.
- that regular burning alone increases Calluna dominance
- that fire kills or significantly damages Sphagnum
- that peatlands are particularly sensitive with regard to fire
- that managed burning helps prevent against future wildfires
- how does managed burning affect landscape scale patterns in flammability?
- does it reduce frequency or burn severity of wildfires?
- how many wildfires actually result from managed burning?
- that fire alone contributes to peatland degradation
International Union for Conservation of Nature UK (2020) Position Statement: Burning and Peatlands
Does not directly address the relationship between muirburn and wildfire. Outlines an overall position that managed burning is a contributing factor to peatland degradation and succession to dry heath dominated by Calluna, a situation claimed to be at risk of wildfire. In contrast it is argued that rewetting restores a naturally resilient state that is less susceptible to wildfire. Claims that wildfire risk does not generally apply to wet blanket bog habitat.
Belcher et al. (2021) UK Wildfires and their Climate Challenges
The report aims to assess the magnitude of present and future wildfire risk in the context of changing climate. It provides various definitions of wildfire and enabling conditions including several UK case studies with photos. Discussion of wildfire data relevant to UK. EFFIS satellite based, only counts greater than 30 ha. FRS data from incident reporting system. Discussion of wildfire modelling studies that have been applied to predict future wildfire risk in UK. Discussion of different types and motivations of management burning with examples - includes managed burning to reduce fuel load. Discussion of mechanical vs. fire based approaches to managing fuel loads - does not identify evidence that managed burning in the moorland habitat context defined for the review influences wildfire occurrence, severity, intensity, extent. Gives argument citing controlled burning studies already included in the review that "well-designed prescribed fires should be capable of removing heavy loads of heather, gorse and Molinia without damaging the ground understory or peat soils."
Gazard et al. (2016) Wildfire Policy and Management in England: An evolving response from Fire and Rescue Services Forestry and Cross Party Groups
They note challenges relating to wildfire statistics.
- Fire statistics have allowed wildfires to be spatially evidenced on a national scale only since 2009.
- Wildfires not reported to the fire and rescue services are not recorded.
- The accuracy of the system depends on data entry.
- Consistent long run statistics are still lacking.
- All vegetation fires must now be reported to central government, this includes georeferencing, however as fires are recorded at a single point, source and spatial extent may not always be determined.
- An analytical challenge is that the incident reporting system follows the financial year, which splits the Spring wildfire season into two years.
- English data does not differentiate between small and large fires.
- Scotland’s wildfire operational guidance has a more flexible definition of large wildfires for recording purposes, using any one of five criteria: involving a geographical area of greater than 1 ha; requiring a committed resource of greater than or equal to four fire appliances; requiring resources to be committed for greater than or equal to 6 h; having a sustained flame length of greater than 1.5 m; or presenting a serious threat to life, environment, property and infrastructure.
They claim that challenges arise from a functional fragmentation of the hazard chain.
- There is no national wildfire agency and responsibility rests with the individual fire and rescue services.
- Wildfire is de jure an FRS problem but de facto a land and people management problem that falls within the scope of many agencies and sectors.
Aylen et al. (2007) The best strategy for mitigating moorland wildfire risk. A Report to Moors for the Future.
Discussion paper. Presents no original results.
Offers perspective on factors influencing fire behaviour. Spatially, there are two important influences on the risk of wildfire outbreaks: flammability and ignition sources. Flammability, or vulnerability to ignition, is a function of weather conditions at the time and fuel loading. Fuel loading is related to habitat type and moorland management. Weather conditions are important to fire behaviour, particularly wind speed and direction.
Claims that vulnerability to fire depends on habitat, fuel characteristics and recreation.
- Older heather stands are vulnerable to fire as they have an increased fuel loading.
- Bare peat interspersed with grass and shrub patches are particularly vulnerable to wild fire.
- In contrast, wetter habitats such as cotton grass are far less likely to be the site of wildfire outbreaks.
- Also notes the effect of drying time, heather and grasses are considered to be “one-hour fuels” as they have a very short drying time due to their small particle sizes.
- Further notes the influence of recreation on ignition, claiming that ignition sources are a function of accessibility and attractiveness of habitat to visitors.
Claims that severity is difficult to define. Area burnt is a limited measure. Depth of burn also matters on peat.
Claims that fire intensity relates to width of burn line, as spread accelerates because the head fire is able to burn more vigorously as the width of a fire increases.
Argues for controlled burning in winter to reduce fuel load in subsequent summers, citing McMorrow et al. (2005) claim that managed heather moorlands are less prone to wildfire outbreaks and are a lower risk vegetation type, and that burning to reduce the fuel load is therefore an effective management solution, widely used in the USA and Europe.
Grau-Andrés et al. (2014) Fuel and Climate Controls on Peatland Fire Severity
Chapter in edited book. Reports same results as Grau-Andrés et al., 2018. "Our results show that the M/L layer has a critical ecosystem function by preventing peat from being exposed to significant temperature pulses during burning."
Davies et al. (2008) The future of fire management in the British uplands
Discussion paper. Presents no original results.
Makes claims that large areas of old heather excluded from rotational burning pose a significant fire risk and wildfires in such areas will be more intense and severe and more likely to ignite peat.
Advocates an ecological approach to management burning, considering 1) vegetation structure and species composition 2) Fuel flammability 3) Plant traits in relation to risk and effects of fire 4) The impacts of both individual fires and the overall management regime. Advocates a more variable habitat mosaic, leaving some areas no burn to become mature heather stands, allowing succession to woodland on eroded slopes. Advocates a participatory approach to developing recommendations and regulations around burning.
Legg and Davies (2009) What determines fire occurrence, fire behaviour and fire effects in heathlands?
Offers perspective on factors influencing fire behaviour: Distinguishes rate of spread (speed) and fireline intensity (heat output per unit length). Both determine the difficulty of putting out a fire. Rate of spread is affected partly by windspeed and partly by the structure and moisture content of the fuel. Rate of spread is greater in fine dry fuel, than coarse damp fuel. Structure of the fuel also affects rate of spread. In older stands where gaps in the canopy the fireline may be sporadic. Increased wind speed can cause the fire to jump over gaps. Fast spreading fires consume fuel at a greater rate and therefore rate of spread and fire line intensity are strongly related. Discussion on fire severity vs. intensity. Fire intensity is a measure of the rate of heat output, whereas fire severity refers to the amount of change that the fire causes to the ecosystem. Amount of change is not easily measured on a single scale. One way to consider severity would be to estimate the amount of time it takes for the system to return to previous state. Challenge is that fire may reoccur before it has returned to the previous state. Can lead to successive changes. Different people may view changes in different ways.
| Paper | Experimental design | Geographical coverage | System diversity coverage | Number of burn events | Burn event coverage |
|---|---|---|---|---|---|
| Alday et al. (2015) | Medium | Low | Low | High | Low |
| Clay et al (2015) | Medium | Low | Low | Unclear/NA | Low |
| Farage et al. (2009) | Low | Low | Low | Unclear/NA | Low |
| Worrall et al (2013b) | Low | Low | Low | Unclear/NA | Medium |
| Milligan et al. (2017) | Medium | Low | Low | High | Unclear/NA |
| Taylor et al. (2021) laboratory study | High | Low | Low | Unclear/ NA | Med |
| Taylor et al. (2022) field study | Med | Low | Low | Unclear/ NA | Med |
| Experimental design - High | The study includes both control sites and is replicated |
|---|---|
| Experimental design - Medium | The study includes control sites or has replication |
| Experimental design - Low | The study does not include control sites or replication |
| Geographical coverage - High | The study includes 3 or more contrasting experimental/study sites |
| Geographical coverage - Medium | The study includes 2 contrasting experimental/study sites |
| Geographical coverage - Low | The study includes 1 experimental/study site |
| System diversity coverage - High | The study includes 3 or more contrasting soil or vegetation types (vegetation structure, Calluna phase) |
| System diversity coverage - Medium | The study includes 2 contrasting soil or vegetation types (vegetation structure, Calluna phase) |
| System diversity coverage - Low | The study includes 1 soil/vegetation type (vegetation structure, Calluna phase) |
| Number of burn events - High | The study includes the monitoring of 3 or more sequential burn events |
| Number of burn events - Medium | The study includes the monitoring of 2 or more sequential burn events |
| Number of burn events - Low | The study includes the monitoring of 1 burn event |
| Burn event coverage - High | The study sampling covers pre, during and post burn |
| Burn event coverage - Medium | The study sampling covers 2 time points (pre & post burn; pre & during burn; during & post burn) |
| L Burn event coverage - Low | The study comprises one sampling time point including samples from during or post burn events only |
| Not enough information provided/unclear or Not Applicable | Not enough information is provided, it is unclear, or it is not applicable |
| Reference | Location | Soil | Vegetation | C-related measurements taken | Main outputs relating to carbon balance (Impact) |
|---|---|---|---|---|---|
| Chen et al. (2008) | Moor House NNR, North Pennines, Northern England | Peat (2–3 m deep) | Calluna vulgaris | CH4 oxidation potential (incubation experiment)
|
|
| Clay et al. (2009) | Trout Beck catchment, Moor House NNR, North Pennines, Northern England | Blanket peat | Dominated by Eriophorum spp. (cotton grass), Calluna vulgaris (heather) and Sphagnum spp. (moss) | Soil water dissolved organic carbon |
|
| Clay et al. (2010b) | Trout Beck catchment, Moor House NNR, North Pennines, Northern England | Blanket peat | Dominated by Eriophorum spp. (cotton grass), Calluna vulgaris (heather) and Sphagnum spp. (moss) | Soil water dissolved organic carbon |
|
| Clay et al. (2010c) | Trout Beck catchment, Moor House NNR, North Pennines, Northern England | Blanket Bog | Dominated by Calluna vulgaris and Eriophorum vaginatum. Sphagnum spp. present in some areas | C budget calculated from measured DOC, POC & dissolved CO2 and estimated primary productivity, net ecosystem respiration & methane |
|
| Clay et al. (2012) | Upland Predation Experiment Emblehope Moor in Redesdale Forest and Ray Demesne moor, near Raylees | Deep peat | Calluna vulgaris, Eriophorum vaginatum, Eriophorum angustifolium, Sphagnum spp., Polytrichum spp., Molinia caerulea, and Rubus Chamaemorus | Soil water dissolved organic carbon (DOC) |
|
| Clay et al. (2015) | Emblehope Moor and Redesdale Forest in Northumberland | Deep peat | Calluna vulgaris, Eriophorum vaginatum, Eriophorum angustifolium, Sphagnum spp., Polytrichum spp., Molinia caerulea | Vegetation C concentration |
|
| Garnett et al. (2000) | Moor House NNR, North Pennines, Northern England | Peat | * | Soil carbon concentration – as assumed to be 50% dry mass |
|
| Grau-Andrés et al. (2019) | Glen Tanar and Braehead Moss. Scotland, UK | Upland dry heath (Glen Tanar) actively managed for red grouse and a lowland raised bog (Braehead Moss) historically experienced low intensity disturbance | Dominated by dense Calluna vulgaris (and a continuous bryophyte layer | Ecosystem respiration, net ecosystem exchange, methane flux and concentration of dissolved organic carbon (raised bog only) measured for up to two years after burning. |
|
| Morton and Heinemeyer, (2019) | Upper Wensleydale, Yorkshire Dales National Park, UK | Blanket bog | Dominated by Sphagnum (moss) species, Eriophorum (cottongrass) species or Calluna vulgaris
| Lab experiment on peat shrinkage 1m field cores in glasshouse experiment
|
|
| Ramchunder et al. (2013) | Pennines, Northern England | Blanket peat | Vegetation dominated by Eriophorum spp. and Calluna. vulgaris, and there was Sphagnum spp. cover at all sites, | Stream dissolved organic carbon concentration
|
|
| Rosenburgh et al. (2013) | Range of sites within North Peak ESA (Bamford, Broomhead and Howdon) | Ombro-trophic mires | dominated by Calluna vulgaris | Soil C:N (g/g) |
|
| Ward et al. (2007) | Moor House NNR, North Pennines, Northern England | Upland banket peat | Calluna vulgaris– Eriophorum vaginatum blanket mire, Empetrum nigrum ssp. nigrum | Soil carbon concentration, Gas emissions (CO2 and CH4), soil solution dissolved organic carbon |
|
| Ward et al. (2012) | Moor House NNR, North Pennines, Northern England | Blanket peat | Calluna vulgaris-Eriophorum vaginatum blanket mire, Empetrum nigrum ssp nigrum | Vegetation, soil and gas measurements after 13CO2 pulse labelling |
|
| Worrall et al. (2007) | Trout Beck, Moor House NNR, North Pennines, Northern England | Blanket peat | Dominated by Eriophorum sp. (cotton grass), Calluna vulgaris and Sphagnum spp. | Dissolved organic carbon |
|
| Worrall et al. (2013a) | Goyt Valley, Derbyshire, Peak District National Park | Deep Peat | dominated by Calluna vulgaris with some Sphagnum spp. In wetter areas | Dissolved organic carbon |
|
| Worrall et al. (2013b) | Peak District National Park, UK | Not stated
| Calluna vulgaris (78%) with some moss (Hypnum cupressiforme and Polytrichum commune) and lichen (Cladonia rangiferina)
| Vegetation C stock (C concentration x vegetation biomass) pre and post prescribed burn
|
|
| Yallop and Clutterbuck (2009) | 50 x small scale catchments, Pennines, and one North York Moors, UK | Categorised into three broad soil types: blanket peat, peaty topsoils and non-peaty soils | Improved grassland, coniferous plantation, ericaceous (predominantly Calluna vulgaris) and grass/sedge dominated moorland were mapped separately | Stream dissolved organic carbon |
|
| Paper | Carbon budget | Experimental design | Geographical coverage | System diversity coverage | Number of burn events | Burn event coverage |
|---|---|---|---|---|---|---|
| Chen et al. (2008) | Low | High | Low | Unclear | High | Unclear |
| Clay et al. (2009)* | Low | High | Low | Unclear | High | Medium |
| Clay et al. (2010b)* | Low | High | Low | Unclear | High | Medium |
| Clay et al. (2010c)* | High | High | Low | Unclear | High | Medium |
| Clay et al. (2012)** | Low | High | Medium | Medium | High | Medium |
| Clay et al. (2015)** | Low | High | Medium | Medium | High | Medium |
| Garnett et al. (2000)* | Low | High | Low | Unclear | High | Medium |
| Grau-Andrés et al. (2018) | Low | High | Medium | Medium | High | Medium |
| Morton and Heinemeyer, (2019) | Low | High | High | High | High | Low |
| Ramchunder et al. (2013) | Low | Medium | High | Unclear | High | Unclear |
| Rosenburgh et al. (2013) | Low | High | High | Unclear | High | Low |
| Ward et al. (2007)* | High | High | Low | Unclear | Medium | Low |
| Ward et al. (2012)* | High | High | Low | Unclear | Medium | Unclear |
| Worrall et al. (2007)* | Low | Medium | Low | Unclear | High | Medium |
| Worrall et al. (2013a) | Low | Medium | Low | Unclear | Medium | High |
| Worrall et al. (2013b) | Low | Medium | Low | Unclear | unclear | Medium |
| Yallop and Clutterbuck (2009) | Low | Unclear | High | Unclear | High | Low |
* Moor House Long Term Experiment: Plots burnt in 1954 then randomly assigned a burning regime (None, every 10 years & every 20 years)
** Both studies use twin site in Northumberland; 1) Emblehope Moor in Redesdale Forest, 2) Ray Demesne moor, near Raylees
| Carbon budget - High | The study includes 3 key areas of a C budget (soil C, vegetation C, water (DOC) and/or gas measurements) |
|---|---|
| Carbon budget - Medium | The study includes 2 key areas of a C budget |
| Carbon budget - Low | The study includes only 1 key area of a C budget |
| Experimental design - High | The study includes both control sites and is replicated |
| Experimental design - Medium | The study includes control sites or has replication |
| Experimental design - Low | The study does not include control sites or replication |
| Geographical coverage - High | The study includes 3 or more contrasting experimental/study sites |
| Geographical coverage - Medium | The study includes 2 contrasting experimental/study sites |
| Geographical coverage - Low | The study includes 1 experimental/study site |
| System diversity coverage - High | The study includes 3 or more contrasting soil or vegetation types |
| System diversity coverage - Medium | The study includes 2 contrasting soil or vegetation types |
| System diversity coverage - Low | The study includes 1 soil/vegetation type |
| Number of burn events - High | The study includes the assessment of 3 or more burn events |
| Number of burn events - Medium | The study includes the assessment of 2 burn events |
| Number of burn events - Low | The study includes the assessment of 1 burn events |
| Burn event coverage - High | The study sampling covers pre, during and post burn |
| Burn event coverage - Medium | The study sampling covers 2 time points (pre & post burn; pre & during burn; during & post burn) |
| Burn event coverage - Low | The study comprises one sampling time point including samples from during or post burn events only |
| Not enough information provided/unclear | Not enough information is provided, or it is unclear |
| Authors | Year | Country | Location | Habitat Type | Species Group/s | Type of Study / Experiment |
|---|---|---|---|---|---|---|
| Hancock et al. | 2011 | Scotland | Abernethy, Scotland | Calluna vulgaris heath | Birds, Invertebrates, Vegetation | Field |
| Robertson et al. | 2017 | England | Northern England | Blanket bog and Calluna vulgaris heath | Birds | Field |
| Smith et al. | 2001 | UK | UK uplands and Langholm | Moorland | Birds | Field |
| Tharme et al. | 2001 | UK | Eastern Scotland, Northern England | Moorland | Birds | Field |
| Bargmann et al. | 2015 | Norway | Norway (Lygra and Lurekalven islands) | Coastal heath | Invertebrates | Field |
| Bargmann et al. | 2016 | Norway | Norway (Lygra and Lurekalven islands) | Coastal heath | Invertebrates | Field |
| Haysom and Coulson | 1998 | UK | UK, Northern England and Southern Scotland | Calluna vulgaris heath | Invertebrates | Field |
| Hochkirch and Adorf | 2007 | Germany | Lower Saxony | Bog | Invertebrates | Field |
| Johnston and Robson | 2018 | UK | UK | Streams (in blanket bog catchments) | Invertebrates | Field |
| Mallik and FitzPatrick | 1996 | Scotland | Aberdeenshire, Scotland | Dry and wet heath | Invertebrates | Field |
| Ramchunder et al. | 2013 | England | Northern England (Upper Teesdale, Wensleydale and Geltsdale) | Streams (in blanket bog catchments) | Invertebrates | Field |
| Davies and Legg | 2008 | Scotland | Mar Lodge, Scotland | Calluna vulgaris heath | Lichens | Field |
| Alday et al. | 2015 | England | Moor House NNR, North Pennines | Blanket bog | Vegetation | Field |
| Davies et al. | 2010 | Scotland | Scottish Highlands | Calluna vulgaris heath | Vegetation | Field |
| Hancock et al. | 2005 | Scotland | Abernethy, Scotland | Calluna vulgaris heath | Vegetation | Field |
| Harris et al. | 2011 | England | Peak District, Northern England | Moorland | Vegetation | Field |
| Lee et al. | 2013a | England | Moor House NNR, North Pennines | Blanket bog | Vegetation | Field |
| Milligan et al. | 2018 | England | Moor House NNR, North Pennines | Blanket bog | Vegetation | Field |
| Noble et al. | 2018b | UK | UK peatlands (95 sites) | Blanket Bog | Vegetation | Field |
| Noble et al. | 2019b | England | Cheviot Hills, North Pennines, Peak District (Northern England) | Blanket Bog | Vegetation | Field |
| Ward et al. | 2007 | England | Moor House NNR, North Pennines | Blanket bog | Vegetation | Field |
| Whitehead and Baines | 2018 | England | Pennines, Northern England | Blanket Bog | Vegetation | Field |
| Whitehead et al. | 2021 | England | Pennines, Northern England | Blanket bog | Vegetation | Field |
| Bargmann et al. | 2014 | Norway | Norway (Lygra island) | Coastal heath | Propagule bank | Glasshouse |
| Lee et al. | 2013b | England | North Peak and Moor House NNR, Northern England | Blanket bog | Propagule bank | Field |
| Grau-Andrés et al. | 2017 | Scotland | Southern Scotland (Braehead Moss) | Raised bog | Sphagnum | Field |
| Noble et al. | 2018a | England | Moor House NNR, North Pennines | Blanket bog | Sphagnum | Field |
| Noble et al. | 2019a | England | Moor House NNR, North Pennines | Blanket bog | Sphagnum | Field |
| Taylor et al. | 2017 | UK | Glasshouse | Peatland | Sphagnum | Glasshouse |
| Curlevski et al. | 2011 | Scotland | Abernethy, Scotland | Calluna vulgaris heath | Soil microbes | Field |
| Ward et al. | 2012 | England | Moor House NNR, North Pennines | Blanket bog | Soil microbes | Field |
| Paper | Experimental design | Geographical coverage | System diversity coverage | Number of burn events | Burn event coverage |
|---|---|---|---|---|---|
| Alday et al. (2015) | High | Low | Low | High | Medium |
| Bargmann et al. (2014) | Medium | Medium | Low | Low | Medium |
| Bargmann et al. (2015) | Medium | High | Low | High | Medium |
| Bargmann et al. (2016) | Medium | High | Low | High | Medium |
| Curlevski et al. (2011) | High | Low | Low | Low | Low |
| Davies and Legg (2008) | Medium | Medium | Low | High | Medium |
| Davies et al. (2010) | Medium | Low | Low | Low | High |
| Grau-Andrés et al. (2017) | High | Low | Low | Low | Medium |
| Hancock et al. (2005) | High | Low | Low | Medium | Medium |
| Hancock et al. (2011) | High | Low | Low | Low | |High |
| Harris et al. (2011) | High | High | Low | High | Medium |
| Haysom & Coulson (1998) | Medium | High | Low | NA | NA |
| Hochkirch and Adorf (2007) | High | High | Low | Low | Medium |
| Johnston and Robson (2018) | High | High | Low | Low | Medium |
| Lee et al. (2013a) | High | Low | Low | High | Medium |
| Lee et al. (2013b) | High | Medium | Medium | High | Medium |
| Mallik and FitzPatrick (1996) | Medium | Medium | Medium | High | Medium |
| Milligan et al. (2018) | High | Low | Low | High | Medium |
| Noble et al. (2017) | High | High | Low | High | Medium |
| Noble et al. (2018) | High | Low | Low | High | Medium |
| Noble et al. (2019) | Medium | High | Low | High | Medium |
| Noble et al. (2019b) | High | High | Low | High | Medium |
| Ramchunder et al. (2013) | High | High | Low | Low | Medium |
| Robertson et al. (2017) | Medium | High | High | Low | Medium |
| Smith et al. (2001) | Medium | High | High | Low | Medium |
| Taylor et al. (2017) | Medium | NA | Low | NA | Medium |
| Tharme et al. (2001) | Medium | High | High | High | Medium |
| Ward et al. (2007) | High | Low | Low | Low | Medium |
| Ward et al. (2012) | High | Low | Low | High | Medium |
| Whitehead and Baines (2018) | High | Low | Low | High | Medium |
| Whitehead et al. (2021) | High | Low | Low | High | Medium |
| Experimental design - High | The study includes both control sites and is replicated |
|---|---|
| Experimental design - Medium | The study includes control sites or has replication |
| Experimental design - Low | The study does not include control sites or replication |
| Geographical coverage - High | The study includes 3 or more contrasting experimental sites |
| Geographical coverage - Medium | The study includes 2 contrasting experimental sites |
| Geographical coverage - Low | The study includes 1 experimental site |
| System diversity coverage - High | The study includes 3 or more contrasting soil or vegetation types |
| System diversity coverage - Medium | The study includes 2 contrasting soil or vegetation types |
| System diversity coverage - Low | The study includes 1 soil or vegetation type |
| Number of burn events - High | The study includes the monitoring of 3 or more burn events (or 3 ages of burned vegetation) or includes the monitoring of short rotation and long rotation burns |
| Number of burn events - Medium | The study includes the monitoring of 2 burn events (or 2 ages of burned vegetation) |
| Number of burn events - Low | The study includes the monitoring of 1 burn event (or 1 age of burned vegetation) |
| Burn event coverage - High | The study includes samples covering pre, during (or immediately after burning), and post burn events |
| Burn event coverage - Medium | The study includes samples covering pre and post burn events or uses a space-for-time substitution study |
| Burn event coverage - Low | The study includes samples covering only post burn events |
| Not applicable | The criterion is not applicable to this study |
Annex 13 – Some additional sources of information that are relevant to the questions but have not been reviewed in this report
Amar, A., Grant, M., Buchanan, G., Sim, I., Wilson, J., Pearce-Higgins, J.W. and Redpath, S. 2011. Exploring the relationships between wader declines and current land-use in the British uplands. Bird Study, 58, 13–26.
Arnell, N., Freeman, A. and Gazzard, R. 2021. The effect of climate change on indicators of fire danger in the UK. Environmental Research Letters, 16(2021), 044027.
Ashby, M.A. 2020. A review of the post-Glaves et al.(2013) Evidence: investigating the effects of managed burning on upland biodiversity, Carbon, greenhouse gas emissions and water. In: Ashby, M., Heinemeyer, A., Stewart, G. and Sotherton, N. 2020. Peatland Protection - The Science: Four key reports. The Uplands Partnership. Report 1.
Belcher, C.A., Brown, I., Clay, G.D, Doerr, S.H, Elliott, A., Gazzard, R., Kettridge, N., Morison, J., Perry, M., Santin, C. and Smith, T.E.L. 2021. UK Wildfires and their Climate Challenges. Expert Led Report Prepared for the Third UK Climate Change Risk Assessment.
Brown, L.E. and Holden, J. 2020. Contextualizing UK moorland burning studies with geographical variables and sponsor identity. Journal of Applied Ecology, 57, 2121–2131.
Burn, W. 2021. The Hidden Half: Blanket Bog Microbial Communities across a Spectrum of Site and Management Conditions and Impacts on Peatland Carbon Cycling. PhD thesis, University of York.
Daplyn, J. and Ewald, J. 2006. Birds, burning and grouse moor management. A Report on Behalf of the Moors for the Future Partnership.
Gazard, R., McMorrow, J. and Aylen, J. 2016. Wildfire policy and management in England: an evolving response from Fire and Rescue Services, forestry and cross-sector groups. Philosophical Transactions of the Royal Society B, 371, 1696.
Grau-Andrés, R., Davies, G.M., Waldron, S., Gray, A. and Bruce, M. 2014. Fuel and climate controls on peatland fire severity. In: Viegas, D.X. (ed). Advances in Forest Fire Research. Imprensa da Universidade de Coimbra. doi.10.14195/978-989-26-0884-6_31
GWCT, 2020. A review of the environmental impacts including carbon sequestration, greenhouse gas emissions and wildfire on peatland in England associated with grouse moor management. In: Ashby, M., Heinemeyer, A., Stewart, G. and Sotherton, N. 2020. Peatland Protection - The Science: Four key reports. The Uplands Partnership. Report 2.
Haworth, P.F. and Thompson, D.B.A. 1990. Factors associated with the breeding distribution of upland birds in the south Pennines, England. Journal of Applied Ecology, 27, 562–577.
Heinemeyer, A., Berry, R. and Sloan, T.J. 2019. Assessing soil compaction and micro-topography impacts of alternative heather cutting as compared to burning as part of grouse moor management on blanket bog. PeerJ, 7, e7298.
International Union for Conservation of Nature National Committee UK, 2020. Position Statement: Burning and Peatlands.
Lees, K.J., Buxton, J., Boulton, C.A., Abrams, J.F. and Lenton, T.M. 2021. Using satellite data to assess management frequency and rate of regeneration on heather moorlands in England as a resilience indicator. Environmental Research Communications, 3, 085003.
Marrs, R.H., Phillips, J.D.P., Todd, P.A., Ghorbani, J. and Le Duc, M.G. 2004. Control of Molinia caerulea on upland moors. Journal of Applied Ecology, 41, 398–411.
McCarroll, J., Chambers, F.M., Webb, J.C. and Thom, T. 2016. Informing innovative peatland conservation in light of palaeoecological evidence for the demise of Sphagnum imbricatum: the case of Oxenhope Moor, Yorkshire, UK. Mires and Peat, 18, 08, 1–24.
Miles, J. 1971. Burning Molinia–dominant vegetation for grazing by red deer. Grass and Forage Science, 26, 247–250.
Moss, D., Joys, A.C., Clark, J.A., Kirby, A., Smith, A., Baines, D. and Crick, H.Q.P. 2005. Timing of breeding of moorland birds. BTO Research Report, 362. Thetford.
Niemeyer, T., Niemeyer, M., Mohamed, A., Fottner, S. and Härdtle, W. 2005. Impact of prescribed burning on the nutrient balance of heathlands with particular reference to nitrogen and phosphorus. Applied Vegetation Science, 8, 183–192.
Picozzi, N. 1968. Grouse bags in relation to the management and geology of heather moors. Journal of Applied Ecology, 5, 483–488.
Reed, M.S., Hubacek, K., Bonn, A., Burt, T.P., Holden, J., Stringer, L.C., Beharry-Borg, N., Buckmaster, S., Chapman, D., Chapman, P.J., Clay, G.D., Cornell, S.J., Dougill, A.J., Evely, A.C., Fraser, E.D.G., Jin, N., Irvine, B.J., Kirkby, M.J., Kunin, W.E., Prell, C., Quinn, C.H., Slee, B., Stagl, S., Termansen, M., Thorp, S. and Worrall, F. 2013. Anticipating and managing future trade-offs and complementarities between ecosystem services. Ecology and Society, 18(1), 5.
Ross, S., Adamson, H. and Moon, A. 2003. Evaluating management techniques for controlling Molinia caerulea and enhancing Calluna vulgaris on upland wet heathland in Northern England, UK. Agriculture Ecosystems & Environment, 97, 39–49.
Stewart, G.B., Glendell, M., McMorran, R., Troldborg, M., Gagkas, Z., Ovando, P., Roberts, M., Maynard, C., Williams, A., Clay, G. and Reed, M.S. 2021. Uplandia: making better policy in complex upland systems – Final Report. Defra and Natural England.
Titterton, P., Crouch, T. and Pilkington, M. 2019. A case study into the estimated amount of carbon released as a result of the wildfire that occurred on the Roaches in August 2018. Moors for the Future Partnership, Edale.
Turner, E.T. and Swindles, G.T. 2012. Ecology of testate amoebae in moorland with a complex fire history: Implications for ecosystem monitoring and sustainable land management. Protist, 163(6), 844–855.
Yallop, A.R., Clutterbuck, B. and Thacker, J. 2010. Increases in humic dissolved organic carbon export from upland peat catchments: the role of temperature, declining sulphur deposition and changes in land management. Climate Research, 45, 43–56.
| Scientific Name | English Name |
|---|---|
| Calluna vulgaris | Heather or Ling |
| Empetrum nigrum | Crowberry |
| Erica cinerea | Bell Heather |
| Erica tetralix | Cross-leaved Heath |
| Eriophorum angustifolium | Common Cottongrass |
| Eriophorum vaginatum | Hare’s-tail Cottongrass |
| Juniperus communis | Common Juniper |
| Molinia caerulea | Purple Moor Grass |
| Nardus stricta | Mat Grass |
| Neottia cordata | Lesser Twayblade |
| Trichophorum germanicum | Deergrass |
| Rubus chamaemorus | Cloudberry |
| Salix aurita | Eared Willow |
| Ulex europaeus | Common Gorse |
| Vaccinium myrtillus | Blaeberry or Bilberry |
| Vaccinium vitis-idaea | Cowberry |
| Scientific Name | English Name |
|---|---|
| Campylopus introflexus | Heath Star Moss |
| Hypnum jutlandicum | Heath Plait Moss |
| Sphagnum capillifolium | Red Bog-moss or Small Red Peat Moss |
| Sphagnum papillosum | Papillose Bog-moss |
| Sphagnum subnitens | Lustrous Bog-moss |
| Scientific Name | English Name |
|---|---|
| Acanthis cabaret | Lesser Redpoll |
| Alauda arvensis | Eurasian Skylark |
| Anthus pratensis | Meadow Pipit |
| Calidris alpina | Dunlin |
| Corvus cornix | Hooded Crow |
| Corvus corone | Carrion Crow |
| Falco columbarius | Merlin |
| Falco tinnunculus | Common Kestrel |
| Gallinago gallinago | Common Snipe |
| Lagopus lagopus scotica | Red Grouse |
| Linaria flavirostris | Twite |
| Numenius arquata | Eurasian Curlew |
| Oenanthe oenanthe | Northern Wheatear |
| Pluvialis apicaria | European Golden Plover |
| Saxicola rubetra | Whinchat |
| Saxicola rubicola | European Stonechat |
| Tetrao urogallus | Capercaillie |
| Tringa totanus | Common Redshank |
| Troglodytes troglodytes | Eurasian Wren |
| Turdus torquatus | Ring Ouzel |
| Vanellus vanellus | Northern Lapwing |
| Scientific Name | English Name |
|---|---|
| Vipera berus | Adder (Common European Adder) |
| Scientific Name | English Name |
|---|---|
| Callophrys rubi | Green Hairstreak |
Disclaimer: Scottish Natural Heritage (SNH) has changed its name to NatureScot as of the 24th August 2020.
At the time of publishing, this document may still refer to Scottish Natural Heritage (SNH) and include the original branding. It may also contain broken links to the old domain.
If you have any issues accessing this document please contact us via our feedback form.
Last updated:







