
NatureScot Research Report 1197 - Seaweed cultivation in Scotland
Published: 2025
Authors: Kerrison, P., Macleod, A. and Thomson, A. (Scottish Association for Marine Science)
Cite as: Kerrison, P., Macleod, A. and Thomson, A. 2025. Seaweed cultivation in Scotland. NatureScot Research Report 1197.
Disclaimer
This report was written in 2019, but not published until 2025. Some information may no longer be current.
Contents
- Keywords
- Background
- Main findings
- 1. Introduction to seaweed cultivation in Scotland/wider UK
- 2. Collection and maintenance of reproductive material
- 3. Hatchery phase
- 4. Approaches to seeding
- 5. Species selection
- 6. Cultivation calendar
- 7. Farm design and management considerations
- 8. Overview of harvesting methods
-
9. Farm siting and environmental interactions
- 9.1 Site selection considerations
- 9.2 Defining the scale of seaweed cultivation projects
-
9.3 Environmental interactions
- 9.3.1 Alteration of ecosystem and creation of habitat
- 9.3.2 Absorption of light
- 9.3.3 Absorption of nutrients
- 9.3.4 Absorption of kinetic energy
- 9.3.5 Release of Dissolved and Particulate Organic Matter (DOM/POM)
- 9.3.6 Release of plastics into the marine environment
- 9.3.7 Entanglement risk
- 9.3.8 Disease and pest management
- 9.3.9 Non-Native Species and Biosecurity Plans
- 9.3.10 Management of environmental interactions
- 10. Final recommendations
Keywords
Seaweed; macroalgae; cultivation; guidance; impacts; environment
Background
The cultivation of macroalgae (seaweed), has recently begun as a new form of aquaculture in coastal Scotland. An established seaweed cultivation industry exists, particularly in East Asia, however the methods used and species grown are different. It is essential to understand how the development of macroalgal aquaculture will affect Scotland’s coastal marine environment.
The guidance provides a summary of the seaweed cultivation in Scotland, including an overview of practical processes, farm design and siting considerations and the possible environmental interactions and biosecurity considerations for this type of aquaculture.
Main findings
- This report summarises the body of knowledge concerning methods in Scottish seaweed cultivation (as reviewed in 2019).
- Farm siting needs to consider the following factors: physicochemical tolerance of the seaweed species, moderate water flow through the site, a typical depth of 20-40 m, operational access to the site as well as other socioeconomic considerations such as social license to operate.
- The scale of farming activity should consider the potential environmental consequences interactions. Small-medium category farms (i.e. less than 50 x 200 m lines) are anticipated to have few detectable impacts; larger scale operations are more likely to cause impacts at a detectable level.
- There are no large-scale seaweed cultivation sites in operation in the UK. Monitoring requirements for future developments will need to be assessed and supported by a more complete understanding of the scale dependent changes likely within receiving environments. Further research on monitoring data will allow the potential magnitude of interactions to be determined, so that appropriate monitoring protocols can be designed for large-scale farming. Key environmental interactions to be considered are detailed below.
- To mitigate light reduction on the benthos, farms should not be sited over sensitive habitats and preferentially in >20 m water depth. Light reduction is unlikely to impact local ecosystems, except at high stocking densities, the effect of which is exaggerated by the scale of the farm size.
- Seaweed farms could be used to remove nutrients generated by anthropogenic activity (bioremediation). At large scales, site specific conditions may result in excessive nutrient removal. In such cases, other cultivated organisms that generate waste nutrients could be co-located or incorporated with the farm as IMTA system (integrated multi-trophic aquaculture). The addition of dissolved inorganic nutrients into the sea should not be allowable.
- Seaweed farms may reduce water flow in the area of the water column where the crop is hanging, and may accelerate the flow underneath the crop. Consideration should be made of site hydrodynamics and benthic topography to understand potential consequences of farm location.
- Seaweeds can release dissolved and particulate organic matter (DOM/POM; mainly carbohydrates) into the surrounding environment. Wild seaweed beds are a major source of naturally occurring DOM/POM. Only larger scale seaweed cultivation is expected to contribute notable inputs to the DOM and POM pool.
- The wear and abrasion of plastic used in farm infrastructure (i.e. ropes, buoys) will be a source of pollution to the marine environment. Although the magnitude of pollution is expected to be far below current sources of marine pollution, new technologies to minimise plastic loss and the use of biodegradable materials should be encouraged.
- The entanglement of marine megafauna (e.g. dolphins, whales) in cultivation infrastructure is possible, but poorly understood. Therefore, farm monitoring protocols should include the documenting and reporting of any entanglement to improve understanding and mitigation of impacts.
- Diseases and pests of cultivated seaweed do occur, but are currently not well understood. Outbreaks may have negative consequences on neighbouring farms and natural seaweed beds in the surrounding ecosystem. Biosecurity planning should utilise a precautionary approach to limit the potential impact from pests and diseases. Planning should be applied at both the farm and hatchery levels of cultivation operations.
- Non-native species (NNS) should not be cultivated. A record should be kept of any non-native or invasive species detected on the farms, which should be included within the biosecurity plans described above. However, this may be a challenge for the average seaweed farmer in terms of practical know-how relating to NNS.
1. Introduction to seaweed cultivation in Scotland/wider UK
1.1 Summary of recent historic activity in the seaweed sector in UK
Seaweed has been utilised by coastal communities for centuries as a human food, animal feed or as fertiliser to support agriculture. Following the industrial revolution, Scotland’s kelp forests were harvested for potash, iodine and alginate extraction in boom and bust cycles (Kenicer et al., 2009). Harvesting for the alginate industry peaked in the 1970s then declined in 1980s due to competition from China, finally ceasing in 2008 (Bailey, 2016).
Modern seaweed cultivation was first established as an industry in 1950s China, and has since grown into an $11 billion worldwide industry, involving at least 27 species totalling 30.5 Mt in 2017 (Food and Agriculture Organization (FAO), 2016; 2019). In recent years, cultivation has begun around Europe as the demand for seaweed has increased and new uses have been developed (Figure 1.1). There is now increasing interest in the development of seaweed cultivation as a new form of aquaculture for coastal Scotland.
There are many different species of seaweed; each with varied uses. These can be broadly split into three groups: the green (Chlorophyta), red (Rhodophyta) and brown (Phaeophyceae) seaweeds. A range of different species are hand-harvested intertidally, predominantly for human food. These are usually dried and ground, but may also be sold fresh or as whole dried blades. Some macroalgae are hard to obtain in sufficient quantities by hand-harvesting, so cultivation is needed to reliably meet the market demand. Currently, the only commercial seaweed cultivation in the UK is for the human food market.
Some seaweeds such as the large brown kelp species have been identified as bioresource feedstocks for products that require a very large volume of biomass, produced at a low cost. For instance (in decreasing value): animal feed (health promotion), bioplastics, biopolymers (e.g. alginate) and biofuel. To be economical, farms for these products would need to be large scale, possibly covering many km2 in area. Operations would be highly mechanised, and most likely be sited far from the coastline. Such farms are currently not feasible due to engineering, logistical and economic constraints.
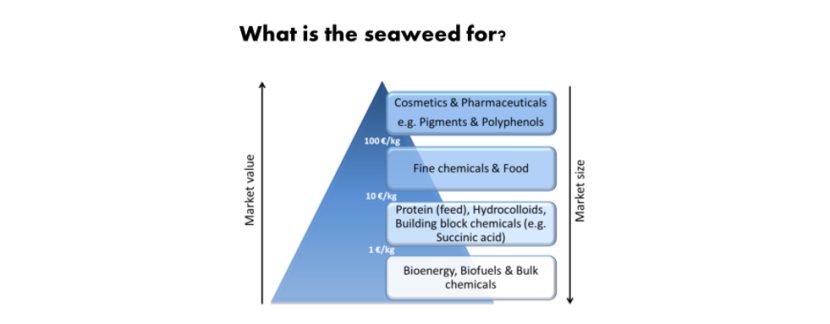
Figure 1.1. Seaweed value pyramid. Modified after Bruton et al. (2009).
Click for a full description
Diagram titled ‘What is the seaweed for?’ Seaweed products are listed in order of increasing market size and decreasing market value (ranging from 100 €/kg through 10 €/kg to 1 €/kg): Cosmetics & Pharmaceuticals e.g. Pigments & Polyphenols; Fine chemicals & Food; Protein (feed), Hydrocolloids, Building block chemicals (e.g. Succinic acid); and Bioenergy, Biofuels & Bulk chemicals.
Certain species may also contain compounds that can be extracted as valuable pharmaceutical/nutraceutical products. At the point of this review, there is no known cultivation of seaweeds for this end-use in Europe. Farms to support the pharmaceutical industry could be any size, ranging from 1 to 1,000 hectares, depending on the market demand for the end-product.
1.2 Recent cultivation activity
The Scottish Association for Marine Science (SAMS) has worked on seaweed cultivation projects for over 15 years. It currently runs two experimental seaweed farms near Oban, Argyll. Trials have also been run in Loch Roag on Lewis, and Loch Fyne and Loch Etive in Argyll. SAMS are actively involved in multi-year research projects involving seaweed cultivation. The commercial arm of SAMS, SAMS Enterprise provides commercial seeded line to UK cultivators from their seaweed hatchery, and offers other associated seaweed consultancy services.
Seaweed aquaculture research has previously been carried out by North Atlantic Fisheries College (NAFC) Marine Centre in Shetland, which ran the Shetland Seaweed Growers project from 2014-2016. This project included farming trials in association with Scottish Sea Farms Ltd., and East Voe Shellfish Ltd. Further seaweed cultivation research has also been carried out by Ardtoe Marine laboratory (now FAI Aquaculture), with farming trials in Loch Sunart.
The Scottish Seaweed Industry Association (SSIA) was created in 2014 and has held a yearly conference since 2016. It is a forum that brings together the majority of entities (both research and industry) carrying out seaweed cultivation, or who have an end-use for the biomass. While the industry is currently dependent on wild harvesting of seaweed, cultivation is recognised by the SSIA as an important step forward for the future. Invited speakers have included the commercial European cultivators OceanRainforest SpF (Faroe Islands) and Islander Kelp Ltd. (N Ireland).
At the point of this report, there is only a single company, New Wave Foods Ltd., which is actively engaged in Scottish seaweed aquaculture. They have two licensed sites near Oban, Argyll, with one operational in 2018-2019. In the wider UK context, Islander Kelp Ltd., based in Northern Ireland, run a profitable business based on seaweed cultivation. Also, the Cornish Seaweed Company Ltd. (Cornwall) are running small-scale trials in collaboration with a local mussel farmer, and were the recent recipients of EMFF grants with Exeter University and Plymouth Marine Laboratory. Seagrown Ltd. (Yorkshire) were awarded £472k by the Coastal Communities Fund in 2019 for the development of a seaweed hatchery and farm. As far as the authors are aware, at least five further UK companies have applied for funding and/or farming licences up to 2019.
2. Collection and maintenance of reproductive material
The lifecycle of seaweeds is highly varied between different species and can be complicated, with multiple free-living stages. This report focuses mainly on the kelps since these are the only group that can be cultivated reliably.
2.1 Seasonality of fertile material collections
Cultivation begins with the collection of fertile wild seaweed to provide a seeding stock. For kelp species, collection is timed to coincide with spring low tides, when most seaweeds are either exposed at the surface or are in very shallow water. Other species that are found higher in the intertidal are available for collection more often. Before undertaking any shore-based activities, an appropriate health and safety assessment should be conducted, including identification of necessary personal protective equipment (PPE) to be worn. A biosecurity plan should be in place to ensure the shore activity does not contribute to the potential spread of non-native species or disease. Adhering to the ‘Check, Clean, Dry’ protocol as recommended by the GB non-native species secretariat should be strongly considered for seaweed cultivation sites. Please see Section 7.3.9 for more information on Biosecurity Plans.
The reproductive peak of many cultivated seaweed species (S. latissima, A. esculenta, P. palmata) occurs from autumn through to early spring, when water nutrient concentrations are naturally higher and temperature and light are lower. This provides a suitable environment for the less tolerant juveniles to begin to grow. In some species such as L. digitata and L. hyperborea, their reproductive peak is during summer/autumn.
Seaweed distribution varies depending on the environmental and habitat requirements of each species. For example, it is unlikely to find A. esculenta in sheltered sandy bays as it is most common on exposed rockfaces. Online resources such as NBN Atlas Scotland, the Marine Life Information Network (MarLIN) and Algaebase, give information on individual species distribution and their site/growth requirements.
2.2 Recommended distance between collection and cultivation site
It is in the interest of the seaweed farming industry to maintain wild genetic diversity and mitigate the impacts of farm to wild interactions (Cottier-Cook et al., 2016; Barbier et al., 2019). Wild diversity not only supports resilience to environmental changes in wild populations, but also offers the genetic resources for sustainable cultivation and future domestication and breeding efforts. Mitigation of farm to wild impacts, including genetic pollution from gene swamping, and the introduction of non-native genotypes leading to the breakdown of local adaptation, also plays a strong role in social license development for the farming sector. Determining a suitable distance from which source material should be collected from the cultivation site is therefore an important consideration to prevent potentially negative ecosystem impacts.
Although spore dispersal in seaweeds is generally thought to be localised (0.1-10 kms), the widespread distribution and the broadcast spawning of seaweed means the effective population size is thought to be relatively large in many species (Schiel and Foster, 2006). In general, broadcast spawners in the marine environment are expected to follow the genetic pattern of isolation by distance, with greater genetic distance, and decreased gene exchange, over increasing geographical distance. Additionally, local topography and environmental differences can add further barriers to gene flow creating complex patterns of diversity and connectivity over heterogeneous landscapes (Selkoe et al., 2016).
At present, the scale and scope of seaweed farming in Scotland is unlikely to lead to impacts on wild population genetics from farm to wild interactions. The large effective population size of wild populations, and the use of local wild strains for cultivation, mean escapes from farm genotypes have a minimal risk of impacting on wild diversity. However, with the anticipated growth of the seaweed industry, the risks of cultivation effects on wild population diversity increase as farm capacity and breeding and domestication efforts expand. A number of simple mitigation steps could be applied to seaweed cultivation practices to reduce the potential impacts of future farming efforts on wild seaweed populations.
In an attempt to mitigate potential damage to wild populations, a condition has been set in Marine Licences that states:
“The licensee must utilise locally sourced stocks for seeding of the cultivation systems”
(Licence No.; 05292/17/0: Port a Bhuiltin, Lynn of Lorn).
Yet, ‘local’ cannot be defined without understanding the genetic population structure. The recent PEGASUS report (Barbier et al., 2019) defines a local strain as:
“A cultivated strain or variety whose genetic background is similar to that of the natural population geographically close. The degree of similarity taken into account is directly dependent on the observed genetic diversity of the species in the considered area, compared to distant populations of the same species. It is a relative parameter (“more or less similar”).”
Population differentiation metrics, based on traditional pairwise population Fst statistics, can offer a statistical means to infer genetic distance between populations. This can vary substantially depending on the species, genetic markers, and statistical approach used, however offers a useful relative measure of genetic distance, and thereby a means of assessing “local” populations within species and study areas. To date, a number of population genetic surveys have been undertaken on S. latissima, including in Denmark (Nielsen et al., 2016), Maine (USA) (Breton et al., 2018), and the Irish Sea (Mooney et al., 2018). Significant genetic differentiation was typically observed at distances of 50-100 km. In Scotland and the Irish sea, populations from Stranraer and Troon showed relatively little differentiation (φ-st = 0.024) over >60 km distance (Mooney et al., 2018). Guideline distances between collection populations and cultivation sites could be suggested based on general trends in existing studies, however the heterogeneous nature of landscapes and population connectivity make inference of population differentiation without localised genetic knowledge difficult. As such it is recommended that relevant regional genetic surveys be undertaken in areas of cultivation interest.
As well as genetic distance, environmental distance between collection and farm site should also be considered when selecting wild populations for cultivation material. Little research exists on local adaptation in seaweeds, however the wide geographical distribution of many species, their ability to colonise a wide range of environments, and the phenotypic diversity observed across those environments, suggests the strong influence of local adaptation in many species (Augyte et al., 2018). As such selecting seedstock from a dissimilar environment to the farm location may prove detrimental to the growth of the outplanted crop in the new environment. Though determining environmental similarity can be difficult, a simple solution may be to sample from multiple locations within the agreed local limits for seedstock in order to encompass a greater genetic diversity and a wider range of environmental tolerances. Selecting a greater number of adult donors from multiple source populations may also offer further mitigating benefits by increasing the effective population size of the farmed crop and reducing the degenerate impact of a small hatchery source population on wild population effective size (Laikre et al., 2010).
Recommendation
- Where possible seedstock should be collected from the immediate vicinity of the farm in order to minimise the genetic and environmental distance between source seedstock and cultivation site.
- Alternatively, seedstock should be collected from multiple locations within the same waterbody, ideally as close to the cultivation site as possible and within < 25 km of the farm (by sea), taking into account major physical and environmental barriers to population connectivity.
- Some cultivation sites may have suitable conditions for seaweed growth, but no natural populations within 25 km of the proposed cultivation site (e.g. offshore or along sandy coastlines). In this situation, it is recommended to use the closest natural population within 150 km (by sea), once again taking into account major physical and environmental barriers to population connectivity. In these situations, it is advised that early dialogue and guidance should be sought from NatureScot.
- Seedstock pools should ideally be sourced from around 10-20 adult donors to increase seed diversity and effective population size and reduce the effects of small hatchery source populations on wild effective population size.
2.3 Quantity of fertile material required and collection method
Kelps have a very high fecundity, and a single adult can release hundreds of millions of spores from their reproductive regions, known as sori. When seeding a farm it is recommended at least 5-10 sorus regions are collected to ensure a mixed genetic diversity in the crop. This could potentially seed 10-20 km of hatchery twine (see Section 4). If very few sori are used, this may cause inbreeding depression in the crop, leading to poor growth on the farm. In Australia, Sea Health Products Ltd. have been granted a licence to collect up to 30 individuals of their common kelp (Ecklonia radiata) per annum, for the purpose of seeding trials, which seems a very reasonable allowance (Jo Lane, pers. comms). Whilst collection of sorus is required, the small quantities required are highly unlikely to lead to any significant impact on the local population.
The sorus region of Laminaria spp. and S. latissima is located towards the distal end of the blade (Figure 2.1A) and can be removed while leaving the adult plant in place to regrow. In A. esculenta the sorus region is located on the sporophylls near the holdfast (Figure 2.1B), and similarly these can be removed while leaving the adult in place. Commercial collections should be carried out in accordance with requirement of a Crown Estate licence to ensure the practice is managed sustainably.

Figure 2.1. Sorus material on kelps appears are slightly raised dark regions. On many species it is located on the frond, usually at the distal end.
Click for a full description
Left (A): Small areas of sori on a portion of a smooth-looking Laminaria digitata blade.
Right (B): Large areas of sori on a wide section of textured S. latissima frond.
The quantity of fertile material required for many other species is currently unknown. In the case of P. palmata, a larger quantity of individuals/ fertile material is necessary due to the smaller spore output in this specie. Based on Werner & Dring (2011), between 6.5-13.1 kg of fertile material per linear km seeded is required. In all species, care should be taken to leave a portion of the adult meristematic tissue (i.e. the growth area of the plant) in place to allow tissue regeneration, and to leave at least half of the fertile wild population undisturbed to allow population recovery following collection. This is particularly true for species that have a patchy distribution, such as P. palmata. Special consideration of the possible environmental impacts of this harvesting would need to be made if large scale cultivation of P. palmata was begun in Scotland.
Guides exist for the preparation and extraction of spores from kelps (Werner & Dring, 2011; Flavin et al., 2013; Rolin et al., 2016). Generally, this involves a cleaning step (see Section 4.2) followed by slight drying overnight in a fridge. The following day, spores are released following re-immersion in seawater (Figure 2.2).

Figure 2.2. Sorus material of S. latissima which was cleaned and desiccated overnight, then re-immersed in seawater. Within 15-30 mins spore release causes the water to become opaque.
2.4 Use of kelp seedbanks
Kelps have an independent microscopic lifestage called the gametophyte. This hardy stage grows as filaments and can be maintained in vegetative culture, and can even survive a year in darkness (tom Dieck, 1993). Gametophyte cultures can act as seedstock for cultivation for many years and are available year-round, negating the need to annually search for and recollect fertile material from the wild. Independent gametophyte cultures for a particular species can be maintained for each location used for cultivation (Figure 2.3). It is not recommended to collect seed from the cultivated seaweed itself, as this will reduce the genetic diversity of the seedstock and may select for earlier reproductive maturity.

Figure 2.3. Kelp gametophytes are maintained in a vegetative stage using red light.
Click for a full description
Left (A): Image shows five stoppered, round-bottomed flasks ¾ full of bubbling liquid. Each flask is connected to tubing and are positioned on a rack under red light.
Right (B): Image depicts several racks of labelled tissue culture flasks under red light.
In the future, commercial seedbanks for kelps may allow the long-term storage of cultures from all across Europe. These can act as a biological bank (biobank) to conserve the natural genetic diversity of seaweed populations, as well as supplying commercial kelp farmers. Such a biobank is currently being established at SAMS for S. latissima from across Europe under the project Genialg.Biobanking may be assisted through the use of cryopreservation for the long-term (decadal) storage of gametophytes. Further study is needed to confirm that growth and fertility can recover following this treatment (Day, 2018), although initial trials are encouraging (Wouter Visch unpublished results).
Porphyra/Pyropia spp. also have an independent filamentous lifestage called the conchocelis. This may be biobanked and cryopreserved, similar to kelp. Methods to biobank many other seaweed species (e.g. P. palmata, Himanthalia elongata) have not yet been developed, and so these species currently require collection of fertile material from the wild every year. An attempt at biobanking with O. pinnatifida was not successful.
3. Hatchery phase
During the hatchery/nursery phase of cultivation, the seaweed seed (usually a microscopic stage), is reared under set light, temperature and nutrients conditions to maximise early growth and survival. The hatchery phase is usually 6-8 weeks, until the juveniles have grown up to 1 cm long. The culture is maintained as cleanly as possible to prevent the inclusion of grazing animals or overgrowth by space competitors such as other macroalgae (Figure 3.1).
3.1 Summary of the process of seeded twine production
Twine seeding is currently the most reliable method for seaweed cultivation (see Section 4). 1-2 mm twine is wound around plastic tubing (spools) in a single layer. The twine on the spools is then seeded with a microscopic phase of the seaweed (usually by spraying or dipping). The spools are then immersed into seawater tanks set up with appropriate lighting and supplied with gentle aeration. The tanks will either be supplied with flow-through seawater or be static with regular refreshment (e.g. weekly). The water (usually natural seawater) should always be filtered and UV sterilised to prevent the introduction of other organisms.

Figure 3.1. Seeded twine production.
Click for a full description
Left (A): Image depicts two rows of artificially illuminated, aerated plastic tanks of seawater. Spools of 1–2mm twine seeded with the microscopic phase of the seaweed are immersed in these tanks for 6–8 weeks.
Right (B): A close-up image of sections of seeded twine showing the juvenile seaweed.
The addition of a typical algal growth medium such as F/2 or Provassoli’s Enriched Seawater (PES) is recommended to accelerate the growth of the juvenile seaweed during this initial vulnerable stage. The chemical germanium dioxide is also recommended during the first seven to nine days of the hatchery to inhibit overgrowth by diatoms and other microorganisms (Kerrison et al., 2016). Open access guides exist that provide more detail on these hatchery methods (Edwards & Watson, 2011; Flavin et al., 2013).
3.2 Hatchery biosecurity
Hatcheries must ensure that appropriate practical biosecurity measures are in place to prevent the accidental movement and spread of invasive non-native species and/or disease. A biosecurity plan should be established to address these concerns.
3.2.1 Cleaning fertile tissue
Fertile seaweed tissue collected from the wild must be cleaned before spores are extracted. This will ensure that other organisms are not accidentally introduced into the culture. This first step involves the manual removal of all macroscopic biota (i.e. mobile animals, other seaweeds, encrusting bryozoan etc. that are visible to the naked eye; see Section 6.2). This is achieved by trimming and repeatedly rubbing with tissue and sterile seawater. A second step may involve a chemical treatment such as dipping in dilute bleach, peroxide or iodine (Rød, 2012; Flavin et al., 2013). The selection of the chemical will depend on the tolerance of the particular species being cultured. Spores extracted from cleaned fertile material are then able to grow without competition from non-target species or grazing. This treatment does not make the material axenic (free of all microbes), as it is beneficial for the seaweed to retain its natural microbial associations.
3.2.2 Maintaining hygiene in the hatchery
All hatchery materials (including twine spools, tanks and bubbling tubes), should be cleaned before and after use. In stagnant culture, the water should be refreshed regularly and the tanks cleaned and all equipment sterilised. The use of multiple, spatially separate tanks are recommended, in case one becomes contaminated. To prevent tank cross-contamination, each tank should be covered and clean gloves worn when handling the spools. Waste water discharge from the hatchery should follow local regulations.
3.2.3 Hatcheries handling non-local seaweed populations
Biosecurity must be stricter when non-local (see Section 2.2) seaweed populations are cultured, or when seaweeds from multiple locations are grown in the same hatchery. Firstly, each population must be cultured in isolated tanks and no equipment moved between tanks without first being sterilised. Secondly, all wastewater must be chemically treated before being discharged as waste. These steps will prevent two potential outcomes:
- The release of seaweed to a non-local/distant cultivation site leading to unknown ecological consequences (e.g. seaweed from Shetland being outplanted in the Firth of Clyde).
- The cross-contamination of seaweed disease/contaminants between populations. Seaweed diseases/ culture contaminants are currently understudied and may be transported into the hatchery with the seedstock (either wild collected spores or gametophyte cultures). Currently the only published work in this area has shown that hatchery raised S. latissima were not infected by a common endophyte (Bernard et al. 2019). It is important that hatcheries do not facilitate disease transmission by preventing cross-contamination of tanks which are then outplanted all around the UK. Detection methods for diseases/contaminants are currently being researched at SAMS.
Figure 3.2 below provides a schematic diagram of the typical phases of kelp cultivation in Europe.
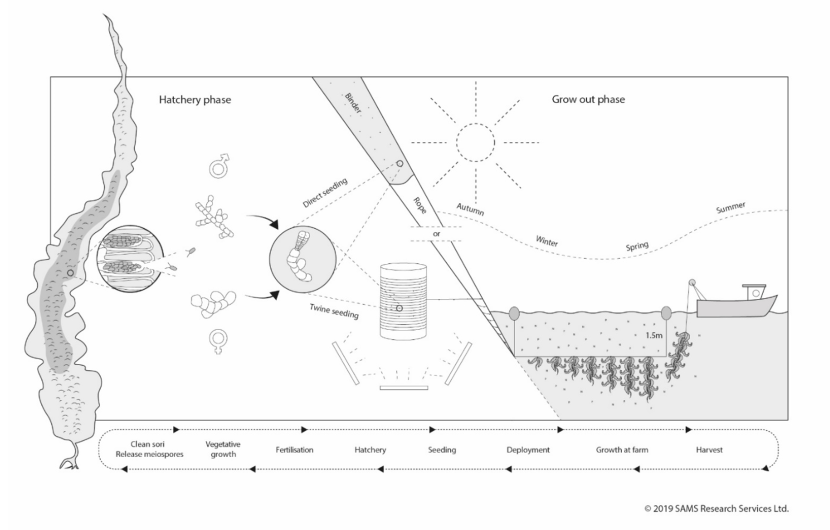
Figure 3.2. Schematic diagram of the typical phases of kelp cultivation in Europe.
Click for a full description
Fertile kelp (sporophyte stage) with darkened areas of sporangial tissue are collected form the wild and cleaned. Meiospore release is stimulated, these germinate and grow into either male or female gametophytes. The male spermatia fertilise the female oogonium and a new juvenile sporophyte grows attached to the female. This culture is then seeded onto twine spools and allowed to grow for 6-8 wk in a hatchery with artificial lighting. The twine is then outplanted at a seaweed farm by helically wrapping it around a rope. An alternative seeding method is that the culture containing juvenile sporophytes is seeded directly onto a rope using the binder method without and extended hatchery phase. Using either method, rope are deployed in autumn suspended at 1.5 m depth. The seaweed grows by intercepting incident light and absorbing nutrients from the surrounding water. The harvesting time is in spring to early summer.
4. Approaches to seeding
4.1 Twine vs. direct seeding
Effective seeding of macroalgae requires an initial hatchery phase, which maximises the survival of early recruits by optimising the conditions for their growth from microscopic spores to macroscopic juveniles (see Section 3). Seeding of spores followed by direct outplanting in the sea without a hatchery phase gives very poor results (Kerrison et al., 2018).
There are currently two approaches to seeding; the established method using twine, and a new method called direct seeding.
4.1.1 Twine
In a typical seaweed (kelp) hatchery, juveniles are reared attached to twine on a spool for 6-8 weeks, until they are, generally, up to 1 cm long. Similar method have been used for dulse (P. palmata).
There are many variants of how this twine is then utilised:
- Rapid method (common in Europe). The twine spools are threaded onto a carrier rope and helically unwound into the lay of the rope (Figure 4.1 A/B). This method allows fairly fast deployment. As the twine is coated in a dense population of juveniles, there is intense competition between the juveniles for space. Thus, a large proportion of the juveniles die. If successful, this method leads to 100% coverage of the rope. If problems arise during the hatchery phase, or the line is not deployed correctly, patchiness can occur. This method is most suitable for lengthy continuous longlines.
- Twine inserts. The twine is cut into short sections (e.g. 10 cm), which are inserted (lazy spliced) into the carrier rope at regular intervals (e.g. 30 cm). By spacing out the twine, this reduces the space competition, giving each individual more space to grow to its maximum size. This method uses less twine, but is more labour intensive. It is often used in dropper systems.
- Individual juvenile inserts. This is the dominant method used in Chile/China. Individual juveniles are picked from the twine and lazy spliced into the carrier rope at set distances (e.g. 30 cm). This method requires that the juveniles are grown to a large size in the hatchery (5-10 cm), and strongly selects for fast growth. It is the most economical use of the juveniles and, due to the spacing, allows each to grow to their maximum size without intraspecific competition. This method is extremely labour intensive and so is not expected to be feasible for European seaweed cultivation.
4.1.2 Direct/binder seeding
An alternative seeding method was developed during the EU FP7 project AT~SEA. The kelp juveniles are grown in tumble culture, detached from any surface, rather than on twine. These juveniles are then mixed with a Binder/Bioglue and applied directly onto the cultivation surface, which can then be immediately deployed into the sea (Figure 4.1C). This method has numerous advantages. The hatchery phase is cheaper, deployment is faster, and growth can occur on materials other than ropes (i.e. net or sheets; Kerrison et al., 2018). It is very suitable for sheltered locations, but it is currently unreliable in exposed sites or during periods of unsettled weather. Further research is underway to improve the formulation of the Binder/Bioglue to allow deployment at any site and time.

Figure 4.1. (A) Rapid twine seeding; (B) Close-up of helical wrapping during an experiment; (C) Direct seeding.
Click for a full description
Top left (A): Two people in a boat unwinding juvenile seaweed on twine onto a rope for deployment in the sea.
Top right (B): Light-coloured twine with dark areas of juvenile seaweed on its surface being wound around a length of thick rope in the laboratory.
Bottom left (C): Two people in a laboratory beside a plastic tank containing a liquid solution of juvenile seaweed and binder, in which ribbon-like material is being immersed.
5. Species selection
Numerous species native to Scotland have been trialled for cultivation. This section provides a brief summary of each species. For further information regarding their use in food, please consult Brunner (2015). The Scottish Government’s Seaweed Cultivation Policy Statement does not allow the cultivation of non-native seaweed species in Scotland (Marine Scotland, 2017).
5.1 Kelps
The group most widely known for cultivation are the kelps. The cultivation method for this group was developed in China in the 1950s, for the species Saccharina japonica and Undaria pinnatifida. In 2017 the industrial cultivation of these two species totalled 12.5 Mt, more than a third of global seaweed production, and 10-fold more than all seaweed gathering from the wild (FAO, 2019). These methods have since been adapted for the cultivation of European kelps. Cultivation has been shown to be successful in Laminaria digitata, Alaria esculenta and Saccharina latissima. Each of these species have different properties making them suitable for different markets.
5.1.1 Laminaria digitata
Common names: Oarweed, Sea tangle
This species is the typical kelp bed species seen exposed at low tide. It has a similar geographical distribution to Saccharina latissima but has slower growth (Figure 5.1A). The species is characterised by an extremely high iodine content, which can be reduced by boiling/blanching. There appears to be less demand for the cultivation of this species, as it can be naturally harvested. Despite this, L. digitata cultivation has been undertaken by Islander Kelp Ltd., and it has previously been harvested in Scotland for its alginate content.
The closely related species Laminaria hyperborea is not used for food (Figure 5.1B). It is very slow growing and has value as a source of high quality alginate. Both species are also of interest for the extraction of microcrystalline cellulose for various industries including printer ink and biodegradable polymer film.

Figure 5.1. (A) Laminaria digitata; (B) The similar species Laminaria hyperborea.
Click for a full description
Left (A): An example of L. digitata, laid out on wet sand. A long stipe is attached to a rock at the lower end by root-like protrusions known as a holdfast and at the other end is the blade, divided into finger-like segments.
Right (B): Two examples of L. hyperborea, laid out on wet sand. The holdfasts are attached to a rock. The blades are divided into finger-like segments like L. digitata.
5.1.2 Saccharina latissima
Common names: Sugar kelp, sea-belt
This fast growing species is widely distributed across Europe (Portugal to Norway) and the West Atlantic coast (Greenland to New Jersey). It has been the focus of many collaborative research projects and is currently the most cultivated European seaweed (Figure 5.2). Commercial cultivation is known in Norway, Denmark, the Netherlands, France, Germany, Portugal, Spain, Ireland, Scotland and the Faroe Islands. It has value as a food due to its sweet taste from a high mannitol content. It can also taste metallic, whilst older material can be bitter. It is related to the East Asian cultivated species S. japonica (Kombu), and so is marketed as Sweet or Royal Kombu. It may have future value for chemical extraction/conversion, but this end market has yet to be commercially realised.

Figure 5.2. Saccharina latissima. (A) Collected from coastal Argyll; (B) Cultivated on a longline at Kerrera seaweed farm, Argyll (SAMS operated).
Click for a full description
Left (A): A single strand of sugar kelp (Saccharina latissima) seaweed lying on flat wet sand. It has a wide, dark central midrib and a flat, undulate blade.
Right (B): Bundles of sugar kelp (Saccharina latissima) seaweed growing on six vertically hanging ropes attached to a longline rope being lifted out of the sea. The kelp is growing densely on the upper parts of the vertical ropes and less densely further down.
5.1.3 Alaria esculenta
Common names: Dabberlocks, Winged kelp
Alaria esculenta is another fast growing species. It is naturally found in exposed locations, but will grow well in sheltered sites when cultivated (Figure 5.3). It is sensitive to high summer temperatures, with a southern limit to its distribution in Brittany. This southern boundary of distribution is expected to move northwards under future climate change scenarios. Alaria esculenta is cultivated in Ireland, Scotland, Norway and the Faroe Islands. The more limited distribution of the species means there is good potential for export to European markets. Its frond has a fresh grassy taste, similar to the East Asia cultivated species Undaria pinnatifida (Wakame), and so is sometimes marketed as Atlantic Wakame. The sporophyll bladelets near the holdfast and its central midrib are tougher and have a stronger flavour.

Figure 5.3. Alaria esculenta. (A) Collected from coastal Argyll; (B) Cultivated on a a longline at Port a Bhuiltin seaweed farm, Argyll (SAMS operated).
Click for a full description
Left (A): Three wet fronds of Alaria esculenta in a coastal shore environment, showing the distinct midribs and unbranched, wavy membranous laminae, which taper towards the top. The stipe is short.
Right (B): A longline of densely cultivated Alaria esculenta supported between two people. The long fronds are about the same height as the people.
5.2 Other species that may be relevant in the future
There is interest to cultivate other European seaweeds, although further research is required to make their cultivation reliable. These species hold potential as future target species for cultivation.
5.2.1 Ulva spp.
This seaweed can be various shades of green, with a number of species that can be hard to distinguish (Figure 5.4). It forms a thin sheet that fragments easily, and so it is not suitable for open water cultivation, except possibly in very sheltered sites. Ulva can grow very quickly in high nutrient conditions resulting in green tides. Tank cultivation is underway in southern Europe (ALGAplus) and Israel.

Figure 5.4. Ulva spp. tank cultured in Scotland.
Click for a full description
Displayed on the side of the culture tank, the frond of this seaweed has a delicate appearance, is light green in colour and has a broad blade with crinkly edges like a lettuce.
5.2.2 Porphyra/Pyropia spp.
This group encompasses a number of species that are found intertidally as thin membranous sheets that range from red to dark purple in colour. They have a complex lifecycle, requiring an expensive hatchery process, but are cultivated in East Asia to produce high value nori sheets (Figure 5.5B). Despite the low biomass yield, over 2.5 Mt were produced in 2017. Cultivation of this species may be possible in Europe and trials are underway in Norway, Ireland, Portugal and Scotland (Figure 5.5A). It is grown using tank cultivation by the Portuguese company ALGAplus.

Figure 5.5. Porphyra spp. (A) Natural growth attached to rocks in the low-intertidal, Iona, Argyll. (B) An example of cultivation in Wando, South Korea.
Click for a full description
Left (A): A length of red seaweed attached to a rock is being lifted out of the water to reveal the red thin, sheet-like appearance of this algae.
Right (B): An area of sea containing a length of net attached to floats which is used to cultivate Porphyra spp., close to the water surface.
5.2.3 Osmundea pinnatifida
This small red seaweed grows slowly in the low intertidal (Figure 5.6). It has a strong garlic-like flavour and so it has value as a condiment. It may also contain other bioactives with pharmaceutical/nutraceutical applications. Cultivation for this species is still in its infancy and has not yet developed commercially. Again, tank cultivation may be the way forward with this species.

Figure 5.6. Osmundea pinnatifida. (A) Growing inter-mixed with other species on an intertidal rockface, Argyll; (B) Growing as biofouling after natural settlement on the Kerrera seaweed farm, Argyll (SAMS operated).
Click for a full description
Left (A): Osmundea pinnatifida among a community of small seaweed species, of various colours growing on a rock in a coastal environment.
Right (B): A hand holding a sample of Osmundea pinnatifida which has become attached to a rope at a seaweed farm. The photograph illustrates the small size of this red seaweed, with its flattened fronds and branched appearance.
5.2.4 Palmaria palmata
Also known as Dulse and Dilisk (Figure 5.7), this red North Atlantic intertidal species is highly valued for food due to its umami flavour. It is currently wild harvested across its distribution, particularly in Ireland and Scotland. Cultivation has been trialled since the 1980s, but there are still problems with the control of the lifecycle making it unreliable. Research is underway in this area due to the market demand.

Figure 5.7. Palmaria palmata biofouling at Kerrera seaweed farm, Argyll (SAMS operated).
5.3 Can seaweed be cultivated to meet the market demand?
Through ongoing discussion with established seaweed companies, both in the UK and across Europe, it is believed that there exists an unfulfilled market demand for cultivated seaweed products in both domestic and foreign markets.
A robust cultivation method exists for the kelps. Both S. latissima and A. esculenta are thought to have a high market demand, as they are hard to harvest from the wild in large quantities. In particular A. esculenta may attract a higher quayside price (Table 5.1). Laminaria spp. gives a lower cultivation yield and may have a lower market demand as these species can be harvested from the wild more easily than S. latissima and A. esculenta.
Other species do not have reliable cultivation methods in Europe, although this may change. In particular, species of Porphyra/Pyropia are cultivated widely in East Asia as they produce a high value product. Cultivation of the European species may become established in the future.
| Seaweed species | Market demand | Market relative value | Cultivation yield | Cultivation reliability |
|---|---|---|---|---|
| Saccharina latissima | High | Low | High | High |
| Laminaria spp. | Low | Low | Moderate | High |
| Alaria esculenta | High | Moderate | High | High |
| Porphyra/Pyropia | High | High | Low | Low |
| Palmaria palmata | High | High | Moderate | Low |
| Ulva spp. | Moderate | Moderate | Low | Low |
| Osmundea pinnatifida | High (?) | High | Very Low | Low |
6. Cultivation calendar
6.1 Outplanting and harvesting timings for different macroalgal groups
Cultivation times are broadly similar across a range of seaweed species. In natural populations, many species are fertile over the winter-spring period, with developing young juveniles able to grow rapidly during spring. A high biomass is then achieved in the early summer (Figure 6.1B).
In the 1950s researchers in China developed the summer hatchery method. In this, juveniles are grown in cooled water in the late summer, and then are outplanted as soon as the water temperature decreases in autumn. This resulted in far higher yields at harvest time in summer. This method can also be applied to European kelp species.

Figure 6.1. (A) Juvenile growth in January (up to 25 cm in length) after deployment in October; (B) A cultivation line at harvest with 10-15 kg per m of linear rope.
Click for a full description
Left (A): A section of rope lifted from the sea to show the growth of kelp after 3 months of cultivation. Kelp fronds of varying length are visible.
Right (B): Five people harvesting a cultivation line completely covered in long, dense kelp fronds.
October outplanting of kelps in Scotland has been shown to give a higher yield compared to deployment in later months (Figure 6.1A). September may also give excellent results but has not been trialled. The latest month for deployment that can potentially give a reasonable harvest yield is February. December appears to be the worst month due to increased storminess and low light. In order of preference, the ideal deployment months are Oct>Nov>Jan>Feb. For other species (i.e. P. palmata) the deployment time in Scotland has not been studied, but is expected to be similar. The fine sheet-like Porphyra/Pyropia spp., grows to a harvestable size very quickly and so deployment for this species may be later in the year, possibly Jan-April. Table 6.1 summaries the seaweed cultivation calendar for Scotland.
| - | S | O | N | D | J | F | M | A | M | J | J | A |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kelps | d | d | d | - | d | d | - | h | h | h | - | - |
| Alaria esculenta | d | d | d | - | d | d | - | h | h | - | - | - |
| Palmaria palmata | d | d | d | - | d | d | - | - | h | h | h | h |
| Porphyra/Pyropia | - | - | - | - | d | d | d | d | h | h | h | h |
In most kelps the harvesting period is between April to June, with the exception of A. esculenta, which tends to degrade earlier and so harvesting may need to be completed before June (Table 6.1). It may be possible to harvest the red seaweed species including P. palmata and Porphyra/Pyropia spp., over the summer, although this needs to be tested.
Four factors influence the chosen harvesting time:
- The growth rate of the crop. This depends on the outplanting time, species and method (e.g. longline, droppers), as well as the environmental conditions over the cultivation cycle. The growth rate is very high over the spring-summer period. The total biomass yield continues to increase over this period and then may peak in mid-summer. After this, the yield may decline due to nutrient limited conditions (see Section 7.4) and overgrowth by biofouling.
- The development of biofouling by other organisms. Many invertebrate animals or other macroalgae will grow on the surface of cultivated seaweed. These may overgrow the frond, graze it away or cause it to fragment. This is an unavoidable natural process, increasing with the time cultivated material stays in the water. The timing and type of biofouling appears to be highly site and species specific, and varies inter-annually. Monitoring of fouling is therefore very important for harvest timing. See Section 6.2 for a summary of the different types of fouling.
The end use of the biomass. For food applications, the seaweed frond needs to be as clean as possible with little or no biofouling organisms. This is particularly true when sold fresh or as the whole frond. Due to the nature of cultivating in the sea, it is (likely) impossible to prevent biofouling from developing or ensure the 100% removal of all contaminating material from the final product. Thus, harvesting a lower crop biomass, before biofouling becomes well established, will be favoured over harvesting later, when peak crop biomass is available. Careful monitoring of the onset of biofouling (intensity and organism type) is therefore very important for food end uses.
As a generalisation for non-food applications, the most important factor determining the harvesting time will be yield achieved; biofouling will have a lower impact on the quality of the bulk biomass harvested, and so a later harvest with higher bulk biomass, but also higher % composition of fouling contamination will likely be favoured. The allowable fouling will vary depend on the particular end use or processing method used.
- Seaweed chemical composition. The chemical composition of seaweeds also varies seasonally (Schiener et al., 2015). For example in kelps, the highest protein is achieved in autumn/winter, while highest carbohydrate content is seen in summer. Harvesting time may therefore vary depending on the end use (e.g. a particular target bioactive may peak in January). For food applications, the flavour profile of seaweed also varies seasonally, and so harvest time may need to be adjusted to a particular time of year.
6.2 Common biofouling on cultivated seaweed in Scotland
The surface of macroalgae provides a habitat for other species. These may be termed as biofouling if they degrade the quality of the seaweed. These include sessile organisms, such as other seaweeds and colonial animals, and also mobile animals including snails, amphipods and small fish. The most important species identified in Scotland are shown in Figure 6.2.

Figure 6.2. Groups of organisms which may cause biofouling problems on cultivated seaweed in Scotland.
Click for a full description
Top left (A): Filamentous brown algae.
These can cause mat-like growth on the ends of seaweed, which hold large quantities of water. They settle during winter and early spring so only cover the oldest material
Top right (B): Colonial hydroids
These animals appear as spider webs that spread diffusely over the frond surface with short (~1 cm) filaments that stick out from the surface. They appear during spring and facilitate the colonisation of other animals.
Middle left (C): Colonial bryozoans
Another colonial organism, bryozoans also appear to settle most during the spring. The colonies totally cover the frond as they grow. By mid-summer they can smother the entire frond.
Middle right (D): Lacuna vincta
These grazing snails settle from the plankton during the spring. 100s may cover each blade. The snails can then grow up to 1 cm long, causing extensive grazing damage in the summer.
Bottom left (E): Jassa falcata
The adults of this omnivorous amphipod form burrows on the surface of the algal frond to brood their young which degrades the quality of the frond. This can become a problem in early to mid-summer.
7. Farm design and management considerations
The goal of the farm infrastructure is to keep the seaweed cultivation material, usually longlines, in a stable position for the duration of the cultivation cycle (i.e. between deployment and harvest). The seaweed is generally held within 1-2 m of the surface, but sometimes to a maximum depth of about 5 m. The farm therefore requires a mooring to the substratum and a floatation system. The structure must be suitably engineered to tolerate storms and be located in sufficient depth of water to prevent it beaching. Many different forms of farm have been developed.
7.1 General farm requirements
Aquaculture sites in Scotland require a Crown Estate licence, a Marine Scotland Marine Licence and must include suitable site marking (see Wood et al., 2017). The farm system used will depend on the volume of seaweed to be produced, the intended end-use of the biomass, and whether a manual or mechanised harvesting method will be used.
7.2 Different farm structures currently used
7.2.1 Adapted mussel longline
Double-header longlines can be an excellent structure for seaweed cultivation when only a small quantity of biomass is required (Figures 7.1 & 7.2). The components and methods to construct these structures are widely available, and mussel farmers can immediately re-purpose unused lines. The header lines at the surface allow the easy attachment of droppers. The mooring are usually drag embedment anchors, which must be tensioned.
There are some drawbacks to these systems. Firstly, they tend to have an excessive amount of buoyancy, as they are designed for growing heavy mussels. This can easily be remedied by reducing the number of floats to one every 10 m, as this should be sufficient for seaweed cultivation. Secondly, the surface lines and large floats attract heavy fouling.

Figure 7.1. Mussel system used for seaweed cultivation in the Sound of Kerrera, Argyll (operated by SAMS).
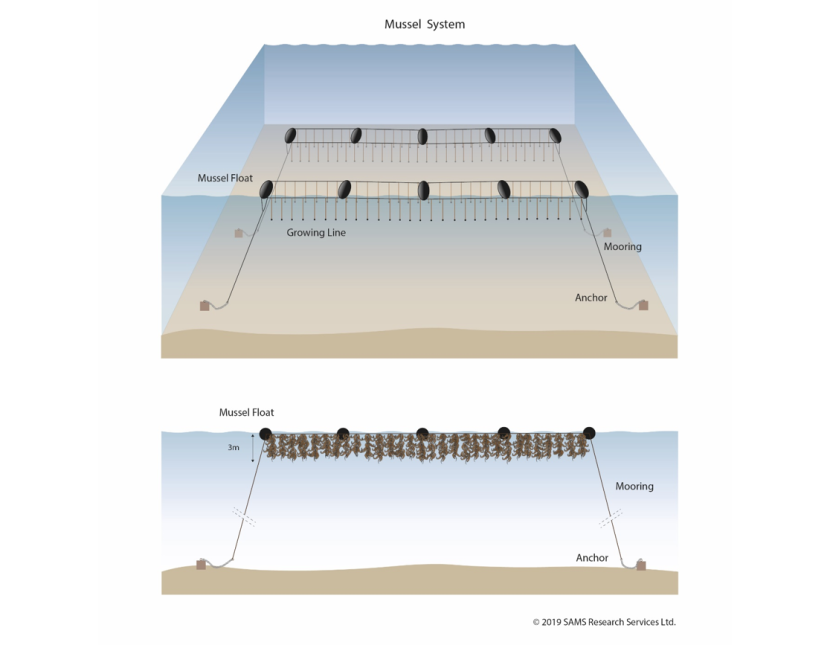
Figure 7.2. Diagrammatic view of a pair of double-header rope mussel systems used for seaweed cultivation.
Click for a full description
Top: Two rows of mussel floats, moored and anchored to the seabed. In each row, each mussel float is connected by two horizontal lines to the next float. These lines support vertical growing lines.
Bottom: Showing how lengths of up to 3 m of seaweed can be supported on the vertical lines in the mussel system.
© 2019 SAMS Research Services Ltd.
7.2.2 Individual longlines
Longline systems have a simple, cheap construction, generally with moorings every 100 m (Figures 7.3 & 7.4). The moorings themselves can be made from various available materials including concrete blocks or eco-anchors containing local stone. A guide to their construction is given by Edwards & Watson (2011). Longlines are excellent for most farms. The growing line (usually at 1.5 m depth) is loose, allowing it to be easily pulled to the surface for inspection. Approximately 10 m spacing is recommended between parallel longlines to prevent interaction during storms. This system is not economical at large scale due to the need for a large number of anchors.

Figure 7.3. Parallel longline systems for seaweed cultivation in Wando, South Korea.
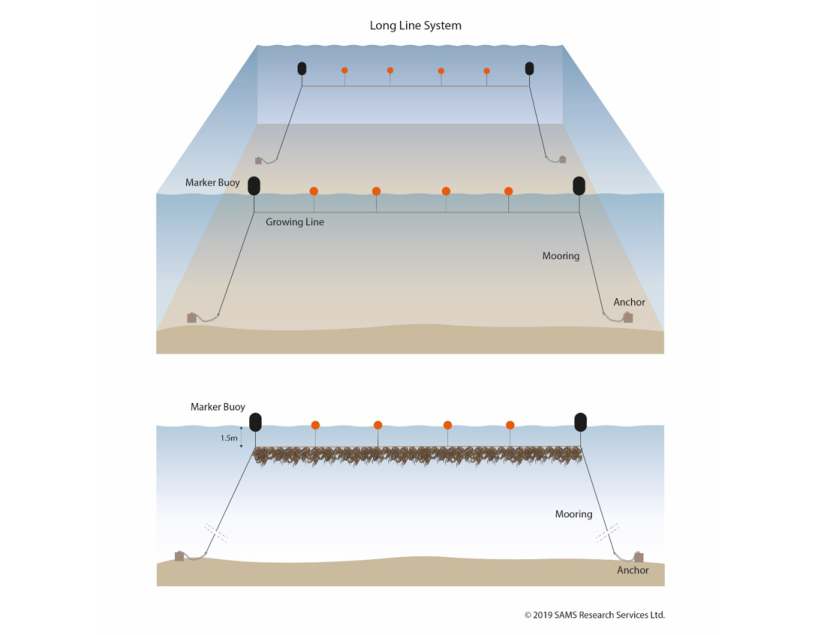
Figure 7.4. Diagrammatic view of a pair of longline systems used for seaweed cultivation.
Click for a full description
Top: The arrangement of a pair of longlines. Each line comprises an anchored marker buoy at either end of a line of floats. The floats mark the location of a single horizontal growing line suspended beneath the sea surface.
Bottom: A cross-section showing how a single growing line, 1.5 m beneath the sea surface, supports seaweed cultivation.
© 2019 SAMS Research Services Ltd.
7.2.3 Grid based system
Grid based systems are likely to be most suitable where larger farms are needed and/or space is restricted (Figures 7.5 & 7.6). A sub-surface rope grid is positioned at a set depth below the surface (e.g. 3 m). This is anchored in all directions using embedment anchors or pilings. Surface buoyancy prevents the grid from sinking. Cultivation lines are then attached onto the grid at a set distance apart. Grids require less anchorage than longline systems, but a disadvantage is that due to the grid being tensioned below the water, it can be difficult to access the growing lines from the surface without a mechanical winch. The rigid nature of this system, resisting water movements, may increase component wear compared to more flexible systems e.g. individual longlines.

Figure 7.5. 100x100 m grid system used for seaweed cultivation at Port a Bhuiltin, Argyll (operated by SAMS).
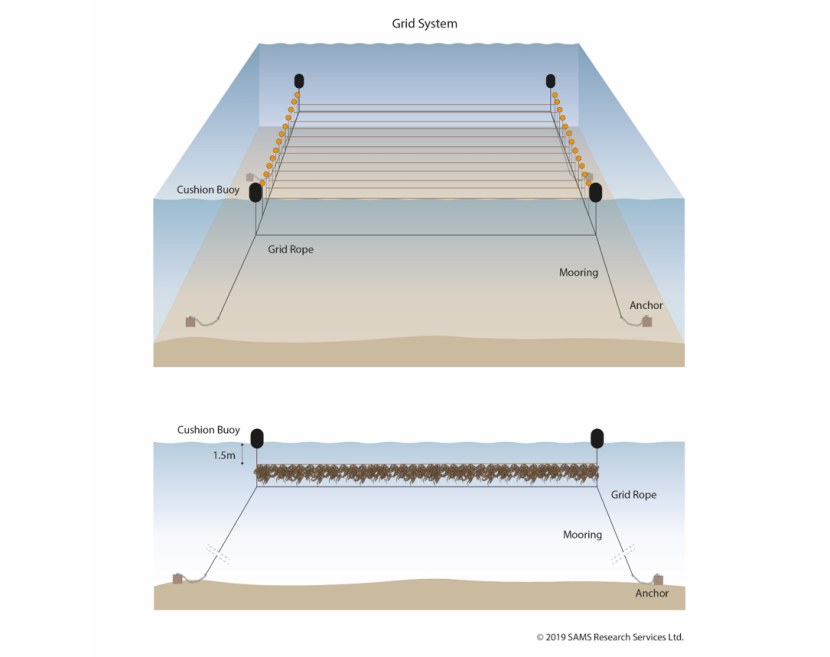
Figure 7.6. Diagrammatic view of a grid based system used for seaweed cultivation.
Click for a full description
Top: Showing a grid, the four corners of which are marked by anchored cushion buoys. These are connected by a sub-surface grid rope, which is attached to surface floats. Cultivation lines are then attached to the grid at defined distances.
Bottom: A cross-section of the grid-based system and illustrates how the seaweed grows at a depth of 1.5 m below the sea surface on the suspended cultivation rope.
© 2019 SAMS Research Services Ltd.
7.2.4 Offshore cultivation rig
This system was developed by OceanRainforest SpF for use in deep oceanic water (Figure 7.7). A sub-surface rope line is loosely moored over 1-2 km, with buoyed vertical ropes that rise to the surface that are used for cultivation. This unique system is only suited where a large sea surface is free for cultivation, as each parallel line must be 20-50 m apart to prevent rope interaction. The loose construction allows the structure to move far more freely with water currents than other systems.

Figure 7.7. Riser growth lines of the OceanRainforest offshore cultivation rig in the Faroe islands. Photo credit OceanRainforest SpF.
7.3 Inspections and maintenance operations
Once deployed, seaweed grows using light from the sun and by absorbing nutrients and dissolved gas from the surrounding water. No further direct intervention by the farmer is necessary (i.e. no fertilisation of the site is needed). Regular site inspections are still essential.
Site inspections are required under the conditions of the Marine Licence to ensure that navigation lighting and the farm infrastructure are maintained in good condition. If the site is deemed to pose a threat to marine traffic due to inadequate lighting, maintenance, drifting or wreck, the licensee is liable for expenses incurred. Criminal charges, such as negligence, or civil action may be pursued if this leads to injury or damage to property. It is recommended that the licensee has suitable insurance in place.
Part 2 12. “The licensee must ensure that a weekly check of the sites marking and lighting equipment….”
Part 2 15. “The licensee must ensure that the works are maintained at all times in good repair.”
Part 2 18. “If in the opinion of the licensing authority the assistance of a Governmental Department, including the broadcast of navigational warnings, is required to deal with any emergency arising from:
a. The failure to mark and light the works as required by licence.
b. The maintenance of the works.
c. The drifting or wreck of the works.
The licensee shall be liable for any expenses incurred in securing such assistance.”
Extract from: Schedule to Licence No.; 05292/17/0: Port a Bhuiltin, Lynn of Lorn, 4/9/17.
Other non-urgent but actionable observations (e.g. partially worn ropes/missing buoys) should be noted, so that time is allocated and suitable equipment can be brought during the next farm inspection to carry out the repair. It is likely that in the future remote sensing methods e.g. unmanned aerial vehicles (commonly known as drones) may be used to assist in these tasks.
At regular intervals during the growth cycle, growth monitoring visits (see Section 7.4) should be used to determine the optimal harvesting time and allow a prediction of the harvesting yield. During these visits, inspections should be made of the accessible below surface infrastructure to:
- Identify and replace worn connections or wear points;
- Monitor for the presence of INNS;
- Remove biofouling where necessary (Figure 7.8);
- Adjust buoyancy of the lines: little buoyancy is necessary when out-planted, but more may need to be added during spring-summer.
It is good practice to also arrange yearly inspections of all moorings using either divers or a remotely operated vehicle (ROV). This will also allow other problems with the sub-surface infrastructure to be identified. It is recommended that a yearly service contract is negotiated with a suitable marine contractor if the farm operators are unable to conduct the activities themselves.

Figure 7.8. Fouling on farm infrastructure by non-target species should be removed regularly to prevent it impacting on the growth of the crop.
During all inspections, the appearance of invasive non-native species should be recorded. Guidance on how to report sightings can be found on the NatureScot Identify and report invasive non-native species webpage. In Scotland, the two most important to look out for are Didemnum vexillum (Dvex, the carpet sea squirt) and Undaria pinnatifida (Japanese Wakame kelp)
7.4 Growth monitoring to predict harvest
After out-planting in October, it is recommended that the site is inspected within 2-3 weeks, and/or following the next storm. This will allow early detection and corrections of problems and allow contingency planning in the case of catastrophic damage. Growth is slow in late autumn and winter due to low light levels, so early growth may not be detectable within this period.
Growth monitoring visits are then typically carried out every 4-6 weeks. The growth rate accelerates during spring as light availability increases and nutrients levels are high (Figure 7.9 A). As the harvest period is approached (see Section 8), monitoring should increase in frequency to once every 1-3 weeks. This will allow the harvestable biomass to be predicted allowing an accurate stock forecast and the organisation of appropriate logistics in preparation for harvest.
The extent of biofouling should also be recorded. Biofouling by other seaweeds and animals tends to become a problem during the late spring – early summer period (Figure 7.9 B). The extent and type of biofouling varies with location and also inter-annually (see Section 6.2). Consistent monitoring is required to allow all cultivators to understand and predict its occurrence. Biofouling can rapidly spread through the crop within 2-3 weeks, degrading the quality of the biomass, particularly for food applications. Biofouling may be allowable for bioresource applications.
The optimal harvesting time is determined by the biomass accumulated (kg) and the level of biofouling. For bioresource applications, chemical analysis may also be employed as seaweed composition is known to vary fluctuate over a yearly cycle (Schiener et al., 2015).
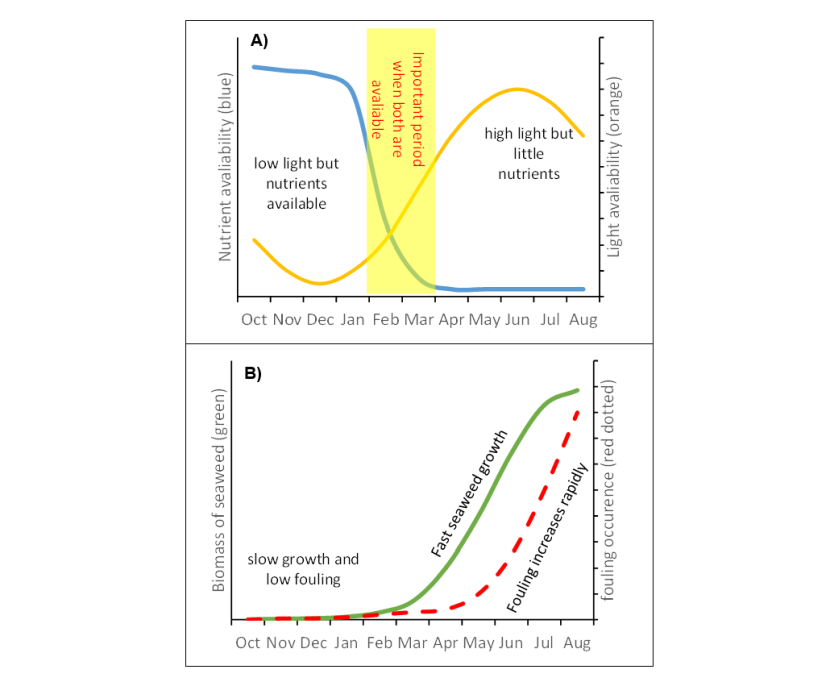
Figure 7.9. An approximate representation of the seasonal changes relevant to seaweed harvesting. (A) The availability of light and nutrients over the cultivation cycle; (B) Typical seaweed biomass accumulation and the occurrence of biofouling.
Click for a full description
Top (A): Graph showing how nutrient availability (left y-axis) and light availability (right y-axis) vary during the seaweed cultivation year (x-axis). From October to January, light levels are low but nutrient levels are high while from April to August, light levels are high but few nutrients are available. February and March represent months when both light and nutrient levels are suitable and available for seaweed cultivation.
Bottom (B): Graph showing how seaweed biomass accumulation and the occurrence of biofouling vary during the year from October to August (x-axis). Seaweed biomass (left y-axis), is at a low level from October to February when it begins to increase rapidly, reaching a peak by August when biomass production begins to slow. The occurrence of biofouling (right y-axis) begins later than seaweed biomass accumulation, rapidly increasing at the beginning of May and continuing until August.
Example monitoring protocol:
- 30 cm length of cultivation line selected at a certain depth, typically 1-1.5 m;
- The five largest blades are collected; frond length and maximum width are measured. The stipe length, stipe maximum diameter and fresh mass may also be recorded.
- Counts/ estimates made of the number of blades >5 cm within the 30 cm length.
- Estimate fouling: 1) % coverage of hydroids, bryozoans and seaweeds (separately); 2) abundance of grazing snails; 3) abundance of all other animals together. Abundance ranked using SACFOR scale: super abundant, abundant, common, frequent, occasional and rare.
- All the biomass stripped from the 30 cm section and bagged – to be weighed onshore.
- Steps 1-5 replicated 3-5 times.
- This process should be completed for each species cultivated. If cultivating using dropper lines you may wish to monitor at multiple depths, e.g., 1, 2, and 3 m.
Note: the 30 cm sections must be selected randomly – do not select the best performing area as this will overestimate the total biomass.
7.5 Monitoring environmental variables
Some monitoring of environmental conditions is recommended at the farm. Although it is understood how the environmental conditions influence seaweed growth, how they influence biofouling may be more important. Environmental monitoring may assist in harvest yield/ fouling onset prediction over multiple years.
It is recommended that temperature and light at the cultivation depth (e.g. 1.5 m) are monitored and recorded. A cost-effective method is to use HOBO Pendant loggers (Onset Corp, USA), which can be set to log every 30 mins and can be downloaded using an Android smart phone. In areas where salinity may be periodically low, an aquarium refractometer can be used during each sampling visit. Monitoring of the nutrient concentrations may also be desirable. Aquarium test strips can give an instant measurement for nitrate, phosphate and ammonium. A more accurate measurement can be achieved using a handheld photometer.
8. Overview of harvesting methods
The harvest should be timed to optimise both the quality (biochemical composition and/or lack of fouling) and quantity of the biomass (discussed in Section 6.1). It is likely that a balance between quality and quantity will need to be decided on to maximise the value of the crop produced. The harvesting method will depend on the species, end-use of the biomass and the scale of the operation. Full or partial harvests and the level of mechanisation are important factors in deciding the appropriate methods to be used (Table 8.1).
- | Full cut | Partial cutting |
|---|---|---|
Manual (no mechinisation) | Suitable for small market operations (< 20 tonne per annum) e.g. aquaculture diversification or high value products where biomass quality must be maximised:
SME branded cosmetics or soaps. | Also suitable for small market operations. Cutting at multiple times a year is likely to increase overall crop yield. Quality may vary between cuttings. This method is used to take multiple harvests from Porphyra/Pyropia spp. cultivation in E Asia, in a single growth season. This may also be possible for some species of kelps. |
Simple mechanised | Full stripping of longlines can be achieved by winching longlines lines through a metal eye/shackle. Alternatively, a lifting apparatus can be used to raise the line onto the boat. A boat with suitable deck space and carrying capacity can be adapted for this. This method is most suitable where a bulk biomass will be required for non-food applications (<1000 tonne per annum):
| Not currently possible. |
Full mechanisation | Full mechanisation will be expensive and designed depending on the farm and species cultivated. It is likely to only be suitable when cultivating >1000 tonne per annum for bioresource applications. Full mechanisation is essential for low value applications such as biofuels. Prototype harvesting machines for kelp have been trialled in Europe in the last few years (Figure 8.2). | Such full mechanisation will also be expensive. A prototype has trialled partial cutting of kelp (Figure 8.2), but is slower than manual cutting For Porphyra/Pyropia spp. cultivated on nets, a specially designed boat is used to perform partial cuts. This is economical due to its high value for the production of Nori sheets for human food. |
8.1 Full or partial harvest
Full harvesting is the most straightforward method. All of the biomass is stripped, and the cultivation line removed from the site. This method is indiscriminate, capturing all fouling organisms within the crop.
Partial harvesting involves either the selection of particular blades (e.g. whole blades for restaurants) or cropping by removing the distal portion of the blade to allow regrow. This method is far more discriminatory and time consuming. It may result in an increased crop yield over the year by reducing space competition (self-shading). Seaweed grows from its meristematic tissue (location varies with species), and so when cropping it is important that this tissue is not cut, and is retained on the cultivation line.
Cropping may carry an additional benefit for kelp cultivation. The distal blade is the oldest and so will tend to have the largest settlement of fouling organisms. Some of these organisms (e.g. bryozoans and hydrozoans) will spread over the biomass in the following months. Spring cropping may remove many of these attached colonies and so reduce biofouling later in the year.
8.2 Manual or mechanised
Manual harvesting is the method currently employed by most companies and research projects around Europe. Generally, the cultivation line is lifted from the water onto a small boat where the seaweed is pulled or cut from the rope. This labour intensive method is only suitable in calm conditions and where a small quantity of biomass is needed (<20 tonnes per annum). It carries the risk of back injury if a poor lifting method is used. It is not recommended for large cultivation structures (i.e. nets and sheets), which are very heavy in the water.

Figure 8.1. Manual harvesting of seaweed lines is possible but is only suitable for small scale operations due to handling risks.
Click for a full description
Left (A): A person holding seaweed on a horizontal cultivation line up out of the sea. Lengths of dark seaweed fronds are visible hanging from the line.
Right (B): Three people manually hauling a heavily laden seaweed line from the sea into the boat they are standing in.
Where a large quantity of biomass is needed (20<1000 tonne per annum), simple mechanisation on a larger boat such as a flatbed will improve the speed of harvesting and the safety of the employees. This may involve a lifting arm to raise the line out of the water for partial harvesting or winching a longline through a metal eye or shackle to strip off the biomass.
Where a very large quantity of kelp biomass (>1000 tonne per annum) is needed at a low cost, high throughput mechanisation is essential. This will require purpose-built boats or machines. Some prototypes have been trialled in Europe, but are not yet used commercially. The cultivation of high value Porphyra/Pyropia spp. in East Asia already employs a purpose-built harvesting boat to carry out multiple partial cuts over the cultivation season.
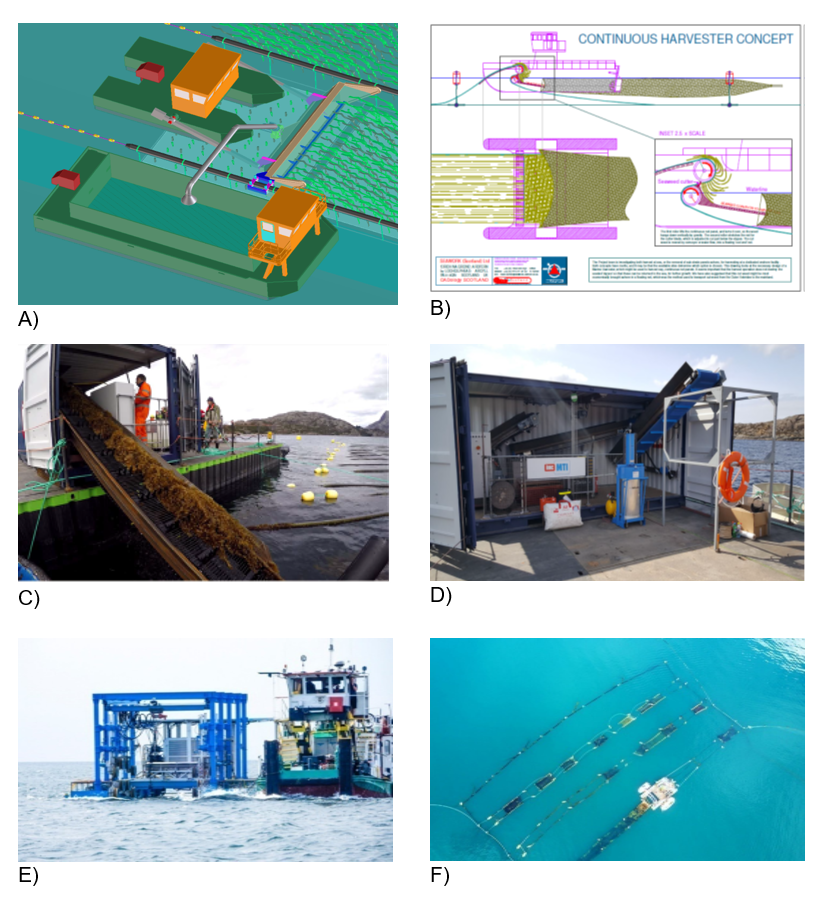
Figure 8.2. Different conceptual or prototype harvesters.
Click for a full description
Top left (A): Diagram of a harvester design by the Energy research Centre of the Netherlands (Lenstra et al., 2011).
Top right (B): Continuous harvester concept by Seawork (Scotland) Ltd. 2012.
Middlfe left (C) and middle right (D): The IHC MTI prototype seaweed harvesting machine (20 ft containerised).
Bottom left (E) and bottom right (F): SeaHarvester I developed by AtseaNOVA.L.
9. Farm siting and environmental interactions
9.1 Site selection considerations
The optimal habitat requirements for many of the candidate seaweed species for Scottish cultivation (Section 5) are well known (Kerrison et al., 2015). Modelling work has also been conducted to predict their natural coastal distribution (Burrows et al., 2012). In general, where there is a stable surface for growth (such as a seaweed farm), many species will grow best in stable physicochemical conditions where summer temperatures do not exceed 18-20 oC, salinity is high (little freshwater influence), turbidity is low (high water clarity) and moderate water flow refreshes the supply of nutrients. These conditions encompass a large swathe of the Scottish coastline, but exclude some environments including estuaries and some sheltered embayments and sea-lochs. Stakeholders may benefit from a strategic modelling approach to identify suitable cultivation sites along Scotland’s coast (van der Molen et al., 2018).
The availability of locations that are otherwise suited to seaweed growth will then be restricted by the engineering considerations of the farm infrastructure. Very shallow locations (<10 m) are generally considered unsuitable due to the risk of equipment beaching, while deep locations (>40 m) require longer mooring lines and so will be more expensive to deploy and maintain, and will require a larger licenced area. Currently in Scotland, 96% of fish and shellfish farms are in depths less than 40 m. New mooring systems are under development (Aquamoor, North Sea Farm and Ocean Rainforest), which allow deployment in deeper water.
Similar to depth, wave exposure limits the locations for possible cultivation around the Scottish coastline. Some degree of wave action is preferable to promote nutrient uptake and facilitate fast seaweed growth (more so than for finfish or shellfish aquaculture). However, very high wave exposure will: (a) increase component wear on the farm structure and necessitate a well-engineered durable design; and (b) decrease the accessibility of the site. Seaweed farms do not require daily access, but the workability of the site during the busy periods of seeding and harvesting should be a principle consideration in site selection. Similar to finfish aquaculture, future ‘off-shore’ cultivation is thought to be crucial to realise the full potential of the industry. Technological advances in remote sensing and the use of autonomous underwater vehicles and unmanned aircraft may help to reduce the operational cost of such off-shore cultivation.
Other restrictions to the site selection may be required depending on the final end-use of the biomass (e.g. avoiding areas enriched with heavy metals or other pollution sources if the seaweed is being used for food). It has been recommended by the Scottish Seaweed Cultivation Policy Document (Marine Scotland, 2017) that seaweed for food should be sited in Class II Shellfish waters; however, this would firstly restrict cultivation to mainly near-shore environments, many of which will be sub-optimal for seaweed growth. Secondly, the monitoring for shellfish waters may not be of relevance to seaweed cultivation i.e. while shellfish filter seawater and so can concentrate harmful microbes or toxins from toxic microalgae, seaweeds do not.
Other operational considerations for seaweed cultivation will include the distance to a boat launching point for inspections, the distance to the landing point (if different) and the location of the processing facility. Seaweed degrades very quickly in air, and so must be processed as soon as possible to avoid loss of biomass.
Although socioeconomic considerations are outside the scope of this report, they are also essential to consider when selecting cultivation sites. These include conflicts with static and mobile fisheries, boating access, visual impact and recreational uses. Of particular note is the opportunity to co-locate seaweed aquaculture with other marine activities, producing environmental, economic and social synergies with other stakeholders (e.g. fish and shellfish aquaculture, scallop ranching, static fisheries, renewable energy). Such synergies can play an important role in the social acceptability of seaweed cultivation and associated industries. Synergies should be encouraged, and are likely to be better received during public consultation events.
Summary and mitigation recommendations
- The optimal habitat requirements for seaweed growth are well documented and much of the Scottish coastline is suitable.
- Areas with depths of 20-40 m with good water exchange due to waves are preferred.
- Fully exposed, off-shore areas in deep water are currently not suitable for cultivation due to engineering limitations (but may become relevant in the future)
- Seaweed cultivation projects have to balance the biological considerations for optimised growth with the practical reality of working at the site.
- Co-location with other marine activities such as animal aquaculture or renewable energy.
9.2 Defining the scale of seaweed cultivation projects
To understand the possible environmental interactions of seaweed cultivation on the Scottish marine environment, a description of the activity must first be provided. Currently, Scottish Government Policy has been to define farm size by direct comparison with mussel cultivation header line structures (Marine Scotland, 2017; also see Figure 7.1). This defines a ‘small-medium’ farm as having less than 50 x 200 m lines where the likelihood of negative environmental interactions is considered low. A 220 m mussel header line can hold 10,000 m of grow-out dropper rope (Poseidon Aquatic Resource Management, 2017). In contrast, it is estimated that only 500-1,000 m of seaweed can be efficiently grown on the same structure, due to differential needs for light and larger individual size compared to mussels. Whilst, differences in productivity between groups using the same mussel-based system are expected, an order of magnitude difference in productivity between existing seaweed cultivation systems is possible depending on the interpretation of scale defined by the Scottish Government.
It is anticipated that most seaweed cultivators will utilise grid-like supporting structures with parallel horizontal growing longlines to maximise the exposure of the seaweed to surface light (see Figure 7.3). Therefore the scales defined by the Government Policy are not relatable to anticipated future seaweed cultivation in Scotland. Further definition of what constitutes a “line” under the existing policy will allow regulators to understand the relative makeup of supporting structures (moorings, grids and longlines) and temporary growing structures (materials used to support seaweed biomass) using different systems.
Rather than referring to the amount of infrastructure installed (number of lines), it is suggested that it would be preferable to use a simple set of site parameters to allow for easy comparison of different cultivation approaches. Such an approach is shown below, defining four metrics that should allow for comparison and contrast between sites utilizing different cultivation approaches. The use of this parameter will be explored in follow on work.
The anticipated (or actual) total site production over the year (kg). This is the total fresh weight produced by the site. It is anticipated that for many scale-dependant environmental interactions, total site production, and not a description on number of ‘lines’, will provide a better description of the activity to be considered.
The anticipated (or actual) fresh weight yield per linear metre of growing line (kg m-1). For kelps, this is likely to range between 4-15 kg m-1, depending on species and harvesting time. This is the quayside weight before processing.
Surface stocking density. This is a measure of the extent of growing structures per meter squared of cultivation area. In the example below uses linear metres of seeded line.
The area used for cultivation (m2). This is to include the full extent of the moorings. So a 100x100m grid containing growing lines = 10,000 m2 (1 ha) + approximately 4 ha of space for mooring lines
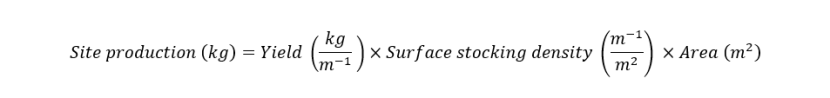
Equation 1
Click for a full description
Site production (kilograms) = Yield (kilograms per linear metre of growing line) × Surface stocking density (linear metres per square metre of cultivation area) × Total cultivation area (square metres).
9.3 Environmental interactions
9.3.1 Alteration of ecosystem and creation of habitat
Any man-made structure deployed in the marine environment will interact with the surroundings; seaweed farms are no exception. Physical interactions will include the interception of light and altering the path/rate of water movement, while biological interactions will include providing a new habitat for the settlement and attachment of organisms, the attraction of pelagic macrofauna such as fish for feeding and shelter, and consequently may also provide foraging opportunities for megafauna such as birds and cetaceans. The growth of the cultivated seaweed itself adds additional biological interactions, including the extraction of nutrients and carbon dioxide from the water, and the release of organic matter (as both dissolved exudates and fragments of the seaweed). This section focuses on the most important drivers of environmental change associated with the establishment of a seaweed farm (Figure 9.1; Campbell et al., 2019).
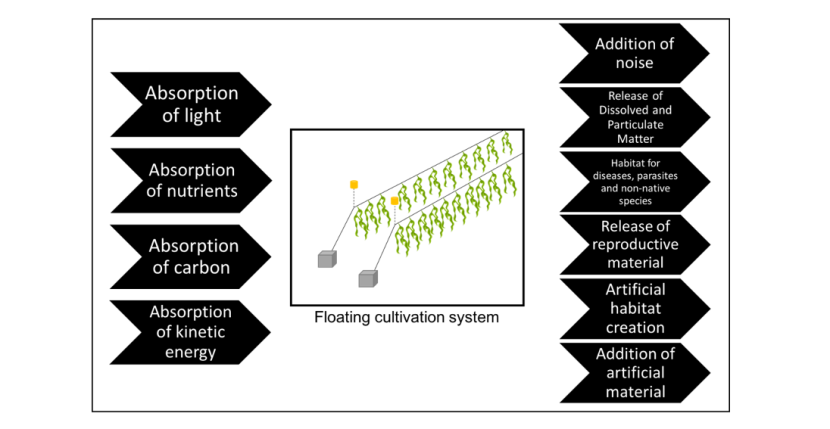
Figure 9.1. Drivers of environmental change in relation to seaweed farm. Figure taken from Campbell et al. (2019).
Click for a full description
The image shows four inputs that are absorbed into a floating seaweed cultivation system:
- light
- nutrients
- carbon
- kinetic energy.
It then lists six potential outputs from the system that may drive environmental changes:
- additional noise
- release of dissolved and particulate matter
- habitat for diseases, parasites and non-native species
- release of reproductive material
- artificial habitat creation
- addition of artificial material.
There is significant uncertainty when predicting the magnitude and consequences of seaweed aquaculture in a European context. All European research studies are being carried out on farms in the small-medium scale, which appear to have little detectable impact. Large-scale farms, with extensive research monitoring, are needed to fully understand the potential impacts of cultivation beyond farms sized at 50 x 200 m lines.
Summary and mitigation recommendations
- Seaweed farming will have a myriad of interactions with the local marine ecosystem, both physical and biological.
- Current research monitoring has been carried out on small-medium European farms (i.e. less than 50 x 200 m lines), with little detectable impact (many of these scientific studies have not yet been published).
- Large scale farms (e.g. ≥10 ha) with extensive research monitoring, are necessary to fully understand the interactions.
9.3.2 Absorption of light
Seaweed absorb light to power photosynthetic primary production for growth. During cultivation, the seaweed is suspended near the surface, resulting in reduced light penetration below the farm. The extent of reduction will be proportional to the density of seaweed at the surface (i.e. are lines close together or far apart) and how much biomass is present (lowest during out-planting and greatest just before harvest).
Dense seaweed farms could be considered to have a negative impact on photosynthetic organisms underneath the farm, such as natural kelp, maerl or seagrass beds. Fortuitously, many such shallow habitats are technically challenging to moor structures in place (due to increased wear from breaking waves and greater likelihood of beaching). To mitigate any potential negative impact, the following measures are suggested: a) sensitive habitats such as maerl and seagrass beds should be avoided; b) only farms with a low density of surface seaweed should be placed in shallow water (e.g. widely spaced lines or small species such as P. palmata, where >90% of the surface is open); and c) preferentially site farms in locations deep enough to avoid these shallow communities e.g. in water depths >25 m. At these depths, changes to the light are expected to have negligible effects.
The partial shading of the water column by a seaweed farm is expected to reduce the productivity of phytoplankton, although measurable effects are only expected when extensive areas are covered by seaweed farming. Reduced phytoplankton abundance is seen inside large cultivation areas during the growing season in China (Shi et al., 2011). In smaller farms, individual phytoplankton would only experience shading for the length of time they traverse a farm; possibly only for minutes.
Summary and mitigation recommendations
- Seaweeds absorb light for photosynthesis and reduce light availability below the farm.
- A simple mitigation measure for benthic impact will include: not siting farms over sensitive habitats, in shallow locations farms at a low stocking density and preferentially site farms in >25 m water depth.
- Phytoplankton impacts are only likely to occur at very large scales.
9.3.3 Absorption of nutrients
Seaweeds are extractive organisms that absorb nutrients for growth from the water that passes across their blade surfaces; this is unlike land plants where uptake is via the roots. Seaweed cultivation is a sink for inorganic nutrients, primarily dissolved inorganic nitrogen (Marinho et al., 2015). In many coastal locations, anthropogenic activities are adding excess nutrients into the marine system, such as the runoff from agricultural land, or treated outflow from cities, which is leading to eutrophication in some areas. Seaweed cultivation can have a positive bioremediation effect, whereby the seaweed removes a portion of these excess nutrients (Seghetta et al., 2016). In the case of animal aquaculture, particularly finfish, seaweed can be cultivated to ‘soak up’ the equivalent amount of released nutrients, as integrated multi-trophic aquaculture (IMTA). This cultivation does not need to be directly adjacent to the finfish farm, but should be in the same water body (within km), preferably connected by tide.
Under typical coastal conditions in Scotland, seaweed cultivation requires no fertiliser addition, as nutrients are replenished by natural processes. However, when the refreshment of water is restricted, such as in some lochs, nutrient removal may exceed replenishment and so limit seaweed growth. Therefore it is always preferable to cultivate seaweed where there is some current flow. At a very large-scale (many km2), such as in China, seaweed cultivation can cause a water body to become nutrient-limited towards the interior of the farm (Shi et al., 2011). This will reduce the natural primary productivity in the area by phytoplankton and wild benthic seaweeds/seagrasses. This has been modelled by Aldridge et al. (2012) in the coastal UK. In such circumstances, the addition of a nutrient source may be necessary to redress the imbalance. While the addition of refined inorganic nutrients has been used in China, a more sustainable method, both ecologically and economically is the use of IMTA. For example in Sango Bay China, roughly 640,000 tonnes of seaweed are grown, along with 132,000 tonnes of oyster, scallop, abalone and sea cucumber.
Summary and mitigation recommendations
- Seaweed absorbs nutrients naturally available in the water with no fertiliser required.
- Position farms in areas with good water refreshment, especially as the scale increases.
- Seaweed cultivation can remove excess nutrients added by human activity (bioremediation).
- Cultivation in water bodies with existing animal aquaculture can improve their environmental credentials
- At very large scales, farms may remove excessive amounts of nutrients (other scales will have negligible impact).
- To redress the imbalance, animal aquaculture could be incorporated into the farm as IMTA (i.e. finfish shellfish).
9.3.4 Absorption of kinetic energy
Both the farming infrastructure and suspended blades of seaweed will absorb and deflect the kinetic energy produced by waves and tides. This will alter the flow of the water body around the farm. While the total tidal exchange volume has been shown to remain unchanged, the surface flow can be reduced, whilst flow below the farm may accelerate (as has been seen in Sanggou Bay, China and mussel aquaculture; Plew et al. 2005; Zeng et al., 2015). Modelled flows in Sanggou Bay show a 20% flow reduction in the open channels, and a 54% reduction among the seaweed lines (Grant & Bacher, 2001). Alteration of water flow will likely be greatest at peak biomass (May-June) and at higher stocking densities.
Seaweed cultivation can therefore alter coastal hydrology, although this is unlikely to cause a measurable impact on the local environment at small-medium scale cultivation. For large-scale projects, a strategic siting (e.g. high flow or offshore) and modelling approach may be required to ensure licensing authorities are able to make informed decisions about the consequences. A similar modelling approach would also be appropriate to understand the potential cumulative impacts of smaller co-located projects.
Summary and mitigation recommendations
- Seaweed farms will absorb kinetic energy and slow water flow within the farm.
- For the water mass to balance, flow will accelerate underneath and to the side of the farm.
- When consenting large farms, some consideration should be given to the likelihood of appreciable changes in water movement around the farm and the extent of habitat and sensitivity of species “down-stream” of the farm.
9.3.5 Release of Dissolved and Particulate Organic Matter (DOM/POM)
Seaweeds naturally release dissolved exudates into the surrounding water. These are mainly sugars/carbohydrates produced by photosynthesis that are surplus to requirements for growth. In natural kelp beds, these can represent 18 to 35% of total carbon fixed from CO2 (Krause-Jensen & Duarte, 2016). This is an important source of DOM to the coastal ecosystem and may be exported far beyond the immediate habitat. Cultivated seaweed will also produce DOM, but it is not yet known if this will differ to natural seaweed communities.
Particulate organic matter (POM) is any larger particle, from tiny specks to whole plants. Many seaweeds naturally fragment at their distal ends as they age. They will continually release small particles into the surrounding environment, and have been shown to contribute over 90% of POM production in Sanggou Bay (Zhang et al., 2011). This release is highly seasonal, increasing as the cultivated biomass increases (i.e. highest in summer near to harvest, lowest in autumn when outplanting). During storms, POM production can dramatically increase, including larger pieces and whole plants. In natural kelp beds, 16-32% of the total fixed carbon may be released as POM (Krause-Jensen & Duarte, 2016).
Due to the low material density of seaweed, the released POM is often carried away from the site suspended in the water column, before settling on the benthos and decomposing (Ren et al., 2014). This means there is unlikely to be any impact of released POM on the benthos immediately surrounding the farm. Research is currently underway, but it appears that much of the larger POM from natural kelp beds settles in seabed depressions further offshore. Similar to DOM, it is not understood if POM differs between cultivated and natural seaweed. Nevertheless, against the background of organic material naturally released in coastal habitats, it is expected that only large to very large scale cultivation would result in any significant change in the release load. Organic matter generated at smaller scale will be readily recycled through natural processes.
Summary and mitigation recommendations
- Cultivated seaweed will release dissolved and particulate organic matter (DOM/POM; mainly carbohydrates) into the surrounding environment.
- This release will increase with the total biomass i.e. highest in summer before harvest.
- Coastal seaweed beds naturally release substantial quantities of DOM and POM to surrounding habitats.
- Seaweed cultivation is unlikely to impact this process, except at large to very large scales.
9.3.6 Release of plastics into the marine environment
Farm infrastructure is usually made of plastic ropes and buoys, as it is a durable material suitable for marine use. This creates a risk of releasing plastic pollution into the environment. When a farm is a) constructed effectively to secure materials in place and prevent wear points; and b) managed correctly, including regular site inspection, there should be no risk of ropes detaching and drifting away from the site. The connection of buoys tend to be a focus for wear, and so there is a risk of buoy loss from the site, particularly during storm events.
Plastic ropes are made from filament strands, and so their wear will cause the release of small plastic particulates/ fibres (mm-cm scale). Furthermore, these strands will be broken down into smaller fragments over time in the marine environment, becoming a source of microplastic pollution. Seaweed growing on plastic rope is not thought to degrade the rope structure (i.e. seaweed growth itself does not appear to cause the detachment of plastic fibres). Nevertheless, the release of plastics from seaweed cultivation has never been quantified. In the absence of evidence, technologies that minimise the loss of plastic and promoting the development of new biodegradable natural fibres such as bioplastics should be encouraged where possible.
In Scotland, the Crown Estate Scotland, has introduced contractual terms (Annual sustainability reporting) which require reporting of the management of plastic used on the leased seabed area, to demonstrate co-existence and stewardship of the marine environment.
Summary and mitigation recommendations
- Seaweed farms introduce plastics into the marine environment, mainly in the form of ropes and buoys.
- The loss of rope from the farm is highly unlikely if properly secured and managed. However, a minor risk of material loss will exist.
- The wear of plastic components will release small particles into the marine environment.
- New technologies to minimise plastic loss and the use of biodegradable materials should be encouraged.
9.3.7 Entanglement risk
The incidental mortality of air breathing marine megafauna (e.g. marine mammals and marine turtles) via entanglement in subsurface mooring lines and fishing gears is a significant conservation concern (Benjamins et al., 2014). The development of seaweed cultivation will create structures that may pose a threat to marine megafauna through entanglement.
The International Council for the Exploration of the Sea (ICES) summarises reports of entanglement events (ICES WGMME, 2014). Post-mortem examinations of strandings are unable to determine whether drowning/entanglement are caused by aquaculture gear rather than fishing gear. However, factors that increase the risk of entanglement include: the behaviour of particular species and their size, the location and depth of moorings, and the presence of low tension or loose lines and other structures that can entangle such as nets.
Until it can be understood how marine mammals use and interact with seaweed cultivation structures, caution should be exercised to ensure cultivation systems are devoid of significant entanglement risks. Any incidences of entanglements should be reported to the UK Cetacean Strandings Investigation Programme (CSIP). It would be prudent to undertake post-mortem examinations of entanglement events at farms to determine the cause of death, to enable lessons to be learnt and changes to recommendations made in an informed way.
Summary and mitigation recommendations
- Entanglement of marine megafauna by seaweed cultivation infrastructure is a possibility, but not well understood.
- Any entanglements should be reported, to inform policymakers on the risk posed.
9.3.8 Disease and pest management
The prevalence of disease and pests affecting aquaculture production is a major global concern (Kim et al., 2014; Stentiford et al., 2017). This issue is intensified by practices that reduce genetic diversity during the domestication of wild seaweed species, making crops more susceptible to abiotic stressors, disease and parasites (Valero et al., 2017). Unlike terrestrial agriculture, reduced crop vigour through domestication of seaweed stocks cannot be supported by the use of pesticides and fertilisers to facilitate growth (due to the open water nature of seaweed cultivation systems) (Barbier et al., 2019). The current approach to sourcing seed in Scotland is precautionary, working to cultivate native ‘locally sourced’ seedstock. The approach traditionally used in many of the major seaweed cultivating nations has been far less precautionary (Barbier et al., 2019). Increased knowledge of the genetic diversity of native species can support the sustainable development of reliable seed in a manner that can ensure sufficient genetic diversity and the promotion of disease resistance.
Cultivated stands will likely experience a large reduction in yield where disease and pests are prevalent, and may also act as a source for disease that could impact natural populations (Loureiro, et al., 2015; Valero et al., 2017). For example, carrageenophyte (Kappaphycus spp.) producing countries have seen a dramatic decline in production following rising sea water temperatures, which causes bleaching of the thallus that in turn makes cultivated individuals more susceptible to infection from viruses and bacteria (Vairappan et al., 2008).
The current knowledge of the epidemiology of seaweed pathogens is very poor, and in many cases the pathogens responsible for disease are difficult to identify and study using current microbial methods (Gachon et al., 2010). Further investigation is required to inform appropriate mitigation measures and prevent significant ecological impacts. Effective biosecurity planning will ensure actions are taken that mitigate risk where practical and will benefit all stakeholders. Ongoing research initiatives such as the Horizon 2020 project GENIALG (GENetic diversity exploitation for Innovative macro-ALGal biorefinery) and the GCRF GlobalSeaweedSTAR programme aim to address this knowledge gap. The research programmes aim to characterise diseases and pests, and to develop appropriate biosecurity protocols to monitor and understand the prevalence of disease in seaweed aquaculture.
Summary and mitigation recommendations
- Disease/pathogens of seaweed exist, but are currently poorly understood.
- Outbreaks are likely to cause reductions in yield and may spread to neighbouring farms and ecosystems.
- Biosecurity planning is necessary to reduce practical risks.
- Seaweed with suspected incidence of disease or pathogens can be sent for testing.
9.3.9 Non-Native Species and Biosecurity Plans
Non-Native Species (NNS) are defined as organisms that have been intentionally or unintentionally introduced outside their native range as a consequence of human activity. NNS may cause ecological damage to the receiving environments and may also be associated with economic losses within affected marine industries, including aquaculture (Pimentel et al., 2001). Once established, species that threaten biodiversity and/or cause economic damage are referred to as Invasive Non-Native Species (INNS). It is widely accepted that once a NNS has been introduced to a new environment, it is very challenging (and in the majority of cases practically unfeasible) to eradicate. Preventing the introduction of new NNS and restricting the likelihood of secondary introductions should be considered alongside the process of disease biosecurity planning and prevention.
Artificial structures used to cultivate seaweed, provide a novel habitat that may favour the establishment of NNS (Glasby et al., 2007; Mineur et al., 2012). In addition, the presence of pathways by which NNS can be further spread (e.g. movement of biofouling attached to vessels and other structures) must be managed to reduce the potential likelihood of occurrence. The intentional cultivation of non-native seaweed species is not allowed in Scotland (Marine Scotland 2017). Although the risk of unintentional introductions can never be fully managed, it is very unlikely that the cultivation of native species will carry a risk of introducing NNS. It is essential that the operation of both the hatchery and farm are managed to reduce any potential risks through biosecurity planning. Any observations of NNS or INNS should be recorded, and reported
The current guidelines for biosecurity planning that NatureScot follow will also apply to seaweed cultivation (Cook et al., 2014). To summarise, the principle component of any Biosecurity Plan is a record of the actions that will be taken in order to minimise the spread of NNS. Biosecurity Plans can be combined with disease management plans to increase efficiency. The ultimate goal of a Biosecurity Plan is to help an organisation manage its activities to minimise the risk of introducing disease, pests and NNS out-with their natural range. A Biosecurity Plan consists of four main stages.
- A Description of the Activity: A description of the planned activities to aid the Risk Assessment and subsequent review of the Biosecurity Plan.
- Risk Assessment: Identification of the level of risk associated with the planned activities (as described in Step 1).
- Resulting actions: List of steps to be followed after consideration of the risks above.
- Contingency plan: What should be done if a problem is identified?
A Biosecurity Plan should contain a set of instructions for personnel to follow including the production of a contingency plan (as part of the Resulting Actions). It should be noted that there is a potential challenge in expecting the average seaweed farmer to possess the practical knowledge to undertake assessments for NNS. Establishing training programmes to raise the awareness and capabilities of farmers could be useful initiatives to ensure that these steps can be conducted appropriately.
Summary and mitigation recommendations
- As with all marine activities, the infrastructure of a seaweed farm may provide a habitat for the establishment and spread of non-native species.
- The cultivation of native seaweed species carries a very low risk of introducing non-natives.
- A record should be kept of any non-native or invasive species and reported where necessary.
- A biosecurity plan should be in place for both seaweed farms and hatcheries. This should be combined with disease management plans.
9.3.10 Management of environmental interactions
If properly managed, seaweed cultivation can provide ecosystem services whilst developing marine resources. Current cultivation sites are typically small-medium scale, and if located and managed with consideration to environmental receptors, pose a low risk to the receiving environment and associated features (Wood et al., 2017; Campbell et al., 2019). However, an expansion of the industry towards large-scale sites will necessitate a more complete understanding of potential impacts in order to fully assess and manage risk. Targeted research and monitoring is required to address knowledge gaps in order to facilitate informed decision making during the consenting process. The production of biologically-coupled hydrodynamic models can be used to support the assessment of risk, help understand the carrying capacity of water bodies, and aid in selecting suitable sites that minimize negative environmental changes. Table 9.1 (from Campbell et al., 2019) illustrates a hypothetical risk assessment and mitigation options for each of the identified impact pathways.
Table 9.1. A risk assessment of the potential environmental impacts from the establishment of a hypothetical large scale seaweed cultivation site (100 ha). Risk scores summarise only negative changes and are based on the potential vulnerability of associated receptors along with the magnitude of potential change (0 = no negative consequences likely, 1 = consequences possible for some receptors, 2 = negative consequences possible for receptors requiring mitigation, 3 = widespread negative consequences possible for receptors requiring mitigation, 4 = negative consequences possible for protected receptors requiring mitigation, 5 = widespread negative consequences possible for protected receptors leading to consent being unlikely). The uncertainty score relates to the current level of uncertainty associated with the predicted risk and assumes that steps have been taken to mitigate risk. Future monitoring, and research needed to address uncertainty are highlighted. Taken and adapted from Campbell et al. (2019).
(a) High impact drivers of environmental change
Drivers of environmental change | Potential impacts to associated receptors | Risk score before mitigation | Possible positive changes | Principle measure(s) to mitigate impacts | Risk score after mitigation | Uncertainty | Future monitoring and research requirements |
|---|---|---|---|---|---|---|---|
| Release of reproductive materials |
| 0-5 | - |
| 0-3 | High |
|
| Facilitation of disease, parasites and non-native species. |
| 0-5 | - |
| 0-3 | High |
|
| Absorption of kinetic energy |
| 0-5 |
|
| 0-2 | High |
|
| Addition of cultivation systems |
| 0-5 |
|
| 0-1 | High |
|
| Nutrient absorption |
| 0-4 |
|
| 0-1 | Low-medium |
|
Drivers of environmental change | Potential impacts to associated receptors | Risk score before mitigation | Possible positive changes | Principle mitigation measure(s) to negative impacts | Risk score after mitigation | Uncertainty score | Future monitoring and research requirements |
|---|---|---|---|---|---|---|---|
| Artificial habitat creation |
| 0-3 |
|
| 0-2 | High |
|
| Absorption of light |
| 0-3 |
|
| 0-1 | Low |
|
Drivers of environmental change | Potential impacts to associated receptors | Risk score before mitigation | Possible positive changes | Principle mitigation measure(s) to negative impacts | Risk score after mitigation | Uncertainty score | Future monitoring and research requirements |
|---|---|---|---|---|---|---|---|
| Release of Particulate and Dissolved Organic Matter |
| 0-2 |
|
| 0-1 | Medium |
|
| Creation of noise |
| 0-2 | - |
| 0-1 | Low | - |
| Addition of cultivation systems |
| 0-1 | - |
| 0-1 | Low | - |
| Absorption of carbon |
| 0 |
| - | 0 | Low | - |
10. Final recommendations
This document provides an overview of the current procedures used for seaweed cultivation in Europe, and how these may develop over time with new advances in research and technology (e.g. new species, binder seeding and harvest mechanisation). Further details on many of these aspects can be found by reading the references given. There is ongoing research into the interactions between seaweed farming and the surrounding marine environment. This document provides reasoned guidance, based on current evidence, into the most important interactions and possible mitigation activities. Many impacts may not be detectable before the scale of the farm reaches large (≥10 ha) to very large (km2) size. The trial of such developments should be granted alongside appropriately designed monitoring programmes to allow quantitative data to be collected to inform policymakers, and to address gaps in knowledge.
Seaweed cultivation in Europe is a rapidly developing field. There are numerous national and transnational research projects ongoing throughout the European area and a growing number of companies are applying to begin commercial seaweed farming around the UK. It is anticipated that this document will require considerable revision within a few years and a substantial rewrite within five years due to both the rapid rate of industry development and increasing knowledge base gained via research programmes.
11. References
Aldridge, J., Van Der Molen, J. & Forster, R. 2012. Wider implications of macroalgal cultivation. Report for the Crown Estate.
Augyte, S., Lewis, L., Lin, S., Neefus, C.D. & Yarish, C. 2018. Speciation in the exposed intertidal zone: the case of Saccharina angustissima comb. nov. & stat. nov. (Laminariales, Phaeophyceae)’, Phycologia, 57(1), 100–112.
Bailey, J. 2016. The History of Alginate Extraction. Grace’s Guide to British Industrial History.
Barbier, M., Charrier, B., Araujo R., Holdt, S.L., Jacquemin, B. & Rebour, C. 2019. Phycomorph European Guidelines for a Sustainable Aquaculture of Seaweeds. COST Action FA1406 (Barbier, M. & Charrier, B. Eds.), Roscoff, France.
Benjamins, S, Harnois, V., Smith, H.C.M., Johanning, L., Greenhill, L.,Carter, C., & Wilson, B. 2014. Understanding the Potential for Marine Megafauna Entanglement Risk from Marine Renewable Energy Developments. Scottish Natural Heritage Commissioned Report No. 791.
Bernard, M., Rousvoal, S., Collet, N., Le Goff, T. L., Jacquemin, B., Peters, A., Potin, P. & Leblanc, C. 2019. A highly prevalent filamentous algal endophyte in natural populations of the sugar kelp Saccharina latissima is not detected during cultivation in Northern Brittany. Aquatic Living Resources, 32 (21), 1-6.
Billot, C., Engel, C.R., Rousvoal, S., Kloareg, B. & Valero, M., 2003. Current patterns, habitat discontinuities and population genetic structure: the case of the kelp Laminaria digitata in the English Channel. Marine Ecology Progress Series, 253, 111–121.
Breton, T.S., Nettleton, J.C., O’Connell, B. & Bertocci, M. 2018. Fine-scale population genetic structure of sugar kelp, Saccharina latissima (Laminariales, Phaeophyceae), in eastern Maine, USA. Phycologia, 57(1), 32–40.
Brunner, L. 2015. Safe harvesting and handling of seaweeds for human consumption: A guide to some edible or food-ingredient seaweed species.
Bruton, T., Lyons, H., Lerat, Y., Stanley, M. & Rasmussen, M.B. 2009. A Review of the Potential of Marine Algae as a Source of Biofuel in Ireland. Sustain. Energy Irel. Dublin 88.
Burrows, M.T. 2012. Influences of wave fetch, tidal flow and ocean colour on subtidal rocky communities. Marine Ecology Proess. Series, 445, 193–207.
Campbell, I., Macleod A., Sahlmann, C., Neves, L., Funderud, J., Øverland, M., Hughes, A.D. & Stanley, M. 2019. The Environmental Risks Associated With the Development of Seaweed Farming in Europe - Prioritizing Key Knowledge Gaps Frontiers in Marine Science, 6, 107.
Cook, E.J., Payne, R. & Macleod, A. 2014. Marine Biosecurity Planning-Identification of Best Practice: A Review. Scottish Natural Heritage Commissioned Report No. 748.
Cottier-Cook, E. J., Nagabhatla, N., Badis, Y., Campbell, M.L., Chopin, T., Dai, W., Fang, J., He, P., Hewitt, C.L., Kim, G.H., Huo, Y., Jiang, Z., Kema, G., Li, X., Liu, X., Liu, H., Liu, Y., Lu, Q., Luo, Q., Mao, Y., Msuya, F.E., Rebours, C., Shen, H., Stentiford, G.D., Yarish,C., Wu, H., Yang, X., Zhang, J., Zhou, Y. & Gachon, C.M.M. 2016. Safeguarding the future of the global seaweed aquaculture industry. United Nations University (INWEH) and Scottish Association for Marine Science Policy Brief.
Day J. 2018. Cryopreservation of Macroalgae. In Charrier B., Wichard T. & Reddy C.R.K. (eds) Protocols for Macroalgae Research. CRC Press, Boca Raton, Florida, 496 p. ISBN-13: 978-1498796422.
Durrant, H.M.S., Barrett, N.S., Edgar, G.J., Coleman, M.A. & Burrridge, C.P. 2018. Seascape habitat patchiness and hydrodynamics explain genetic structuring of kelp populations. Marine Ecology Progress Series, 587, 81–92.
Edwards, M. & Watson, L. 2011. Aquaculture Explained No.26: Cultivating Laminaria Digitata. BIM: Irish Sea Fisheries Board, Ireland. 71 pp.
FAO. 2016. The State of World Fisheries and Aquaculture 2016. Contributing to Food Security and Nutrition for All. Rome, Italy. 200 pp.
FAO, 2019. Fisheries and Aquaculture Information and Statistics Services.
Flavin, K., Flavin, N. & Flahive. B. 2013. Kelp Farming Manual. A Guide to the Processes, Techniques, and Equipment for Farming Kelp in New England Waters. Ocean Approved.
Gachon, C.M.M., Sime-Ngando, T., Strittmatter M., Chambouvet, A. & Kim G.H. 2010. Algal Diseases: Spotlight on a Black Box. Trends in Plant Science, 15(11), 633–640.
Glasby, T.M., Connell, S.D., Holloway, M.G. & Hewitt, C.L. 2007. Nonindigenous Biota on Artificial Structures: Could Habitat Creation Facilitate Biological Invasions? Marine Biology, 151(3), 887–895.
Grant, J., & Bacher, C. 2001. A Numerical Model of Flow Modification Induced by Suspended Aquaculture in a Chinese Bay. Canadian Journal of Fisheries and Aquatic Sciences, 58(5), 1003–11.
Kenicer, G., Bridgewater, S. & Milliken, W. 2009. The Ebb and Flow of Scottish Seaweed Use. Botanical Journal of Scotland, 52(2), 119–48.
Kerrison, P.D., Stanley, M.S., Edwards, M.D., Black, K.D. & Hughes. A.D. 2015. The Cultivation of European Kelp for Bioenergy: Site and Species Selection Biomass and Bioenergy, 80, 229–42.
Kerrison, P.D., Stanley, M.S., Maeve, K., MacLeod, A., Black, K.D., Hughes, A.D. 2016. Optimising the settlement and hatchery culture of Saccharina latissima (Phaeophyta) by manipulation of growth medium and substrate surface condition. Journal of Applied Phycology, 28(2), 1181-1191.
Kerrison, P.D, Stanley, M.S. & Hughes, A.D. 2018. Textile Substrate Seeding of Saccharina Latissima Sporophytes Using a Binder: An Effective Method for the Aquaculture of Kelp Algal Research, 33 (Jul), 352–57.
Kim, G.H., Moon, K-H. Kim, J-Y., Shim, J. & Klochkova, T.A. 2014. A Revaluation of Algal Diseases in Korean Pyropia (Porphyra) Sea Farms and Their Economic Impact. Algae, 29 (4), 249-265
Krause-Jensen, D. & Duarte, C.M 2016. Substantial Role of Macroalgae in Marine Carbon Sequestration. Nature Geoscience, 9 (10), 737.
Kusumo, H.T., Pfister, C.A. & Wootton, J.T., 2006. Small-scale genetic structure in the sea palm Postelsia palmaeformis Ruprecht (Phaeophyceae). Marine Biology, 149(4), 731–742
Lenstra, WJ., van Hal, J.W. & Reith, H. 2011. Economic aspects of open ocean seaweed cultivation.
Laikre, L., Schwartz, M.K., Waples, R.S. & Ryman, N.. 2010 Compromising genetic diversity in the wild: Unmonitored large-scale release of plants and animals. Trends in Ecology and Evolution, 25(9), 520–529.
Loureiro, R., Gachon, C.M.M., & Rebours C. 2015. Seaweed Cultivation: Potential and Challenges of Crop Domestication at an Unprecedented Pace. New Phytologist, 206(2), 489–492.
Marine Scotland, 2017. Seaweed Cultivation Policy Statement. Edinburgh, UK.
Marinho, G.S., Holdt S.L, Birkeland, M.J. & Angelidaki. I. 2015. Commercial Cultivation and Bioremediation Potential of Sugar Kelp, Saccharina latissima, in Danish Waters. Journal of Applied Phycology, 27(5), 1967-1973.
Mineur, F, Cook, E.J., Minchin, D., Bohn, K., MacLeod A. & Maggs, C.A. 2012. Changing Coasts: Marine Aliens and Artificial Structures. Oceanography and Marine Biology, 50, 189–234.
Molen, J van der, Ruardij, P., Mooney, K., Kerrison, P., O’Connor, N.E., Gorman, E., Timmermans, K., Wright, S., Kelly, M.& Hughes, A.D 2018. Modelling Potential Production of Macroalgae Farms in UK and Dutch Coastal Waters. Biogeosciences, 15(4), 1123.
Mooney, K.M., Beatty, G.E., Elsäßer, B., Follis, E.S., Kregting, L., O’Connor, N.E., Ridden, G.E. & Provan, J. 2018. Hierarchical structuring of genetic variation at differing geographic scales in the cultivated sugar kelp Saccharina latissima. Marine Environmental Research. Elsevier, (June), pp. 108–115.
Nielsen, M.-M. et al., 2016. Genetic diversity of Saccharina latissima (Phaeophyceae) along a salinity gradient in the North Sea-Baltic Sea transition zone. Journal of Phycology, 52(4), 523–531.
Pimentel, D., McNair, S., Janecka, J., Wightman, J., Simmonds, C., O’Connell, C., Wong, E., Russel, L., Zern, J., Aquino, T. & Tsomondo, T. 2001. Economic and Environmental Threats of Alien Plant, Animal, and Microbe Invasions. Agriculture, Ecosystems & Environment, 84 (1), 1–20.
Plew, D.R., Stevens, C.L., Spigel, R.H. & Hartstein, N.D., 2005. Hydrodynamic implications of large offshore mussel farms. IEEE J. Ocean. Eng. 30, 95–108.
Poseidon Aquatic Resource Management, 2017. Scottish Shellfish Development Critical Mass Model. Report Submitted to Crown Estate Scotland & Association of Scottish Shellfish Growers (ASSG), Nov 2017. Lymington, UK.
Ren, L., Zhang, J., Fang, J., Tang, Q., Zhang, M. & Du. M. 2014. Impact of Shellfish Biodeposits and Rotten Seaweed on the Sediments of Ailian Bay, China Aquaculture International, 22 (106), 811–19.
Rød, K.K. 2012. Sori Disinfection in Cultivation of Saccharina Latissima, Evaluation of chemical treatments against diatom contamination Norwegian University of Science and Technology. PhD thesis.
Robuchon, M., Le Gall, L., Mauger, S. & Valero, M.2014. Contrasting genetic diversity patterns in two sister kelp species codistributed along the coast of Brittany, France. Molecular Ecology, 23(11), 2669–2685
Rolin, C., Inkster, R., Laing, J., Hedges, J., & Mcevoy, L., 2016. Seaweed cultivation manual. Shetland Seaweed Growers Project 2014-16. Shetl. Seaweed Grow. Proj. 2014-16. NAFC, UHI.
Sanderson, J.C. 2006. Reducing the environmental impacts of sea-cage fish farming through cultivation of seaweed. PhD thesis, The Open University, UK & UHI Millenium Institute.
Schiel, D. R. & Foster, M. S. 2006. The population biology of large brown seaweeds : Ecological consequences life histories of multiphase in dynamic coastal environments. Annual Review of Ecology, Evolution and Systematics, 37(2006), 343–372
Schiener, P., Black, K.D., Stanley, M.S. & Green, D.H. 2015. The Seasonal Variation in the Chemical Composition of the Kelp Species Laminaria Digitata, Laminaria Hyperborea, Saccharina Latissima and Alaria Esculenta Journal of Applied Phycology, 27(1), 363–73.
Seghetta, M., Tørring, D., Bruhn, A. & Thomsen, M. 2016. Bioextraction Potential of Seaweed in Denmark - An Instrument for Circular Nutrient Management Science of the Total Environment, 563–564, 513–29.
Selkoe, K. A., D’Aloia, C.C., Crandall, E.D., Iacchei, M., Liggins, L., Puritz, J.B., von der Heyden, S.& Toonen, R.J. 2016. A decade of seascape genetics: Contributions to basic and applied marine connectivity. Marine Ecology Progress Series, 554, 1–19.
Shi, J., Wei, H., Zhao, L., Yuan, Y., Fang, J. & Zhang J. 2011. A Physical–Biological Coupled Aquaculture Model for a Suspended Aquaculture Area of China Aquaculture, 318(3–4), 412–24.
Stentiford, G.D., Sritunyalucksana, K., Flegel, T.W., Williams, B.A.P., Withyachumnarnkul, B., Itsathitphaisarn, O. & Bass, D. 2017. New Paradigms to Help Solve the Global Aquaculture Disease Crisis. PLoS Pathogens, 13(2), e1006160.
tom Dieck, I. 1993. Temperature Tolerance and Survival in Darkness of Kelp Gametophytes (Laminariales, Phaeophyta): Ecological and Biogeographical Implications Marine Ecology Progress Series, 100(3), 253–64.
Vairappan, C.S., Chung, C.S., Hurtado, A.Q., Soya, F.E., Lhonneur, G.B. & Critchley, A. 2008. Distribution and Symptoms of Epiphyte Infection in Major Carrageenophyte-Producing Farms. Journal of Applied Phycology, 20(5), 477–483.
Valero, M., Guillemin, M.L., Destombe, C., Jacquemin, B., Gachon, C.M.M., Badis, Y. Buschmann, A.H., Camus, C. & Faugeron, S. 2017. Perspectives on Domestication Research for Sustainable Seaweed Aquaculture. Perspectives in Phycology, February.
van der Molen, J., Ruardij, P., Mooney, K., Kerrison, P., O’Connor, N.E., Gorman, E., Timmermans, K., Wright, S., Kelly, M., Hughes, A.D. & Capuzzo, E. 2018. Modelling potential production of macroalgae farms in UK and Dutch coastal waters. Biogeosciences, 15, 1123–1147.
Werner, A. & Dring. M.J. 2011. Aquaculture Explained No.27: Cultivating Palmaria Palmata. BIM, Irish Sea Fisheries Board. Ireland. 75 pp.
Wood, D., Capuzzo, E., Kirby, D., Mooney-McAuley, K. & Kerrison, P. 2017. UK Macroalgae Aquaculture: What Are the Key Environmental and Licensing Considerations? Marine Policy, 83, 29–39.
Zeng, D., Huang, D., Qiao, X., He, Y. & Zhang, T. 2015. Effect of Suspended Kelp Culture on Water Exchange as Estimated by in Situ Current Measurement in Sanggou Bay, China. Journal of Marine Systems, 149, 14–24.
Zhang, J., Fang, J., Wang, W., Du, M., Gao, Y. & Zhang. M. 2011. Growth and Loss of Mariculture Kelp Saccharina Japonica in Sungo Bay, China. Journal of Applied Phycology, 24(5), 1209–16.





