Scottish Wildcat Action (SWA) Specialist Report - Ecology
This report should be cited as: Kilshaw, K., Campbell, R.D., Kortland, K. and Macdonald, D.W. (2023). Scottish Wildcat Action final report: Ecology. NatureScot, Inverness.
Authors and institutions: Dr Kerry Kilshaw, Wildlife Conservation Research Unit, Dr Roo Campbell, Scottish Wildcat Action / NatureScot, Kenny Kortland, Forestry and Land Scotland, Prof. David Macdonald, Wildlife Conservation Research Unit.
Year of publication: 2023

Keywords
Wildcat hybrid, GPS radio tracking, ecology, rest sites, habitat use, conservation threats.
Background to SWA
The Scottish Wildcat Conservation Action Plan (SWCAP) was published in 2013 following the input of over 20 partner organisations. This led to the establishment of the Scottish Wildcat Action (SWA) project that ran from 2015-2020, funded by the Scottish Government, National Lottery Heritage Fund and others. SWA operated with a team of project staff managed by NatureScot, and associated work was carried out by various partner organisations. The overall work programme was steered by a group made up of ten of the partners. The International Union for Conservation of Nature (IUCN) Cat Specialist Group reviewed the work of the SWA, and other ongoing wildcat conservation work, and provided recommendations for future action (Breitenmoser et al., 2019). A wide range of topics relevant to wildcat conservation were covered during the SWA, and have now been published as a series of reports, of which this is one. These outputs will inform the next phase of wildcat conservation work in Scotland, including the SavingWildcats ‘SWAforLife’ project that runs from 2019-2026.
Wildcats hybridise with domestic cats and we use a combination of morphology and genetics to distinguish wildcats from domestic cats and their hybrids. The method applied is generally determined by the practicalities of management. For example, it is much easier to have high confidence in the pelage scores from a sedated cat, than from a camera-trap image taken at night. Genetic and pelage results can only be generated jointly in certain scenarios. Therefore, identifications under different situations have different levels of confidence associated with them. We therefore set different thresholds for identification of wildcats based on the ability to distinguish pelage and genetic characteristics in different situations. The confidence hierarchy, and the definitions used in this report, are given below.

Wildcat definition
Click for a full description
A graphic showing increasing certainty of wildcat identity over five levels as more information on pelage and genetic status are available.
It reads:
Unless described otherwise, a wildcat in this report is defined as a cat with a 7PS pelage score of 19+ and/or that passes the genetic threshold for the genetic analyses. The consequences of applying different thresholds in different situations and management regimes are also examined.
A ‘wildcat’ in this report is defined at level 4 by a cat with a pelage threshold of 17+. Genetic information is not available for all cats and so is not used in population size assessments.
Background
Data on the basic ecology of the wildcat in Scotland is limited because difficulties in identification have meant much of the previous research focussed mainly on defining the wildcat (Hubbard et al., 1992, Daniels et al., 1998, Daniels et al., 2001, Kitchener et al., 2005, Macdonald et al., 2004, Yamaguchi et al., 2004, Beaumont et al., 2001, Driscoll et al., 2007, McEwing et al., 2011, Senn and Ogden, 2015, Kitchener and Easterbee, 1992). The study by Campbell (2015) using GPS collars enhanced the available literature on habitat, natal den use and locations of rest sites, previously collated using standard VHF radio tracking techniques (e.g. Corbett, 1979, Daniels, 1997, Scott et al., 1993), but was limited by the longevity of the collar. Here we use the most recently available GPS technology to collect detailed information on fine scale habitat use and spatial ecology of individuals, offering an insight into how they use the landscape daily and seasonally, which is not possible to obtain using any other method. Increasing our knowledge on ecological behaviour can also offer insights into how hybridisation occurs and identify threats and factors important for future conservation efforts of the wildcat.
Summary of work
- As part of the SWA fieldwork to collect samples for disease screening and genetic testing of wild-living cats within the SWCAP priority areas, we fitted some of the individuals sampled with GPS radio-tracking collars. This was to improve our understanding of wildcat ecology and behaviour and how this affects conservation management for wildcats.
- Camera trap survey work carried out by SWA identified several putative wildcats based on pelage. Trapping was carried out during Feb - Mar 2018, Nov 2018 - Mar 2019 and Dec 2019 - Mar 2020. Overall, 14 individuals were fitted with GPS collars, across four of the five priority areas; Morvern, Strathpeffer, Strathbogie and Angus Glens. Individuals were classified as wildcats based on their pelage score (Kitchener et al., 2005); cats with a pelage score ≥17 (up to a maximum score of 21) are classified as wildcats. Here, mean pelage score was 18 (range = 17-21).
- All of the collared individuals had a hybrid score (Q = proportion of wildcat genes) <0.75, failing to meet the genetic threshold of a wildcat; indicating they were wildcat hybrids. Mean Q score = 0.54 (range = 0.285-0.71). Although identified as wildcat hybrids genetically, collared individuals exhibited behaviour similar to wildcats’ radio-tracked in mainland Europe and Scotland, suggesting wildcat hybrids may occupy a similar ecological niche as wildcats here. Data collected is still therefore relevant for wildcat conservation management.
- Collars have lasted to date an average 281 days (range = 113 - 333) [not including data from two young males; Brodie, whose collar stopped working/came off after one month and Finley, whom we lost track of during COVID-related fieldwork restrictions and have only collected 15 days of data to date]. Two collars fell off earlier than anticipated and were redeployed on different individuals. Two males (Bogie and Clash) caught in Feb/March 2018 were re-collared in the 2018/2019 trapping season. Clash was collared for a third time in Jan 2020 and a female (Doll) was also recollared in January 2020. Two individuals (Finley and Woody) are still currently collared.
- Mean total home range size based on 95% Minimum Convex Polygons was 13.94 km2 for females (range = 4.42-28.59 km2, N= 4) and 18.15 km2 for males (range = 8.32-29.27 km2, N=8). Home range size for females was larger than previously reported and for both sexes, larger than reported for some mainland European wildcat populations.
- Home range size varied monthly and was largest for males between February and August (mean monthly home range size Feb-August= 12.79 km2) and for females between February and April (mean monthly home range size Feb-April = 11.93 km2). Female home range size decreased from the end of April due to decreased ranging behaviour when rearing kittens (mean monthly home range size May - Oct = 3.48 km2).
- Three individuals were collared more than once to examine home range size and use over a longer period. Annual home range size for Bogie (Strathbogie Priority Area) did not appear to vary notably during the two consecutive years he was collared for. In comparison, Clash’s home range size (Strathbogie Priority Area) decreased in 2020 compared to 2018 and 2019 and also shifted to more open farmland in the third consecutive year he was tracked for, likely in relation to the establishment of a home range by another collared individual, Emma, within part of his territory. Doll’s home range size was also notably smaller in 2020 compared to 2019. This could be explained in part by the presence of Finley, a new young male not previously seen in her home range and also by the increased presence of visitors and wild campers to the area in 2020 as a result of COVID-related travel restrictions.
- Two individuals in the Angus Glens (Markie and Doll) showed some seasonal shift in home range from forested areas in winter to more open heathland and upper grasslands in spring/summer, possibly in relation to prey availability.
- Two females showed evidence of giving birth; Fairburn (Strathpeffer Priority Area) around the 30th April/1st May 2018, in a derelict stone barn with piles of wood leaning against the walls. A second female, Emma (Strathbogie Priority Area) around the 5th/6th May 2019 on the edge of an area of clear-fell. Emma showed a marked change in activity patterns for several weeks after this period. As has been found by other studies, Emma moved kittens to other den sites after a few weeks. We think Fairburn lost her kittens after a few weeks based on her activity patterns.
- Wildcat hybrids exhibited habitat selection within their home ranges. Although habitat use varied by individual cat across the different Priority Areas, overall, collared individuals were significantly more likely to be found close to edge habitats (except for Fairburn, where distance to edge was not included as a predictor variable in the model). Pooling the individual RSF model data showed that wildcat hybrids in general were more likely to be found close to or in arable landscapes (especially areas with rabbits), broadleaved woodland, edge habitats and using low use roads/forest tracks. But differences also existed between different Priority Areas. For example, in the Angus Glens, wildcat hybrids were significantly likely to be found in or close to coniferous woodland, which was not the case in the other Priority Areas. Here, collared individuals regularly used areas of wind blow within coniferous woodland, forested tracks through the trees or followed the edges of coniferous woodland up the glens. In Strathbogie, all six collared individuals were often found close to or within farm buildings or around the edges of rural properties and showed a significant decrease in probability of a fix further away from these structures. In Morvern, both Sprat and Stonker showed a significant decrease in occurrence further away from mixed woodland. In this Priority Area in particular, there is a relatively substantial amount of mature mixed woodland that is well connected and highly structured, providing excellent cover and prey for cats.
- Wildcat hybrids used many different habitats and structures as rest sites including patches of gorse, hedgerows, wind blow, stone cairns, hay barns, dense vegetation, and woody debris in clear-fell. Across all collared individuals, several different rest sites were identified where cats spent three hours or more. Many of these were used more than once (mean = 40, SD = 33), some of which were very regularly used on an almost daily basis (e.g. a favourite hay barn), the remainder were only used once (mean = 73, SD = 45). Rest sites were spread across the individuals’ home ranges, often using several different “locations” within the same general area (e.g. a large area of gorse or farmyard). The number of rest sites used increased as the number of days a cat was monitored for increased, suggesting that rest sites are not a limiting factor. Note this may not be the case for natal dens where areas suitable for rearing kittens could be limited.
- The wildcat is a European protected species and is fully protected under the Conservation (Natural Habitats, &c.) Regulations 1994 (as amended). Under this legislation, with respect to natal dens (breeding den) or rest sites (dens or other structures used for shelter and resting), it is an offence to deliberately or recklessly “disturb a wildcat in a den or any other structure or place it uses for shelter or protection”, “disturb a wildcat while it is rearing or otherwise caring for its young”, or “obstruct access to a den or other structure or place wildcats use for shelter or protection or otherwise deny the animal use of that place”, it is also an offence to “damage or destroy a breeding site or resting place of such an animal (whether or not deliberately or recklessly). Current advice for developers is to look for natal dens or dens (rest sites) by initial walking over the site, then using camera traps (under licence) or genetic testing of samples such as hair or scats at any potential dens (rest sites) or natal dens to identify whether a wildcat is using them or not. However, experience from this and other studies shows that finding rest sites and natal dens is often difficult even with collared individuals, particularly if the individual is using an area of wind blow or large patch of gorse which is difficult to get into. In addition, the large number of locations used means that it is likely that some are being missed by the current survey methods. We therefore recommend that current advice to developers, foresters and land managers is revised.
- Regular use of farm structures by wildcat hybrids highlights the importance of neutering feral farm cats to reduce hybridisation, and the risk of potential secondary rodenticide poisoning through the consumption of rodents found in many farm buildings.
- Some individuals used heathland/grouse moorlands. Site visits to the areas used indicated they were not intensively managed and consequently had relatively high vegetation heights and a higher diversity of plants, providing cover and potentially supporting more prey species (e.g. mountain hare, water voles, ground nesting birds and field voles). But wildcats in grouse moorlands are at risk of accidental persecution as a result of difficulties in wildcat identification. Continued work with the game keeping industry to improve identification techniques and increase wildcat-friendly practices is therefore essential.
- Roads are a major cause of mortality for wildcats and the use of roads by wildcat hybrids here highlights an area of concern for wildcat populations in Scotland. Initiatives to identify areas of potential high risk for wildcats in Scotland should be undertaken to try and reduce road-related wildcat mortality.
Acknowledgements
The SWA project was supported by the National Lottery Heritage Fund.
It was also funded by the Scottish Government, NatureScot and the Royal Zoological Society of Scotland. Support and resources for associated work was also provided by all the Steering Group members: Cairngorms National Park Authority, Forestry and Land Scotland, National Museums Scotland (NMS), Scottish Land & Estates, NatureScot, Scottish Wildlife Trust, The National Trust for Scotland, The Royal (Dick) School of Veterinary Studies, The Royal Zoological Society of Scotland (RZSS), The Scottish Gamekeepers Association, The Scottish Government, Wildlife Conservation Research Unit (WildCRU). In addition, Jenny Bryce and Fiona Strachan helped develop the project while Eileen Stuart, Alan Bantick and Andrew Kitchener chaired the Steering Group.
Additional sponsors, funders and contributors to SWA work included Aigas Field Centre, Ailsa Black, British Association for Shooting and Conservation, Cairngorm Brewery Company, Chester Zoo, Roy Dennis Wildlife Foundation, John Muir Trust, Loch Lomond & The Trossachs National Park, Lukas Ruiter Production, MyPetOnline, NFU Scotland, petDetect, RSPB, Scotland The Big Picture, Swift Ecology, the Highland Council and Robertsons Trust.
We are also very grateful to the many academics, landowners and managers, farmers, foresters, keepers, captive collection managers, veterinary specialists and others who contributed valuable time and effort, members of the public who contributed funds and support, and in particular the many volunteers who got involved in the project.
Due to data protection/privacy, we are not able to name everyone we would like to, but we want you to know how much we have appreciated your help. This project would not have been possible without the support of the many, many dedicated SWA volunteers in collecting data on the location of wild-living cats in the priority areas, many of whom were committed to the project for several years and continue to be involved in wildcat conservation. Special thanks to you all. Without this background data, trapping and collaring of cats would have been much more difficult. Furthermore, the logistic support of the SWA and FLS field staff in getting collars on the cats was vital. We would like to thank the following, Dr. Keri Langridge, Nicola Tallach, Callum Hislop, Hebe Carus and Emma Rawling (SWA), Dr Roisin Palmer-Campbell, Kelsey Wilson, Kirsten Mair, Dr Alice Bacon and Matthew Wilson for their enormous efforts in trapping and collaring cats, also Dr Philip Bacon and Norman Davidson. Thanks also to the wonderful veterinary support (Huntly, Kirriemuir, Dingwall), the local FLS staff (Jackie Cumberbirch, Gareth Ventress, Gareth Mason, Karen Patterson, Henry Dobson amongst others), the rangers at Glen Doll, for all their assistance in access and interest in the wildcat. Other thanks to the gamekeepers and landowners who allowed access to their land for trapping and collaring purposes.
Other researchers providing information, discussion or useful comments over the past few years include Dr. Mike Daniels (John Muir Trust), Dr. David Hetherington (Cairngorms National Park Authority), Dr. Ro Scott, Adrian Davies (Naida Ecology), Dr. Andrew Hearn (WildCRU) and members of the EuroWildcat group, in particular, Dr Stefano Anile (Cooperative Wildlife Research laboratory, Southern Illinois University), Dr. Malte Gotz (Deutsche Wildtier Stiftung), Dr. Matthias Herman (OEKO-LOG), Dr. Lea Maronde (KORA) and Dr. Bernd Klutz for discussion on GPS collaring and Dr Sam Cushman.
This project was primarily funded through the Robertson Trust, with logistical and vehicular support from NatureScot and some additional collars funded by FLS. We’d also like to thank Dr. Marc Büntjen (e-obs) for his continued support in answering questions about the collars, Dr. Paul Johnson (WildCRU) for comments on the RSF models and Dr Christos Astaras and Despina Migli for discussions about analysis of the GPS and accelerometer data for rest site identification.
We are very grateful to Dr Marcus Rowcliffe, who undertook an independent review of an earlier version of this report and whose comments greatly improved this final version.
1. Introduction
Understanding the ecological behaviour of a species is important for its conservation management, especially in relation to understanding how landscape management practices might influence an individual’s behaviour and overall survival of the population. Collating data on fine scale habitat use for example can help identify habitats that are important to the species and habitats that are avoided and how the composition of the landscape can influence the way an individual uses it. In addition, information on spatial use can help identify individual-specific and species-specific threats and provide clues as to how to mitigate these.
The wildcat in Scotland, a sub-population of the European wildcat (Felis silvestris), is Britain’s only surviving native felid (Macdonald et al., 2004) and while currently classified under the International Union for Conservation of Nature (IUCN) Red List of Threatened Species as Least Concern (Yamaguchi et al., 2015), its status is not healthy by historical standards. Previously widespread across the UK, the decline of the wildcat commenced several centuries ago and is attributed largely to hunting for fur and sport/persecution (Kitchener, 1995, Lovegrove, 2007). Records of wildcats being hunted for sport or as vermin are found as far back as the 12th century (Hamilton, 1897) and persecution of all species considered vermin was particularly rife during the 16th – 18th centuries (Lovegrove, 2007). By the 1800s, the wildcat had become restricted to parts of Northern Scotland (Balharry and Daniels, 1998, Davies and Gray, 2010, Easterbee et al., 1991) with Harvie-Brown (1881) stating it was already extinct to the south and east of a line running from Oban, north east across to Glas Maol, north to Tomintoul and north west to Inverness; and was still present only in Northern Argyll, the mountainous western comers of Aberdeen and Banff, and more widely in Inverness-shire, Ross and Cromarty, Sutherland and Caithness. Langley and Yalden (1977) concluded that the early decline of the wildcat in England and Wales may have been due to a combination of deforestation and hunting, with the later decline in Scotland coinciding with the later stages of re-afforestation and most closely with the development of sporting estates from the mid-1800s because wildcats were considered destructive to game and lambs and were consequently controlled (Langley and Yalden, 1977, McOrist and Kitchener, 1994, Tapper, 1992).
Re-colonisation is thought to have been aided by the decline in gamekeeping during the first and second world wars and perhaps reforestation by the Forestry Commission (established in 1919). By the 1950s the wildcat had recolonised most of the area north of the central belt of Scotland (Jenkins, 1962, Kitchener, 1995), but it was not clear how many of these individuals were now hybrids. By the time the wildcat received legal protection in 1988 it is likely that the population was already in a perilous state. Via extrapolation of wild-living cat samples collected in the 1990s, the Scottish population was previously estimated to be roughly 400 (Macdonald et al., 2004). Estimates based on extensive camera trap surveys placed the population between 157 - 311 individuals (Kilshaw, 2015) and consequently Kilshaw (2015) suggested the Scottish population should have its status revised. More recently, Mathews et al. (2018) placed the population size at 200 (95% Confidence Intervals 30-430). Note however that both estimates use a wildcat definition that encompasses some hybridisation (see below) and are therefore likely to overestimate the population size.
Across their range European wildcats are thought to be primarily threatened by hybridisation with the feral domestic cat (Hubbard et al., 1992, Nowell and Jackson, 1996, Macdonald et al., 2004) and wildcat hybrids which threatens the genetic integrity and evolutionary persistence of wildcat populations (Randi et al., 2001, Pierpaoli et al., 2003). There are high levels of introgression at some localities in Europe, e.g. in Hungary (26% of sampled cats, Pierpaoli et al., 2003), in western Germany (43% of sampled cats, Hertwig et al., 2009) and in France (31% of sampled cats, Say et al., 2012), but appears to be greatest in Scotland (50% of sampled cats; Beaumont et al., 2001). However, in a re-analysis of hybridisation across their range (but with no samples from Hungary and very few from central Italy), Tiesmeyer et al. (2020) found levels of introgression were variable but generally low to moderate in most populations (≤5% in western, central and south east Europe, 13-16% in alpine areas). Exceptions to this were southern Spain and Scotland with 20% of the Iberian (n=53) and 100% of the Scottish wild-living cat samples (n=15) identified as hybrids (Tiesmeyer et al., 2020). This high value for Scotland is confirmed in the larger sample of cats tested during the current project (Kitchener and Senn, 2023). Wildcats have potentially been hybridising with domestic cats for 2-3000 years in the UK but evidence of hybrids was first documented in 1820 (Bewick, 1820) and later by Darwin (1875). The potentially negative impact of hybridisation was recognised by Cocks (1876), with later evidence that interbreeding occurred (Pitt, 1939, Gray, 1971, Corbett, 1979). Hybridisation may also however be considered a symptom rather than a cause of wildcat decline (Balharry et al., 1994), with wildcat declining due to other factors (Sumption and Flowerdew, 1985, Macdonald et al., 2004), so that they became greatly outnumbered by domestic and feral cats and hybridisation became unavoidable. In Scotland, hybridisation has resulted in complications in surveying and monitoring for the wildcat and as a result, limited data exists on both its basic ecology and the factors that might influence hybridisation (O'Brien et al., 2009, Macdonald et al., 2004).
1.1 Home range
Home range size estimates for the wildcat in Scotland are few. Based on VHF radio tracking data, Corbett (1979) estimated a mean home range size of 1.76 km2 (males) and 1.75 km2 (females) on Glen Tanar estate, in Deeside, where at the time rabbit populations were extensive. Whereas Scott et al. (1993) found male and female home range sizes to be larger on the Ardnamurchan peninsula in the west of Scotland (9.9 km2 and 6.3 km2 for males and females respectively) where rabbits were not present. Daniels' (1997) study in the Angus Glens looked at wild-living cats with different pelage markings and found mean home range sizes of 6.06km2 (males) and 1.72 km2 (females). More recently, using GPS radio tracking Campbell (2015) estimated mean home range size for wild-living cats across different study sites as 6.34 km2 (males) and 5.73 km2 (females).
Corbett (1979) found that there was some overlap in home range between males and females but none between females, whereas Daniels’ (1997) study found adult females overlapped home ranges by ~10%. Campbell (2015) also noted no contemporaneous overlap between adjacent adult females, except where one individual shifted home range ahead of denning. Females appear to remain faithful to their home ranges, with monthly fluctuations centred on a core area (Corbett, 1979, Daniels, 1997) indicating some degree of territoriality. Corbett (1979) noted that female home ranges were largest between February - April.
Home range use varies between individuals, with some traversing most of their ranges during nightly activity sessions, whilst others alternate between pine forest and scrub habitats within their range, spending a few days in each habitat type (Corbett, 1979). During winter wildcats may move down to valley floors to shelter within hay barns (Corbett, 1979).
1.2 Habitat use
Across its range the European wildcat has traditionally been associated with forests, reaching their highest densities in broad-leaved or mixed forests (Parent, 1975, Schauenberg, 1981, Stahl and Leger, 1992). Despite being referred to as a “forest cat”, studies across Europe and in Scotland have indicated that this is not always the case (Langley and Yalden, 1977, Campbell, 2015, Daniels, 1997, Scott et al., 1993), and that wildcats use a wide variety of habitats ranging from Mediterranean scrubland (Ragni, 1981, Lozano et al., 2003, Lozano, 2010), riparian forest, marsh boundaries and sea coastal habitat (Lozan and Korcmar, 1965, Heptner and Sludskii, 1972, Dimitrijevic and Habijan, 1977, Scott et al., 1993, Nowell, 1996) and display individual, sex-based and seasonal variation in habitat selection (Wittmer, 2001, Beugin et al., 2016, Ballesteros-Duperón et al., 2005, Oliveira et al., 2018b).
In general, wildcats require areas with two basic habitat types; the first are areas that provide shelter and resting places, normally a closed structure habitat such as woodland. The second consists of open patches such as grasslands or riparian areas for hunting (Wittmer, 2001, Corbett, 1979, Stahl et al., 1988, Biro et al., 2004). Riparian areas within forests as well as edge habitats often provide a higher diversity and abundance of small prey mammals than interior forest (e.g. Doyle, 1990, Crandall et al., 1994, Gomez and Anthony, 1998).
In Scotland, wildcats appear to prefer woodland and stream edges, pine plantations <30 years or >100 years old are preferred over middle-aged plantations, other habitats that provide cover and prey such as unimproved grassland and shrub habitat are also preferred (Corbett, 1979, Daniels, 1997, Scott et al., 1993, Campbell, 2015). Daniels (1997) found that wild-living cats avoided open grassland and heather moorland and used clear-fell in relation to its availability. In Glen Tanar however, Corbett (1979) noted that some wildcats made nocturnal forays over open heather moorlands adjacent to forests and sometimes became temporary residents in open moorland valleys during summer months. Such valleys always contained rabbits and rocky cairns, which wildcats used as dens. Easterbee et al. (1991) showed that wildcats selected mosaics consisting of open fields and reforested patches and are generally absent from areas of intensive cultivation.
Wildcats are also generally not found (either seasonally or at all) in areas where snow cover is greater than 50%, or more than 20-30cm deep (e.g. Dotterer and Bernhart, 1996, Mermod and Liberek, 2002), or where snow remains for more than 100 days of the year (Ragni, 1991). Under these conditions, wildcats will move to forested areas or lower altitudes where snow cover is less. For example, Corbett's (1979) study showed that if the snow cover was >0.5m deep then wildcats would hunt under the tree canopy of mature pine forests and gorse.
1.3 Natal dens and resting sites
The wildcat is a European protected species and is fully protected under the Conservation (Natural Habitats, &c.) Regulations 1994 (as amended). Under this legislation it is an offence to deliberately or recklessly “disturb a wildcat in a den or any other structure or place it uses for shelter or protection”, “disturb a wildcat while it is rearing or otherwise caring for its young”, or “obstruct access to a den or other structure or place wildcats use for shelter or protection or otherwise deny the animal use of that place”. It is also an offence to “damage or destroy a breeding site or resting place of such an animal (whether or not deliberately or recklessly). To reduce confusion, in this report, breeding sites are referred to as ‘natal dens’ and dens or any other structures or places used for shelter or protection or as a resting place are referred to as ‘rest sites’.
In Scotland, wildcat kittens are born between end of March – May with a second litter or later litter sometimes born between July – August (Daniels, 1997, Campbell, 2015, Matthews, 1941). Litter sizes range between 1-8 young, although more typically they give birth to between 2-6 kittens (Muntyanu et al., 1992, Kitchener, 1995, Campbell, 2015). Kittens emerge from cover between 4-6 weeks old and by 10-12 weeks they are hunting with their mother, natal sites are moved closer to hunting grounds as the kittens grow (Lindemann, 1953, Campbell, 2015). Using GPS collars, Campbell (2015) identified several wild-living cat natal dens including in clear-fell (under woody debris), wind blow, brash piles, boulder scree and rabbit burrows. In mainland Europe, litters have also been observed being born above ground without the use of nesting material (Condé and Schauenberg, 1974).
Corbett (1979) found that most wildcats had 2-3 regularly used day-time resting sites. The Campbell (2015) study found wild-living cats had a mean of 11 rest sites (range 1 – 34) that they used and that they tended to be closer to edge habitats. The Campbell (2015) study also highlighted how important gorse was as a resting site with 35% of rest sites being found within gorse habitat, followed by 27% in coniferous plantations. Also used were wind blow (patches of conifer plantation felled by winds, 5% of rest sites in the two individuals that used them) and farm buildings (half of the rest sites in the two individuals that used them). In western Scotland, Scott et al. (1993) found wildcats would rest by day in gorse thickets or young forestry plantations.
1.4 Activity patterns
Corbett (1979) found that although wildcats could be active in every hour of a 24hr period, most activity (93%) occurred between 1600 – 0200 hours, particularly in summer, whereas in winter, wildcats were active outside of this main period of activity as well, presumably in relation to daylight length. In general, Corbett (1979) concluded that wildcats were mainly nocturnal hunters. Daniels (1997) found females were more active than males and all cats were also more active at night than during the day, whereas Campbell (2015) noted only a slight tendency towards nocturnal behaviour. In Ardnamurchan Scott et al. (1993) found that wildcats could be active at any time but that daytime activity was dependent on having good cover (young forestry plantation, gorse). During the night wildcats would emerge from cover and make forays across open ground and around farmlands, often covering 10km or more.
There is some evidence that wildcat activity is negatively influenced by poor weather conditions with wildcats remaining inactive for up to 28 hours during bouts of heavy snow (Corbett, 1979) and reduced activity during heavy rainfall (RD Campbell, Unpublished data).
1.5 Diet
In the north east of Scotland, where rabbit density is greater than in the north west of Scotland, rabbits form a major prey item (Corbett, 1979, Daniels, 1997, Delahay et al., 1998) making up to 90% of the diet in some cases. Where rabbits occur, juvenile rabbits are preferentially taken in spring (Corbett, 1979). In areas such as the west of Scotland, where rabbits are absent or less abundant, their diet is mainly rodent based (Hewson, 1983, Scott et al., 1993) with birds making up a small percentage (~11%) of the overall diet (Delahay et al., 1998). Lozano et al. (2006) reviewed the diets of wildcat across their European range and concluded that wildcat are facilitative specialists, taking rabbits where available, rodents if not and then other prey.
2. Aims and objectives
Under the Scottish Wildcat Conservation Action Plan, “ecology” encompasses the following actions:
3.4 Improve our understanding of wildcat ecology and behaviour as affects their conservation.
3.4.1 Improving our understanding of where and why hybridisation occurs.
To date only four radio tracking studies have been carried out on wildcats and wild-living cats in Scotland over the past 40 years. Three of these studies were carried out using traditional VHF radio tracking (Corbett, 1979, Daniels, 1997, Scott et al., 1993) and have provided a huge amount of data on wildcats and wild-living cats as detailed above. More recently, Campbell (2015) tested the use of GPS collars on wild-living cats and showed how important GPS technology could be in detailing fine scale ecological behaviour. In particular, the Campbell (2015) study significantly improved on the available literature on Scottish wild-living cat habitat and natal den / rest site use. The study was limited to some extent however by the longevity of the collars (mean = 33 days; range = 7-134 days).
Here we use the most recent GPS technology to collect longer scale (~9-12 months) detailed information on the fine scale habitat use and spatial ecology of individuals, offering a unique insight into how they use the landscape daily and seasonally, which is not possible to obtain using any other method. In addition to increasing our knowledge of the wildcat’s ecological behaviour and how this potentially influences hybridisation, we aim to use the data collated to identify specific threats / issues that are relevant for wildcat conservation.
3. Study Location / Methodology
Wildcat hybrids were collared in four of the six Scottish Wildcat Action Priority Areas (see Figure 1); a brief description of the four areas is provided below.
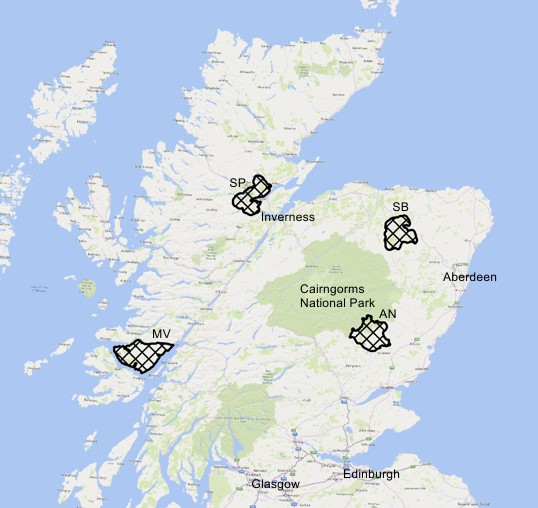
Figure 1. Map of northern Scotland showing the four Priority Areas wildcat hybrids were collared in (AN – Angus Glens, SB – Strathbogie, SP – Strathpeffer and MV – Morvern).
AN = Angus Glens
The Angus Glens Priority Area (56°48’N, 3°09’W) has a low human population and road density and relatively high elevations (up to 900m). Characterised by high mountains bordering parallel lowland glens, much of the area is either open heathland managed for grouse and scattered areas of woodland, including several areas managed by Forestry and Land Scotland for timber. Farmland can be found in the lowland areas with mostly rough grazing used for sheep and cattle grazing, with scattered villages and rural farms. In the lower parts of the glens, rabbits can be found in some areas at very high density.
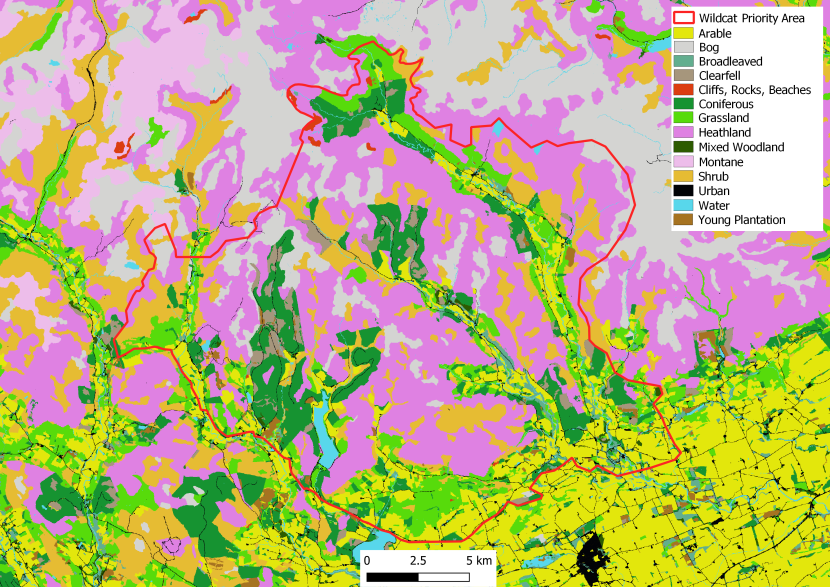
Figure 2. Landcover for the Angus Glens Priority Area.
Click for a full description
Landcover reclassed from the EUNIS dataset (obtained under Open Government Licence according to Table 1, cliffs and scree added but not included in Table 1. This is a visual representation of the habitat described for the Angus Glens Priority Area above.
SB = Strathbogie
The Strathbogie Priority Area (57°23’N, 2°50’ W) stretches east and west of the River Bogie and south of the A96. Edging the Grampian Mountains, the area has several high peaks (up to ~600m) and fertile lowlands, much of which is either grazed or used for arable farming. Large patches of woodland managed by Forestry Land and Scotland for timber dominate the area, with scattered patches of broadleaved woodland. Rabbits can mainly be found to the south and east of the Priority Area. Small holdings and farms are found throughout and the main town of Huntly sits on the north of the Priority Area. There is good mosaic of habitat with lots of cover and corridors for cats in the form of hedgerows, fences and stone walls. There is a relatively dense network of mostly low - medium use roads crossing the area but the A96, a busy A road, cuts across the northern end of the Priority Area.
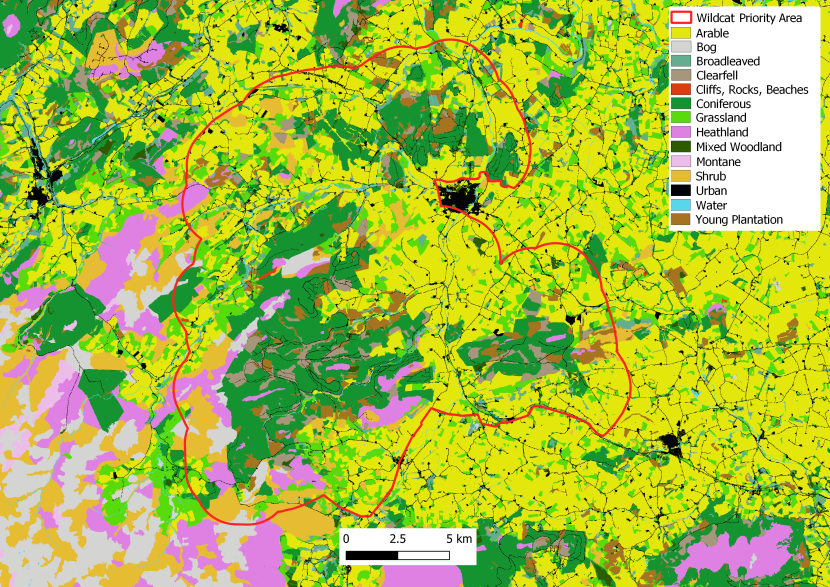
Figure 3. Landcover for the Strathbogie Priority Area.
Click for a full description
Landcover reclassed from the EUNIS dataset (obtained under Open Government Licence) according to Table 1, cliffs and scree added but not included in Table 1. This is a visual representation of the habitat described for Strathbogie Priority Area above.
SP = Strathpeffer
Strathpeffer Priority Area (57°30’N, 4°31’W) has high human population densities and a busy network of roads, some quite high use, crossing the area. Historically many wildcats were found here. Several large villages are found throughout including Beauly and Strathpeffer, both are surrounded by wooded hills, either coniferous or broadleaved, some high areas (peaks up to ~1000m) are characterised by open heathland and lowlands are mainly farmed. A few small rabbit populations still exist, but most have been wiped out by disease.
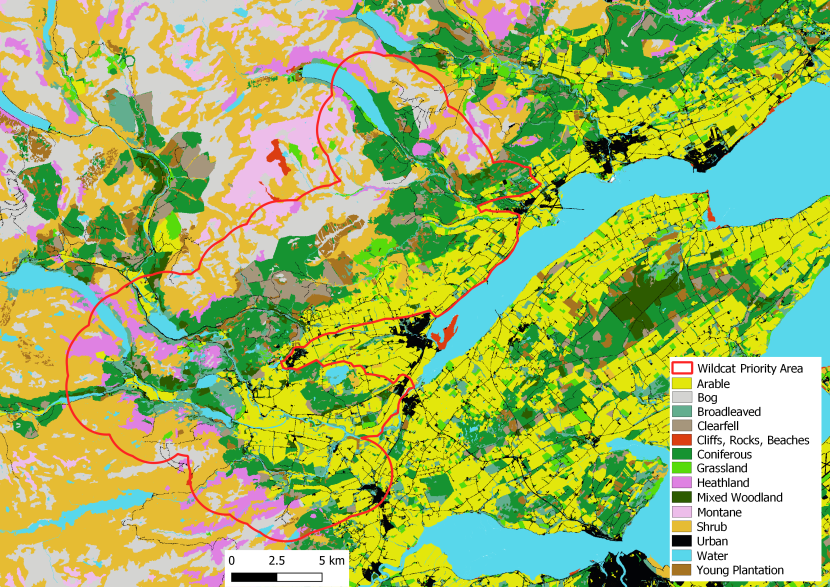
Figure 4. Landcover for the Strathpeffer Priority Area.
Click for a full description
Landcover reclassed from the EUNIS dataset (obtained under Open Government Licence) according to Table 1, cliffs and scree added but not included in Table 1. This is a visual representation of the habitat described for Strathpeffer Priority Area above.
MV = Morvern
Morvern Priority Area (56°37’N, 5°46’W) has also historically been good for wildcats, although the rabbit population disappeared in the 1980s due to disease. A large part of the Priority Area is open hillside (up to ~ 750m), covered in shrub heathland. The south western edge of the Priority Area borders the sea, along which is dense patches of broadleaved woodland, shrub and gorse with scattered rural dwellings. Areas of low intensity grassland are found towards the village of Lochaline and a large patch of coniferous woodland managed by Forestry and Land Scotland is found on the SW as well. Morvern has low human population and low road densities.
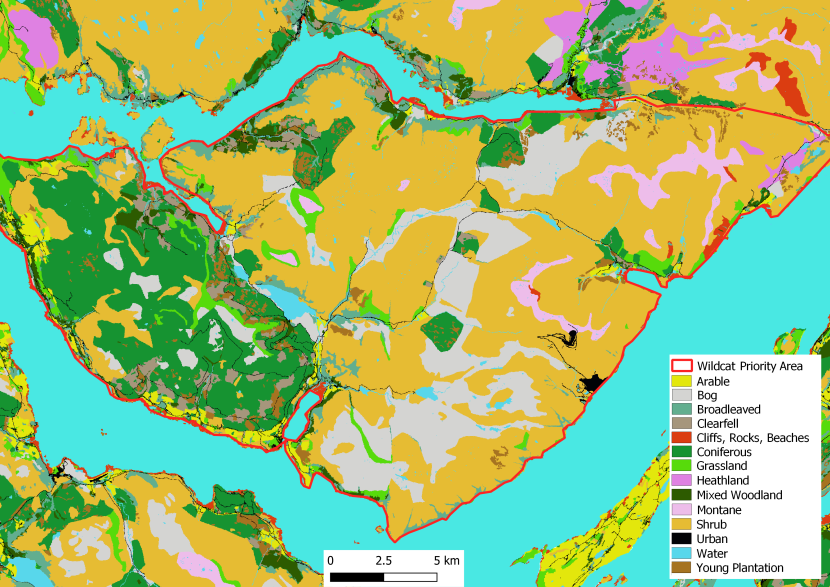
Figure 5. Landcover for the Morvern Priority Area.
Click for a full description
Landcover reclassed from the EUNIS dataset (obtained under Open Government Licence) according to Table 1, cliffs and scree added but not included in Table 1. This is a visual representation of the habitat described for Morvern Priority Area above.
In collaboration with the SWA field team and local Forestry and Land Scotland staff, we aimed to collar wildcats within the priority areas so that the data collected would be directly relevant for management being carried out on the ground as part of the SWCAP actions.
Wildcats to be targeted for trapping and collaring were initially identified from the camera trap surveys being carried out by volunteers and SWA staff across 5 of the 6 priority areas (Strathbogie, Strathspey, Angus Glens, Morvern and Strathpeffer) in the winters of 2017/2018, 2018/2019 and 2019/2020. Individuals were given a pelage score following the criteria developed by Kitchener et al. (2005). In line with SWA guidelines, cats that scored 17 or more were classified as wildcats unless genetic data could verify otherwise (Campbell et al., 2023a, Campbell et al., 2023b).
3.1 Trapping and collaring
Once potential wildcats had been identified based on their pelage score, traps were set and pre-baited at / near locations the individuals had been detected regularly using camera traps. Exact details on the trapping protocol for wildcats can be found in Campbell et al. (2021aa,b), but we provide a brief summary here. Traps were pre-baited for at least 5-7 days using pheasant/quail bait and valerian root as a lure. Traps were carefully covered with plastic sheeting and camouflaged using surrounding vegetation to a) ensure trapped cats did not get wet/cold, b) reduce the risk of tampering. Whilst pre-baiting, traps were locked open using a padlock and/or the back door was removed so that traps could not be accidentally set. This was important to ensure no animals were trapped whilst the traps were not being regularly monitored. Camera traps were used to determine whether or not the target individual was entering the trap and being attracted to the bait. Provided this was occurring, and ensuring veterinary support was on hand and the weather was fair, then traps were set and baited for five consecutive days and nights at a time over the winter trapping period. Following guidance, traps were checked every eight hours unless wireless remote trap monitoring units could be used (e.g. Mink Police Units; www.minkpolice.com), in which case traps could be checked within ~ 20 mins of being triggered. Mink Police Units are placed on the open door of the trap and notify you when the trap door has shut, however, they are dependent on mobile phone coverage, so were only used where mobile reception was consistently good (predominantly Morvern). Trapping was carried out under NatureScot licences 129331, 154280 and 181833.
Although the project commenced in January 2018, trapping did not start until mid-February 2018 because of poor weather conditions (heavy snowfall and temperatures regularly below 0C). Trapping was carried out between Feb-mid March 2018; Nov 2018-mid March 2019 and December 2019-mid March 2020 to deploy GPS collars. If a cat was trapped then an initial assessment was briefly carried out to determine whether it was a wildcat to be collared or a hybrid/feral to be neutered; if it had already been neutered as identified by a tipped ear, it was released. Feral cats and tabby cats with a pelage score <17 were neutered by a vet at a local veterinary surgery or in the case of Morvern, in a temporary neutering clinic, and kept overnight to recover before being released at the site of capture (see Campbell et al., 2021b for the exact methodology). Any cats captured that had not been previously identified from camera trap photographs were also pelage scored and sampled (condition, weight, disease and genetics).
Cats considered to be potential wildcats were checked over and blood samples were taken for genetic analysis and health screening. Provided they weighed 2.3kg or more (collars weighed 69g and should be ≤3% of the animal’s total body weight to avoid impacting their behaviour and well-being; Kenward, 2001), and were in good health, they were fitted with GPS collars (e-obs, GmBH). The collars have both GPS and tri-axial accelerometer sensors (ACC) to not only obtain locational GPS fixes but also to collect data on cat behaviour. The batteries are designed to last up to 24 months depending on how the collars are programmed. These collars were specifically chosen because they have successfully been used on the European wildcat in Switzerland, Germany and Greece (e.g. Jerosch et al., 2017; L. Maronde pers comm., M. Gotz pers comm., Migli et al., 2021). The collars also contain a weak link composed of a segment of leather joining the two ends of the collar so that in the event that any of the cats are not successfully re-trapped for collar removal, they will eventually break off. One major benefit of these collars is that data is downloaded remotely in the field, allowing a more frequent, real-time assessment of cat movement and making management decisions easier, instead of having to wait for the collars to drop off before the data can be collected.
Once each wildcat had been collared, relevant samples and measurements had been taken, it had recovered from anaesthesia and was deemed ready and fit for release by the vet, it was then released at site of capture. Non-target species (e.g. badger [Meles meles], pine marten [Martes martes]) were released at the site of capture after a brief visual check of their condition. The chances of trapping non–target species was greatly reduced by ensuring traps were not set near sites that had previously photographed non-target species.
3.2 GPS radio tracking
Each e-obs GPS collar has a unique UHF transponder frequency which was used to locate individuals and download the data. Limitations of the collar were that in order to maximise their battery life, UHF transmission of the signal was restricted to 2-3 hours a day, making locating the cats quite difficult initially. In the first two weeks after release, collared individuals were relocated once a week if possible to ensure collars were functioning correctly and individuals were behaving as normal (based on previous radio tracking studies e.g. Corbett, 1979, Scott et al., 1993, Campbell, 2015). In the event of a problem, for example if the individual had not really moved after 24-36 hours, then they would have been recaptured and the collar removed. However, this was not necessary.
Individuals were initially located by going to the point of release and searching in increasing circles around this area, ideally from a nearby high point because the transmitter signal was greatly influenced by line of sight. In some cases, it took several days to initially locate a cat. Cats were then relocated using receivers every 3-4 weeks until they were able to be re-trapped for collar removal. Relocating the cat was generally easier once the ranging pattern of each individual had been identified from the data. Once located, data from the collars was remotely downloaded onto a hand-held base station if the individual was within range (50-300m). The base station was used to download and store the data from collared individuals during fieldwork, data was remotely extracted from the collar using the antennae via the UHF radio waves and stored on an SD card inside the base station for later extraction on a computer. By regularly downloading the data from the collars, not only did this prevent data loss if the collar fell off and was not able to be retrieved or the cat could not be relocated at a later date, but also enabled the field staff to examine the data on a regular basis and act on any potential issues. Data download could take up to 40-60 minutes and was affected by poor weather conditions such as rain, blizzards or fog.
3.3 Data analysis
We utilised the ACC-informed GPS function on the collars to maximise battery life. This allowed the activity of the cat to be linked to the frequency of GPS fixes. In this case, when the cat was active, GPS fixes were taken every 15 minutes and when inactive (e.g. resting) every four hours. In addition to GPS data, the ACC data collected by the collars was used to help identify rest locations. The ACC measures the animal’s acceleration along three dimensions (X, Y and Z) to reflect both static acceleration (changes in body posture due to gravity) and dynamic acceleration (changes in speed due to animal movements). The ACC recorded data in four-second bursts every two minutes (10Hz).
Data was uploaded into a GPS depository called Movebank (www.movebank.org), a freely available platform which allows data holders to share their GPS data with relevant stakeholders. This allowed SWA and Forestry and Land Scotland (FLS) staff to access it to help with on the ground management decisions.
Data was analysed in R version 4.0.0 (The R Foundation for Statistical Computing, 2019) and QGIS version 3.4.2 (QGIS.org, 2020, QGIS Geographic Information System, Open Source Geospatial Foundation Project. http://qgis.org). Details on the exact R packages used for each analysis are described in the relevant sections below. Note that this is a preliminary analysis of the data and further analysis will be carried out when the remaining data has been collected from currently collared individuals.
3.3.1 Home range size
Home range sizes were calculated using the package adehabitatHR (Calenge, 2011) in R. To allow comparison with other radio tracking studies carried out in Scotland and other parts of Europe, the Minimum Convex Polygon (MCP) estimate was used (Burt, 1943). This is one of the most widely used estimation methods for home range size and involves calculation of the smallest convex polygon enclosing all the relocations of the animal to estimate the 100% MCP, and is simple to compute, but has been observed to be sensitive to outliers and sample sizes (Downs and Horner, 2008) and may over-estimate home range size (Burgman and Fox, 2003) but not necessarily significantly so compared to other methods (Adam et al., 2015). The 95% and 100% MCPs are provided here for comparison with other studies. Data from Brodie was not included in this part of the analysis because only one month of data was collected before his collar fell off and total home range sizes appeared to stabilise after ~three months. Data from Finley was also excluded in some of the home range analysis as only 15 days’ worth of data has been recovered so far.
To aid visualisation of the data, heatmaps were generated using the interpolation function in QGIS. This creates a density raster map (heatmap) from the GPS points using kernel density estimation. Density is based on the number of points at a location and allows easy identification of hotspots and clustering of GPS fixes.
3.3.2 Seasonal and annual home range size and use
Monthly home range sizes were also calculated using adehabitatHR; only months with 10 or more days of GPS data were included in this analysis. We examined seasonality in home range size (monthly 95% MCPs) using a mixed effects model in the R package lmer (Bates et al., 2015) using a random intercept for cat ID. The global model included sex, a cubic term for month (e.g. Jan = 1, Dec = 12) and a month*sex interaction. We compared this model with those containing quadratic and linear terms for month, with and without the month*sex interaction, just month, just sex and a null model (see Annex 1, Table A1). We used AICc to compare models in the R package MuMIn (Barton, 2019), selecting the model with the lowest AICc value. Brodie and Finley’s data were included here.
Seasonal shift in home range size and location was examined visually in QGIS to determine whether collared individuals moved to use different areas at different times of year. Two individual males in the Strathbogie Priority Area (Bogie and Clash) and one female (Doll) in the Angus Glens Priority Area were collared for > one year so differences in annual home range size (95% MCPs) was also compared.
3.3.3 Rest sites identification
We used the ACC data and GPS locations to identify rest sites. Following Migli et al., (2021) we extracted the overall dynamic body acceleration (ODBA) value for each two-minute interval of the ACC data using the Movebank Acceleration Viewer (Wikelski et al.). The ODBA value is the sum of the dynamic body acceleration from the ACC three axis (X, Y and Z), and can be used as a proxy for animal energy expenditure (Wilson et al., 2006). To identify locations of rest sites we first calculated the mean ODBA for 15 minutes either side of each GPS fix. A rest site was then assumed if the following criteria were met:
- mean ODBA + 2xSD ODBA was <2000
- maximum two min ODBA value recorded in the 30 min period was <2000
Based on Migli et al., (2021) observations and our own from collared individuals before they were released from the trap and therefore “resting”, this ODBA value of <2000 appears to be robust. For example, in the 10 minutes prior to his release Bogie’s mean ODBA value was 835 and leapt up to 30,943 when he was released and sprinted out of the trap.
Using QGIS, the rest site data were double checked against known rest sites discovered during fieldwork to assess the methodology. The number of sites was calculated by using the point cluster function in QGIS so that points within a 15m radius of each other were clumped together and considered to be one location. Then the number of clusters and individual locations was counted. The EUNIS landcover data for each of these fixes was also extracted using the point sampling tool plugin in QGIS. Because the EUNIS data is at a resolution of 10m x 10m, it does not always identify certain features that are often used by the cats such as patches of gorse within an open field or small patches of woodland or hedgerows (see Figure 6) so the habitat data extracted for the rest site locations were double checked using aerial photography and ground-truthing.
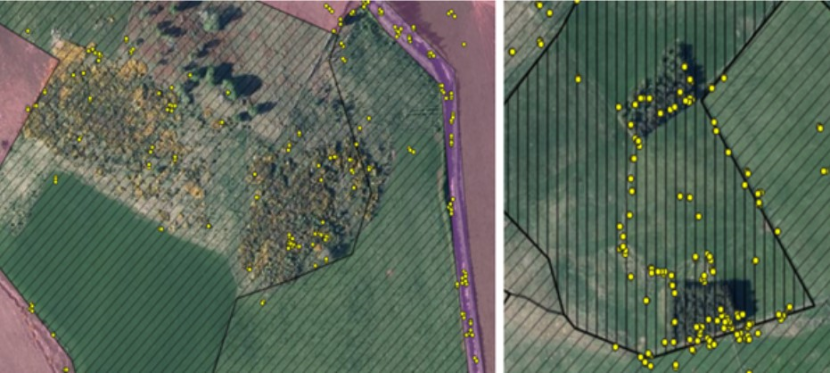
Figure 6. Examples of some of the habitat used by collared individuals that is sometimes missed using 10m x 10m landcover sources such as the EUNIS Landcover for modelling and analysis purposes.
Click for a full description
On the left, the areas with diagonal stripes are all classified as grassland (Grasslands and lands dominated by forbs, mosses or lichens). The yellow points show the GPS fixes for one of the male wildcat hybrids which are largely concentrated on a patch of gorse (shrub). On the right-hand map, the area with the vertical lines is classified as “heath, shrub and tundra” even though it is actually grassland and EUNIS fails to identify the two small patches of coniferous woodland here as well. This is a visual representation of some of the text mentioned in 4.3.3.
3.3.4 Rest site habitat preference
Jacobs Index D (Jacobs, 1974) was used to evaluate the degree of preference for each habitat type as a rest site.
Jacobs index D = (r - p)/(r + p – 2rp)
where r = represents the proportion of use of a habitat as a rest site and p = the proportional availability of that habitat within the wildcat hybrid’s home range. The selectivity index varies from -1 (indicating a lower use of that habitat as a rest site in relation to the availability of the habitat) through 0 (indicating random use or a proportional use of that habitat for resting in relation to its availability) to 1 (indicating cats are showing a strong preference for resting in this habitat in relation to its availability).
3.3.5 Natal den identification
Natal dens were identified from the two females who were captured on camera trap looking heavily pregnant at the end of April. The GPS data and activity data (ACC) were examined to identify likely times of birth. For example, for Emma, her average daily distance moved was 3787m ±2226m /24 hr period (classified as 12:00hrs – 12:00hrs) but on 5th May 2019, she moved only 176m in total suggesting this was the date of birth. This was followed by a change in activity patterns. Clusters of GPS fixes to which the females returned on a daily basis for several hours at a time and for several weeks were initially identified using the programme DYNAMOvis (Dodge et al., 2018). These locations were considered natal dens for the 10-12 weeks post-birth when the kittens were young. Natal dens were further validated when several consecutive field visits to download the data placed the females in the same areas when this was not their normal behaviour in the pre-birth period. Note we did not attempt to locate the exact den site in the field to reduce the risk of disturbing a female with kittens.
3.3.6 Resource Selection Function model (RSF)
The mean horizontal accuracy of the GPS fixes for all cats was 11.13m (SD ± 11.99, range = 1-98m). Before the data was analysed for the Resource Selection Function models, it was first examined to determine what value of horizontal accuracy could be retained to minimise data loss but maximise accuracy in identifying which habitats wildcat hybrids were using. Following this initial assessment, fixes with an accuracy >60m were removed from the dataset, as were extreme and/or unusual outliers (for example, Fairburn had 3 GPS fixes on the same day over a 1-hour period that were 150km away from all her other fixes, which were extreme and did not fit with the rest of the data, so were removed). This retained 99.1% of the total GPS fixes with a mean accuracy of 9.8m (SD ±1.62m). When cats were active the collars took three consecutive fixes every 15 minutes when a GPS location was taken, when not active only one GPS fix was obtained every 4 hours. Because these consecutive fixes were generally close together spatially (<10m), we only retained the third fix (normally the highest accuracy (m)) to avoid pseudo-replication in the RSF modelling.
We wanted to examine how wildcat hybrids were using different habitats within their home ranges (Third order selection) according to the hierarchical structure of resource selection detailed by Johnson (1980). Initial analysis of the data indicated that habitat selection varied significantly by individual cat and Priority Area for all predictors. We therefore developed individual resource selection function (RSF) models for each collared wildcat hybrid using binomial general linear models (GLMs). Following (Klar et al., 2008) random points equal to the number of GPS fixes for each wildcat hybrid were generated within that cat’s 100% MCP. These two layers of points (GPS and Random) were then merged in QGIS to produce a single vector layer per cat with both “GPS” and “Random” points in it. Using the merged point data our Response variable was binary, with GPS fixes (Use) coded 1 and random points (Available) coded 0, leading to a logistic model of habitat use.
Predictor variables
Predictor variables were chosen based on previous studies and our own field experience, which showed that distance to roads, buildings, habitat edges and habitat type are some important predictors of habitat use for wildcats (e.g. Campbell, 2015, Klar et al., 2008).
The primary source of landcover data was the EUNIS (European Nature Information System) Landcover Scotland data obtained from NatureScot under an Open Government Licence (EUNIS, 2020). This is a 10m raster based Landcover map amalgamated from 13 different sources and classified according to EUNIS. Landcover categories selected for use in the model are shown in Table 1, summaries of the main predictors for each Priority Area are also shown in Table 2. Separate 1 x 1m raster files were created for each landcover type used in the analysis in QGIS. In addition to the landcover data from EUNIS, other predictor variables included in the RSF models were:
- Distance (m) to nearest change in cover type (edge) (dEdge); to create an edge map, the EUNIS Landcover categories used for the RSF models were reclassified into three cover types; “open”, “mixed” and “closed” (see Table 1) and converted into separate 1 x 1m raster files.
- Distance (m) to low use roads (dRoadL) (minor roads, forest tracks, some quiet B roads) and distance (m) to medium to high use roads (dRoadH) (A roads, motorways and any roads that had a name representing a residential street likely to be used on a regular basis, B roads); data was extracted from the OS Open Roads database (licensed under an Open Government Licence, contains OS data © Crown copyright and database right 2021) and converted to 1 x 1m raster file.
- Distance (m) to the nearest building (dBuilding); a Settlements shapefile for Scotland was obtained from the Office for National Statistics (licensed under the Open Government Licence v.3.0. Contains OS data © Crown copyright and database right [2020]) and converted to a 1 x 1m raster file.
The raster proximity tool was used to produce raster proximity maps from the relevant 1 x 1m raster files for; 1) each of the landcover types used in the analysis, 2) cover type (closed, mixed or open) to estimate distance to nearest edge (change in cover type), 3) roads, 4) buildings. The point sampling tool plugin in QGIS was used to extract the distance from each GPS fix/Random point to each of the predictor variables using the raster proximity maps.
| Cover type for dEdge in RSF models | Landcover categories for RSF models | Habitat classification (EUNIS) |
|---|---|---|
| Closed | Coniferous woodland | G3: Coniferous woodland G3.4 Pinus sylvestris woodland south of the taiga G3.F Highly artificial coniferous plantations |
| Closed | Broadleaved Woodland | G1: Broadleaved deciduous woodland G1.8 Acidophilous Quercus-dominated woodland G1.9 Non-riverine woodland with Betula Populus tremula or Sorbus aucuparia |
| Closed | Mixed Woodland | G4: Mixed deciduous and coniferous woodland G Woodland forest and other wooded land G4 Mixed deciduous and coniferous woodland G6 Exotic woodland and scrub |
| Mixed | Clear-fell (CF) | G5.8 Recently felled areas |
| Mixed | Young plantation | G5.7 Coppice and early-stage plantations G5.6 Early-stage natural and semi-natural woodlands and regrowth |
| Mixed | Shrub | E5 Woodland fringes and clearings and tall forb stands F4 Temperate shrub heathland F3.1 Temperate thickets and scrub F9.2 Salix carr and fen scrub |
| Mixed | Heathland | F2 Arctic alpine subalpine and extensive scrub F4.1 Wet heaths F4.2 Dry heaths |
| Open | Arable | I1 Arable land and market gardens E2.6 Agriculturally-improved re-seeded and heavily fertilised grassland including sports fields and grass lawns |
| Open | Bog | D1: Raised and Blanket bog |
| Open | Grassland | E Grasslands and lands dominated by forbs mosses or lichens E4 Alpine subalpine and extensive grasslands E1.2 Perennial calcareous grassland and basic steppes E4.3 Acid alpine subalpine and extensive grassland E4.4 Calcareous alpine subalpine and extensive grassland |
Further details on the EUNIS categories can be found online.
| Landscape features | Angus Glens | Morvern | Strathbogie | Strathpeffer |
|---|---|---|---|---|
| Arable | 0.1 | 0.02 | 0.4 | 0.2 |
| Bog | 0.07 | 0.1 | 0.02 | 0.1 |
| Coniferous woodland | 0.1 | 0.2 | 0.2 | 0.2 |
| Broadleaved woodland | 0.02 | 0.04 | 0.02 | 0.06 |
| Mixed woodland | 0.004 | 0.02 | 0.01 | 0.02 |
| Clear-fell | 0.03 | 0.02 | 0.04 | 0.04 |
| Young plantation | 0.01 | 0.03 | 0.05 | 0.03 |
| Grassland | 0.1 | 0.03 | 0.1 | 0.02 |
| Montane | 0.001 | 0.02 | 0 | 0.01 |
| Heathland | 0.4 | 0.004 | 0.04 | 0.06 |
| Shrub | 0.1 | 0.5 | 0.07 | 0.2 |
| Road/track length - low use (km) | 214 | 224 | 392 | 342 |
| Road length medium - high use (km) | 44.35 | 66.2 | 80.33 | 62.7 |
| Urban (m2) | 0.0005 | 0.0002 | 0.002 | 0.002 |
| Edge length (km) | 7135 | 7871 | 9647 | 9323 |
| Total area (km2) | 375 | 486 | 347 | 336 |
RSF Models
We used the RSF models to examine habitat selection and predict the relative probability of an individual using different habitats within its home range based on the distance (d) to the edge of the predictor variables from each GPS fix / random point (locations falling within the habitat type were given a value of d = 0m) using the following model;
- Global RSF model 3rd Order Selection: Use (0,1) = dArable + dBroadleaved woodland + dClear-fell + dConiferous woodland + dGrassland + dHeathland + dMixed woodland + dShrub + dYoung plantation + dEdge + dRoadL + dRoadH + dBuilding
The predictor variables were normalised to zero mean and one standard deviation using the Standardize function in Excel prior to being fitted in the model to reduce convergence issues. Prior to running the RSF models, correlations between predictor variables were assessed on the raw data using the cor function and the corrplot (Wei and Simko, 2017) and PerformanceAnalytic (Peterson et al., 2020) packages in R. If two or more predictor variables were highly correlated (r>0.8) and/or had a VIF >10 (Variable Inflation Factor; detects multicollinearity in regression analysis) then the predictor that made the most sense biologically was retained for that individual cat’s model.
The RSF models were run in R using the glm function in the lme4 (Bates et al., 2015) and lmerTest packages (Kuznetsova et al., 2017). Beta coefficients (i.e. selection coefficients) were considered to be presentative of relative habitat selection. Predicted values (with 95% confidence intervals calculated from the model output using the ggpredict function in the package ggeffects in R) (Lüdecke, 2018) and plotted out.
To identify any patterns in habitat use overall and across Priority Areas, we also calculated averaged population-level selection coefficients from the individual RSFs. Here significant individual beta coefficient estimates were averaged using an inverse variance weighting (IVW) method. This method accounts for the variation in coefficients that may result from differences in sample sizes within a specific habitat type and gives a higher weight to individuals with larger samples sizes and lower standard errors (i.e. Takahata et al., 2014, Thomas et al., 2021).
4. Results
Camera trap survey work carried out by the SWA field team identified several putative wildcats based on pelage. Overall, 14 individuals (4 females and 10 males) were fitted with GPS collars, across four of the five priority areas; Morvern, Strathpeffer, Strathbogie and Angus Glens. Mean pelage score was 18 (range = 17-21). See information on cats collared including SWA ID (for reference to other reports), name, sex and location (PA = Priority Area) cat was collared, date first collared, battery life (and total number of GPS fixes), pelage score (adapted from Kitchener et al., 2015, cats scoring 17 or more out of a possible 21 for 7 key pelage characteristics were classified as wildcat, Q = hybrid score (i.e. how hybridised an individual is), lower (LBQ) and upper (UBQ) range of Q are also shown, cats scoring 0.75 for LBQ are classified as wildcat, mitochondrial DNA (mtDNA; Wc = wildcat, Dc = domestic cat) and the final status of the collared individual is also shown. (Two males, Fergyl and Brodie, had their pelage scores later downgraded when recaptured for collar removal to <17).
Genetic analysis (Senn et al., 2019) indicated that all of the collared individuals had a hybrid score Q<0.75 (Q = % wildcat genes a cat has), failing to meet the genetic threshold of a wildcat (Q ≥ 0.75), indicating they were wildcat hybrids (Table 3).
Collars have lasted to date an average 281 days (range = 113 – 333). This does not include data from two young males, Brodie and Finley; see Table 3. Brodie’s collar fell off at some point, and only a month’s worth of data was collected from him so this was excluded from some of the analyses. Finley was collared at the end of February 2020, only 15 days of data were collected before COVID enforced lockdown in March 2020 prevented data collection. He has not yet been relocated, so his data is excluded from some of the analyses. The number of GPS fixes obtained per cat varied according to battery life, individual collar differences and in relation to the overall activity of the cat (Table 3). Two collars fell off before the batteries had expired and were re-deployed on different individuals. Two males caught in Feb/March 2018 (Bogie and Clash) were re-collared in the 2018/2019 trapping season. Clash was re-collared again in the 2019/2020 trapping season and a female cat, Doll, was also recollared in the 2019/2020 trapping season.
During the trapping period 2019/2020 one of the main aims was to trap and remove collars no longer working and re-collar some of the individuals. The winter trapping season was very successful, with all expired collars removed/accounted for. In the winter of 2021, attempts were made to trap and remove the collars of the four remaining collared individuals. Collars were successfully removed from Clash and Doll, but Finley and Woody in the Angus Glens have not yet been relocated following COVID related fieldwork restrictions and efforts continue to find them. This was the first time these collars had been used on wild-living cats in Scotland so it was important to assess the cats when the collars were removed to determine if the collars would be suitable for use in the future. The batteries on the collars are designed to last up to two years depending on the settings and as a result the “weak link” did not appear to have been weakened much after nine months. Consequently, the new batch of collars that were deployed in the winter of 2019/2020 was produced with a weaker link than the previous batch. The performance of these links did not appear to be greatly different to the previous models and this is an area that would ideally be improved for shorter term studies so that individuals are not wearing collars that are no longer collecting data unnecessarily. Two males (Brodie and Fergyl), who only just passed the pelage threshold when trapped and collared, were re-caught and neutered as both had very low genetic scores (Q = <0.3) and subsequent pelage assessment resulted in a downgrading of their pelage scores from 17 to 15.5 and 14.5 respectively (Table 3).
Although identified as wildcat hybrids genetically, the GPS data from this and previous radio tracking studies (Campbell, 2015, Daniels, 1997, Scott et al., 1993, Corbett, 1979) indicates that they were behaving in a similar manner to European wildcats in mainland Europe. Therefore, the data still offer a valuable insight into how wild-living cats are using the different landscapes across the Priority Areas and the implications this may have for wildcat conservation in Scotland.
Table 3. Information on cats collared including SWA ID (for reference to other reports), name, sex and location (PA = Priority Area) cat was collared, date first collared, battery life (and total number of GPS fixes), pelage score (adapted from Kitchener et al., 2015, cats scoring 17 or more out of a possible 21 for 7 key pelage characteristics were classified as wildcat), Q = hybrid score (i.e. how hybridised an individual is), lower (LBQ) and upper (UBQ) range of Q are also shown, cats scoring 0.75 for LBQ are classified as wildcat),mitochondrial DNA (mtDNA; Wc = wildcat, Dc = domestic cat ) and the final status of the collared individual is also shown.
4.1 Behaviour of cats immediately post release
Using direct observation and the GPS/Accelerometer data we were able to get a good indication of how collared individuals behaved immediately post release after collaring. Both direct observations and video footage of released cats show them sprinting out of the traps upon release. The accelerometer (ACC) and GPS data was then examined to determine how far and where they went post release.
- Bogie
2018 - Initially trapped within the edge of grassland alongside the river Bogie, when released he ran for several minutes and his next GPS location 15 minutes later was 2.2km away. He then appeared to go underground for several hours because GPS signal was lost, the GPS signal before and after this period indicates he was on top of a rabbit warren (from a later field visit), so we assume he went down into one of these for this period while no GPS signal was getting picked up. We know he was running for the first few minutes because we watched him sprint out of the trap along a field edge and leap over a small river. Looking at his accelerometer data it shows high speed movement with a sharp peak when he leapt over the river before acceleration speed dropped slightly (presumably still running/trotting) before slowing right down at ~11:10am (Figure 7). GPS signal was picked up again when he must have moved slightly and then he rested another 3.5hrs. He stayed within the same area until 23:30 then started to move. The location he ran to turned out to be a nearby haybarn, with plenty of rabbits and cover in the form of shrub and hedgerows, which he continued to use on a regular basis as a rest site over the next two years.
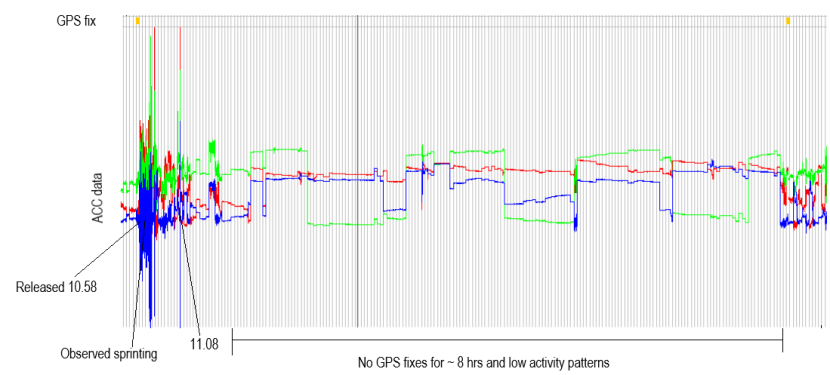
Figure 7. Screenshot of the accelerometer data at Bogie’s release.
Click for a full description
He was released from the trap at 10:58 and was seen sprinting across a large field. From the pattern of the accelerometer data we can conclude that he ran for 10 minutes before slowing down. The triaxial accelerometer (ACC) axes are shown in green (X), red (Y) and blue (Z). The ACC measures acceleration along three dimensions (X, Y and Z) to reflect both static acceleration (changes in body posture due to gravity) and dynamic acceleration (changes in velocity due to animal movements). Here we can see how the ACC data looks when Bogie is sprinting versus when he is not moving much. Based on the fact that there were no GPS fixes taken for nearly 8hrs we concluded that he must have gone underground during this period where no signal could be picked up. This is a visual representation of some of the text mentioned above.
2019 - When he was recollared he was trapped in a small patch of woodland near a hay barn that he used on a very regular basis. After release, he ran 100m to the hay barn where he stayed for 3 hours until 15:15. He then moved 700m to a large hillside of gorse where he stayed overnight for 11 hours before normal activity resumed. He continued to return to the hay barn/woodland where he was caught on a regular basis, suggesting trapping him had not influenced his use of this area significantly.
- Brodie – Initially trapped within a small barn, when released he ran ~2.5km to a burrow within some gorse at the edge of a field where he stayed for 10 hours until evening before moving out.
- Brook – Trapped, he ran 100m to a nearby croft where he stayed for 9 hrs before starting to move around.
- Clash
2018 – Initially trapped on the edge of coniferous woodland near a forestry track, when released he ran ~500m then rested within coniferous plantation for 2hrs before moving a further 400m to the edge of a field where he stayed for an hour within scrub. He then moved another 400m to a small coniferous plantation where he stayed for 7hrs.
2019 - Trapped along a fence line within a patch of gorse on farmland, when released he ran 400m to an area of gorse where he spent 6 hours resting. He then moved around for an hour or so before GPS signal disappeared for ~ 20 hrs between 20:00 on the day of release until 16:00 hrs the following day in an area he was regularly detected within a large rabbit warren. It’s possible that he was underground in a burrow during this period. He then resumed normal activity.
2020 – Initially trapped along a stone wall in open grassland, he went straight to his regular hay barn where he stayed for 7 hours during the daytime before heading out at dusk as per normal behaviour.
- Doll
2019 – Initially trapped along a fence line bordering woodland and open heathland, when released she ran ~ 700m along the fence line uphill, then rested within coniferous plantation for 4hrs. She then moved 250m then stayed there for a further 4 hrs before moving again ~150m where she stayed for 4 hours before starting to move around.
2020 - Trapped at the same location as 2019, ran along the same fence line before heading into coniferous woodland where she stayed for 15 hours before moving around again.
- Emma – Trapped within coniferous woodland bordering a small forestry track, when released she ran 800m then rested within coniferous plantation for 12 hrs.
- Fairburn – Ran ~ 2km, then stopped for 6.5 hrs within broadleaved woodland on the edge of farmland before moving around again.
- Fergyl – Trapped just outside a small barn, when released he ran ~ 2km to an area of gorse then stayed there for 22hrs before moving.
- Finley – Trapped within a coniferous plantation near the edge of open grassland. When released he ran towards the grassland then uphill along a fence line for ~500m before staying in a small patch of birch / scrub for 19 hours before moving.
- Lochie – Initially trapped within a small coniferous plantation on a steep hill, when released he ran ~ 1.5km to the neighbouring small patch of coniferous plantation (possibly within a rabbit burrow) opposite a farm and stayed there for almost 24 hrs before moving.
- Markie – Trapped on the edge of coniferous woodland next to a forestry track, when released, he ran 800m up a steep hill to the edge of a coniferous plantation where he stayed for 16hrs. He then moved 100m to the edge of the plantation in some heathland/scrub where he stayed for a further 8 hrs.
- Sprat – Trapped in a rural garden, when released she ran 10m away to the edge of a nearby field where she stayed within some broadleaved scrub for 16 hrs. This area later proved to be one of her main rest sites.
- Stonker – Moved over 2km then rested in broadleaved woodland on the edge of a field for 12.5 hrs before moving as normal.
- Woody – Trapped along the side of a stone wall in larch woodland. When released he ran uphill to an area of wind blow where he stayed for 1.5 hrs before moving along the edge of clear-fell to another patch of coniferous woodland where he stayed for 6 hours before moving around for the night.
In summary, all the cats that were collared appeared to be behaving normally by 24 hrs post collaring and release. In addition, during the winter of 2019/2020, all the cats that were caught and assessed to have their collars removed turned up on camera trap within ~48 hours, suggesting no observable adverse effects of being re-trapped and re-anesthetised for collar removal.
4.2 Body condition of cats collared
When initially collared, cats weighed a mean of 4.6kg, with females on average weighing less than males (3.6kg vs 4.9kg). Post collaring mean weight of the cats was 4.7kg (includes Morvern cats, see below; 2.3-6.4kg) with females weighing an average of 3.3kg and males 5.7kg post collar removal (or a mean of 3.8kg vs 5.9 kg for females and males respectively excluding the Morvern cats). The increase in weight for the male cats can be explained by the fact that three were young adults at the time of collaring and had matured at recapture (see Table 4). For comparison, low scoring hybrids that were neutered had a mean body weight of 3.6 kg (mean of 3.3kg vs 4.3kg for females and males respectively) (Table 5).
With the exception of the two cats collared in Morvern, all of the cats collared appeared to be in good body condition as assessed by the vets involved when their collars were removed (see Table 4). Importantly there were no obvious issues under and around the collar area such as skin irritation or sores indicating that the collars had been properly fit to the cats and had not stretched out of shape as can sometimes occur with some makes of collars.
The two cats collared in Morvern unfortunately lost a significant amount of body weight during the study (see Table 4). Both cats had underlying conditions when collared, although at the time they were not deemed to be causing any issues with overall fitness based on long term camera trap survey work. Stonker had an old eye injury which meant he was blind in one eye but he almost nightly moved ~10km across his home range. Sprat had an abdominal hernia which did not appear to be causing any clinical signs but had enlarged during the course of the study. One of the local volunteers managed a video camera during the project which provided regular observations of the two cats and provided evidence that both suffered from a leg injury during the study. Sprat in particular was seen limping for several weeks, before apparently recovering. The leg injuries would have made it difficult to hunt, and both cats unusually appeared at the volunteer’s back garden during the summer months. Although they did not appear to be obviously underweight on the videos, the behaviour was sufficiently strange, and the evidence of a limp meant that they were fed supplementary cat food whenever they appeared, until they were able to be trapped. Because of the weight loss the cats experienced, the collar on Sprat was slightly loose and had rubbed her skin underneath causing some irritation which was treated. Bloods were taken and assessed for Stonker when he was trapped but there was nothing obvious to explain the weight loss (see Table 4). It may have been a combination of underlying conditions, leg injuries, and the effect of the collar and potentially low prey availability that resulted in both these cats losing weight. The coincidence of two cats in the same Priority Area both suffering weight loss when no other collared cats did, indicates something else might also be contributing to this issue. Unfortunately, no other cats in Morvern were caught during this period for comparison so it’s difficult to determine the exact causes, but some uncollared hybrids in the Strathbogie priority area were also noted to be underweight this winter, which was attributed to parasitic worms (Table 5). Other uncollared hybrids are also shown in Table 5 for comparison.
Table 4. Details on the condition of collared cats at fitting (PreC) and removal (CR) of collars including weight (kg) and body condition score (cs 1-5; 1 = very poor, 5 = excellent). Also shows condition of the three cats (Bogie, Clash and Doll) that were re-collared (ReC). Priority Area = PA, SB = Strathbogie, AG = Angus Glens, SP = Strathpeffer and MO = Morvern. Procedure ID (Proc ID) is the unique number given when the cat is processed by the vet for reference to other SWA reports. Sex = F for female and M for male cats.
Table 5. Details on the condition of non-collared cats in the different Priority Areas (AG = Angus Glens, SP = Strathpeffer, MO = Morvern and SB = Strathbogie) for comparison with collared individuals, including weight (kg) and body condition score (cs 1-5; 1 = very poor, 5 = excellent). Procedure ID (Proc ID) is the unique number given when the cat is processed by the vet for reference to other SWA reports. Sex = F for female and M for male cats. NR = Not Recorded.
4.3 Home range size
Data from the GPS collars was only included in the analysis of home range size if 10 or more days of data existed for that particular month. Two young males were excluded from the overall home range size analysis. Brodie, collared in the Strathbogie Priority Area, was excluded because we lost track of him for several months; he was re-trapped during the winter of 2019/2020 without his collar on, indicating it had fallen off (possibly at a location that impeded the UHF transponder signal) so we only had 1 months’ worth of data for him. Similarly, Finley, in the Angus Glens Priority Area had only 15 days of data downloaded before we lost track of him as a result of COVID related fieldwork restrictions. He has not yet been relocated. They are both, however, included in the monthly home range size analysis.
One young male (Lochie) collared in the Angus Glens had a relatively small home range, before undergoing what appeared to be a dispersal event five months after being collared and was detected some 17km away from where he was first collared and we lost track of him (Figure 8). He turned up on camera traps again where he was initially trapped during the subsequent winter without his collar, indicating it had fallen off. Not all data from cats collared in 2020 are included in the analysis because fieldwork restrictions due to the coronavirus means these data are currently incomplete. These will be analysed at a later date. The results relating to the specific questions asked are detailed below.

Figure 8. Return of Lochie (young male) without his collar on to the area he was originally trapped and collared in. This is a visual representation of some of the detail described in 5.3 for Lochie’s home range.
One male, Bogie (408 days of data) had a superficially large home range size (100% MCP = 83.56 km2) because of an excursion he underwent during 6th April – 8th April 2019 which saw him moving a distance of ~ 35 km over a 72hr period outside of his “normal” home range (Figure 9).
Figure 9. Home range of Bogie, x and y axis show distance in km, clear circles show GPS fixes and lines show the line between consecutive fixes.
Click for a full description
Blue triangle shows location where he was first trapped on 18/2/2018 and red square shows the location of the last GPS fix recorded on the 19/12/2019. This is a visual representation of some of the results discussed in 5.3 above for Bogie’s home range.
4.3.1 Overall home range size
Mean overall home range size (95% MCP) of males was 18.15 km2 (range = 8.32 -29.72 km2, N=8), larger but not significantly so than females, which was 13.94 km2 (range = 4.42-28.59 km2, N= 4) (Welsh two-sample T-test; t = 0.721, df = 4.67, p = 0.505, Figure 10).
However, within males, the younger males (1-2 years old as estimated by veterinary clinicians; Lochie, Fergyl and Brook) had smaller mean 95% home range areas than the older males (Bogie, Clash, Stonker, Markie and Woody) regardless of the priority area they were located in (t = 5.258, df = 5.37, p = 0.003). However, comparing the older males with females did not reveal a significant difference in home ranges (t = 1.591, df = 4.068, p = 0.186). For females, Emma had a larger home range size than the other three female cats collared (Figure 10).
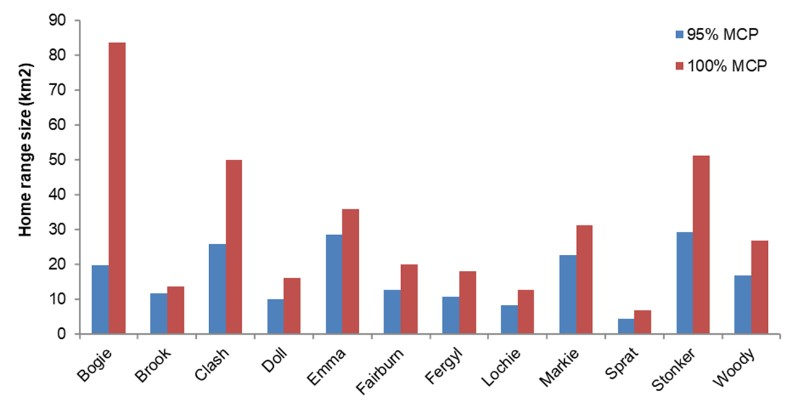
Figure 10. 95% and 100% Minimum Convex Polygon (MCP) estimates of home range size of the 12 individuals included in the overall home range analysis
Click for a full description
(Finley and Brodie not included in this analysis) and tracked for various time periods between Feb 2018 and March 2021. This is a visual graphical representation of the results described in 5.3.1.
4.3.2 Annual home range variation in 95% monthly MCPs
Three individuals were re-collared during the project; two male cats in the Strathbogie Priority Area, Bogie and Clash, were first collared in early 2018 and re-collared in late 2018/2019. Clash was recollared for a third time in 2020 but his dataset has not yet been fully analysed. In the Angus Glens, a female, Doll, was collared in 2019 and re-collared in 2020. This long-term GPS data provides an insight into whether home ranges remained stable over a longer period.
Comparing overall home range size (95% MCP) between consecutive years, we can see that Bogie’s home range size increased slightly between 2018 and 2019 (17.21 km2 vs 20.67 km2) and covers mostly the same area (Figure 11). In comparison Clash’s overall home range size appears to be substantially smaller in 2020 (11.06 km2) than in 2018 (20.89 km2) and 2019 (19.17 km2). Clash also appears to have shifted his home range slightly in 2019 when he seemed to avoid an area that he previously used in 2018, and this appears to correspond to both the location and period over which Emma had her kittens. Emma established a home range in this area and in 2020 Clash appears to have shifted even further away to the south – east (Figure 11 and see Figure 13), although he still retained his most frequently used rest site despite Emma also appearing to use it on several occasions. Doll’s overall home range size in 2020 (4.96 km2) was much smaller than in 2019 (11.12 km2, Figure 12).

Figure 11: Shows the 95% MCP’s for Bogie (2018 and 2019) and Clash (2018, 2019, 2020)
Click for a full description
and the cover type as described in Table 1 in the Strathbogie Priority Area. Bog habitat, not considered to be used extensively by wildcats, are shown in white. This is a visual representation of the results discussed in 5.3.2.
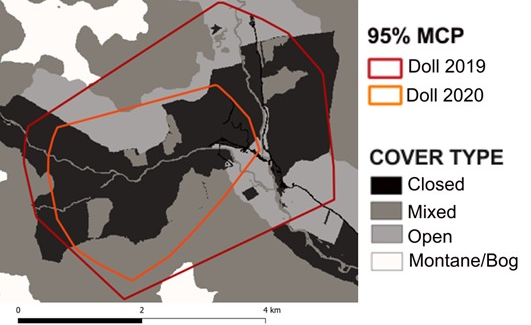
Figure 12. Shows the 95% MCP’s for Doll in 2019 and 2020 and the cover type as described in Table 1 in the Angus Glens Priority Area.
Click for a full description
Bog and montane habitats, not considered to be used extensively by wildcats, are shown in white. This is a visual representation of the results discussed in 5.3.2.
4.3.3 Home range overlap
Individual 95% MCP’s overlapped with each other. In Strathbogie 6 cats were collared, 5 within range of each other; Emma’s home range overlapped with two of the collared males (Clash and Brodie) and a young male (Fergyl) overlapped with Bogie’s home range (Figure 13). Camera trap surveys carried out during the GPS study by the SWA also indicated that several other uncollared wildcat hybrids within the Strathbogie Priority also overlapped with the collared cats’ home ranges. In Morvern, Sprat’s home range was almost completely encompassed by Stonker’s range and three other non-collared hybrid males were also regularly seen on the same camera trap within her home range. Similarly, in the Angus Glens, Doll’s home range in 2020 overlapped with Finley’s (Figure 14) and Woody was using areas known to be occupied by an uncollared female with kittens, another mature male (possibly Markie) and two younger males.
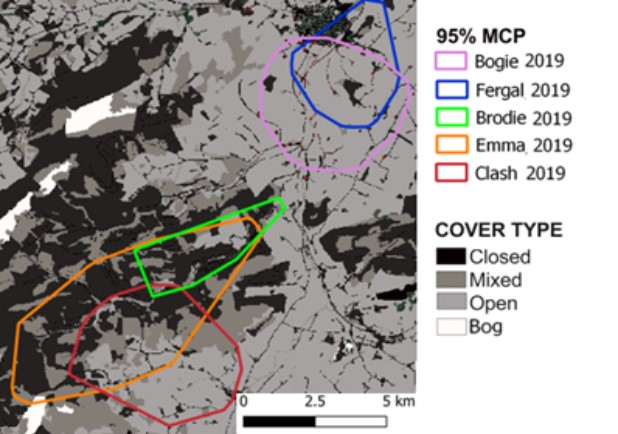
Figure 13. 95% MCP overlap between five of the collared cats in the Strathbogie Priority Area in 2019.
Click for a full description
Note Brodie’s data was only for 1 month, his actual 95% MCP is likely to be larger. Also shows the cover type encompassed by each cat’s 95% MCP. Bog habitat, not considered to be used much by wildcats, is shown in white. This is a visual representation of the results discussed in 5.3.3
4.3.4 Monthly home range use
The most informative model for seasonality in home range contained a linear term for month, sex and a month*sex interaction (M5; Annex 1, Table A2). The next most informative model (M6; Annex 1, Table A2) had an AICc value 0.91 higher and contained only month (linear) and sex but no interaction. However, AICc weighted model averaged estimates using both these models indicated that the month*sex interaction was still significant (estimate 0.060, 95% CIs 0.031 - 0.166) and therefore we provide the results from the most informative model here.
| Parameter | Co-efficient Estimate | Std.Error | t-value | Pr(>|t|) |
|---|---|---|---|---|
| Intercept | 2.38549 | 0.25056 | 9.521 | 5.12e-10 |
| Month | -0.14002 | 0.02533 | -5.528 | 2.58e-07 |
| Sex | 0.19451 | 0.30901 | 0.629 | 0.53 |
| Month * Sex | 0.09834 | 0.03411 | 2.883 | 0.005 |
The difference between male and female 95% MCP home range size varied significantly with month (χ2 (1) = 8.2415, p = 0.004). Home range size was largest for males between February - August (mean monthly home range sizeFeb-August = 12.79 km2) and for females between February - April (mean monthly home range sizeFeb-April = 11.93 km2). Female home range size decreased from the end of April presumably due to decreased ranging behaviour when rearing kittens (mean monthly home range sizeMay - Oct = 3.48 km2) (Figure 15).
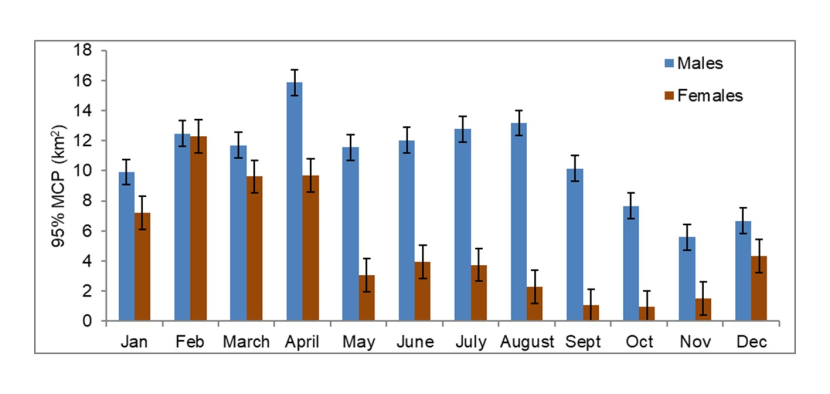
Figure 15. Mean monthly 95% Minimum Convex Polygon (MCP) home range sizes (km2) for males and females with vertical black bars showing standard errors of the mean (s.e).
Click for a full description
This is a visual graphical representation of the results described in 5.3.4..
4.3.5 Seasonal home range
In the Angus Glens, some of the collared wildcat hybrids appeared to shift their home range seasonally, to use the more open dry heath (heather moorland/heathland) in the spring/summer months and forest and other covered habitats in autumn/winter. For example, both Doll and Markie stayed in open heathland up to 2km away from the nearest forest edge during much of May to July (Figure 16 and Figure 17). Markie in particular showed a shift from using forested areas in the winter to the more open heathland from the end of April, where he spent a significant amount of time resting in rocks and cliff faces (Figure 17). However, another male, Woody, who overlapped some of the same area as Markie (but in 2020), did not move up into the hills and instead used more open farmland towards the southern end of his home range in the summer months (Figure 18).
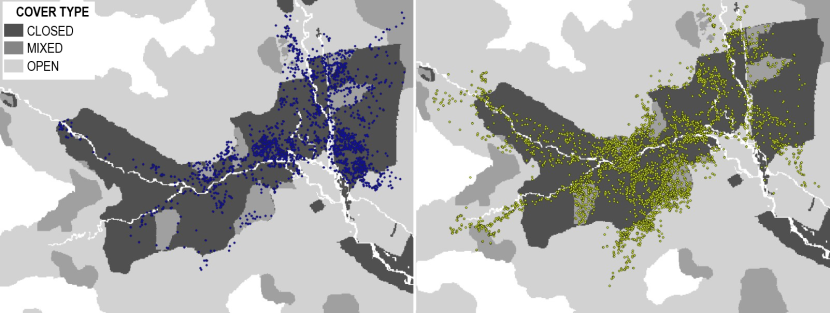
Figure 16. Doll’s home range during autumn/winter 2019 (blue circles) and spring/summer 2019 (yellow circles) showing use of more mixed cover habitat (in this case dry heathland) and open grassland during warmer months where she moved up to 1.5km away
Click for a full description
from the nearest forest cover (data from 10th January 2019 to 8th August 2019).White areas represent bog and white lines show rivers and roads. This is a visual representation of the results discussed in 5.3.5
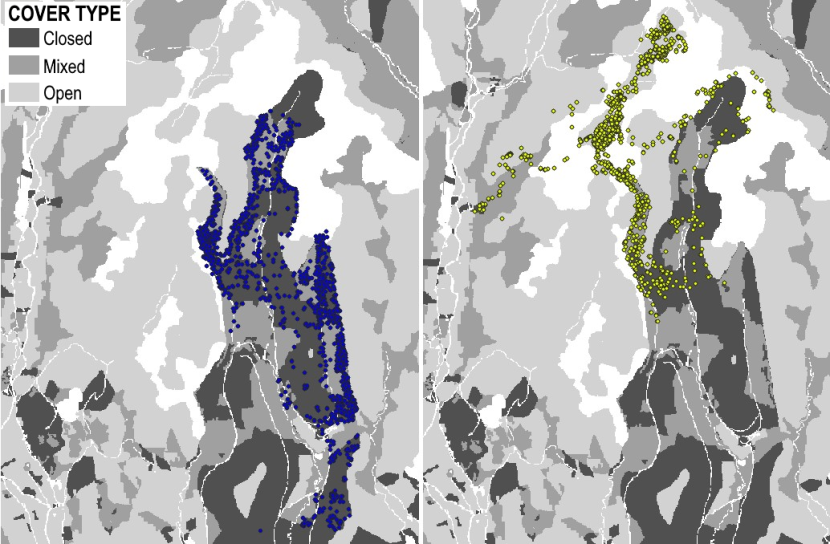
Figure 17. Markie’s home range during autumn/winter 2018 (blue circles) and spring/summer (yellow circles) showing use of more mixed cover habitat (in this case dry heathland) during warmer months where he moved up to 3km away from the nearest forest
Click for a full description
cover (data from 20th February 2018 to 7th July [when his collar fell off]). White areas represent bog and white lines show rivers and roads. This is a visual representation of the results discussed in 5.3.5.

Figure 18. Woody’s home range during autumn/winter 2020 (blue circles) and spring/summer (yellow circles) showing similar use of habitat across these periods (data 9th March to 31st October 2020).
Click for a full description
White areas represent bog and white lines show rivers and roads. This is a visual representation of the results discussed in 5.3.5.
In Strathbogie, the cats appeared to use all parts of their home range across the year (e.g. Figure 20 and Figure 19), except for Emma, who reduced her home range during the denning period (see Figure 27).
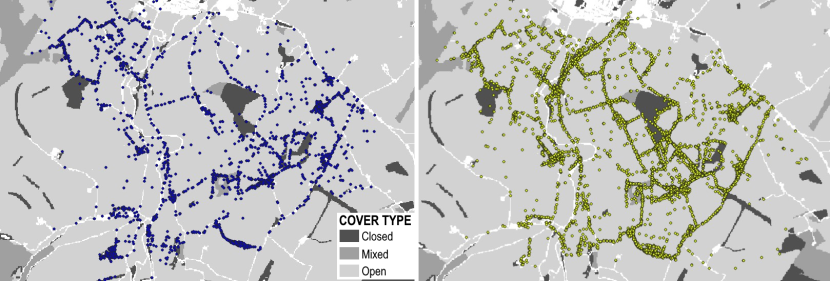
Figure 19. Bogie’s home range during autumn/winter 2018 (blue circles) and spring/summer (yellow circles) showing similar use of habitat across these periods (data 16th February 2018 to 24th November 2018).
Click for a full description
White lines show rivers and roads. This is a visual representation of the results discussed in 5.3.5.
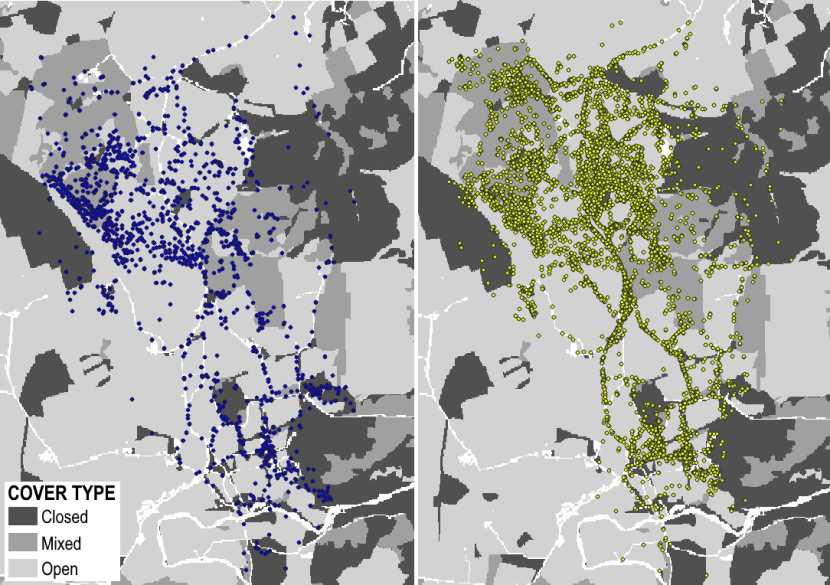
Figure 20. Brook’s home range during autumn/winter 2019 (blue circles) and spring/summer (yellow circles) showing similar use of habitat across these periods (data 15th March to 8th November 2019).
Click for a full description
White lines show rivers and roads. This is a visual representation of the results discussed in 5.3.5.
In Strathpeffer, like Emma, Fairburn ranged widely in the weeks before birth before concentrating her home range in agricultural land, but this still fell within the area she had used in winter/early spring (Figure 21).
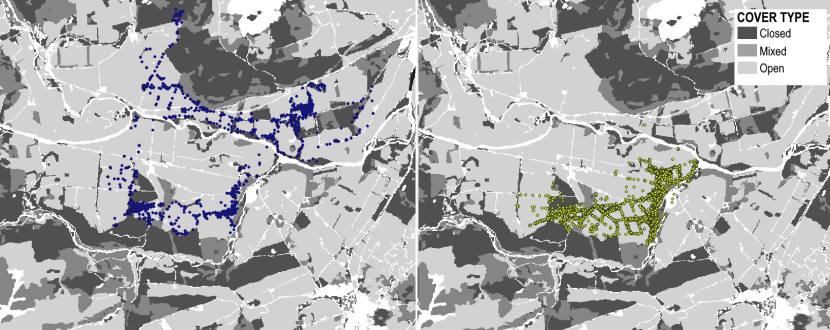
Figure 21. Fairburn’s home range during autumn/winter 2018 (blue circles) and spring/summer (yellow circles) showing similar use of habitat across these periods (data 14th March to 7th September 2018 when her collar fell off).
Click for a full description
This is a visual representation of the results discussed in 5.3.5.
4.4 Rest sites
Across all collared individuals several different rest sites were identified where cats spent three hours or more (e.g. see Bogie’s rest sites in Figure 22). Individuals used many of these rest sites more than once (mean = 40, SD = 33), some of which were very regularly used on an almost daily basis, the remainder were only used once (mean = 74, SD = 46) (Table 7). Rest sites were spread across the individuals’ home ranges. The number of rest sites used increased as the number of days a cat was monitored for increased, suggesting that rest sites are not a limiting factor (there are many suitable locations). Approximately 21% (range 1 – 258) of the rest sites identified by the ODBA data did not have GPS co-ordinates associated with them. For this collar make, the most likely scenario for this is because the cat was underground and the collar could not connect to the satellites at the 15min period.
| Cat ID | Priority Area | Total No rest periods identified | No. with XY and habitat data | No. without XY | Rest sites used more than once | Rest sites used once | Days GPS data collected for |
|---|---|---|---|---|---|---|---|
| Doll | AG | 538 | 352 | 186 | 54 | 90 | 483 |
| Lochie | AG | 52 | 52 | 0 | 11 | 17 | 113 |
| Woody | AG | 313 | 217 | 96 | 25 | 99 | 236 |
| Finley | AG | 25 | 5 | 20 | 0 | 5 | 15 |
| Markie | AG | 185 | 146 | 39 | 30 | 43 | 138 |
| Bogie | SB | 807 | 633 | 144 | 112 | 104 | 583 |
| Brook | SB | 345 | 301 | 44 | 50 | 117 | 239 |
| Brodie | SB | 45 | 34 | 11 | 1 | 8 | 29 |
| Clash | SB | 898 | 640 | 258 | 77 | 122 | 592 |
| Fergyl | SB | 197 | 197 | 0 | 21 | 40 | 138 |
| Emma | SB | 139 | 139 | 0 | 20 | 69 | 170 |
| Sprat | MO | 633 | 491 | 142 | 89 | 139 | 333 |
| Stonker | MO | 300 | 233 | 67 | 37 | 114 | 174 |
| Fairburn | SP | 216 | 215 | 1 | 33 | 56 | 178 |
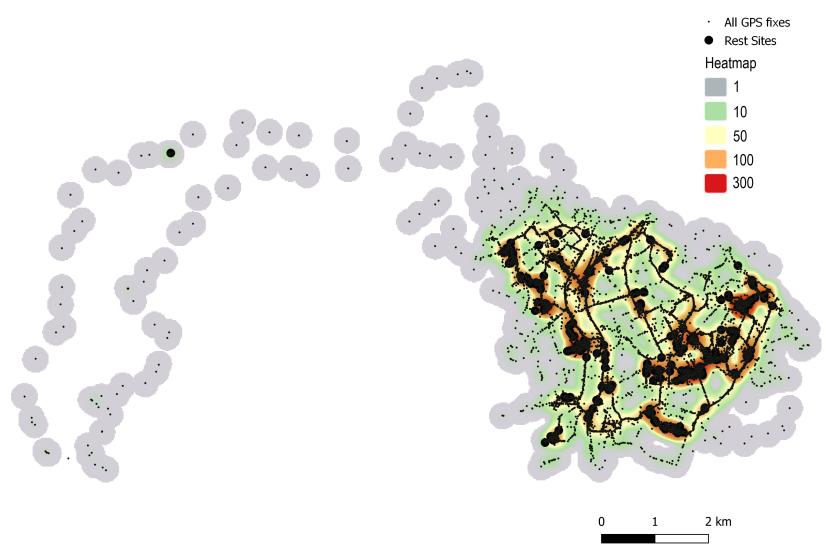
Map of Bogie, Strathbogie Priority Area showing all GPS locations
Click for a full description
Figure 22. Bogie (Strathbogie Priority Area) – showing all GPS locations, heatmap representing the total number of GPS fixes across his home range and rest sites between February 2018 and December 2019. The rest site at a farm during his 72hr exploration outside his normal home range can also be seen. This is a visual representation of some the results discussed in 5.4.
Cats showed a preference for resting in cover, in particular broadleaved and mixed woodland, gorse, clear-fell and young plantation, wind blow in coniferous woodland, as well as farm buildings in relation to their availability within the cats’ 100% MCPs, although there were individual differences in the proportion of rest sites each cat had in the different habitats (Figure 23 and Figure 24). For example, Doll, a female in the Angus Glens whose habitat was predominantly within FLS-owned land showed a preference for coniferous woodland, in particular wind blow but also clear-fell and rocky scree within areas of clear-fell and open dry heath. Fairburn, a female cat in the Strathpeffer Priority Area which is dominated by farmland, mostly located her rest sites within small patches of broadleaved woodland, patches of gorse, hay barns on farms, hedgerows on the edges of fields and areas of unimproved grassland along field boundaries. Markie, a male cat in the Angus Glens, was the only cat who showed some selection of heathland as a rest site, where he predominantly rested in rocky cairns within the heathland. In comparison, Woody, who overlapped some of Markie’s range, tended to select wind blow within coniferous woodland, and patches of clear-fell and young plantation to rest in. Bogie’s home range consisted predominantly of farmland. Favoured rest sites for him were hay barns on farms which had an abundance of rabbits, patches of gorse, unimproved grassland and hedgerows along field edges and a preference for the limited patches of broadleaved woodland that were scattered across his home range.
The majority of cats occupying areas with a lot of arable/grassland showed a strong selection of gorse habitats, farm buildings and broadleaved woodland in relation to the availability of these features. In Morvern, Sprat and Stonker often used hedgerows on the edges of grassland and areas of unimproved grassland to rest in and both these individuals showed a strong selection of broadleaved woodland.
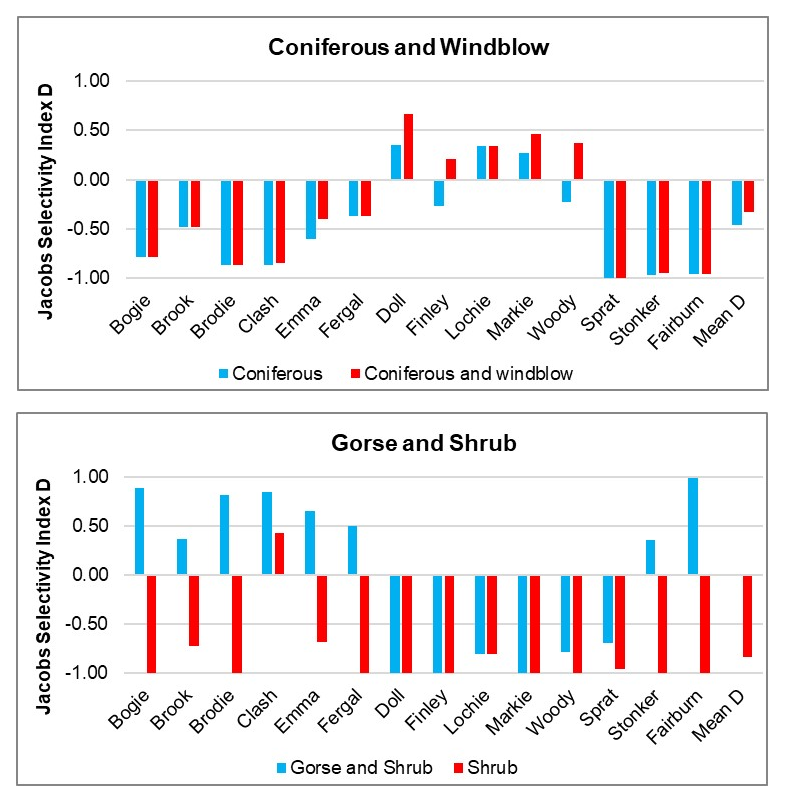
Figure 23. Selection of different habitats as rest sites in relation to their availability in an individual’s home range using Jacobs Index D. Values at 0 represent random use, 1 = strong selection of habitat as a rest site, -1 = avoidance of habitat
Click for a full description
for resting in. Gorse was often unidentified by the EUNIS dataset and was typically found bordering arable and grassland and woodland so the proportion of rest sites (r) in gorse was combined with the shrub data for comparison. Similarly, wind blow was combined with coniferous woodland for comparison with the proportion of rest sites in coniferous alone. Mean D across all cats is also shown for interest. This is a visual graphical representation of some the results discussed in 5.4.
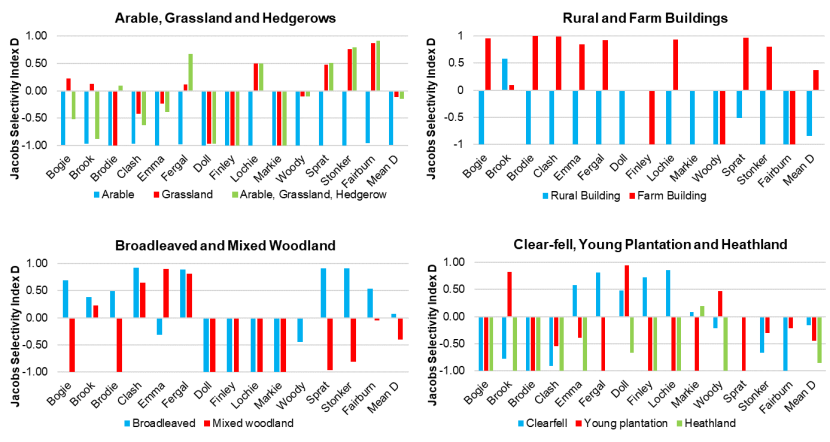
Figure 24. Selection of different habitats as rest sites in relation to their availability in the individual’s home range using Jacobs Index D.
Click for a full description
Values at 0 represent random use, 1 = strong selection of habitat as a rest site, -1 = avoidance of habitat for resting in. Hedgerows were often unidentified by the EUNIS dataset and were typically found bordering arable and grassland so the proportion of rest sites (r) in hedgerows was combined with the grassland and arable data for comparison. Mean D across all cats is also shown for interest. This is a visual graphical representation of some the results discussed in 5.4.
4.4.1 Natal dens
Two of the female cats collared were pregnant during the project:
- Emma was collared on 14th January 2019. During the summer, two young juveniles were seen crossing a farmyard close to where she was regularly detected using the GPS collars, which were likely her offspring. Based on movement patterns in the GPS data, it looks probable that she gave birth between 5th and 6th May 2019 as there was a decrease in her overall activity during this time (ranging only a total of 176m over a 24hr period compared to moving an average of 4000m / 24 hrs in the days preceding birth) and she remained in the same location for 17.5 hrs from 13:45 on the 5th May 2019 until 07:00 on the 6th May 2019. She continued to return to the same area within a small patch (0.004 km2) of mixed woodland/scrub running alongside a small stream every day for several weeks, spending several hours at a time (7-16hrs) not moving much, suggesting this is where she had the kittens.
On the 24th June 2019 she moved ~1.6km away to a patch of wind blow on the edge of a forestry track. She stayed in this area for two days, then moved again to a small patch of shrub on the edge of some clear-fell, returning to this area until the 29th June 2019 before moving again towards an area of gorse behind a rural property. From 13th July to 17th July 2019, she was located in a hedgerow between two fields of unimproved grassland. Between 18th July and the end of August 2019, she was located predominantly around a small farm which had a large rabbit population (Figure 27). Two sub-adults were trapped in this area later in winter and appeared on the same camera traps that she did. This is also where juvenile kittens were seen in late summer, so most likely these sub-adults were her kittens. She was ~ 6 weeks pregnant when she was re-trapped in early March 2020 to have her collar removed.
In addition to a reduced ranging behaviour, there was also a change in her activity patterns post birth when she became predominantly active between dawn and dusk instead of between dusk - dawn from the 5th May which continued until late July (~10 weeks) (Figure 25 & Figure 26).
- Fairburn was approximately six weeks pregnant when she was collared on the 14th March 2018. She was caught on camera on the 24th April looking heavily pregnant. She appears to have given birth between the 27th – 28th April 2018 in a wooden lean to against a ruined stone cottage, where she returned daily for the next four weeks for several hours (up to 16 hrs) at a time, suggesting she had the kittens there. After locating her at this site several times during May 2018 she then appeared to move again but no kittens were ever seen on camera before her collar came off, so it is unclear if they survived or not. Note, the collar allows data to be downloaded from a distance so den sites were not approached closely (<50m) to avoid disturbing a female with young kittens. Unlike Emma, there was also no obvious change in activity patterns during this period and instead she followed a similar pattern to the other cats collared; predominantly resting during the day (08:00 – 16:00) and being much more active at night (Figure 26). It is of note that even though she was detected within the structure using the GPS collar there was no discernible evidence of a cat or any other species using the area (e.g. flattened vegetation, tracks, scats), highlighting just how difficult it is to locate and identify wildcat natal sites.

Figure 25. Emma’s daily activity patterns between 8th April and 1st June 2019 pre and post birth period.
Click for a full description
Darker areas represent higher activity levels. There was a shift in activity patterns post birth from activity spread across a 24-hr period but more active between 16:00 – 08:00 to a distinct lack of activity during the night (18:00 – 06:00) for approximately 2 weeks (red rectangle). Instead, during this period, Emma was mostly active during daylight hours. This is a visual representation of some of the results for Emma’s activity patterns as detailed in section 5.4.1.
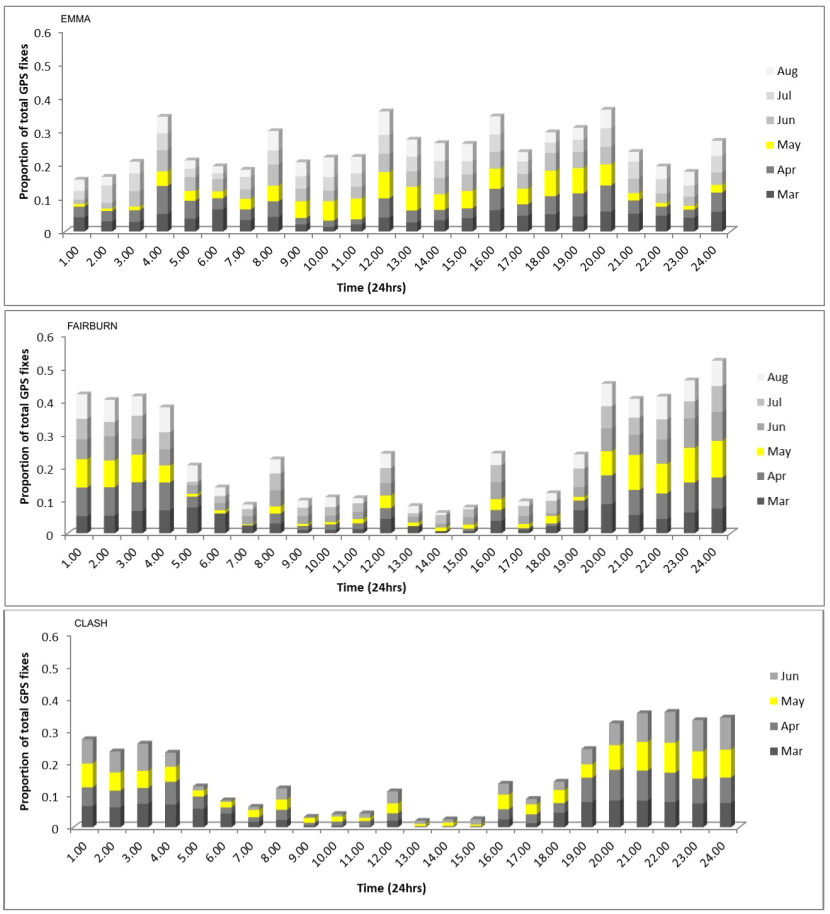
Figure 26. Comparison of proportion of GPS fixes obtained during different periods of the day for Emma, Fairburn and Clash (a male cat using much of the same home range as Emma).
Click for a full description
Clash and Fairburn both show a greater number of fixes obtained between 18:00 – 04:00 hrs, in comparison Emma shows a higher number of GPS fixes between 09:00-20:00 during May, post birth. Peaks in GPS fixes can be seen at 08:00, 12:00 and 16:00 hrs during the daytime, which generally correspond to the fixes taken every 4 hrs when the cat is inactive. This is a visual graphical representation of some of the results described in section 5.4.1.
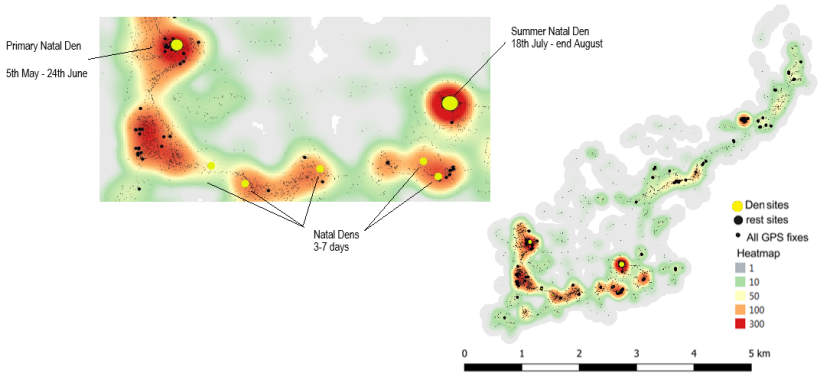
Figure 27. Emma – showing all GPS locations after being corrected, heatmap representing the number of GPS fixes to highlight areas of greatest density of use, rest sites between January – October 2019 and probable natal den sites
Click for a full description
between 5th May to end of August 2019.This is a visual representation of some of the results discussed for Emma’s rest sites and natal dens sites as discussed in section 5.4.1.
4.5 Resource Selection Function models
There were strong correlations (r>0.8) between one or more of the predictor variables in each individual cat model, the predictor/s that made the greatest biological sense for that individual and had the lowest VIF value (<10) were retained in each model respectively. Model results for each individual can be found in the following sections but are summarized below.
4.5.1 3rd order habitat selection (use of specific habitats within the home range)
Pooling the data showed that overall, the 14 wildcat hybrids were selecting arable landscapes, broadleaved woodland, areas of clear-fell, edge habitats, farm and rural buildings and low use roads and forest / farm tracks. However, as can be seen from Table 8 and the following individual RSF’s models, habitat use sometimes varied between different Priority Areas and between individuals within a Priority Area. For example, all collared cats in the Angus Glens (Doll, Finley, Lochie, Markie and Woody) had a significantly higher than random probability of using clear-fell and coniferous woodland with the probability of a GPS fix decreasing as the distance from these habitats increased. However, only Markie showed a significant selection of heathland where he spent a large proportion of the summer months (Table 12 and Figure 32).
Individuals found in Strathbogie and Strathpeffer all had a high probability of occurring within 0 - 50m of farm and rural buildings (Figure 28 and Table 8). Collared individuals within these Priority Areas often used hay barns on farms, not just as rest sites but as meeting points with other cats during the breeding months and as areas to hunt as well. Particularly in the Strathbogie Priority Area, most of the farms visited by the collared cats had rabbits in close vicinity. Individuals would also use the trees and hedgerows bordering rural properties. However, only Emma and Brook were significantly selecting coniferous woodland in the Strathbogie Priority Area. In several instances across all four Priority Areas, there was a medium to low use road running through the home range of a cat, and these individuals regularly used these roads to move across their home ranges. Stonker in particular had a high tendency to use a main B road bisecting his home range and the habitat either side of this road, which was primarily broadleaved and mixed woodland and patches of gorse and grassland. His behaviour could be explained in part by the fact that he was blind in one eye and may have found it easier to primarily traverse alongside a road.
All cats, with the exception of Fairburn, were significantly selecting edge habitat. In some cases, they were never more than 200m from an edge (dEdge was not included as a predictor variable in Fairburn’s model because it was highly correlated with distance to dArable (r = 0.93) and had a higher VIF value, but we note regular use of edges by this cat).
| Predictor | All cats | Angus Glens | Strathbogie | Morvern | Strathpeffer |
|---|---|---|---|---|---|
| dArable | -0.36556 | -0.89937 | -0.13962 | -0.83782 | -0.85389 |
| dBog | 0.56164 | 0.54081 | 0.38571 | 0.81488 | - |
| dBroadleaved | -0.29575 | -0.43347 | -0.20527 | -0.44776 | -0.58595 |
| dClear-fell | -0.06325 | -0.64835 | 0.29789 | -0.69561 | 0.25432 |
| dConiferous woodland | 0.11379 | -0.70988 | 0.22937 | 0.39934 | - |
| dGrassland | 0.06839 | 0.45525 | -0.02740 | -0.54485 | - |
| dHeathland | 0.31731 | -0.05599 | 0.44871 | - | - |
| dMixed Woodland | 0.26421 | -0.13115 | 0.42378 | -0.69929 | - |
| dShrub | 0.17111 | 0.45776 | 0.10505 | 0.29134 | -0.31485 |
| dYoung Plantation | 0.19037 | 0.25973 | 0.12216 | 0.45408 | 0.42712 |
| dBuilding | -0.57091 | 0.44709 | -0.60858 | -0.79639 | -0.81282 |
| dEdge | -0.26254 | -0.18189 | -0.27118 | -1.40665 | - |
| dRoadL | -0.33360 | -0.28218 | -0.36578 | -0.80315 | -0.08908 |
| dRoadH | 0.12148 | 0.70584 | 0.09821 | -0.56057 | 0.26022 |
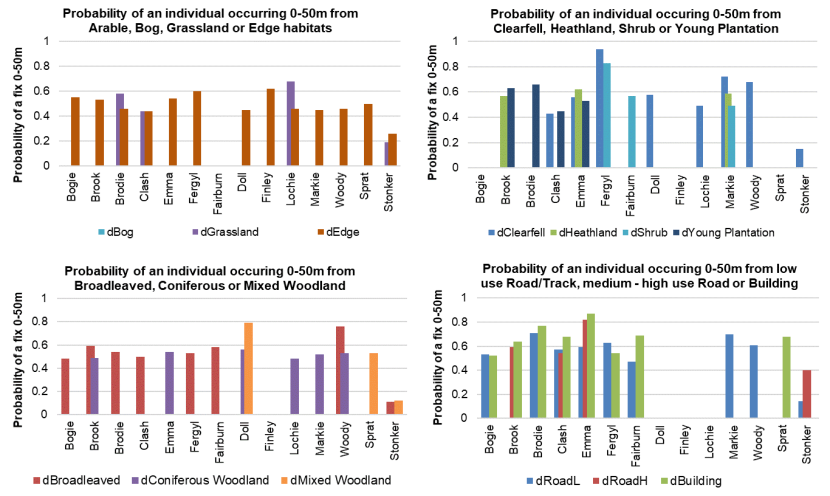
Figure 28. Summarises the probability of an individual occurring within 0-50m of the different habitats.
Click for a full description
Values are only shown where the probability of getting a fix decreased significantly from random (p<0.05) as the cat moved further away from these habitats. This is a visual graphical representation of some of the results described in section 5.5.1.
Angus Glens
A total of five individuals were collared in the Angus Glen Priority Area.
Doll
The model for 3rd order selection (habitat use within the home range) for Doll retained eight of the 14 variables. The co-efficient values indicate that Doll is most likely to be found in or close to clear-fell, coniferous woodland, mixed woodland and edge habitats with predicted probability of occurrence decreasing as distance from these landcover types increased (Table 9; Figure 29).
The model supports observations in the field that Doll was using clear-fell, particularly areas with good scree cover, patches of wind blow within coniferous woodland, areas of mixed woodland along the rivers and edge habitats, particular the edges of grassland bordering the forested areas on the upper slopes of the glen.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.942 | 0.037 | -25.287 | 2e-16 | - |
| Arable | Not incl. | Not incl. | Not incl. | Not incl. | 0.05 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0.03 |
| Broadleaved woodland | 0.076 | 0.030 | 2.549 | 0.01 | 0.002 |
| Coniferous woodland | -1.322 | 0.052 | -25.386 | 2e-16 | 0.3 |
| Clear-fell | -0.565 | 0.020 | -28.162 | 2e-16 | 0.04 |
| Grassland | 0.613 | 0.027 | 22.814 | 2e-16 | 0.1 |
| Heathland | 0.347 | 0.028 | 12.562 | 2e-16 | 0.3 |
| Mixed woodland | -1.068 | 0.042 | -25.287 | 2e-16 | 0.001 |
| Shrub | 0.552 | 0.021 | 26.724 | 2e-16 | 0.01 |
| Young plantation | Not incl. | Not incl. | Not incl. | Not incl. | 0.002 |
| Building | Not incl. | Not incl. | Not incl. | Not incl. | 0.0002 |
| Edge | -0.139 | 0.029 | -4.871 | 1.11E-06 | 187.4 km |
| RoadL | 0.142 | 0.034 | 4.130 | 3.63E-05 | 7 km |
| RoadH | Not incl. | Not incl. | Not incl. | Not incl. | 0 km |
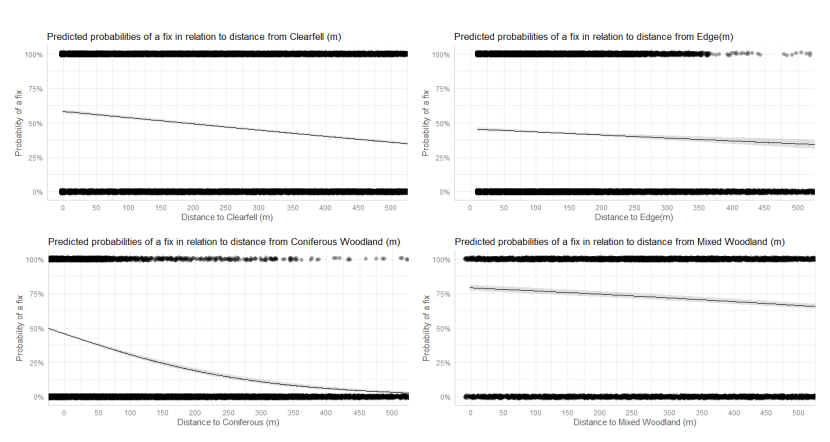
Figure 29. Shows the four main predictor variables Doll was most likely to be found in / within 50m of.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from clear-fell, coniferous woodland, edge habitat and mixed woodland. Black dots (jittered slightly on the Y axis to enable easier viewing) show the raw data for Doll (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the predictions. This is a visual graphical representation of some of the results described in Table 9.
Finley
The model for 3rd order selection (habitat use within the home range) for Finley retained eight of the 14 variables. The co-efficient values indicate that he is most likely to be found in or close to edge habitats with predicted probability of occurrence decreasing significantly as distance from edges increased (Table 10; Figure 30). This supports observations in the field that Finley was primarily using the edges of coniferous woodland and the higher grassy slopes on the edges of open heathland, but note we only have 15 days’ worth of data for this individual to date.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.10468 | 0.06819 | -1.535 | 0.12 | - |
| Arable | Not incl. | Not incl. | Not incl. | Not incl. | 0.06 |
| Broadleaved woodland | Not incl. | Not incl. | Not incl. | Not incl. | 0.002 |
| Bog | 0.89293 | 0.14748 | 6.055 | 1.41E-09 | 0.05 |
| Coniferous woodland | Not incl. | Not incl. | Not incl. | Not incl. | 0.3 |
| Clear-fell | 2.13E-01 | 8.38E-02 | 2.539 | 0.01 | 0.04 |
| Grassland | 0.23843 | 0.07866 | 3.031 | 0.002 | 0.2 |
| Heathland | 3.57E-02 | 1.00E-01 | 0.356 | 0.72 | 0.3 |
| Mixed woodland | Not incl. | Not incl. | Not incl. | Not incl. | 0.001 |
| Shrub | 5.23E-02 | 8.33E-02 | 0.628 | 0.53 | 0.007 |
| Young plantation | 8.61E-02 | 6.84E-02 | 1.259 | 0.21 | 0.001 |
| Building | Not incl. | Not incl. | Not incl. | Not incl. | 0.002 |
| Edge | -8.00E-01 | 1.29E-01 | -6.223 | 4.88E-10 | 205 km |
| RoadL | 2.42E-01 | 1.44E-01 | 1.683 | 0.09 | 8.41 km |
| RoadH | Not incl. | Not incl. | Not incl. | Not incl. | 0 km |

Figure 30. Shows the main predictor variable Finley was most likely to be found in / within 50m of habitat edges.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from edge habitat. Black dots (jittered slightly to enable easier viewing) show the raw data for Finley (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 10.
Lochie
The model for 3rd order selection (habitat use within the home range) for Lochie retained 13 of the 14 variables. The co-efficient values indicate that he is most likely to be found in or close to arable, coniferous woodland, clear-fell, grassland and edge habitats with predicted probability of occurrence decreasing significantly as distance from these landcover types increased (Table 11; Figure 31).
The model supports observations in the field that Lochie was using farmland and patches of gorse within open grassland where there were high densities of rabbits; he was also using small, isolated patches of clear-fell and coniferous woodland scattered across his home range.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.32963 | 0.04678 | -7.046 | 1.84E-12 | - |
| Arable | -1.99413 | 0.10448 | -19.087 | 2E-16 | 0.2 |
| Broadleaved woodland | -0.10533 | 0.07737 | -1.361 | 0.17 | 0.03 |
| Bog | -0.01015 | 0.08265 | -0.123 | 0.90 | 0.01 |
| Coniferous woodland | -0.20344 | 0.05928 | -3.432 | 0.0006 | 0.1 |
| Clear-fell | -0.15477 | 0.07436 | -2.081 | 0.04 | 0.02 |
| Grassland | -0.69845 | 0.09257 | -7.545 | 4.52E-14 | 0.3 |
| Heathland | 0.42758 | 0.08899 | 4.805 | 1.55E-06 | 0.08 |
| Mixed woodland | 1.77128 | 0.11341 | 15.619 | 2E-16 | 0.001 |
| Shrub | 0.43883 | 0.05897 | 7.442 | 9.90E-14 | 0.2 |
| Young plantation | Not incl. | Not incl. | Not incl. | Not incl. | 0.04 |
| Building | -0.03649 | 0.06637 | -0.55 | 0.58 | 0.0008 |
| Edge | -0.17466 | 0.06801 | -2.568 | 0.010 | 301.7 km |
| RoadL | 0.89279 | 0.0555 | 16.087 | 2E-16 | 2.2 km |
| RoadH | 0.70584 | 0.07699 | 9.168 | 2E-16 | 14.3 km |
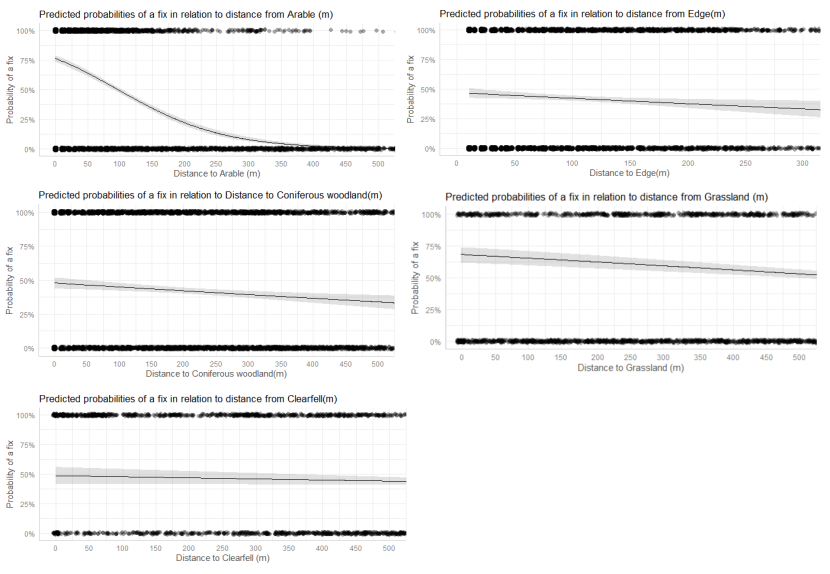
Figure 31. Shows the five main predictor variables Lochie was most likely to be found in / within 50m of.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from arable, coniferous woodland, clear-fell, edge habitat and grassland. Black dots (jittered slightly to enable easier viewing) show the raw data for Lochie (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 11.
Markie
The model for 3rd order selection (habitat use within the home range) for Markie retained 11 of the 14 predictor variables. The co-efficient values indicate that he is most likely to be found in or close to coniferous woodland, clear-fell, heathland, shrub, edge habitat and low use roads/tracks (in this case forestry and estate tracks) with predicted probability of occurrence decreasing significantly as distance from these predictors increased (Table 12, Figure 32).
The model supports observations in the field that Markie was using clear-fell, areas of wind blow within coniferous woodland, the forest tracks through coniferous woodland, patches of gorse and the rock cairns within the open heathland and edges of woodland and clear-fell and small rivers.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.42087 | 0.03667 | -11.478 | 2E-16 | - |
| Arable | 0.08916 | 0.06791 | 1.313 | 0.19 | 0.03 |
| Broadleaved woodland | Not incl. | Not incl. | Not incl. | Not incl. | 0.003 |
| Bog | 0.4835 | 0.0595 | 8.127 | 2E-16 | 0.18 |
| Coniferous woodland | -0.6609 | 0.0752 | -8.789 | 4.42E-16 | 0.24 |
| Clear fell | -1.44782 | 0.07844 | -18.457 | 2E-16 | 0.08 |
| Grassland | 0.30296 | 0.05584 | 5.425 | 5.80E-08 | 0.01 |
| Heathland | -0.96536 | 0.0393 | -24.561 | 2E-16 | 0.35 |
| Mixed woodland | Not incl. | Not incl. | Not incl. | Not incl. | 0.0002 |
| Shrub | -0.26498 | 0.05781 | -4.584 | 4.57E-06 | 0.09 |
| Young plantation | 1.5407 | 0.08834 | 17.441 | 2E-16 | 0.007 |
| Building | 0.44709 | 0.05007 | 8.929 | 2E-16 | 0.0001 |
| Edge | -0.26398 | 0.05398 | -4.89 | 1.01E-06 | 538.6 km |
| RoadL | -1.79612 | 0.06181 | -29.059 | 2E-16 | 32.3 km |
| RoadH | Not incl. | Not incl. | Not incl. | Not incl. | 0 km |
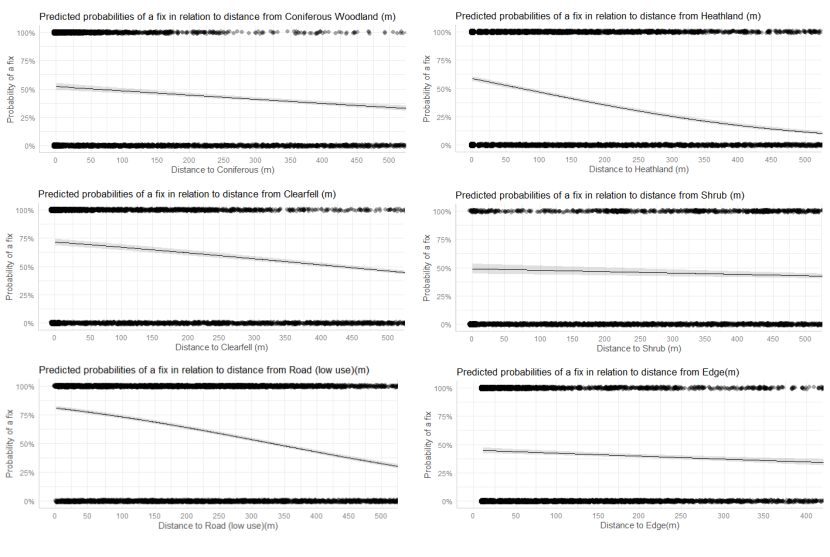
Figure 32. Shows the six main predictor variables Markie was most likely to be found in / within 50m of.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from coniferous woodland, clear-fell, low use roads/tracks, heathland and shrub. Black dots (jittered slightly to enable easier viewing) show the raw data for Markie (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 12.
Woody
The model for 3rd order selection (habitat use within the home range) for Woody retained 9 of the 14 predictor variables. The co-efficient values indicate that he is most likely to be found in or close to arable, broadleaved woodland, coniferous woodland, clear-fell, edge habitats and low use roads/tracks with predicted probability of occurrence decreasing significantly as distance from these predictors increased (Table 13: Figure 33).
The model supports observations in the field that Woody was using clear-fell, areas of wind blow within coniferous woodland, the forest tracks through coniferous woodland, patches of broadleaved woodland around some of the rural residences and along the river edges, small isolated patches of coniferous/broadleaved woodland in open grassland and the edges of arable land. As well as forest tracks, he was also regularly moving along a low use B road that ran along the southern edge of his home range.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.25963 | 0.02428 | -10.694 | 2E-16 | - |
| Arable | -0.74226 | 0.03958 | -18.751 | 2E-16 | 0.08 |
| Broadleaved woodland | -1.1406 | 0.03512 | -32.473 | 2E-16 | 0.03 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0.01 |
| Coniferous woodland | -0.60496 | 0.03792 | -15.953 | 2E-16 | 0.1 |
| SClear-fell | -0.95199 | 0.03245 | -29.341 | 2E-16 | 0.02 |
| Grassland | Not incl. | Not incl. | Not incl. | Not incl. | 0.3 |
| Heathland | -0.02346 | 0.02867 | -0.818 | 0.41 | 0.08 |
| Mixed woodland | 0.53462 | 0.04202 | 12.723 | 2E-16 | 0.001 |
| Shrub | Not incl. | Not incl. | Not incl. | Not incl. | 0.2 |
| Young plantation | 0.15366 | 0.02542 | 6.044 | 1.50E-09 | 0.04 |
| Building | Not incl. | Not incl. | Not incl. | Not incl. | 0.0006 |
| Edge | -0.15864 | 0.04512 | -3.516 | 0.0004 | 503.6 km |
| RoadL | -0.65014 | 0.03272 | -19.869 | 2E-16 | 29.8 km |
| RoadH | Not incl. | Not incl. | Not incl. | Not incl. | 0.7 km |
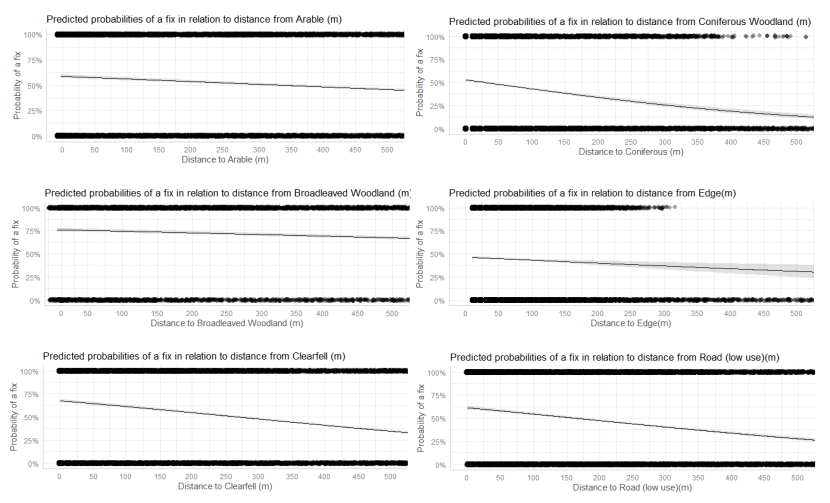
Figure 33. Shows the six main predictor variables Woody was most likely to be found in / within 50m of.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from arable, broadleaved woodland, clear-fell, coniferous woodland, low use roads/tracks and edge habitats. Black dots (jittered slightly to enable easier viewing) show the raw data for Woody (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 13.
Strathbogie
Six individuals were collared in the Strathbogie Priority Area.
Bogie
The model for 3rd order selection (habitat use within the home range) for Bogie retained 13 of the 14 variables plus an additional predictor dRailway. The co-efficient values indicate that Bogie is most likely to be found in or close to (<50m) arable, broadleaved woodland, edge habitats, buildings and low intensity use roads/ tracks with predicted probability of occurrence decreasing as distance from these predictors increased (Table 14; Figure 34).
The model supports observations that Bogie was predominantly using farmland, hay barns, patches of gorse/shrub and hedgerows and field margins and fields with high density of rabbits in.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.25459 | 0.01346 | -18.916 | 2E-16 | - |
| Arable | -0.16042 | 0.01986 | -8.078 | 6.58E-16 | 0.7 |
| Broadleaved woodland | -0.12005 | 0.02004 | -5.989 | 2.11E-09 | 0.02 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Coniferous woodland | 0.41178 | 0.0187 | 22.019 | 2E-16 | 0.02 |
| Clear-fell | 0.73356 | 0.02051 | 35.758 | 2E-16 | 0.002 |
| Grassland | 0.0965 | 0.01801 | 5.358 | 8.41E-08 | 0.1 |
| Heathland | 0.93213 | 0.01942 | 48.004 | 2E-16 | 0.0002 |
| Mixed woodland | 0.50514 | 0.02311 | 21.858 | 2E-16 | 0.02 |
| Shrub | 0.13375 | 0.01696 | 7.885 | 3.15E-15 | 0.03 |
| Young plantation | 0.91819 | 0.02083 | 44.089 | 2E-16 | 0.003 |
| Building | -0.32071 | 0.02108 | -15.216 | 2E-16 | 0.01 |
| Edge | -0.37495 | 0.01889 | -19.854 | 2E-16 | 574.3 km |
| RoadL | -0.32839 | 0.0171 | -19.208 | 2E-16 | 42.7 km |
| RoadH | 0.49607 | 0.0184 | 26.966 | 2E-16 | 35.8 km |
| Railway | -0.01408 | 0.02253 | -0.625 | 0.53 | 5.1 km |
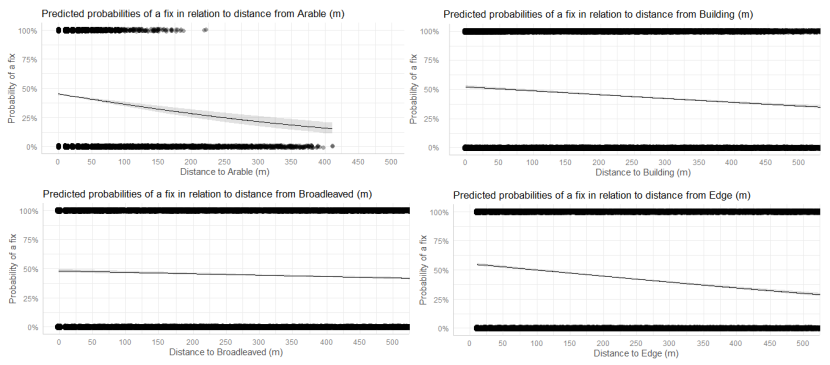
Figure 34. Shows the four predictor variables Bogie was most likely to be found in / within 50m of based on the small-scale model.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from arable, broadleaved woodland, edge habitat and buildings. Black dots (jittered slightly to enable easier viewing) show the raw data for Bogie (Use = 1) and Random points (Use = 0) used in the model. Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 14.
Brodie
The model for 3rd order selection (habitat use within the home range) for Brodie retained 11 of the 14 variables. The co-efficient values indicate that Brodie is most likely to be found in or close to (<50m) broadleaved woodland, grassland, young plantation, buildings (in this case he regularly used farm hay barns), edge habitats, and low use roads (in this case he often used a single track along the forest edges) with predicted probability of occurrence decreasing as distance from these predictors increased. Brodie was never found >150m away from edge habitats (Table 15; Figure 35).
The model corresponds to observations that this individual was predominantly using edges of farmland, hay barns, areas of young plantation, patches of broadleaved woodland along river edges and patches of gorse/shrub within open grassland and predominantly moving through his home range along a quiet low use road.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.67295 | 0.08481 | -7.935 | 2.11E-15 | - |
| Arable | Not incl. | Not incl. | Not incl. | Not incl. | 0.2 |
| Broadleaved woodland | -0.71616 | 0.09606 | -7.456 | 8.95E-14 | 0.03 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Coniferous woodland | 0.24683 | 0.08692 | 2.84 | 0.005 | 0.5 |
| Clear-fell | 0.1773 | 0.08294 | 2.138 | 0.03 | 0.09 |
| Grassland | -0.5484 | 0.09299 | -5.898 | 3.68E-09 | 0.03 |
| Heathland | 0.27156 | 0.10831 | 2.507 | 0.01 | 0.03 |
| Mixed woodland | 0.52113 | 0.10936 | 4.765 | 1.89E-06 | 0.01 |
| Shrub | 0.95945 | 0.10608 | 9.044 | 2E-16 | 0.01 |
| Young plantation | -0.73894 | 0.08518 | -8.675 | 2E-16 | 0.04 |
| Building | -1.58046 | 0.11103 | -14.235 | 2E-16 | 0.001 |
| Edge | -0.59763 | 0.09953 | -6.004 | 1.92E-09 | 265.2 km |
| RoadL | -1.60305 | 0.12092 | -13.257 | 2E-16 | 22.5 km |
| RoadH | Not incl. | Not incl. | Not incl. | Not incl. | 1.8 km |
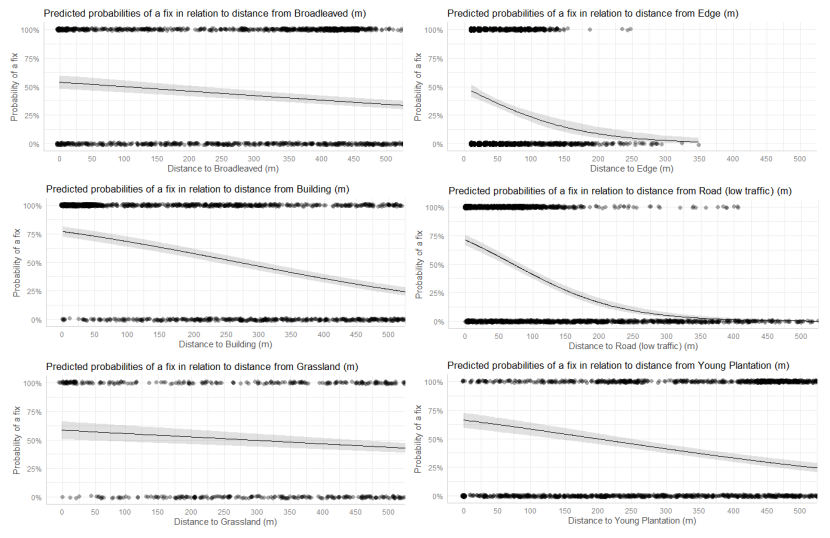
Figure 35. Shows the six predictor variables Brodie was most likely to be found in / within 50m of based on the small-scale model.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from broadleaved woodland, buildings, grassland, edge habitat, low use roads/tracks and young plantation. Black dots (jittered slightly to enable easier viewing) show the raw data for Brodie (Use = 1) and Random points (Use = 0) used in the model. Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 15.
Brook
The model for 3rd order selection (habitat use within the home range) for Brook retained 13 of the 14 variables. The co-efficient values indicate that Brook is most likely to be found in or close to (<50m) arable, broadleaved woodland, coniferous woodland, heathland, young plantation, buildings (he regularly used the areas adjacent to farm buildings and rural residential houses), edge habitats, and medium to high use roads with predicted probability of occurrence decreasing as distance from these predictors increased (Table 16; Figure 36).
The model corresponds to observations that Brook was predominantly using hedgerows along field edges, large areas of Christmas tree plantations and patches of gorse/shrub in grassland, the edges of coniferous woodland and farmland, he was also using large areas of mature broadleaved (Beech; Fagus sylvatica and Oak; Quercus spp.) woodland and regularly using the hedgerows and ditches running alongside a low use B road that bisected his home range.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.09355 | 0.01981 | -4.723 | 2.33E-06 | - |
| Arable | -0.27475 | 0.03968 | -6.924 | 4.38E-12 | 0.5 |
| Broadleaved woodland | -0.30389 | 0.02636 | -11.527 | 2E-16 | 0.05 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Coniferous woodland | -4.91E-02 | 2.38E-02 | -2.064 | 0.04 | 0.1 |
| Clear-fell | 4.98E-01 | 2.81E-02 | 17.753 | 2E-16 | 0.004 |
| Grassland | -0.07385 | 0.03884 | -19.016 | 0.25 | 0.05 |
| Heathland | -1.61E-01 | 4.56E-02 | -3.538 | 0.0004 | 0.02 |
| Mixed woodland | 0.05627 | 0.03607 | 1.56 | 0.12 | 0.02 |
| Shrub | 1.22E-01 | 2.59E-02 | 4.709 | 2.49E-06 | 0.02 |
| Young plantation | -0.65745 | 0.02858 | -23.001 | 2E-16 | 0.2 |
| Building | -0.51639 | 0.03336 | -15.48 | 2E-16 | 0.002 |
| Edge | -0.24151 | 0.02547 | -9.484 | 2E-16 | 300.1 km |
| RoadL | 1.65E-01 | 3.25E-02 | 5.099 | 3.42E-07 | 15.2 km |
| RoadH | -0.33834 | 0.03856 | -8.774 | 2E-16 | 6.7 km |
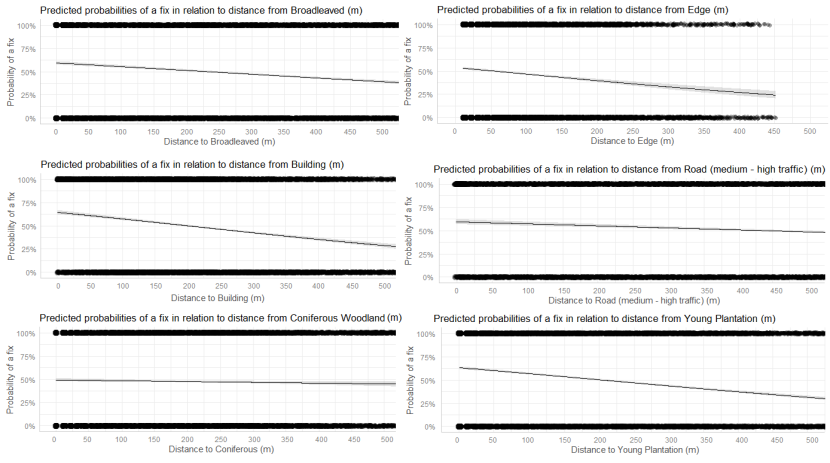
Figure 36. Shows six of the eight predictor variables Brook was most likely to be found in / within 50m of based on the small-scale model.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from broadleaved woodland, buildings, coniferous woodland, edge habitat, medium use roads, young plantation, grassland, and heathland. Black dots (jittered slightly to enable easier viewing) show the raw data for Brook (Use = 1) and Random points (Use = 0) used in the model. Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 16.
Clash
The model for 3rd order selection (habitat use within the home range) for Clash retained 13 of the 14 variables. The co-efficient values indicate that Clash is most likely to be found in or close to (<50m) broadleaved woodland, clear-fell, grassland, young plantation, buildings, edge habitats, and both low and medium to high use roads with predicted probability of occurrence decreasing significantly as distance from these predictors increased (Table 17; Figure 37).
The model supports observations in the field that Clash regularly used several different hay barns within his home range, hedgerows and field edges, patches of clear-fell and grassy areas with rabbits in (Table 17). He also used small isolated patches of young plantation and broadleaved woodland within more open grassland. He regularly used forest and farm tracks to move across his home range and used the verges of a medium use B road that bisected his home range on a regular basis.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.38278 | 0.01754 | -21.827 | 2E-16 | - |
| Arable | -0.05474 | 0.03907 | -1.401 | 0.16 | 0.2 |
| Broadleaved woodland | -0.38653 | 0.03333 | -11.597 | 2E-16 | 0.007 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0.01 |
| Coniferous woodland | 0.4121 | 0.02267 | 18.176 | 2E-16 | 0.4 |
| Clear-fell | -0.07831 | 0.02506 | -3.125 | 0.002 | 0.06 |
| Grassland | -0.06272 | 0.02211 | -2.836 | 0.005 | 0.1 |
| Heathland | 0.21148 | 0.02362 | 8.953 | 2E-16 | 0.1 |
| Mixed woodland | 0.27343 | 0.02398 | 11.403 | 2E-16 | 0.006 |
| Shrub | 0.05459 | 0.03361 | 1.624 | 0.10 | 0.08 |
| Young plantation | -0.12074 | 0.0213 | -5.668 | 1.45E-08 | 0.06 |
| Building | -1.16994 | 0.03579 | -32.694 | 2E-16 | 0.001 |
| Edge | -0.17413 | 0.01674 | -10.401 | 0.02 | 852.2 km |
| RoadL | -0.66045 | 0.0262 | -25.213 | 2E-16 | 45 km |
| RoadH | -0.52993 | 0.02953 | -17.943 | 2E-16 | 10.7 km |
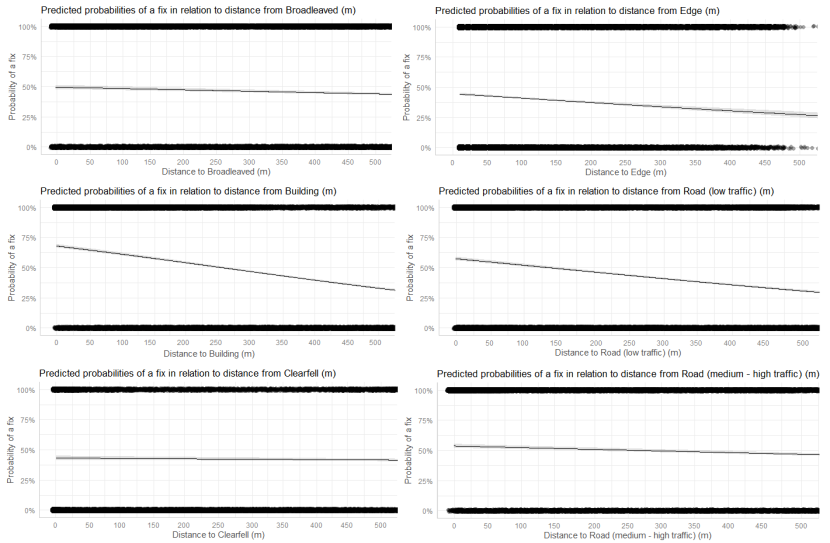
Figure 37. Shows six of the eight predictor variables Clash was most likely to be found in / within 50m of based on the small-scale model.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from broadleaved woodland, buildings, clear-fell, edge habitat, low and medium use roads, young plantation and grassland. Black dots (jittered slightly to enable easier viewing) show the raw data for Clash (Use = 1) and Random points (Use = 0) used in the model. Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 17.
Emma
The model for 3rd order selection (habitat use within the home range) for Emma retained all 14 variables. The co-efficient values indicate that Emma is most likely to be found in or close to (<50m) coniferous woodland, clear-fell, heathland, young plantation, buildings (in this case she spent several months within close vicinity of a farm where she moved her kittens), edge habitats, and both low and medium-high use roads with predicted probability of occurrence decreasing as distance from these predictors increased (Table 18; Figure 38).
The model corresponds to observations that Emma was largely using field edges, patches of clear-fell and gorse/shrub, areas of young plantation, edges of heathland and wind blow within coniferous woodland. She also used forest and farm tracks and the verges of a medium use B road that ran along the southern edge of her home range. During the mating season Emma travelled long distances to spend time in several different hay barns where two collared males (Clash and Brodie) were also detected.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.05865 | 0.027 | -2.173 | 0.03 | - |
| Arable | 0.17787 | 0.05732 | 3.103 | 0.002 | 0.1 |
| Broadleaved woodland | 0.27524 | 0.06186 | 4.45 | 8.60E-06 | 0.01 |
| Bog | 0.38571 | 0.06769 | 5.698 | 1.21E-08 | 0.002 |
| Coniferous woodland | -0.46287 | 0.03649 | -12.685 | 2E-16 | 0.5 |
| Clear-fell | -0.33772 | 0.03536 | -9.55 | 2E-16 | 0.09 |
| Grassland | 0.30526 | 0.03872 | 7.883 | 3.20E-15 | 0.06 |
| Heathland | -0.31418 | 0.03518 | -8.93 | 2E-16 | 0.04 |
| Mixed woodland | -0.01919 | 0.05326 | -0.36 | 0.72 | 0.007 |
| Shrub | 0.48867 | 0.05586 | 8.748 | 2E-16 | 0.04 |
| Young plantation | -0.15039 | 0.0301 | -4.997 | 5.83E-07 | 0.1 |
| Building | -1.38498 | 0.04578 | -30.256 | 2E-16 | 0.001 |
| Edge | -0.26759 | 0.03251 | -8.232 | 2E-16 | 755.7 km |
| RoadL | -0.41529 | 0.035 | -11.865 | 2E-16 | 45.2 km |
| RoadH | -0.99428 | 0.07996 | -12.435 | 2E-16 | 5.3 km |
Fergyl
The model for 3rd order selection (habitat use within the home range) for Fergyl retained 13 of the 14 variables plus an additional predictor dRailway. The co-efficient values indicate that Fergyl is most likely to be found in or close to (<50m) arable, broadleaved woodland, clear-fell, shrub, edge habitat, buildings and low use roads/tracks with predicted probability of occurrence decreasing as distance from these predictors increased (Table 19; Figure 39).
The model corresponds to observations that Fergyl was largely using farmland, isolated patches of broadleaved / mixed woodland (including along a railway running through his home range), hedgerows and field margins, clear-fell and large areas of gorse/shrub within open grassland. He also spent a lot of time around farm buildings, particularly hay barns and in the mixed/broadleaved woodlands surrounding these buildings. Fergyl’s home range was bisected by a very busy B road which he occasionally crossed; he regularly used farm tracks and low use roads to move across his home range. He spent a lot of time in a small patch of mixed woodland adjacent to this road that had a high rabbit density.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.07464 | 0.03301 | -2.261 | 0.02 | - |
| Arable | -0.07976 | 0.03778 | -2.111 | 0.03 | 0.7 |
| Broadleaved woodland | -0.13837 | 0.05432 | -2.547 | 0.01 | 0.03 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Coniferous woodland | 0.59212 | 0.05227 | 11.328 | 2E-16 | 0.02 |
| Clear-fell | -1.43329 | 0.1024 | -13.997 | 2E-16 | 0.007 |
| Grassland | -0.0344 | 0.09769 | -0.352 | 0.72 | 0.08 |
| Heathland | 1.13576 | 0.10045 | 11.307 | 2E-16 | 0 |
| Mixed woodland | 1.08133 | 0.08069 | 13.4 | 2E-16 | 0.02 |
| Shrub | -1.00063 | 0.05843 | -17.126 | 2E-16 | 0.02 |
| Young plantation | 0.6019 | 0.08015 | 7.51 | 5.93E-14 | 0 |
| Building | -0.21455 | 0.05012 | -4.28 | 1.86E-05 | 0.02 |
| Edge | -0.48409 | 0.05448 | -8.886 | 2E-16 | 351.6 km |
| RoadL | -0.57412 | 0.05166 | -11.114 | 2E-16 | 18.2 km |
| RoadH | 0.12204 | 0.04975 | 2.453 | 0.01 | 28.8 km |
| Railway | 0.53243 | 0.06275 | 8.485 | 2E-16 | 6.2 km |
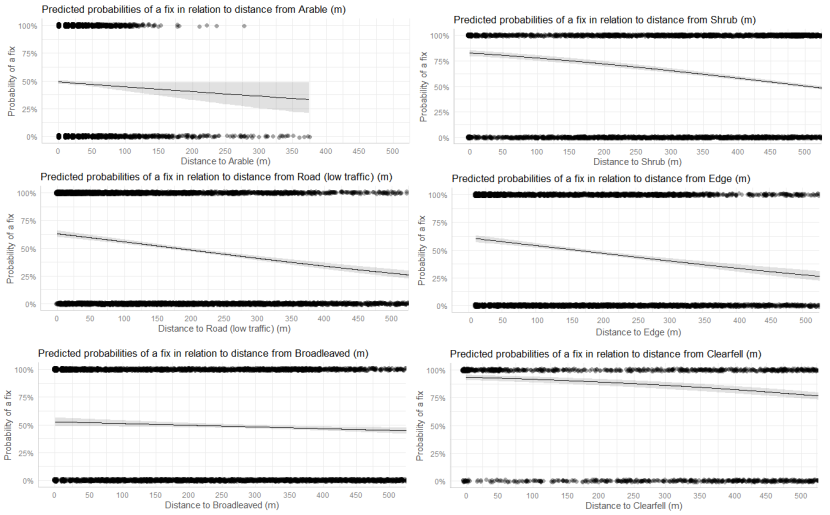
Figure 39. Shows six of the seven predictor variables Fergyl was most likely to be found in / within 50m of based on the small-scale model.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from broadleaved woodland, buildings, clear-fell, low use roads, edge habitat and shrub. Black dots (jittered slightly to enable easier viewing) show the raw data for Fergyl (Use = 1) and Random points (Use = 0) used in the model. Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 19.
Strathpeffer
Only one individual was collared in the Strathpeffer Priority Area.
Fairburn
The model for 3rd order selection (habitat use within the home range) for Fairburn retained 10 of the 14 variables. The co-efficient values indicate that Fairburn is most likely to be found in or close to (<50m) arable, broadleaved woodland, shrub, buildings, and low use roads/tracks with predicted probability of occurrence decreasing as distance from these predictors increased (Table 20; Figure 40).
The model corresponds to observations that Fairburn was largely using farmland, especially the hedgerows bisecting the fields, farm buildings such as hay barns, patches of broadleaved woodland, particularly around farms and river edges, clear-fell and large areas of gorse/shrub on the edges of heathland and grassland. She also used the farm tracks and low use roads within her home range.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.23121 | 0.02899 | -7.975 | 1.52E-15 | - |
| Arable | -0.85389 | 0.06608 | -12.922 | 2E-16 | 0.6 |
| Broadleaved woodland | -0.58595 | 0.03614 | -16.215 | 2E-16 | 0.08 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Coniferous woodland | 0.06676 | 0.03676 | 1.816 | 0.07 | 0.2 |
| Clear-fell | 0.25432 | 0.03369 | 7.55 | 4.36E-14 | 0.04 |
| Grassland | Not incl. | Not incl. | Not incl. | Not incl. | 0.01 |
| Heathland | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Mixed woodland | 0.02491 | 0.03293 | 0.757 | 0.45 | 0.04 |
| Shrub | -0.31485 | 0.02982 | -10.557 | 2E-16 | 0.01 |
| Young plantation | 0.42712 | 0.03084 | 13.849 | 2E-16 | 0.01 |
| Building | -0.81282 | 0.03926 | -20.703 | 2E-16 | 0.002 |
| Edge | Not incl. | Not incl. | Not incl. | Not incl. | 480.1 km |
| RoadL | -0.08908 | 0.03802 | -2.343 | 0.02 | 28.5 km |
| RoadH | 0.26022 | 0.02937 | 8.86 | 2E-16 | 10.2 km |
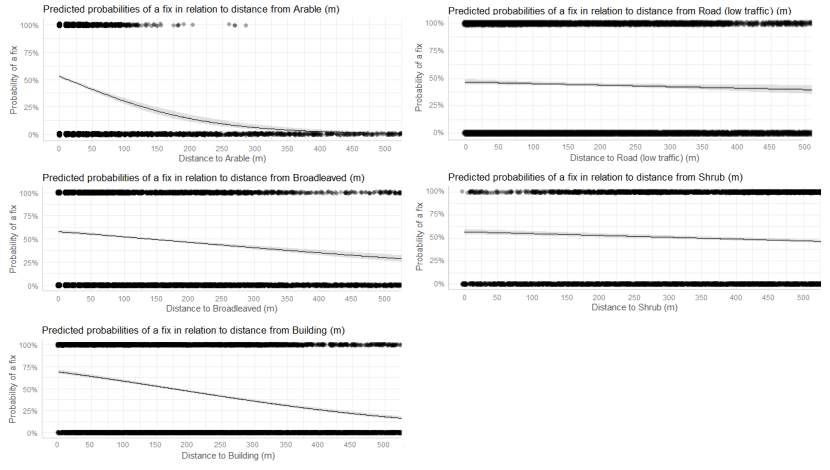
Figure 40. Shows the five main predictor variables Fairburn was most likely to be found in / within 50m of. Probability of getting a fix for this individual decreased significantly further away from arable, broadleaved woodland, buildings
Click for a full description
low use roads/tracks and shrub. Black dots (jittered slightly to enable easier viewing) show the raw data for Fairburn (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 20.
Morvern
Two individuals were collared in the Morvern Priority Area.
Sprat
The model for 3rd order selection (habitat use within the home range) for Sprat retained 10 of the 14 predictor variables. The co-efficient values indicate that she is most likely to be found in or close to arable, mixed woodland, buildings, and edge habitats with predicted probability of occurrence decreasing significantly as distance from these predictors increased. Sprat was never found >100m from mixed woodland or >50m from edge habitats (Table 21; Figure 41).
The model corresponds to observations that Sprat tended to be found in the mixed woodland around the edges of gardens and rural properties, bordering streams, rivers and fields. She also used hedgerows and field margins and stuck close to edge habitats.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -0.9336 | 0.3169 | -2.9460 | 0.003 | - |
| Arable | -0.856 | 0.034 | -25.285 | 2E-16 | 0.3 |
| Broadleaved woodland | 0.06781 | 0.0508 | 1.335 | 0.18 | 0.06 |
| Bog | Not incl. | Not incl. | Not incl. | Not incl. | 0.01 |
| Coniferous woodland | 0.39934 | 0.02847 | 14.025 | 2E-16 | 0.07 |
| Clear-fell | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Grassland | 0.03428 | 0.03239 | 1.058 | 0.29 | 0.1 |
| Heathland | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Mixed woodland | -1.756 | 0.25264 | -6.95 | 3.64E-12 | 0.2 |
| Shrub | 1.68242 | 0.12636 | 13.314 | 2E-16 | 0.2 |
| Young plantation | Not incl. | Not incl. | Not incl. | Not incl. | 0.01 |
| Building | -0.79639 | 0.05888 | -13.525 | 2E-16 | 0.001 |
| Edge | -0.57428 | 0.15686 | -3.661 | 0.0003 | 185.9 km |
| RoadL | -0.05573 | 0.08442 | -0.66 | 0.51 | 11.1 km |
| RoadH | 0.84686 | 0.09207 | 9.198 | 4.44E-09 | 2.7 km |
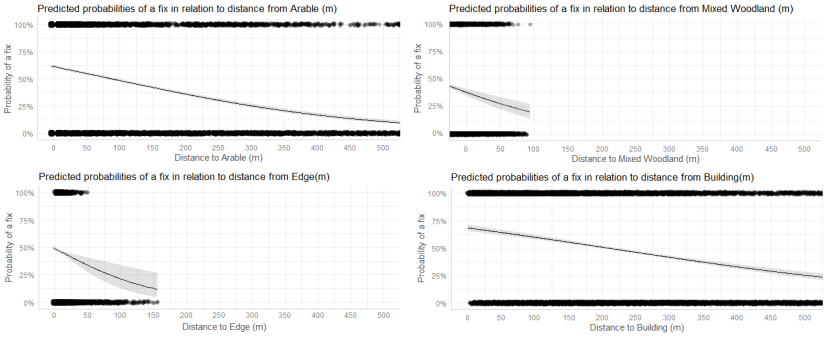
Figure 41. Shows the four main predictor variables Sprat was most likely to be found in / within 50m of.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from arable, buildings, edge habitat and mixed woodland. Black dots (jittered slightly to enable easier viewing) show the raw data for Sprat (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 21.
Stonker
The model for 3rd order selection (habitat use within the home range) for Stonker retained 12 of the 14 predictor variables. The co-efficient values indicate that he is most likely to be found in or close to arable, broadleaved woodland, clear-fell, grassland, mixed woodland, low use roads/tracks (in this case mainly single estate tracks), medium/high use roads and edge habitat with predicted probability of occurrence decreasing significantly as distance from these predictors increased (Table 22; Figure 42).
The model corresponds to observations that Stonker was often located in the patches of woodland, open grassland fields and gorse/shrub that bordered a medium use B road that bisected the length of his home range. He primarily spent his time moving along this road from one end of his home range to another. He also used field boundaries including hedgerows, small areas of clear-fell and low use private tracks and roads.
| Parameter | Co-efficient Estimate | Std.Error | Z.Value | Pr(>|z|) | Proportion / length (km) of predictor variables within home range |
|---|---|---|---|---|---|
| (Intercept) | -2.49592 | 0.1034 | -24.138 | 2E-16 | - |
| Arable | -0.58516 | 0.12498 | -4.682 | 2.84E-06 | 0.08 |
| Broadleaved woodland | -0.44776 | 0.11129 | -4.023 | 5.74E-05 | 0.03 |
| Bog | 0.81488 | 0.07482 | 10.891 | 2E-16 | 0.03 |
| Coniferous woodland | -0.02824 | 0.03846 | -0.734 | 0.46 | 0.5 |
| Clear-fell | -0.69561 | 0.04982 | -13.964 | 2.88E-16 | 0.06 |
| Grassland | -0.54485 | 0.06662 | -8.178 | 2.88E-16 | 0.04 |
| Heathland | Not incl. | Not incl. | Not incl. | Not incl. | 0 |
| Mixed woodland | -0.5695 | 0.08854 | -6.432 | 1.26E-10 | 0.04 |
| Shrub | 0.15041 | 0.04022 | 3.74 | 0.0002 | 0.2 |
| Young plantation | 0.45408 | 0.04663 | 9.738 | 2E-16 | 0.03 |
| Building | Not incl. | Not incl. | Not incl. | Not incl. | 0.0005 |
| Edge | -2.03457 | 0.13624 | -14.933 | 2E-16 | 840.3 km |
| RoadL | -0.80315 | 0.08653 | -9.282 | 2E-16 | 41.5 km |
| RoadH | -2.62583 | 0.11153 | -23.543 | 2E-16 | 17.9 km |
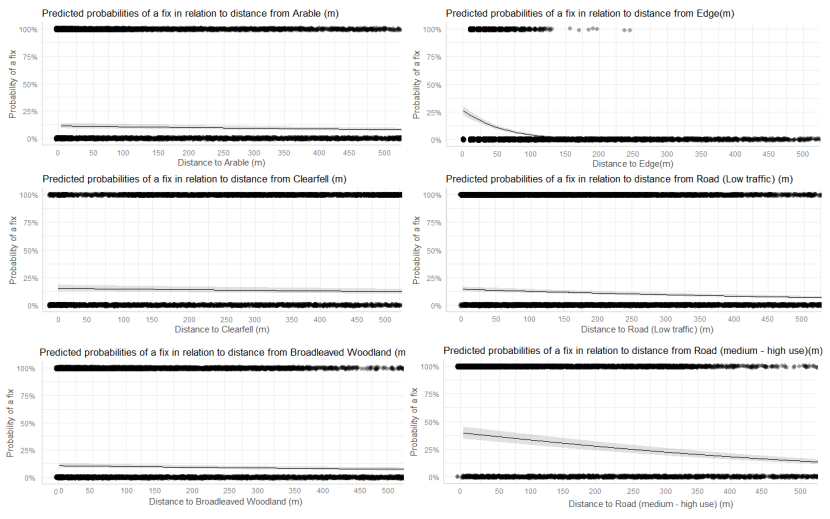
Figure 42. Shows the six main predictor variables Stonker was most likely to be found in / within 50m of.
Click for a full description
Probability of getting a fix for this individual decreased significantly further away from arable, buildings, edge habitat, mixed woodland and low use roads/tracks. Black dots (jittered slightly to enable easier viewing) show the raw data for Stonker (Use = 1) and Random points (Use = 0). Grey area shows upper and lower confidence levels for the prediction. This is a visual graphical representation of some of the results described in Table 22.
4.5.2. Other 4 habitat features used by wildcat hybrids
From the GPS data it was clear that the wildcat hybrids were also using some specific features in the landscape that were not included in the RSF models primarily because datasets were not available for all of them. But they are mentioned briefly here because of their perceived importance/interest in how wildcat hybrids use the landscape.
Linear features such as fence lines and stone walls (dykes) were regularly used by individuals moving across open pasture (e.g. grassland grazed by sheep/horse/cattle) and arable land (Figure 44) and along the edges of woodland and open habitat. Isolated groups of trees within open fields and lines of trees going across grassland or agriculture were also used by individuals to move across the landscape and trees bordering the edges of rural gardens and around farms, such as mature hedgerows were also often used (Figure 45).
In addition to these linear features, as modelled for Bogie and Fergyl, in the Strathbogie Priority Area, these two males used the railway line to move up and down sections of their home range and as a route across a major river in the area. Although as noted in the RSF models for these two individuals, they did not tend to use the railway line more than other habitats in their home range. The railway in this Priority Area offered good cover along either side in the form of shrub/gorse, unimproved grassland and clumps of broadleaved/mixed woodland along its edges and also appeared to be used as a route across a major river in the area (Figure 43).
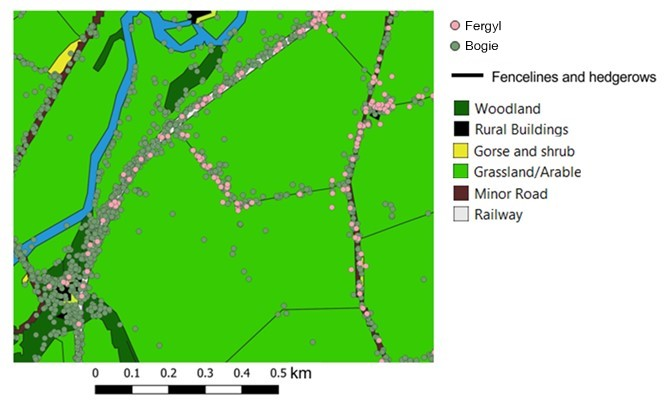
Figure 43. Shows the use of some of the railway line in the Strathbogie Priority Area by two males, Bogie and Fergyl.
Click for a full description
This figure also features some of the use of minor (low use) roads, farm buildings and patches of mixed woodland/broadleaved woodland around rural buildings, as well as a fence line through an open field. This is a visual representation of some of the text detailed in 5.5.2.

Figure 44. Shows the use of a stone wall and hedgerows bordering arable fields by Bogie in the Strathbogie Priority Area.
Click for a full description
This is a visual representation of some of the text detailed in 5.5.2.
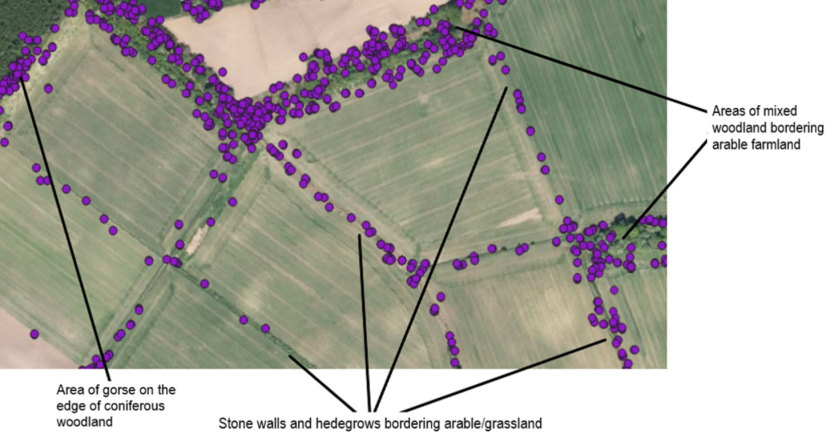
Figure 45. Shows the use of a stone wall and hedgerows bordering arable and pasture fields by Fairburn in the Strathpeffer Priority Area, as well as the use of small patches of mixed
Click for a full description
and broadleaved woodland around the edges of some of these fields and a patch of gorse at the edge of coniferous woodland. This is a visual representation of some of the text detailed in 5.5.2.
5. Discussion
Below we discuss the findings of the preliminary analysis of the GPS data in relation to both other radio tracking studies that have been carried out on the European wildcat and the implications for future wildcat conservation.
5.1 Home range size
The mean home range sizes of wildcat hybrids in this study (95% MCP, 13.94 km2 and 18.15 km2 for females and males respectively) are larger than has previously been reported for both male and female wildcats and other wild-living cats with wildcat pelage in Scotland. Radio tracking studies in the 1970s and 1990s estimated home range sizes of 1.72 – 6.3 km2 for females and 1.75 – 9.9 km2 for males, with home ranges larger for both sexes in the west where rabbits were limited or non-existent (Scott et al., 1993, Corbett, 1979, Daniels, 1997). As noted in the introduction, these early studies were carried out using VHF radio tracking which provides less detail than GPS tracking. More recently, using GPS radio tracking, Campbell (2015), using GPS fixes every 10 minutes, estimated a mean 95% MCP for wild-living cats across different study sites as 5.73 km2 (females) and 6.34 km2 (males) but with the largest home range sizes reaching 8.98 km2 for a female and 17.9 km2 for a male. Campbell’s study was limited by collar longevity, however. So, the mean home range is likely smaller than it would have been if some of the collars had lasted longer, as noted by the large variations in home range size in relation to battery duration.
Home range sizes (95% MCP) reported here are larger than some produced by studies on wildcats in continental Europe, for example; 2.83 km2 and 9.5 km2 females and male wildcats in Croatia (Popović et al., 2017), 2.28 km2-13.71 km2 for wildcats in Portugal (Monterroso et al., 2009) and 4.78 km2 – 6.53 km2 and 6.17– 22.74 km2 in Sicily (Anile et al., 2017) but comparable to others. For example, the mean home range size of male wildcats in parts of Sicily, and home range sizes of male wildcats in Germany (6.77 km2 – 22.38 km2 for females - males) (Jerosch et al., 2017) and both sexes in parts of Spain 17.55 km2 – 20.09 km2 (Oliveira et al., 2018a) and females in the Iberian Peninsula ranged from 3.14 -59.78 km2 (mean ± SE: 13.56 ± 7.25 km2; Olivera et al., 2018). Home range sizes of the wildcat hybrids in Strathbogie were also much larger than that of some domestic cats that were GPS radio tracked in the same Priority Area during 2013-2014 (95% MCP 0.007-0.179 km2) by Campbell (2015) and 2018-2019 (95% MCP 0.002-0.17 km2) by White (2019). Our findings are consistent with the Germain et al. (2008) study in France who found that there was no significant difference in home range sizes of wildcats and wildcat hybrids but that both had larger home range sizes than domestic cats.
Home range size is known to be influenced by resources, in particular prey availability, which has been shown to be inversely correlated with home range size, such that at lower prey densities, home range sizes are larger (Litvaitis et al., 1986, McNab, 1983, Schmidt, 2008, Herfindal et al., 2005). As discussed in more detail in Campbell et al., (2021a), densities of rabbits (a key prey species) can be affected by different factors including disease (Aebischer et al., 2011), red deer abundance (Lozano et al., 2007) and land management practices such as the legal control of rabbit populations. Rodent densities can also be greatly affected by red deer grazing (Smit et al., 2001, Flowerdew and Ellwood, 2001) and forest structure and management (Ecke et al., 2001, Michał and Rafał, 2014, Suchomel et al., 2012). Other landscape management practices also have a large impact on small mammals, such as the connectivity of woodland patches on farmland (Fitzgibbon, 1997), livestock grazing intensity, particularly sheep grazing in upland habitats (Evans et al., 2006, Wheeler, 2008) and agricultural intensification or changes (e.g. Halliez et al., 2015). Rabbits were found in several of the Priority Areas, including Strathbogie and parts of the Angus Glens, and in fact rabbit densities were extremely high where the young male cat Lochie was found, which could account for his relatively small home range size. Rabbit populations in Strathbogie were often clustered around farms and both Bogie and Clash had their main rest sites at farms with high rabbit densities. However, both these males had large home ranges suggesting that prey densities were not the only factor influencing home range size. Other important resources are mates, and these two males in particular roamed widely visiting most of the farms within their home ranges, with more than one cat often visiting the same farm. Other uncollared individuals (both wildcats and hybrids) were noted to be appearing at some of these locations during the breeding season as well.
Population density can also have a large effect on home range size; for example, Benson et al. (2006) found that bobcats (Lynx rufus) reduced their home range sizes as population density increased, and Dahle et al. (2006) found a similar trend in Scandinavian brown bears (Ursus arctos). During this project, long term camera trapping surveys carried out by the SWA field team and a large number of volunteers, have established that population densities of wild-living cats in general, including wildcats and wildcat hybrids are low in all the Priority Areas (Campbell et al., 2021a). As noted for wildcats with large home range sizes in Spain (Oliveira et al., 2018b), the large home range sizes of wildcat hybrids in Scotland could be highlighting a combination of potential issues for wildcats including limited prey availability, low population densities/mate availability and suboptimal habitat quality for wildcats in general.
One individual, Sprat, in Morvern had the smallest home range overall (4.43 km2), which is more comparable to that of female wildcats in mainland Europe, as noted above. One explanation for this is that her range encompassed a relatively large proportion of well-connected mixed woodland including mature hazel (Corylus avellane), oak (Quercus sp.), ash (Fraxinus excelsior) and beech (Fagus sylvatica) trees that had good understorey. This type of habitat offers good cover and prey availability. This compares to the home range of another female wildcat hybrid radio collared in a previous study within the Morvern Priority Area, but in an area with less mixed deciduous woodland and more coniferous plantation and open grassland which was twice the size of Sprat’s (8.98 km2; Campbell (2015)). In addition, there were at least four male wildcat hybrids using parts of her range, including Stonker, indicating lack of mates was not a potential issue for her, further reducing the need for a large home range. We should also note that she suffered a leg injury for several weeks during the summer, but noted no significant decrease in daily distance travelled during this period so we cannot attribute her smaller home range size to this.
5.1.1 Monthly and sex variation in size
Monthly and sex variation in home range sizes of wildcat hybrids shows a similar pattern to that reported for the European wildcat. Female home ranges were smaller during the breeding months and male home ranges largest during the winter/spring mating period, as noted by other studies (Wittmer, 2001, Campbell, 2015, Liberek, 2002, Sarmento et al., 2006, Corbett, 1979, Daniels, 1997). Interestingly, Anile et al. (2017) noted that sub adult females in their study in Sicily had a similar home range size to males, suggesting that this might be due to exploration of the surrounding habitat in order to find a “free” home-range for their first gestation and parturition. Here, Emma, a young female (estimated to be 1-2 years old when collared), had a large home range size of almost 30km2 and it may be that she was a dispersing female looking for her own territory. Her home range size peaked during the breeding season and it could be seen from the data that she was visiting several farm buildings that were encompassed by the home ranges of at least one of the four male cats collared in the area. She had kittens in the spring/summer. It is likely therefore that the large home range size was a combination of looking for breeding opportunities and either searching for a suitable area to birth or establishing a home range when collared. She was not recollared due to the difficulty in her re-trapping to remove the first collar, so we don’t know if she would have continued to have such a large home range. Similar behaviour was observed in a breeding female nearby during Campbell’s (2015) earlier GPS study, but again no post-breeding data were available. The older male wildcat hybrids in this study also showed similar patterns to mature male wildcats by having larger home range sizes than male sub-adults (Anile et al., 2017), something Daniels et al. (2001) also noted in Scotland. Other studies have reported that adult male wildcats have larger home ranges than females (Daniels et al., 2001, Biro et al., 2004, Germain et al., 2008, Monterroso et al., 2009, Anile et al., 2017) and younger male home range sizes tend to be a similar size to mature female home ranges (Daniels et al., 2001). These previous studies were carried out using VHF radio tracking, but we note that Jerosch et al. (2017), who also used GPS radio collars, found that although male seasonal home range sizes were larger than females, it was not significantly so.
5.1.2 Season variation
Seasonal shift in home range occurred in some Priority Areas but not in others. For example, in Strathbogie, collared individuals were observed to maintain relatively stable home range sizes covering a mixture of farmland and coniferous woodland. But there was some shift in home range use by Clash, likely in response to a female, Emma, establishing a territory and having kittens during the same period. In the Strathpeffer Priority Area, although Fairburn ranged widely during the winter/early spring months, the area she settled in (an agricultural area with crops of mainly barley) once she had given birth was still an area she had used during winter/early spring. Both collared individuals in Morvern showed no obvious spatial shift between winter and summer months.
In comparison, in the Angus Glens, two of the five individuals collared in this Priority Area showed a seasonal shift in range. Markie moved from the edges of coniferous plantation in the winter, up into open heathland (in this case dry heath) in the summer, presumably to predate on ground nesting birds, young leverets and possibly water voles, where he rested in patches of gorse and rock cairns. Doll also used dry heath in the spring/summer around the edges of the coniferous woodland she used in the winter months. (A third individual, Finley, resided in the same area as Doll, but we lost track of him after 15 days so did not observe this behaviour.) These three individuals resided at altitudes of >700m, the highest home ranges of all 14 wildcat hybrids collared and some of the areas used by them were composed of open dry heath with a good mixture of mid-height heather, alpine grassland and rocky cairns.
During the field work, in addition to the ground nesting birds observed, leverets were regularly seen and both water vole and field vole signs found, suggesting good prey availability. This behaviour of switching ranges to the heathland (or more open habitats) was noted by Corbett (1979) with some of the wildcats he studied becoming temporary residents in grouse moorlands during the summer months, and it is one of the reasons camera trap surveys are normally carried out during winter months in Scotland, to maximise the chances of detecting these cats when they come down off the open hills. Although upland dry heathlands are often managed as grouse moorlands, putting the cats at some risk of accidental or deliberate persecution, we should note that the landowners and gamekeepers involved in the SWA project and areas the collared cats roamed were very positive about wildcats and happy to have them on their land.
During this project we were able to notify the neighbouring grouse estates of the presence of potential wildcats on their land with the help of the SWA field staff, which also provided an opportunity to remind them how to distinguish wildcats from feral cats based on the pelage criteria. This provides a good example of how data from the GPS collars could be used to help direct reactive conservation management actions on the ground. The use of grouse moorland by wildcats has been known for many years. Prior to receiving legal protection in 1988, wildcats were considered a pest species and were legally controlled by gamekeepers. For example, in 1979 on one estate alone, snaring accounted for 58% of recorded wildcat deaths (Kitchener, 1995). During Daniels (1997) radio tracking study of wild-living cats (many of which did not display wildcat phenotypes) in the Angus Glens, 42% of radio collared cats were killed during predator control procedures. And during a study in Glen Tanar (Corbett, 1969), 74% (eight individuals) of his radio collared wildcats died as a result of persecution. Control and persecution of the wildcat over the past century has undoubtedly had a significant impact on its population status (Langley and Yalden, 1977). This work, however, highlights how important non-intensively managed grouse moorlands could be for wildcats in some parts of Scotland. Clearly this type of habitat could have an important role to play in wildcat conservation in Scotland in terms of providing areas of potentially suitable summer habitat if it is managed favourably and low densities of feral cats are maintained. However, continued work with grouse estates is required to further reduce persecution risks and promote wildlife conservation.
5.1.3 Annual variation and home range fidelity
The longevity of the collars and re-collaring of some individuals provided an opportunity to examine annual variation in home range use by two males, Bogie and Clash and a female, Doll. Bogie exhibited stable home range sizes and location over the two years he was followed for, excluding the three-day venture outside his normal home range. Here he travelled for ~35 km over a 72-hour period stopping at a farm during one day before returning to his normal home range area. Similar behaviour was noted by Corbett (1979) who found one male wildcat travelled over 40km during a six-week period during spring. In comparison, Clash, who was collared for three consecutive periods, had a similar home range size and location in 2018 and 2019 but in 2020 there was a marked shift where he moved further south, away from the coniferous woodland towards more open farmland. His home range size also significantly decreased in 2020. The most obvious explanation for this behaviour was the establishment by Emma of her home range and natal dens in an area Clash had previously used on a regular basis. Despite his shift in range, he still continued to use one of his most frequented rest sites, a hay barn which was situated within 500m of where Emma moved with her young towards the end of the summer.
Doll also showed a decrease in home range size in 2020 compared to 2019; one possible explanation for this was the presence of the young male, Finley, who had appeared in the winter of 2019/2020 and seemed to have replaced an older neutered feral male cat who had previously been the only other known resident in Doll’s range. Another reason for her decreased size in range might have been the higher number of visitors to the area during COVID-19 lockdown compared to previous years, with local staff and rangers reporting a huge increase in hikers, dog walkers and wild campers. Previous studies have shown that the European wildcat is sensitive to high levels of human presence, which can cause elevated levels of stress, especially during the breeding season and reduced occupancy (Piñeiro et al., 2012, Anile et al., 2019). Indeed, Piñeiro et al. (2012) recommended that tourist numbers should be controlled during the breeding period and areas of restricted public use maintained to reduce the impact of high visitor numbers on wildcats.
5.1.4 Home range overlap
ome range overlap occurred and varied between individuals. Two females, Emma and Sprat, overlapped with several males and two young males, Fergyl and Brodie, overlapped with the older males Bogie and Clash. This follows previous studies in Scotland which found that there was some overlap in home range between males and females (Corbett, 1979, Daniels, 1997), with adult males overlapping ~20-30% with each other, females ~10% with each other and females with males ~70% (Daniels, 1997). Jerosch et al. (2017) found an adult female wildcat overlapped other cats by 86%, which is similar to Emma’s overlap with the three males (Brodie, Clash and Bogie) and Sprat’s overlap with Stonker. There were also several other non-collared cats (both wildcat and hybrid) caught on camera trap within the home ranges of all individuals collared, indicating overlapping amongst many different cats was occurring. As noted by Daniels (1997) and Corbett (1979), wildcats are seldom seen in the same area at the same time, and this was also the case here with wildcat hybrids. In particular, Emma and Clash shared a large part of their home ranges and also used the same hay barn to rest in during the daytime on regular occasions but not at the same time, indicating some temporal partitioning was occurring.
5.2 Rest sites and natal dens
5.2.1 Rest sites
Wildcat hybrids in this study used several different rest sites across their home range where they spent three hours or more. We found that female hybrids (N = 4) used 20-89 rest sites on a regular basis and a further 56-139 sites just once across their range over a period of 6-11 months. We found that male hybrids (N = 10) used 1-112 rest sites on a regular basis and a further 5-122 sites just once across their range over a period of 15 days - 20 months. This compares to other studies on the European wildcat using the same type of e-obs GPS collars, where, for example, in Germany, a study found that male wildcats (n=16) used 76-330 and females (n=14) 53-162 rest sites over a period of 10 – 14 months (Simon et al., 2021).
In Greece, Migli et al. (2021) found that male wildcats (N = 2) used 9-23 regular rest sites and females (N = 2) 19-23 regular rest sites over 8-9 months. In Scotland, Campbell (2015) found wild-living cats had a mean of 11 rest sites per cat (range 1 – 34) and noted the number detected depended on the time interval over which data were gathered. Here we also found that the number of rest sites increased in relation to the longevity of collars, which lasted much longer than Campbell’s study (with cats followed for 5-98 days) which would explain the larger number found here. Presumably if suitable rest sites were rare these cats would return to use the same restricted number over time. This suggests that in general rest site availability is not a limiting factor on wildcat presence. However, this does not necessarily mean that secure shelters suitable for denning kittens (see below) are not limiting since the above rest sites can also include open-air lie-ups in cover habitat. Furthermore, a variety of different natal den sites in different habitats in close proximity may be required by breeding females to provide protection for kittens as they develop (Campbell 2015, see also below).
Here, collared individuals used a mixture of different habitats to rest in during the day. Cats showed a preference for resting in cover, in particular broadleaved woodland, gorse, clear-fell, wind blow and coniferous woodland, mixed woodland, as well as farm buildings in relation to their availability. This is similar to findings by other studies in Scotland (Campbell, 2015, Corbett, 1979, Tomkies, 1991) and for wildcats in mainland Europe (e.g. Gotz and Roth 2005; Potočnik et al., 2005; Jerosch et al., 2010). Simon et al., (2021) highlights the importance of wind blow as both breeding (natal den) and rest sites for wildcats and as hunting grounds. Jerosch et al. (2010) highlight the importance of deadwood structures as rest sites and recommended that forest management strategies enhance the structural heterogeneity by (natural) gap dynamic and with a high amount of deadwood, such as crowns or brushwood.
As also noted in Campbell's (2015) study, gorse was frequently used as a resting site. Gorse supports many prey species, provides good cover for cats, and protection from the weather throughout the year. Scrub habitat including gorse is known to be important to wildcats across parts of Europe, providing cover and supporting high prey densities (Ferreira 2003; Monterroso et al., 2009; Lozano 2010; Silva et al., 2013a), particularly for rabbits when it is located on the edge of grasslands. In Scotland, Scott et al. (1993) noted the importance of scrub for wildcats on the west coast. This potentially poses a significant issue for wildcats in Scotland because gorse is often cleared for tree planting, in particular as part of forest management activities and to improve livestock grazing. Retaining gorse on the edges of forestry/grassland and agricultural land could therefore help support wildcat conservation in some areas and ideally gorse removal would not be carried out during the main denning period April – August in areas with wildcat present.
Several of the male collared cats each had a hay barn they regularly returned to during the daytime. The use of farm buildings has also been noted by previous studies in Scotland (Corbett, 1979, Daniels, 1997, Campbell, 2015). This exposes wildcats to several potential risks including hybridisation with farm cats, highlighting the importance of neutering farm cat colonies, risk of rodenticide poisoning and risk of disturbance/persecution. High levels of exposure to rodenticides in some wild-living cats have been identified that could impact individual survival (Bacon et al., 2021) and farms often use rodenticides to control rodent populations. During this study, cats resting in hay barns were often potentially disturbed during normal farm practices, especially during the baling season, but the farmers were aware the cats used their hay barns and tried very hard to minimise disturbance where possible.
Farming activities did not appear to stop the cats from using these structures. Many of the farmyards had farm cats (neutered) and/or dogs which did not seem to affect the collared individual’s behaviour. Bogie spent several months using the hay barn of a farm which had a lot of chickens, along with high densities of rabbits. The owners noted no predation on their chickens, but regular rabbit carcasses were found. In the Angus Glens Priority Area, Lochie regularly used parts of a grousing estate, but the keepers reported no obvious predation of grouse by him, which they put down to the very high rabbit densities. Bogie was also collared just before sustained periods of unusually heavy and sustained snowfall beginning in March 2018. Studies on wildcats have shown that when snow depth is above 10cm, wildcats tend to stay in cover or move to lower elevations (Heptner and Sludskii, 1972, Corbett, 1979, Mermod and Liberek, 2002, Jerosch et al., 2017). In this case, Bogie spent four days sheltering in a hay barn on a farm which had a small patch of woodland next to the barn where there were plenty of rabbits. Some studies have found that snowy winters are a critical period for the survival of wildcats, with young animals dying more often than others (Heptner and Sludskii, 1972, Muntyanu et al., 1992), partially due to the difficulty in hunting small mammals that are under snow cover.
Hay barns not only provide shelter for cats in both cold and hot weather but also tend to support large populations of rodents, providing a source of food even in harsh weather conditions (and rabbits are often present nearby). Given that the weather in Scotland is likely to become more extreme due to climate change, retaining old buildings present in the landscape that will provide shelter may be a feasible step to mitigate some of the impacts of heavy snowfall for wildcats away from farms and small holdings where they are at potential risk of hybridisation, secondary rodenticide poisoning and persecution. For example, Muntyanu et al. (1992) found that in Romania, wildcats were often killed during winter months when they entered poultry houses (87% of anthropogenic deaths). However, Tomkies (1991) noted predation on chickens by a wildcat when the local rabbit population suddenly crashed due to myxomatosis. Emma was caught on camera with a chicken in her mouth at one end of her home range that lacked rabbits, highlighting a risk of potential conflict in rural areas following local rabbit population crashes or in areas of low rabbit numbers.
5.2.2 Natal dens
Two females, Fairburn and Emma, gave birth during late April/early May 2018, the timing consistent with other studies (Daniels, 1997, Campbell, 2015, Matthews, 1941). Both cats appeared to roam very widely before giving birth. For example, Fairburn crossed a major road and a large river using a very narrow but busy road bridge. One author has seen similar behaviour previously when tracking a mother before she gave birth (Campbell, 2015). Emma moved natal dens after a few weeks, then used several natal dens for shorter periods when presumably the kittens became more mobile and were old enough to start hunting (Lindemann, 1953, Campbell, 2015), a behaviour also noted by other authors. Simon et al., (2021) noted a total of 59 natal den sites for 14 female wildcats in Germany over the 8-week period post birth. Here Emma used a mixture of dense mixed woodland, areas of wind blow and shrub and hedgerows as natal den sites. As noted above, wind blow is considered an important denning habitat for wildcats because it is highly structured and complex, providing plenty of cover and prey (Simon, 2021) and Gotz et al., (2009) and Simon (2021) found that 89% of natal dens after the birth location were in dense deadwood structures on the ground.
We suspect that the other female, Fairburn, might have lost her kittens early on because no juveniles were seen on camera traps later on in the year. She had her litter during a particularly dry and hot summer; her natal den in an old stone barn under piles of wood was adjacent to a small ditch, which dried up. The area was also full of ticks, more so than other Priority Areas visited, so it may be that some combination of the lack of accessible water and heavy tick burdens might have resulted in the loss of her litter. What is of note, however, is that despite detecting her on a regular basis within the natal den using the radio tracking equipment and picking her up on camera traps located nearby, there were no visible signs that the area was being used at all. Campbell (2015) also noted the difficulties in identifying the natal den without using VHF equipment because of the lack of obvious signs. In that case, the natal den was found because the kittens were heard calling. Tomkies (1991) also states that wildcat natal dens are difficult to find because they tend to leave no obvious traces outside and that when the kittens are about two months old, the mother will use temporary natal dens near hunting grounds for a few days at a time while the kittens are learning to hunt, thus moving around regularly. Regular moving of kittens by female wild-living cats was noted during the Scottish Wildcat Action project and by Campbell (2015).
As a European protected species, protection of the wildcat in Scotland comes under the Conservation (Natural Habitats, &c.) Regulations 1994 (as amended). Part of this legislation includes the protection of natal dens and rest sites from disturbance. Currently, prior to any potential disturbance such as through forestry activities or developments, field surveys are undertaken to locate and identify potential rest sites and natal dens. However, the difficulty in identifying these sites as noted above, makes it difficult to protect them from disturbance, especially during the breeding season (see Stahl and Artois, 1995). For example, in Belgium, in one forest alone, three cases of litters being destroyed by the removal of log piles were recorded in 1989/1990 and this was identified as one of the main threats to wildcats in Belgium (Stahl and Artois, 1995).
More recently, Simon (2021) has made recommendations for improved wildcat conservation measures in forest management in Germany to reduce the impact of forest activities on breeding females between March – August including not removing log piles during this period, minimising the use of heavy machinery during this period, not clearing wind blow and layering deadwood piles away from paths to reduce disturbance. Many large-scale forestry operators such as FLS are aware of the need for extra care to be taken during this period in areas where wildcats exist. For example, during this study kittens were found in a logging stack due for removal in one of the Priority Areas and work was immediately suspended while the area was monitored by cameras.
In another site, harvesting was stopped during the breeding season due to the presence of wildcats. However, even small-scale forestry activities pose a potential risk to breeding wildcats as do large developments that involve extensive land clearance such as a road or housing or wind farm construction within areas where wildcats are present (see Stahl and Artois, 1995). More research needs to be carried out into helping identify wildcat rest sites and natal dens, such as through the use of scent dogs, which are currently used in parts of Germany before forest operations. In the meantime, given the large number of dens and resting sites used by individual cats, the most practical approach is to identify areas that are used by wildcats using camera traps set at an appropriate density and over an appropriate time-period; minimise activity in these high-risk areas during the breeding season; pre-emptively protect likely den structures when found; and promote awareness among forestry workers and land management staff. Revising the current guidelines for land development/management is therefore recommended.
5.3 Habitat use
Wildcat hybrids used certain habitats and landscape features significantly more than expected by chance, but this varied between individuals, Priority Areas and also between individuals within the same Priority Area. The landscape varies widely across Scotland, with many areas containing very low forest cover but having historical or current records of wildcat. Despite being previously considered a forest cat, wildcats are capable of utilising a variety of habitats. They show a preference for broadleaved or mixed forests (Parent, 1975, Schauenberg, 1981, Stahl and Leger, 1992, Sarmento et al., 2006), scrubland (Ragni, 1981, Lozano et al., 2003, Lozano, 2010), riparian areas and grassland for hunting (Wittmer, 2001, Corbett, 1979, Stahl et al., 1988, Biro et al., 2004) and edge habitats (Corbett, 1979, Daniels, 1997, Scott et al., 1993, Campbell, 2015). Observations from this study support those carried out on wildcats elsewhere, with wildcat hybrids showing a preference for cover habitats such as woodland, in particular broadleaved woodland, clear-fell and edge habitats.
Wildcat hybrids also showed a significant selection for arable landscapes. The farmland in the Priority Areas is generally low intensity with high densities of rabbits in parts of the Angus Glens and Strathbogie and is interspersed with small patches of woodland and hedgerows, allowing cats to make use of arable land while remaining close to cover. Recent studies have also highlighted the use of agriculturally dominated landscapes by wildcats in Germany where the farmland is also characterised by high structural diversity with a mixture of crops, hedgerows and pockets of woodland (Jerosch et al., 2017). Jerosch and Malte (2021) discuss the importance of long linear shelter habitats such as hedgerows and small patches of woodland as crucial in allowing wildcats to establish home ranges in open agricultural landscapes. We also note in this study, wildcat hybrids not only used the hedgerows for moving around but as mentioned above, for resting in. The use of features such as hedgerows/stone walls and small patches of woodland in otherwise open landscapes offers an insight into how habitat management could be applied for improving landscape connectivity for wildcats and also as a way to improve wildcat survey efforts and identification of wild-living cats for neutering by targeting camera trap surveys along these types of features.
Some of the wildcat hybrids in this study used low-medium traffic roads (B roads) to move up and down their home range and linear features such as hedgerows were followed to the point where they crossed busier A roads. Roads are used by many carnivores as a means to increase speed of movement around their home range. This is common for many meso-carnivores (e.g. Andersen et al., 2017, Frey and Conover, 2010) and has been postulated to improve foraging success by increasing search rates (e.g. Dickie et al., 2017). Camera trap footage also documented two individuals using culverts and small underpasses to go under roads instead of across them. This behaviour has also been noted by the European wildcat (Yanes et al., 1995). Road traffic accidents are a significant factor affecting mortality rates across Europe (Grilo et al., 2009, Riols, 1988, Lüttmann et al., 1997) and for reintroduced wildcats (Ruiz-Olmo and Mino, 1992, Hartmann, 2006) and in fact, have recently been identified as the main source of mortality for the European wildcat (Bastianelli et al., 2021).
Road traffic accidents are also a known mortality risk across Scotland. For example, when Balharry and Daniels (1998) collected samples, 147 cats (n=581) had been run over. Over the course of the SWA field work (five years), 61 road traffic accident cat carcasses were collected of which six were wildcat and 43 wildcat hybrids (Campbell et al., 2021a). Campbell et al. (2021) also found that males made up the majority of road casualties and that there was a significant peak in the number of cats being killed on roads in late summer/early autumn around the time of sub adult dispersal. This data combined with the use of roads by wildcat hybrids here highlights an area of concern for wildcat populations in Scotland. Initiatives to identify areas of potential high risk for wildcats in Scotland could be undertaken for existing roads and as a consideration for future road infrastructure through wildcat suitable habitat (e.g. identified through GPS data or road casualty data) and potential mitigation proposed. For example, in Germany Klar et al., (2009) found that installing roadside fencing designed to keep wildcats off the road reduced road-related wildcat mortality significantly and wildcats often used wildlife bridges and road culverts.
In addition to roads, of interest was the regular use of a railway and railway bridges to get across rivers and a busy A road in the Strathbogie Priority Area by two male wildcat hybrids. As well as providing a linear feature to follow, the railway passed through low intensity farmland and often followed a large river. Habitat on either side of the railway was characterised by scrub, patches of broadleaved woodland, hedgerows, gorse and unimproved grassland, all providing cover and prey for cats using the railway. Wildcats have anecdotally been reported to use railway lines in Scotland previously (Anonymous, pers comm, 2010), they are routinely cleared for snow in the winter months and in addition to railway banks providing prey and cover, railways may also provide an easier route for a cat to hunt across its home range during heavy snowfall. Studies in Europe and Scotland have shown that wildcats tend to either move to lower elevations or stay within covered habitat such as woodland during winter months (Corbett, 1979, Mermod and Liberek, 2002).
Despite being one of the most dominant landcover types in the different Priority Areas, the use of mature coniferous plantation varied depending on the Priority Area. Individuals in the Angus Glens Priority Area were significantly selected for this habitat type, whereas this was not the case for other areas in general, where broadleaved and/or mixed woodland was preferred. Only two cats in Strathbogie significantly selected coniferous woodland, Emma and Brook. In Emma’s case, most of her use of coniferous woodland was either in wind blow or using the forest tracks running through coniferous woodland between patches of clear-fell and in Brook’s, the coniferous woodland he was using was not mature.
Other studies have noted that wildcats do not spend much time in mature pine plantations (Corbett, 1979, Sarmento et al., 2006, Campbell, 2015, Scott et al., 1993, Daniels, 1997, Easterbee et al., 1991, Lozano et al., 2003), particularly if there is no shrub layer underneath (Puzachenko, 1992) with monospecific plantations of conifers (or even broadleaved) resulting in a drastic reduction in shelter and hunting grounds for wildcats (Stahl and Artois, 1995). Snow tracking of wildcats in the Polish Carpathian Mountains showed that wildcats rarely moved through old even-aged woodland, especially coniferous plantations, but would often move through younger deciduous or mixed forest stands with dense undergrowth (Okarma et al., 2002). Indeed, Stahl and Artois (1995) noted that the creation of a patchwork of stands of differing ages, including dense regeneration areas, would be beneficial for wildcats. Simon (2021) also notes the importance of developing a forestry practice that encourages wildcat habitat including non-clearing of wind blow (an important habitat for breeding and prey as noted above) and allowing natural reforestation in some areas. Here we observed that cats using timber plantations tended to use areas of clear-fell, patches <20 years old or young plantation and edges of mature coniferous stands, particularly edges alongside forest tracks which corresponds to observations by Campbell (2015). All collared individuals here (except Fairburn) selected edge habitats, so increasing the amount of ecotone habitat within coniferous plantations could also aid wildcat habitat creation.
5.4 Conclusions
The results from this study indicate that wildcat hybrids in Scotland are behaving in a similar manner to European wildcats in mainland Europe. As a result, wildcat hybrids in Scotland appear to be fulfilling the same ecological niche as a wildcat would and these data provide us with invaluable information on how wild-living cats, and potentially wildcats and re-introduced wildcats, might use the landscape, as well as highlighting some potential threats for wildcats in Scotland.
- This data and other studies from mainland Europe all highlight the difficulties in identifying and locating the many different rest sites and natal den sites that wildcats and wildcat hybrids appear to be using. It is evident from this study that protecting some habitat features where wildcats are present could be more effective and easier to implement in many cases than trying to identify every single site. Based on evidence from European studies, improved forest management could also reduce the impact of forest activities during the kitten rearing period (March – August) and changes to other landscape management practices such as the extent and timing of gorse/shrub clearance in areas that wildcats are present. We recommend that the current guidance for developers, foresters and land managers is re-evaluated with the existing data in mind and in the meantime further research should be carried out on rest site and natal den characteristics.
- Based on the comparatively large home range sizes exhibited here, the GPS data also indicates that parts of Scotland are potentially sub-optimal habitat for wildcats unless steps can be taken to improve prey densities, connectivity and cover through careful land management. Such changes to land management would likely benefit other species thus providing wider biodiversity gains. There are many initiatives being proposed across Scotland, including increased planting of native woodland, that may be beneficial, particularly if they are managed in a way to create complexity both internally (i.e. a structurally diverse understorey) and at the landscape scale (i.e. avoiding single species and age monocultures). Improvements to landscape connectivity could be made through planting of hedgerows and small patches of woodland / scrub and increasing the amount of edge habitat within plantations. We would emphasise the apparent importance of gorse as a rest site and recommend that future initiatives to increase native woodland cover in Scotland both ensure a sufficient level of vertical stratification and do not occur at the expense of gorse, particularly adjacent to rabbit populations. As noted, wind-blow appears to be particularly important for wildcats across their range and should be retained as much as possible.
- The use of farm barns by many of the collared individuals highlights a route for continued hybridisation in Scotland and we recommend that efforts are continued to neuter farm cats to reduce this threat.
- The use of heathland by some of the individuals and the known historic use of this habitat type by wildcats indicates the importance of continuing to improve wildlife conservation initiatives across grouse moorlands.
- Roads are a major cause of mortality for wildcats and the use of roads by wildcat hybrids here highlights an area of concern for wildcat populations in Scotland. Initiatives to identify areas of potential high risk for wildcats in Scotland should be undertaken to try and reduce road-related wildcat mortality.
6. References
Adam, A. M. A., Kropil, R. & Pichler, V. 2015. Differences in evaluation of three different approaches in home range sizes of red deer Cervus elaphus in Western Carpathians. Central European Forestry Journal, 61, 12-18.
Aebischer, N. J., Davey, P. D. & Kingdon, N. G. 2011. National Gamebag Census: Mammal Trends to 2009 [Accessed].
Andersen, G. E., Johnson, C. N., Barmuta, L. A. & Jones, M. E. 2017. Use of anthropogenic linear features by two medium-sized carnivores in reserved and agricultural landscapes. Scientific Reports, 7, 11624.
Anile, S., Bizzarri, L., Lacrimini, M., Sforzi, A., Ragni, B. & Devillard, S. 2017. Home-range size of the European wildcat (Felis silvestris silvestris): a report from two areas in Central Italy. Mammalia, 82.
Anile, S., Devillard, S., Ragni, B., Rovero, F., Mattucci, F. & Lo Valvo, M. 2019. Habitat fragmentation and anthropogenic factors affect wildcat Felis silvestris silvestris occupancy and detectability on Mt Etna. Wildlife Biology, 1, 1-13.
Bacon, A., Beckmann, K. M., Anderson, N. E., Alves, B. S. G., Ogden, R. & Meredith, A. L. 2023. Scottish Wildcat Action Final Report: Disease Surveillence. NatureScot, Inverness.
Balharry, D. & Daniels, M. J. 1998. Wild living cats in Scotland. Scottish Natural Heritage Research, Survey and Monitoring Report No. 23. Edinburgh, Scotland
Balharry, E., Staines, B. W., Marquiss, M. & Kruuk, H. 1994. Hybridisation in British Mammals. JNCC Report No. 154. Joint Nature Conservation Committee, Peterborough.
Ballesteros-Duperon, E., Barea-Azcon, J. M., Moleon, M., Virgos, E., Gil-Sanchez, J. M. & Chirosa, M. Preliminary results on habitat preferences of a wildcat (Felis silvestris Schreber, 1777) population in an area of south of Spain: influence of rabbit distribution and abundance. In: Herrmann, M., ed. Symposium: Biology and Conservation of the European Wildcat (Felis silvestris silvestris), 2005 Germany. January 21st –23rd 2005, 24 - 25.
Barton, K. A. 2019. Mu-MIn: Multi-model inference. R Package Version 0.12.2/r18 [Accessed].
Bastianelli, M. L., Premier, J., Herrmann, M., Anile, S., Monterroso, P., Kuemmerle, T., Dormann, C. F., Streif, S., Jerosch, S., Gotz, M., Simom, O., Moleon, M., Gil-Sanchez, J. M., Biro, Z., Dekker, J., Severon, A., Krannich, A., Hupe, K., Germain, E., Pontier, D., Janssen, R., Ferreras, P., Diaz-Ruiz, F., Lopez-Martin, J. M., Urra, F., BizzarriI, L., Bertros-Martin, E., Dietz, M., Trinzen, M., Ballesteros-Duperon, E., Barea-Azcon, J. M., Sforzi, A., Poulle, M.-L. & Heurich, M. 2021. Survival and cause-specific mortality of European wildcat (Felis silvestris) across Europe. Biological Conservation, 261, 109239.
Bates, D., Machler, M., Bolker, B. & WALKER, S. 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software, 67, 1-48.
Beaumont, M., Barratt, E. M., Gottelli, D., Kitchener, A. C., Daniels, M. J., Pritchard, J. K. & Bruford, M. W. 2001. Genetic diversity and introgression in the Scottish wildcat. Molecular Ecology, 10, 319-336.
Benson, J. F., Chamberlain, M. J. & Leopold, B. D. 2006. Regulation of space use in a solitary felid: Population density or prey availability? Animal Behaviour, 71, 685-693.
Beugin, M., Leblanc, G., Queney, G., Natoli, E. & Pontier, D. 2016. Female in the inside, male in the outside: insights into the spatial organization of a European wildcat population. Conservation Genetics, 17, 1405-1415.
Bewick, T. 1820. A general history of British quadrupeds, John Van Voorst, London.
Biro, Z., Szemethy, L. & Heltai, M. 2004. Home range size of wildcats (Felis sylvestris) and feral domestic cats (Felis sylvestris f. catus) in a hilly region of Hungary. Mammalian Biology, 69, 302-310.
Burgman, M. A. & Fox, J. C. 2003. Bias in species range estimates from minimum convex polygons: implications for conservation and options for improved planning. Animal Conservation, 6, 19-28.
Calenge, C. 2011. Home range estimation in R: the adehabitatHR package. Office national de la classe et de la faune sauvage, Saint Benoist, Auffargis, France.
Campbell, R. D. 2015. Spatial ecology of the Scottish wildcat. Report to PTES. Wildlife Conservation Research Unit, Oxford, U.K. (2015)
Campbell, R. D., Langridge, K., Kilshaw, K., H., C., Hislop, C., Rawling, E. & Tallach, N. 2023a. Scottish Wildcat Action final report: Monitoring and Surveys. NatureScot, Inverness.
Campbell, R. D., Rawling, E., Langridge, K. V. & Tallach, N. 2023b. Scottish Wildcat Action final report: Trap Neuter Vaccinate Return Programme. NatureScot, Inverness.
Cocks, A. H. 1876. Wild cats: period of gestation. The Zoologist, 5038-5039.
Conde, B. & Schauenberg, P. 1974. Reproduction du Chat forestier (f. silvestris Schr.) dans le nord-est de la France. Revue suisse de zoologie., 81, 45-52.
Corbett, L. K. 1979. Feeding ecology and social organization of wild cats (Felis silvestris) and domestic cats (Felis catus) in Scotland. PhD, University of Aberdeen, PhD Thesis, 334 pp.
Crandall, R., Crandall, C., Nayar, J. & Doyle, T. 1994. Resistance and disease in Brugia malayi infection of ferrets following prior infection, injection of attenuated infective larvae and injections of larval extracts. Parasite Immunology, 16, 425-433.
Dahle, B., Støen, O. G. & Swenson, J. E. 2006. Factors influencing home-range size in subadult brown bears. Journal of Mammalogy, 87, 859-865.
Daniels, M. J. 1997. The biology and conservation of the wildcat in Scotland. PhD, University of Oxford.
Daniels, M. J., Balharry, D., Hirst, D., Aspinall, R. J. & Kitchener, A. C. 1998. Morphological and pelage characteristics of wild living cats in Scotland: Implications for defining the 'wildcat'. Journal of Zoology, 244, 231-247.
Daniels, M. J., Beaumont, M. A., Johnson, P. J., Balharry, D., MacDonald, D. W. & Barratt, E. 2001. Ecology and genetics of wild-living cats in the north-east of Scotland and the implications for the conservation of the wildcat. Journal of Applied Ecology, 38, 146-161.
Darwin, C. 1875. Variation of animals and plants under domestication (2nd Edition), New York: D. Appleton & Co.
Davies, A. R. & Gray, D. 2010. The distribution of Scottish wildcats (Felis silvestris) in Scotland (2006-2008). Scottish Natural Heritage Commissioned Report No. 360.
Delahay, R. J., Daniels, M. J., MacDonald, D. W., McGuire, K. & Balharry, D. 1998. Do patterns of helminth parasitism differ between groups of wild-living cats in Scotland? Journal of the Zoological Society of London, 245, 175-183.
Dickie, M., Serrouya, R., McNay, R. S. & Boutin, S. 2017. Faster and farther: wolf movement on linear features and implications for hunting behaviour. Journal of Applied Ecology, 54, 253-263.
Dimitrijevic, S. & Habijan, V. 1977. Spreading and numbers of the wild cat Felis silvestris Schreber, 1777 in Vojvodina. Arhiv Bioloskih Nauka (Arh.biol.nauka), Beograd 29, 175-178.
Dodge, S., Xavier, G. & Wong, W. Y. 2018. DynamoVis - Dynamic Visualization of Animal Movement Data. Retrieved from the Data Repository for the University of Minnesota, [Online]. [Accessed].
Dotterer, M. & Bernhart, F. 1996. The occurrence of wildcats in the southern Swiss Jura Mountains. Acta Theriologica, 41, 205-209.
Doyle, A. T. 1990. Use of riparian and upland habitats by small mammals. Journal of Mammalogy, 71, 14-23.
Driscoll, C. A., Menotti-Raymond, M., Roca, A. L., Hupe, K., Johnson, W. E., Geffen, E., Harley, E. H., Delibes, M., Pontier, D., Kitchener, A. C., Yamaguchi, N., O'Brien, S. J. & MacDonald, D. W. 2007. The Near Eastern Origin of Cat Domestication. Science, 317, 519-523.
Easterbee, N., Hepburn, L. V. & Jefferies, D. J. 1991. Survey of the status and distribution of the wildcat in Scotland, 1983-1987. Edinburgh: Nature Conservancy Council for Scotland.
Ecke, F., Lofgren, O., Hornfeldt, B., Eklund, U., Ericsson, P. & Sorlin, D. 2001. Abundance and Diversity of Small Mammals in Relation to Structural Habitat Factors. Ecological Bulletins, 49, 49, 165-171.
EUNIS 2020. EUNIS habitat data for Scotland.
Evans, D. M., Redpath, S. M., Elston, D. A., Evans, S. A., Mitchell, R. J. & Dennis, P. 2006. To graze or not to graze? Sheep, voles, forestry and nature conservation in the British uplands. Journal of Applied Ecology 43, 499-505.
Fitzgibbon, C. D. 1997. Small Mammals in Farm Woodlands: The Effects of Habitat, Isolation and Surrounding Land-Use Patterns. Journal of Applied Ecology, 34, 530-539.
Flowerdew, J. R. & Ellwood, S. A. 2001. Impacts of woodland deer on small mammal ecology. Forestry, 74, 277-287.
Frey, S. N. & Conover, M. R. 2010. Habitat Use by Meso‐Predators in a Corridor Environment. The Journal of Wildlife Management, 70, 1111-1118.
Germain, E., Benhamou, S. & Poulle, M.-L. 2008. Spatio-temporal sharing between the European wildcat, the domestic cat and their hybrids. Journal of Zoology, 276, 195-203.
Gomez, D. M. & Anthony, R. G. 1998. Small mammal abundance in riparian and upland areas of five seral stages in western Oregon. Northwest Science, 72, 293-302.
Gray, A. 1971. Mammalian hybrids: a checklist with bibliography. Commonwealth Bureau of Animal Breeding Genetics Technical communication 10.
Grilo, C., Bissonette, J. A. & Santos-Reisa, M. 2009. Spatial–temporal patterns in Mediterranean carnivore road casualties: Consequences for mitigation. Biological Conservation, 142, 301-313.
Halliez, G., Renault, F., Vannard, E., Farny, G., Lavorele, S. & Giraudoux, P. 2015. Historical agricultural changes and the expansion of a water vole population in an Alpine valley. Agriculture, Ecosystems and Environment, 198-206.
Hamilton, E. 1897. The wildcat of Scotland. The Annals of Scottish Natural History, 6, 65-78.
Hartmann, M. 2006. Reintroduction of captive bred wildcats in Germany. Iberian Lynx Ex-situ Conservation reintroduction project.
Heptner, V. H. & Sludskii, A. A. 1972. [Mammals of the Soviet Union. Vol III: Carnivores (Feloidea).] Vyssha Shkola, Moscow (in Russian). Smithsonian Institute and the National Science Foundation, Washington DC.
Herfindal, I., Nilsenn, E. B., Andersen, R., Linnell, J. D. C. & Odden, J. 2005. Prey density, environmental productivity and home-range size in the Eurasian lynx (Lynx lynx). Journal of Zoology, 265, 63-71.
Hertwig, S., Schweizer, M., Stepanow, S., Jungnickel, A., Bohle, U. & Fischer, M. 2009. Regionally high rates of hybridization and introgression in German wildcat populations (Felis silvestris, Carnivora, Felidae). Journal of Zoological Systematics and Evolutionary Research, 47, 283-297
Hewson, R. 1983. The food of wild cats (Felis silvestris) and red foxes (Vulpes vulpes) in west and north-east Scotland. Journal of the Zoological Society of London, 200, 283-289.
Hubbard, A. L., McOrist, S., Jones, T. W., Boid, R., Scott, R. & Easterbee, N. 1992. Is survival of European wildcats Felis silvestris in Britain threatened by interbreeding with domestic cats? Biological Conservation, 61, 203-208.
Jacobs, J. 1974. Quantitative measurement of food selection:a modification of the forage ratio and Ivlev’s Electivity Index. Oecologia, 14, 413-417.
Jenkins, D. 1962. The present status of the wild cat (Felis silvestris) in Scotland. Scottish Naturalist, 70, 126-138.
Jerosch, S., Gotz, M. & Roth, M. 2017. Spatial organisation of European wildcats (Felis silvestris silvestris) in an agriculturally dominated landscape in Central Europe. Mammalian Biology, 82, 8-16.
Kenward, R. E. 2001. A Manual for Wildlife Radio Tagging, London, Academic Press.
Kilshaw, K. 2015. Introgression and the current status of the Scottish wildcat. PhD, University of Oxford.
Kitchener, A. C. 1995. The Wildcat, The Mammal Society.
Kitchener, A. C. & Easterbee, N. 1992. The taxonomic status of black wild felids in Scotland. Journal of Zoology, 277, 342-346.
Kitchener, A. C. & Senn, H. 2023. Scottish Wildcat Action final report; wildcat genetics and taxonomy. NatureScot, Inverness.
Kitchener, A. C., Yamaguchi, N., Ward, J. M. & MacDonald, D. W. 2005. A diagnosis for the Scottish wildcat (Felis silvestris): A tool for conservation action for a critically-endangered felid. Animal Conservation, 8, 223-237.
Klar, N., Fernandez, N., Kramer-Schadt, S., Herrmanne, M., Trinzenf, M., Buttnerf, I. & Niemitz, C. 2008. Habitat selection models for European wildcat conservation. Biological Conservation, 141, 308-319.
Klar, N., Hermann, M. & Kramer-Schadt, S. 2009. Effects and Mitigation of Road Impacts on Individual Movement Behavior of Wildcats. The Journal of Wildlife Management, 73, 631–638.
Kuznetsova, A., Brockhoff, P. B. & Christensen, R. H. B. 2017. lmerTest Package: Tests in Linear Mixed Effects Models. Journal of Statistical Software, 82, 1-26.
Langley, P. J. W. & Yalden, D. W. 1977. The decline of the rarer carnivores in Great Britain during the nineteenth century. Mammal Review, 18, 741-760.
Liberek, M. 2002. Distribution, home range, activity periods and habitat use of four wildcats (Felis s. silvestris) in Swiss Jura mountains: first results. Säugetierk. Inform.d. Naturschutz Niedersachs, 5, 233-238.
Lindemann, W. 1953. Einiges über die Wildkatze der Ostkarpaten (Felis s. silvestris Schreber, 1777). [Some remarks on wildcats in the East Carpathian mountains]. Säugetierk. Mitt., 1, 73-74.
Litvaitis, J. A., Sherburne, J. A. & Bissonette, J. A. 1986. Bobcat habitat use and home range size in relation to prey density. Journal of Wildlife Management, 50, 110-117.
Lovegrove, R. 2007. Silent Fields; The Long Decline of a Nation's Wildlife, Oxford University Press.
Lozan, M. N. & Korcmar, N. D. 1965. Materialy po ekologii nekotorych chriscnych zverej v plavnijack Pruta.](in Russian). Ochrana prirody Moldavii, 3, 140-149.
Lozano, J. 2010. Habitat use by European wildcats (Felis silvestris) in central Spain: what is the relative importance of forest variables? Animal Biodiversity and Conservation, 33, 143-150.
Lozano, J., Moleon, M. & Virgos, E. 2006. Biogeographical patterns in the diet of the wildcat, Felis silvestris Schreber, in Eurasia: Factors affecting the trophic diversity. Journal of Biogeography, 33, 1076-1085.
Lozano, J., Virgos, E., Cabezas-Diaz, S. & Mangas, J. G. 2007. Increase of large game species in Mediterranean areas: Is the European wildcat (Felis silvestris) facing a new threat? Biological Conservation, 138, 321-329.
Lozano, J., Virgos, E., Malo, A. F., Huertas, D. L. & Casanovas, J. G. 2003. Importance of scrub-pastureland mosaics for wild-living cats occurrence in a Mediterranean area: Implications for the conservation of the wildcat (Felis silvestris). Biodiversity and Conservation, 12, 921-935.
Ludecke, D. 2018. ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. Journal of Open Source Software, 3, 772.
Luttmann, J., Hochhardt, W. & Hoßfeld, E. 1997. Entwicklung eines für Wildkatzen unüberwindbaren Wildschutzzaunes an Straßen auf Basis von Verhaltensstudien als Beitrag zum Artenschutz bei Säugetieren in German (Development of a Protective Fence for Wildcats (Felis silvestris silvestris) at Roads based on Behaviour Studies. A contribution to the conservation of species. Nature + Landscape (mimeograph).
MacDonald, D. W., Daniels, M. J., Driscoll, C., Kitchener, A. & Yamaguchi, N. 2004. The Scottish Wildcat: Analyses for Conservation and an Action Plan: Wildlife Conservation Research Unit, University of Oxford.
Mathews, F., Kubasiewicz, L. M., Gurnell, J., Harrower, C. A., McDonald, R. A. & Shore, R. F. 2018. A Review of the Population and Conservation Status of British Mammals: Technical Summary. A report by the Mammal Society under contract to Natural England, Natural Resources Wales and Scottish Natural Heritage. Natural England, Peterborough. .
Matthews, H. L. 1941. Reproduction of the Scottish wild cat, Felis silvestris grampia Miller. Proceedings of the Zoological Society of London, Series B, 111, 59-77.
McEwing, R., Kitchener, A. C., Holleley, C., Kilshaw, K. & O'Donoghue, P. 2011. An allelic discrimination SNP assay for distinguishing the mitochondrial lineages of European wildcats and domestic cats. Conservation Genetic Resources, Published online DOI 10.1007/s12686-011-9499-z.
McNab, B. 1983. Energetics, body size, and the limits to endothermy. Journal of Zoology, 199, 1-29.
McOrist, S. & Kitchener, A. C. 1994. Current Threats to the European Wildcat, Felis-Silvestris, in Scotland. Ambio, 23, 243-245.
Mermod, C. P. & Liberek, M. 2002. The role of snow cover for European wildcat in Switzerland. Zeitschrift fur Jagdwissenschaft, 48, 17-24.
Michal, B. & Rafal, Z. 2014. Responses of small mammals to clear-cutting in temperate and boreal forests of Europe: a meta-analysis and review. European Journal of Forest Research, 133, 1-11.
Migli, D., Astaras, C., Boutsis, G., Diakou, A., Karantanis, N.-E. & Youlatos, D. 2021. Spatial Ecology and Diel Activity of European Wildcat (Felis silvestris) in a Protected Lowland Area in Northern Greece. Animals, 11, 3030.
Monterroso, P., Brito, J. C., Ferreras, P. & Alves, P. C. 2009. Spatial ecology of the European wildcat in a Mediterranean ecosystem: dealing with small radio-tracking datasets in species conservation. Journal of Zoology, 279, 27-35.
Muntyanu, A. I., Vasiliev, A. G. & Chegorka, P. T. Distribution and number of the European wildcat (Felis silvestris Schreber, 1777) in R. Moldova. Seminar on the biology and conservation of the wildcat (Felis silvestris), 1992 Nancy, France, 23-25 September 1992. Council of Europe, Strasbourg, 59-61.
Nowell, K., & Jackson, P. 1996. Wild cats, Status survey and conservation action plan, Gland, Suiza, IUCN.
Nowell, K. & Jackson, P. (eds.) 1996. Status Survey and Conservation Action Plan: Wild Cats, Gland, Swtzerland: IUCN.
O'Brien, J., Devillard, S., Say, L., Vanthomme, H., Leger, F., Ruette, S. & Pontier, D. 2009. Preserving genetic integrity in a hybridising world: are European Wildcats (Felis silvestris silvestris) in eastern France distinct from sympatric feral domestic cats? Biodiversity and Conservation, 18, 2351–2360.
Oliveira, T., Urra, F., Lopez-Martin, J. M., Ballesteros-Duperon, E., Barea-Azcon, J. M., Moleon, M., Gil-Sanchez, J. M., Alves, P. C., Diaz-Ruiz, F., Ferreras, P. & Monterroso, P. 2018a. Females know better: Sex‐biased habitat selection by the European wildcat. Ecology and Evolution, Published online 29 August 2018.
Oliveira, T., Urra, F., Lopez-Martin, J. M., Ballesteros-Duperon, E., Barea-Azcon, J. M., Moleon, M., Maria, J., Gil-Sanchez, J. M., Alves, P. C., Diaz-Ruiz, F., Ferreras, P. & Monterroso, P. 2018b. Females know better: Sex‐biased habitat selection by the European wildcat. Ecology and Evolution, 8, 9464-9477.
Parent, G. H. 1975. La migration récente à caractère invasionnel du chat sauvage Felis silvestris Schreber en Lorraine belge. [Recent colonization of the wildcat Felis silvestris silvestris Schreber in Belgian Lorraine.] (in French). Mammalia, 39, 251-288
Peterson, B. G., Carl, P., Boudt, K., Bennett, R., Ulrich, J., Zivot, E., Cornilly, D., Hung, E., Lestel, M., Balkissoon, K., Wuertz, D., Christidis, A. A., Martin, R. D., Zhou, Z. & Shea, J. M. 2020. PerformanceAnalytics: Econometric Tools for Performance and Risk Analysis (version 2.0.4).
Pierpaoli, M., Randi, E., Ragni, B., Bir, Z. S., Szemethy, L., Herrmann, M., Hupe, K. & Fernandes, M. 2003. Genetic distinction of wildcat (Felis silvestris) populations in Europe, and hybridization with domestic cats in Hungary. Molecular Ecology, 12, 2585-2598.
Pineiro, A., Barja, I., Silvan, G. & Illera, J. C. 2012. Effects of tourist pressure and reproduction on physiological stress response in wildcats: management implications for species conservation. Wildlife Research, 39, 532-539.
Pitt, F. 1939. Wild Animals in Britain, B.T. Batsford, London.
Popovic, R., Sver, L., Grguric, Z. & Guzvica, G. 2017. Analysis of the wildcat’s (Felis sivestris silvestris L.) preference to the habitat types (by NCH) in the Bilogora mountain, Croatia.
Puzachenko, A. Y. The European wildcat (Felis silvestris) in Armenia, Azerbaijan, Belarus, Georgia, Moldova, Russia and Ukraine. Seminar on the biology and conservation of the wildcat (Felis silvestris), 1992 Nancy, France, 23-25 September 1992. Council of Europe, Strasbourg, 64-71.
Ragni, B. P. I. 1981. Gatto selvatico, Felis silvestris Schreber, 1977 [Forest wildcat, Felis silvestris Schreber, 1977] (In Italian). Distribuzione e biologia di 22 specie di mammiferi in Italia [Distribution and biology of 22 mammal species in Italy](in Italian). C.N.R. Rome.
Randi, E., Pierpaoli, M., Beaumont, M., Ragni, B. & Sforzi, A. 2001. Genetic identification of wild and domestic cats (Felis silvestris) and their hybrids using bayesian clustering methods. Molecular Biology and Evolution, 18, 1679-1693.
Riols, C. 1988. Etude du régime alimentaire du chat forestier (Felis silvestris) dans l’Est de la France [Study of the diet of the forest wildcat (Felis silvestris) in eastern France.] (in French). . Bull. Mens. O.N.C., 121, 22-27.
Ruiz-Olmo, J. R. & Mino, A. Reintroduction of wildcats in Catalonia (Spain). Seminar on the biology and conservation of the wildcat (Felis silvestris), 1992 Nancy, France, 23-25 September 1992. Council of Europe, 111 - 114.
Sarmento, P., Cruz, J., Tarroso, P. & Fonseca, C. 2006. Space and habitat selection by female European wild cats (Felis silvestris silvestris). Wildlife Biology in Practice, 2, 79-89.
Say, L., Devillard, S., Leger, F., Pontier, D. & Ruette, R. 2012. Distribution and spatial genetic structure of European wildcat in France. Animal Conservation, 15, 18-27.
Schauenberg, P. 1981. Elements d'ecologie du Chat forestier d'Europe (Felis sylvestris Schreber, 1777) [Elements of ecology of the wild cat of Europe] (in French). Revue D Ecologie-La Terre Et La Vie, 35, 3-36.
Schmidt, K. 2008. Behavioural and spatial adaptation of the Eurasian lynx to a decline in prey availability. Acta Theriologica, 53, 1-16.
Scott, R., Easterbee, N. & Jefferies, D. A radio-tracking study of wildcats in western Scotland. Proc. seminar on the biology and conservation of the wildcat (Felis silvestris), 1993 Nancy, France. Council of Europe, Strasbourg, 94-97.
Senn, H. V., Ghazali, M., Kaden, J., Barclay, D., Harrower, B., Campbell, R. D., MacDonald, D. W. & Kitchener, A. C. 2019. Distinguishing the victim from the threat: SNP‐based methods reveal the extent of introgressive hybridization between wildcats and domestic cats in Scotland and inform future in situ and ex situ management options for species restoration. Evolutionary Applications, 399-414.
Senn, H. V. & Ogden, R. 2015. Wildcat Hybrid Scoring For Conservation Breeding under the Scottish Wildcat Conservation Action Plan. Royal Zoological Society of Scotland.
Simon, O. Wildcat conservation measures in forest management and consideration of species in impact assessment procedures. In a good way? The current situation of the Wildcat in Germany and Europe, 2021 Germany. Deutsche Wildtier Stiftung, 148-152.
Simon, O., Dietz, M., Gotz, M., Herrmann, M., Klenk, B., Krannich, A., Neumann, G. & Trinzen, M. Impacts on the European wildcat of anthropogenic interventions in forest habitats, with particular reference to wind turbines. Initial findings of a research project by the German Wildlife Foundation. In a good way? The current situation of the Wildcat in Germany and Europe, 2021 Germany. Deutsche Wildtier Stiftung.
Smit, R., Bokdam, J., Den Ouden, J., Olff, H., Schot-Opschoor, H. & Schrijvers, M. 2001. Effects of introduction and exclusion of large herbivores on small rodent communities. Plant Ecology, 155, 119-127.
Stahl, P. & Artois, M. 1995. Status and Conservation of the wildcat (Felis silvestris) in Europe and around the Mediterranean rim. Nature and Environment, No. 69. Council of Europe.
Stahl, P., Artois, M. & Aubert, M. F. A. 1988. Organisation spatiale et déplacements des chat forestiers adultes (Felis silvestris, Schreber, 1777 en Lorraine. [Spatial organization and displacement of adult forest wildcats (Felis silvestris Schreber, 1777) in Lorraine.] (in French). Terre et Vie 43, 113-132.
Stahl, P. & Leger, F. 1992. Le chat sauvage (Felis silvestris, Schreber, 1777). In: Artois, M. & Maurin, H. (eds.) Encyclopédie des Carnivores de France. Société Française pour l’Etude et la Protection des Mammifères (S.F.E.P.M.), Bohallard, Puceul, France.
Suchomel, J., Purchart, L. & Čepelka, L. 2012. Structure and diversity of small-mammal communities of lowland forests in the rural central European landscape. European Journal of Forest Research, 131, 1933-1941.
Sumption, K. J. & Flowerdew, J. R. 1985. The ecological effects of the decline in Rabbits (Oryctolagus cuniculus L.) due to myxomatosis. Mammal Review, 15, 151-186.
Takahata, C., Nielsen, S. E., Takii, A., Izumiyama, S. & • 2014. Habitat Selection of a Large Carnivore along Human-Wildlife Boundaries in a Highly Modified Landscape. PLOS ONE.
Tapper, S. 1992. Game Heritage, The Game Conservancy: Fordingbridge.
Thomas, J. P., Larter, N. C. & Jung, T. S. 2021. Individual-based seasonal habitat selection in a forest-dwelling population of reintroduced bison <i>Bison bison</i>. Wildlife Biology, 2021.
Tiesmeyer, A., Ramos, L., Manuel, L., J. , Steyer, K., Alves, P. C., Astaras, C., Brix, M., Cragnolini, M., Domokos, C., Hegyeli, Z., Janssen, R., Kitchener, A. C., Lambinet, C., Mestdagh, X., Migli, D., Monterroso, P., Mulder, J. P., Schockert, V., Youlatos, D., Pfenninger, M. & Nowak, C. 2020. Range-wide patterns of human-mediated hybridisation in European wildcats. Conservation Genetics, 21, 247–260.
Tomkies, M. 1991. Wildcats, London: Whittet Books.
Wei, T. & Simko, V. 2017. R package "corrplot": Visualization of a Correlation Matrix (version 0.84).
Wheeler, P. 2008. Effects of sheep grazing on abundance and predators of field vole (Microtus agrestis) in upland Britain. Agriculture Ecosystems and Environment, 123, 49-55.
White, F. 2019. Drivers of domestic cat movement in Scotland. MSc.
Wikelski, M., Davidson, S. & Kays, R. Movebank: Archive, Analysis and Sharing of Animal Movement Data (accessed 11 November 2021) [Online]. [Accessed].
Wilson, R. P., White, C. R., Quintana, F., Halsey, L. G., Liebsch, N., Martin, G. R. & Butler, P. J. 2006. Moving towards acceleration for estimates of activity-specific metabolic rate in free-living animals: The case of the cormorant. Journal of Animal Ecology, 75, 1081-1090.
Wittmer, H. U. 2001. Home range size, movements, and habitat utilization of three male European wildcats (Felis silvestris Schreber, 1777) in Saarland and Rheinland-Pfalz (Germany). Mammalian Biology, 66, 365-370.
Yamaguchi, N., Driscoll, C. A., MacDonald, D. W., Kitchener, A. C. & Ward, J. M. 2004. Craniological differentiation between European wildcats (Felis silvestris silvestris), African wildcats (F. s. lybica) and Asian wildcats (F. s. ornata): Implications for their evolution and conservation. Biological Journal of the Linnean Society, 83, 47-63.
Yamaguchi, N., Kitchener, A., Driscoll, C. & Nussberger, B. 2015. Felis silvestris. The IUCN Red List of Threatened Species 2015: e.T60354712A50652361 [Online]. [Accessed 2021].
Yanes, M., Velasco, J. M. & Suarez, F. 1995. Permeability of roads and railways to vertebrates the importance of culverts. Biological Conservation, 71, 217-222.
7. ANNEX 1
| m1 <- lmer(I(log(mcp+1))~month + I(month^2) + I(month^3) + sex + sex*month+(1|cat ID) |
|---|
| m2 <- lmer(I(log(mcp+1))~month + I(month^2) + I(month^3) + sex + (1|cat ID) |
| m3 <- lmer(I(log(mcp+1))~month + I(month^2) + sex + sex*month + (1|cat ID) |
| m4 <- lmer(I(log(mcp+1))~month + I(mon^2) + sex + (1|cat ID) |
| m5 <- lmer(I(log(mcp+1))~month + sex + sex*month + (1|cat ID) |
| m6 <- lmer(I(log(mcp+1))~month + sex +(1|cat ID) |
| m7 <- lmer(I(log(mcp+1))~month + (1|cat ID) |
| m8 <- lmer(I(log(mcp+1))~sex + (1|cat ID) |
| m9 <- lmer(I(log(mcp+1))~(1|cat ID) |
RSF model | (Int) | month | month^2 | month^3 | sex | month*sex | df | logLik | AICc | delta | weight |
|---|---|---|---|---|---|---|---|---|---|---|---|
| m5 | 2.385 | -0.14 | - | - | Yes | Yes | 6 | -95.703 | 204.2 | 0 | 0.564 |
| m6 | 2.077 | -0.08604 | - | - | Yes | - | 5 | -97.271 | 205.1 | 0.91 | 0.357 |
| m7 | 2.587 | -0.08866 | - | - | - | - | 4 | -99.997 | 208.4 | 4.18 | 0.07 |
| m3 | 2.192 | -0.05832 | -0.0068 | - | Yes | Yes | 7 | -99.22 | 213.5 | 9.3 | 0.005 |
| m4 | 1.888 | -0.00631 | -0.00662 | - | Yes | - | 6 | -100.841 | 214.4 | 10.27 | 0.003 |
| m8 | 1.585 | - | - | - | Yes | - | 4 | -105 | 218.4 | 14.19 | 0 |
| m1 | 1.591 | 0.3916 | -0.09225 | 0.004581 | Yes | Yes | 8 | -101.244 | 219.8 | 15.65 | 0 |
| m2 | 1.356 | 0.3769 | -0.07852 | 0.003857 | Yes | - | 7 | -103.952 | 222.9 | 18.76 | 0 |
| m9 | 2.124 | - | - | - | - | - | 3 | -108.433 | 223.1 | 18.91 | 0 |
For further information on this report please contact: Dr Kerry Kilshaw
Address: Wildlife Conservation Research Unit, Department of Zoology, University of Oxford, Recanati-Kaplan Centre, Tubney House, Abingdon Road, Tubney, Oxfordshire, OX13 5QL
Telephone: +44 1865 611 100
Email: [email protected]
ISBN: 978-1-78391-978-9
This publication is part of a series of specialist reports on the work of the Scottish Wildcat Action (SWA) project that ran from 2015-2020. The work was led and steered by a partnership of organisations: Cairngorms National Park Authority, Forestry and Land Scotland, National Museums Scotland, Scottish Land & Estates, NatureScot, Scottish Wildlife Trust, The National Trust for Scotland, The Royal (Dick) School of Veterinary Studies (R(D)SVS), The Royal Zoological Society of Scotland, The Scottish Gamekeepers Association, The Scottish Government, Wildlife Conservation Research Unit (WildCRU).
The many other partners and funders are listed in the SWA Summary report.
For more information, including access to the other reports, contact Martin Gaywood at NatureScot.
This report, or any part of it, should not be reproduced without the permission of NatureScot or the relevant authors. This permission will not be withheld unreasonably. The views expressed by the author(s) of this report should not be taken as the views and policies of NatureScot. © NatureScot 2023.






