Scottish Wildcat Action (SWA) Specialist Report - Monitoring and Surveys
This report should be cited as: Campbell, R.D., Langridge, K., Kilshaw, K., Carus, H., Hislop, C., Rawling, E., and Tallach, N. (2023). Scottish Wildcat Action final report: Monitoring and Surveys. NatureScot, Inverness.
Authors and institutions: Roo Campbell - Scottish Wildcat Action, Keri Langridge - Scottish Wildcat Action, Kerry Kilshaw - WildCRU, Hebe Carus - Scottish Wildcat Action, Calum Hislop - Scottish Wildcat Action, Emma Rawling - Scottish Wildcat Action and Nicola Tallach - Scottish Wildcat Action. HC, CH, KL, ER and NT conducted fieldwork for this report. RC, KL and KK conducted the statistical analyses and wrote the text.
Year of publication: 2023
Reviewed: 2026

Background to SWA
The Scottish Wildcat Conservation Action Plan (SWCAP) was published in 2013 following the input of over 20 partner organisations. This led to the establishment of the Scottish Wildcat Action (SWA) project that ran from 2015-2020, funded by the Scottish Government, National Lottery Heritage Fund and others. SWA operated with a team of project staff managed by NatureScot, and associated work was carried out by various partner organisations. The overall work programme was steered by a group made up of ten of the partners. The International Union for Conservation of Nature (IUCN) Cat Specialist Group reviewed the work of the SWA, and other ongoing wildcat conservation work, and provided recommendations for future action (Breitenmoser et al., 2019). A wide range of topics relevant to wildcat conservation were covered during the SWA, and have now been published as a series of reports, of which this is one. These outputs will inform the next phase of wildcat conservation work in Scotland, including the ‘SWAforLife’ project that runs from 2019-2026.
Wildcats hybridise with domestic cats and we use a combination of morphology and genetics to distinguish wildcats from domestic cats and their hybrids. The method applied is generally determined by the practicalities of management. For example, it is much easier to have high confidence in the pelage scores from a sedated cat, than from a camera-trap image taken at night. Genetic and pelage results can only be generated jointly in certain scenarios. Therefore, identifications under different situations have different levels of confidence associated with them. We therefore set different thresholds for identification of wildcats based on the ability to distinguish pelage and genetic characteristics in different situations. The confidence hierarchy, and the definitions used in this report, are given below.

Wildcat definition
Click for a full description
A graphic showing increasing certainty of wildcat identity over five levels as more information on pelage and genetic status are available.
It reads:
Unless described otherwise, a wildcat in this report is defined as a cat with a 7PS pelage score of 19+ and/or that passes the genetic threshold for the genetic analyses. The consequences of applying different thresholds in different situations and management regimes are also examined.
A ‘wildcat’ in this report is defined at level 4 by a cat with a pelage threshold of 17+. Genetic information is not available for all cats and so is not used in population size assessments.
Background
Under the SWCAP, monitoring encompasses the following projects:
2.6 Monitoring of population trends in Priority Areas.
- 2.6.1 Develop and implement monitoring protocols to inform approaches in Priority Areas.
3.5 Standardise wildcat records
- 3.5.1 Agree standards for wildcat records / criteria for National Biodiversity Network (NBN) records.
3.6 Develop a protocol for wildcat samples.
- 3.6.1 Agree a protocol for the collection and archiving of wildcat specimens/ samples/ post-mortem and disease screening.
3.7 Monitor national wildcat distributions and population trends.
- 3.7.1 Develop and implement a programme of monitoring for wildcats.
Following this, the National Lottery Heritage Fund asked us:
“To continue monitoring and surveying of wildcat populations in the six Priority Areas. Camera trap images will be screened and analysed. Samples of genetic materials from wildcats and feral cats will be collected to increase knowledge of hybridisation and genetic diversity, as well as disease. Habitat quality and prey availability will be assessed, and conservation professionals will make recommendations for reintroduction sites.”
Note that the recommendations for reintroduction sites fall outside the scope of this report and will be reported elsewhere.
Summary of work
- We collected data on cat populations within the designated SWA Priority Areas (PAs) and more widely using baited camera trap surveys, collection of road casualty cats, live-trapping of cats during ‘trap neuter vaccinate and return’ (TNVR, non-wildcats) or for genetic screening and collaring (wildcats), and through the collection of public sightings.
- We conducted standardised camera trap surveys over winter in year one (2015-2016) for all PAs excluding Morvern, year two (2016-2017) for Morvern, and repeated winter surveys in year three (2017-2018) for the Angus Glens, Strathbogie, Strathpeffer and Strathspey. In each Priority Area during standardised surveys, volunteers (between three and 46, mean 26 per survey), staff and contractors set out between 60 and 142 (mean 82 per survey) baited camera traps for over 60 days each.
- We conducted additional targeted surveys in all PAs at other times, with the main aim of locating cats for trapping.
- For in-situ conservation, wildcats were distinguished from hybrids and domestic cats based on a standardised visual assessment of their pelage characters, with cats scoring 17-21 on the “Kitchener 7PC pelage score” classified as ‘wildcat’. This threshold is lower than the recommended “strict” definition of 19/21 proposed by Kitchener et al. (2005) but was deemed precautionary. Genetic samples provided additional complementary information regarding wildcat identification, but could not be used to direct the immediate decision-making required in the field, hence all population data reported here is based on the pelage system.
- In total, we invested 50,644 camera-trap days on standardised PA surveys, 35,755 camera-trap days on targeted surveys and over 10,492 camera-trap days on surveys by partners and affiliates outside priority areas (total = 96,891), collecting over one million images and videos.
- Images were assessed by project staff and cat images stored and catalogued. Individual cats were visually identified based on appearance, given a unique ID number, and recorded in a spreadsheet for each PA. Individual cats were pelage-scored from photos (and sometimes video) by two trained independent observers, and classified as wildcat (17+ and no single character scores of 1), hybrid (pelage score 10.5-16) or domestic cat (pelage score below 10.5 with no scores of 3).
- Across the project years (including data collected out-with standardised surveys) we detected a total of 31 individual wildcats, 162 individual hybrids and 151 individual domestic cats, excluding known pets (total 356 including 12 un-categorised cats). Overall, hybrid and domestic cats together outnumbered wildcats 10:1. Only two wildcats were detected outside our PAs during camera surveys.
- During standardised surveys and summed across sites, we detected one individual cat for every 201 camera days and one individual wildcat for every 1,746 camera days. Where we repeated surveys, effort per individual cat dropped by 43% but effort per wildcat increased by 12%.
- In Strathavon, the first standardised PA survey in year one and a follow-up PA survey in year two failed to detect any wildcats. In all other areas we detected between one and five wildcats during the first standardised PA survey and between zero and five wildcats in subsequent years, with evidence of turnover in the wildcat population during the project.
- Spatially explicit capture recapture (SECR) modelling was run on the standardised survey data only, providing population estimates given in the table below:
| PA | Survey Year | Population Size estimate of all cats (N) using SECR | Standard error | Lower confidence limit | Upper confidence limit | Actual number cats detected | Detections |
|---|---|---|---|---|---|---|---|
| Angus | 1 | 42 | 8.96 | 30.88 | 69.12 | 24 | 142 |
| Strathbogie | 1 | 31 | 1.35 | 29.27 | 34.75 | 27 | 249 |
| Strathpeffer | 1 | 48 | 9.59 | 35.13 | 74.40 | 21 | 228 |
| Strathpeffer | 3 | 60 | 8.52 | 47.97 | 82.34 | 30 | 88 |
- Overall we detected too few wildcats in our Priority Areas (mean density of 0.01 (± 0.005) wildcats/km2) to be sustainable in the short-term without contribution from any wildcats that might exist outside the PAs, and too few wildcats nationally to be viable in the long-term.
- In areas where standardised PA surveys were repeated over more than one winter, we detected no change in the wildcat populations, except in Strathpeffer where the survey in year three found no wildcats. This was potentially due to significant weather differences with the first winter (year one) very mild compared to year three, affecting cat movements.
- The robustness of repeated standardised PA surveys was compromised to some degree by the conflicting needs of systematic monitoring, maximising detection for management intervention, and baiting/trapping/removing cats during the survey.
- Some individual cats survived the duration of the project, and we were able to identify several individuals detected during research conducted in 2013-2014. Only one of those cats was confirmed as surviving until the end of this project, however, with the oldest known individual surviving until at least seven-years-old.
- Organising camera trap deployment, camera trap and volunteer management, data handling and data analysis took considerable staff time that, during full survey years, severely compromised the amount of time we could invest in TNVR. Balancing the requirement for robust monitoring with conservation actions needs to be considered carefully during future wildcat conservation work. Volunteer management for any field activity requires significant staff time. Streamlining is needed for the handling of data collected by volunteers.
- We live-captured a total of 228 cats, including 20 wildcats, and obtained 13 hair samples and two saliva samples through non-invasive means. We obtained a further 58 samples (mostly hair) from live cats not captured by the project, including from five wildcats. We obtained samples from 51 cats killed on the roads.
- No other cats (scoring below the pelage threshold) passed the genetic threshold for a wildcat; and with only five of 73 cats captured further than 50m from buildings testing as domestic cat. These results suggest that in the Priority Areas almost all wild-living cats are hybrids. The situation appears to be similar elsewhere in northern Scotland (see the Genetics and Morphology report for more details).
- We collected 769 records through the sightings page on the SWA website and the app, of which 317 were described by the observer as a wildcat. We were only able to verify 45 of these as wildcat, with an additional 106 reports verified as ‘plausible’ wildcats. Excluding plausible wildcats, no wildcats were verified north of Lairg, indicating either very low numbers or their extirpation from the far north.
Acknowledgements
The SWA project was supported by the National Lottery Heritage Fund.
It was also funded by the Scottish Government, NatureScot and the Royal Zoological Society of Scotland. Support and resources for associated work was also provided by all the Steering Group members: Cairngorms National Park Authority, Forestry and Land Scotland, National Museums Scotland (NMS), Scottish Land & Estates, NatureScot, Scottish Wildlife Trust, The National Trust for Scotland, The Royal (Dick) School of Veterinary Studies, The Royal Zoological Society of Scotland (RZSS), The Scottish Gamekeepers Association, The Scottish Government, Wildlife Conservation Research Unit (WildCRU). In addition, Jenny Bryce and Fiona Strachan helped develop the project while Eileen Stuart, Alan Bantick and Andrew Kitchener chaired the Steering Group.
Additional sponsors, funders and contributors to SWA work included Aigas Field Centre, Ailsa Black, British Association for Shooting and Conservation, Cairngorm Brewery Company, Chester Zoo, Roy Dennis Wildlife Foundation, John Muir Trust, Loch Lomond & The Trossachs National Park, Lukas Ruiter Production, MyPetOnline, NFU Scotland, petDetect, RSPB, Scotland The Big Picture, Swift Ecology, the Highland Council and Robertsons Trust.
We are also very grateful to the many academics, landowners and managers, farmers, foresters, keepers, captive collection managers, veterinary specialists and others who contributed valuable time and effort, members of the public who contributed funds and support, and in particular the many volunteers who got involved in the project.
Dr Stefano Anile undertook an independent review of this report prior to publication, for which the authors are sincerely grateful. Our volunteers worked extremely hard in collecting camera data for the project over multiple years, many covering several locations and some investing several days each week during winter, working in all weathers to cover cameras for us. We also hosted several full-time volunteers who contributed hugely to the monitoring. These were Laura Braunholtz, Jen Clark, Claire Hill, Ana Luisa Barros, Kirstin Mair, Ana Morales González, Fiona Pamplin, Héctor Ruiz, Lara Semple, Laura Sessions, and James Walker. Various contractors (Dr Roisin Campbell-Palmer, Kelsey Wilson, Kirstin Mair, Matt Wilson and Rhian Evans) also conducted camera monitoring and, in some instances, trapping. Local veterinary practitioners and Dr Alice Bacon collected samples for genetic screening. The RZSS’ WildGenes lab conducted the genetic screening presented in this report and various other partners contributed to our monitoring by conducting surveys on their own ground (see the main content). Dr Phil Bacon assisted in organising the database linking the myriad of records collated during cat sampling. Jamie Sneddon helped with organising camera trap images and cataloguing cats. Lastly, several hundred members of the public took the time to submit their sightings of wildcats, hybrids and feral cats that helped build the wider picture of the cat population in Scotland. Our profound thanks to all.
So many others contributed to the monitoring work in some form that errors of omission are likely, for which we apologise.
We would like to extend special thanks to colleagues at SWA for their dedication and hard work throughout the project: Emma Rawling, Nicola Tallach, Calum Hislop, Hebe Carus, Duncan MacKenzie and Vicky Burns. Also Matt Wilson, Dr Alice Bacon, Dr Philip Bacon, Dr Roisin Palmer-Campbell, Kelsey Wilson, Kirstin Mair and Norman Davidson.
1. Introduction
Monitoring is essential to evaluate the efficacy of any conservation action (e.g. Lindenmayer et al., 2006, Mills et al., 2006). However, monitoring is a broad term and we should begin by defining it. Here we use the term monitoring to mean establishing, through surveys or reports from the public, the presence or population metrics of the species or animal of interest and, where possible, using such records to improve knowledge on the ecology of the species and either estimating their numbers or arriving at an index of abundance.
By establishing presence of the species of interest, monitoring allows the targeting of conservation actions. The aims of such targeting may not align with the other aims of monitoring outlined above. For example, adopting improvements in detection methods is desirable for wildlife management, but changes in methodology will complicate the analysis of population change from repeated surveys. Therefore, it is a tool with multiple and sometimes conflicting uses.
Monitoring of the wildcat Felis silvestris in Scotland is complicated by the presence of widespread introgressive hybridisation with the introduced domestic cat F. catus (Macdonald et al., 2004). Hybridisation occurs when individuals from closely related but genetically distinct species/populations (in this case, wildcat × domestic cat) interbreed and produce hybrid offspring. If the first generation (F1) hybrids are viable and fertile, they can interbreed (backcross) with one or both parent species resulting in gene flow or ‘introgression’ between the two populations and can result in introgression hybridisation when a hybrid (e.g. wildcat hybrid) goes on to interbreed (backcrosses) with one of the parent species (e.g. wildcat or domestic cat) resulting in gene flow between the two populations (Rhymer and Simberloff, 1996, Woodruff, 1973, Anderson, 1949). Introgression is a long-term process, taking many generations of hybrids backcrossing and results in individuals with a complex mixture of genes and morphological characteristics. Continuous and repeated introgression can gradually dilute the genetic integrity of the rarer species, resulting in a hybrid population with a complex mixture of genes and morphological characteristics. Hybridisation in the British wildcat population was first documented 200 years ago (Bewick, 1820), then by Darwin (1875) and Cocks (1876) noted the potentially negative impact of hybridisation. Interbreeding between wildcats and domestic cats in Scotland was known (Pitt, 1939, Gray, 1971, Corbett, 1979) and the first genetic work carried out by Beaumont et al. (2001) and Daniels et al. (2001) noted that introgression appeared to be occurring in Scotland with two main groups existing, those closest to wildcat (based on phenotype and genotype) and those closest to domestic cat. Introgression is now so extensive in the Scottish wildcat population that it has resulted in a genetic and phenotypic continuum from wildcat to domestic cat known as a ‘hybrid swarm’ (Daniels et al., 1998, Daniels et al., 2001, Macdonald et al., 2004, Senn and Ogden, 2015).
Hybridisation presents significant challenges for conservationists, for example in defining and identifying what is to be conserved (a problem compounded by the continuous nature of the hybrid swarm). A standardised method of identification was developed by Kitchener et al. (2005) using a pelage scoring system to distinguish between wildcats, hybrids and domestic cats based on their external appearance. It uses seven pelage characteristics (7PC), scoring each from 1 (domestic cat) to 3 (wildcat), giving a possible final score from 7 to 21. Kitchener et al. (2005) set the threshold for a ‘strict’ wildcat definition at 19 and above and a more ‘relaxed’ wildcat definition at 14 and above with no individual characteristic scoring 1.
SWA employed a precautionary working threshold of 17/21 to define a wildcat, to balance the somewhat conflicting aims of maximising retention of the wildcat gene pool while minimising continued introgression. This lower threshold also recognised that in a field situation there may be uncertainty in the scoring of pelage characters. Although a genetic test was developed under the SWCAP (see the Genetics and Morphology report and Senn et al., 2019), it was not practical to rely on genetic scores to guide our fieldwork in the Priority Areas because the turnaround time for test results was generally too long, especially where immediate decisions were required. For example, a typical in-situ conservation action might include a staff member trapping a cat, confirming it is not a wildcat based on a pelage score <17, and transferring the cat to a veterinary clinic for neutering and vaccination before releasing it at the site of capture within 24 hours (see the Trap Neuter Vaccinate Return Programme report).
Below we summarise methods used by others for surveying wildcats, before describing the agreed actions under the monitoring work programme in more detail. For consistency throughout this report, we refer to any cat that scores 17 or more on the 7PC score as a ‘wildcat’, irrespective of subsequent genetic test results. This reflects the current in-situ situation where management and monitoring of wild-living cats has to be done reactively using the pelage definition, until a more rapid genetic test can be developed. Genetic scores of the cats within the Priority Areas are provided in the Genetics and Morphology report and will be used in discussion here for context.
1.1 Methods available for wildcat monitoring
1.1.1 Public sightings, predator control activity, and collection of road casualties
As is the case with many monitoring programmes for mammals in the UK, most early surveys of wildcats in Scotland relied on reports or survey responses from members of the public and stakeholders (e.g. Jenkins, 1962, Easterbee et al., 1991, Davies and Gray, 2010, Hetherington and Campbell, 2012), although these records are confounded by geographical variation in effort and recording (Easterbee et al., 1991).
Prior to legal protection of the wildcat in 1988, records from game bags were used to assess the number and distribution of wildcats killed by gamekeepers during predator control activity between 1971-1987 (Easterbee et al., 1991). With changes in public perception regarding lethal control of feral cats, records of feral cats killed by gamekeepers have become more difficult to obtain. For example, Hetherington and Campbell (2012) invested considerable effort in securing such data from just five estates in the Cairngorms National Park, only obtaining records through anonymising the estates. In comparison, data collected prior to protective legislation by Easterbee et al. (1991) and Balharry and Daniels (1998) indicated high levels of both legal control of feral cats and persecution of wildcats.
Another monitoring method is the collection of road-casualty cats, which has the benefit of allowing phenotypic, genetic and other morphological assessments (as well as other procedures, such as disease screening). Although these methods have the advantage of requiring relatively little effort with widespread coverage, they may be biased in their coverage, with a relatively higher rate of reporting likely from areas with higher population densities (and thus more eyes to see and more traffic to collide with), or from land managed for game birds rather than for other purposes. Additionally, hybridisation has complicated the accurate identification of wildcats versus hybrids and tabby domestic cats, resulting in survey data that is of uncertain quality. Nevertheless, in the context of a wider monitoring programme, all these methods can and have contributed to the wider picture.
1.1.2 Live-trapping
Physical capture of cats allows sedation and detailed assessment of phenotype along with genetic sampling and disease screening. The process is invasive because cats have to be injected to be sedated, which is inevitably stressful for the captured individual. It also requires a veterinary clinician or, if the work falls within the Animals (Scientific Procedures) Act 1986, a Home Office licence holder. Furthermore, a licence to trap is needed from NatureScot because of the wildcats’ status as a European Protected Species. This method has been successfully employed by previous studies (e.g. Scott et al., 1993, Daniels, 1997, Littlewood et al., 2014, Campbell, 2015), however considerable effort is needed. For example, Daniels (1997) recorded 155 trap-nights and Potocnik et al. (2002) 59 trap-nights of effort per capture, although this was reduced greatly to 26 trap-nights by Littlewood et al. (2014) through the use of camera traps for identifying the best live-trapping locations (see below). The physical effort of frequent trap checks can be further reduced by employing a remote trap-trigger alarm system, such as MinkPolice or GPRS cameras, that sends images of the trap to the trapper (Campbell and Griffith, 2015), although this is dependent on the local mobile network coverage. The advantage of live-capture is that biases inherent in sightings from the public or road casualties can be eliminated with careful selection of trapping areas, while phenotype, genetics and disease assessments can be conducted.
1.1.3 Camera surveys
Camera traps are remote cameras that are triggered by a sensor that detects changes in temperature and motion. When an object (e.g. an animal) with a different temperature from the air around the camera (e.g. a warm blooded mammal) moves across the camera’s “detection zone” the camera is triggered to take a photograph or video. Camera trap technology has advanced considerably in recent decades, with the development of low-cost digital models that can be left in-situ for months at a time, continuously collecting images for review or even sending them immediately to researchers via a mobile network. Digital remote cameras have revolutionised the study of rare, elusive and often nocturnal species around the world (Karanth, 1995, Mudappa, 1998, Gompper et al., 2006, Soisalo and Cavalcanti, 2006), including the wildcat in Scotland (Kilshaw and Macdonald, 2011, Kilshaw et al., 2015, Kilshaw et al., 2016, Hetherington and Campbell, 2012, Littlewood et al., 2014) and other parts of Europe (Anile et al., 2012, Can et al., 2011, Gil-Sánchez et al., 2020). Many wildcat species have unique individual markings (e.g. stripes, spots) allowing researchers to identify individuals and apply statistical analyses such as spatially-explicit-capture-recapture (SECR) models to estimate population densities and other population parameters. Such data can allow other population parameters to be measured in addition, such as population growth rate, turnover (recruitment and mortality) and even maximum longevity (e.g. Anile et al., 2020).
In Scotland, camera traps were first tested as a method to survey for wildcats in 2009 (Kilshaw and Macdonald, 2011, Kilshaw et al., 2015) and later used to systematically survey for wildcats as part of the Cairngorm Wildcat Project (Hetherington and Campbell, 2012) and across the known range of the wildcat in Scotland (Kilshaw et al., 2016). Both latter studies used pairs of cameras with bait at 20 locations set roughly one km apart to form a grid, with the bait refreshed about every two weeks. The combined survey map from these two studies can be found in Kilshaw et al. (2016). As with live-trapping, camera traps do not have the same location bias inherent in public sightings and road-casualty cats, although other biases may arise. For example, if cameras are sited in known preferred habitat instead of randomly, the use of bait, and landowner access permission, may also impact the coverage of the trail camera survey. In addition, the detection success can vary in relation to camera model and set up (Driessen et al., 2017, Urlus et al., 2014). However, relatively detailed assessments of phenotype can be made from the images, provided cameras are set up correctly (e.g. see ANNEX 1), enabling pelage scoring under the Kitchener 7PC system, although sometimes it is not possible to see all pelage characters on individual cats. The method requires considerably less effort than live-trapping and does not need such a high level of training. Indeed, camera traps are also a useful engagement tool, because the cameras can be set up easily by volunteers and images of cats shared with other volunteers and the public.
1.1.4 Hair posts
Hair traps, consisting of roughened wooden posts, sprayed with valerian tincture (Valeriana officinalis), were first used to monitor wildcats in Germany (Weber, 2008, Hupe and Simon, 2007) and are still the primary method of surveying the European wildcat there (Steyer et al., 2013). Valerian causes a similar behavioural response in some cats to catnip (Nepeta cataria), attracting the cat in close proximity and inducing it to rub against the wooden posts, although there may be a bias in using scent lures because the response to them is genetic (Waller et al., 1969). For example, a study comparing the catnip response in a variety of wild–living cats found that some species reacted very strongly in a similar manner to a domestic cat (e.g. lion, Panthera leo and jaguars, P.onca) and others barely at all (e.g. tigers – P. tigris, bobcats – Lynx rufus and cougars – Puma concolor) (Hill et al., 1976). Bol et al. (2017) found that one in three domestic cats did not respond to catnip but of these 75% respond to silver vine (Actinidia polygama) and 50% to Tatarian honeysuckle (Lonicera tatarica) and Valerian. Kilshaw and Macdonald (2011) and Campbell (2015) found a limited response of wild-living cats in Scotland to valerian tincture or catnip extract. Similarly, a study in Sicily also found that wildcats there did not react much to valerian tincture (Anile et al., 2009). However, during this project we tested dried valerian root and found this to be very successful in attracting some cats. This could be because the root retains its scent longer than the tincture does in Scotland’s wetter climate. We suggest testing other scents, such as the silver vine, might improve monitoring methods for the wildcat in Scotland.
When the cat rubs against the wooden post, hairs are often trapped in the splinters and genetic material can then be extracted for screening. If a sufficient number of hairs are extracted (≥20), genetic analysis to identify species and individuals can be carried out (Steyer et al., 2016). SNPs (Single Nucleotide Polymorphisms) can also be used to identify wildcats from hybrids and domestic cats if sufficient high-quality genetic material can be extracted (Nussberger et al., 2013, Senn and Ogden, 2015). The use of this method in conjunction with cameras could allow non-invasive pelage and genetic assessments of cats. However, the limitations of this method are the difficulties in identifying which individual the hairs come from if more than one cat visits the station, regular checking of posts to remove hairs and insufficient hairs being left for genetic analysis. During this project, some of the volunteers (NW and PB) developed a prototype hair trap that worked well and could be developed further. One volunteer (PB) also noted that visiting cats sometimes chewed the cotton pouches in which valerian root was held at these hair posts. The chewed pouch yielded sufficient genetic material for the 35 SNP test (see the Genetics and Morphology report) and this approach was used successfully to genotype two individuals during the project.
1.1.5 Scat surveys
Scats can potentially provide information on species presence, as well as data on diet and material for genetic screening (Stahl et al., 1992, Sarmento, 1996, O'Reilly et al., 2008, Mucci and Randi, 2007, Adams et al., 2003). However, distinguishing between a wildcat, hybrid or domestic cat scat on appearance alone is impossible. Even distinguishing reliably between scats from wild-living cats and from other carnivores, in particular the red fox (Vulpes vulpes) can be difficult without genetic analysis (e.g. Lozano et al., 2013). Kilshaw (2015) attempted to extract DNA from wild-living cat scats in Scotland but found that 57% of scats did not yield DNA of sufficient quality for testing for mtDNA (mitochondrial DNA) and none for higher resolution testing. This is likely a product of the age of the scats used as studies in Europe have managed to extract DNA from fresher scats (e.g. Velli et al., 2015). Indeed, in two separate studies, Stefano Anile improved the success of extracting genetic information from scats for microsatellite analysis from 38% (Anile et al., 2014) to 66% (Anile et al., 2019) through changes to sampling protocols. Scat collection in Scotland could also be improved by using scat dogs (e.g. Long et al., 2007a, Long et al., 2007b); fresher scats would therefore increase the quality of the DNA that can be extracted.
1.2 Measuring population change from camera surveys
Before monitoring began, NatureScot commissioned a modelling study to assess the survey design and the effort needed to measure any population changes in wildcats (Newey et al., 2015). Newey et al. (2015) modelled the ability to detect a 25% change in population density with 80% power over a range of scenarios, including different numbers of camera traps, trap spacing, survey duration, capture probabilities (g0=0.01 and g0=0.02) and sigma (σ, a movement parameter representing home range size). Assuming a value of sigma =700m (the highest value they modelled), these findings suggested a minimum requirement of 132 cameras spaced in a grid of one km squares for 60–80 days (the range of survey durations modelled). Unfortunately, deploying such a large number of cameras in each of the Priority Areas was financially and logistically impractical. In addition, because there was some uncertainty around the actual movement parameters and detectability of wildcats during the modelling (Newey et al., 2015), and because we required the information on the locations of cats for TNVR that camera traps gather, we decided to attempt camera surveys of the Priority Areas using as many cameras as possible, taking into account Newey et al. (2015) recommendations on survey spacing and duration and drawing from experience gained from previous surveys (Kilshaw and Macdonald, 2011, Campbell, 2015, Littlewood et al., 2014).
Moreover, a 25% change in the wildcat population over the relatively short duration of the project was deemed unlikely due to the nature of conservation strategy, which focused on neutering and returning domestic and hybrid cats and not removing them. The implications of neutering hybrid and domestic cats for wildcat, hybrid and domestic cat populations were unclear, as too little is currently known about behavioural and ecological interactions between the different species that could influence conservation outcomes. For example, neutered cats could potentially survive and continue to hold territories for several years, which in the short term could result in fewer opportunities for wildcat recruitment (or hybrid and domestic cat recruitment) into the Priority Areas, unless young wildcats could out-compete incumbent adult non-wildcats for home ranges.
2. Methods
2.1 Surveyed areas
National records were collected through public sightings data from across Scotland. Road casualty specimens collected by the public or stakeholders, and standardised camera trap surveys led by project partners, covered the near-historical range of the wildcat in Scotland. Project-led surveys and live-trapping were focused on the six Priority Areas (Table 1, Figure 1) that were selected following recommendations from Littlewood et al. (2014).
| Priority Area | AG | MO | SA | SB | SP | SS |
|---|---|---|---|---|---|---|
| Location at centre (OS) | NO2866 | NM7353 | NJ2024 | NJ4835 | NH4760 | NH9122 |
| Extent (Ha) | 37460 | 48697 | 34319 | 34725 | 33626 | 20576 |
| Extent exc freshwater (Ha) | 37215 | 48312 | 34319 | 34721 | 32497 | 20471 |
| High quality habitat (Ha) | 11068 | 12899 | 9885 | 11994 | 10904 | 7520 |
| Number of households | 347 | 246 | . | 1068 | 2543 | 1068 |
| % land managed by project partner | 8% | 16% | . | 27% | 15% | 1% |
Household and project partner land management were not calculated for Strathavon. AG = Angus Glens, MO = Morvern, SA = Strathavon, SB = Strathbogie, SP = Strathpeffer and SS = Strathspey

Figure 1. Priority Area boundaries.
Click for a full description
A map of Scotland excluding the south and the far north. The map shows the outline of the Wildcat Priority Areas: Angus Glens (southern Cairngorms); Morvern (west coast NE of Mull); Strathpeffer (NW of Inverness); Strathspey, Stratrhavon and Strathbogie (three areas in a line from Aviemore, through Tomintoul to around Huntly in Aberdeenshire).
2.2 Data collection
2.2.1 Public sightings
Throughout the project SWA promoted the reporting of wild-living cat sightings as one of the key ways that the public could contribute to wildcat conservation. Initially the public could report sightings of cats directly to project staff, but shortly after the project began a sightings page was created on the SWA website, followed later by a smartphone app. Both reporting methods inputted directly into the Biological Recording Centre’s iRecord website, which are periodically transferred to NBN Atlas Scotland. Sightings submitted to SWA or directly into iRecord are verified by project staff, following a protocol that clearly sets out the criteria of proof (e.g. details of description or photographic evidence) needed for the different available verification categories (see ANNEX 2). Cats were either defined as wildcat, domestic cat, or hybrid using the agreed SWA pelage threshold of 17/21 and above for wildcats based on the scoring system developed by Kitchener et al. (2005). More detail can be found on the implementation of the sightings webpage and smartphone app in the Communications report. Completion of this work addressed SWCAP action point 3.5 (Agree standards for wildcat records / criteria for NBN records).
2.2.2 Road casualties and other dead-found cats
Dead cats were collected throughout the project from Priority Areas and across northern Scotland by SWA and partner organisations. Most were found next to roads (some in barns and outbuildings) by members of the public, who alerted project staff for collection where possible. Where project staff could not respond immediately, people were advised to either remove and store the cat for future collection, or to take a hair/tissue sample and send to SWA with details of the location and date found. Carcases were stored in a freezer at one of the partner offices used by project staff or transported to RZSS Highland Wildlife Park/Edinburgh Zoo until such time as they could be transported to National Museums Scotland in Edinburgh for processing. Cats were initially labelled with locations and dates using cardboard luggage labels, and the information was recorded in a spreadsheet by SWA staff. In 2016 SWA developed a Cat Procedure Form (CPF, see ANNEX 3) that allowed detailed and standardised data to be recorded for every processed cat or sample, along with initial pelage assessments and other veterinary procedures in quadruplicate. Each set of forms came with a unique ID number (‘SWA UID’’) to allow tracking of the samples. Three copies were posted with each carcass for eventual distribution of samples to NMS, RZSS and R(D)SVS and one kept as a SWA record.
2.2.3 Live-trapping
Live-trapping was conducted for either the purposes of capturing potential wildcats for sampling and GPS collaring or for the capture of domestic cats under the trap, neuter, vaccinate and return (TNVR) programme (see the Trap Neuter Vaccinate Return Programme report for detailed TNVR methodology). Since TNVR was targeted in areas with wildcats, the two different trapping programmes were usually operating simultaneously, with different outcomes for individual cats dependent on the pelage score. Where trapping for TNVR involved cats living around human habitations and farms, cats living with one or more other cats were noted as coming from larger colonies of ≥5 individuals (‘colony’) or smaller colonies of <5 (‘few’). A detailed wildcat trapping protocol is provided in ANNEX 4. Data from any capture event were recorded on our Cat Procedure Form.
2.2.3.1 Timing
Targeted trapping for wildcats only took place over winter in years three to five of the project (2017-18, 2018-19 and 2019-20). In year two (2016-2017), TNVR trapping for domestic cats and phenotypic hybrids deliberately aimed to avoid the incidental capture of wildcats wherever possible, but any that were trapped were health screened and genetically sampled as per the protocol (see 2.2.3.5 Sampling). Trapping in areas frequented by wildcats was halted by early-mid March to reduce the chances of trapping a pregnant female cat or pregnant pine martens (Martes martes). TNVR trapping of domestic and hybrid cats could be done at any time of year, but winter was preferred due to the improved capture rates and to reduce the chances of affecting pregnant females or dependent young.
2.2.3.2 Bait
Traps that were set in remote areas, relatively further away from farms or houses (and likely targeting more ‘feral’ or ‘wild-living’ cats) were usually baited with meat, including game-birds (quail or pheasant), fish, chicken or rabbit, and a scent lure (valerian root and/or salmon oil). Domestic cat food was more often used as bait for trapping cats living in or around farms and human habitations. The different baits were used to maximise trapping success, by replicating the probable diet of target cats, and also because cat food is more perishable than ‘fresh’ baits and more difficult to replace regularly in remote trapping sites.
2.2.3.3 Traps
A variety of live traps was used for TNVR and wildcat trapping. ‘Trapman’ standard and large ‘cat traps’ have spring-loaded doors that close from the inside. The large model was particularly suitable for trapping larger and more wary cats, including wildcats. MDC Export cat traps (both PF and Eezicatch traps) were more suitable for TNVR, being slightly smaller and without spring-loaded door mechanisms. In several instances, where target cats were particularly large or trap-shy, we used Tomahawk-style fox traps and modified Albion Manufacturing fox traps with a one-inch-square gauge mesh. The larger size and externally-closing door helped to push the cat inside and increase trapping success. Tomahawk-style fox traps were only used in combination with trap alarms (MinkPolice) because the large rectangular mesh (1” x 2”) could possibly have caused tooth and claw injuries if cats were not released quickly after capture.
2.2.3.4 Targeting trapping effort
Live-trapping was almost always preceded by some form of standardised or targeted camera survey, except when project staff were reacting to detailed information from members of the public, usually relating to domestic or hybrid cats for TNVR. Aside from TNVR of feral cat colonies, all live-traps were monitored with camera traps prior to and after the capture of target individuals.
2.2.3.5 Sampling of captured individuals
Trapped cats were blood sampled for genetic and disease screening, with up to 3ml of blood transferred to 0.5 – 1ml EDTA tubes. Buccal, oropharyngeal, conjunctival and rectal swabs were also taken. Samples were only collected from a proportion of cats when conducting TNVR on large farm colonies. Hair or (very rarely) faecal samples were taken from cats that were trapped but had to be released without treatment (for example, incidental capture of wildcats during TNVR, or by a gamekeeper conducting predator control). Whisker samples were taken from February 2018 onwards in order to assess diet through stable isotope analysis as part of a PhD project in conjunction with the University of Exeter. Analysis of the whiskers has not yet been completed and so will not be reported here. Sample collection data was recorded on the Cat Procedure Form (see 2.2.2 above) and all samples were labelled with the respective SWA UID number. Wildcats were injected with a PIT tag (microchip) prior to release, and the unique tag reference number recorded to aid future identification. Some individuals (n = 14) were fitted with GPS collars (see the Ecology report) by project staff. All sample data was subsequently entered onto a central SWA spreadsheet, and Cat Procedure Forms were scanned and stored.
2.2.4 Camera trap surveys
Camera traps were used to conduct standardised surveys of the wild-living cat population in each Priority Area over the winter (‘Standardised PA Survey’, see Annex 5 for maps of camera and cat locations). They were also used more informally to locate cats and target the conservation activity throughout the year, but primarily also over the winter (‘Targeted Survey’). Camera traps deployed during standardised PA surveys were predominantly managed by local volunteers, each managing a small number of cameras (often just one or two), with the remainder managed by project staff, staff from partner organisations (e.g. Forestry and Land Scotland) or ‘residential’ volunteers, recruited to assist the Project Officer for the duration of the survey season. The more remote, less populous (therefore fewer volunteers available) and logistically difficult Priority Areas of Morvern and Angus Glens required additional assistance from paid contract staff during the winter field season, as detailed below:
- The standardised PA camera trap survey in Morvern during year two relied mainly on a local contractor (126 camera locations), with local volunteers covering 12 camera trap locations, and four camera locations covered by a consultant for Aggregate Industries to access the restricted Glensanda quarry area
- Targeted camera trap surveys in the Angus Glens in years two and five, where a contractor was carrying out the TNVR and wildcat trapping
Standardised PA camera trap surveys were carried out in each of the six Priority Areas, excluding Morvern in years one and two, and in four areas (Angus Glens, Strathbogie, Strathpeffer and Strathspey) in year three (see Table 2). In other years, targeted camera trap surveys were carried out with the principal motive of detecting cats, either for TNVR or wildcat genetic screening and GPS collaring. Following the cessation of work in Strathavon at the end of year two, the increased staff time available in Strathbogie allowed the recruitment and management of a relatively large number of volunteers to assist in camera surveys. Therefore, even in the most targeted survey years, the number, duration and re-baiting schedule of cameras in Strathbogie were equivalent to those in standardised survey years.
Most camera trap deployment occurred during winter because previous camera trap surveys have shown that detectability of wild-living cats is higher during these months (Kilshaw et al., 2015, Kilshaw and Macdonald, 2011). Occasionally, cameras were deployed during the summer and autumn, mainly to coordinate TNVR. Attempts were also made to monitor a potential breeding den of a GPS-collared female wildcat. Following licensing restrictions, no bait was used, the camera was deployed at least three metres (usually further) from the den and set-up time near the den was minimised. We contacted landowners (and ground staff) and secured permission to place the cameras prior to deployment of all camera traps.
2.2.4.1 Camera trap models
We used a mixture of camera trap models for survey work. SWA inherited 135 Cuddeback (Black Flash Ambush and Black Flash E3 models) from previous wildcat surveys, and acquired an additional 297 Spypoint Force 12 models with project funds at the start of the fieldwork. Following a number of camera losses from failures (approx. <10% per year over the project’s duration) and a small number of thefts, Spypoint cameras were replaced with 40 Spypoint Force 11d models, six Spypoint Force 10 models, four Bushnell HD Aggressor models and ten Reconyx Hyperfire 2 (the latter donated by Siemens). Spypoint Force 12 and 11d models were chosen because at the time they were two of just a few available camera traps with integrated viewing screens, allowing easy set-up and review of images by volunteers. The Bushnell and Reconyx cameras were not used for any of the full surveys; Bushnell HD cameras were primarily used to gather high-quality video footage of cats for use in communications.
2.2.4.2 Standardised surveys
Camera placement
In contrast to past wildcat camera trap surveys in Scotland (Kilshaw et al., 2015, Kilshaw et al., 2016, Hetherington and Campbell, 2012), we only used one trail camera at each location, following Littlewood et al. (2014). This allowed greater coverage in terms of area, but increased the risk that a visiting cat would either be missed (see Pease et al., 2016; but note the use of bait in this study would have counteracted the use of a single camera to some degree) or not identified where too few images of both sides of a cat were captured. Based on knowledge of cat habitat use from past camera surveys (Kilshaw et al., 2015, Kilshaw et al., 2016, Hetherington and Campbell, 2012) and radio tracking studies (Corbett, 1979, Scott et al., 1993, Daniels, 1997, Campbell, 2015), we directed volunteers to place cameras near the interface between cover and open habitat, or along linear features such as walls and fence lines. We also encouraged them to look for ‘pinch-points’ for cats moving through the landscape, such as narrow corridors of cover habitat, water courses or bridges over water, breaks in walls, or holes in otherwise animal-proof fencing.
Project officers suggested potential camera locations based on a mapping exercise (using aerial satellite and Ordnance Survey [OS] maps) ahead of visiting field sites. Volunteers were trained in the field by Project Officers (or occasionally by trained residential volunteers) to locate and set the camera-traps using standardised methodology. We instructed volunteers to set cameras at approximately cat head-height from ground-level (20-40cm), between 1.5-3m from the target detection area (bait attached to a post or tree, or from the path or trail), and facing away from direct sunlight and the direction of inclement weather. We asked volunteers to remove vegetation between the camera and the bait as this can obscure photographs of the cats, cause false triggers or lead to under-exposure of flash-lit images during darkness. In addition to volunteer training, we created a set of online videos describing the camera set-up, and later in the project we also produced a wildcat camera-trapping leaflet (ANNEX 1). Results from the camera surveys, including by-catch of other species, were fed back to the landowners at the end of the survey if requested.
| Year | Detail | Angus Glens | Morvern | Strathavon | Strathbogie | Strathpeffer | Strathspey | Total |
|---|---|---|---|---|---|---|---|---|
| 1 | Survey type | SD | None | SD | SD | SD | SD | - |
| 1 | No. camera locations | 63 | - | 60 | 80 | 82 | 69 | 354 |
| 1 | No. camera-days | 3,800 | - | 4,394 | 6,070 | 4,997 | 3,974 | 23,235 |
| 1 | No. volunteers | 20 | - | 20 | 40 | 40 | 19 | 139 |
| 1 | Timing MM (peak MM) | 12-04 (01-03) | N/A | 12-05 (01-03) | 12-05 (01-03) | 12-05 (01-03) | 12-03 | - |
| 2 | Survey type | TNVR | SD | TNVR | TNVR | TNVR | TNVR | |
| 2 | No. camera locations | 124 | 142 | 40 | 80 | 33 | 40 | 459 |
| 2 | No. camera-days | 4,621 | 10,086 | 2948 (30) | 7,241 (35) | 1,848est | 2,240est | 28,984 |
| 2 | No. volunteers | 0 | 3 | 12 | 38 | 20 | 12 | 85 |
| 2 | Timing MM (peak MM) | 12-03 | 12-03 (01-02) | 12-03 | 12-03 | 09-08 (01-03) | 12-02 | - |
| 3 | Survey type | SD | TNVR | None | SD | SD | SD | - |
| 3 | No. camera locations | 54 | 42 | - | 84 | 89 | 96 | 365 |
| 3 | No. camera-days | 3,159 | 2,352est | - | 5,057 | 4,250 | 4,857 (91) | 19,675 |
| 3 | No. volunteers | 23 | 3 | - | 46 | 30 | 16 | 118 |
| 3 | Timing MM (peak MM) | 12-03 (01-03) | 01-03 | - | 12-04 (01-03) | 01-04 (01-03) | 12-04 (01-03) | - |
| 4 | Survey type | TNVR | WC trapping | None | TNVR | None | None | - |
| 4 | No. camera locations | 82 | 40 | - | 37 | - | - | 159 |
| 4 | No. camera-days | 5,002 | 2,240est | - | 2,657 (34) | - | - | 9,899 |
| 4 | No. volunteers | 7 | 1 | - | 20 | - | - | 28 |
| 4 | Timing MM (peak MM) | 12-03 | NA | - | 08-11 (01-04) | - | - | - |
| 5 | Survey type | WC trapping | None | None | WC trapping | None | None | - |
| 5 | No. camera locations | 16 | - | - | 69 | - | - | 84 |
| 5 | No. camera-days | 777 | - | - | 3,829 (65) | - | - | 4,606 |
| 5 | No. volunteers | 0 | - | - | 18 | - | - | 18 |
| 5 | Timing MM (peak MM) | 12-03 | - | - | 10-03 (12-03) | - | - | - |
| Total | No. camera days | 17,359 | 14,678 | 7,342 | 24,854 | 11,095 | 11,071 | 86,399 |
Camera spacing
For standardised surveys, we aimed to deploy approximately 70 or more cameras in each of the Priority Areas. Exceptions to this were Strathspey, due to its relatively small size, and the Angus Glens, where we were unable to secure sufficient land access in suitable habitat. We created a grid for each Priority Area with a cell size of 1.25km and used this to guide the camera spacing, aiming to have no more than one camera per grid cell. The exception to this was Morvern, because of the large areas of open habitat. In Morvern cameras were instead placed at high density around habitat edges and pinch-points (see Annex 5).
Camera bait
Cameras were baited with meat that was usually frozen quail or sometimes other gamebirds, such as partridge or pheasant. Volunteers occasionally used other meat that was available, such as rabbit. Bait was stored frozen and distributed to volunteers when needed. Some volunteers were able to store bait at home, while for others, bait was stored by SWA and distributed throughout the survey. The bait was tied to a tree or a post in front of the camera trap. In year one, pheasant wings were added to the bait set-up as a visual lure tied 1-2m above ground from an overhanging branch. In years one and two, salmon oil (SOS Salmon Oil for Dogs) was also used as a scent lure. This was poured onto a sponge inserted into a drilled hole on a wooden post and/or poured over the wooden post. We later switched to using dried valerian root (BuyWholefoodsOnline brand) as a scent lure, following trials that showed it to be an effective cat attractant that encouraged rubbing. The root was tied into a small cotton/jute cloth pouch and secured to a post or tree trunk. Due to logistical constraints, we did not run experiments to measure the difference in cat detectability when using salmon oil versus valerian root.
Camera checks and survey duration
Volunteers were asked to visit their cameras every two weeks to replace the bait and switch SD cards to recover the images. Including set up and camera removal, this meant a total of five visits over the trapping season of 60 days. If no cats visited the camera in the first 30 days and the surveyor subsequently thought that the location was sub-optimal, they could move the camera to a nearby location (i.e. within 50-100m) and this information was recorded. The camera trap stations were removed after 60 days, though some cameras remained out for longer, for example to try and collect more images of individual cats for accurate pelage scoring, or if the camera had malfunctioned. During full surveys conducted in year three, we placed cage traps out at some locations once cats were detected (see 2.2.3 above).
2.2.4.3 Targeted surveys
Targeted surveys to find cats for trapping followed the same general placement and baiting methods outlined in 2.2.4.2 above. However, there were no restrictions on the spacing of cameras, the duration that a camera was deployed at a location, or on the frequency of checks/rebaiting.
2.2.4.4 Other surveys
We encouraged partner organisations and other interested groups or individuals to conduct camera surveys outside Priority Areas following our methodology. We created an instruction leaflet describing our methods (ANNEX 1), which was available in hard copy and on the website and provided training or advice. A summary of these additional surveys is provided in Table 8.
2.2.5 Non-invasive genetic sampling
During standardised PA camera surveys and for some targeted surveys, wooden hair-trap posts were set out at each camera trap station, with salmon oil or dried valerian root as a lure (see 2.2.4.2 above). The posts were rough wood onto which we stapled strips of hook-side Velcro to catch hair as the cats rubbed the post. These did not prove to be effective, and only yielded a handful of hair samples from Strathbogie, which only gave partial genetic results. After year one, we used valerian pouches only, and attempted to collect hairs from the rough wooden posts (onto which the pouches were tied). Later in the project we attempted to collect saliva from valerian pouches that had been chewed by a cat, following the discovery by a volunteer that these could yield DNA suitable for analysis. Both methods provided only a very small number of samples. Another volunteer experimented with alternative hair sampling methods, including tunnels with a hair-trap (a lint roller or a coiled spring) fixed to the underside of the roof. This proved more effective in collecting suitable samples than the passive ‘rubbing’ methods described above, but because of their complexity we could not adopt this method widely and it was only used occasionally by volunteers in Strathbogie. We also occasionally collected scats for genetic sampling, and collected hairs from cats that were trapped by the project or by gamekeepers and which could not be sedated for blood sampling. Non-invasive samples were not analysed unless we were confident that they had come from a single individual. All samples were recorded on our CPF and accompanying spreadsheet.
2.3 Data analyses
2.3.1 Identification of cats
Volunteers were asked to immediately send images of cats captured by their cameras to the Project Officer. Where the volunteer was unable to email images, the Project Officer would collect the SD cards in person. Volunteers were asked to send a few of the best images that allowed identification and pelage scoring of the individual. After the full surveys, all camera-trap images were reviewed by the Project Officer to record by-catch and ensure that no cat capture events were missed.
Individual cats were identified based on appearance, particularly pelage markings, but size and overall morphology were also used for cats that looked alike (especially black cats, see below). Tail markings and the pattern of striping and spots on the fore- to mid-flanks were particularly useful for diagnostics. We also used markings on the forelimbs, head and face, and nape and shoulders. Some markings vary in appearance depending on the angle of view: for example, the dorsal line down the back could vary within individuals depending on the lay of the fur and position of the tail/haunches. Cats also appear very different when they are wet, and the coat markings can look different in the summer and winter. The use of black-flash camera traps meant that night-time images (taken using infra-red light) artificially enhance the contrast in the wildcat pelage, such that the pattern stands out more and makes the individuals easier to identify (although this may not represent the degree of contrast visible in the pelage with the naked eye). Multiple traits (including the cat’s location) were ultimately used to determine individual identification. Where possible (and especially if the first assessor was not confident), one or more additional assessors reviewed the images to ensure accurate identification (particularly for wildcats).
Low-quality images (e.g. from very grainy night-time images or motion blur) often precluded accurate identification of individuals, and these were marked as unidentified and not used in statistical analyses. Black cats were particularly problematic to identify as individuals. For cats with less distinct markings, sometimes the gross appearance allowed identification, such as a distinctive tail shape, but sometimes we could not be confident of identity. Sometimes, underlying pelage markings are visible on black cats in infrared (IR) camera trap images (c.f. Hedges et al., 2015), but not clear enough to aid individual identification.
Identification was a notable issue in Strathbogie, where there was a significant number of black cats around Gartly Moor. It was decided that visits to cameras by black cats that looked appreciably the same and were within 3km from each other, were the same cat. This value was chosen because it is the approximate diameter of a circle of 6.36km2, which was found by Campbell (2015) to be the mean 95% MCP (Minimum Convex Polygon) home range of GPS-collared wild-living cats. Individual identifications were updated as further images of those individuals were collected, within and across survey years.
For classification of cats as wildcat, hybrid and domestic, we used the Kitchener et al. (2005) 7PS pelage scoring system, but adopted the following more precautionary categories; <10.5 for a domestic cat; >17 for a wildcat and cats scoring in between classed as hybrid. Following this method, if a cat scored one on any trait then it could not be classed as a wildcat, and a cat scoring two for all features would be classed as a hybrid. In addition, a cat scoring three for any trait, but with a total score <10.5, is classified as a hybrid. Note that this method of classification is based on appearance only and cats scoring either domestic or wildcat on pelage often show genetic evidence of hybridisation (see Senn et al., 2019, and the Genetics and Morphology report). Pelage scores were updated as more data became available from surveys undertaken during the project.
2.3.2 Identification of by-catch
In year one, the Project Officers reviewed all camera trap images and recorded by-catch for each estate or major landowner where this information was requested (most landowners). This activity took a substantial amount of time, and so in subsequent years we asked a small number of dedicated volunteers and students reviewing camera-image sets from the full surveys to record other species that visited the cameras. Where possible this was done to species level, but for some images we were only able to record species groups (e.g. ‘deer’, ‘rodent’, ‘hare’, or even ‘lagomorphs’). This information was also particularly important for volunteer engagement, because the volunteers running the camera traps enjoyed seeing all the species that were captured during the volunteer ‘feedback events’ at the end of each survey season. The images were also used in social media communications. Later in the project we began uploading survey images to MammalWeb to obtain a more consistent approach to identification of by-catch, as well as reducing the time staff spent handling data.
2.3.3 Statistical analyses
We conducted statistical analyses in R (versions 3.5.3 and 4.4.3, R Core Team 2019) and RStudio (version 1.1.463).
2.3.3.1 Population density using SECR
Data from the standardised camera surveys were analysed to estimate population densities. Data available for the analysis were restricted by excluding cameras where metadata (e.g. deployment dates; camera location) were incomplete. Visits by known pet cats were excluded. An encounter was defined as a visit to a camera by an identifiable individual within a 24-hour period, running from noon until noon.
We conducted spatially-explicit-capture-recapture (SECR) analysis using the R package SECR (Efford, 2020) to estimate population densities of wildcats, hybrids and domestic cats in the different Priority Areas. SECR uses data collected from ‘detectors’ (e.g. camera traps, hair traps, live captures etc) from which individuals can be identified to estimate animal abundance by combining a state model (distribution of animal home ranges in the landscape) and an observation model (spatial detection model). The detection model describes the decline in detection probability with distance (d) from the home range centre. We used a half normal detection function:
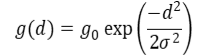
where d is the distance between a detector location and the centre of an animal’s home range, g0 is the baseline encounter probability, and σ (sigma) is the spatial scale parameter of the detection function (or the rate at which detection declines with distance) (Efford, 2015). This formula describes the decaying probability of detecting a cat at a given detector as a function of the distance between the detector location and the centre of an individual’s home range. SECR methods use a mask that consists of a set of systematic points that cover the camera trap grid and the surrounding area that might contain home range centres of an animal (ANNEX 6).
Density is then estimated for each mask point. We created a mask grid with points spaced 250m apart; the grid covered the trapping area plus a 12,000m buffer. The 12,000m buffer was calculated following preliminary SECR analyses of years one to two data that found a minimum buffer of 11,304m for wildcats in Strathbogie (the maximum recommended buffer for any cat-type in any area). The size of the buffer is to ensure any individuals within the mask grid stood a chance of being caught on camera.
Three parameters are modelled in SECR; density/ha (D), detection probability (g0) and a movement parameter, sigma (σ). For all models, we assumed no variation in density across each site (Priority Area: D~1), no difference in movement between individuals (σ~1), and that detection probability (g0) would vary (Table 3). We assessed support for the best model based on AICc value (Akaike Information Criteria, corrected for small sample size). If a model provided obviously erroneous results (very high population estimates or failure to compute confidence intervals), then we removed this model from the candidate list. Model averaging was carried out if two or more models were within two AICc values using the model.average function.
Population estimates, calculated within SECR using N.region, were based on the area of the Priority Area at a cell resolution of 250m. Data were based on 248 detected cats during the standardised surveys, of which 29 were classed as wildcat, 127 as hybrid and 92 as domestic. The number of cats detected during the standardised surveys is provided as MNA (Minimum Number Alive). Results are provided in 3.4.2.
| Model | Description | Note |
|---|---|---|
| Model_1 | g0~1 | Detection (g0) is constant across individuals, occasions and detectors |
| Model_2 | g0~b trap happy/shy | Step change after first detection |
| Model_3 | g0~Cat | Detection varies between cat types |
| Model_4 | g0~Cat + b | Detection varies between cat types and after first detection |
| Model_5 | g0~bk | Detection varies as a result of an individual becoming trap shy/ trap happy after first capture |
Model_6
| g0~Cat + bk
| Detection varies between cat types and as a result of an individual becoming trap shy/ trap happy after first capture |
| Model_7 | g0~t | Detection varies over time |
| Model_8 | g0~Cat + t | Detection varies between cat types and varies over time |
2.3.3.2 Genetic status of cats in Priority Areas
For this we used genetic data from all samples collected within Priority Areas (SWA_DB_06_Snapshot_20190815). The genetic test provides an estimate of the proportion of the genotype that is of wildcat origin (Q), along with a 90% upper and lower confidence boundary for the estimate (UBQ and LBQ respectively, see Senn et al., 2019). We use an LBQ > 0.75 to denote a wildcat and a UBQ < 0.25 to denote a domestic cat. See Senn et al. (2019) and the Genetics and Morphology report for full details of the genetic test. We compared genetic scores of cats sampled in Priority Areas using 187 samples that were from within a Priority Area and from which genetic results have been obtained (Table 4, results are provided in 3.4.3.).
Differences in numbers or focus on capturing cats living in colonies might influence the mean genetic scores for each area. Therefore, we recoded the variable ‘Colony’ to denote cats living alone (0) and cats living gregariously, either ‘few’ or ‘colony’. We ran a General Linear Model (GLM) describing genetic score Q, with Priority Area and colony as predictors. We reclassified two individuals in the dataset from initial assessments as wildcat to hybrid, following a review of their pelage scores and in light of their genetic scores. These were individuals with SWA sample IDs of 232 and 482, both from Strathbogie. Results are provided in 3.4.3.
| Column | Row selection | N rows | Total rows |
|---|---|---|---|
| Initial dataset | - | - | 426 |
| swaPA | All exc nonPA OR Zoo | -91 | 335 |
| Q | All exc NA | -126 | 209 |
| PERCENT_MISSING | Exc profile <50% | -5 | 204 |
| Q_loci | Exc 'fail' | -6 | 198 |
| DateCapt | Exc pre 2016 | -6 | 192 |
| Notes | Mention of pet. Exc | -5 | 187 |
| Notes | Microchip implanted already (assume pet) | -1 | 186 |
| Duplicate record | - | -1 | 185 |
| Updated genetic information | - | +1 | 186 |
| Final sample size | - | - | 186 |
2.3.3.3 Genetic status and distance from habitations
We assessed the genetic status of cats in relation to distance from human habitations (including operational farm buildings), using a Generalised Linear Mixed Models (GLMM). For this we used genetic data from samples taken during TNVR, wildcat live-trapping, and non-invasive sampling, either from accidental capture of cats by members of the public or from hair traps (SWA_DB_06_Snapshot_20190815). We also included samples collected in the Priority Areas from prior research under SWCAP (Littlewood et al., 2014, Campbell, 2015). Road casualties were not included because location information was often insufficiently accurate for the purposes of this analysis. We also excluded samples from outside the Priority Areas. See Table 5 for details of sample selection. Where the sample was recorded as coming from a cat colony (‘Colony’ = “Col”, n=20, or “Few”, n=15), we set the distance to human habitation to 0m. For the remaining records we calculated the straight-line distance from the location of the sample to the nearest inhabited building, defined using OS and aerial photography maps.
None of the pre-project records were from colonies. Two hair samples provided only partial genotype profiles (see the Genetics and Morphology report), both suggesting possible wildcat status (LBQ > 0.75), but with some uncertainty. No other samples tested as wildcat. Most trapping occurred over winter, with different aims in different years. For example, a focus on trapping cats for neutering may invest more effort in trapping cats nearer human habitations than for a season where we focused on trapping wildcats, potentially leading to a bias in different seasons. Therefore we included ‘year’ in the analysis as a random effect, using financial year (‘FY’, 1st April-31st March) to define years. The data included only one individual trapped in the 2019-2020 financial year (September 2019) and so we included this in the prior year. Both Priority Area (PA) and the sex of the individual may influence the relationship between genetic score and distance from habitation and so these were included in the model as random effects. In particular, the Angus Glens Priority Area included no cats with genetic scores of domestic cats and no cats where distance from habitation was 0m. Because the distance data exhibited a skew towards low values, we used ln distance +1m (lnd).
To model the relationship, we used mixed-effects models fitted using Restricted Maximum Likelihood estimation (REML). For step one we initially built a null model with random slopes for PA, FY and sex and compared changes to the random effect structure such as dropping random effects or specifying one or more of PA, FY or sex as random intercepts using AICc. This led to 26 candidate random effects structures. We rejected random models that resulted in singular fits (potentially indicating overfitting) following Matuschek et al. (2017). For step two, using the random effect model with the most support, we then added lnd, comparing this to the null model using AICc. Again, models that resulted in singular fits were rejected and we then compared models using the next most supported random effects structure. To assess the influence of colony cats and those caught closest to buildings, we removed data from individuals where lnd<4 (<54m) and repeated steps one and two above. Statistical analyses were conducted in R (version 3.5.3, R Core Team 2019) using the packages lme4 (Bates et al., 2015) with model selection using MuMIn (Barton, 2019) and significance calculations in lmerTest (Kuznetsova et al., 2017). Results from these are provided in 3.4.4.
| Column | Row selection | N rows | Total rows |
|---|---|---|---|
| Initial dataset | - | - | 426 |
| swaPA (Priority Area) | Exc. nonPA OR Zoo | -91 | 335 |
| GridRef (location) | Exc. Blank OR Unk. | -2 | 333 |
| Q (genetic score) | Exc. NA | -126 | 207 |
| AccKm (accuracy of location in Km) | Exc. values >0.02km | -19 | 188 |
| Scheme | Exc. RTA | -31 | 157 |
| Notes | Exc. Pets | -3 | 154 |
| Notes | Exc. Prior microchip implant (suggests pet) | -1 | 153 |
| Other | Exc. Error in location | -3 | 150 |
| Other | Genetic profile very poor (>50% missing) | -10 | 140 |
| Add one updated genetic profile | - | +1 | 141 |
| Added pre-project data (Littlewood et al., 2014, Campbell, 2015) | - | +19 | 160 |
| Removed individuals duplicated pre-project and SWA | - | -1 | 159 |
| Final dataset | - | - | 159 |
2.3.3.4 Road-casualty cats
We examined overall seasonality in the rate of cats killed on roads that we collected following reports to us or that members of the public handed in. We further investigated seasonality in relation to age-class, sex and genetic status (see Senn et al., 2019) and the Genetics and Morphology report for full details of the genetic test. We determined age-class as adult or juvenile (<1 year), further subdividing this into kittens (younger than approx. four months) or older juveniles for display purposes only. There were too few data to treat these two groups separately during statistical analysis. Sex and age-class were recorded either during collection of the cat or during post mortem. For some individuals, these details were not recorded or recordable, though sex was always established where genetic tests were conducted. Data are from the SWA dataset, see Table 6 for details of sample selection.
To ensure search effort was consistent, we only used cats collected after the project launched in the Priority Areas in April 2015. The last record in this dataset was from April 2019. The total sample included 42 individuals classed as adult (16 female, 22 male, four adults of unspecified sex), nine individuals with no recorded age-class (six male and three with no recorded sex) and 10 juveniles. For statistical analysis of seasonality in road deaths and adult sex, we assumed any individual for which sex was recorded, but age-class was not, that the cat was an adult. This is because there is likely to be a bias towards juveniles in recording age-class (i.e. that the individual was a juvenile is notable).
| Column | Row selection | N rows | Total rows |
|---|---|---|---|
| General analysis | - | Initial | 426 |
| Scheme | RTA or Public only | -363 | 63 |
| Location | Any mention of RTA or found on a road | +12 | 75 |
| Notes | Any mention of RTA or found on a road | +4 | 79 |
| pmCauseDeath | Exc. Shot | -1 | 78 |
| DateCapt | After 31st March 2015 | -17 | 61 |
| Final dataset | - | - | 61 |
| For genetic × season analysis | - | Initial | 61 |
| Q | Exc. NA | -19 | 42 |
| Final dataset | - | - | 42 |
Seasonality in numbers of carcases collected: to test whether there was significant variation between months, we organised the data into cat classes of adults of unknown sex (U adult, including individuals where age-class was not noted), adult females (F adult), adult males (M adult) and juveniles (juv. including kittens). For each cat class and month combination (from Apr 2015 to Apr 2019 inclusive, leading to 196 rows), we noted the number of individuals collected (range zero to four). Month was an ordinal variable and year a factor. Initial analyses indicated that the data structure did not support mixed-effects modelling. We ran a GLM with a Poisson fit, including quadratic terms for month (month2 + month3), terms for year and cat class, and interaction terms for month×year, cat class×month and cat class×year. From this global model, we ran alternative models of all possible combinations of these variables, noting that interaction terms are dependent on their parent variables and quadratic terms are dependent on their lower order equivalents. Model selection was based on AICc using the R package MuMIn. We report below only for models within AICc = 4 (ΔAICc ≤ 4) of the most supported model. The most supported model was assessed for over-dispersion using the R package AER. Results are provided in 3.3.
Seasonality and genetic score: we compared genetic scores (Q, proportion of SNPs that are of wildcat origin) over the year by grouping the data into quarters (Jan-Mar, Apr-Jun, Jul-Sep, Oct-Dec), as there were too few samples in some months to assess by month. We compared the differences between quarters using a GLM with a Gaussian fit.
3. Results
3.1 Public sightings
In total, 769 cat records were received via iRecord between 1st April 2015 and 31st March 2020 (Table 7). These included 29 cat (18 wildcat) records that were sighted prior to the start of the project, of which 17 (eight wildcat) were since 2010, seven (five wildcat) from 2000-2010, two (both wildcat) from the 1990s and three (all wildcat) from the 1980s. Of the 769 cat records, 317 were wildcat records, of which only 45 (14%) were verified. Of the wildcats sighted prior to the project, only four were verifiable. The distribution of the 45 verified wildcat sightings was dominated by records from the north east (Aberdeenshire and Moray), Strathspey, Angus, Inverness-shire and Lochaber (Figure 2).
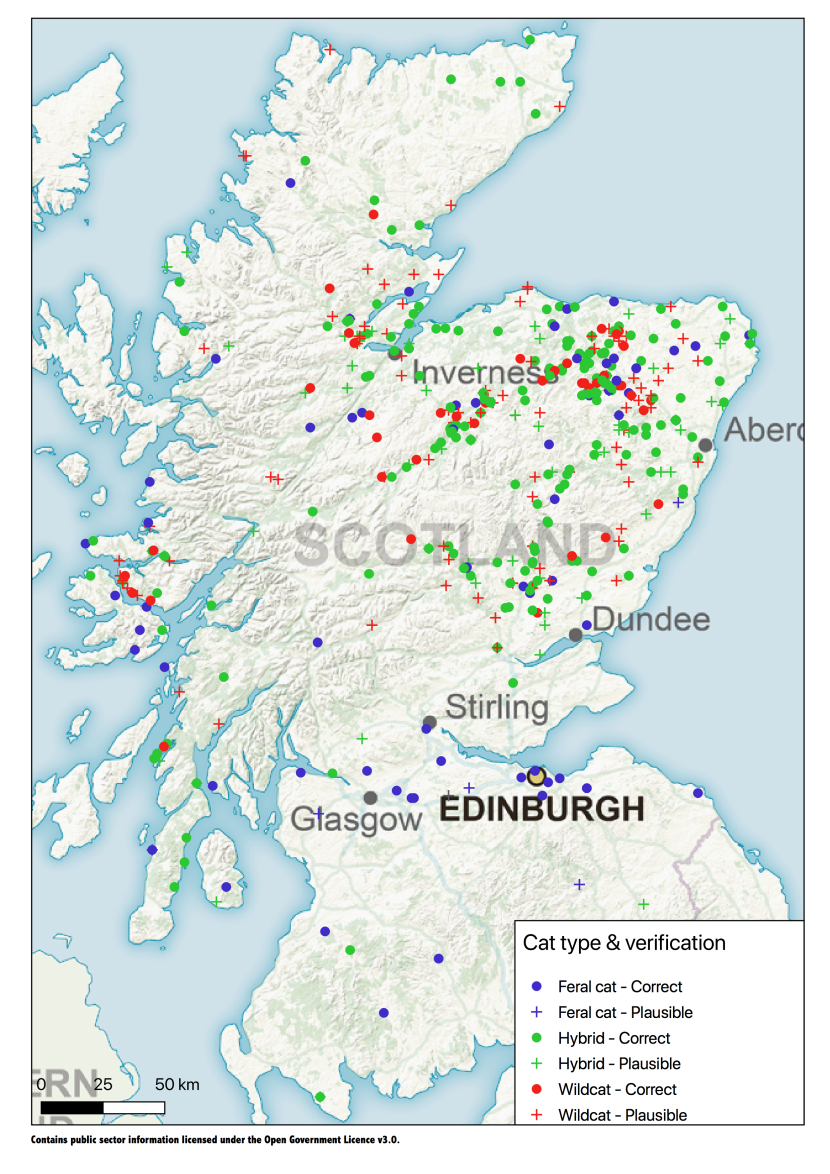
Figure 2. Public sightings of cats submitted to iRecord and verified as correct / considered correct or plausible.
Click for a full description
Plausible records are where the general description suggested the cat could have been a wildcat but no photographs or videos were provided to enable the record to be confirmed. Only records sighted between 1st April 2015 and 31st March 2020 are shown.
A map of mainland Scotland showing the locations of cat records submitted by the public to iRecord. Only records that were verified as ‘correct’, ‘considered correct’ or ‘plausible’ are shown. There is a significant overlap in points, particularly between wildcats and hybrids. Records for both are concentrated in the North East (Aberdeenshire and Angus), Badenoch and Strathspey, Easter Ross and Lochaber (mainly Morvern and Ardnamurchan). Hybrids were also reported from Caithness and Sutherland, but excluding plausible records, no wildcats were recorded north of Lairg.
This could be explained, in part, by the focus of the project on the Priority Areas, but we received a number of sightings of wildcats outside Priority Areas, particularly in the north-east, Angus and Inverness-shire. Notably absent are verified records since 2015 for the far north of Scotland (the most northerly record was from near Lairg) and only one record for the south west (Argyll to the Trossachs). For one third of the wildcat records submitted by the public, we were unable to verify their accuracy with any certainty and these were recorded as ‘plausible’. The distribution of these was similar to the distribution of wildcat records, except there were a few records for northern Perthshire, the far north and the south-west. Verified records of hybrid cats were distributed across the near-historical range of the wildcat (north and west of the Highland Boundary Fault) and outnumbered verified wildcat records nearly 5:1. We received a small number of verified hybrid records outside the former range of the wildcat, notably in Galloway and Mull, that may point to past informal translocations of wildcats or hybrids. Feral cat records were distributed similarly to hybrid records and in addition we received feral cat records outside the near-historical range of the wildcat such as in the central belt of Scotland. Overall, we received far fewer records of feral cats than of hybrids or wildcats, possibly because they are of less note to the public or because they are more often assumed to be domestic pets.
Over the duration of the project, the number of records submitted to iRecord (Figure 3) increased from a very low number (usually one-two / month) to a typical rate of over 10 per month by the second year of the project. There was some variation across the project (e.g. a lull in late 2017 to 2018) that may have been related to the effort we placed on public communications. There was also a notable shift over the project in the proportion of submitted sightings, with wildcats forming a larger portion of the submitted records later in the project. This may have arisen from some of the messaging put out by SWA around the need for wildcat records (more commonly highlighted than the need for records of hybrids), but perhaps also reflected a decline in the clear messages put out by ourselves and others on the extent of hybridisation and the likelihood that a tabby cat sighting in the wild is a hybrid and not a wildcat. There is no reason to believe that the change in proportion here reflects a real change in the population, i.e. there was not a relative increase in wildcats versus hybrids since 2017. Feral cat records declined over the project, again most likely reflecting a shift in how we communicated the importance of submitting records of these cats.
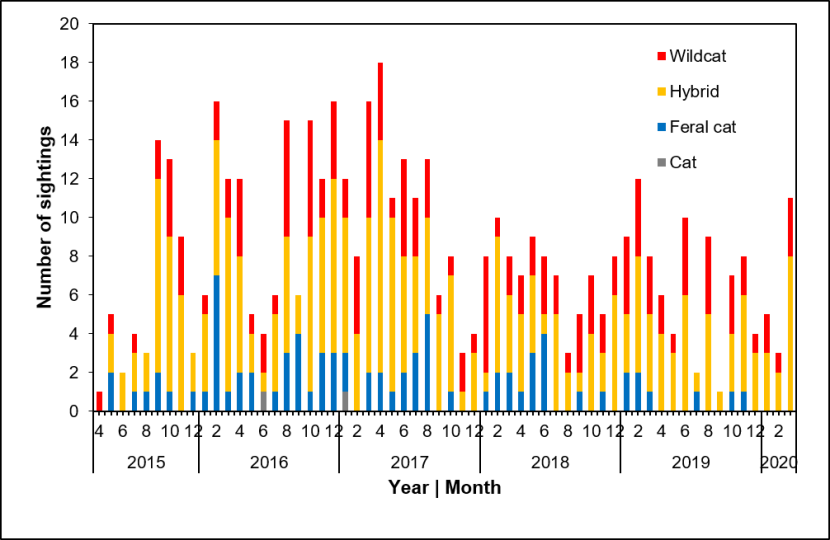
Figure 3. Changes in monthly sightings of cats reported to iRecord over the duration of the project. Note these are dates the cats were sighted, which may be different from the dates the record was submitted.
Click for a full description
Bar chart showing number of sightings of each type of cat during the project. Reporting for all cats increased over 2015, peaked in 2016-2017 and began to decline thereafter. Sightings were dominated by hybrids
| Accepted | - | - | - | - |
|---|---|---|---|---|
| Species | Considered correct | correct | accepted | Total |
| Cat spp. | 1 | . | 1 | 2 |
| Feral cat | 30 | 22 | 21 | 73 |
| Hybrid | 82 | 73 | 49 | 204 |
| Wildcat | 13 | 13 | 19 | 45 |
| Total | 126 | 108 | 90 | 324 |
| Rejected | - | - | - | - |
| Species | Incorrect | Unable to verify | rejected | Total |
| Feral cat | 1 | 3 | . | 4 |
| Hybrid | 18 | 51 | . | 69 |
| Wildcat | 30 | 93 | 3 | 126 |
| Total | 49 | 147 | 3 | 199 |
| Unconfirmed | - | - | - | Grand Total |
| Species | Queried | Plausible | Total | - |
| Cat spp. | . | . | . | 2 |
| Feral cat | 3 | 5 | 8 | 85 |
| Hybrid | 20 | 72 | 92 | 365 |
| Wildcat | 40 | 106 | 146 | 317 |
| Total | 63 | 183 | 246 | 769 |
3.2 Camera surveys other than SWA-led Priority Area surveys
Several organisations, mainly SWA partners and some in collaboration with SWA staff, conducted camera trap surveys totalling over 10,000 camera days of survey effort before and during the SWA project (see Table 8 for details). Survey locations ranged from Balmacara (near Kyle of Lochalsh) in the north west, to Hill of Tarvit (Fife, out-with the known range of the wildcat) in the southeast, Loch Lomond and Trossachs National Park in the southwest and several locations northeast of the Strathbogie Priority Area.
These surveys yielded a total of 34 cats, including two cats confirmed as wildcat based on pelage, one within one of our Priority Areas and one at the edge of one of our Priority Areas. Another cat detected not far from a Priority Area might have been a wildcat, but the images were not sufficient for scoring. All three were from Aberdeenshire or Tayside (Perthshire and Angus), broadly matching the results from public sightings. However, survey effort was low for the north, west and southwest compared with Aberdeenshire and Tayside.
| Organisation | Survey location | Time | No. camera locations (CD) | Results |
|---|---|---|---|---|
| National Trust for Scotland (NTS)* | Balmacara; Ben Lawers; Craigievar Craigower; Crarae; Dunkeld; Glencoe; Hill of Tarvit; House of Dun; Inverewe; Leith Hall; Linn of Tummel; Torridon | Dec 2014 to Dec 2015 | 112+ (2,405+ CD). Note camera effort not available for Crarae, Inverewe, and Torridon | One possible wildcat (within an SWA Priority Area) 12 other cats |
| Forestry and Land Scotland (FLS) | Tayside District forests outside of Angus Glens Priority Area, including Errochty, and Rannoch†‡ | Winter 2014/2015 | 11+ (1,000+ CD) | No confirmed wildcats One unidentified cat |
| Forestry and Land Scotland (FLS) | Strathardle (Kindrogan, Blackcraig and Dalrulzion)† | 2016/2017 | 26 | No wildcats No other cats |
| Forestry and Land Scotland (FLS) | Strathardle (Kindrogan, Blackcraig and Dalrulzion)† | Winter/spring 2018 | 22 (2,618 CD) | One possible wildcat/hybrid Two other cats |
| Forestry and Land Scotland (FLS) | Glen Loy | Jan-Mar 2019 | 11 (308 CD) | No wildcats No other cats |
| Royal Society for the Protection of Birds (RSPB) | Insh Marshes† Abernethy/Craigmore† Ballinlaggan† Loch Ruthven† | Feb-Mar 2016 | 12 (c.360 CD) | No wildcats No other cats |
| Loch Lomond and Trossachs National Park Authority (LLTNPA), FLS, Woodland Trust Scotland, RSPB | Trossachs and Breadalbane | Jan-Mar 2018 Jan-Mar 2019 | 21 (c.1,260 CD) 20 (840+ CD) | No wildcats No other cats |
| SWA / FLS / Universidad de Burgos | Aberdeenshire (Bennachie, Pitfichie, Correen Hills and Corrennie) | Jul - Nov 2018 | 69 (1,641 CD) | One possible wildcat 16 other cats |
| Scottish Woodlands | Loch Arkaig | Autumn/winter 2018 | 7 (c. 900 CD) | No wildcats |
| SWA/RSPB | Abernethy | - | - | No wildcats |
| TOTAL | - | - | 268+ (10,492+ CD) | 2-3 possible wildcats, 31-32 other cats |
* Detecting wildcats was not the sole aim of these surveys.
† These surveys did not follow exact SWA methodologies but methods were similar.
‡ The survey also included forest sites in the Angus Glens Priority Area. However, though wildcats were detected in those sites, not enough information was provided to establish numbers. Therefore, in this report we rely on results from surveys conducted by Kilshaw (2015, Glen Isla) and Scottish Wildcat Action (most areas in the Angus Glens Priority Area, see 3.4.1.1 below).
3.3 Road casualties
We collected a total of 61 road casualties. Based on pelage assessments, these included six wildcats, 43 hybrids and nine domestic cats. Three cats were not ascribed to any category based on pelage. Thirty-three of the cats were recorded as male compared with 18 female (22 adult males and 16 adult females), a ratio of 1.8:1. Sex was not recorded at time of collection or found later from genetic analysis for the remaining 10 specimens. Methods used for data collection are in 2.2.2 and for statistical analyses in 2.3.3.4. Data are available for download.
3.3.1 Seasonality in numbers of carcases collected
A pronounced peak in the number of cats collected (mostly killed on roads) is evident during autumn, with a smaller peak also visible in January (Figure 4 and Figure 5). Monthly averages were highest in 2015 (1.78) and lowest in 2017 and 2018 (1.0). Overall, we collected 1.24 carcases per month.
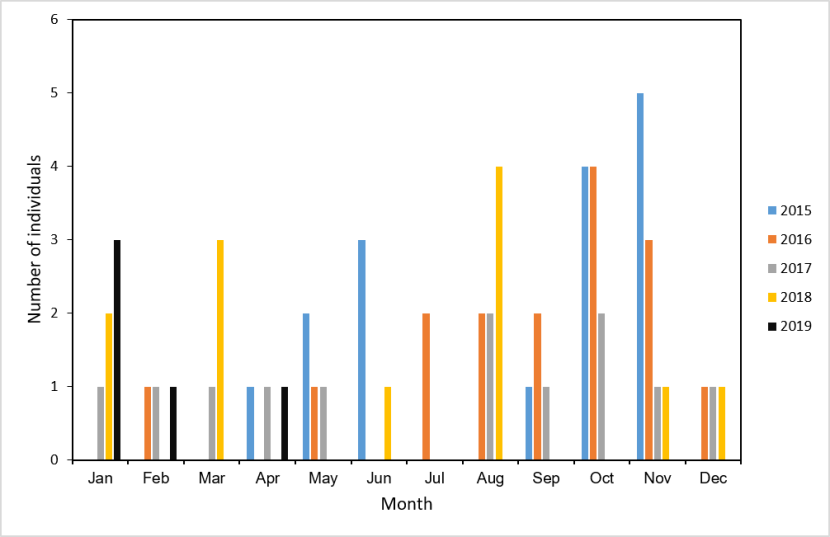
Figure 4. Number of carcases collected by month between April 2015 and April 2019 inclusive.
Click for a full description
Bar chart of the total numbers of cat carcasses collected each month by the project between April 2015 and April 2019. There was some variation between years months.
We found statistical support for seasonality in road casualty rate. The most supported model included the full quadratic term for month and cat class only (Table 9). The next most supported model contained the linear term for month and cat class only (ΔAICc 1.33). Of the top five models within AICc = four, only one retained a cat class×month interaction and none retained year or year×month (Table 9). Therefore, there is an effect of cat class (i.e. some sex / age classes were more frequent than others) and season (more casualties in some months), but no evidence for variation in seasonality between the cat classes or variation between years. This was consistent with the result from the most supported model, where month and cat class were significant (Table 10). The most supported model showed no evidence of over-dispersion in the data (alpha = 0.040, z = 0.458, p=0.324).
To examine whether the low numbers of individuals classed under cat class as ‘U adult’ were influencing these results, we re-ran the above analysis excluding these rows. This did not change the results, with the same model finding the most support (AICc 231.2) and with the next most supported model (ΔAICc 0.84,), including just month as a linear term and cat class, and only three models containing a cat class×month interaction in the seven models with ΔAICc <4 (combined AICc weight 0.254). Examining the most supported model containing the interaction term, there was no evidence of a statistically significant interaction between month and cat class (P(Chisq)=0.30). Again, the most supported model showed no evidence of over-dispersion in the data (alpha = 0.015, z = 0.152, p=0.440).
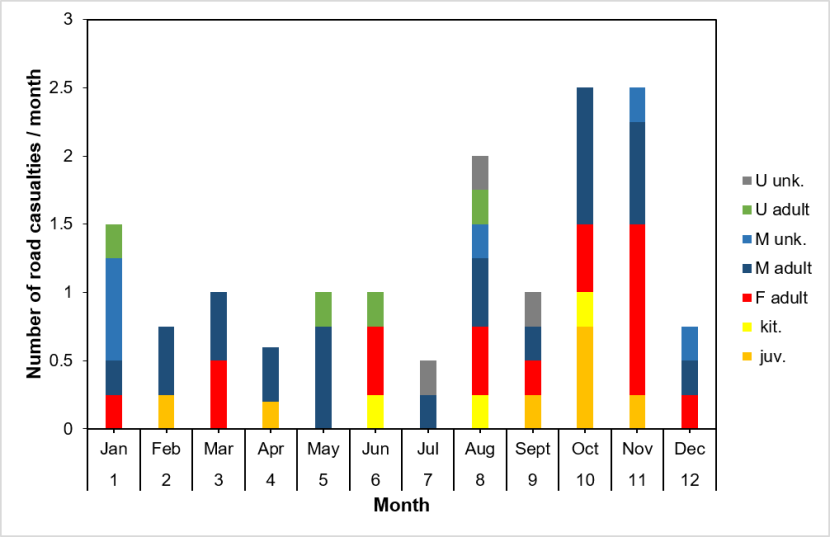
Figure 5. Mean monthly number of carcasses collected over all years, separated by sex (adults only, F = female, M = male, U adult = unknown) and age class (adult, kit. = kitten, juv. = juvenile, U unk. = unknown).
Click for a full description
Bar chart of mean number of cat carcasses collected by the project each month. These are colour coded by sex and age-class. Oct-Nov, Aug and Jan shows the highest mean values. Oct had more juvenile animals collected and Nov more adult females. Overall, males were more frequently collected then females.
| (Int) | Mnth | Mnth2 | Mnth3 | Cat class | class × month | df | logLik | AICc | delta | AICc weight |
|---|---|---|---|---|---|---|---|---|---|---|
| -0.02 | -1.06 | 0.19 | -9.5 e-3 | Y | N | 7 | -130.7 | 276 | 0 | 0.450 |
| -1.56 | 0.07 | N | N | Y | N | 5 | -133.5 | 277 | 1.33 | 0.231 |
| -1.12 | N | N | N | Y | N | 4 | -135.0 | 278 | 2.25 | 0.146 |
| -1.34 | -0.03 | 6.66e-3 | N | Y | N | 6 | -133.3 | 279 | 3.17 | 0.092 |
| -0.76 | -0.95 | 0.19 | -9.5 e-3 | Y | Y | 10 | -129.1 | 279 | 3.43 | 0.081 |
| Predictor | Estimate | SE | Df | Deviance | Resid. Df | Resid. Dev | P(Chi) |
|---|---|---|---|---|---|---|---|
| (Intercept) | -0.019 | 0.731 | - | - | 195 | 183.31 | |
| month | -1.064 | 0.475 | 1 | 3.03 | 194 | 180.29 | 0.082 |
| month2 | 0.195 | 0.083 | 1 | 0.30 | 193 | 179.99 | 0.586 |
| month3 | -0.010 | 0.004 | 1 | 5.32 | 192 | 174.67 | 0.021 |
| class = juv | -0.470 | 0.403 | 3 | 16.22 | 189 | 158.45 | 0.001 |
| class = M adult | 0.560 | 0.313 | 3 | 16.22 | 189 | 158.45 | 0.001 |
| class = U adult | -0.827 | 0.453 | 3 | 16.22 | 189 | 158.45 | 0.001 |
3.3.2 Seasonality in genetic scores
There was an apparent increase in the genetic score of road-casualty cats in Apr-Jun (Figure 6). Using all data, we found no evidence of a significant difference in genetic scores between quarters (F 1.833, df 3, 38, P=0.158). However, there was a notable outlier (Q=0.017) in the Apr-Jun cats, with all the other cats from those months exhibiting genetic scores from 0.545 to 0.673. Excluding that individual, there were significantly higher genetic scores (estimate 0.19 ± 0.09SE) for cats killed on the roads during Apr-Jun (F 3.915, df 3, 37, P=0.016). Sample sizes are small for the Apr-Jun period (total eight cats) and therefore these results should be interpreted with caution.
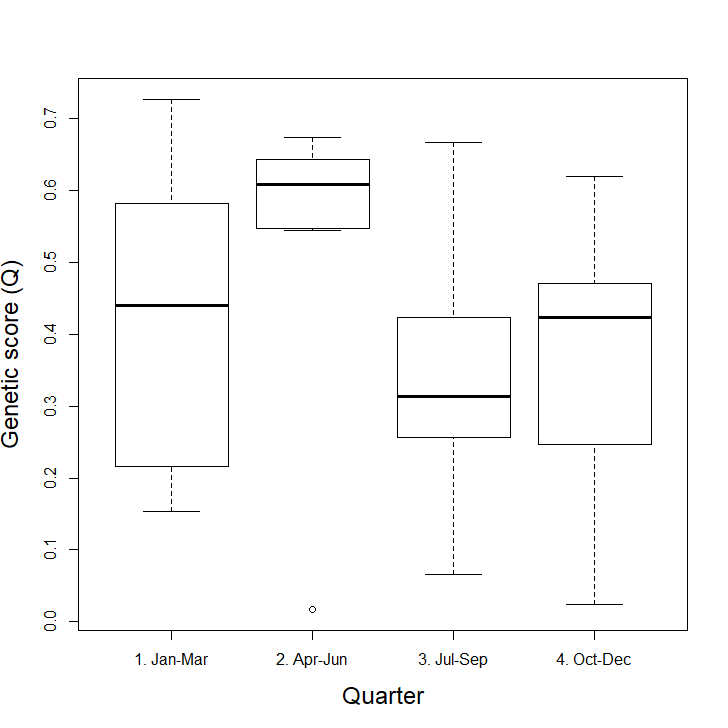
Figure 6. Means and interquartiles of the genetic scores of cats killed on roads during each quarter.
Click for a full description
Box and whisker plot of the genetic scores of cats killed on roads during each quarter. Man genetic score was around 0.30 to 0.45 for three of the quarters but higher (0.6) in Apr-Jun, with the interquartiles suggesting this different could be significant.
3.4 Cat populations in Priority Areas
3.4.1 Detected cats
Encounter histories (years of the project in which the cat was seen) for cats that were neither pets nor associated with a single farm in each area are provided in ANNEX 7 and are also available for download along with the photographic catalogues.
3.4.1.1 Angus Glens
We detected a total of 102 cats in years one to five (see Table 11), of which 61 were detected during standardised surveys in years one and three (Figure 7), 67 individual + year combinations. An additional seven individual cats in total were detected outside the standardised survey period in both years (five of them in year one and three in year three), either on camera or captured in cage traps. The remaining 33 individuals were detected in other years. During the standardised surveys, four wildcats were detected in the year one survey and five in the year three survey, alongside 20 and 38 non-wildcats respectively. This gives a ratio in the two standardised surveys of 6.0 and 8.6 (mean 7.6) non-wildcats for every wildcat (noting here that our definition of a wildcat is based on a pelage threshold of 17 and does not include the genetic score). Over the two standardised surveys 3/18 (17%) domestic, 3/34 (9%) hybrids and 0/9 (0%) wildcats were detected in both years. Two additional wildcats (both male) were detected outside the standardised surveys, both in year five. Therefore the total number of wildcats detected during the project was 11.
3.4.1.2 Morvern
We detected a total of 21 cats (Table 11). Of these, 11 cats, including two wildcats (both male) were detected during the year two standardised survey (Figure 7). During the course of our work in subsequent years we detected one additional female wildcat, although we were concerned we may have been missing another wildcat or hybrid that produced kittens at one location in two of the years. The total number of detected wildcats in Morvern was therefore three. All cats with domestic cat phenotypes detected on survey cameras were associated with settlements, and almost all turned out to be pet cats or known feral cats that were being fed regularly. All wildcats and (with one exception) all non-wildcats were found on the southwest of the peninsula, along with the primary human settlements and the main area of coastal woodland, furthest from the northeast land bridge connecting the peninsula to the mainland. Despite following the female wildcat closely after year two (she often returned to the garden of a local volunteer) and her being tagged with a GPS collar in 2018, we never detected evidence of breeding.
One of the male wildcats had also been recorded during the scoping survey (Littlewood et al., 2014), but was found dead and underweight in May 2018. The other male wildcat was almost certainly breeding with one-three hybrid females distributed along the southwest coast, as stripy kittens were seen along the road in this location every year (although never caught on camera trap, despite intensive efforts). One kitten was caught, tested and found to be a hybrid (he was subsequently socialised to people and rehomed). One female hybrid collared during 2014 (see Campbell, 2015) was still occupying the survey area in 2019 and probably breeding with the wildcat male.
3.4.1.3 Strathavon
No wildcats were detected during the standardised survey in year one, with only 12 cats detected in total (Figure 7; Table 11). Follow-up targeted surveys during the following winter included new locations but still failed to detect any wildcats. Two potential wildcats or hybrids were live-trapped in a gamekeeper’s trap at the edge of the Priority Area in 2016. Hair samples were taken for genetic testing before release, but no full pelage score is available for either individual due to the difficulty of scoring a cat in a trap, with estimated scores for both being 16-17. Neither genetic test result confirmed these as wildcats, although they were the two with the highest genetic scores from the cats we sampled in Strathavon (see Figure 13).
3.4.1.4 Strathbogie
A total of 104 cats were detected over the project (Table 11), 52 of which were detected during the standardised surveys in years one and three (Figure 7, 63 individual + year combinations). Five wildcats were detected in both year one and year three surveys, alongside 22 and 31 non-wildcats respectively. However, different individuals were detected in each survey with only 10% of wildcats (1/10), 20% domestic (4/20) and 26% of hybrids (6/23) detected in either survey were detected in both years. All but 13 of cats new to year three occupied locations within 1.25km of the known ranges of cats seen in year one, indicating recruitment and turnover. Of those 13 cats, six were at locations not surveyed in year one. One additional wildcat was detected outside the standardised surveys and so the total number of detected wildcats was 10.
3.4.1.5 Strathpeffer
We detected a total of 66 cats over the first three years (Table 11), 47 of which were detected during the standardised PA surveys in years one and three (Figure 7, 52 individual + year combinations). During the standardised surveys five wildcats were detected in year one and none in year three, alongside 17 and 30 non-wildcats respectively. Over these two surveys only 13% (3/23) domestic and 11% (2/19) hybrids were detected in both surveys. No additional wildcats were detected outside the full surveys and therefore the total number of wildcats detected was five.
3.4.1.6 Strathspey
We detected a total of 48 cats over the first three years (Table 11), 46 of which were detected during the standardised PA surveys in years one and three (Figure 7, 47 individual + year combinations). During the standardised PA survey, only one wildcat was detected in year one and two in year three, alongside 12 and 32 non-wildcats respectively. One wildcat was detected in both surveys (50%), but none of the 15 domestic and 29 hybrid cats were detected in both years. No additional wildcats were detected outside the full surveys and therefore the total number of wildcats detected was two.
| Pelage type | AN | MO | SA | SB | SP | SS | Total |
|---|---|---|---|---|---|---|---|
| wildcat | 11 | 3 | 0 | 10 | 5 | 2 | 31 |
| hybrid | 49 | 8 | 6 | 50 | 21 | 31 | 165 |
| domestic cat | 31 | 10 | 11 | 44 | 37 | 15 | 148 |
| uncertain | 11 | 0 | 0 | 0 | 1 | 0 | 12 |
| Total | 102 | 21 | 17 | 104 | 64 | 48 | 356 |
3.4.1.7 Overview
Overall we invested 50,644 camera trap days in full surveys in the first three years of the project and a further 35,755 camera trap days during our other work (Table 2), detecting 356 individual cats, including 31 wildcats (Table 11). The Priority Area mean number of camera trap days per individual cat was 323 (range 170-699), and 3,342 (range 1,578-4,893) camera trap days per wildcat. Variation in the total numbers of cats detected between the Priority Areas in part reflects variation in the effort expended in monitoring (Figure 8). Just considering the standardised surveys, there was a survey mean of 270 (range 73-917, total 201) camera days per individual cat and 2,032 (range 632-5,043, total 1,746) per individual wildcat (the equivalent to running 34 cameras for a full 60-day survey). During the standardised surveys, higher numbers of cats were not related to greater survey effort (Figure 9).
Focusing only on the four Priority Areas surveyed in both years one and three, we see no clear trend in the numbers of domestic cats or wildcats; with a mean of nine domestic cats and three wildcats detected per Priority Area (though within areas there were fluctuations, see above and Figure 7). There were a small number of individuals we were unable to reliably classify as hybrid or wildcat from camera trap images; all of these were in the year three surveys. This also occurred during the year one survey, but we were able to obtain more images to accurately score pelage in subsequent surveys (see 2.3.1). However, the number of detected hybrid cats increased in all areas (see Figure 7) from a mean of nine to 19, leading to an overall increase in the mean number of cats of all types from 18 to 34 individuals per Priority Area. Our survey effort did not increase between these surveys (total 18,841 camera days in year one and 17,070 days in year three) and so we detected individual cats in fewer camera days (119 days per cat in year three versus 209 in year one). Of the 219 cats detected in the four areas we surveyed in years one and three, 23 (9.1%) were detected in both years.
The total number of cats detected in each Priority Area during all types of survey was related positively to the survey effort (Figure 8). However, this was not the case for cat numbers detected during within standardised surveys (Figure 9), suggesting that the effect of survey effort on cat numbers was largely due to recruitment and immigration into the local populations outside the standardised survey periods.
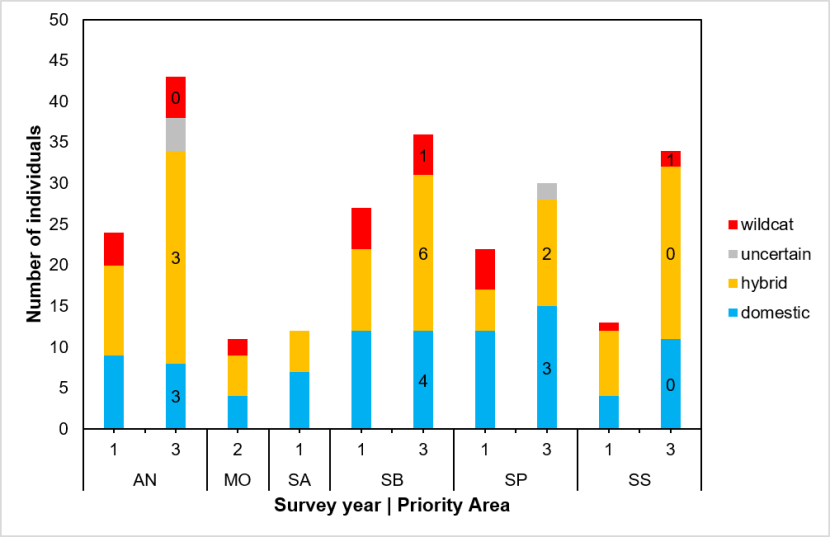
Figure 7. Number of individual cats detected during standardised surveys across the Priority Areas. First surveys were conducted in years one or two and repeated in four areas in year three.
Click for a full description
Numbers in the columns indicate the number of individuals that were re-sighted from the previous full survey. Priority Area codes are provided in Table 2. Bar chart showing the number of individual cats detected during standardised surveys across the Priority Areas. Numbers detected ranged from 11 to 43. More cats were detected in the second survey, where conducted. Hybrids and domestic cats clearly outnumbered wildcats (as defined by pelage score). Morvern and Strathavon had the fewest cats detected, with no wildcats detected in Strathavon.
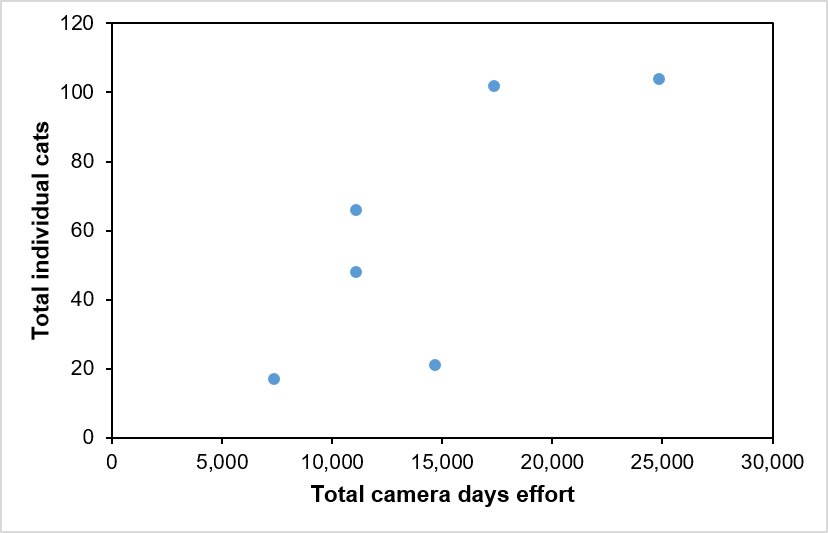
Figure 8. Total number of individual cats detected over the project duration versus total camera effort (camera days) summed across all years in each Priority Area.
Click for a full description
A scatter plot showing the total number of individual cats detected over the project duration versus total camera effort (camera days) summed across all years in each Priority Area. Though there was some variation, in general the greater the camera effort, the greater the number of cats detected.
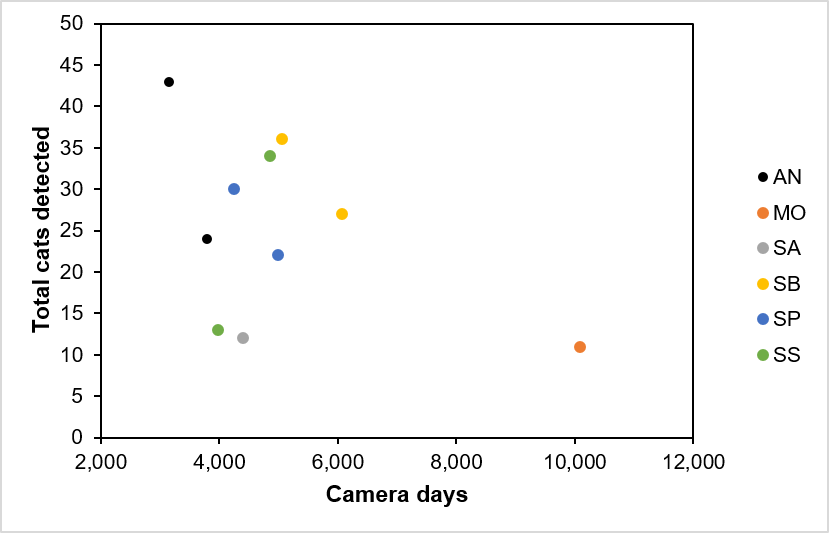
Figure 9. Number of individual cats detected versus survey effort within each standardised survey in each Priority Area.
Click for a full description
A scatter plot showing the number of individual cats detected versus survey effort within each standardised survey in each Priority Area, colour coded by Priority Area. The one standardised survey in Morvern shows the greatest effort but the fewest cats. In the remaining Priority Areas, there was no clear pattern between effort and numbers of cats detected.
3.4.2 Population estimates and SECR results
Methods are described in 2.3.3.1. The mean number of camera traps used per survey was 81 (range = 51-148) and the mean camera trap spacing was 966m (range = 610-1463m) (Table 12). A version of the survey data with coordinates ‘desensitised’ by convertion to zero mean and random rotation are available to download.
For all sites and cat types, Model_1 (D~1, g0~1, σ ~1) or Model_3 (D~1, g0~Cat Type, σ ~1) were the most supported model (Table 13). Results from the SECR analysis are shown in Table 13 and summarised below.
| Area | AN | AN | MO | SA | SS | SS | SB | SB | SP | SP |
|---|---|---|---|---|---|---|---|---|---|---|
| Year | 1 | 3 | 2 | 1 | 1 | 3 | 1 | 3 | 1 | 3 |
| Cameras | 63 | 51 | 142 | 60 | 69 | 93 | 80 | 84 | 82 | 91 |
| Mean camera spacing (m) | 900 | 1463 | 885 | 1209.3 | 899 | 610 | 1138 | 991 | 858 | 712 |
| Mask area (km2) | 1,618 | 2,108 | 1,031 | 1,470 | 1,358 | 1,358 | 1,547 | 1,421 | 1,356 | 1,476 |
| Detections | 142 | 200 | 228 | 38 | 51 | 315 | 249 | 340 | 118 | 90 |
| MNA (All cats) | 24 | 43 | 11 | 12 | 13 | 34 | 27 | 36 | 22 | 30 |
| PA size (km2) | 372 | 372 | 483 | 343 | 205 | 205 | 347 | 347 | 325 | 325 |
| PA | Year | Cat type | g0 | σ/m | D/ha | s.e. | LCL | UCL | D/km2 | Top Model |
|---|---|---|---|---|---|---|---|---|---|---|
| AN | 1 | All | 0.0547 | 979.49 | 0.0013 | 0.0004 | 0.0008 | 0.0022 | 0.129 | 3 |
| AN | 1 | Wildcat | - | - | - | - | - | - | - | - |
| AN | 1 | W+H | 0.1116 | 969.06 | 0.0009 | 0.0003 | 0.0004 | 0.0018 | 0.089 | 3 |
| AN | 1 | Hybrid | 0.1105 | 980.47 | 0.0005 | 0.0001 | 0.0003 | 0.0008 | 0.045 | 1 |
| AN | 1 | Domestic | 0.0508 | 1016.48 | 0.0004 | 0.0001 | 0.0002 | 0.0008 | 0.04 | 1 |
| AN | 3 | All* | 0.0285 | 1945.17 | 0.0007 | 0.0001 | 0.0005 | 0.001 | 0.072 | 3 |
| AN | 3 | Wildcat | 0.0312 | 4542.68 | 0 | 0 | 0 | 0.0001 | 0.004 | 1 |
| AN | 3 | W+H | 0.0309 | 2112.75 | 0.0005 | 0.0001 | 0.0004 | 0.0008 | 0.053 | 3 |
| AN | 3 | Hybrid | 0.0761 | 1013.64 | 0.0008 | 0.0002 | 0.0005 | 0.0011 | 0.077 | 1 |
| AN | 3 | Domestic | 0.1762 | 530.42 | 0.0004 | 0.0002 | 0.0002 | 0.0009 | 0.042 | 1 |
| MO | 2 | All | 0.061 | 1693.13 | 0.0002 | 0.0001 | 0.0001 | 0.0003 | 0.019 | 3 |
| MO | 2 | Wildcat | - | - | - | - | - | - | - | - |
| MO | 2 | W+H | 0.0858 | 1722.32 | 0.0001 | 0 | 0.0001 | 0.0002 | 0.012 | 1 |
| MO | 2 | Hybrid | - | - | - | - | - | - | - | - |
| MO | 2 | Domestic | - | - | - | - | - | - | - | - |
| SA | 1 | All | 0.0497 | 467.03 | 0.001 | 0.0003 | 0.0005 | 0.0019 | 0.099 | 1 |
| SA | 1 | Wildcat | - | - | - | - | - | - | - | - |
| SA | 1 | W+H | - | - | - | - | - | - | - | - |
| SA | 1 | Hybrid | - | - | - | - | - | - | - | - |
| SA | 1 | Domestic | - | - | - | - | - | - | - | - |
| SS | 1 | All | 0.0165 | 427.38 | 0.0009 | 0.0002 | 0.0005 | 0.0015 | 0.089 | 3 |
| SS | 1 | Wildcat | - | - | - | - | - | - | - | - |
| SS | 1 | W+H | 0.1504 | 434.34 | 0.0006 | 0.0002 | 0.0003 | 0.0012 | 0.061 | 3 |
| SS | 1 | Hybrid | 0.2887 | 254.73 | 0.001 | 0.0004 | 0.0005 | 0.0022 | 0.104 | 1 |
| SS | 1 | Domestic | - | - | - | - | - | - | - | - |
| SS | 3 | All | 0.2311 | 668.03 | 0.0015 | 0.0003 | 0.0011 | 0.0022 | 0.154 | 1 |
| SS | 3 | Wildcat | - | - | - | - | - | - | - | - |
| SS | 3 | W+H | 0.2219 | 706.98 | 0.0011 | 0.0002 | 0.0007 | 0.0016 | 0.108 | 1 |
| SS | 3 | Hybrid | 0.2311 | 668.01 | 0.0015 | 0.0003 | 0.0011 | 0.0022 | 0.154 | 1 |
| SS | 3 | Domestic | 0.175 | 466.78 | 0.0006 | 0.0002 | 0.0003 | 0.0012 | 0.062 | 1 |
| SB | 1 | All* | 0.1982 | 937.91 | 0.0009 | 0.0002 | 0.0006 | 0.0013 | 0.087 | 3 |
| SB | 1 | Wildcat | 0.0348 | 1568.42 | 0.0001 | 0 | 0 | 0.0002 | 0.011 | 3 |
| SB | 1 | W+H | 0.0187 | 1406.54 | 0.0004 | 0.0001 | 0.0002 | 0.0006 | 0.037 | 3 |
| SB | 1 | Hybrid | 0.0247 | 1196.91 | 0.0003 | 0.0001 | 0.0001 | 0.0005 | 0.026 | 1 |
| SB | 1 | Domestic | 0.4924 | 556.26 | 0.0004 | 0.0001 | 0.0003 | 0.0008 | 0.045 | 1 |
| SB | 3 | All | 0.0758 | 1094.24 | 0.0011 | 0.0002 | 0.0008 | 0.0016 | 0.113 | 1 |
| SB | 3 | Wildcat | 0.0178 | 1631.65 | 0.0001 | 0.0001 | 0.0001 | 0.0003 | 0.014 | 1 |
| SB | 3 | W+H | 0.0689 | 1290.33 | 0.0007 | 0.0001 | 0.0005 | 0.001 | 0.069 | 3 |
| SB | 3 | Hybrid | 0.0983 | 1215.79 | 0.0005 | 0.0001 | 0.0004 | 0.0008 | 0.054 | 1 |
| SB | 3 | Domestic | 0.4604 | 376.45 | 0.0007 | 0.0002 | 0.0004 | 0.0012 | 0.068 | 1 |
| SP | 1 | All | 0.0813 | 545.62 | 0.0014 | 0.0004 | 0.0009 | 0.0023 | 0.14 | 3 |
| SP | 1 | Wildcat | - | - | - | - | - | - | - | - |
| SP | 1 | W+H | 0.1276 | 472.11 | 0.0007 | 0.0002 | 0.0003 | 0.0013 | 0.333 | 3 |
| SP | 1 | Hybrid | 0.326 | 426.99 | 0.0003 | 0.0001 | 0.0001 | 0.0007 | 0.028 | 1 |
| SP | 1 | Domestic | 0.0557 | 642.6 | 0.0006 | 0.0002 | 0.0004 | 0.0011 | 0.063 | 1 |
| SP | 3 | All* | 0.0529 | 485.12 | 0.0017 | 0.0003 | 0.0012 | 0.0025 | 0.174 | 3 |
| SP | 3 | Wildcat | - | - | - | - | - | - | - | - |
| SP | 3 | W+H | - | - | - | - | - | - | - | - |
| SP | 3 | Hybrid | 0.1039 | 502.55 | 0.0008 | 0.0002 | 0.0004 | 0.0014 | 0.078 | 1 |
| SP | 3 | Domestic | 0.1003 | 435.48 | 0.0011 | 0.0003 | 0.0006 | 0.0018 | 0.109 | 1 |
*additional cats included in the ALL cats analysis but could not be identified to type so were excluded from other analysis
3.4.2.1 All cats
To obtain population size estimates with confidence, a minimum of 20 individuals and 30 recaptures were used. This means population size estimates for all cats across some of the PAs could be calculated but not all areas (Table 14). Mean population size for these PAs was 45 wild-living cats (±12).
| PA | Survey Year | Population Size estimate (N) | s.e | lcl | ucl | MNA | Recaptures |
|---|---|---|---|---|---|---|---|
| Angus | 1 | 42 | 8.96 | 30.88 | 69.12 | 24 | 142 |
| Strathbogie | 1 | 31 | 1.35 | 29.27 | 34.75 | 27 | 249 |
| Strathpeffer | 1 | 48 | 9.59 | 35.13 | 74.40 | 21 | 228 |
| Strathpeffer | 3 | 60 | 8.52 | 47.97 | 82.34 | 30 | 88 |
Mean density of all cats across all Priority Areas was 0.11 cats / km2 ± 0.04 (Table 13). Density estimates varied significantly between cat types (One-Way ANOVA; F = 20.051, d.f = 2, p= 0.0003), with wildcats found at a significantly lower density than hybrids across all Priority Areas (Tukeys Test p = 0.03; see Figure 10)
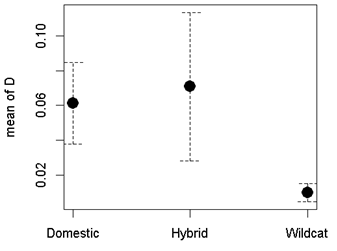
Figure 10. Means plot of density/km2 (D) of different cat types as classified by pelage across all PAs, dotted lines show standard deviation of the mean D.
Click for a full description
Graph showing the mean and standard deviation (SD) of density (individuals per km2) of domestic, hybrid and wildcats. Domestic and hybrid cats show a similar mean density (0.06-0.07/km2), though the SD for hybrids is greater. Wildcats show a lower density (mean of approximately 0.01km2) with SD limits that do not cross those of the other cat types.
However, density estimates/km2 for all cats combined per PA did not differ significantly between Priority Areas (Kruskal-Wallis Test for small sample size used; chi-squared = 5.5636, df = 5, p-value = 0.35).
The mean movement parameter (σ) for all cats was 924m ± 529.06m and although it varied between Priority Areas (Table 13) with cats in Strathavon, Strathpefffer and Strathspey all having lower mean movement parameters than the other PAs, this difference was not significant (Kruskal-Wallis chi-squared = 7.3636, df = 5, p = 0.2).
However σ did vary between cat types (One-way ANOVA; F = 6.5355, d.f. =2, p = 0.04), with wildcats having a larger mean σ than hybrids (Tukeys Test p = 0.005) and domestic cats (Tukeys Test p = 0.001) (see Figure 11. and Table 13). In particular, σ for all cats in Strathspey year one (427m), Strathavon (467m) and Strathpeffer (545m and 485m for years one and three respectively) was much smaller than the mean distance between camera traps (899m; 1209m; 858m and 712m for Strathspey year one, Strathavon and Strathpeffer year one and year three respectively) suggesting there was a chance not all individuals within these areas may have been detected.
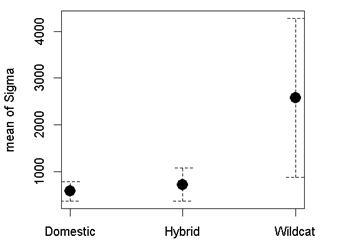
Figure 11. Mean sigma (σ) moved (m) by different cat types across all the PAs and survey years. Wildcats move significantly further than hybrids or domestic cats.
Click for a full description
Graph showing the mean and standard deviation (SD) of sigma (σ) moved (m) of domestic, hybrid and wildcats. Domestic and hybrid cats show a similar mean sigma (approximately 500), though the SD for hybrids is slightly greater. Wildcats show a much higher sigma (mean 2,580), with SD limits that do not cross those of domestic cats.
Overall the detection rate (g0) for all cats across all Priority Areas was 0.08 ± 0.07 and did not vary significantly between Priority Areas (Kruskal-Wallis chi-squared = 3, df = 5, p= 0.07) but did between cat type (One-way ANOVA; F = 12.939, d.f. = 2, p= 0.002) with wildcats less likely to be detected than hybrids (Tukeys Test p = 0.04) or domestic cats (Tukeys Test p = 0.02) (see Figure 12).
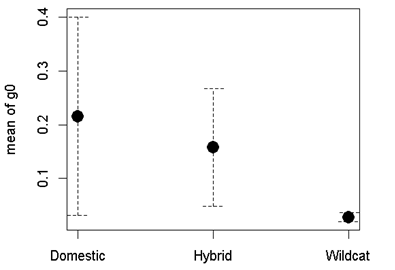
Figure 12. Mean detection probability (g0) of different cat types across all the PAs and survey years. Wildcats were significantly less likely to be detected than hybrids or domestic cats.
Click for a full description
Graph showing the mean and standard deviation (SD) of detection probability (g0) for domestic, hybrid and wildcats. Domestic and hybrid cats show a similar detection probability (0.15-0.25), though the mean and SD for domestic cats is slightly greater. Wildcats show a much lower detection probability (mean 0.03), with SD limits that do not cross those of hybrids.
3.4.2.2 Wildcats
There was an insufficient number of wildcats and wildcat recaptures to estimate reliably a population size for wildcats in any of the PAs. No wildcats were detected at all in Strathavon or in Strathpeffer in year three. The mean Minimum Number Alive across all PAs was 2.9 wildcats (±2; range = 1-5 wildcats) with a mean density of 0.01 (± 0.005) wildcats/km2. Mean σ for wildcat was 2,580m ± 1,387m and mean g0 was 0.03 ± 0.009.
3.4.2.3 Hybrids
Mean detectability of hybrids was 0.15 ± 0.19 and did not vary significantly between different PAs (Kruskal-Wallis chi-squared = 4.5, df = 3, p = 0.21). The mean Minimum Number Alive across all PAs was 14.4 hybrids (±10.2; range = 1-5 wildcats) with a mean density of 0.07 (± 0.04) hybrids/km2. Density did not vary significantly between PAs (Kruskal-Wallis chi-squared = 4.5, df = 3, p = 0.21).
Only one PA had a sufficient number of hybrids and recaptures to estimate population size with a good degree of confidence; in year three there was an estimated population size of 28.69 hybrids (s.e. = 53.88, 95%CI 19.28 – 42.70) in the Angus Glens. The movement parameter σ for hybrids was 782m ± 368 but didn’t vary significantly between different PAs (Kruskal-Wallis chi-squared = 6, df = 3, p = 0.11).
3.4.2.4 Domestic cats
The average MNA of domestic cats across the Priority Areas was 9 ± 3 cats with a mean density of 0.06 ± 0.02 cats / km2. Density estimates did not vary greatly between PAs (Kruskal-Wallis chi-squared = 4.5, df = 3, p = 0.21).
One PA had a sufficient number of domestic cats and recaptures to estimate population size with a good degree of confidence; in Strathbogie, year three, there was an estimated population size of 23.68 hybrids (s.e. = 5.02, 95%CI 17.22 – 38.19). Mean detectability of domestics was 0.21 ± 0.28 and did not vary significantly between different PAs (Kruskal-Wallis chi-squared = 0.32143, df = 3, p= 0.9559). Mean σ for domestic cats was 574m ± 213m (Table 13) and did not differ significantly between PAs (Kruskal-Wallis chi-squared = 6, df = 3, p = 0.11).
3.4.3 Genetic status of cats
Methods are provided in 2.3.3.2. Data are available for download. From live-trapping, 160 samples were collected within Priority Areas, alongside 12 hair and two saliva samples through non-invasive sampling, 62 samples from outside the project (‘public’, mainly from stray and feral cats) and 34 road casualty cats (total = 270, Table 15). Two hair samples gave partial profiles (80% and 58% of a full profile) with LBQ above the 0.75 threshold, suggesting possible wildcat. However, full samples are needed to confirm these results. Two other partial profiles from hair samples showed high estimates (Q = 0.87 and 0.92) but LBQ fell below 0.75 (0.47 and 0.65 respectively), reflecting considerable uncertainty around these results and again highlighting the importance of full profiles from better/more complete samples. These latter two samples are not included here due to the unreliability of the genetic scores. Overall, 38% of hair samples (n = 5) collected using lure posts failed to produce a profile better than 50%, and all faecal samples (n = 10) failed to produce any profile. Both saliva samples obtained from the cotton of valerian root pouches provided profiles. One fifth of samples (n = 80) had not been processed at the time of analysis (April 2020), though genetic results processed subsequent to this analysis have not changed the overall pattern nor confirmed any samples as from a wildcat.
From the samples tested from Priority Areas that yielded full genotype profiles, all Priority Areas showed similar patterns of hybridisation (Figure 13), with any differences likely due to variation in total sampling effort or different focus of cat trapping. For example, no cats from the Angus Glens tested as domestic cat (LBQ < 0.25), although this was one of the only Priority Areas (along with Strathavon) where none of the sampled cats was recorded as living gregariously. The genetic score of colony cats was typically lower than other cats, but otherwise there was no clear difference in mean genetic scores of cats between Priority Areas (Table 16, Figure 14). The GLM of genetic score in relation to Priority Area and colony status confirmed this, with the most supported model containing both terms (AICc=-88.2) and the next most supported model, without colony, showing much less support (ΔAICc=40.3). There was no effect of PA (Type II SS ANOVA, F = 1.46, df = 6,179, P = 0.194) but a significant effect of colony was found (Estimate = -0.256 ± 0.035SE, F = 54.96, df = 1,179, P<0.001). The relationship between cat type (pelage score) and genetic score is explored further in the Genetics and Morphology report.
| Source | AG | MO | SA | SB | SP | SS | PA total | Non-PA | Total | Failed | Not run |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Public | 3 | 6 | 7 | 32 | 8 | 6 | 62 | 33 | 95 | 8 | 24 |
| RTA | 6 | 3 | - | 2 | 15 | 8 | 34 | 28 | 62 | 0 | 19 |
| SWA_Survey | - | - | 1 | 13 | - | - | 14 | 11 | 25 | 15 | 0 |
| Faecal | - | - | - | - | - | - | 0 | 10 | 10 | 10 | 0 |
| Hair | - | - | 1 | 11 | - | - | 12 | 1 | 13 | 5 | 0 |
| Saliva | - | - | - | 2 | - | - | 2 | - | 2 | 0 | 0 |
| TNVR | 37 | 7 | 2 | 109 | 24 | 23 | 202 | 5 | 207 | 0 | 34 |
| Blood | 37 | 7 | 2 | 49 | 23 | 20 | 138 | 5 | 143 | - | 32 |
| DeadCat | - | - | - | - | 1 | - | 1 | - | 1 | - | 0 |
| Hair | - | - | - | 2 | - | 3 | 5 | - | 5 | - | 2 |
| Not sampled | - | - | - | 58 | - | - | 58 | - | 58 | - | - |
| WildcatTrap | 2 | 3 | - | 8 | - | 3 | 16 | 1 | 17 | 0 | 3 |
| MouthSwab | - | - | - | - | - | 1 | 1 | - | 1 | - | 0 |
| Blood | 2 | 3 | - | 8 | - | 2 | 15 | 1 | 16 | - | 3 |
| Total | 48 | 19 | 10 | 164 | 47 | 40 | 328 | 78 | 406 | 23 | 80 |
| Priority Area | Lone | Lone | Lone | Colony | Colony | Colony | All | All |
|---|---|---|---|---|---|---|---|---|
| - | Q | SD | N | Q | SD | N | Q | SD |
| Angus Glens | 0.477 | 0.142 | 37 | - | - | 0 | 0.477 | 0.142 |
| Morvern | 0.471 | 0.139 | 6 | 0.217 | 0.208 | 7 | 0.334 | 0.217 |
| Strathavon | 0.268 | 0.257 | 6 | - | - | 0 | 0.268 | 0.257 |
| Strathbogie | 0.461 | 0.217 | 58 | 0.093 | 0.101 | 22 | 0.360 | 0.253 |
| Strathpeffer | 0.378 | 0.179 | 24 | 0.252 | 0.153 | 9 | 0.343 | 0.179 |
| Strathspey | 0.399 | 0.185 | 13 | 0.364 | 0.121 | 5 | 0.389 | 0.167 |
| All areas | 0.438 | 0.194 | 144 | 0.178 | 0.163 | 43 | 0.378 | 0.216 |
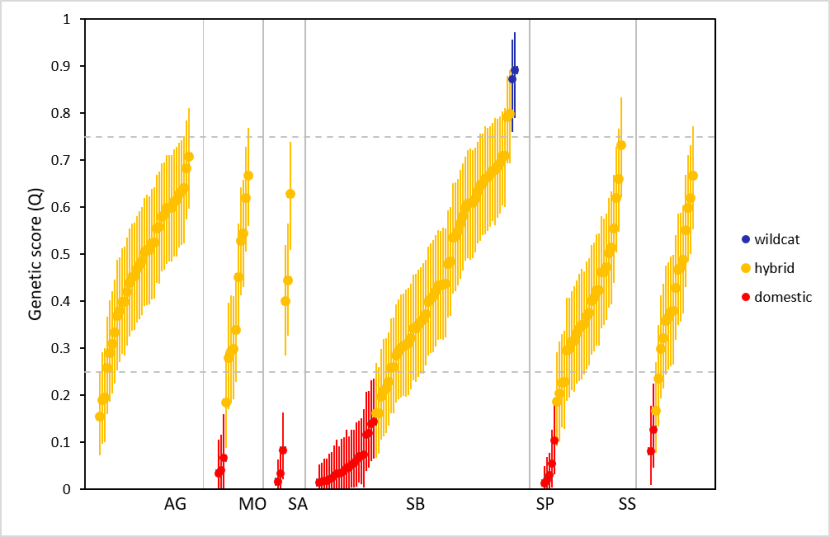
Figure 13. Genetic scores of cats in the Priority Areas (Angus Glens AG; Morvern MO; Strathavon SA; Strathbogie SB; Strathpeffer SP; Strathspey SS).
Click for a full description
Samples are from all sources, including TNVR of wild-living and farm-living cats, wildcat live-capture, non-invasive sampling and samples sourced from outside the project. Dashed horizontal line at 0.25 indicates the UBQ boundary between domestic and hybrid genetic classes, and 0.75 indicates the LBQ boundary between hybrid and wildcat genetic classes. Note that the two individuals with genetic scores indicative of wildcat are partial profiles from non-invasive samples and cannot be considered definitive. We found no statistical differences in the mean genetic scores between Priority Areas.
A plot of genetic scores (Q) with lower and upper confidence boundaries for cats sampled in the Priority Areas. Data are organised by Priority Area and then from low to high Q scores. Most samples were collected from Strathbogie. Only two cats had genetic scores that were above the threshold for wildcat, though both were partial profiles and so not definitive. In all areas except the Angus Glens and excluding the two possible wildcat genetic scores, data show a spectrum of genetic scores from domestic cat to hybrids with Q values at approximately 0.7. The Angus Glens showed a similar pattern except no cats with genetic scores indicating domestic cat were sampled.
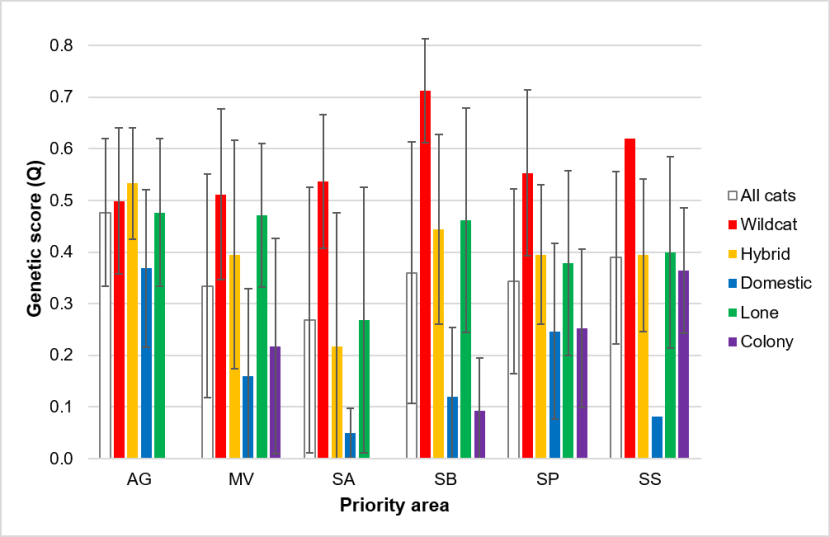
Figure 14. Means with standard deviations of genetic scores for cats in each Priority Area classed as wildcat, hybrid or domestic feral from pelage, and as living alone or in a colony.
Click for a full description
Note that two of the nine wildcats sampled in Strathbogie yielded partial profiles which may inflate the results in that area. Excluding these two samples, the mean genetic score for wildcats in Strathbogie was 0.66 ± 0.03 SD.
A bar chart showing the mean with standard deviations of genetic scores (Q) for cats in each Priority Area classed as wildcat, hybrid or domestic feral from pelage, and as living alone or in a colony. In most cases, the mean Q for wildcats was slightly higher than for hybrids, which were in turn higher than for domestic cats. The only exception to this was the Angus Glens where Q scores for wildcats and hybrids were very similar. Cats living alone tended to have higher Q scores than those living in colonies.
3.4.4 Genetic status and distance from habitations
Details on the methods used in this analysis are provided in 2.3.3.3. Examining the influence of Priority Area, sex and year on the recorded genetic score, the model with random intercepts only for Priority Area (PA) and financial year (fyear) was the most supported (AICc = -46.7). The next most supported model contained a random intercept for fyear only and had a ΔAICc = 1.8. The third most supporting contained random intercepts for fyear and sex and had a ΔAICc = 2.2 indicating only limited evidence for a difference between the sexes in genetic scores. Adding ln distance from nearest inhabited building (lnd) resulted in an AICc = -93.3 (ΔAICc = 46.36), indicating strong support for an effect of distance. The relationship between distance and genetic score was significant (Table 17).
Excluding individuals captured closest to buildings (<54m), a random slope for fyear had the most support (AICc = -45.7). However, adding lnd as a fixed effect led to a singular fit. Using the next most supported random effects structure (random intercept for fyear, AICc = -43.9), a model with lnd as a fixed effect had more support (AICc = -47.6, ΔAICc = 3.7). The relationship between distance and genetic score remained significant (Table 17).
Thus there is evidence for an increase in genetic score with increasing distance from inhabited buildings (Figure 15), though there is considerable variation around it. This pattern is driven by the existence of both a group of genetically low-scoring cats living around inhabited buildings (78% in colonies or small groups), and a group of relatively high-scoring cats (Q~c.0.60) living more than c.400m from inhabited buildings (lnd>6).
| - | - | Estimate | SE | DF | F | P |
|---|---|---|---|---|---|---|
| a) All | Intercept | 0.261 | 0.048 | - | - | - |
| a) All | Ind | 0.05 | 0.006 | 1,117 | 71.63 | <0.001 |
| b) Ind>4 | Intercept | 0.172 | 0.103 | - | - | - |
| b) Ind>4 | Ind | 0.061 | 0.017 | 1,82 | 12.99 | <0.001 |
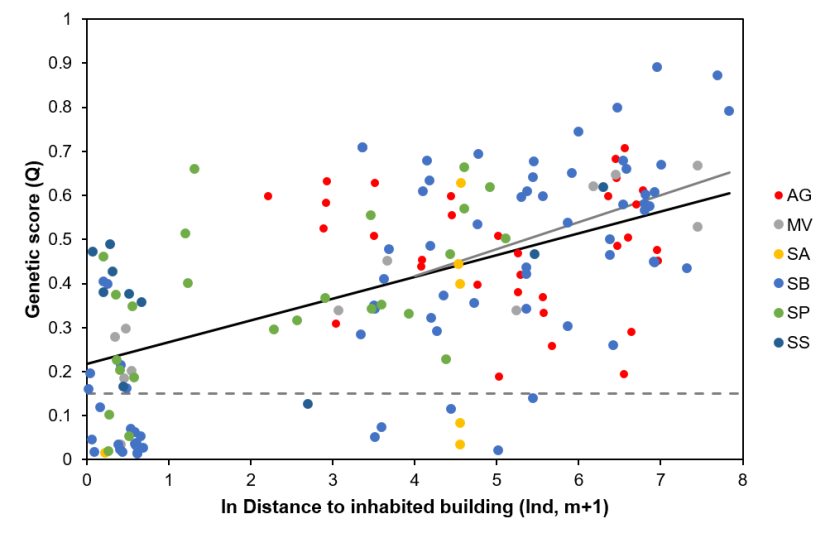
Figure 15. The relationship between genetic score and distance (on a log scale) to the nearest habitation. Data are coloured differently for each Priority Area.
Click for a full description
Data points are present with jitter (0<x<1m) along the x axis to separate overlapping points at 0m. The black fitted line was calculated from the most supported mixed-effects model using all data and the solid grey fitted line from the most supported model using data only from individuals trapped over 54m (lnd>4) from buildings. Note that data points under the dashed grey line (Q<0.15) all scored as domestic cat (UBQ<0.25). The furthest we captured a domestic cat from an inhabited building was 229m (lnd=5.44), while the furthest for a cat of any type was 2.5km.
A scatter plot showing distance to inhabited building (on a log scale) on the X axis and genetic score of cats on the Y axis. Points are colour coded by Priority Area. The data show a weak but significant positive relationship between the two variables. This pattern is visible for all Priority Areas except the Angus Glens, where no cats were sampled next to inhabited buildings, nor were any cats with very low genetic scores indicative of domestic cat.
4. Discussion
4.1 Cat populations within Priority Areas
4.1.1 Status of wildcats in Priority Areas
Based on pelage assessments alone, overall we detected too few wildcats in any of the Priority Areas to constitute viable populations, with a mean of 2.9 wildcats (±2; range = 1-5 wildcats) found in a Priority Area in any one year. In the Strathavon Priority Area we did not detect any wildcats over two survey years. In contrast, we detected significant numbers of hybrids and domestic cats in all Priority Areas. From the total number of individuals detected (Minimum Number Alive) during full PA surveys in all areas, domestic and hybrid cats outnumbered wildcats by 7:1 and the SECR analysis also indicated that wildcats were found at significantly less densities than either hybrids or domestic cats. Detection probabilities were much lower for wildcats (0.03 ± 0.009) in Scotland than have been noted for wildcats in other countries using camera traps. For example, in Sicily, Anile et al. (2014) had a detection probability for wildcats as 0.14 ± 0.03, although this is likely a reflection of the much lower wildcat densities in Scotland rather than a difference in applying the methodology, because detection probabilities of all cats in this study were similar (0.08 ± 0.07) and mean camera trap spacing was comparable (966 ± 78.5m this study vs 759.9 ± 24.5 m in Sicily).
Overall, wildcats as assessed by pelage exist in the Priority Areas at a mean density of 0.01 (± 0.005) wildcats/km2 (one wildcat per 100km2). This is lower than population density estimates reported by other recent studies for wildcats across Europe and in Scotland (see Table 13). But it should be noted that all these studies have used slightly different methodologies and definitions of a wildcat, which makes comparisons between studies complicated. Indeed, across the range of the European wildcat, different definitions of how to define a wildcat exist but steps to standardise this are currently in discussion between researchers.
| Country | Density/km2 | Method | Source |
|---|---|---|---|
| Germany | 0.45 | Live trapping | Strief et al., 2016 |
| Switzerland | 0.25 | Hair lures | Kery et al., 2011 |
| Switzerland | 0.2-0.5 | Radio tracking | Herrmann & Vogel 2005 |
| Switzerland | 0.35 | Hair lures | (Weber, 2008) |
| France | 0.32 | Radio tracking, genetics | Beugin et al. 2016 |
| Polish Carpathians | 0.1 | Snow tracking, observations, pelts | (Okarma et al., 2002a) |
| Spain | 0.07 | Camera trapping | (Gil-Sánchez et al., 2020) |
| Sicily | 0.28 | Camera trapping | (Anile et al., 2012) |
| Sicily | 0.32 | Camera trapping | (Anile et al., 2014) |
| Turkey | 0.22 | Camera trapping | (Can et al., 2011) |
| NE Scotland | 0.3 | Radio tracking | (Corbett, 1979) |
| W Scotland | 0.01 | Radio tracking | (Scott et al., 1993) |
Nonetheless, wildcat densities in the Priority Areas are extremely low, and even densities of all cats captured in the PAs can be considered low in general at a mean of 0.11 cats / km2 ± 0.04. This indicates that some factor or combination of factors may be influencing population densities of wild-living cats in Scotland. Carbone and Gittleman (2002) showed that prey density is a fundamental determinant of carnivore density both within and between species. When Corbett (1979) carried out his radio tracking study in Glen Tanar, Scotland in the mid-1970s he reported a wildcat density of 0.3 wildcats / km2 (or 30 wildcats/100 km2),which is comparable to recent estimates from Sicily, Switzerland, Germany and Turkey (see Table 18). He also reports that rabbits were the preferred prey (92% occurrence in wildcat scats analysed) and that rabbits were abundant. Later camera trap surveys (Hetherington and Campbell, 2012), however, detected no wildcats on Glen Tanar, but also no rabbits, which had been practically wiped out due to disease. Rabbit populations in Scotland have shown a huge decline since the 1950s when myxomatosis arrived (Aebischer et al., 2011). They had started to make some recovery but further declined as a result of rabbit haemorrhagic disease (RHD), with rabbit bags in Scotland decreasing 85% between 1995 and 2009 (Aebischer et al., 2011). Local decreases in wildcat populations were observed in the Strathspey PA by Hetherington and Campbell (2012) from previously higher numbers (Kilshaw et al. 2010). This decline was attributed to a severe crash in the local rabbit population as a result of extremely heavy and prolonged snow cover in the winters of 2009/2010 and 2010/2011. Across the PAs, areas with rabbits (e.g. Angus Glens, Strathbogie and Strathpeffer) had higher wildcat densities and cat captures and many studies have noted the importance of rabbits for wildcat populations (e.g. Corbett, 1979, Lozano et al., 2006, Delahay et al., 1998, Anile et al., 2012). In addition to disease and persecution/control, rabbit densities can be negatively affected by the abundance of red deer, Cervus elephus (Lozano et al., 2007).
Low densities of rabbits may limit the abundance of wildcat populations in multiple environments at a regional scale, although wildcats are also able to maintain high abundances in areas where rabbits are absent by eating rodents (Lozano et al., 2007, Malo et al., 2004, Moleon and Gil Sanchez, 2003, Lozano et al., 2006). Rodents can exist in high numbers and diversity if the habitat is suitable, but rodent densities are also negatively affected by red deer (Smit et al., 2001, Flowerdew and Ellwood, 2001) and can be greatly influenced by forest structure and management practices (Ecke et al., 2001, Michał and Rafał, 2014, Suchomel et al., 2012). It is possible that the current landscape in Scotland, with highly fragmented woodland largely dominated by conifer plantations, low prey diversity and low densities of some key prey species in some areas, in combination with predator control and dramatic fluctuations in rabbit numbers, has had a negative impact on wildcat densities. See also Anile and Devillard (2020) for an interesting insight into the variability in population density of smaller felids more generally. We recommend that further research be carried out on the carrying capacity of different habitat types within the Scottish highlands, in order to identify steps that could be taken to increase local prey population densities to the benefit of the wildcat. For example, a lot of research has been carried out on how rabbit management could benefit reintroduced Iberian lynx (Lynx pardinus) populations including habitat management, building artificial warrens and rabbit restocking (e.g. Ferreira and Delibes-Mateos, 2011).
While the number of wildcats in each area is low, the mean total population sizes of all wild-living cats of 45±12 per Priority Area (as estimated by SECR where possible) are close to the target set out in the scoping survey (Littlewood et al., 2014) of 40 individuals per Priority Area for most areas. However, evidence from both the camera trap data and the GPS data (Ecology report) show that wild-living cats, including wildcats, show a strong tendency to utilise the edges of forest (where there is more prey such as rabbits and small mammals) rather than stay exclusively within the coniferous forest. Riparian areas within forests as well as edge habitats often provide a higher diversity and abundance of small prey mammals than interior forest (e.g. Doyle, 1990, Gomez and Anthony, 1998, Osborne, 2005). Unfortunately, forest edges across the PAs are in general close to farms and rural human populations, putting wildcats in these areas at high risk of further hybridisation (as is evidenced by the high number of hybrids) and exposure to disease (see the Disease Surveillance report). This highlights the need for the neutering and vaccinating of surrounding domestic cats and hybrids (see the Trap Neuter Vaccinate Return Programme report).
Studies from Europe have shown that wildcats reach their highest densities in broadleaved, highly structured forests compared to coniferous plantations (Parent, 1975, Schauenberg, 1981, Stahl and Artois, 1994), which are considered a marginal habitat for wildcats (Heptner and Sludskii, 1972). For example, wildcats appear to prefer areas of windthrow and young succession stages with dense undergrowth and avoid more mature coniferous stands (Okarma et al., 2002b, Hötzel et al., 2007). Mixed habitats that provide a mixture of good prey and shelter are also important (Stahl et al., 1988, Wittmer, 2001, Biro et al., 2004, Klar et al., 2008, Lozano et al., 2003, Jerosch et al., 2017). Forest cover appears to be particularly important for female wildcats (Beugin et al., 2016, Oliveira et al., 2018). In addition to extensive neutering therefore, we recommend further investigation into how coniferous plantations could be improved to the benefit of female wildcats in particular. For example, studies in European forests have shown that small changes in forest management can lead to big changes in small mammal diversity and abundance (e.g. Suchomel et al., 2012, Krojerová-Prokešová et al., 2016, Gasperini et al., 2016) in particular retaining scrub and woody debris (Krojerová-Prokešová et al., 2016, Converse et al., 2006) and retaining areas of wind throw for wildcats. Stahl and Artois (1995) noted that “maintenance of a vertical stratification of vegetation is important” for wildcats, including the development of shrubby habitat that humans can’t get into and creation of patches of stands of different ages.
Results from genetic testing revealed evidence of a complete hybrid swarm in all Priority Areas so that, with the exception of two partial genetic profiles, all phenotypic wildcats tested displayed genetic scores below the defined threshold (see 3.4.3). The results from pelage scores and genetic testing are consistent with a hybrid-swarm population structure, with virtually no cats at the ‘wildcat’ end of the spectrum for both pelage and genetic scores. In three of our Priority Areas (Angus Glens, Strathbogie and Strathspey), despite the low number and some evidence of turnover in individuals (probably from mortality and recruitment), we noted no overall decline in the number of wildcats (see 4.1.2 below). It is highly probable that cats assessed from pelage as wildcats are breeding freely with other cats and the individual wildcats recruited into the populations during the project are cats with wildcat-like phenotypes drawn from a larger population that includes more obviously hybridised individuals.
However, we did find some evidence that wildcats (as classified by pelage) moved further (mean = 2580m ± 1387m) than hybrids (mean =782m ± 368m) and domestic cats (mean = 574m ± 213m). This might be linked to these cats living further from settlements where a reduced or zero reliance on human-sourced food requires larger home ranges.
4.1.2 Changes in the Priority Area cat populations during the project
As mentioned above, wildcat numbers (as assessed from pelage) remained stable in the Angus Glens, Strathbogie and Strathspey, despite some turnover of individuals. In Strathpeffer, the initial five wildcats detected during the first survey were not detected two years later and no new wildcats were detected either. See 4.2.1.5 (below) for further discussion on this.
We observed a general increase in the densities of hybrid individuals between the repeat surveys in years one and three. This increase may reflect a genuine increase in the cat population, or an improvement in our survey efficiency. Volunteers may have got better at camera placement during the survey and we changed (and probably improved) the scent lure used between these years. In addition, volunteers were more likely to move their cameras if they had not detected a cat in the previous year, positively biasing detection probability. It is also possible that the increased numbers of cats with hybrid pelage may reflect the neutering of the most obviously hybridised or domestic phenotypes earlier on, meaning that remaining wildcats or those with scores close to 17 were more likely to breed. However, two years may not be long enough to see any visible changes in the cat population from the activities of the project. We observed more variation in the change in domestic cat numbers, increasing in two areas and declining in others. Some of this might arise from variation in our pelage scoring for individuals that are borderline between hybrid and domestic. Given that the genetic results show almost all wild-living cats are actually hybrid, we can consider treating wild-living domestic and hybrid cats together.
4.2 Priority Area specific comments
4.2.1.1 Angus Glens
The Angus Glens yielded relatively high numbers of wildcats and wild-living cats, potentially as a result of the high density of rabbits found across some parts of this PA. In the second standardised survey in particular, the overall number of cats (mainly hybrids) caught on camera almost doubled. However, the two standardised surveys were not repeated at all the same locations due to changes in access and volunteers between the surveys, so the difference in figures should be treated with caution. There was a large increase in the mean spacing between camera traps between year one and year three (900m – 1463m) resulting in a large increase in the overall area surveyed. Detection probability also decreased from 0.05 in year one to 0.02 in year three, highlighting how changes in distance between camera traps/size of area surveyed could have an impact on detection of cats (e.g. Newey et al., 2015).
We noticed a relatively high proportion of the cats were spottier (a high proportion of broken stripes and more spots on the flanks) and scored close to the 17 pelage threshold, making management decisions on whether to neuter or not difficult. The Angus Glens were unusual in that there was no difference in genetic score for cats that scored as wildcat or hybrid from pelage, indicating that pelage is a particularly poor indicator of introgression status here. There appears to be a higher proportion of cats that are wild-living in the area (in contrast to farm colony cats, for example), but this may reflect a bias in the surveyed area. There are reports of considerable feral/farm cat issues just to the south of the Priority Area (E. Stirling, pers. com) and local gamekeepers have noted regular issues with pet cats being abandoned at the foot of the Glens, possibly from the relatively nearby larger cities and towns.
4.2.1.2 Morvern
We detected little change in the wildcat population in Morvern. Although only one full survey was conducted, all of the cats were found to occupy the same area (the southwest coastal edge of the peninsula), and hence targeted surveys could effectively monitor this section of the PA for years three, four and five. Several of the cats also regularly visited local volunteers, where they have been fed for many years, and so their progress could be monitored. The southwest coastal strip is the most forested area of Morvern, with a large proportion of deciduous coastal hazel woodland. It is also the area with largest coverage of forestry plantations. Almost all the human population live in the southwest, between the villages of Lochaline and Drimnin, and hence this is where most of the domestic cats can be found. The climate in the southwestern corner is generally warmer and drier than most other parts of the Morvern peninsula. However, we know from recent historical records and specimens that wildcats did used to occupy other lowland areas across Morvern. Rabbits were also historically present in Morvern but populations were reduced in 1978 due to myxomatosis (Hewson, 1984) and were thought to have been wiped out 20+ years ago (pers. comm., M. Wilson, 2020). Rabbit populations seem to be starting to make a very small recovery in parts of Morvern (pers. comm., M. Wilson, 2020). In addition to the lack of rabbits, cats in Morvern were being controlled throughout the 1990s and 2000s. Breeding was detected in at least two years (stripy kittens are regularly seen on the road between Lochaline and Drimnin), but not from the one known female wildcat. The one remaining male wildcat probably bred with several hybrid females (including one that was trapped and GPS-collared in 2014; see Campbell, 2015) living along the southwestern edge of the peninsula.
Given the extremely low numbers of wildcats, their age (all were adults in 2016) and the remoteness of individuals from other wildcats, any future successful breeding seems unlikely and it is probable that all individuals are closely related. There were also few hybrid and feral domestic cats. It is not possible to state whether the low total numbers of wild-living cats are due to the carrying capacity of the area or some other factor (such as a low number of cats to begin with, or high infant mortality), but changes in habitat management could potentially improve the carrying capacity. Such changes could include the reintroduction of potential prey species currently absent, such as rabbits or water vole Arvicola amphibius, planting of deciduous scrub and woodland, and reducing Molinia in grassland and moorland to improve habitat for voles.
4.2.1.3 Strathavon
Strathavon was the only Priority Area selected where earlier scoping surveys (Littlewood et al., 2014) did not detect any cat classified as a wildcat using the Kitchener et al. (2005) strict classification (pelage score ≥19). Only one cat detected during that scoping survey had a pelage score of 17. Therefore, the area was the least supported of the six areas for in situ conservation of wildcats. Our subsequent failure to detect definitively any other wildcats with a pelage score ≥17 in the first two years of this project confirmed this. Two possible wildcats were trapped by a gamekeeper, but we were unable to obtain a definitive pelage score and subsequent genetic results were below the genetic threshold. Following the second survey, we reallocated staff resources to our work in Strathbogie.
All cats caught on camera in Strathavon appeared to be moving much less than in other PAs (mean σ = 467m). Sollmann et al. (2012) recommended trap distances be less than 2σ. Mean camera trap spacing in Strathavon was 1470m so in this case it seems possible that some cats were being missed by the camera trap layout.
4.2.1.4 Strathbogie
Strathbogie had the most staff resources and a very committed volunteer team. In addition, the area contained a higher proportion of land under management by a project partner and a higher proportion of colony-living farm cats than other areas. These factors together are reflected in the higher and more consistent survey coverage and large number of cats trapped and sampled, including some of the highest genetic scores recorded in the Priority Areas (though no score from a full genetic profile exceeded the threshold for wildcats). Strathbogie cats classified as wildcat from pelage had among the highest ranging behaviour (sigma) and lowest detectability (g0) estimated from SECR analysis, suggesting they had large home ranges (which is supported by the GPS data; see the Ecology report). Most wildcats were found in and around the periphery of the Clashindarroch Forest managed by Forestry and Land Scotland, though one male (Bogie) was living entirely in farmland (see the Ecology report ). Because surveys were relatively consistent in terms of effort and locations covered, we were able to track better occupancy of locations (posited home ranges) by individuals in some areas. From this we identified replacement of individuals by new cats on a few occasions. Therefore, in Strathbogie we can be more confident that changes in individuals between the full surveys two years apart are partly because of turnover of individuals in the population and not wholly from the low detectability of wildcats.
4.2.1.5 Strathpeffer
In Strathpeffer there was an abrupt decline in the number of wildcats detected between the two standardised PA surveys in year one and year three, from five to zero individuals. One wildcat was killed on the road, but the reason for not detecting the remaining individuals is uncertain. The probability of detection (g0) was low for all cats in Strathpeffer over year one and year three, but not as low as in the Angus Glens, although cats in Strathpeffer had quite low σ (mean = 515m). Camera trap spacing was also quite small during both standardised surveys (mean = 785m) suggesting cats stood a good chance of being caught on camera. However, cats in this PA also had low re-capture rates, generally visiting the camera once or twice and then never again, suggesting a highly ephemeral population with individuals moving in and out of the surveyed areas. The habitat itself is very diverse landscape with lots of hedgerows and movement corridors, so the likelihood of placing a camera on exactly the right path was quite low. There was also a local decline in rabbit populations between the two survey periods due to severe weather conditions, which is very likely to have had an effect on local wildcats, but possibly less so on feral domestic cats and hybrids, which may utilise human based resources more readily. In addition, since more cats were detected relatively close to habitations than for most other areas (e.g. see Figure 15), cats of all types were possibly gaining sufficient food in winter from human hand-outs and therefore were less interested in our bait.
4.2.1.6 Strathspey
Strathspey had few wildcats, indicating that the population has not recovered from the decline tracked within part of the Priority Area by Hetherington and Campbell (2012) from previously higher numbers (Kilshaw et al. 2010, see 4.1.1). We redetected very few cats between the two standardised surveys in year one and year three. As with the Angus Glens, access permission varied between survey years as did the volunteers involved. Of all the Priority Areas, Strathspey contains the lowest proportion of land managed by organisations partnered to the project and so also has potentially the lowest consistency in access. Therefore, the lack of redetection is due in part to surveys not covering all the same locations in both years, but it may also reflect population turnover.
4.3 Overall status of the wildcat in Scotland
4.3.1.1 Previous wildcat distribution
Past surveys of wildcats in Scotland have suggested they occupy areas north of the Highland Boundary Fault, excluding islands (Easterbee et al., 1991, Balharry and Daniels, 1998, Davies and Gray, 2010). These surveys were based on game bag records, cat carcases collected from roads, live-capture, interviews and public sightings. Generally evident from these surveys is also a lack of records from the tip of north-eastern Aberdeenshire around Peterhead, and fewer records running in a northwest-southeast line from around Kyle of Lochalsh, through Fort William and Rannoch Moor to Stirling. This latter pattern may have arisen from observer bias due to lower human population density. Davies and Gray's (2010) assessment, collected from 2006 to 2008 and based on public sightings and a questionnaire survey, also showed a gap in records around Caithness inland from the north coast. The most recent assessment by the Mammal Society (Mathews et al., 2018), modelled from public sightings and other records collected between 1995 and 2016, therefore overlapped with Davies and Gray (2010). Their distribution estimates match this historical range, including an apparent gap in distribution from Kyle to Stirling (again possibly from observer bias and survey effort, see for example Figure 16) and the more recent gap in Caithness.
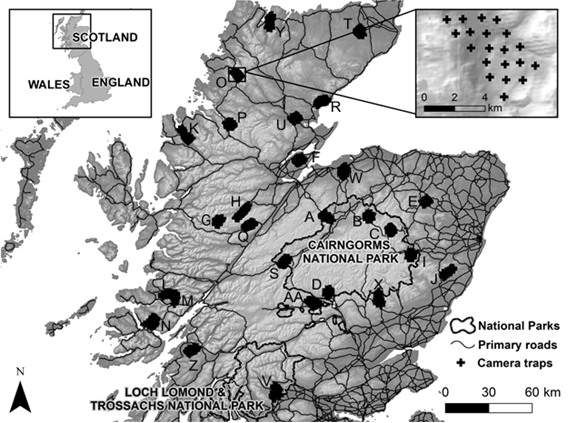
Figure 16. Intensive (60-70 days using 40 cameras at 20 points) baited camera trap survey locations, set between 2010 and 2013. Figure from Kilshaw (2015), reproduced with permission.
Click for a full description
A map of northern mainland Scotland showing 26 locations where intensive camera-trap surveys were conducted by Kilshaw (2015). These show locations spread across northern Scotland within the range of the wildcat. There is an inset showing an example of the spread of 20 points within a site where two cameras were placed at each.
Since the earliest of these surveys, there has been a growing recognition of the problem of hybridisation, both in terms of the status of the wildcat and the problems hybridisation creates for reliable verification of wildcat records. Since 2005 there has been a relatively objective method for assessing whether a cat is a wildcat based on its appearance (Kitchener et al., 2005). However, it is not an easy method, requiring some training and usually close examination or good quality photographs. Kilshaw et al. (2016) was the first large-scale survey relying solely on camera-trap technology able to apply this definition. Data for this were collected using intensive long-term (60-day) camera surveys at 27 locations (including data from Hetherington and Campbell (2012)) across the near-historical range of the wildcat from 2010 to 2013 (Figure 16). A ‘relaxed’ pelage score of 14 and above with no scores of 1 (domestic cat traits) was the threshold for wildcats during this survey, although all cats, except for one individual (PS = 16) classified as wildcat, had pelage scores >17 so are comparable to the data collected in this study. The survey found wildcats at seven of these sites, all in the central-east Highlands (Cairngorms National Park, Aberdeenshire and Angus) or west coast (Morvern). Seven sites in the northern Highlands yielded no wildcats, nor did a site in Argyll and one in the Trossachs.
4.3.1.2 Current wildcat distribution
Since the launch of the SWA project, we worked to tighten the standard at which wildcat records from the public are accepted (see ANNEX 2), while retaining records as plausible where the evidence did not allow definitive verification. Compared with Kilshaw et al. (2016), we used a higher pelage score threshold for wildcats of 17/21 with no scores of one (though note this is still below the strict threshold of 19 set by Kitchener et al. (2005)). We also raised the profile of the wildcat with the public (see the Communications report) and called for public sightings, resulting in a significant increase in records reported to iRecord. The distribution of verified records from the public since 2015 generally confirm the results of Kilshaw et al. (2016), with no records in the far north and only one record from Argyll. However, we received some plausible records in both those areas and so we would encourage reporting of further records should there be sightings. As with these past surveys, we also see much less evidence for wildcats around Peterhead and around that northwest-southeast line from Kyle to Stirling. Camera surveys by partner organisations during the period of this project further support this distribution, with most notably no records from two surveys in the Loch Lomond and Trossachs National Park.
Overall, it appears that the range of the wildcat (as defined by pelage) has contracted from the far north of Scotland and possibly the south-west (Argyll and the Trossachs), a trend hinted at in records prior to the project (Davies and Gray, 2010, Mathews et al., 2018). In contrast, records of hybrids came from throughout the current and former range of the wildcat and notably there was no region free from hybrids. Here, and in the Genetics and Morphology report, we see multiple lines of evidence that point in the same direction: obvious phenotypic signs of hybridisation in almost all individuals in all areas where cats were recorded; a genetic signature of hybridisation in all samples collected from multiple sources (live-capture and road casualties), including from cats that showed the fewest phenotypic indicators of hybridisation; and extremely low numbers of cats with few phenotypic signs of hybridisation from public sightings and camera surveys conducted by project staff and partners (see also Breitenmoser et al., 2019). Based on this, we believe it is unlikely that there is a viable pocket of non-introgressed wildcats remaining undetected in Scotland. The mostly likely scenario is that the population consists of a hybrid swarm (see the Genetics and Morphology report) in all areas within the near-historic range of the wildcat, with very few individuals that pass pelage or genetic thresholds remaining.
4.3.1.3 Causes of the recent decline
Some of this apparent recent range contraction could result as a by-product of our improved knowledge. We better understand the extent of hybridisation and there has been an increased use of camera traps that allows better verification (compared with tracks and scats, for example), where records of hybrids accepted as wildcat in the past are now recorded as hybrid. However, the range decrease also likely reflects real changes in distribution.
Introgressive hybridisation is a gradual and cumulative process: if the number of wildcats is low and continues to decrease, then the relative proportion of hybrid cats with significant wildcat ancestry will decrease over time, to be replaced by an increasing number of hybrid cats with greater domestic cat genetic component and subsequently lower pelage scores (i.e. a genetic dilution of the wildcat population) [see the Genetics and Morphology report]. Introgressive hybridisation can be driven by many factors, particularly the reduction in wildcat numbers, leading to fewer breeding opportunities between individual wildcats. Notably, alongside a lack of wildcat records, there are few records of hybrids in the far north or south-west, suggesting that these areas became less suitable for cats generally, perhaps due to a mixture of poor or declining habitat quality, sudden reduction in rabbit populations (known to occur in several locations where wildcat records exist prior to but not after the rabbit population crashed) and/or persecution of the remaining wild-living cats.
An additional cause of wildcat population declines may be the widespread historic and recent crashes in rabbit populations (Oryctolagus cuniculus) from viral disease, including repeated myxomatosis outbreaks and several strains of rabbit haemorrhagic disease (Mathews et al., 2018).
The series of unusually snowy winters in 2009-2011 may have also played a role since deep snow would have made the hunting of alternative prey such as field vole Microtus agrestis more difficult. For example, the decline in wildcats on an estate in Strathspey (see 4.2.1.6 above) occurred over the 2009-2011 snowy winters and concurrent rabbit declines, while Assynt had records of both rabbit and wildcat prior to 2010, but the Kilshaw et al. (2015) survey in 2012 found neither. Several Assynt landowners remarked to one of the authors on the disappearance of cats when local rabbit populations crashed. In contrast, the same survey series found plentiful rabbits concentrated in a small corner at a site further north, but less suitable habitat for cats because of the general lack of cover. No cats of any type were found here, suggesting various factors are also involved in determining whether cats are present in an area or not.
Prey and weather may have operated in conjunction with other pressures such as persecution. While many land managers support the conservation of wildcats, our impression from speaking with land managers across the range of the wildcat is that this support is not universal. Note that hybridisation may act in conjunction with many other factors. The legitimate killing of hybrid and feral cats by land managers during predator control may have helped to slow the pace of hybridisation once it had started, but a land manager targeting hybrids is more likely to accidentally kill a wildcat through misidentification, particularly when indiscriminate methods are used. Several factors may act together since smaller populations are less resilient to negative effects (Caughley 1994). Other drivers have been identified during the project proposed to us and we summarise these in Table 19.
| Driver | Evidence |
|---|---|
| Persecution | Intensive and widespread persecution, particularly by gamebird shooting estates, aided by improved technology such as lamping and snaring prior to legal protection in 1988. Legal protection ineffective because hybrids cannot be reliably distinguished and therefore a higher risk of wildcats being mistaken for hybrids now that hybridisation is widespread, particularly in conjunction with the increased use of infra-red technology, which cannot distinguish between cat types. There is general support from land managers for wildcat conservation (see the Land Management report), but this is not universal and often does not translate into positive action. However, there are limited recent data on the extent of accidental or intentional killings of wildcat. Anecdotal discussions between land managers and Project Officers suggests that persecution remained intensive over recent decades. |
| Hybridisation | Very strong evidence that wildcats are part of a hybrid continuum, see the Genetics and Morphology report. Introgressive hybridisation is driven by, and may operate in a negative feedback loop with, other factors. |
| Rabbit declines | Correlative declines in both rabbits and wild-living cats have been observed (Hetherington and Campbell 2013; Kilshaw et al., 2015), but no causal effect established. |
| Weather | Observed snowy winters in 2009-2011 coincide with wild-living cat declines (Hetherington and Campbell, 2012, Kilshaw et al., 2015), but no causal effect established. Snow may affect cats indirectly, through the impact of long-lying snow on prey populations, in particular rabbits, or directly by making movement more difficult and shielding prey such as voles. Wildcats in Switzerland have been observed previously to dislike deep snow (Dotterer and Bernhart, 1996, Mermod and Liberek, 2002). |
| Habitat change | There are no data on changes in habitat availability. For a relatively generalist carnivore, such as the wildcat, this is difficult to measure. In the past 15 years in the Highlands there has been an increase in developments overall, which can involve the removal of scrub habitats (possibly supporting rabbits), so there may have been some level of continued habitat change/loss. Over a longer timescale, the reduction in mixed farming over the past several decades may have reduced the diversity of prey and cover types for wildcats in some areas, though no data are available to examine this. Similarly, in the late 1800s/early 1900s, the increase in grouse estates will have also resulted in a significant change in the landscape. Recent afforestation projects, particularly of native woodland, could potentially provide new habitat for wildcats provided it is not planted at the expense of shrub habitats such as gorse (see the Ecology report). |
| Increasing road network / traffic | Wildcats and other cats are killed on roads. It is not possible to assess whether this has a population-level impact but given the very low numbers of wildcats thought to remain in Scotland, and the disproportionate impact vehicle collisions have on young dispersing individuals, it is feasible that road mortality has a significant negative impact on local populations. Hartmann et al. (2013) found some evidence that motorways act as a barrier to gene-flow in wildcats in Germany. See 4.4.2 for further discussion. |
| Disease | A consequence of hybridisation is that there are fewer barriers to interactions between domestic cats and wildcats. Our disease data suggest that epidemiologically, wildcats, hybrids and domestic cats are the same population and may be susceptible to the same diseases (see the Disease Surveillance report). Currently there is no evidence that disease has a population-limiting impact on wildcats in Scotland. |
| Secondary poisoning | We identified high levels of exposure to rodenticides in some cats (see the Disease Surveillance report). It is unclear whether this has played a role in the wildcat’s decline. |
| Competition with other carnivores | No data are available on the impacts of intra-guild competition on wildcats. An initial analysis of some camera data collected during the project found no evidence other carnivore affected cats (Forbes, 2019). It is likely that competition only has an impact when wildcat numbers are already very low. In Germany, fox predation on kittens is thought to be a significant cause of infant mortality (Anonymous pers comm, 2019). |
4.4 Other findings
4.4.1 Are hybrids more ‘invasive’ than domestic cats?
During the course of the project, it was unusual for wild-living cats to score as domestic cats under genetic testing (i.e. UBQ < 0.25). Analysis of the combined results confirmed this to be accurate with most cats testing as domestic trapped in colonies or in close proximity to inhabited buildings. Moreover, the most wild-living individuals (as measured by trap distance from inhabited buildings) also displayed the highest genetic scores. We cannot establish cause and effect for this relationship and clearly, with the distances involved (maximum distance from habitation was 2.5km), individuals living closest to habitats could travel far from habitations and vice versa. Nevertheless, trapping location probably reflects the core home range for most individuals. Possibly, cats living furthest away from inhabited buildings exhibited generally higher genetic scores because hybridisation risk is lower. Alternatively, there is selection pressure against cats with high domestic ancestry away from human habitations due to environmental constraints or because they cannot compete in territorial contests with cats with more wildcat ancestry.
There may also be changes in behavioural traits associated with some introgressed alleles, such as boldness (Montague et al., 2014) or behaviours relating to differences in colour vs nocturnal vision (surprisingly perhaps, a much greater density of colour sensitive cones has been noted in the retina of wildcats, Kitchener et al., 2010). What is clear from the genetic scores is that the wild-living cat populations (i.e. those that were trapped at least 50m from human habitation) in our Priority Areas are made-up almost entirely of genetically hybrid cats with a near complete absence of feral domestic cats, even though some of those cats scored as domestic based on pelage. An extreme example of this situation was reported in the media in 2018 (e.g. BBC, 2018), where two to three kittens, one of which clearly showed hybrid traits (it had blotchy-tabby markings), were filmed at a very remote location near Lochnagar, 800m above sea level and around 6km to the nearest building. The surrounding habitat was dry heath and bog, with no forest cover nearer than 2.5km. It seems unlikely that a feral domestic could survive or breed successfully at such a location.
A useful future study would be to examine the pattern of feral domestic cat distribution in areas of Britain with similar environments that do not contain wildcats or hybrids. If domestic cats live away from human habitations in those areas, this would be evidence that domestic cats in our areas are constrained by the presence of hybrids and wildcats and not by the environment. Conversely, if domestic cats don’t appear to live as independently in other areas as hybrid cats do in Scotland, introgressive hybridisation may have led to a situation where cats we would class as non-native are able to live in more remote locations where population management is difficult to enact. The long-term conservation of wildcats will need to address the management of wild-living hybrid cats across Scotland (including locations outside the former range of the wildcat where possible hybrids were detected, such as in Galloway). Long-term conservation for the wildcat in Scotland entails that the wildcat population is self-sustaining without the need for future interventions to reduce introgressive hybridisation. The existing hybrid population may present a risk to this outcome by maintaining a reproductive ‘bridge’ between wildcats and domestic cats (sensu Germain et al. 2009).
4.4.2 Road risk
Carcases were collected from roads of all types and so we should not assume that larger roads present a higher risk than smaller roads. Possibly, this is because cats use smaller quieter roads to travel along (Campbell, 2015), also see the Ecology report, exposing themselves to greater risk than when crossing the road. This differential use of different sized roads could be taken into consideration when designing mitigation. For example, larger roads will have a higher risk from crossing and so structures designed to funnel individuals to safe crossing points may be effective (Klar et al., 2009). But the same mitigation measure may not work as effectively on smaller roads, where deploying road signs could be a better strategy. Males made up the majority of road casualties, possibly being killed nearly twice as often. This may be because the greater ranging behaviour of males puts them at greater risk and data from the GPS collars suggest males are much more inclined to use potentially busy roads than females (Campbell, 2015); also see the Ecology report). There is a significant peak in cat road casualties in late summer and autumn, coinciding with dispersal, and a smaller peak in January coinciding with the peak breeding season. The sample size is too small to establish whether the winter peak is driven by males moving around more in their search for females (see the Ecology report) and there is no clear evidence that the autumn peak is driven solely by dispersing juveniles, though there is a clear peak in the number of juveniles killed on the road during October. It is possible that autumn dispersal leads to more movement by all age classes as dispersing individuals attempt to establish home ranges and current incumbents increase their territorial patrol behaviours. A study of road casualty seasonality for otters Lutra lutra (Philcox et al., 1999) noted a similar peak in road casualties in autumn to early winter, along with a peak in March. Macdonald et al. (2010) suggested that this peak was driven by the coincidence of peak daily activity for otters and peak vehicle traffic on roads during rush hour. Cats are similarly crepuscular and it is therefore probable that the increase in traffic at certain times of the day plays a role here too.
4.4.3 Lessons learned on monitoring
4.4.3.1 Volunteer management and data handling
This project initially employed two full-time staff covering two Priority Areas each and a further two part-time staff each covering one Priority Area. Their task in the first winter was to recruit and train volunteers and set up the winter camera trap monitoring that would inform further conservation work in these areas. Cameras and bait had to be delivered to volunteers, the volunteers trained in the field and supervised for the duration of the survey, and data and equipment collected afterwards. Staff and volunteers together collected an impressive amount of data on the cat populations in each area during this first winter. However, the time and effort involved in completing an effective survey delayed implementation of other intensive activities, such as the Trap, Neuter, Vaccinate and Return programme. It was not possible to run these activities simultaneously during the first winter given the limited staff resources. The workload involved in collating the survey data, extracting the cat images from each camera and then identifying the cat individually and pelage scoring it took additional months following the end of the survey.
Working with a contractor in the Angus Glens, we streamlined the process so that cats were pelage scored, but not necessarily identified as individuals ahead of targeting for neutering. A consequence of not identifying cats individually is that we could not then rely on images collected of the individual at other cameras to improve the pelage scoring in advance of trapping. Therefore we often adopted a trap-and-assess policy, where cats were trapped and then pelage scored under sedation. However, because we did not identify the individual in advance, we ran the risk of unnecessarily targeting an individual we had recently trapped at a different location. This technique improved the overall efficiency of TNVR (see the Trap Neuter Vaccinate Return Programme report), but still required time later to identify individuals. Overall, monitoring at the level achieved during this project is resource-intensive. Future projects should look at ways to make the management of equipment, allocation of bait and data handling more efficient. We experimented in the final winter (2019-2020) with allowing the, by then experienced, volunteers in Strathbogie to manage their own survey. This was led by a small team of volunteer coordinators, who passed back to the trappers any records of cats that could be targeted for capture. We encouraged them to upload the results to MammalWeb, a bottom-up citizen science platform for sharing and identifying camera-trap images. Data collection and transmission of the relevant information to project staff was successful, but few of the volunteers were willing to upload the images to MammalWeb. Possibly the primary motivations for some project volunteers (to conserve wildcats) are different to many other camera trap volunteers (to collect mammal records).
4.4.3.2 Methodological notes
In the first year, we discovered occasional problems where the trail cameras set by volunteers were placed too close to the bait, or too high up, so that not all of the cat was visible in the images. This frustrated pelage scoring and prompted us to clarify the advice we gave to volunteers. The opposite issue was sometimes that the camera was too far away, resulting in not enough detail of the pelage being visible in night-time images.
Our monitoring methods attempted to balance scientific rigour with utility for other conservation activities (particularly, locating cats for targeting). We adjusted baiting and allowed cameras to be repositioned in subsequent surveys, improving our efficiency (e.g. having more cats identified per unit effort), but at a cost of making comparisons between years more difficult. One particular advance was employing dried valerian root, bound-up in cotton cloth, at each camera. This enabled, on occasion, non-invasive genetic testing from the collection of saliva samples. The lure also sometimes created social ‘nodes’ for cats, especially if one individual began marking the post to which the valerian lure was attached. Where this occurred, the camera was often visited subsequently by several different cats. The valerian root also had the advantage of being effective in summer, when we have noted that meat bait is much less effective. Salmon oil as a scent lure may still be effective in some areas, possibly more in coastal areas where the cats have experienced feeding on fish.
Overall, we would recommend valerian root tied to a post and feathers (e.g. a pheasant wing) hung above the reach of other carnivores, with meat bait (game bird, such as quail, partridge or pheasant) added in winter. Cutting open entire gamebirds used for bait helps by increasing the smell of the bait. However, wherever bait is used consideration should be taken to disease transmission risks (see Sorensen et al., 2014). If multiple individuals are visiting the same bait station and there is a risk of saliva exchange such as from chewing on the same piece of meat or the same valerian pouch, diseases could be transmitted between individual cats. During this project relatively few wild-living cats tested positive for diseases that can be transferred in this way, though they are prevalent. The number of different cats visiting the same locations within days of each other was also low except at a few locations, and so both these factors will reduce this risk. Ensuring valerian pouches are changed regularly (also helpful to obtain a genetic sample from just one individual) and taking away used food bait will further reduce such risks.
We also recommend that future camera trap surveys do not space camera traps further apart than 1km in suitable habitat, to maximise the chances of capturing individuals that have comparatively small movement parameters. This is thought to be particularly important for improving the detection of female cats (e.g. Anile and Devillard, 2018) and that camera trap arrays are as large as possible to account for the wider ranging wildcats.
Please note that this series of specialist reports was written in parallel and thus minor discrepancies between the sizes of datasets used in different parts of the reports may occur. This document reports on genetic records generated by the Royal Zoological Society of Scotland (RZSS) during the course of the SWA project, joined to metadata as provided by NatureScot in August 2019 from the database SWA_DB_06_Snapshot_20190815. Any variation from this database is described in the text. Furthermore, reference to a cat as ‘domestic’ is based on pelage or genetic scores and not lifestyle. A domestic cat can be a pet cat, farm cat, stray or feral. Feral cats may be reliant to some extent on human food sources or may be living wild. Throughout we have highlighted where we refer to a subset of these domestic cat types.
5. REFERENCES
Adams, J. R., Waits, L. P. & Kelly, B. T. 2003. Using faecal DNA sampling and GIS to monitor hybridisation between red wolves (Canis rufus) and coyotes (Canis latrans). Molecular Ecology, 12, 2175-2186.
Aebischer, N. J., Davey, P. D. & Kingdon, N. G. 2011. National Gamebag Census: Mammal Trends to 2009.
Anderson, E. 1949. Introgressive Hybridisation, Wiley, New York.
Anile, S., Amico, C. & Ragni, B. 2012. Population Density Estimation of the European Wildcat (Felis silvestris silvestris) in Sicily Using Camera Trapping. Wildlife Biology in Practice, 8, 1-12.
Anile, S., Bizzarri, L. & Ragni, B. 2009. Camera trapping the European wildcat (Felis silvestris silvestris) in Sicily (southern Italy): preliminary results. Hystrix (Italian Journal of Mammalogy), 20, 55-60.
Anile, S. & Devillard, S. 2018. Camera‐trapping provides insights into adult sex ratio variability in felids. Mammal Review, 48, 168-179.
Anile, S. & Devillard, S. 2020. Spatial variance-mass allometry of population density in felids from camera-trapping studies worldwide. Scientific Reports volume 10, 14814.
Anile, S., Ragni, B., Randi, E., Mattucci, F. & Rovero, F. 2014. Wildcat population density on the Etna volcano, Italy: a comparison of density estimation methods. Journal of Zoology, 293, 252 - 261.
Balharry, D. & Daniels, M. J. 1998. Wild living cats in Scotland. Scottish Natural Heritage Research, Survey and Monitoring Report No. 23. . Edinburgh, Scotland
Barton, K. A. 2019. Mu-MIn: Multi-model inference. R Package Version 0.12.2/r18. [Online]. [Accessed].
Bates, D., Mächler, M., Bolker, B. & Walker, S. 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software, 67, 1-48.
Beaumont, M., Barratt, E. M., Gottelli, D., Kitchener, A. C., Daniels, M. J., Pritchard, J. K. & Bruford, M. W. 2001. Genetic diversity and introgression in the Scottish wildcat. Molecular Ecology, 10, 319-336.
Beugin, M., Leblanc, G., Queney, G., Natoli, E. & Pontier, D. 2016. Female in the inside, male in the outside: insights into the spatial organization of a European wildcat population. Conservation Genetics 17, 1405–1415
Bewick, T. 1820. A general history of British quadrupeds, John Van Voorst, London.
Biro, Z., Szemethy, L. & Heltai, M. 2004. Home range size of wildcats (Felis sylvestris) and feral domestic cats (Felis sylvestris f. catus) in a hilly region of Hungary. Mammalian Biology, 69, 302-310.
Bol, S., Caspers, J., Buckingham, L., Anderson-Shelton, G. D., Ridgway, C., Buffington, C. A. T., Schulz, S. & Bunnik, E. M. 2017. Responsiveness of cats (Felidae) to silver vine (Actinidia polygama), Tatarian honeysuckle (Lonicera tatarica), valerian (Valeriana officinalis) and catnip (Nepeta cataria). 13, 70 (2017). . BMC Vet Res, 13, 70.
Breitenmoser, U., Lanz, T. & Breitenmoser-Würsten, C. 2019. Conservation of the wildcat (Felis silvestris) in Scotland: Review of the conservation status and assessment of conservation activities. 2019.
Campbell, R. D. 2015. Spatial ecology of the Scottish wildcat. Report to PTES. Wildlife Conservation Research Unit, Oxford, U.K. (96 pages)
Campbell, R. D. & Griffith, M. 2015. The use of remote cameras for monitoring cage traps during animal trapping. Ecological research, 30, 963-967.
Can, C. E., Kandemir, I. & Togan, I. 2011. The wildcat Felis silvestris in northern Turkey: assessment of status using camera trapping. Oryx, 45, 112-118.
Carbone, C. & Gittleman, J. L. 2002. A common rule for the scaling of carnivore density. Science, 295, 2273-2276.
Cocks, A. H. 1876. Wild cats: period of gestation. The Zoologist, 5038.
Converse, S. J., White, G. C. & W.M., B. 2006. Small Mammal Responses to Thinning and Wildfire in Ponderosa Pine-Dominated Forests of the Southwestern United States. The Journal of Wildlife Management, 70, The Journal of Wildlife Management.
Corbett, L. K. 1979. Feeding ecology and social organization of wild cats (Felis silvestris) and domestic cats (Felis catus) in Scotland. PhD, University of Aberdeen, PhD Thesis, 334 pp.
Daniels, M. J. 1997. The biology and conservation of the wildcat in Scotland. PhD, University of Oxford.
Daniels, M. J., Balharry, D., Hirst, D., Aspinall, R. J. & Kitchener, A. C. 1998. Morphological and pelage characteristics of wild living cats in Scotland: Implications for defining the 'wildcat'. Journal of Zoology, 244, 231-247.
Daniels, M. J., Beaumont, M. A., Johnson, P. J., Balharry, D., Macdonald, D. W. & Barratt, E. 2001. Ecology and genetics of wild-living cats in the north-east of Scotland and the implications for the conservation of the wildcat. Journal of Applied Ecology, 38, 146-161.
Darwin, C. 1875. Variation of animals and plants under domestication (2nd Edition), New York: D. Appleton & Co.
Davies, A. R. & Gray, D. 2010. The distribution of Scottish wildcats (Felis silvestris) in Scotland (2006-2008). Natural Heritage Commissioned Report No. 360.
Delahay, R. J., Daniels, M. J., Macdonald, D. W., Mcguire, K. & Balharry, D. 1998. Do patterns of helminth parasitism differ between groups of wild-living cats in Scotland? Journal of the Zoological Society of London, 245, 175-183.
Dotterer, M. & Bernhart, F. 1996. The occurrence of wildcats in the southern Swiss Jura Mountains. Acta Theriologica, 41, 205-209.
Doyle, A. T. 1990. Use of riparian and upland habitats by small mammals. Journal of Mammalogy, 71, 14-23.
Driessen, M. M., Jarman, P. J., Troy, S. & Callander, S. 2017. Animal detections vary among commonly used camera trap models. Wildlife Research, 44, 291-297.
Easterbee, N., Hepburn, L. V. & Jefferies, D. J. 1991. Survey of the status and distribution of the wildcat in Scotland, 1983-1987. Edinburgh: Nature Conservancy Council for Scotland.
Ecke, F., Löfgren, O., Hörnfeldt, B., Eklund, U., Ericsson, P. & Sörlin, D. 2001. Abundance and Diversity of Small Mammals in Relation to Structural Habitat Factors. Ecological Bulletins
49, 49, 165-171.
Efford, M. 2020. secr: Spatially explicit capture-recapture models. R package version 4.3.3
Efford, M. G. 2015. secr: Spatially explicit capture-recapture models. . R package version 2.9.4
Ferreira, C. C. & Delibes-Mateos, M. 2011. Wild Rabbit Management in the Iberian Peninsula: state of the art and future perspectives for Iberian lynx conservation. Wildlife Biology in Practice, 6, 48-66.
Flowerdew, J. R. & Ellwood, S. A. 2001. Impacts of woodland deer on small mammal ecology. Forestry, 74, 277-287.
Forbes, H. 2019. Multi-species occupancy modelling of carnivores detected through camera trapping. BSc, University of Aberdeen.
Gasperini, S., Mortelliti, A., Bartolommei, P. & Bonacchi, A. 2016. Effects of forest management on density and survival in three forest rodent species. Forest Ecology and Management, 382, 151-160.
Gil-Sánchez , J. M., Barea-Azcón, J. M., Jaramillo, J., Herrera-Sánchez, F. J., Jiménez, J. & Virgós, E. 2020. Fragmentation and low density as major conservation challenges for the southernmost populations of the European wildcat. PLOS one.
Gil-Sánchez, J. M., Barea-Azcón, J. M., Jaramillo, J., Herrera-Sánchez, F. J., Jiménez, J. & Virgós, E. 2020. Fragmentation and low density as major conservation challenges for the southernmost populations of the European wildcat. PLOS ONE, 15, e0227708.
Gomez, D. M. & Anthony, R. G. 1998. Small mammal abundance in riparian and upland areas of five seral stages in western Oregon. Northwest Science, 72, 293-302.
Gompper, M. E., Kays, R. W., Ray, J. C., Lapoint, S. D., Bogan, D. A. & Cryan, J. A. 2006. A comparison of non-invasive techniques to survey carnivore communities in Northeastern North America. Wildlife Society Bulletin, 34, 1142-1151.
Gray, A. 1971. Mammalian hybrids: a checklist with bibliography. Commonwealth Bureau of Animal Breeding Genetics Technical communication 10.
Hartmann, S., Steyer, K., Kraus, R. S., Segelbacher, G. & Nowak, C. 2013. Potential barriers to gene flow in the endangered European wildcat (Felis silvestris). Conservation Genetics, 14, 413-426.
Hedges, L., Lam, W. Y., Campos‐Arceiz, A., Rayan, D. M., Laurance, W. F., Latham, C. J., Saaban, S. & Clements, G. R. 2015. Melanistic leopards reveal their spots: Infrared camera traps provide a population density estimate of leopards in malaysia. The Journal of Wildlife Management, 79, 846-853.
Heptner, V. H. & Sludskii, A. A. 1972. [Mammals of the Soviet Union. Vol III: Carnivores (Feloidea).] Vyssha Shkola, Moscow (in Russian). Smithsonian Institute and the National Science Foundation, Washington DC.
Hetherington, D. & Campbell, R. 2012. The Cairngorms Wildcat Project Final Report. Report to Cairngorms National Park Authority, Scottish Natural Heritage, Royal Zoological Society of Scotland, Scottish Gamekeepers Association and Forestry Commission Scotland.
Hewson, R. 1984. Scavenging and predation upon sheep and lambs in west Scotland. Journal of Applied Ecology, 21, 843-868.
Hill, J. O., Pavlik, E. J., Smith, G. L., Burghardt, G. M. & Coulson, P. B. 1976. Species-characteristic responses to catnip by undomesticated felids. Chem Ecol 239-253
Hötzel, M., Klar, N., Schröder, S., Steffen, C. & Thiel, C. 2007. Die Wildkatze in der Eifel: Habitate, Ressourcen, Streifgebiete, Laurenti, Bielefeld.
Hupe, K. & Simon, O. 2007. Die Lockstockmethode – eine nicht invasive Methode zum Nachweis der Europäischen Wildkatze (Felis silvestris silvestris) (The Hair Lure - a non-invasive method for the detection of the European wildcat (Felis silvestris silvestris). In German). Inform.d.Naturschutz Niedersachs., 1, 66-69.
Jenkins, D. 1962. The present status of the wild cat (Felis silvestris) in Scotland. Scottish Naturalist, 70, 126-138.
Jerosch, S., Götz, M. & Roth, M. 2017. Spatial organisation of European wildcats (Felis silvestris silvestris) in an agriculturally dominated landscape in Central Europe. Mammalian Biology, 82, 8-16.
Karanth, K. U. 1995. Estimating tiger Panthera tigris populations from camera-trap data using capture-recapture models. Biological conservation, 71, 333-338.
Kilshaw, K. 2015. Introgression and the current status of the Scottish wildcat. PhD, University of Oxford.
Kilshaw, K., Johnson, P. J., Kitchener, A. C. & Macdonald, D. W. 2015. Detecting the elusive Scottish wildcat Felis silvestris silvestris using camera trapping. Oryx, 49, 207-215.
Kilshaw, K. & Macdonald, D. W. 2011. The use of camera trapping as a method to survey for the Scottish wildcat. In: 479:, S. C. R. (ed.). Scottish Natural Heritage.
Kilshaw, K., Montgomery, R. A., Campbell, R. D., Hetherington, D. A., Johnson, P. J., Macdonald, D. W., Kitchener, A. & Millspaugh, J. 2016. Mapping the spatial configuration of hybridization risk for an endangered population of the European wildcat (Felis silvestris silvestris) in Scotland. Mammal Research, 61, 1-11.
Kitchener, A. C., Yamaguchi, N., Ward, J. M. & Macdonald, D. W. 2005. A diagnosis for the Scottish wildcat (Felis silvestris): A tool for conservation action for a critically-endangered felid. Animal Conservation, 8, 223-237.
Klar, N., Fernandez, N., Kramer-Schadt, S., Herrmanne, M., Trinzenf, M., Buttnerf, I. & Niemitz, C. 2008. Habitat selection models for European wildcat conservation. Biological Conservation, 141, 308-319.
Klar, N., Hermann, M. & Kramer-Schadt, S. 2009. Effects and mitigation of road impacts on Individual Movement Behavior of Wildcats. The Journal of Wildlife Management, 73, 631–638.
Krojerová-Prokešová, J., Homolka, M., Barančeková, M., Heroldová, M., Baňař, P., Kamler, J., Purchart, L., Suchomel, J. & Zejda, J. 2016. Structure of small mammal communities on clearings in managed Central European forests. Forest Ecology and Management, 367, 41-51.
Kuznetsova, A., Brockhoff, P. B. & Christensen, R. H. B. 2017. lmerTest Package: Tests in Linear Mixed Effects Models. Journal of Statistical Software, 82, 1-26.
Lindenmayer, D. B., Franklin, J. F. & Fischer, J. 2006. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biological Conservation, 131, 433-445.
Littlewood, N. A., Campbell, R. D., Dinnie, L., Gilbert, L., Hooper, R., Iason, G., Irvine, J., Kilshaw, K., Kitchener, A., Lackova, P., Newey, S., Ogden, R. & Ross, A. 2014. Survey and Scoping of Wildcat Priority Areas. Scottish Natural Heritage Commissioned Report No. 768.
Long, R., Donovan, T., Mackay, P., Zielinski, W. & Buzas, J. 2007a. Comparing scat detection dogs, cameras, and hair snares for surveying carnivores. Journal of Wildlife Management, 71, 2018-2025.
Long, R., Donovan, T., Mackay, P., Zielinski, W. & Buzas, J. 2007b. Effectiveness of scat detection dogs for detecting forest carnivores. Journal of Wildlife Management, 71, 2007-2017.
Lozano, J., Moleon, M. & Virgos, E. 2006. Biogeographical patterns in the diet of the wildcat, Felis silvestris Schreber, in Eurasia: Factors affecting the trophic diversity. Journal of Biogeography, 33, 1076-1085.
Lozano, J., Virgós, E. & Cabezas-Díaz, S. 2013. Monitoring European Wildcat Felis Silvestris Populations Using Scat Surveys in central Spain: are population trends related to wild rabbit dynamics or to landscape features? Zoological Studies, 52, 1-13.
Lozano, J., Virgos, E., Cabezas-Dıaz, S. & Mangas, J. G. 2007. Increase of large game species in Mediterranean areas: Is the European wildcat (Felis silvestris) facing a new threat? Biological Conservation, 138, 321-329.
Lozano, J., Virgos, E., Malo, A. F., Huertas, D. L. & Casanovas, J. G. 2003. Importance of scrub-pastureland mosaics for wild-living cats occurrence in a Mediterranean area: Implications for the conservation of the wildcat (Felis silvestris). Biodiversity and Conservation, 12, 921-935.
Macdonald, D. W., Daniels, M. J., Driscoll, C., Kitchener, A. & Yamaguchi, N. 2004. The Scottish Wildcat: Analyses for Conservation and an Action Plan.: Wildlife Conservation Research Unit, University of Oxford.
Macdonald, D. W., Newman, C., Buesching, C. D. & Nouvellet, P. 2010. Are badgers ‘Under The Weather’? Direct and indirect impacts of climate variation on European badger (Meles meles) population dynamics. Global Change Biology, 16, 2913-2922.
Malo, A. F., Lozano, J., Virgos, E. & Huertas, D. L. 2004. A change of diet from rodents to rabbits (Oryctolagus cuniculus). Is the wildcat (Felis silvestris) a specialist predator? Journal of Zoology, 263, 401-407.
Mathews, F., Kubasiewicz, L. M., Gurnell, J., Harrower, C. A., Mcdonald, R. A. & Shore, R. F. 2018. A Review of the Population and Conservation Status of British Mammals: Technical Summary. . A report by the Mammal Society under contract to Natural England, Natural Resources Wales and Scottish Natural Heritage. Natural England, Peterborough. .
Matuschek, H., Kliegl, R., Vasishth, S., Baayen, H. & Bates, D. 2017. Balancing type I error and power in linear mixed models. Journal of Memory and Language, 94, 305-315.
Mermod, C. P. & Liberek, M. 2002. The role of snowcover for European wildcat in Switzerland. Zeitschrift fur Jagdwissenschaft, 48, 17-24.
Michał, B. & Rafał, Z. 2014. Responses of small mammals to clear-cutting in temperate and boreal forests of Europe: a meta-analysis and review. European Journal of Forest Research, 133, 1-11.
Mills, A., Morkel, P., Amiyo, A., Runyoro, V., Borner, M. & Thirgood, S. 2006. Managing small populations in practice: Black rhino Diceros bicornis michaeli in the Ngorongoro Crater, Tanzania. Oryx, 40, 319-323.
Moleon, M. & Gil Sanchez, J. M. 2003. Food habits of the wildcat (Felis silvestris) in a peculiar habitat: The Mediterranean high mountain. Journal of Zoology, 260, 17-22.
Montague, M. J., Li, G., Gandolfi, B., Khan, R., Aken, B. L., Searle, S. M. J., Minx, P., Hillier, L. W., Koboldt, D. C., Davis, B. W., Driscoll, C. A., Barr, C. S., Blackistone, K., Quilez, J., Lorente-Galdos, B., Marques-Bonet, T., Alkan, C., Thomas, G. W. C., Hahn, M. W., Menotti-Raymond, M., O’Brien, S. J., Wilson, R. K., Lyons, L. A., Murphy, W. J. & Warren, W. C. 2014. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. PNAS, 111, 17230-17235.
Mucci, N. & Randi, E. 2007. Sex identification of Eurasian otter (Lutra lutra) non-invasive DNA samples using ZFX/ZFY sequences. Conservation Genetics, 8, 1479-1482.
Mudappa, D. 1998. Use of camera-traps to survey small carnivores in the tropical rain forest of Kalakad-Mundanthurai Tiger Reserve, India. Small Carnivore Conservation, 18, 9-11.
Newey, S., Potts, J. & Irvine, R. J. 2015. Simulation study to inform the design of wildcat camera trap monitoring protocols. Scottish Natural Heritage Commissioned Report No. 899. .
Nussberger, B., Greminger, M. P., Grossen, C., Keller, L. F. & Wandeler, P. 2013. Development of SNP markers identifying European wildcats, domestic cats, and their admixed progeny. Molecular Ecology Resources, 13, 447–460.
O'Reilly, C., Statham, M., Mullins, J., Turner, P. D. & O'Mahony, D. 2008. Efficient species identification of pine marten (Martes martes) and red fox (Vulpes vulpes) scats using a 5' nuclease real-time PCR assay. Conservation Genetics, 9, 735-738.
Okarma, H., Sniezko, S. & Olszanska, A. 2002a. Occurence of wildcat in the Polish Carpathian Mountains. Acta Theriologica, 47, 499-504.
Okarma, H., Sniezko, S. & Olszanska, A. 2002b. The occurrence of wildcat in the Polish Carpathian Mountains. Acta Theriologica, 47, 499-504.
Oliveira, T., Urra, F., López‐Martín, J. M., Ballesteros‐Duperón, E., Barea‐Azcón, J. M., Moléon, M., Gil‐Sánchez, J. M., Alves, P. C., Díaz‐Ruíz, F., Ferreras, P. & Monterroso, P. 2018. Females know better: Sex‐biased habitat selection by the European wildcat. Ecology and Evolution, Published online 29 August 2018.
Osborne, P. E. 2005. Key issues in assessing the feasibility of reintroducing the great bustard Otis tarda to Britain. Oryx, 39, 22–29.
Parent, G. H. 1975. La migration récente à caractère invasionnel du chat sauvage Felis silvestris Schreber en Lorraine belge. [Recent colonization of the wildcat Felis silvestris silvestris Schreber in Belgian Lorraine.] (in French). Mammalia, 39, 251-288
Pease, B. S., Nielsen, C. K. & Holzmueller, E. J. 2016. Single-Camera Trap Survey Designs Miss Detections: Impacts on Estimates of Occupancy and Community Metrics. PloS One, 11, e0166689.
Philcox, C. K., Grogan, A. L. & Macdonald, D. W. 1999. Patterns of otter Lutra lutra road mortality in Britain. Journal of Applied Ecology, 36, 748-762.
Pitt, F. 1939. Wild Animals in Britain, B.T. Batsford, London.
Potocnik, H., Kljun, F., Racnik, J., Skrbinsek, T., Adamic, M. & Kos, I. 2002. Experience obtained from box trapping and handling wildcats in Slovenia. Acta Theriologica, 47, 211-219.
Rhymer, J. M. & Simberloff, D. 1996. Extinction by hybridization and introgression. Annual Review of Ecology and Systematics, 27, 83-109.
Sarmento, P. 1996. Feeding ecology of the European wildcat Felis silvestris in Portugal. Acta Theriologica, 41, 409-414.
Schauenberg, P. 1981. Elements d'ecologie du Chat forestier d'Europe (Felis sylvestris Schreber, 1777) [Elements of ecology of the wild cat of Europe] (in French). Revue D Ecologie-La Terre Et La Vie, 35, 3-36.
Scott, R., Easterbee, N. & Jefferies, D. A radio-tracking study of wildcats in western Scotland. Proc. seminar on the biology and conservation of the wildcat (Felis silvestris), 1993 Nancy, France. Council of Europe, Strasbourg, 94-97.
Senn, H. V., Ghazali, M., Kaden, J., Barclay, D., Harrower, B., Campbell, R. D., Macdonald, D. W. & Kitchener, A. C. 2019. Distinguishing the victim from the threat: SNP‐based methods reveal the extent of introgressive hybridization between wildcats and domestic cats in Scotland and inform future in situ and ex situ management options for species restoration. Evolutionary Applications, 399-414.
Senn, H. V. & Ogden, R. 2015. Wildcat Hybrid Scoring For Conservation Breeding under the Scottish Wildcat Conservation Action Plan. Royal Zoological Society of Scotland.
Smit, R., Bokdam, J., Den Ouden, J., Olff, H., Schot-Opschoor, H. & Schrijvers, M. 2001. Effects of introduction and exclusion of large herbivores on small rodent communities. Plant Ecology, 155, 119-127.
SNH 2013. Scottish Wildcat Conservation Action Plan. Scottish Natural Heritage.
Soisalo, M. K. & Cavalcanti, S. M. C. 2006. Estimating the density of a jaguar population in the Brazilian Pantanal using camera-traps and capture-recapture sampling in combination with GPS radio-telemetry. Biological Conservation, 129, 487-496.
Sollmann, R., Gardner, B. & Belant, J. L. 2012. How Does Spatial Study Design Influence Density Estimates from Spatial Capture-Recapture Models? PLoS ONE, 7, 34575 10.1371/journal.pone.0034575
Sorensen, A. A., Van Beest, F. M. & Brook, R. K. 2014. Impacts of wildlife baiting and supplemental feeding on infectious disease transmission risk: A synthesis of knowledge. Preventive Veterinary Medicine, 113, 356-363.
Stahl, P. & Artois, M. Status and conservation of the wildcat (Felis silvestris) in Europe and around the Mediterranean rim. Convention on the Conservation of European Wildlife and Natural Habitats Standing Committee, 1994. Strasbourg, France: Council of Europe Pressey, R. L., 1-76.
Stahl, P. & Artois, M. 1995. Status and Conservation of the wildcat (Felis silvestris) in Europe and around the Mediterranean rim. Nature and Environment, No. 69. Council of Europe.
Stahl, P., Artois, M. & Aubert, M. F. A. 1988. Organisation spatiale et déplacements des chat forestiers adultes (Felis silvestris, Schreber, 1777 en Lorraine. [Spatial organization and displacement of adult forest wildcats (Felis silvestris Schreber, 1777) in Lorraine.] (in French). Terre et Vie 43, 113-132.
Stahl, P., Aubert, M. F. A. & Artois, M. 1992. Assessment of two methods of estimating the proportion of rodents in the wild cat Felis silvestris silvestris diet. Mammalia, 56, 15-24.
Steyer, K., Kraus, R. H. S., Mölich, T., Anders, O., Cocchiararo, B., Frosch, C., Geib, A., Götz, M., Herrmann, M., Hupe, K., Kohnen, A., Krüger, M., Müller, F., Pir, J. B., Reiners, T. E., Roch, S., Schade, U., Schiefenhövel, P., Siemund, M., Simon, O., Steeb, S., Streif, S., Streit, B., Thein, J., Tiesmeyer, A., Trinzen, M., Vogel, B. & Nowak, C. 2016. Large-scale genetic census of an elusive carnivore, the European wildcat (Felis s. silvestris). Conservation Genetics, 17, 1183-1199.
Steyer, K., Simon, O., Kraus, R. S., Haase, P. & Nowak, C. 2013. Hair trapping with valerian-treated lure sticks as a tool for genetic wildcat monitoring in low-density habitats. European Journal of Wildlife Research, 59, 39-46.
Suchomel, J., Purchart, L. & Čepelka, L. 2012. Structure and diversity of small-mammal communities of lowland forests in the rural central European landscape. European Journal of Forest Research, 131, 1933-1941.
Urlus, J., Mccutcheon, C., Gilmore, D. & Mcmahon, J. 2014. The effect of camera trap type on the probability of detecting different size classes of Australian mammals. In: Meek, P. & Fleming, P. (eds.) Camera trapping: Wildlife management and research. Melbourne, Vic., Australia: CSIRO Publishing.
Velli, E., Bologna, M. A., Silvia, C., Ragni, B. & Randi, E. 2015. Non-invasive monitoring of the European wildcat (Felis silvestris silvestris Schreber, 1777): comparative analysis of three different monitoring techniques and evaluation of their integration. European Journal of Wildlife Research, 65, 657-668.
Waller, G. R., Price, G. H. & Mitchell, E. D. 1969. Feline attractant, cis,trans-nepetalactone: metabolism in the domestic cat. Science, 164, 1281-1282.
Weber, D. 2008. Monitoring Wildcats (Felis silvestris silvestris) Guidance for a systematic survey of the distribution of wildcats and for monitoring population changes over time (Translated by Hannes Schnell, Helen Armour and Ruth Maier).
Wittmer, H. U. 2001. Home range size, movements, and habitat utilization of three male European wildcats (Felis silvestris Schreber, 1777) in Saarland and Rheinland-Pfalz (Germany). Mammalian Biology, 66, 365-370.
Woodruff, D. S. 1973. Natural hybridization and hybrid zones. Systematic Zoology, 22, 2123-218.
ANNEX 1 - Camera trapping guidance leaflet
GUIDE TO CAMERA TRAPPING SCOTTISH WILDCATS (also available to download at the foot of this report)
STEP 1: SELECTING A LOCATION USING AERIAL MAPS
Before going into the field look at aerial maps and OS maps (available on Bing maps) of your target survey area. Try and site a camera with the aim that, even if the particular cat isn’t attracted by the baits and lures, it’ll happen to walk past at some point.
Look for:
- Edges between open habitat (such as unmanaged grassland) and cover habitat (such as woodland or scrub).
- L inear features that cats might walk along such as tracks, fences, walls, rivers/ burns, gulleys, woodland belts or lines of scrub, and habitat edges.
- B ottlenecks and habitat bridges such as a narrow strip of cover habitat between two separate woodland patches.
If you have more than 1 camera,
Avoid:
- Wet ground and open moorland with little cover, especially where there is deep heather.
Tips:
- Cats are unlikely to be found in the centre of dense forestry blocks, but they may use the edges, especially next to good prey habitat.
- Clear-fell is good prey habitat, and wind-throw provides good shelter. • Ruins, rocky cairns, or woodland patches are good habitat targets in an otherwise open landscape.
STOP!
Please make sure you have the permission of the landowner before siting trail cameras.
try to space them at least 1km apart.
STEP 2: SELECTING A LOCATION (MICRO-SITING) ON THE GROUND
Once on the ground, look for field signs of cats, and other smaller features that act as bottlenecks for cat movement, including the following:
- Cat scats (be aware that misidentification is easy).
- Cat tracks.
- Animal trails, especially junctions of trails.
- Holes in otherwise animal-proof fencing.
- Posts or trees to secure a camera to. Ideally, these should stand straight with respect to the ground slope.
- An even ground surface.
Tips:
- Old ruins, rocky cairns, hollow trees, brash piles wind-throw, and empty rabbit warrens or badger setts/fox dens can be potential wildcat den sites. But don’t place a camera right next to a den site as this is illegal unless you have a licence from SNH. • Wildcats may stakeout rabbit warrens, although be careful not to place your camera too close or you will get many pictures of rabbits. Avoid:
- Areas with intensive badger activity
- Fields with livestock grazing
- Areas with lots of human activity
- Active forestry felling operations
- Heather
- The edge of a river that might flood during winter
STEP 3: SETTING UP THE CAMERA STATION
Once you have found a good location, you need to set up your baited camera station. Ideally, a clear area just off the path or track, preferably with a tree or post to attach the camera and bait/lures.
Trail Camera Setup Checklist
CAMERA
- Full battery power (lithium type best).
- Empty SD card (at least 4GB if possible).
- Correct time: Use GMT not British Summer Time.
- Correct date: use DD/MM/YY (Cameras often default to US format).
- Correct setting: photo only with 0–5 second delay. If possible, set camera to take a burst of 3 images per trigger.
POSITION
- Cat head height (30-40cm) and level with ground.
- 1 .5-3m away from the trail or bait.
- Oblique angle to path.
- No vegetation in the foreground.
- Face camera away from the sun.
- Bait securely tied to a post or tree at least 1m off the ground. We recommend game birds but chicken legs can work.
- Pheasant wing or feathers (weighted with small stick) hanging above bait can act as a visual lure.
- Valerian root (dried) can be placed in hessian pouch and tied at cat head height to provide additional lure.
WHEN CAMERA AND BAIT ARE IN POSITION
- Test camera: if your camera has a ‘Test’ mode, check that it can detect you moving at the distance/height of a cat.
- Check your view: if possible, review image (e.g. on a digital camera with SD card slot); you should be in lower half of image, with no vegetation in foreground.
BAIT
- Arm the camera and wait until it has finished counting down.
- Make sure all fastenings are tightly closed and the door seals are clean so water/ moisture cannot get in.
- Secure/padlock your camera if necessary.
- Walk IN FRONT of camera and hold up a sheet of paper with the camera number, date, time and location (grid reference).
STEP 4: RE-VISITS AND CHECKS
Return to check the camera station approximately every two weeks, in order to swap the SD card and renew the bait.
- Turn the camera OFF before removing SD cards, or you will corrupt the data.
- Remove SD card and replace with an empty card.
- Check battery and time/date.
- Replace bait, and other lures if necessary.
- Arm the camera and remember to take a photo of yourself with an information sheet as the first image.
STEP 5: DATA
- At home copy your SD card to your computer immediately for back-up, noting location on filename / tags.
- Review all your images to find cats, but do not delete any images (cat or otherwise) collected during the survey.
- If you don’t get a clear enough view, for pelage scoring, of a cat that is visiting your camera, re-angle or perhaps relocate the camera on your next visit (if you are able to review images in the field you can do this immediately). Remember, we need to see the whole cat.
- You can also try placing the bait higher up to encourage the cat to stretch up and show off its nape, shoulders, back and dorsal surface of tail (all useful for pelage scoring).
- I f you are a volunteer on our priority area survey, your project officer will have explained how to share the data with us.
- If you are participating in our national survey, then please visit saving wildcats to upload the cat sightings and images.
IDENTIFYING A SCOTTISH WILDCAT
To enable accurate pelage scoring of a potential wildcat Guide can be found we need clear photos of: on our website here
- Each side of that animal, showing the flank and rump.
- The back of the head and shoulders.
- The dorsal/back stripe and tail.
ANNEX 2 - Public sightings iRecord verification protocol
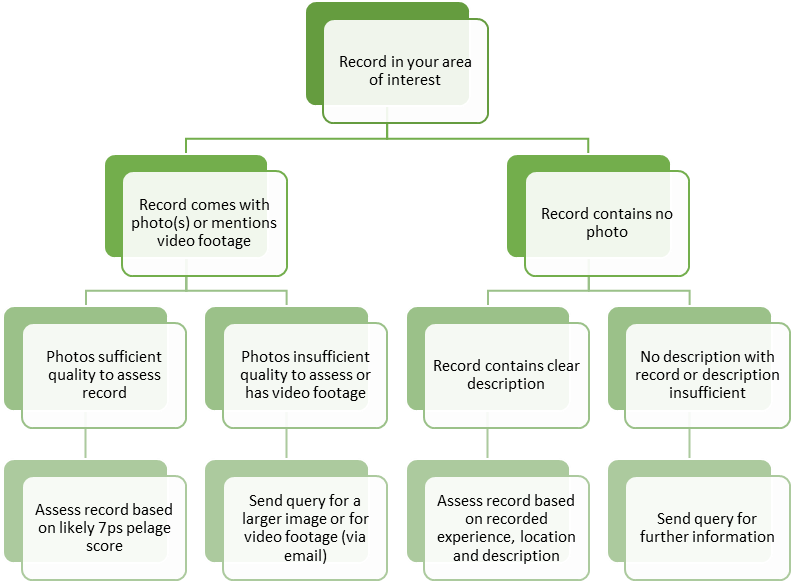
Annex 2 flow chart
Click for a full description
iRecord guidance: http://www.brc.ac.uk/irecord/sites/default/files/guides/verifiers.pdf
Set a ‘Where’ filter, or two, in iRecord to cover your Priority Areas and save the filter. You only need to verify sightings in those areas.
Whatever the record quality, you can confer with other verifiers using the send to expert option in iRecord. You can also write notes in the comment box (see 5.1 in the iRecord verification guidance).
- Where photo or video footage is available
- Score all visible markings from the Kitchener 7PS score
- Follow the project definition of a wildcat (pelage score of ≥ 17 / 21), but see points 1.3 and 1.6
- Use additional information, such as presence of white feet / patches to further grade the cat. E.g. A cat that scores >17 on the 7PS score but has a white paw should be classed as a hybrid
- Where markings are not visible and therefore not scored, take the total score (based on what you can see), divide by the number of features scored and multiply by 7. E.g. A cat scoring 3 for tail shape, 2 for dorsal line, 3 for tail bands, 2 for stripes and 1 for spots on flank and hindquarters, but with missing information for shoulder and nape stripes would score (3+2+3+2+1) / 5 x 7 = 15.4. However, fewer than four pelage characteristics can’t be scored and no view of the tail is possible, the pelage score cannot be relied on and you may have to class the record as Plausible.
- Based on the score and any other defining characteristics (see 1.3), accept the record as correct (see 1.6) or considered correct if it is the right assessment, or redetermine (redet) it (again, using the same above classifications) if the assessment by the recorder is wrong (e.g. the pelage score is likely below 17 and the recorder classed it as a wildcat).
- Kitchener et al. (2005) seven-point (7PS) classification of wildcat based on pelage marking gave a cut-off of 19/21 below which a cat likely contains some domestic cat heritage. As a project, we accept that pelage is an imperfect tool for identifying wildcat and have therefore reduced this threshold (but retained it above the 14 point relaxed threshold) to ensure cats that contain a high proportion of wildcat heritage are not unnecessarily neutered. However, to keep classification consistent with published work, for wildcat only accept a record as correct (instead of considered correct) if you can assess all 7 pelage features and the cat scores ≥19.
- Where only a description is available
- If the recorder reports a wildcat, never accept as correct, only considered correct
- If the recorder reports a wildcat and the description clearly identifies the main characteristics of the cat as a wildcat (fat / blunt-tipped tail, ringed tail, very stripy cat, no white feet), then you can class as considered correct
- If the recorder reports a wildcat and the description misses out some of this information and the Details tab for the recorder indicate limited experience, we recommend you redet the record to hybrid cat and add a note in the comment box that opens when you click the considered correct button. There will be some blurring between points 2.2 and 2.3 in terms of quality of description and ultimately, you may need to use your own judgement. But you can confer with other staff members using the send to expert option.
- If the description is of a feral cat or a hybrid, it is likely that the record won’t need redet, though be aware that misidentification between the two is still possible. A very friendly hybrid is likely to be someone’s pet cat and so can be not accepted. A tabby feral cat may be misidentified as a hybrid cat, though preliminary genetic work found that many feral cats living away from cities in the Highlands show genetic evidence of past hybridisation.
- Where no description is available
- Ask for further information, but avoid leading questions.
- Ask: What made them think it was a wildcat? What were the markings on the flank like? What did the tail look like? How did the cat behave?
- Don’t ask: Was the tail-tip blunt? Was the cat very stripy?
- If the recorder doesn’t reply with more detail the record will remain under query.
- Ask for further information, but avoid leading questions.
You will be faced with a choice of 1) redet a cat reported as a wildcat but which comes with insufficient evidence or 2) accepting the record as Plausible. Plausible records are not passed to NBN but will be available to the project and will still be useful (e.g. for follow up with trail cameras). If the recorder appears convinced the animal was a wildcat, it is better to class as Plausible than to re
|
Verification status 2 |
Explanation |
|---|---|
|
Correct |
The verifier is able to confirm that the species has been identified correctly, usually on the basis of photo/s within iRecord (or specimen/s outside iRecord) |
|
Considered correct |
The verifier has not seen photo/s or specimen/s but has a high degree of confidence that the record is likely to be correct, based on difficulty of ID, date, location and recorder skills/experience etc. |
|
Unable to verify |
The verifier has a high degree of confidence that the record is likely not to be correct based on difficulty of ID, date, location and recorder skills/experience (and where no photo/s or specimen/s are available); or photos are available but do not show enough detail to confirm the identification; and/or the record is not sufficiently well documented to confirm (e.g. location is vague) |
|
Incorrect |
The verifier is able to confirm that the species has not been identified correctly, or the record is erroneous in other respects, on the basis of photo/s or specimen/s, or on information from the recorder |
|
Plausible |
The record is plausible based on species, date and location, but there is not enough supporting evidence for the possibility of misidentification to be ruled out |
|
Not reviewed |
The record is in the system but has either not been looked at, or has been looked at but a decision not yet taken |
ANNEX 3 - Cat procedure form
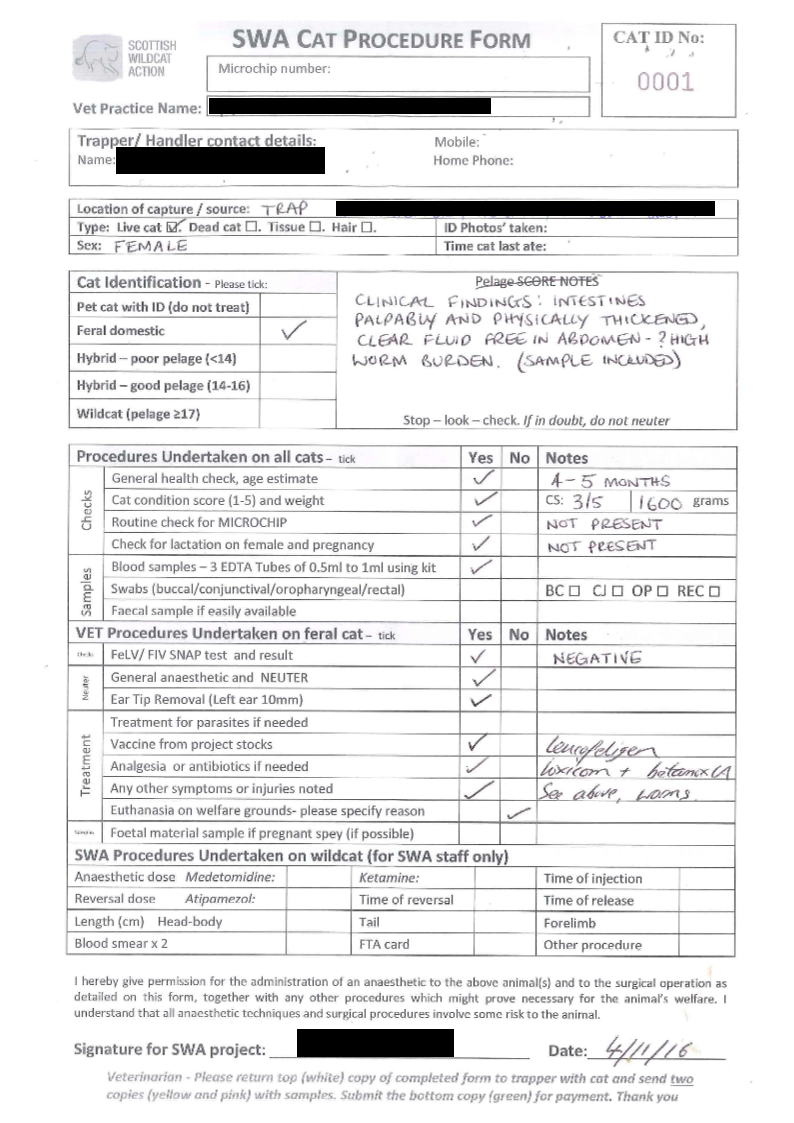
ANNEX 3 - CAT PROCEDURE FORM
Click for a full description
Example of a top copy of the quadruplicate form, filled in for a feral cat captured during TNVR. The Notes box contained, in faint text to allow overwriting, pelage score guidance. Cat ID No. is unique for every quadruplicate set of forms.
ANNEX 4 - Wildcat trapping protocol
For a detailed protocol for live-trapping cats in general, please refer to our TNVR guidance. Details specific to wildcat are provided in this document. Version date 31st Jan 2018.
Licence restrictions
The SWA EPS (European Protected Species) licence for the trapping of wildcat permits:
- A maximum interval between trap checks of 12 hours and a total holding period (time from last trap check to release) of 18 hours. This maximum is by exception and staff or contractors should seek to keep the time spent holding a wildcat (from initial capture to handling and release) to a minimum wherever possible.
- Trapping in all locations (subject to landowner permission) in our five Priority Areas or outside these if within one mile of the boundary of a Priority Area.
All named staff and contractors should carry a copy of our EPS licence whenever conducting wildcat trapping.
Some procedures conducted on wildcats, including sedation/anaesthesia and blood sampling, fall within the Veterinary Surgeons Act. Therefore all such procedures must be undertaken by a qualified veterinary surgeon. If the vet has concerns about the rate of recovery of a sedated wildcat, this takes precedence over the above EPS licence restrictions. Note however there is a trade-off between extending recovery times to benefit the cat and increasing stress for the individual from being held captive for longer.
Prior to trapping
- Secure permission to trap from the landowner, making it clear that this may involve visiting the trap location during night-time.
- Secure the services of a vet to conduct sedation and blood sampling. Our order of preference is
a. The vet is able to attend at or near the trap location at any hour of the day or night
b. The vet is able to process the cat at the veterinary clinic at any hour of the day or night
c. The vet is able to process the cat at the veterinary clinic during opening hours
For option a) an alternative to using a local vet is to contract a vet from elsewhere or secure a volunteer vet.
Options b) and c) should be considered last resorts and every effort should be made to organise a vet to attend in the field. Moreover, the clinic must be within 20 miles or 45 minutes journey time from the trap location.
For c), trap opening time and check intervals must be scheduled so that the licence restrictions are not breached. E.g. this means, assuming an 8am opening time for the clinic, the trap could be set at no earlier than 7:30-8pm and checked by 7:30-8am the next day. A later trap set time should be considered with a target of an 8-hour check interval (i.e. opening the trap at 11:30 – 12pm).
Note that while we would encourage a vet to attend in the field to reduce stress to the wildcat from transport, there are circumstances where taking the wildcat to the clinic could lead to improved welfare because of better facilities. For example, in the unlikely event of a complication during anaesthesia.
- If possible, pre-bait the cage-trap for a period before commencing active trapping. This entails the same trap set-up as detailed below but with the trap door locked open using a padlock. You can also remove the rear door on some traps to create a tunnel.
- Assess the GPRS mobile phone signal strength at the trap location. A good signal suggests you may be able to use an Acorn GPRS/mobile network camera (with working sim card) to assist in remote trap monitoring while a poor (but not absent) signal suggests you may be able to use the Acorn camera with booster antenna or a MinkPolice trap monitoring unit. If you are planning on using these devices, consider testing them during pre-baiting.
- Check from prior trail camera images for visiting wildcat showing any indication of pregnancy or dependent young. This is unlikely during our winter field period. If there is a clear risk of capturing a wildcat with dependent young, then do not proceed.
During trapping
Follow our TNVR guide for methods on trapping and biosecurity, excluding trap check intervals. Aim to check the trap as frequently as possible without causing undue disturbance to the area (which could scare off the wildcat). Technology such as GPRS cameras or MinkPolice (see below) allow very frequent checks with no disturbance. But where these aren’t available or are unusable due to lack of mobile signal then aim to physically check every eight hours. In some situations this may not be practical. For example, where a vet is not available until 8am and the wildcat is likely to visit the trap before it was opened at midnight. In such situations, trap check intervals can exceed eight hours but must not exceed 12 hours.
- Ensure the trap mechanisms are working correctly. The treadle triggers the door release when pressed but that it’s not likely to trigger by itself or that it doesn’t require excessive pressure to trigger. And the trap door locks when closed. You can lubricate the mechanisms with salmon oil, vegetable oil or similar but do not use synthetic oil as the scent may be off-putting to cats.
- Ensure the trap is level and stable and won’t rock when a cat enters. Consider pinning the trap between two posts.
- Ensure the trap is protected from the elements. As a minimum (in very sheltered locations) this should consist of a roof with material placed against the sides of the trap. In exposed locations at least the two long sides of the trap should be additionally protected with windproof covering. Covering the rear of the trap may also be necessary. A plastic trap cover can be used for this, but if you judge this could put off a wildcat from entering, then wall the sides with turf or tightly packed vegetation. Do not use turf or vegetation on the roof in lieu of waterproof material. Make sure the covering doesn’t foul the trap mechanisms.
- Bait the trap with a piece of gamebird (e.g. quail), chicken and/or scent lure (e.g. valerian root). Do not provide excessive food bait as a full stomach could result in a cat vomiting under anaesthesia. The rate at which the bait needs refreshed if not taken will depend on the temperature.
Physical trap checks
- Check the trap at least every 12 hours. A more frequent trap check is preferable, with eight hours being a reasonable target. If possible, check the trap without approaching the trap (e.g. from a nearby vantage point) as long as you can see the door of the trap and the bait inside. If checking remotely, ensure you take a close look at the trap on some visits to ensure no problems have developed with the set-up.
Remote monitoring and using cameras
Details below assume that a vet is available out-of-hours and traps are not being closed and opened daily.
Check you have sufficient network signal for using the camera. At least two bars is recommended for the Acorn cameras. A small number of booster antennae are available with camo-tape to disguise them.
- Place the camera directly facing the trap door at approximately trap height. Wrap a piece of white tape on the door so that it’s obvious when the door is closed.
- Set the camera to send an image every four hours and the PIR sensor is on. Set intervals between PIR triggered images to at least 10 minutes and not more than an hour. The camera should send images every 3.5-4 hours on this setting and also send an image when triggered by a visiting animal.
- The camera will send an image when switched on. Check you are receiving images from the trap before leaving the area (subject to you having sufficient signal on your phone). The front of the trap should be clearly visible in the images.
- Images sent from the camera should allow you to assess the status of the trap door and ideally also the bait inside. Network variability means that sometimes a camera may miss sending an image. If you have not received any images from the camera over eight hours, check the trap physically and check the camera is working (are there unsent images on the SD card or are the batteries dead?).
- If you are at all unsure of the status of the trap from the camera images (e.g. the camera is sending images but these are fogged), check the trap physically within the 12 hour period since the last clear image was taken.
- Physically check the trap at least once per week even if the camera images indicate no issues.
Remote monitoring of traps using MinkPolice
- Ensure you have network signal (the MinkPolice unit will emit a beep to tell you it has found a network).
- With string or braided wire, attach the magnet to the door so that the closing of the trap door pulls the magnet from the MinkPolice unit.
- Enclose the string or wire in a length of pipe/duct so that rodents do not chew through it. Make sure this pipe doesn’t stop the door closing fully.
- The MinkPolice will send a text message when switched on and also provide regular updates as to its status. If the unit fails to send a text, check the MinkPolice website to assess whether contact has been made. If you are at all unsure of the status of the trap, check the trap physically within the 12 hour period since the last ‘all ok’ message was sent.
- For the first 24 hours, physically check the trap every 12 hours. Thereafter, physically check the traps every 24 hours. Over time we will review this and may extend this period as we get more confident of the MinkPolice system.
Cat handling and anaesthesia
When a wildcat is trapped, do not approach the trap until you absolutely have to. This minimises stress to the animal. You can approach the trap:
- To make an initial assessment. E.g. to check it’s not a cat captured previously, a hybrid or feral cat to be taken for neutering, or a pet. In this case, keep the assessment as short as possible and unless the cat is to be released immediately, re-cover the trap and move away.
- When you are ready to transport the cat or when the vet arrives and is ready to proceed
See our TNVR guide for methods to transfer and handle the cat. The vet will judge the most appropriate methods for anaesthesia and sampling. Note that for healthy individuals our veterinary advisor Prof Anna Meredith recommends, injected into the thigh muscle for field anaesthesia, a combination of ketamine (5mg/kg) and medetomidine (40µg/kg). This is a light sedation used without incident previously, giving approximately 5-10 minutes to anaesthesia and 20 minutes before the cat wakes. Reversal is atipamezole at the same dosage as medetomidine. Using this combination, our recommendations and experiences are:
- Cover the cage after injection and keep in a quiet area, only approaching the cage to check anaesthesia.
- There is a low risk of vomiting under anaesthesia and it is better to begin anaesthesia as soon as you are ready rather than subjecting the cat to additional stress by holding it for several hours to ensure an empty stomach.
- Time to anaesthesia is approximately 5-10 minutes.
- The cat will naturally begin to wake after an additional 20 minutes.
- The dose can be topped-up with half the initial dose should the cat not achieve anaesthesia under the initial does (i.e. after 15-20 minutes), should it wake early or wake before the procedure is complete.
- Reversal should be injected even if the cat awakes naturally.
- After reversal, the cat can be released safely after between 20-45 minutes (or 30-80 minutes after the initial ketamine injection). If conditions are particularly cold and snowy, consider holding the cat for longer and use a solid box for holding.
Cat transport and holding
In the event that the wildcat needs transported to a veterinary clinic or other location to complete the procedure, a solid wooden box is preferable over a transfer basket for transport to reduce the risk of injury during transport. If the transfer basket or carry cage must be used, then it should be covered. Lining the base of the transfer basket with soft material (e.g. ‘vetbed’) may reduce rubbing. Do not use the radio or play music in the vehicle and keep conversation to a minimum.
While holding the cat prior to and after the procedure, ensure noises are kept to a minimum and the cat is not left in a room with the scent of other cats and dogs. There is also a risk of exposure to disease at a vet clinic therefore consider holding the cat in a vehicle.
Samples and measurements
Take photos of the cat under anaesthesia for pelage records. Follow the protocol for photographing dead or anesthetised cats written by Dr Andrew Kitchener. The mains points are:
- Smooth fur flat and make sure fur is dry.
- Include a label in as many of the photos as possible.
- Take photos of the cat alongside a measuring tape, showing:
a. Complete dorsal view
b. Complete ventral view
c. Right lateral view
d. Left lateral view
e. Close-up of nape and shoulders
f. Close-up of lower back and tail
g. Lower abdomen and underside of tail
h. Face
Ensure that the complete tail is visible in all images except e. and h.
Measure and record:
- Body length (nose to base of tail following curve of body)
- Tail length (base to tip, excluding fur)
- Upper fore limb-length (humerus)
- Lower fore limb-length (top of ulna to bottom of carpal bones)
All samples should be labelled with the unique ID number found at the top right of the SWA cat procedure form. We require these sample and storage methods:
- Blood (max 3ml), stored in 0.5, 1 or 2ml EDTA tubes (always ensure tubes are filled to the line and inverted several times once sealed to prevent clotting).
- Buccal, oropharyngeal, conjunctival and rectal swabs, stored in swab tubes.
- Whiskers: snip two whiskers with scissors as close to the muzzle as possible. Store dry at room temperature in a zip-lock bag or small envelope. This will allow us to examine diet through isotope analysis. Whiskers regrow and so the procedure has no lasting impact on the cat.
- Faeces if available. Store in a freezer.
- Hair sample – three tufts (>20 follicles) taken from the rump and stored as whiskers.
Postage of samples
Send in an envelope marked ‘Wildcat Project’, addressed to:
For the Urgent Attention of
Professor Anna Meredith
Royal (Dick) School of Veterinary Studies,
The University of Edinburgh,
Easter Bush Veterinary Centre,
Roslin,
Midlothian,
EH25 9RG
PIT tags
Mark the cat for future reference using an ISO-compatible PIT tag (microchip). Record the tag number on the cat procedure form but do not add the number to a pet database. As with other consumables such as medication, if the vet uses PIT tags from their own stock, SWA will pay for them. Check the PIT tag is functioning before releasing the animal.
GPS collar system and attachment
Some cats will have a GPS telemetry collar attached. This should only be done where the vet assesses the cat is healthy and can bear the additional weight of the collar. If a collar is to be fitted, it is recommended that blood samples not be taken from the jugular. The cephalic vein on the front of the fore-limb can be used instead. The e-obs collars weigh around 69g. The traditional rule of thumb is to ensure the collar does not exceed 5% of the body weight of a terrestrial animal. SWA policy is that the collar should not exceed 3% body weight, therefore the e-obs collars should not be fitted on cats with body weight of less than 2.3kg. Most adult wildcats will weight over around 3.5kg, meaning in most cases, the collar will represent less than 2% total body weight.
All collars should have redundancies built in to ensure the collar does not remain on the individual, including a break-away mechanism in case of entanglement. Usually, this will mean the collar containing:
- A weak link, usually 6-12 strands of linen thread that will weaken over time to allow the collar to drop off and act as the break-away mechanism, and
- A method for relocating the individual to allow trapping for collar removal (usually a VHF transmitter), or
- A timed mechanical break-away system
Collars should be sized to the individual and fitted so they are sufficiently snug to minimise collar rubbing but loose enough not to restrict movement. Usually, this can be tested by seeing whether one to two fingers can be inserted between the neck and the collar. Once fitted, rotate the collar to ensure it moves freely and no fur is caught.
Ensure the GPS and VHF transmitter (if it has one) are both active prior to release.
Ensure the collar unique ID number is recorded on the cat's record.
If a possible wildcat is reassessed as a hybrid
Once sedated, a cat that pelage-scored 17+ based on camera images may be reassessed as scoring <17. In this case, the cat should brought around (around 20 minutes after initial injection) and transported to a vet for neutering and vaccinating following our TNVR protocol.
SNAP test failures
If a wildcat's SNAP test shows indication of FeLV or FIV, run a second SNAP test. After a second failure, arrange for the wildcat to be taken to holding facilities at the Highland Wildlife Park near Kingussie, where it can await results from an accurate PCR test for these diseases. Wildcats should be held for no more than a week. A wildcat should be returned and released if it passes the second SNAP test or PCR test. Should these diseases be confirmed, veterinary advice is euthanasia on welfare grounds. The presence of either disease cannot be disentangled from that individual’s future contributions to the wildcat population; therefore the wildcat also presents a risk to other wildcats. If holding facilities cannot be found, the SNAP test results can be assumed accurate (c.1% false positive rate from two tests) and the cat euthanised. However, the final decision on this should remain with the vet.
Risk-assessed approach to trapping
Through the work of SWA, we must balance the risks to the welfare and survival of individual wildcats against the need to conduct conservation work crucial to the survival of the population. All activities carry some risk and it is SWA’s policy to minimise all such risks. The risks detailed below are not exhaustive and the SWA team should review and update this regularly. See Risk Matrix below. Risks to wildcats during trapping include:
- Death of dependent young when the mother is contained in a trap or during a procedure (L=2, C=3). Mitigation: see below for mitigation of this risk.
- Death of individual during a procedure. This has not happened to date. (L=1, C=3). Mitigation: see cat handling and anaesthesia guidance above to mitigate this risk.
- Damage to individuals while contained in the trap (L=2, C=2). Mitigation: follow guidance above to minimise time spent in trap and avoid using 1x2” (rectangle) gauge mesh traps as there may be an increased risk of tooth damage. Instead use square 1x1” gauge traps.
- Death of individual in trap that is accidentally set (L=1, C=3). Mitigation: when pre-baiting with traps, always ensure the trap door is locked open and/or the rear door is removed. This avoids the trap being accidentally set by someone tampering with it.
Risk to dependent young
Are you trapping at a time when a (potentially) female wildcat may have young dependents? This is generally mid-March to late August, though in the west of Scotland the breeding season can begin two weeks earlier.
- If yes, do not set a trap where there is a risk of capturing a female wildcat. Away from human habitations, use trail cameras prior to trapping to look for the presence of female wildcats.
- If no, note that there is a possibility that a wildcat could have dependent young at other times of year. Where possible, use trail cameras prior to trapping and if there is evidence that a female wildcat could have dependent young out of season. For example, look for visible signs of lactation, gravid appearance (swollen bellies) or young kittens themselves. In these circumstances, do not set the trap.
| - | Consequences (C) | Consequences (C) | Consequences (C) |
|---|---|---|---|
| Likelihood (L) | 1 | 2 | 3 |
| 1 | Low - Broadly acceptable risk level - Monitor and reduce. | Low - Broadly acceptable risk level - Monitor and reduce. | Mod - Tolerable risk level - Risk must be reduced. |
| 2 | Low - Broadly acceptable risk level - Monitor and reduce. | Mod - Tolerable risk level - Risk must be reduced. | High - Intolerable risk level - Immediate action required |
| 3 | Mod - Tolerable risk level - Risk must be reduced. | High - Intolerable risk level - Immediate action required | High - Intolerable risk level - Immediate action required |
Equipment and responsibilities
If the procedure is to occur in the field, the vet is responsible for providing the medication and needs to bring:
- Anaesthetics
- Syringes and needles
- Stethoscope or other means of monitoring heart rate
- PIT microchip and applicator (backup supply)
- Vaccine (Tricat and FeLV)
- Emergency drugs: injectable antibiotics in case of injuries, Euthasol, in case of FIV/FeLV positive result and no available holding facilities for further PCR testing
The project staff are responsible for providing all other equipment, which includes:
- Traps and transfer basket
- Cordless clippers
- EDTA tubes
- Swab tubes
- FIV/FeLV test kits and reagent
- Envelopes/zip-lock bags for sample storage
- Camera
- Pelage scoring chart
- Measuring tape
- Battery operated scales / spring balance
- SWA Cat procedure form
- Pop-up tent (in case of poor weather)
- Heat pads (charged), or hot water bottle and vacuum flask with hot water
- Lamp/torch
- Towels/padding and basket covers
- Surgical gloves
- Disposable towels
- Surgical cleaner
- Iodine/alcohol swabs
- PIT tags
- PIT tag reader
Project staff and volunteers are responsible for:
- Erecting the tent
- Transferring the trapped cat
- Taking photographs and measurements
- Confirming pelage score
- Inverting EDTA tubes once filled
- Storing and labelling samples
- Testing PIT tags once injected
- Collecting faecal samples if available
- Releasing the cat once recovered
- Recording details on the SWA cat procedure form
If trained, project staff may also be responsible for the fitting of the GPS collars.
The vet is responsible for
- Directing immobilisation during injection
- Application of eye lubricant
- Directing the monitoring of the cat under anaesthesia
- Sampling of blood, whiskers, swabs and hair (blood not to be taken from jugular if cat is to be collared)
- Processing and interpreting FIV/FeLV tests
- Injecting and checking PIT tag/chip
- Injecting full vaccine (Tricat and FeLV)
- Assessing recovery and clearing the cat for release
ANNEX 5 - Maps showing camera trap placement and where individual cats were detected during the standardised surveys
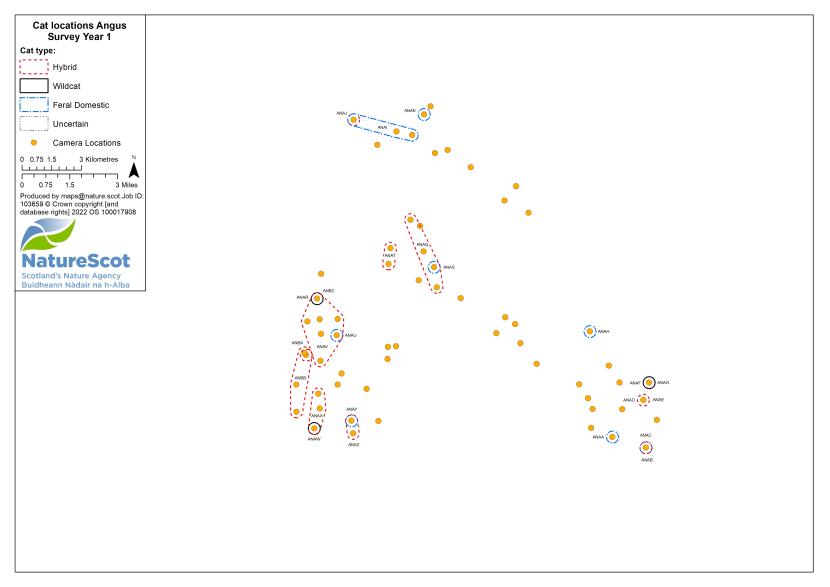
Figure A5.1. Distribution of camera traps and the locations of cats for the standardised survey in the Angus Glens in winter 2015-2016
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
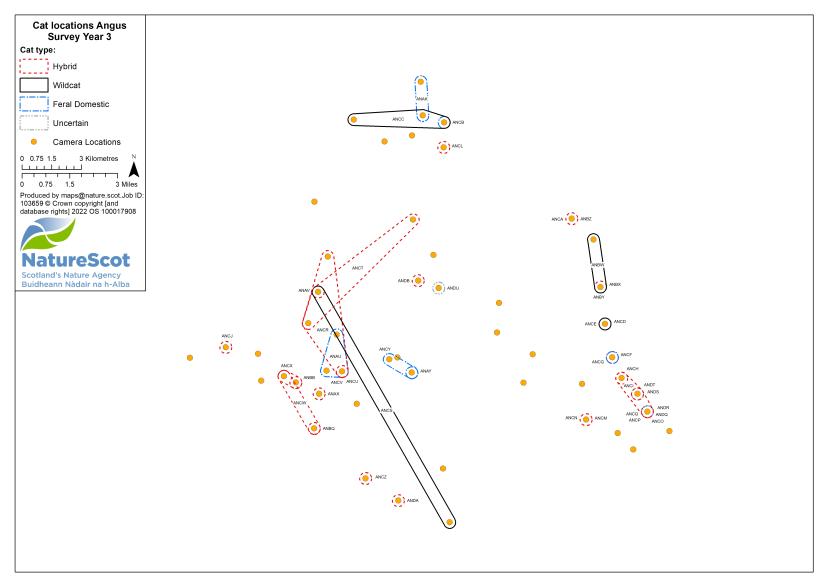
Figure A5.2. Distribution of camera traps and the locations of cats for the standardised survey in the Angus Glens in winter 2017-2018
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
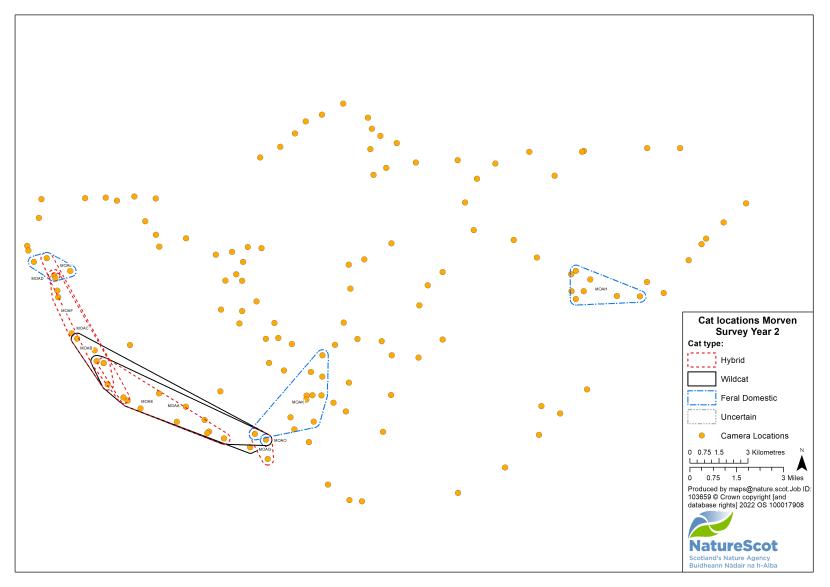
Figure A5.3. Distribution of camera traps and the locations of cats for the standardised survey in the Morvern in winter 2016-2017.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
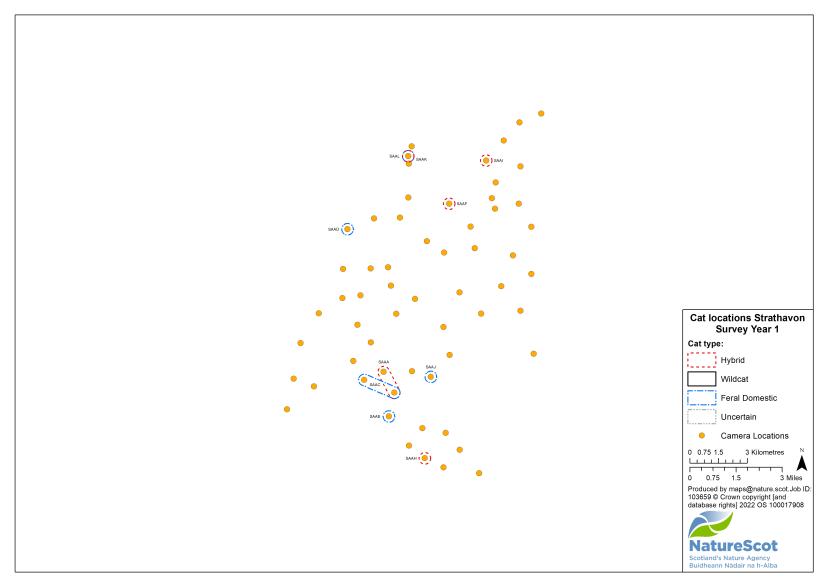
Figure A5.4. Distribution of camera traps and the locations of cats for the standardised survey in the Strathavon in winter 2015-2016.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
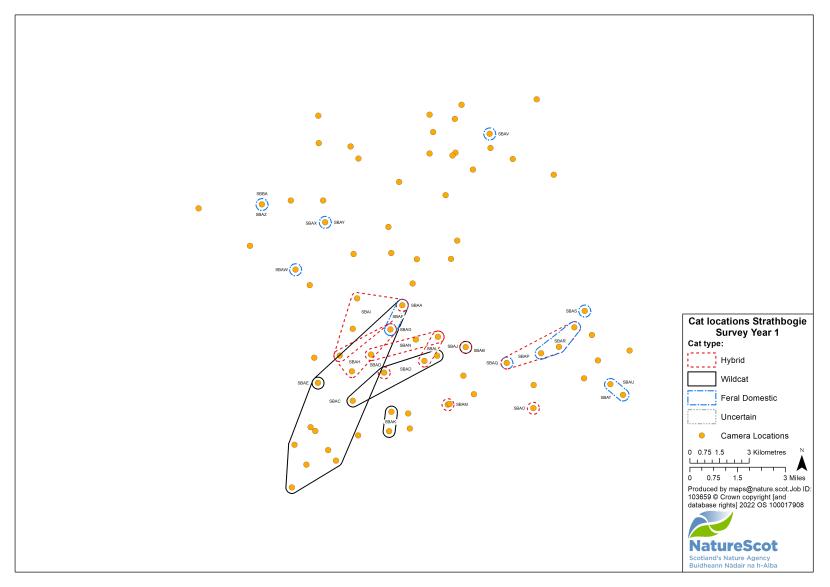
Figure A5.5. Distribution of camera traps and the locations of cats for the standardised survey in the Strathbogie in winter 2015-2016.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
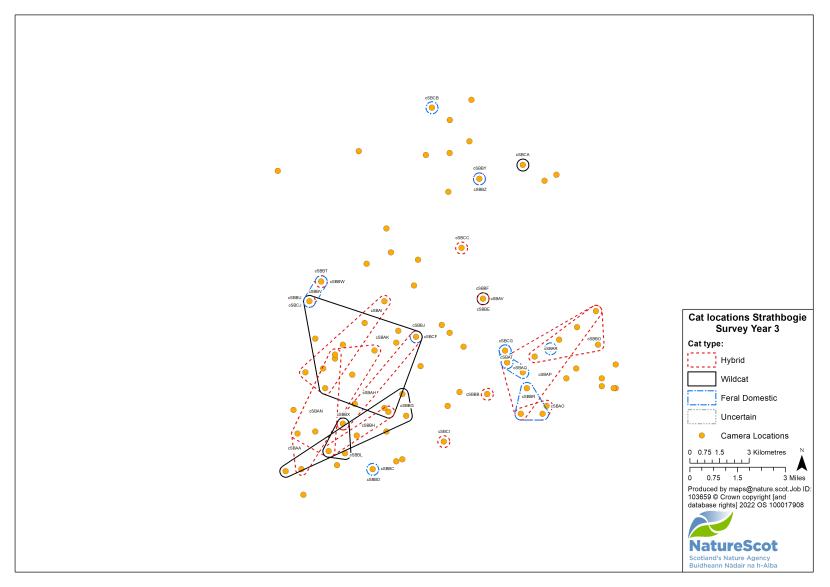
Figure A5.6. Distribution of camera traps and the locations of cats for the standardised survey in the Strathbogie in winter 2017-2018.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
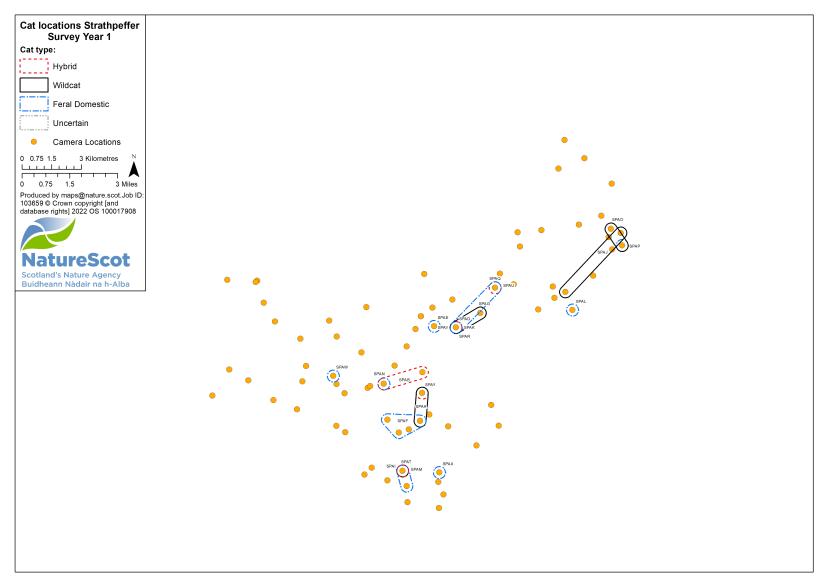
Figure A5.7. Distribution of camera traps and the locations of cats for the standardised survey in the Strathpeffer in winter 2015-2016.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
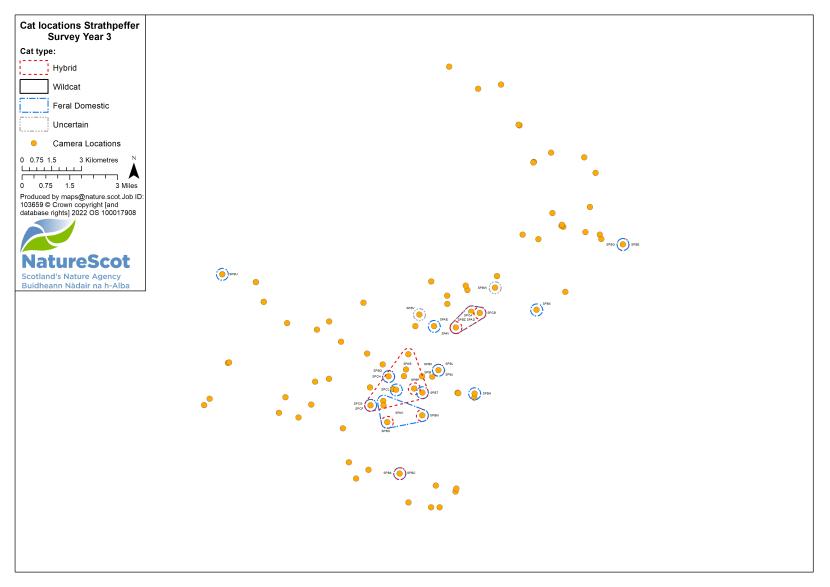
Figure A5.8. Distribution of camera traps and the locations of cats for the standardised survey in the Strathpeffer in winter 2017-2018.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
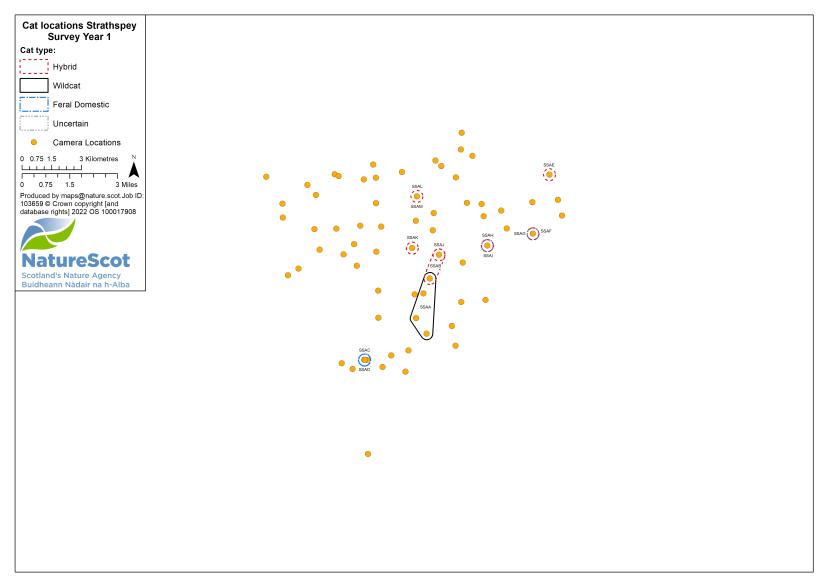
Figure A5.9. Distribution of camera traps and the locations of cats for the standardised survey in the Strathspey in winter 2015-2016.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
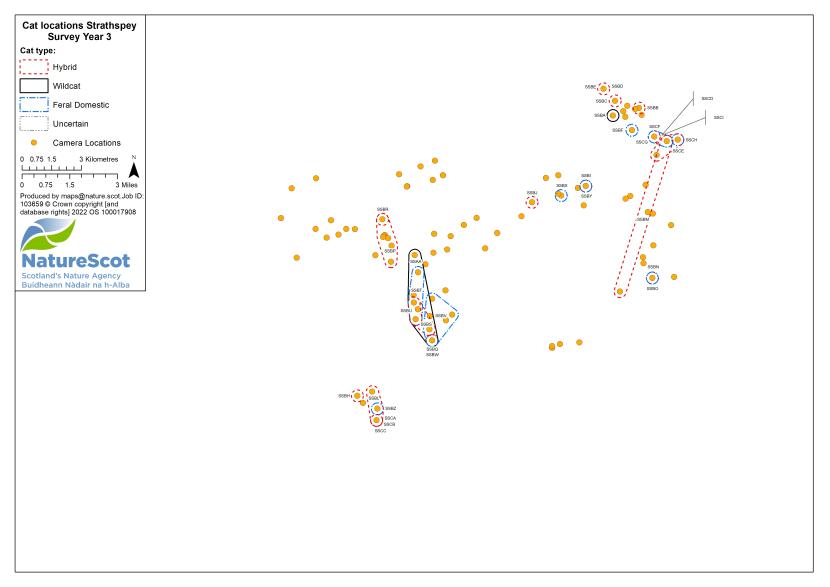
Figure A5.10. Distribution of camera traps and the locations of cats for the standardised survey in the Strathspey in winter 2017-2018.
Click for a full description
Cat locations are shown as minimum convex polygons buffered around the outlying cameras at which each individual was detected and are colour coded by type as assessed by pelage.
ANNEX 6 - EUNIS (European Nature information system) classification for cat habitat
|
EUNIS_code |
EUNIS_desc |
Note_ |
Suitable Wildcat habitat |
|
|---|---|---|---|---|
|
1 |
A |
Marine habitats |
Original EUNIS habitat |
0 |
|
2 |
B |
Coastal habitats |
Original EUNIS habitat |
0 |
|
3 |
C |
Inland surface waters |
Original EUNIS habitat |
0 |
|
5 |
E |
Grasslands and lands dominated by forbs, mosses or lichens |
Original EUNIS habitat |
1 |
|
7 |
G |
Woodland, forest and other wooded land |
Original EUNIS habitat |
1 |
|
11 |
J |
Constructed, industrial and other artificial habitats |
Original EUNIS habitat |
0 |
|
12 |
K |
Montane habitats |
Added due to lack of information |
0 |
|
101 |
A1 |
Littoral rock and other hard substrata |
Original EUNIS habitat |
0 |
|
102 |
A2 |
Littoral sediments |
Original EUNIS habitat |
0 |
|
201 |
B1 |
Coastal dunes and sandy shores |
Original EUNIS habitat |
0 |
|
202 |
B2 |
Coastal shingle |
Original EUNIS habitat |
0 |
|
203 |
B3 |
Rock cliffs, ledges and shores, including supralittoral |
Original EUNIS habitat |
0 |
|
401 |
D1 |
Raised and blanket bogs |
Original EUNIS habitat |
0 |
|
504 |
E4 |
Alpine, subalpine and extensive grasslands |
Extended to cover HAP 3 + 4 areas |
1 |
|
505 |
E5 |
Woodland fringes and clearings and tall forb stands |
Original EUNIS habitat |
1 |
|
602 |
F2 |
Arctic, alpine, subalpine and extensive scrub |
Extended to cover HAP 3 + 4 areas |
1 |
|
604 |
F4 |
Temperate shrub heathland |
Original EUNIS habitat |
1 |
|
701 |
G1 |
Broadleaved deciduous woodland |
Original EUNIS habitat |
1 |
|
703 |
G3 |
Coniferous woodland |
Original EUNIS habitat |
1 |
|
704 |
G4 |
Mixed deciduous and coniferous woodland |
Original EUNIS habitat |
1 |
|
706 |
G6 |
Exotic woodland and scrub |
Added |
1 |
|
901 |
I1 |
Arable land and market gardens |
Original EUNIS habitat |
1 |
|
902 |
I2 |
Cultivated areas of gardens and parks |
Original EUNIS habitat |
0 |
|
1103 |
J3 |
Extractive industrial sites |
Original EUNIS habitat |
0 |
|
1104 |
J4 |
Transport networks and other constructed hard-surfaced areas |
Original EUNIS habitat |
0 |
|
1105 |
J5 |
Highly artificial man-made waters and associated structures |
Original EUNIS habitat |
0 |
|
1201 |
K1 |
Montane vegetation |
Added due to lack of information |
0 |
|
6001 |
FA |
Hedgerows |
Original EUNIS habitat |
1 |
|
10205 |
A2.5 |
Coastal saltmarshes and saline reedbeds |
Original EUNIS habitat |
0 |
|
25028 |
X28 |
Blanket bog complexes |
Original EUNIS habitat |
0 |
|
50102 |
E1.2 |
Perennial calcareous grassland and basic steppes |
Original EUNIS habitat |
1 |
|
50206 |
E2.6 |
Agriculturally improved, re-seeded and heavily fertilised grassland, including sports fields and grass lawns |
Original EUNIS habitat |
1 |
|
50403 |
E4.3 |
Acid alpine, subalpine and extensive grassland |
Extended to cover HAP 3 + 4 areas |
1 |
|
50404 |
E4.4 |
Calcareous alpine, subalpine and extensive grassland |
Extended to cover HAP 3 + 4 areas |
1 |
|
50503 |
E5.3 |
Pteridium aquilinum fields |
Original EUNIS habitat |
1 |
|
50701 |
E7.1 |
Atlantic parkland |
Original EUNIS habitat |
1 |
|
60301 |
F3.1 |
Temperate thickets and scrub |
Original EUNIS habitat |
1 |
|
60401 |
F4.1 |
Wet heaths |
Original EUNIS habitat |
1 |
|
60402 |
F4.2 |
Dry heaths |
Original EUNIS habitat |
1 |
|
60902 |
F9.2 |
Salix carr and fen scrub |
Original EUNIS habitat |
1 |
|
70108 |
G1.8 |
Acidophilous Quercus-dominated woodland |
Original EUNIS habitat |
1 |
|
70109 |
G1.9 |
Non-riverine woodland with Betula, Populus tremula or Sorbus aucuparia |
Original EUNIS habitat |
1 |
|
70304 |
G3.4 |
Pinus sylvestris woodland south of the taiga |
Original EUNIS habitat |
1 |
|
70506 |
G5.6 |
Early-stage natural and semi-natural woodlands and regrowth |
Original EUNIS habitat |
1 |
|
70507 |
G5.7 |
Coppice and early-stage plantations |
Original EUNIS habitat |
1 |
|
70508 |
G5.8 |
Recently felled areas |
Original EUNIS habitat |
1 |
|
90102 |
I1.2 |
Mixed crops of market gardens and horticulture |
Original EUNIS habitat |
1 |
|
110402 |
J4.2 |
Road networks |
Original EUNIS habitat |
0 |
|
110403 |
J4.3 |
Rail networks |
Original EUNIS habitat |
0 |
|
110405 |
J4.5 |
Hard-surfaced areas of ports |
Original EUNIS habitat |
0 |
|
110407 |
J4.7 |
Constructed parts of cemeteries |
Original EUNIS habitat |
0 |
|
701001 |
G1.A |
Meso- and eutrophic Quercus, Carpinus, Fraxinus, Acer, Tilia, Ulmus and related woodland |
Original EUNIS habitat |
1 |
|
703005 |
G3.E |
Nemoral bog conifer woodland |
Original EUNIS habitat |
1 |
|
703006 |
G3.F |
Highly artificial coniferous plantations |
Original EUNIS habitat |
1 |
|
9940001 |
D2/D4/D5 |
Fens, mires, sedge- and reedbeds |
Combined due to lack of information |
0 |
|
9980001 |
H2/H3 |
Screes, inland cliffs, rock pavements and outcrops |
Combined due to lack of information |
1 |
|
11040201 |
J4.21 |
Motorway |
Added |
0 |
|
11040202 |
J4.22 |
A Road |
Added |
0 |
|
11040203 |
J4.23 |
B Roads, local streets and minor roads |
Added |
0 |
|
11040204 |
J4.24 |
Private roads, publicly accessible and restricted |
Added |
0 |
|
99110001 |
J1/J2 |
Buildings of cities, towns and villages / Low density buildings |
Combined |
1 |
|
99110002 |
J3/J6 |
Extractive industrial sites / Waste deposits |
Combined |
0 |
|
997010002 |
G1.4/G1.5 |
Broadleaved swamp woodland |
Combined |
1 |
ANNEX 7 - Cat encounter histories
The following tables provide details on the cats detected during the project within each area. These table may include the following: The unique identification code for each cat (UID); the approximate location within the Priority Area; which year(s) (where year 1 is 1st April 2015 to 31st March 2016) the cat was detected (D=detected, ND= not detected, T = trapped; N = neutered; C = collared with GPS tracker; X = found deceased); pelage score (the Kitchener 7PC score, range 7-21); coat type (appearance); cat type (wildcat, hybrid, domestic cat or uncertain); a procedure form number if the cat was sampled or trapped; whether the cat was identified during a standardised survey (‘in survey’); sex of cat. These table are also available for download.
|
Cat UID |
Location |
Year 1 |
Year 2 |
Year 3 |
Year 4 |
Year 5 |
Pelage score |
Coat type |
Cat type |
procedure form |
In survey |
|---|---|---|---|---|---|---|---|---|---|---|---|
|
ANAA |
Glen Clova/Prosen |
Y |
ND |
ND |
ND |
ND |
7 |
black |
domestic cat |
NA |
1 |
|
ANAB |
Glen Clova/Prosen |
Y |
ND |
ND |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANAC |
Glen Clova/Prosen |
Y |
ND |
ND |
ND |
ND |
NA |
tabby |
uncertain |
NA |
1 |
|
ANAD |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
16.6 |
tabby |
hybrid |
NA |
1 |
|
ANAE |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
14.1 |
tabby |
hybrid |
NA |
1 |
|
ANAF |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANAG |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
17 |
tabby |
wildcat |
NA |
1 |
|
ANAH |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
7 |
black |
domestic cat |
NA |
1 |
|
ANAI |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
8 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANAJ |
Glen Clova |
Y |
Y |
ND |
ND |
ND |
16.3 |
tabby |
hybrid |
NA |
1 |
|
ANAK |
Glen Clova |
Y |
Y |
Y |
Y |
Y |
8.2 |
blotchy tabby |
domestic cat |
754 |
1 |
|
ANAL |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
7 |
black |
domestic cat |
NA |
0 |
|
ANAM |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
16.9 |
tabby |
hybrid |
NA |
0 |
|
ANAN |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
15.3 |
tabby |
hybrid |
NA |
0 |
|
ANAO |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
0 |
|
ANAP |
Glen Clova |
Y |
ND |
ND |
ND |
ND |
16.9 |
juv. tabby |
uncertain |
NA |
0 |
|
ANAQ |
Glen Prosen |
Y |
ND |
ND |
ND |
ND |
16.6 |
tabby |
hybrid |
NA |
1 |
|
ANAR |
Glen Isla |
Y |
ND |
ND |
ND |
ND |
17.5 |
tabby |
wildcat |
NA |
1 |
|
ANAS |
Glen Prosen |
Y |
ND |
ND |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANAT |
Glen Prosen |
Y |
ND |
ND |
ND |
ND |
16 |
tabby |
hybrid |
NA |
1 |
|
ANAU |
Glen Isla |
Y |
Y |
Y |
Y |
ND |
10.5 |
blotchy tabby |
domestic cat |
755 |
1 |
|
ANAV |
Glen Isla |
Y |
Y |
Y |
Y |
Y |
16.5 |
tabby |
hybrid |
NA |
1 |
|
ANAW |
Glen Isla |
Y |
ND |
ND |
ND |
ND |
17.9 |
tabby |
wildcat |
NA |
1 |
|
ANAX |
Glen Isla |
Y |
Y |
Y |
Y |
ND |
15.8 |
tabby |
hybrid |
507 |
1 |
|
ANAY |
Glen Isla |
Y |
ND |
Y |
ND |
ND |
8.5 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANAZ |
Glen Isla |
Y |
Y |
ND |
Y |
ND |
16 |
tabby |
hybrid |
NA |
1 |
|
ANBA |
Glen Isla |
Y |
ND |
ND |
ND |
ND |
15.8 |
tabby |
hybrid |
NA |
1 |
|
ANBB |
Glen Isla |
Y |
ND |
Y |
ND |
ND |
11 |
tabby |
hybrid |
NA |
1 |
|
ANBC |
Glen Isla |
Y |
Y |
ND |
ND |
ND |
18 |
tabby |
wildcat |
NA |
1 |
|
ANBD |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
12.5 |
juv. tabby |
uncertain |
NA |
0 |
|
ANBE |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
12.5 |
juv. tabby |
uncertain |
NA |
0 |
|
ANBF |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
15.5 |
tabby |
hybrid |
753 |
0 |
|
ANBG |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
NA |
tabby |
uncertain |
NA |
0 |
|
ANBH |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
11.2 |
tabby |
hybrid |
NA |
0 |
|
ANBI |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
13.5 |
tabby |
hybrid |
758 |
0 |
|
ANBJ |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
10 |
blotchy tabby |
domestic cat |
752 |
0 |
|
ANBK |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
NA |
tabby |
hybrid |
NA |
0 |
|
ANBL |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
7 |
black |
domestic cat |
NA |
0 |
|
ANBM |
Glen Isla |
ND |
Y |
ND |
Y |
ND |
15 |
tabby |
hybrid |
759 |
0 |
|
ANBN |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
13.3 |
tabby |
hybrid |
NA |
0 |
|
ANBO |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
15.2 |
tabby |
hybrid |
NA |
0 |
|
ANBP |
Glen Clova |
ND |
Y |
ND |
ND |
ND |
NA |
tabby |
uncertain |
NA |
0 |
|
ANBQ |
Glen Isla |
ND |
Y |
Y |
ND |
ND |
13.8 |
tabby |
hybrid |
761 |
1 |
|
ANBS |
Glen Isla |
ND |
Y |
ND |
Y |
ND |
14 |
juv. tabby |
hybrid |
509 |
0 |
|
ANBT |
Glen Clova/Prosen |
ND |
Y |
ND |
ND |
ND |
14 |
tabby |
hybrid |
NA |
0 |
|
ANBU |
Glen Clova/Prosen |
ND |
Y |
ND |
ND |
ND |
NA |
tabby |
uncertain |
NA |
0 |
|
ANBV |
Glen Isla |
ND |
Y |
ND |
ND |
ND |
14 |
juv. tabby |
hybrid |
NA |
0 |
|
ANBW |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
17 |
tabby |
wildcat |
NA |
1 |
|
ANBX |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
10.5 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANBY |
Glen Clova |
ND |
ND |
Y |
Y |
ND |
7 |
black |
domestic cat |
NA |
1 |
|
ANBZ |
Glen Clova |
ND |
ND |
Y |
Y |
ND |
8.1 |
blotchy tabby |
domestic cat |
NA |
1 |
|
ANCA |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
14.5 |
tabby |
hybrid |
NA |
1 |
|
ANCB |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
NA |
black |
domestic cat |
NA |
1 |
|
ANCC |
Glen Clova |
ND |
ND |
Y |
Y |
Y |
19 |
tabby |
wildcat |
510,717 |
1 |
|
ANCD |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
15.3 |
tabby |
hybrid |
NA |
1 |
|
ANCE |
Glen Clova |
ND |
ND |
Y |
Y |
Y |
17.5 |
tabby |
wildcat |
511 |
1 |
|
ANCF |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
10.2 |
tabby |
domestic cat |
NA |
1 |
|
ANCG |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
7 |
black |
domestic cat |
NA |
1 |
|
ANCH |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
15.2 |
tabby |
hybrid |
NA |
1 |
|
ANCI |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
10.5 |
blotchy tabby |
hybrid |
NA |
1 |
|
ANCJ |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
16.1 |
tabby |
hybrid |
NA |
1 |
|
ANCK |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
16 |
tabby |
hybrid |
NA |
0 |
|
ANCL |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
14.8 |
tabby |
hybrid |
NA |
1 |
|
ANCM |
Glen Clova/Prosen |
ND |
ND |
Y |
ND |
ND |
14 |
tabby |
hybrid |
NA |
1 |
|
ANCN |
Glen Clova/Prosen |
ND |
ND |
Y |
ND |
ND |
12 |
white and tabby |
hybrid |
769 |
1 |
|
ANCO |
Glen Clova |
ND |
ND |
Y |
Y |
ND |
17 |
tabby |
wildcat |
NA |
1 |
|
ANCP |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
7.5 |
black |
domestic cat |
NA |
1 |
|
ANCQ |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
13.4 |
tabby |
hybrid |
NA |
1 |
|
ANCR |
Glen Isla |
ND |
ND |
Y |
Y |
Y |
16 |
tabby |
hybrid |
782 |
1 |
|
ANCS |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
17.3 |
tabby |
wildcat |
774 |
1 |
|
ANCT |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
13 |
tabby |
hybrid |
NA |
1 |
|
ANCU |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
13.5 |
tabby |
hybrid |
779 |
1 |
|
ANCV |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
10.5 |
tabby |
domestic cat |
778 |
1 |
|
ANCW |
Glen Isla |
ND |
ND |
Y |
Y |
ND |
16.3 |
tabby |
hybrid |
NA |
1 |
|
ANCX |
Glen Isla |
ND |
ND |
Y |
Y |
ND |
13.5 |
tabby |
hybrid |
776 |
1 |
|
ANCY |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
8 |
black |
domestic cat |
NA |
1 |
|
ANCZ |
Glen Isla |
ND |
ND |
Y |
Y |
ND |
11 |
blotchy tabby |
hybrid |
775 |
1 |
|
ANDA |
Glen Isla |
ND |
ND |
Y |
ND |
ND |
14.3 |
tabby |
hybrid |
NA |
1 |
|
ANDB |
Glen Prosen |
ND |
ND |
Y |
ND |
ND |
15 |
tabby |
hybrid |
NA |
1 |
|
ANDO |
Glen Clova/Prosen |
ND |
ND |
Y |
ND |
ND |
7 |
black and white |
domestic cat |
771 |
0 |
|
ANDP |
Glen Clova/Prosen |
ND |
ND |
Y |
ND |
ND |
7 |
black |
domestic cat |
770 |
0 |
|
ANDQ |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
NA |
tabby |
uncertain |
NA |
1 |
|
ANDR |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
NA |
tabby |
uncertain |
NA |
1 |
|
ANDS |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
NA |
tabby |
uncertain |
NA |
1 |
|
ANDT |
Glen Clova |
ND |
ND |
Y |
ND |
ND |
15.3 |
tabby |
hybrid |
NA |
1 |
|
ANDU |
Glen Prosen |
ND |
ND |
Y |
ND |
ND |
NA |
tabby |
uncertain |
NA |
1 |
|
ANDC |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
8.5 |
black |
domestic cat |
508 |
0 |
|
ANDD |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
15.8 |
tabby |
hybrid |
509 |
0 |
|
ANDE |
Glen Clova |
ND |
ND |
ND |
Y |
ND |
7 |
juv. black |
domestic cat |
513 |
0 |
|
ANDF |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
7.5 |
black |
domestic cat |
514 |
0 |
|
ANDG |
Glen Prosen |
ND |
ND |
ND |
Y |
ND |
15.4 |
tabby |
hybrid |
NA |
0 |
|
ANDH |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
0 |
|
ANDI |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
12 |
tabby |
hybrid |
NA |
0 |
|
ANDJ |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
9 |
black |
hybrid |
NA |
0 |
|
ANDK |
Glen Clova/Prosen |
ND |
ND |
ND |
Y |
ND |
7 |
black |
domestic cat |
NA |
0 |
|
ANDL |
Glen Clova |
ND |
ND |
ND |
Y |
ND |
7 |
juv. black |
domestic cat |
NA |
0 |
|
ANDM |
Glen Clova |
ND |
ND |
ND |
Y |
ND |
8 |
blotchy tabby |
domestic cat |
NA |
0 |
|
ANDN |
Glen Isla |
ND |
ND |
ND |
Y |
ND |
16.5 |
tabby |
hybrid |
NA |
0 |
|
ANDV |
Glen Isla |
ND |
ND |
ND |
ND |
Y |
NA |
blotchy tabby |
hybrid |
NA |
0 |
|
ANDW |
Glen Isla |
ND |
ND |
ND |
ND |
Y |
NA |
tabby |
hybrid |
NA |
0 |
|
ANDX |
Glen Doll |
ND |
ND |
ND |
ND |
Y |
17 |
tabby |
wildcat |
517 |
0 |
|
ANDY |
Glen Isla |
ND |
ND |
ND |
ND |
Y |
18 |
tabby |
wildcat |
788 |
0 |
|
Cat UID |
Year 2 |
Year 3 |
Year 4 |
Pelage score |
Coat type |
Cat type |
procedure form |
In survey |
sex |
|---|---|---|---|---|---|---|---|---|---|
|
MOAA |
D |
ND |
X |
19.5 |
tabby |
wildcat |
865 |
1 |
male |
|
MOAB |
D |
D |
T |
17 |
tabby |
wildcat |
873 |
1 |
male |
|
MOAC |
D |
D |
T |
16.8 |
tabby |
hybrid |
872 |
1 |
male |
|
MOAD |
D |
ND |
ND |
14 |
tabby |
hybrid |
NA |
1 |
female |
|
MOAD2 |
D |
D |
T |
18 |
tabby |
wildcat |
870 |
0 |
female |
|
MOAE |
D |
ND |
ND |
14.5 |
tabby |
hybrid |
NA |
1 |
female |
|
MOAF |
N |
ND |
ND |
11 |
blotchy tabby |
hybrid |
360 |
1 |
female |
|
MOAG |
D |
ND |
ND |
11 |
blotchy tabby |
hybrid |
NA |
1 |
suspected female |
|
MOAH |
N |
ND |
ND |
8 |
black |
domestic cat |
361 |
1 |
male |
|
MOAI |
D |
ND |
ND |
8.4 |
blotchy tabby |
domestic cat |
NA |
0 |
male |
|
MOAJ |
N |
ND |
ND |
9 |
ginger |
domestic cat |
359 |
0 |
female |
|
MOAK |
D |
ND |
ND |
10 |
ginger |
domestic cat |
NA |
1 |
male |
|
MOAL |
D |
ND |
ND |
7 |
black |
domestic cat |
NA |
1 |
NA |
|
MOAO |
D |
ND |
ND |
10 |
blotchy tabby |
domestic cat |
NA |
1 |
NA |
|
MOAP |
D |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
0 |
NA |
|
MOAQ |
TX |
ND |
ND |
7 |
grey blotchy tabby |
domestic cat |
356 |
0 |
NA |
|
MOAS |
N |
ND |
ND |
11.7 |
blotchy tabby |
hybrid |
362 |
0 |
NA |
|
MOAT |
ND |
D |
ND |
7 |
spotted tabby |
domestic cat |
NA |
0 |
NA |
|
MOAV |
ND |
X |
ND |
12.8 |
blotchy tabby |
hybrid |
866 |
0 |
NA |
|
MOAY |
ND |
D |
ND |
7 |
white and black |
domestic cat |
NA |
0 |
NA |
|
MOBB |
ND |
N |
ND |
NA |
tabby |
hybrid |
951 |
0 |
male |
N = neutered, T = trapped, X = deceased
|
Cat UID |
Year 1 |
Year 2 |
Coat type |
Cat type |
Pelage score |
|---|---|---|---|---|---|
|
SAAA |
D |
ND |
tabby |
hybrid |
15.2 |
|
SAAC |
D |
D |
black |
domestic cat |
8 |
|
SAAD |
D |
ND |
black |
domestic cat |
7 |
|
SAAE |
D |
ND |
ginger |
domestic cat |
8.4 |
|
SAAF |
D |
ND |
tabby |
hybrid |
15.4 |
|
SAAH |
D |
ND |
tabby |
hybrid |
11.1 |
|
SAAI |
D |
ND |
tabby |
hybrid |
14 |
|
SAAJ |
D |
ND |
black |
domestic cat |
7 |
|
SAAK |
D |
D |
black |
domestic cat |
7 |
|
SAAL |
D |
ND |
tabby |
hybrid |
14 |
|
SAAM |
ND |
N |
ginger |
domestic cat |
7 |
|
SAAN |
ND |
N |
black |
domestic cat |
7 |
|
SAAO |
ND |
N |
ginger |
domestic cat |
9 |
|
SAAP |
ND |
N |
black |
domestic cat |
7 |
|
SAAQ |
ND |
N |
tabby |
hybrid |
11.2 |
N = neutered
|
CAT UID |
Year 1 |
Year 2 |
Year 3 |
Year 4 |
Year 5 |
Pelage score |
Coat type |
Cat type |
form no. |
Procedure |
|---|---|---|---|---|---|---|---|---|---|---|
|
SBAA |
D |
T |
D |
D |
D |
12 |
white and tabby |
hybrid |
318 |
TNVR |
|
SBAI |
D |
D |
T |
ND |
ND |
15 |
tabby |
hybrid |
223 |
TNVR |
|
SBAK |
D |
D |
D |
ND |
ND |
18 |
tabby |
wildcat |
256 |
Hair sample |
|
SBAN |
D |
T |
D |
ND |
ND |
11.7 |
white and tabby |
hybrid |
319 |
TNVR |
|
SBAO |
D |
D |
D |
D |
ND |
14 |
tabby |
hybrid |
NA |
NA |
|
SBAP |
D |
D |
D |
D |
ND |
14 |
tabby |
hybrid |
613 |
Hair sample |
|
SBAQ |
D |
D |
D |
ND |
D |
9.8 |
black |
domestic cat |
407 |
Hair sample |
|
SBAR |
D |
D |
D |
D |
D |
8.4 |
black |
domestic cat |
262 |
Hair sample |
|
SBAT |
D |
D |
T |
D |
ND |
8.4 |
white and tabby |
domestic cat |
221, 368 |
TNVR |
|
SBAV |
D |
D |
D |
ND |
D |
7 |
black and white |
domestic cat |
NA |
NA |
|
SBAB |
D |
D |
ND |
ND |
ND |
19 |
tabby |
wildcat |
251 |
Hair sample |
|
SBAC |
D |
D |
ND |
ND |
ND |
17 |
tabby |
wildcat |
260 |
Hair sample |
|
SBAD |
D |
D |
ND |
ND |
ND |
18 |
tabby |
wildcat |
258 |
Hair sample |
|
SBAE |
D |
D |
ND |
ND |
ND |
17.2 |
tabby |
wildcat |
259 |
Hair sample |
|
SBAG |
D |
D |
ND |
ND |
ND |
<10.5 |
ginger/white |
domestic cat |
NA |
NA |
|
SBAY |
D |
T |
ND |
D |
ND |
8.2 |
LH blotchy tabby |
domestic cat |
320 |
TNVR |
|
SBAH |
D |
ND |
D |
D |
D |
11 |
white and tabby |
hybrid |
NA |
NA |
|
SBAF |
D |
ND |
ND |
ND |
ND |
9 |
blotchy tabby |
domestic cat |
NA |
NA |
|
SBAJ |
D |
ND |
ND |
ND |
ND |
8.7 |
tortoiseshell and tabby |
domestic cat |
252 |
Hair sample |
|
SBAL |
D |
ND |
ND |
ND |
ND |
15 |
tabby |
hybrid |
NA |
NA |
|
SBAM |
D |
ND |
ND |
ND |
ND |
14 |
tabby |
uncertain |
NA |
NA |
|
SBAS |
D |
ND |
ND |
ND |
ND |
7 |
black |
domestic cat |
NA |
NA |
|
SBAU |
D |
ND |
ND |
ND |
ND |
10.5 |
tabby |
domestic cat |
NA |
NA |
|
SBAW |
D |
ND |
ND |
ND |
ND |
9.8 |
tabby |
domestic cat |
NA |
NA |
|
SBAX |
D |
ND |
ND |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
NA |
|
SBAZ |
D |
ND |
ND |
ND |
ND |
8 |
LH blotchy tabby |
domestic cat |
NA |
NA |
|
SBBF |
ND |
D |
T |
ND |
ND |
16.5 |
tabby |
hybrid |
222 |
TNVR |
|
SBBO |
ND |
T |
D |
D |
ND |
9 |
black |
hybrid |
323 |
TNVR |
|
SBBR |
ND |
D |
D |
D |
ND |
7 |
black |
domestic cat |
NA |
NA |
|
SBCP |
ND |
D |
ND |
ND |
ND |
19.8 |
tabby |
wildcat |
NA |
NA |
|
SBCR |
ND |
D |
ND |
ND |
ND |
16 |
tabby |
hybrid |
NA |
NA |
|
SBCS |
ND |
D |
ND |
ND |
ND |
16 |
tabby |
hybrid |
NA |
NA |
|
SBCT |
ND |
X |
ND |
ND |
ND |
10 |
NA |
domestic cat |
366 |
RTA |
|
SBCU |
ND |
D |
ND |
ND |
ND |
16 |
tabby |
hybrid |
NA |
NA |
|
SBCV |
ND |
D |
ND |
ND |
ND |
14 |
NA |
hybrid |
NA |
NA |
|
SBCW |
ND |
D |
ND |
ND |
ND |
12 |
NA |
hybrid |
NA |
NA |
|
SBCX |
ND |
D |
ND |
ND |
ND |
NA |
tabby |
uncertain |
NA |
NA |
|
SBCY |
ND |
D |
ND |
ND |
ND |
NA |
tabby |
hybrid |
NA |
NA |
|
SBCZ |
ND |
T |
ND |
ND |
ND |
12 |
NA |
hybrid |
322 |
TNVR |
|
SBDA |
ND |
D |
ND |
ND |
ND |
9 |
white and tabby |
domestic cat |
NA |
NA |
|
SBBE |
ND |
ND |
C |
C |
D |
17 |
tabby |
wildcat |
292, 299 |
Trap |
|
SBBG |
ND |
ND |
D |
C |
D |
18 |
tabby |
wildcat |
0481, 0298 |
0481, 0298 |
|
SBBI |
ND |
ND |
C |
C |
C |
17 |
tabby |
wildcat |
295, 231 |
Trap |
|
SBBB |
ND |
ND |
D |
ND |
ND |
15 |
tabby |
hybrid |
NA |
NA |
|
SBBC |
ND |
ND |
D |
ND |
ND |
7 |
LH tabby |
domestic cat |
NA |
NA |
|
SBBD |
ND |
ND |
D |
ND |
ND |
7 |
tabby |
domestic cat |
NA |
NA |
|
SBBH |
ND |
ND |
D |
ND |
D |
16 |
tabby |
hybrid |
NA |
NA |
|
SBBJ |
ND |
ND |
D |
ND |
ND |
15.5 |
tabby |
hybrid |
452 |
Hair sample |
|
SBBK |
ND |
ND |
D |
ND |
ND |
14 |
tabby |
hybrid |
NA |
NA |
|
SBBL |
ND |
ND |
D |
ND |
ND |
17 |
tabby |
wildcat |
NA |
NA |
|
SBBM |
ND |
ND |
D |
ND |
ND |
7 |
black |
domestic cat |
NA |
NA |
|
SBBN |
ND |
ND |
D |
ND |
ND |
15.8 |
tabby |
hybrid |
NA |
NA |
|
SBBQ |
ND |
ND |
D |
ND |
ND |
12 |
tabby |
hybrid |
NA |
NA |
|
SBBS |
ND |
ND |
D |
ND |
ND |
8 |
LH black |
domestic cat |
NA |
NA |
|
SBBT |
ND |
ND |
D |
ND |
ND |
13 |
tabby |
hybrid |
NA |
NA |
|
SBBU |
ND |
ND |
T |
ND |
ND |
9.8 |
tabby |
domestic cat |
291 |
TNVR |
|
SBBV |
ND |
ND |
T |
D |
ND |
8 |
black |
domestic cat |
290 |
TNVR |
|
SBBW |
ND |
ND |
D |
ND |
ND |
15.8 |
tabby |
hybrid |
NA |
NA |
|
SBBX |
ND |
ND |
D |
ND |
ND |
7 |
white and tabby |
domestic cat |
NA |
NA |
|
SBBY |
ND |
ND |
T |
ND |
ND |
13 |
tabby |
hybrid |
293 |
TNVR |
|
SBBZ |
ND |
ND |
D |
ND |
ND |
8 |
black |
domestic cat |
NA |
NA |
|
SBCA |
ND |
ND |
T |
ND |
ND |
14 |
tabby |
hybrid |
294 |
TNVR |
|
SBCB |
ND |
ND |
D |
ND |
D |
7 |
black |
domestic cat |
NA |
NA |
|
SBCC |
ND |
ND |
D |
ND |
ND |
16.5 |
tabby |
hybrid |
NA |
NA |
|
SBCD |
ND |
ND |
D |
ND |
D |
15.2 |
tabby |
hybrid |
NA |
NA |
|
SBCE |
ND |
ND |
D |
ND |
ND |
14.7 |
tabby |
hybrid |
451 |
Hair sample |
|
SBCF |
ND |
ND |
D |
D |
ND |
7 |
ginger/white |
domestic cat |
NA |
NA |
|
SBCG |
ND |
ND |
D |
ND |
ND |
7 |
black |
domestic cat |
NA |
NA |
|
SBCH |
ND |
ND |
D |
ND |
ND |
7 |
blotchy tabby |
domestic cat |
NA |
NA |
|
SBCI |
ND |
ND |
D |
ND |
ND |
12.8 |
white and tabby |
hybrid |
NA |
NA |
|
SBCJ |
ND |
ND |
D |
ND |
ND |
10 |
tabby |
domestic cat |
NA |
NA |
|
SBCN |
ND |
ND |
ND |
C |
T |
14.5 |
tabby |
hybrid |
232 |
Trap then TNVR |
|
SBCO |
ND |
ND |
ND |
T |
D |
16 |
tabby |
hybrid |
469, 652, 707 |
652 & 707 are samples |
|
SBCQ |
ND |
ND |
ND |
C |
T |
15.5 |
tabby |
hybrid |
482, 655(?) |
Trap |
|
SBCK |
ND |
ND |
ND |
D |
D |
7 |
black |
domestic cat |
NA |
NA |
|
SBCL |
ND |
ND |
ND |
D |
D |
NA |
white and tabby |
domestic cat |
NA |
NA |
|
SBCM |
ND |
ND |
ND |
D |
ND |
13.3 |
white and tabby |
hybrid |
NA |
NA |
|
SBDF |
ND |
ND |
ND |
D |
D |
8 |
ginger/white |
domestic cat |
NA |
NA |
|
SBDL |
ND |
ND |
ND |
D |
ND |
12.6 |
tabby |
hybrid |
NA |
NA |
|
SBDM |
ND |
ND |
ND |
D |
D |
8 |
black |
domestic cat |
NA |
NA |
|
SBDB |
ND |
ND |
ND |
ND |
D |
8 |
black and white |
domestic cat |
NA |
NA |
|
SBDC |
ND |
ND |
ND |
ND |
T |
14 |
tabby |
hybrid |
240 |
TNVR |
|
SBDD |
ND |
ND |
ND |
ND |
T |
13 |
tabby |
hybrid |
244 |
TNVR |
|
SBDE |
ND |
ND |
ND |
ND |
T |
15 |
tabby |
hybrid |
242 |
TNVR |
|
SBDG |
ND |
ND |
ND |
ND |
D |
7 |
black and white |
domestic cat |
NA |
NA |
|
SBDH |
ND |
ND |
ND |
ND |
D |
12.8 |
tabby |
hybrid |
NA |
NA |
|
SBDI |
ND |
ND |
ND |
ND |
T |
12 |
tabby |
hybrid |
249 |
TNVR |
|
SBDJ |
ND |
ND |
ND |
ND |
T |
9.3 |
tabby |
domestic cat |
250 |
TNVR |
|
SBDK |
ND |
ND |
ND |
ND |
D |
15.2 |
tabby |
hybrid |
484 |
Blood sample |
|
SBDN |
ND |
ND |
ND |
ND |
D |
16 |
tabby |
hybrid |
NA |
NA |
|
SBDO |
ND |
ND |
ND |
ND |
D |
13 |
tabby |
hybrid |
NA |
NA |
|
SBDP |
ND |
ND |
ND |
ND |
D |
11.7 |
tabby |
hybrid |
NA |
NA |
|
SBDQ |
ND |
ND |
ND |
ND |
D |
7 |
tabby |
domestic cat |
NA |
NA |
|
SBDR |
ND |
ND |
ND |
ND |
D |
8 |
white and tabby |
domestic cat |
NA |
NA |
|
SBDS |
ND |
ND |
ND |
ND |
D |
11.2 |
tabby |
hybrid |
NA |
NA |
|
SBDT |
ND |
ND |
ND |
ND |
D |
7 |
black and white |
domestic cat |
NA |
NA |
|
SBDU |
ND |
ND |
ND |
ND |
D |
7.5 |
ginger/white |
domestic cat |
NA |
NA |
|
SBDV |
ND |
ND |
ND |
ND |
D |
7.5 |
white and blotchy tabby |
domestic cat |
NA |
NA |
|
SBDW |
ND |
ND |
ND |
D |
D |
17 |
tabby |
hybrid |
654 |
Hair sample |
|
SBDX |
ND |
ND |
ND |
ND |
D |
8.4 |
blotchy tabby |
domestic cat |
NA |
NA |
|
SBDY |
ND |
ND |
ND |
ND |
D |
8.2 |
blotchy tabby |
domestic cat |
NA |
NA |
|
SBDZ |
ND |
ND |
ND |
ND |
D |
17 |
tabby |
wildcat |
236 |
Blood sample |
|
SBEA |
ND |
ND |
ND |
ND |
T |
10 |
blotchy tabby |
domestic cat |
239 |
TNVR |
|
SBEB |
ND |
ND |
ND |
D |
ND |
NA |
Unknown |
Unknown |
65 |
Saliva sample |
|
Cat UID |
Year 1 |
Year 3 |
Phenotype |
Pelage score |
In survey |
|---|---|---|---|---|---|
|
SPAA |
D |
ND |
wildcat |
17.5 |
1 |
|
SPAB |
D |
D |
hybrid |
14.5 |
1 |
|
SPAC |
D |
ND |
hybrid |
16 |
0 |
|
SPAD |
D |
D |
domestic cat |
7 |
1 |
|
SPAE |
D |
D |
domestic cat |
7 |
1 |
|
SPAF |
D |
ND |
domestic cat |
7 |
1 |
|
SPAG |
D |
ND |
wildcat |
18 |
1 |
|
SPAH |
D |
ND |
hybrid |
14 |
0 |
|
SPAI |
D |
ND |
wildcat |
17.5 |
1 |
|
SPAJ |
D |
ND |
wildcat |
17 |
1 |
|
SPAK |
D |
D |
domestic cat |
7 |
1 |
|
SPAL |
D |
ND |
domestic cat |
6.4 |
1 |
|
SPAM |
D |
ND |
domestic cat |
7 |
1 |
|
SPAN |
D |
ND |
domestic cat |
9 |
1 |
|
SPAO |
D |
ND |
wildcat |
18 |
1 |
|
SPAP |
D |
ND |
domestic cat |
7 |
1 |
|
SPAQ |
D |
ND |
hybrid |
14 |
1 |
|
SPAR |
D |
ND |
domestic cat |
7 |
1 |
|
SPAT |
D |
ND |
hybrid |
12.8 |
1 |
|
SPAU |
D |
ND |
domestic cat |
10 |
1 |
|
SPAV |
D |
D |
hybrid |
13 |
1 |
|
SPAW |
D |
ND |
domestic cat |
7 |
1 |
|
SPAX |
D |
ND |
domestic cat |
7 |
1 |
|
SPAY |
D |
ND |
hybrid |
15.4 |
1 |
|
SPAZ |
D |
ND |
domestic cat |
7 |
0 |
|
SPBA |
D |
ND |
domestic cat |
7 |
1 |
|
SPBC |
ND |
D |
hybrid |
14.3 |
1 |
|
SPBD |
ND |
D |
domestic cat |
7 |
0 |
|
SPBE |
ND |
D |
hybrid |
16.6 |
1 |
|
SPBF |
ND |
D |
uncertain |
7 |
0 |
|
SPBG |
ND |
D |
domestic cat |
7 |
1 |
|
SPBH |
ND |
D |
domestic cat |
7 |
1 |
|
SPBI |
ND |
D |
domestic cat |
7 |
1 |
|
SPBJ |
ND |
D |
hybrid |
11.5 |
1 |
|
SPBK |
ND |
D |
domestic cat |
7 |
1 |
|
SPBL |
ND |
D |
domestic cat |
7 |
1 |
|
SPBM |
ND |
D |
hybrid |
14.7 |
1 |
|
SPBO |
ND |
D |
hybrid |
16.9 |
1 |
|
SPBP |
ND |
D |
hybrid |
16.8 |
1 |
|
SPBQ |
ND |
D |
hybrid |
10.8 |
1 |
|
SPBR |
ND |
D |
hybrid |
15.5 |
0 |
|
SPBS |
ND |
D |
domestic cat |
7 |
0 |
|
SPBT |
ND |
D |
domestic cat |
7 |
1 |
|
SPBU |
ND |
D |
domestic cat |
7 |
1 |
|
SPBV |
ND |
D |
uncertain |
7 |
1 |
|
SPBW |
ND |
D |
uncertain |
7 |
1 |
|
SPBX |
ND |
D |
domestic cat |
10 |
1 |
|
SPBZ |
ND |
D |
hybrid |
15.5 |
1 |
|
SPCA |
ND |
D |
hybrid |
14 |
1 |
|
SPCB |
ND |
D |
hybrid |
14 |
1 |
|
SPCD |
ND |
D |
hybrid |
13.1 |
0 |
|
SPCE |
ND |
D |
hybrid |
14 |
0 |
|
SPCF |
ND |
D |
hybrid |
14 |
1 |
|
SPCG |
ND |
D |
domestic cat |
7 |
1 |
|
SPCH |
ND |
D |
domestic cat |
7 |
1 |
|
SPCI |
ND |
D |
domestic cat |
7 |
0 |
|
SPCJ |
ND |
D |
domestic cat |
7 |
1 |
|
SPCK |
ND |
D |
domestic cat |
10.5 |
0 |
|
SPCL |
ND |
D |
domestic cat |
7 |
0 |
|
SPCM |
ND |
D |
domestic cat |
7 |
0 |
|
SPCN |
ND |
D |
domestic cat |
7 |
0 |
|
SPCO |
ND |
D |
domestic cat |
7 |
0 |
|
SPCP |
ND |
D |
domestic cat |
7 |
0 |
|
SPCR |
ND |
D |
domestic cat |
7 |
0 |
|
SPCS |
ND |
D |
domestic cat |
7 |
0 |
|
SPCT |
ND |
D |
domestic cat |
7 |
0 |
|
Cat UID |
Year 1 |
Year 3 |
Coat type |
Cat type |
pelage score |
In survey |
|---|---|---|---|---|---|---|
|
SSAA |
D |
D |
tabby |
wildcat |
17 |
1 |
|
SSAB |
D |
ND |
tabby |
hybrid |
10.8 |
1 |
|
SSAC |
D |
ND |
tabby |
domestic cat |
10.5 |
1 |
|
SSAD |
D |
ND |
black |
domestic cat |
7 |
1 |
|
SSAE |
D |
ND |
tabby |
hybrid |
16.3 |
1 |
|
SSAF |
D |
ND |
tabby |
domestic cat |
7 |
1 |
|
SSAG |
D |
ND |
tabby |
hybrid |
16.8 |
1 |
|
SSAH |
D |
ND |
tabby |
domestic cat |
7 |
1 |
|
SSAI |
D |
ND |
tabby |
hybrid |
10.9 |
1 |
|
SSAJ |
D |
ND |
tabby |
hybrid |
16.2 |
1 |
|
SSAK |
D |
ND |
tabby |
hybrid |
15.8 |
1 |
|
SSAL |
D |
ND |
tabby |
hybrid |
13.1 |
1 |
|
SSAM |
D |
ND |
grey tabby |
hybrid |
16.9 |
1 |
|
SSBA |
ND |
D |
tabby |
wildcat |
17 |
1 |
|
SSBB |
ND |
D |
tabby |
hybrid |
15 |
1 |
|
SSBC |
ND |
D |
white and tabby |
hybrid |
14 |
1 |
|
SSBD |
ND |
D |
tabby |
hybrid |
11.7 |
1 |
|
SSBE |
ND |
D |
tabby |
hybrid |
15.8 |
1 |
|
SSBF |
ND |
D |
ginger |
domestic cat |
9 |
1 |
|
SSBG |
ND |
D |
tabby |
hybrid |
16.5 |
0 |
|
SSBH |
ND |
D |
tabby |
hybrid |
15 |
1 |
|
SSBI |
ND |
D |
tabby |
hybrid |
12 |
1 |
|
SSBJ |
ND |
D |
tabby |
hybrid |
15.6 |
1 |
|
SSBK |
ND |
D |
tabby |
hybrid |
14.9 |
0 |
|
SSBL |
ND |
D |
tabby |
hybrid |
10.8 |
1 |
|
SSBM |
ND |
D |
white and tabby |
hybrid |
13 |
1 |
|
SSBN |
ND |
D |
tabby |
hybrid |
15.5 |
1 |
|
SSBO |
ND |
D |
white and tabby |
domestic cat |
7.5 |
1 |
|
SSBP |
ND |
D |
tabby |
hybrid |
14 |
1 |
|
SSBQ |
ND |
D |
black |
domestic cat |
7.8 |
1 |
|
SSBR |
ND |
D |
tabby |
hybrid |
11.1 |
1 |
|
SSBS |
ND |
D |
tabby |
hybrid |
11 |
1 |
|
SSBT |
ND |
D |
black |
domestic cat |
9 |
1 |
|
SSBU |
ND |
D |
tabby |
hybrid |
11.3 |
1 |
|
SSBV |
ND |
D |
ginger |
domestic cat |
8.5 |
1 |
|
SSBW |
ND |
D |
tabby |
hybrid |
16 |
1 |
|
SSBX |
ND |
D |
tortoiseshell |
domestic cat |
7 |
1 |
|
SSBY |
ND |
D |
black |
domestic cat |
7 |
1 |
|
SSBZ |
ND |
D |
white and tabby |
domestic cat |
9.3 |
1 |
|
SSCA |
ND |
D |
tabby |
hybrid |
11.3 |
1 |
|
SSCB |
ND |
D |
white and tabby |
hybrid |
13.5 |
1 |
|
SSCC |
ND |
D |
tabby |
hybrid |
11.2 |
1 |
|
SSCD |
ND |
D |
tabby |
domestic cat |
9.3 |
1 |
|
SSCE |
ND |
D |
tabby |
hybrid |
11.4 |
1 |
|
SSCF |
ND |
D |
tabby |
hybrid |
12.4 |
1 |
|
SSCG |
ND |
D |
tabby |
hybrid |
12.3 |
1 |
|
SSCH |
ND |
D |
ginger |
domestic cat |
7 |
1 |
|
SSCI |
ND |
D |
ginger |
domestic cat |
9 |
1 |
For further information on this report please contact:
Roo Campbell, NatureScot, Great Glen House, Leachkin Road, Inverness, IV3 8NW.
Telephone: 01463 725130
Email: [email protected]
ISBN: 978-1-78391-976-5
This publication is part of a series of specialist reports on the work of the Scottish Wildcat Action (SWA) project that ran from 2015-2020. The work was led and steered by a partnership of organisations: Cairngorms National Park Authority, Forestry and Land Scotland, National Museums Scotland, Scottish Land & Estates, NatureScot, Scottish Wildlife Trust, The National Trust for Scotland, The Royal (Dick) School of Veterinary Studies (R(D)SVS), The Royal Zoological Society of Scotland, The Scottish Gamekeepers Association, The Scottish Government, Wildlife Conservation Research Unit (WildCRU).
The many other partners and funders are listed in the SWA Summary report.
For more information, including access to the other reports, contact Martin Gaywood at NatureScot.
This report, or any part of it, should not be reproduced without the permission of NatureScot or the relevant authors. This permission will not be withheld unreasonably. The views expressed by the author(s) of this report should not be taken as the views and policies of NatureScot. © NatureScot 2023.
Document downloads
Last updated:






