NatureScot Research Report 1275 - Time series analysis to determine change in maerl beds
Year of publication: 2024
Authors: Doggett, M., Morris-Webb, L. and Baldock, L.
Cite as: Doggett, M., Morris-Webb, L. and Baldock, L. 2024. Time series analysis to determine change in maerl beds. NatureScot Research Report 1275.
Keywords
maerl; management; fishing; monitoring; recovery; impact; drop imagery
Background
Maerl beds comprise a group of calcareous red algae which can cover extensive areas and support diverse benthic communities and are considered a ‘non-renewable resource’ due to their extremely slow growth and sensitivity to disturbance. Maerl beds are an OSPAR threatened and declining habitat across Europe, a Priority Marine Feature (PMF) and protected within 11 Marine Protected Areas (MPA) in Scotland. This project was commissioned by NatureScot to assess best practice for monitoring maerl beds and to suggest methods to detect maerl bed recovery and assess the effects of management measures over relatively short time frames (years rather than decades). The study focused on drop imagery data from two case study areas: the first from the sheltered beds around the Isle of Arran, now South Arran MPA, and the second from more exposed open coast beds off the west coast of Scotland, now Wester Ross MPA. This report explores the possibility of analysing semi-quantitative species data to determine community change.
To inform future monitoring and assessment, NatureScot sought input from experts to develop and pilot analytical techniques to describe variation in maerl beds, using a subset of existing video and imagery data collected during MPA monitoring surveys. The long-term aim is to use the methods developed to detect maerl bed recovery and assess the effects of management measures. Results presented here are for proof-of-concept only.
Main findings
The literature review of previous maerl research found that despite a long-standing interest in the protection and monitoring of maerl beds, there is no published evidence of the success of any long-term monitoring approach backed up by repeat surveys to evaluate change. The change of functional diversity metrics of benthic communities subject to different pressures has potential in helping to assess impacts of different management scenarios.
This report goes on to assess the case study data from around the Isle of Arran and Wester Ross. Semi-quantitative (SACFOR) taxa / taxon-group data derived from drop imagery were converted using methods suggested by Strong and Johnson (2020) and proved robust for multivariate analyses.
Analysis of case study data – Arran 2014 & 2018
- Significant differences in the converted SACFOR community data were found between 2014 and 2018 from both Pladda and Holy Isle around Arran. Differences were driven by changes in abundance of some taxa (e.g. kelp and tunicates) between years rather than a significant change in the list of taxa. Given that the results were based on only two years of data it is not possible to attribute the change to either natural or anthropogenic influences.
Analysis of case study data – Wester Ross
- Available transect data available for the maerl habitats were deemed insufficient for statistical temporal or spatial comparison due to differences in data acquisition methods, data seasonality, differences in maerl bed type and insufficient sample replication.
- Two almost contiguous transects were identified in the Wester Ross MPA; one surveyed in November 2014 and the second in July 2018. Qualitative observations highlighted differences between these surveys that could help guide future assessments; the abundance of algae was higher during summer, confirming that seasonal timing of surveys is critical; and maerl bed physical structure differed between transects.
Finally the report provides recommendations for future monitoring survey design using drop imagery to provide sufficient sample replication to detect changes in community composition in selected maerl beds. The recommendations address the statistical problems of spatial autocorrelation and aim to determine real ecological change compared to variation inherent in the data. The following recommendations are made:
- Standardise data collection and analytical methods to reduce ‘procedural noise’ in data.
- A maerl-specific stratified, random sampling design is proposed.
- Transects >100 m long of video and high-quality stills for analysis can generate empirical and SACFOR data. Ideally two methods: one broader scale forward-facing imagery analysed qualitatively, and one using finer scale downward-facing images analysed robustly, will assist in providing context for the resulting data.
- Results obtained here indicate that in many instances 10 still images per transect can provide a good representation of the benthic community for maerl habitats.
- Statistical power of univariate data from maerl biotopes around Holy Isle explored the number of replicates required to be to detect change at various probabilities for nine key ‘indicator variables’. For some common taxa an order of magnitude change may be detectable from 4-9 transects per maerl bed, for other more rare or cryptic taxa this would rise to 25-200+ transects for sufficient statistical power.
- Annual, standardised sampling over a minimum of five years will enhance understanding of variability in the data and may then provide more robust evidence pertaining to any changes observed following the implementation of management measures.
- Procedures and protocols should be developed to ensure that data collection and analysis adheres to rigorous, repeatable practices.
- Expert opinion will always be required to sensibly interpret the statistics to determine what is significant ecological change.
The report concludes that in designing any monitoring programme it is important to have clearly specified aims: are you wanting to look at community change (multivariate data) or change in populations of selected taxa (univariate data) in relation to management measures. Well-grounded definitions of what is change (procedural/natural), at what level of significance, and which direction is “good”, are essential before designing the monitoring protocol, and before beginning any analysis. Future maerl monitoring programs need careful experimental design in terms of transect replication and location, and careful selection of sampling methods depending on the metric to be monitored.
Introduction
Background
This project was commissioned under the NatureScot and JNCC ‘Framework Agreement for Marine Survey, Analysis and Interpretation to Support Nature Conservation’. NatureScot is developing analytical techniques to assess change in protected benthic habitats in the Scottish marine environment in the context of management measures introduced through the Scottish Marine Protected Area (MPA) Programme.
The Scottish MPA network includes sites for nature conservation, protection of biodiversity, demonstrating sustainable management and protecting natural heritage. Sites designated to protect marine habitats include Nature Conservation MPAs and Special Areas of Conservation (SACs). Inshore MPAs and SACs with benthic features have been split into two groups to allow for phased implementation of fisheries management measures. A summary guide was produced for measures introduced under the first phase in February 2016 (Marine Scotland, 2016) while phase two measures are still under discussion.
The Scottish Marine Protected Area monitoring strategy (Marine Scotland, 2017) was developed by Marine Scotland, Scottish Natural Heritage (now NatureScot) and the JNCC to create a long-term approach to MPA-related survey and monitoring effort with a goal to report on the status of MPAs every six years with the next report due in 2024.
Maerl beds are both a Priority Marine Feature (PMF) and an OSPAR threatened and declining habitat (Hall-Spencer et al., 2010) and are a protected feature in 11 MPAs in Scotland. New fisheries management measures to protect maerl beds more effectively outside MPAs are also currently under discussion (Marine Scotland, 2019). Maerl beds are highly sensitive to physical disturbance, particularly in the form of abrasion and habitat removal / change, organic enrichment, siltation and changes in local hydrodynamics. In particular, research has shown that heavy, bottom towed gears have the potential for inflicting significant negative impacts on maerl beds.
Maerl is a Bretton name given to a group of calcareous red algae which can, under certain circumstances, form extensive beds of varying topography. The best of these maerl beds are defined by the presence of a complex three dimensional structure, created by accumulations of branched maerl pieces (dead or alive) which can form deposits of considerable depth, though only the top layers which are exposed to light consist of live maerl. Beds vary in appearance and can form continuous carpets, patches of dense maerl on or among other sediments, or have a linear appearance, following the troughs or ridges of sediment waves on the seabed (Peña et al., 2014). The size, shape and structure of maerl pieces themselves can vary from the most common ‘twiglet’ form to a ‘hedgehog’ form or ‘medallions’ and there is a long history in the literature of attempts to classify these forms and relate them to the local hydrodynamic regime and depth (Cabioch, 1966, Peña & Bárbara, 2009). Maerl pieces break down into maerl gravel composed of small dead maerl fragments without much remaining complexity and three dimensional structure. Maerl is considered a non-renewable resource due to its’ extremely slow growth rates and the diverse benthic communities supported by beds.
The epifaunal and infaunal communities associated with maerl beds can also vary considerably. This can be due to natural variation in environmental parameters or due to anthropogenic activity, which means that understanding the drivers of the condition of maerl beds as a protected feature can be difficult. NatureScot have been monitoring maerl beds in various locations using survey techniques including drop down video (DDV) and stills imagery for many years. Samples have been collected for a range of purposes (e.g. biotope assignment, general dive site overview, targeted long-term monitoring) and over a number of years using variable video quality and camera resolutions and in different environmental conditions (underwater visibility, vessel speed etc.).
Project aims and scope
To inform future monitoring and assessment, NatureScot sought input from experts in the field to develop and pilot analytical techniques to describe variation in maerl beds, using a subset of existing video and imagery data collected during MPA monitoring surveys. The long-term aim is to use the methods developed to detect maerl bed recovery and assess the effects of management measures. The focus of this contract is on visual survey techniques, primarily drop-down imagery. Data from two case-study areas were provided for these proof-of-concept analyses, the areas being around the Isle of Arran and off Wester Ross.
The project objectives were to:
- Provide a review of existing peer reviewed literature and agency reports with the aim to establish best practices for monitoring changes in maerl;
- Explore the existing information available as baseline data and investigate best practice for future monitoring
- Test methods that may be applicable on a time series of data in the case-study area;
- Report ‘best practice advice’, considering methods for data collection, imagery analysis and best statistical analysis available for the data type over time.
This report explores the possibility of using at least a semi-quantitative approach to the analysis of data collected from maerl beds. It is likely that the best assessment of the condition of maerl beds will be a combination of such analysis combined with expert opinion.
Methods
Literature review
Methods for surveying and monitoring maerl beds using drop imagery for assessing biotopes and epibiota were evaluated using available peer-reviewed and some grey literature. The review was approached under a series of headings as follows:
- Maerl bed types
- Monitoring methods
- Indicator species – flora and fauna
- Impacts on maerl beds
- Recommendations for field data collection
- Analytical approach
- Application of GIS
Statistical analysis of case study data
Data were provided for a number of survey areas around south Arran and Wester Ross where video transects have been recorded in multiple locations and formats and at irregular time intervals since 2010 over a range of different habitats, some of which have been identified as maerl bed biotopes. Management measures were introduced in both areas at the end of 2014 to reduce impacts from bottom damaging activities such as dredging and trawling and to protect the communities identified.
One of the aims of this report was to determine whether or not certain statistical methods could be employed in the future to assess the degree and direction of change in maerl beds toward ‘richer/better’ or ‘poorer/worse’ condition.
Here we investigate whether or not community composition data derived using the SACFOR abundance scale can be used to detect differences in maerl bed community composition within a site over time, which might be indicative of changes in condition.
Data limitations and suitability of data sets provided
Case study data sets were provided for surveys of maerl biotopes around the island of Arran and off Wester Ross. Data are available for four areas of maerl and maerl gravel inside and outside the South Arran MPA. One area with dense live maerl (Pladda, D6), two with maerl gravel and rare live maerl (Holy Isle D1 and Holy Isle North, D5) inside the MPA, and one with maerl gravel outside the MPA on the Arran east coast east of Corrie (D3).
There is some video available from before and after introduction of management measures in late 2014 for the areas inside the South Arran MPA. One area with relatively dense live maerl (Pladda) is believed not to have received much fishing pressure even before the introduction of management measures because it is difficult to access due to natural barriers. Therefore, it may not be appropriate to describe data from this area as “before” and “after” impact data.
Data for Wester Ross generally do not provide replicate coverage and the videos show that there is considerable variation in maerl bed topography as well as seasonal differences in survey dates. This makes it difficult to assess the condition of the maerl beds in relation to fishing pressures and management of the MPA.
It is important to remember that the data obtained to date have been collected for different purposes using a variety of techniques and equipment. Tow lengths, number of tows, area coverage and locations have varied thus far across the broader areas monitored. The replicate video tows gathered to date, particularly around Arran, have been collected within set ‘monitoring boxes’ which have been designated based on areas of similar environmental parameters e.g. sediments or depths and fishing pressures. Video tows were more or less uniformly distributed throughout the monitoring boxes in any given year but were not set at fixed locations; occasionally tows taken in different years have been in close proximity. The intention of this approach was to be able to extrapolate the video data to represent the benthic communities present throughout each monitoring box. This report considers whether or not sufficient replicate video data tows have been collected from within maerl habitats within these wider monitoring boxes to be able to detect changes in these maerl communities and whether the sampling design is appropriate for the questions being asked of the data.
It is also important to consider that many of the surveys undertaken to date were not designed to gather data with species-specific analyses in mind. On this basis, the statistical methods used here are presented for proof of concept or testing analytical power only; the results should not be taken as direct evidence of real differences in maerl bed condition, community assemblage or species abundance at this stage. For robust statistical conclusions to be drawn regarding the impact of management policies on the recovery or quality of maerl communities and/or selected species over time, further sampling data need to be collected and must continue to follow a robust approach to sampling design.
Introduction to analyses
The SACFOR scale has been used extensively across numerous sampling and survey programmes throughout the UK since its inception more than 25 years ago. The use of SACFOR data is often limited to that of determining biotopes encountered during surveys. The ordinal (or categorical) format with its set scale has historically made SACFOR data incompatible with most common statistical analyses (Strong & Johnson, 2020). Furthermore, the combination of count and percentage cover data using different logarithmic base scales and the use of different density classes for organisms of different body sizes can complicate any analyses of full community datasets. However, recent developments in applying and testing a process for converting SACFOR codes have resulted in a method (see Section 2.2.3) to convert SACFOR data into a more ‘analysable end product’ that can be used to track differences and/or changes in patterns of community composition using multivariate techniques (Strong & Johnson, 2020). Although the broad abundance classes in the SACFOR scale lack precision, it is this breadth (i.e. a single value represents an order of magnitude) that ensures a high level of repeatability between surveyors / analysts, thereby helping to reduce sampling variation and improve consistency of results compared with using raw count data (Strong & Johnson, 2020). Furthermore, the process enables existing SACFOR data to be analysed without the potentially high costs associated with revisiting all historical survey footage to generate community composition data in a different format.
The conversion process allows the full taxon complement of semi-quantitative community data used to assign biotopes and describe biological communities to be analysed together, rather than having to exclude certain taxa based solely on where they are located on the MNCR abundance recording scale i.e. separate analysis of taxa recorded by either density or percentage cover. Converting the data in this way allows whole-community trends or responses to environmental, biological or anthropogenic influences to be observed together. Strong & Johnson (2020) do note that more power may be gained from statistical analyses that stop short of the final stage in the conversion process using fixed numerical values on either the log2 or log10 base scales (thereby retaining a more quantitative element in the data) and then analyse data separately based on either body size or growth form. This perhaps would be a suitable second step to any analytical process in the event that analysis of the entire, fully converted data suggests that community changes may be occurring.
Strong & Johnson (2020) specifically state that “SACFOR data, that includes information on multiple taxa, in well replicated surveys from large areas of marine habitat, provides sufficient power that these data sets should be considered useful for monitoring studies in areas lacking quantitative observations”. The main point in the authors’ conclusions regarding their conversion process for SACFOR data, and alluded to at the outset of this report, is that conversion of SACFOR data is a simplification of the community structure and brings its own inherent limitations. Over-analysis of the data should therefore be avoided. Rather, the data can be used to indicate taxa or taxon-groups worthy of more detailed investigation using more refined sampling or recording methods designed to test particular hypotheses, or the data could be used to further qualify expert opinion on the overall condition / improvement / degradation of any Priority Marine Feature (PMF).
Moore et al. (2019) have evaluated a number of approaches for the analysis of benthic communities from drop imagery. Natural Resources Wales conducts in situ monitoring of rocky reef communities by SCUBA divers in Milford Haven using 0.25 m2 quadrats with a 5x5 grid. Surveyors record each taxon or taxon-group on a scale of 1 to 25 based on the number of grid squares it occupies (Irving et al., 2012). Whilst this method puts each variable on the same, limited scale, it does not account for differences in physical size of organisms and how this contributes to the overall community structure. This can result in some species being over- or under-represented relative to one another e.g. two taxa occurring just once each may be rated at equivalent ‘abundance’ whether they occupy a full grid cell (e.g. massive sponge) or just a tiny fraction of a cell (e.g. zooanthid). Although the technique assesses cell frequency, resolution of taxon frequency or percentage cover is not lost entirely but is reduced, particularly for small taxa which may be clumped but occur in high numbers. In quadrat monitoring elsewhere, Natural England monitors subtidal habitats in situ by recording percentage cover of all taxa or taxon-groups in target habitats within multiple 0.25 m2 quadrats (e.g. Bessell et al. (2016)). Whilst this loses resolution of taxon density, a greater element of dominance is retained in terms of physical space occupied by each taxon / taxon-group.
None of these recording techniques provides the perfect solution to representing entire epibenthic communities where organisms vary greatly in lifeform, size and even mobility, but each method allows an analysis of overall community structure (which is not possible with a mix of count and percent cover raw data) and has the potential to indicate changes occurring over time. Changes suggested by the multivariate analyses with regard to particular taxa or taxon-groups may then require further, more targeted investigation, to assess the significance of the changes. One advantage of using SACFOR data generated from still images is that in order to generate the SACFOR values, species must first be enumerated or assessed for percentage cover; this approach provides quantitative data which may, or may not be useful in univariate analyses (see Section 4.4).
The analyses presented here have been undertaken to determine if existing data (in SACFOR format) are sufficient to demonstrate changes in maerl bed communities over time. Data for transects surveyed around Arran in 2014 were available in SACFOR format. Comparative SACFOR data for 2018 surveys were generated for transects across the same broad scale areas by analysis of the survey imagery. Taxa were initially counted and then converted to the SAFOR scale (see details in Section 2.2.3 below). These count data were also used directly when considering what statistical analysis might be appropriate for taxon-specific (univariate) data. Further details are provided in the following section.
Analytical methods
Data were analysed from video and still images taken during seabed video tows at several survey sites in the Wester Ross MPA and around Arran in 2014 and 2018. Data for Arran from 2014 and 2018 were considered the most comparable in terms of sampling design and spatial coverage and for most locations, offered the best temporal comparison for before / after the introduction of management measures. SACFOR data from 2014 were obtained from the original combined video and stills analyses undertaken by Marine EcoSol (Morris-Webb & Stamp, 2015), while data for 2018 were generated from either multiple still images taken at the time of survey or from multiple screen grabs at the points when the video sled paused over the seabed (details below).
Image analysis
Taxa or taxon-groups identified in each image were either counted or assessed for percentage cover based on the size / growth form categories provided by the MNCR Phase II SACFOR scale (Connor et al., 2004). The approximate area of seabed covered by each image was calculated using the laser scale (100 mm) visible in each image. The approximate abundance per m2 for each taxon per transect was then calculated by summing the abundances from each image and calculating the average percentage cover or density per m2 of seabed for each transect (sample – In this case a ‘sample’ is a single surveyed transect used in statistical analysis. More generally, for Marine Recorder data entry, a single video transect, or tow, would be referred to as an ‘event’, and a ‘sample’ would instead refer specifically to an analysed biotope, subsection or even a single image); the appropriate SACFOR rating was then assigned to each taxon. This resulted in a single column of taxon / taxon-group data per sample based on multiple images (n = 10 to 15). Finally, each transect video was watched in full and any conspicuous taxa not already identified and quantified from the still images were added to the list of taxa and recorded as ‘present’. These mainly included large and conspicuous mobile taxa such as echinoderms, fish and octopi and occasional sedentary taxa such as the colonial bryozoan Omalosecosa ramulosa. Using the still images in this way provided a more accurate way to enumerate individual taxa or taxon-groups for subsequent univariate analyses and power analysis. Using video imagery the field of view can constantly change along a transect and could lead to variable accuracy when estimating densities. While this may have less influence on assigning biotopes to areas surveyed it could have implications for quantitative statistical analyses aiming to assess changes in abundance patterns.
For each image, the seabed sediment composition was quantified based on the percentage cover of the visible area of each sediment type present e.g. sand, shell gravel, stone gravel, dead/live maerl, pebbles, cobbles etc. This was standardised for this pilot study by placing a 5x5 grid across each image to aid accurate estimation of percentage cover wherever practical. The values recorded per image were then averaged to give a one value to represent the whole transect.
A sample of video data for 2014 from each survey site was reviewed to confirm that the SACFOR ratings applied in the original analysis were considered correct and appropriate for comparison with the 2018 data.
Taxonomic nomenclature was standardised across the survey years. This avoided potential repetition of variables across different rows owing either to spelling errors, changes in nomenclature or recording to different taxonomic levels. The World Register of Marine Species (WoRMS) ‘Taxon-match’ function was used to automate this process and reduce the likelihood of errors in the data.
Multivariate analyses
Multivariate community composition data were analysed in PRIMER 7TM.
SACFOR scale ratings were converted to values ranging from 0.3 to 8 depending on organism size class or classification (‘crust/meadow’ or ‘massive/turf’) based on the methodology developed and described by Strong & Johnson (2020). In addition, occasional taxa identified from the video footage and assigned a rating of ‘present’ or ‘P’ were assigned a value of 0.1. The outcome of the conversion process gave count and percentage cover data equal weighting and removed any influence of body size or growth form prior to analysis (Strong & Johnson, 2020).
The workflow of the conversion process is outlined in Figure 1 of Strong & Johnson (2020) and briefly described here.
- Each species observation is attributed to either the SACFOR ‘count’ or ‘percentage cover’ scale and also to body size / growth form respectively.
- Numeric conversion of the counts / cover is based on the lowest possible density or cover value per class using log10 transformation values for density data. Percent cover data were first log10 transformed before being antilog (log2) transformed. The numeric conversion values are not abundance or percent cover values but “are conversion numbers that will align with the converted cover and count values onto an ordinal scale after transformation” (Strong & Johnson, 2020).
- Finally the converted numeric count and cover values are aligned along a ‘new’ ordinal value range from 0.3 to 8. These are the values used in the multivariate analyses.
In practice, using the detail provided in Strong & Johnson (2020), the conversion process is rapid and entails simple additions and subtractions to the SACFOR data generated depending on their abundance class.
Converted data were imported into PRIMER and Bray-Curtis resemblance matrices were generated to compare sample similarities. From the resemblance matrices, multidimensional scaling (MDS) plots were used to visualise the data whilst analysis of similarity (ANOSIM) tests determined whether significant differences existed between pre-defined sample groups i.e. between sites or between time periods within a single site. Where significant differences were identified, SIMPER tests highlighted which taxa or taxon-groups contributed to similarity within or dissimilarities between sample groups.
Physical data (substrate composition and depth) were analysed to see how these might influence the results observed (bubble plots and BEST analysis). In certain cases, BVSTEP analysis was used to test whether any patterns observed in the data were due to specific subsets of taxa or taxon-groups within the wider set of sample variables.
Power analyses
The power of a statistical test to detect change is an important consideration in the design and execution of any experiment or monitoring programme. The collection of too few samples might mean that incorrect conclusions are reached if data cannot demonstrate significant differences when they are known to exist (Type II errors), whilst the collection of too many samples can be a waste of resources. Power analysis therefore is important in predicting future survey and analytical costs while ensuring that data collected are fit for purpose.
Power analysis of multivariate tests (specifically ANOSIM) on community composition data are not commonly employed owing to the problematic and complex nature of such a task. However, multivariate tests are sensitive to differences in community composition, as investigated and discussed with regard to the examples from Arran (see Section 4.1) and other studies cited (Section 5.2), whereby significant statistical differences might be detected despite significant ecological change being considered not to have occurred.
The quantitative data generated for several of the taxa and taxon-groups from the 2018 images from the Holy Isle maerl sites were used to complete univariate power analyses. Power analyses calculate the probability of detecting a change in a variable on the basis that the effect or change really is occurring. Power analyses are often used to determine the optimal number of samples required to detect a significant change in their populations based either on dummy values or a restricted number from a pilot study or similar dataset which comprises values likely to represent the anticipated range and magnitude of those in the planned experiment or monitoring programme.
The univariate power analysis results are based on the outputs from the MONITOR programme which uses simulation procedures to evaluate how each component of a monitoring program influences its power to detect change (Gibbs & Ene, 2010). The programme is devised with population monitoring in mind and allows the user to define the planned sampling design. Outputs indicate the power to detect the specified levels of change over the specified time range based on the known or estimated population mean and standard deviation. Further explanation of the tests is provided alongside the outputs in Section 4.3.
Literature Review
Maerl bed types
Scottish maerl beds are dominated by two, probably three, widely distributed, free living species: Phymatolithon calcareum, Phymatolithon lusitanicum and Lithothamnion glaciale and have been classified to include one main biotope and two sub-biotopes dominated by P. calcareum and a second main biotope with L. glaciale (JNCC, 2015):
- SS.SMp.Mrl.Pcal: Phymatolithon calcareum maerl beds in infralittoral clean gravel or coarse sand.
- SS.SMp.Mrl.Pcal.R: Phymatolithon calcareum maerl beds with red seaweeds in shallow infralittoral clean gravel or coarse sand.
- SS.SMp.Mrl.Pcal.Nmix: Phymatolithon calcareum maerl beds with Neopentadactyla mixta and other echinoderms in deeper infralittoral clean gravel or coarse sand.
- SS.SMp.Mrl.Lgla: Lithothamnion glaciale maerl beds in tide-swept variable salinity infralittoral gravel.
A third main maerl biotope may prove to be present in Scotland with the recent discovery of Lithothamnion corallioides in the Sound of Barra (Bunker et al., 2018):
- SS.SMp.Mrl.Lcor: Lithothamnion corallioides maerl beds on infralittoral muddy gravel
Bunker et al. (2018) also demonstrated mixed populations in the Barra maerl beds including the recently described species Phymatolithon lusitanicum (Peña et al., 2015) which was dominant at some sites. However, it is not always possible to identify the species of maerl present, particularly when surveyors do not have maerl ‘in hand’, and without using molecular identification techniques.
A recently described species, Lithothamnion erinaceum, has been shown to occur at low densities with L. glaciale in Scottish maerl beds (Melbourne et al., 2017). The proportion of maerl species in mixed beds has been shown to change over time (Cabioch, 1969).
It is generally accepted that maerl forming species propagate largely by asexual fragmentation of the branched phase (Pardo et al., 2019) which has a bearing on the genetic diversity of populations. These authors showed for P. calcareum that genetic differentiation was more marked between populations in partially enclosed estuaries and embayments compared with those from the more open coast, a feature which needs to be kept in mind when managing and maintaining connectivity of MPAs for maerl. The genetic diversity of Scottish maerl beds and their connectivity with consequent implications for conservation strategy are the subjects of an on-going study by C. Simon Nutbrown (Heriot-Watt University).
Blake & Maggs (2003) studied growth of two maerl species in Strangford Lough, Northern Ireland obtaining rates of 0.9 mm yr-1 for P. calcareum and only 0.7 mm yr-1 for L. glaciale. As a consequence of the slow growth rate some large maerl thalli may be over 100 years old. This research suggested that there was photo inhibition of growth at water depths of 5 m and shallower (Blake & Maggs (2003). Given these very slow growth rates it is only possible to detect maerl growth in the field over relatively long time periods (>20 years) and other measures of bed condition must be found to detect change in the short to medium term.
Scottish maerl beds show a variety of topographies and it may be necessary to develop different monitoring techniques to take account of these differences, for example Peña et al. (2014) recognised several types, such as maerl beds with ripples or mega-ripples (>20 cm high), with varying densities of algae, a dense scatter of live maerl fragments among cobbles and pebbles or occurring between rocky outcrops. The sediment matrix around the maerl may be either biogenic (comprising broken shell and other biologically derived fragments), or geogenic (composed predominantly of stone pebbles, gravels or coarse sand). The case study data set includes the following topographic types:
- Mega-ripples with maerl devoid of algae, biogenic matrix: deep sites off Melvaig, Wester Ross.
- Mega-ripples with maerl and dense stands of algae, especially Desmarestia spp. – Wester Ross MPA
- Variably dense stands of kelp (specially Saccharina latissima) with maerl beneath on either biogenic or geogenic sediments - Arran (Pladda)
- Maerl between boulders and cobbles – Wester Ross (Inner Sound).
In the literature reviewed, considerable attention has been given to the morphology of the maerl fragments making up the beds with some evidence to suggest that the shape is related to the local hydrodynamic regime (wave exposure, currents) (Cabioch, 1966, Peña & Bárbara, 2009). This might be a useful feature to quantify for Scottish maerl beds using a technique for representing particle shape described by Graham & Midgley (2000). Hall-Spencer & Moore (2000) showed that maerl fragments collected from a bed in the Clyde Sea area in 1891 were much larger compared with samples collected in 1995 after intensive scallop dredging impacts. In a more recent one-off assessment, Kamenos et al. (2003) collected larger fragments from an unimpacted maerl bed in Loch Sween with much smaller thalli gathered from an impacted site on the Isle of Bute. A range of morphotypes may occur in the same maerl bed as shown below for a bed in Guirdal Bay, Rum at about 20 m below CD (Figure 1), however it is possible that these two forms may represent different species in this instance.

Figure 1. Different maerl morphologies from the same maerl bed at Guirdal Bay, northwest Rum at 20 m depth. © L. Baldock
The structural heterogeneity of a maerl bed is crucial to the diversity of micro-niches available for infauna. The loose, open structure of beds, such as those around the Small Isles (Wester Ross MPA) at a depth of about 12 m, supports high biodiversity both within the maerl matrix and supporting diverse epiphytes on it. Deeper beds (20-30 m) off Melvaig (Wester Ross) lack this 3-dimensional structure with the maerl fragments partly buried in the surrounding sediment (Figure 2), lack epiphytic algae and support only crinoids as obvious epifauna.
Joshi et al. (2017a) studying the hydrodynamic forces required to move maerl thalli into suspension found that beds with a high branch density were the most unstable, mobilising en masse once the critical sheer stress was exceeded, the interlocking fragments resisting entrainment up to this point. These authors also demonstrated that live maerl fragments are less dense than dead maerl and therefore settle out differentially from suspension. Kamenos et al. (2003) have shown that un-impacted maerl grounds are more structurally complex than those which have been affected by bottom-towed fishing gear, so some measure of structural complexity could be useful in assessing change in maerl beds.

Figure 2. Structurally complex, shallow maerl bed in the Small Isles (A- left) compared with simple maerl fragments partly buried in sediment on deeper beds off Melvaig (B-right) – Wester Ross. Images taken from video reported by: (A) Moore et al. (2011)
Monitoring methods
The term monitoring in the context of this report refers to a “procedure by which a series of surveys is conducted in a sufficiently rigorous manner for changes in the attributes of a site (or species) to be detected over a period of time” (Hiscock, 1998). It is well recognised that Scottish maerl beds are valuable habitats requiring protection and effective management (Simon-Nutbrown et al., 2020) and hence need for effective monitoring methods, but to date very little monitoring has been undertaken. Recently Hernandez-Kantun et al. (2017) stated that “there is a lack of long-term monitoring of these wonderful habitats [maerl beds] throughout the North Atlantic” and the review of available literature confirms a dearth of any long term monitoring data sets for maerl habitats which are independent of perceived impact studies. Hall-Spencer et al. (2010), reviewing the status of maerl in the OSPAR area, stressed the need for establishing long-term monitoring protocols with programmes designed to target site specific threats to the habitat and to include as a minimum: the extent of the bed, proportion of live maerl and some measure of biodiversity. It has been demonstrated that MPAs with regular monitoring programmes were the most likely to achieve quality objectives (Álvarez-Fernández et al., 2017).
The most extensive monitoring work on northeast Atlantic maerl beds has been carried out around Brittany in France, with early work by Cabioch (1969) followed by a formal monitoring approach developed under the REBENT programme where maerl beds are reviewed every six years (REBENT, 2010). Methods here depend largely on analysis of grab samples (beyond the scope of the current report) together with extent mapping of the beds using remote techniques. A search of available literature failed to find subsequent reports of the results of these monitoring surveys.
Sparse maerl beds of P. calcareum off Swanage on the Dorset coast have been monitored since 1999 (K.J. Collins, Department of Ocean & Earth Science, University of Southampton) using replicate (destructive) samples collected by divers from a 0.5 m2 quadrat (Mitchell & Collins, 2004). This technique is probably not applicable to the denser and often much more patchy maerl beds found around Scottish coasts, and a non-destructive monitoring method needs to be sought to minimise survey damage to these long lived beds. Published results for the Dorset study over the long term programme are not available to assess whether or not there has been significant change in the maerl beds there.
Wilson et al. (2007) undertook an extensive survey of maerl beds off the Antrim coast in Northern Ireland covering a relatively large area using drop video and diver surveys. The drop imagery was collected from sample stations laid out in a grid which allowed identification of seabed communities and maerl biotopes. Algae were sampled using a grab and diver surveys groundtruthed an area of 10 m x 10 m at selected stations with data reported using MNCR survey forms. Repeat surveys have not been conducted at this site so the effectiveness of this approach for monitoring change in a maerl bed cannot be evaluated.
Peña & Bárbara (2010) studied maerl beds off the Galician coast of Spain over a period of just over a year where divers collected replicate samples of algae from 25 cm x 25 cm quadrats for detailed analysis. At the same time the percentage cover of algal species classified into three life-forms and seven functional groups was estimated as well as an assessment made of the proportion of live to dead maerl at the site. The method was successful in identifying that the greatest differences between Galician maerl beds were seen in summer demonstrating that the timing of monitoring surveys is critical. The three areas studied were different in terms of their floral composition. Subsequent surveys to assess long term change in this area have not been conducted.
Diver surveys undertaken on Scottish maerl beds have used targeted MNCR Phase II techniques described by Hiscock (1996) combined with quantification of maerl densities using small quadrats (Bunker et al., 2018, Mercer et al., 2018). Maerl beds in the Fal in Cornwall have been extensively surveyed and reviewed over the years using divers and remote imagery, but to date no routine monitoring programme has been developed (Perrins et al., 1995, Howson et al., 2004, Allen et al., 2014). The most recent survey (Allen et al., 2014) used a combination of drop imagery and diver surveys and obtained broadly similar results to previous studies. One issue that was noted was the inconsistent classification of biotopes: Howson et al. (2004) recorded biotopes with a significant proportion of dead maerl gravel as maerl biotopes whereas Allen et al. (2014) more appropriately classified these as gravel (coarse sediment) biotopes. Aside from this discrepancy of biotope classification, qualitative assessment of differences between surveys conclude that there had not been any substantial change in the maerl beds.
Given the lack of published evidence from repeat surveys for any monitoring programme on a maerl bed, it is not possible to draw conclusions as to which, if any of the above approaches provides the best measure of change. Carefully tailored monitoring methods will need to be developed depending on the purpose of the monitoring programme (assessment of impact, recovery etc.), the nature of the maerl bed concerned together with constraints on available survey resources.
Indicator species - taxa that may denote change in maerl bed condition
During the development of the Marine Habitat Classification for Britain and Ireland by the JNCC (Connor et al., 2004) biological comparative tables were provided allowing a comparison of lists of key taxa occurring in a user-defined set of biotopes. For the purposes of the present study the taxa occurring in the core records for six maerl biotopes were extracted to a spreadsheet (Annex 1) and reviewed to assess whether any of these might represent useful indicator species for maerl bed monitoring. The taxa listed are all very widely distributed and not dependant on the presence of maerl in the habitat.
It may be possible to make a list of taxa together with selected functional traits which would be useful to monitor in maerl beds. The research of Peña & Bárbara (2010) suggested common algal species which occur widely both in maerl beds and other habitats should be monitored (e.g. Cryptopleura ramosa, Dictyota dichotoma, Plocamium spp, Scinaia sp), as well as species more or less confined to maerl beds and which are sensitive to disturbance (e.g. Lychaete rhodolithicola, Lychaete pygmaea, Cruoria cruoriaeformis, Gelidiella calcicola, Gelidium maggsiae).
However, species level identification of algae requires a high level of expertise within the survey team and it is essential that there is consistency across surveys. Trials reported by Moore et al. (2015) showed that it is particularly difficult to standardise an approach to field recording even with expert participants. Allocation of taxa to functional groups requires that different taxa are distinguished but it is not necessary then to identify each one to species level. Provided that a careful record is kept for each survey (lists, photographs) of which taxa are allocated to which functional group it should be possible to use this method consistently for algae across surveys.
Peña & Bárbara (2010) classified maerl bed algal flora into three life form groups following the approach of Garbary (1976): annual, perennial, and semi-perennial (where only part of the frond persists either as a reduced erect frond, basal creeping stolons or a crust). The flora was then further sub-divided into functional groups (Table 1): crustose, foliose, filamentous, corticated foliose, terete, articulated calcareous and leathery. Steneck & Dethier (1994) first proposed such a classification to describe algal communities from a wide range of marine environments, this classification has been refined and additional traits added by Mauffrey et al. (2020) for macroalgae which could then be used in further analysis.
Table 1. Functional groups for algae (after Steneck & Dethier, 1994).
| Functional Group | Example taxa |
|---|---|
| Crustose | This will include some life stages of common maerl bed taxa: e.g. Cruoria rosea tetrasporophyte of Halarachnion as well as purely crustose forms e.g. Peyssonnelia spp. |
| Filamentous | Cladophora, Ceramium spp, Gelidium spp |
| Foliose | Ulva spp, Porphyra spp |
| Corticated Foliose | Dictyota, Phyllophora, Kallymenia |
| Terete | Chondrus, Gracilaria spp. |
| Articulated calcareous | Corallina spp, Jania spp. |
| Leathery macrophytes | Kelps, Dilsea |
The classifications of Steneck & Dethier (1994) and Peña & Bárbara (2010) have been applied to the results of two Scottish maerl bed surveys which included detailed information on algal taxa and undertaken by broadly the same team of experts: in the Sound of Barra (Bunker et al., 2018) and in the South Arran MPA (Mercer et al., 2018) (Annex 3). The two maerl beds supported relatively high algal diversity with 86 and 53 taxa respectively but with rather different communities. A functional traits analysis of the algal and invertebrate community of maerl beds could prove useful in assessing impacts of changes in management (Mason et al., 2005, Mouillot et al., 2013).
The algal flora associated with maerl beds can be very varied, with 30% of the total seaweed diversity of the northeast Atlantic represented in maerl habitats (Peña et al., 2014) hence providing a potentially effective monitoring tool representing an important component of the community. The flora tends to be dominated by Rhodophyta (Peña & Bárbara, 2008, Bunker et al., 2018), of which many species typically have heteromorphic life histories with a cryptic, crustose phase which occurs on maerl (Maggs, 1984). Some species of algae with ramifying rhizoids and tendrils play an important role in stabilising maerl beds by binding together adjacent maerl fragments (Bárbara et al., 2004), examples include Gelidiella calcicola (Maggs & Guiry, 1987), Spermothamnion spp. and Vertebrata byssoides (Wilson et al., 2007).
The suite of taxa comprising marine benthic communities of has long been used to assess marine benthic systems for spatial variation and to evaluate possible anthropogenic impacts on these communities such as from the use of demersal fishing gear (Bremner et al., 2003a). Bremner et al. (2003b) considered the advantages of using the diversity or richness of biological traits to evaluate a community to include:
- difficulties of geographical variation inherent in taxonomic community composition are avoided, ecosystem structure then becomes independent of geographical location;
- the distribution of traits within a community varies on a smaller spatial scale than does taxonomic composition, thus allowing for the possibility of better discrimination between sites;
- there are strong links between biological traits and ecosystem processes, thus changes in the proportion of selected traits represented in the community over time can serve as a reflection of changes in ecological function which may have been mediated by changes in environmental drivers including differences in management measures.
Characteristics of likely suitable taxa
The following taxon attributes are likely to be useful for assessing change in maerl beds:
- Relatively long lived, but not excessively so;
- Somewhat limited dispersal capabilities;
- Resilience to disturbance – this requires a fine balance. For example populations of the burrowing bivalve Dosinia are likely to take decades to recover (too long), the burrowing anemone Cerianthus lloydii up to two years (useful), while the polychaete worm Chaetopterus has tremendous regenerative powers and a very tough tube so may not appear to be impacted by disturbance at all (Hall-Spencer et al., 2003);
- Binding growth forms among the algae (e.g. Gelidium spp, Cladophoraceae);
- Some taxa are conspicuous and therefore easily recorded from drop imagery such as species of starfish (e.g. Marthasterias glacialis, Porania pulvillus, Luidia spp, Asterias rubens).
A list of taxa commonly occurring in maerl beds is provided in Annex 1, together with information on longevity, burial depth, tube length of the organism and other characters important to the structure of maerl beds. This list of taxa is derived from the Marine Habitat Classification for Britain and Ireland using the biological comparative tables which allow a comparison of lists of key species occurring in a user-defined set of biotopes. For the purposes of the present study the species occurring in the core records for six maerl biotopes listed in the classification were extracted. Biological traits for 40 of these taxa are available from the Biological Traits Information Catalogue (BIOTIC) developed by MarLIN (2006) and are provided in Annex 2, traits likely to be relevant in terms of assessing change in management practice need to be selected from this list for further analysis while details for other taxa can be added with further research. Both ordinal and numeric values can be accommodated in the traits list and R scripts are available for further analysis.
When selecting characterising taxa for monitoring, it must be kept in mind that there need to be sufficient data from a practical number of replicate samples to detect significant statistical differences. For example, very rare or cryptic taxa are unlikely to be useful for detecting significant community change from drop down imagery. Data sufficiency in maerl bed monitoring is discussed further in relation to power analysis (Section 4.4). This section goes on to list some of the potentially relevant indicator taxa in terms of their statistical suitability for ongoing monitoring work.
Impacts on maerl beds
It is important to identify the likely impacts to the maerl bed concerned so that a suitably targeted monitoring programme can be set up – details of the precise monitoring strategy will be site specific and will depend on the impact being monitored e.g. fishing impacts (current / historic), aquaculture development and general drivers of change e.g. climate change. The survey plan must provide information for comparison with subsequent surveys without confounding differences due to survey and analytical methodology (procedural differences) with natural or other causes of change in the maerl bed community. Seasonal timing of the survey is also important, particularly so where algal communities are a critical component of the complex being monitored.
Impacts to be considered will include:
- Anthropogenic impacts – bottom towed fishing (Hall-Spencer & Moore, 2000, Hauton et al., 2003, Kamenos et al., 2003, Barberá et al., 2017), aquaculture (Hall-Spencer et al., 2006). Un-impacted maerl grounds are more structurally complex than those which have been affected by dredging for example (Kamenos et al., 2003, Bernard et al., 2019) so a measure of relevant substrate complexity would be useful. Three dimensional imaging techniques have been trialled on an undisturbed maerl bed in Loch Sween and in this instance gave a qualitative impression of the structure of the maerl bed. There is scope for improving the resolution of the source imagery and using fractal analysis to quantify the structural complexity of the community (Jon Chamberlain pers. com.). Evidence of damage to epibenthos such as loss of arms in echinoderms and the relationship with fishing intensity has been explored (Kaiser, 1996, Rogers et al., 2001).
- Siltation and smothering by fine sediment Wilson et al. (2004) showed that burial by fine sediments was lethal to maerl while a Dorset study of Phymatolithon calcareum demonstrated differences in the rate of calcium deposition between spring/neap tide cycles. The differences were related to changes in light attenuation due to higher levels of suspended solids in the water column during spring tides (K.J. Collins, Department of Ocean & Earth Science, University of Southampton, pers. comm.).
- Climate change mediated by either or both increased temperature and increased ocean acidity has been identified as a potential serious threat to maerl communities (Brodie et al., 2014). Increased acidity will lead to increasingly brittle maerl thalli and therefore a greater likelihood of the complex three-dimensional structure of a prime maerl bed breaking down. Changes in temperature regime may result in a change in the balance of maerl species within a given bed. Simon-Nutbrown et al. (2020) have modelled the distribution of maerl in Scottish waters in the context of future climate change.
- Increase in the frequency of extreme weather events: Joshi et al. (2017b) have shown that storm events are critical in governing maerl distribution in Galway Bay, Ireland.
Recommendations for field data collection
A number of publications provide recommendations regarding the use of underwater drop imagery for monitoring subtidal benthic habitats with useful summaries provided by Coggan et al. (2007) and Hitchin et al. (2015) with best-practice guidance by Noble-James et al. (2017). Any monitoring plan needs to be carefully considered in order to provide sufficient replication for robust statistical analysis and adequate coverage for GIS deliverables.
These publications indicate that the main requirements for obtaining high quality imagery are:
- High quality drop video, well-lit and with some means of scaling the image.
- High quality well-lit still, relatively close-up images of the seabed collected at regular intervals – these are particularly useful in assessing the sediment properties and proportions of live/dead maerl (e.g. EMFF 2018 Arran survey). Still frames extracted from drop-video where the unit rests on the seabed may also be suitable (e.g. 2018 Melvaig herring survey).
- Relatively slow speed over ground, ideally <0.5 knots.
- Relatively short deployments are better, each set of drop video and still imagery providing a “sample” for later analysis. A survey area of at least 25 m2 should be the aim for each video transect (sample), the minimum area required to define a biotope with confidence. Most transects around Arran in 2014 and 2018 were ~100 m2 (equivalent to 7-10 minutes of video on seabed under the prevailing conditions).
- Accurate geo-referencing of imagery is crucial and requires a GPS track log with positions recorded at 5-10 s time intervals and ideally also a continuously updating displayed on the video stream together with depth and with these data recorded on the saved video file. It is possible to geo-reference video screen grabs retrospectively but this is less accurate and is also time consuming. Still images can be automatically geo-referenced to the nearest GPS track log position. For this to be as accurate as possible it is essential that cameras are synchronised with GPS time, or at least the time difference is precisely known and recorded, even if this is only a few seconds. Deriving points given just the start and end positions of the survey track is very much a last resort.
- Side-scan sonar - relatively inexpensive units are now available and can be run concurrently with the acquisition of drop imagery. These data from selected manufactures’ units can be readily imported to a GIS using proprietary software: for example ReefMaster which has a multi-channel sonar viewer as well as a method for compiling sonar mosaics. Hawes et al. (2014) working in St Austell Bay, Cornwall, showed that bed forms visualised this way could be combined with maerl distribution derived from drop imagery and used to elucidate changes in bed form between monitoring visits.
- Other data to collect might include: recent severe weather events, known incursions by fishing vessels, pollution incidents etc.
Analysis of drop imagery
There have long been detailed recommendations available for standardising analysis of drop imagery (Turner et al., 2016) and Marine EcoSol has produced a standard operating procedure which adheres to these guidelines (Annex 4). It is crucial that image analysis provides consistent data sets between surveys in order that results are comparable across surveys. Various resources must be provided with each contract to ensure consistency between surveys; however it is important not to view these as static assets but evolving as various maerl beds are surveyed and more experience is gained. Items to include:
- Image analysis guidance document (Turner et al., 2016).
- A Standard Operating Procedure (SoP) see for example that provided in Annex 4.
- Illustrative resources such as representative sediment scales, % cover scales, list of taxa indicating the SACFOR/enumeration method used for each one, proforma spreadsheet listing all features to be recorded for every survey etc. Moore et al. (2019) have reviewed the relative merits of different data extraction methods from drop imagery in the analysis of benthic communities.
- Reference collection of taxon images recorded during monitoring of Scottish maerl beds.
- If functional traits of taxa are to be used these can be defined a posteriori with full details provided, including the definition of terms, so that the same criteria are used throughout monitoring.
- Illustrated categories of maerl morphology (if to be used) (Cabioch, 1966).
Analytical methods to determine changes in maerl communities
Prior to the analysis of the 2014 and 2018 south Arran maerl community data, the potential for the use of SACFOR data to detect differences in epibenthic communities was tested on the 2014 data for the whole of south Arran using the data from Morris-Webb & Stamp (2015). This was solely to determine whether or not the analytical method was capable of differentiating between broad scale areas supporting different biological communities; if this was not possible then the technique would unlikely be sensitive enough to detect more subtle shifts within biotopes and a different investigative approach would be required. The results showed that analysis of SACFOR data could detect differences in the communities and are presented in brief in Annex 5. The results were not discussed in detail as they were not directly related to the main aims of this report.
In the following multivariate analyses, most mobile taxa and taxon-groups were removed from the variable lists prior to analysis. Many of these taxa might either actively avoid the survey equipment as it passed overhead (fish, larger crustaceans, cephalopods) or might not be recorded efficiently by the camera’s limited field of view (larger, widely dispersed echinoderms); their inclusion would only serve to increase variation in the data.
Analysis of temporal differences within maerl bed communities around Arran 2014 vs 2018
Pladda maerl beds, south Arran
Of all the sampling data available for south Arran, the transects around the island of Pladda held the best examples of maerl beds with a high percentage of live maerl so transect samples from in this area were selected for the first analysis of converted SACFOR data, as per Strong & Johnson (2020) (see Section 2.2.3). The analyses aimed to determine whether multivariate analysis of converted SACFOR community data could show differences in maerl communities from the same site between two time periods, September 2014 and September 2018.
Around Pladda, the live maerl biotope occurs to the east and west of a shallow rocky reef running between the north of the island and the south coast of Arran (Figure 3). Analysis of seabed images taken in 2018 (see Section 2.2.3) generated further SACFOR data specifically for the sites with maerl biotopes (O'Dell et al., in prep.). Existing sample data identified as maerl biotopes from 2014 were selected for comparison (Morris-Webb & Stamp, 2015).
A one-way ANOSIM test indicated significant differences in the converted SACFOR community data between the two sampling years (ANOSIM R = 0.665, P<0.001). Separation of the data by sampling date was evident in an MDS plot (Figure 4).
SIMPER analysis of the data (Annex 6) demonstrated that the taxa or taxon-groups responsible for the top 70% of sample dissimilarity between the years were recorded within the survey area in both years (with two exceptions: Didemnidae in 2014 and Spirorbidae in 2018). Therefore, it was mainly differences in the recorded abundances of the majority of the variables that generated most of the between-year dissimilarity, as opposed to presence vs. absence of any prominent taxa between years. This result suggests that analysis of maerl community data using converted SACFOR scores has sufficient sensitivity to detect temporal changes in community composition despite few differences in the overall species inventory.
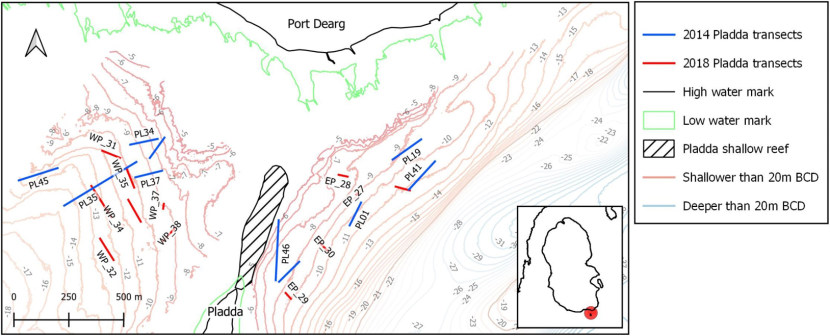
Figure 3. Locations of transects with maerl biotopes around Pladda south Arran sampled in 2014 and 2018. (Contains public sector information, licensed under the Open Government Licence, from Marine Scotland and OS data
Click for a full description
© Crown copyright [and database rights] 2021. Not to be used for navigation). Around Pladda, the live maerl biotope occurs to the east and west of a shallow rocky reef running between the north of the island and the south coast of Arran.
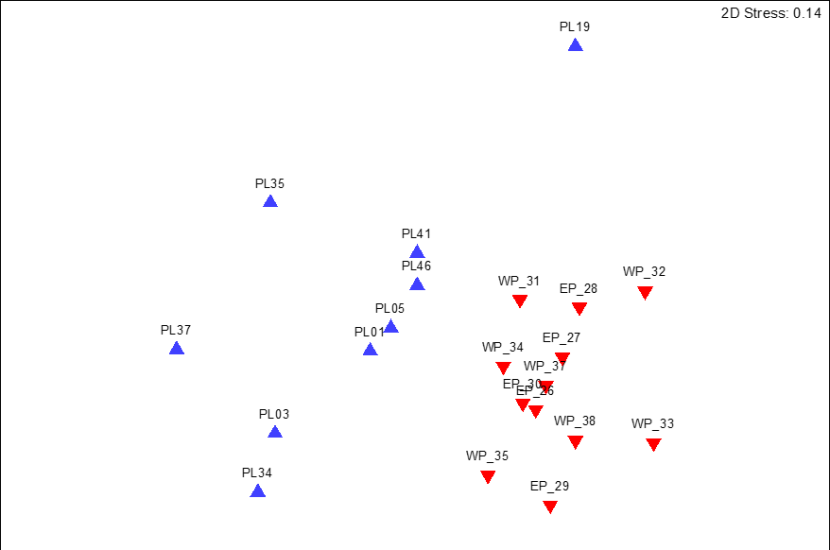
Figure 4. MDS plot of transect community composition data collected from maerl biotopes around Pladda off south Arran in September 2014 (▲) and 2018 (▼)(n = 21).
Click for a full description
The points for 2014 cluster clearly apart from the points of 2018.
Although the survey area around Pladda contained the best examples of live maerl biotopes around south Arran on which to test the application of multivariate analysis on SACFOR community data, the area may not provide the best example to demonstrate potential effects from changes in management measures. The shallow waters together with the associated rocky reef and tidal conditions around Pladda coupled with the visually good condition of the maerl bed in that area suggest it is unlikely that towed fishing gears were used extensively here prior to the introduction of management measures in 2014. Consequently converted SACFOR data from a second maerl bed off south Arran (Holy Isle) were also analysed to determine if similar differences between surveys undertaken in 2014 and 2018 also existed at this site.
Holy Isle maerl beds, south Arran
Across the shallow plateau off the south coast of Holy Isle are areas of dense maerl gravel albeit only with rare live maerl. This area was fished regularly prior to the introduction of management measures in 2014 and therefore has the potential to demonstrate recovery. Video data from 2014 and 2018 offer some means of community comparison before and after the implementation of management measures (Figure 5).
Ten transects from 2014 that contained high percentages of dead maerl gravel were identified as of interest. Eight transects from the same general survey area were identified in 2018, also containing maerl gravel data. Some of these were in close proximity to those from 2014 whilst others were more distant but were selected on the basis of their maerl content or designation as a maerl biotope. In total 106 images were analysed to generate the 2018 data for the Holy Isle sites with 10-15 images contributing to the sample data per replicate transect.
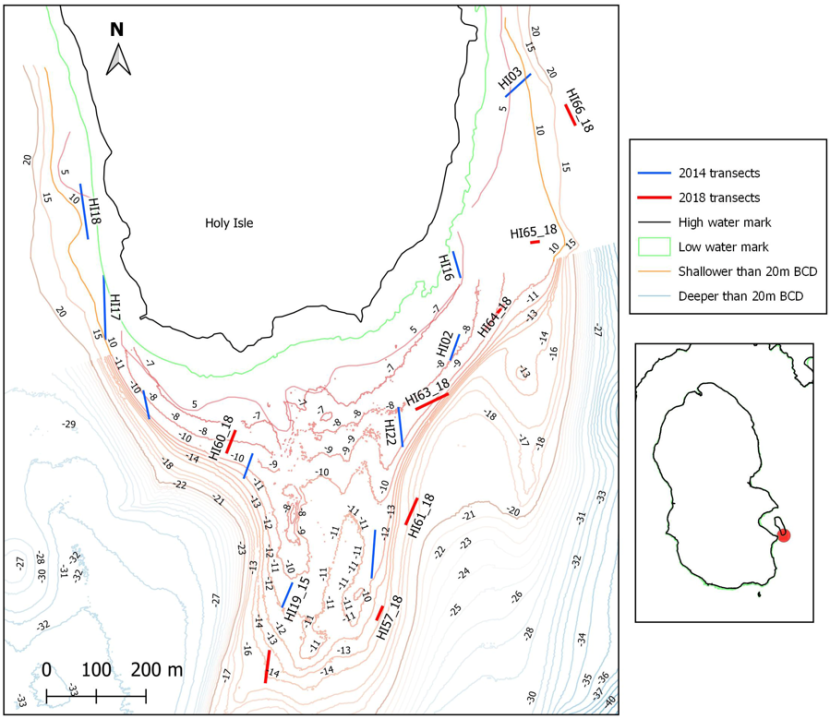
Figure 5. Locations of transect samples classified as maerl biotopes and therefore selected for analysis of maerl community changes off Holy Isle, south Arran in 2014 and 2018.
Click for a full description
(Contains public sector information, licensed under the Open Government Licence, from Marine Scotland and OS data © Crown copyright [and database rights] 2021. Not to be used for navigation) A map displaying the south end of Holy Isle, off Arran, with depth contours and survey stations transects.
A one-way ANOSIM test indicated there were significant differences in the converted SACFOR community data from the Holy Isle maerl beds between the two sampling years (ANOSIM R=0.937, P<0.001). Strong separation of the 2014 vs 2018 data based on sampling date was evident in the MDS plot of the sample data (Figure 6).
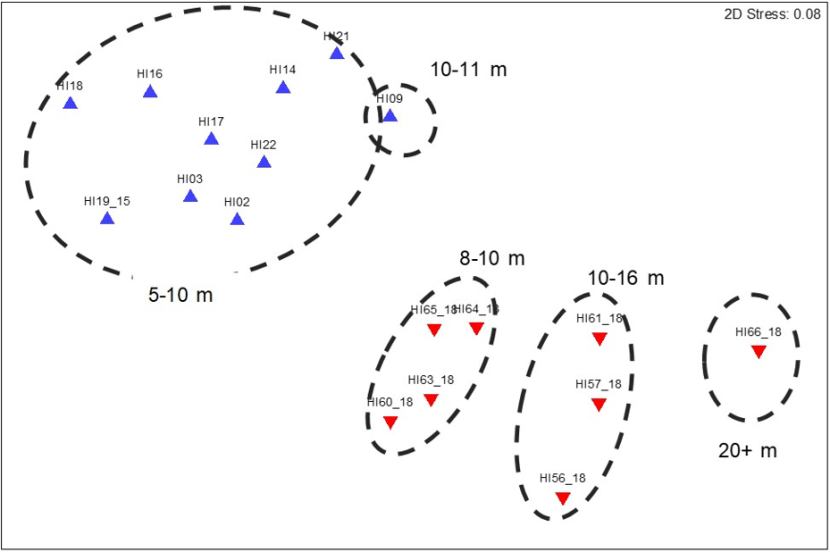
Figure 6. MDS plot of transect community composition data collected from maerl biotopes around Holy Isle off south Arran in September 2014 (▲) and 2018 (▼)(n = 18). Approximations of average transect depths are indicated by dashed lines.
Click for a full description
There are four discreet clusters, one for all 2014 stations, and three for 2018.
A SIMPER analysis of the data (Annex 7) demonstrated that many, but not all of the taxa or taxon-groups responsible for the top 70% of sample dissimilarity between years were recorded in both years. As with the Pladda data, for most variables it was the difference in the recorded abundances that generated most of the between-year dissimilarity. Some variables were conspicuous by their absence in one year or the other e.g. Toxisarcon sp., bivalve siphons, Neopentadactyla mixta were absent from the 2014 data, whilst ‘Rhodophyta – Heterosiphonia japonica’ was absent from the 2018 sample data. Possible reasons for these observed differences are discussed later.
Figure 5 shows the locations of the transect samples taken over maerl biotopes around Holy Isle in 2014 and 2018 in relation to depth. The influence of depth over the communities recorded seems likely to be important: in 2014 all transects identified as maerl biotope were located in shallow water with transects all <11 m BCD. Whilst this might explain some of the interannual dissimilarities, data from the shallower sites in 2018 were distinctly separate on the MDS plot from those recorded in 2014 (Figure 6). In this instance, the average depth of each transect was approximated from the depths provided at the start and end of each transect which varied by no more than 4m, most being less. Any final analysis of the wider data sets available might be able to gain a better average depth estimate from complete tow depth profiles over the maerl habitats. Depth has been included here as a possible factor affecting sample composition as suggested by the 2018 data and is worthy of further investigation should a data set exist with a greater number of replicate maerl transects across the depth ranges encountered in the monitoring boxes.
Potential influence of abiotic variables
The ‘biota and / or environment matching’ (BEST) analysis in PRIMER selects the biotic and abiotic variables from those available that ‘best’ explain the community patterns observed. Substrate composition and average transect depth were used to determine whether or not these might explain the patterns observed in the data. The variables with the highest correlation with the observed data were subtidal sand and subtidal fine sand (correlation coefficient = 0.832) (full results in Annex 8). These two environmental variables (subtidal sand and subtidal fine sand) were not recorded in 2014, due to the lower quality of the recorded footage and speed over ground of the video recordings (Morris-Webb & Stamp, 2015) but were available in 2018. In 2014 coarser sediments were recorded along the transects, occasional notes suggest some finer sediments were occasionally present but they were unquantifiable from the available footage. Perhaps surprisingly, transect depth below chart datum did not factor as a highly contributing variable despite there appearing to be a relationship between depth and percentage sand / fine sand in 2018.
The bubble plots below (Figure 7 and Figure 8) overlay the MDS ordination from Figure 6 with the percentage of sand/fine sand (where data were available) and depth to illustrate how these varied during each survey event. The plots show that in 2018 the sand fraction recorded from the images increased with depth and indicate the potential influence of depth and substrate on the community data at each site but accurate determination of the relationships is hindered by the available data, particularly with regard to sediment fractions. The examples provided here illustrate the analyses that can be undertaken using both abiotic and community data but results should not be taken as conclusive at this stage.

Figure 7. Bubble plot MDS overlays of percentage composition of sand (blue) and fine sand (red)of each survey transect over maerl habitat off Holy Isle in 2014 (left hand side clusters, sand not reported) and 2018 (right hand side clusters).
Click for a full description
Bubble size indicates the percentage composition of the sediment fractions between the ranges denoted on the figure.
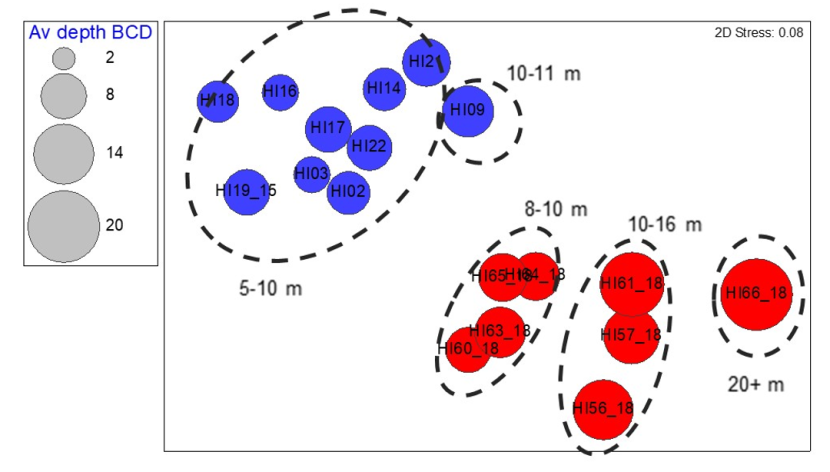
Figure 8. Bubble plot MDS overlay of estimated depth below chart datum of survey transects over maerl habitat off Holy Isle in 2014 (blue) and 2018 (red).
Click for a full description
Bubble size indicates depth in meters BCD with broader depth classes denoted by dashed lines.
Main taxa responsible for community patterns
BVSTEP analysis is a step-wise process capable of determining whether subsets of species variables might best explain patterns in the community. In summary, a subset of 17 taxa/taxon-groups was selected which produced a resemblance matrix with a correlation coefficient of 0.939 with that which included all taxa or taxon-groups. The taxa or taxon-groups selected were those most frequently recorded in sample images in either one or both the survey years and included:
- Alcyonium diaphanum
- Cerianthus lloydii
- Porania pulvillus
- Clatharina sp.
- Gibbula sp.
- Toxisarcon sp.
- Chorda filum
- Bryozoa indet. crust
- Corallinaceae
- Desmarestia sp.
- Fucus sp.
- Melobesioideae (dead)
- Melobesioideae (live)
- Saccharina latissima
- Bryozoa (turf)
- Rhodophyta - filamentous
- Scrupocellaria sp.
A number of other subsets of taxa with correlation coefficients of 0.936 were also identified by the analysis. Whilst most of the taxa in those groups were the same as those listed above, they also contained Antedonidae, Ophiuroidea, Amphilectus fucorum, Laminariaceae, Membranipora membranacea, Abietinaria abietina, Carpomitra costata, Cryptopleura ramosa, Hydrozoa (fine), Obelia sp., Phaeophyceae (filamentous), Rhodophyta – branched, Rhodophyta (Heterosiphonia japonica), Sertulariidae, Ciona intestinalis, Neopentadactyla mixta.
The lists above go some way to defining the core taxa and taxon-groups responsible for the underlying patterns of similarity and dissimilarity between samples (see Figure 6). The BVSTEP analysis may therefore be useful for selecting particular taxa or taxon-groups for specific monitoring purposes if they are understood to form some of the main drivers behind community patterns in particular maerl beds. Some of these taxa and taxon-groups are examined further in Section 4.4 with regard to power analysis and the potential to use existing data for univariate measures of change. The full results of the BVSTEP analysis are provided in Annex 9.
Analysis of temporal differences within maerl bed communities around Wester Ross
The transect surveys undertaken around Arran in 2014 and 2018 provided a high number of replicate tows throughout targeted survey areas (monitoring boxes). Available transect data for maerl habitats around Wester Ross in both the Summer Isles and the reference locations north and south of Loch Gairloch were fewer in number and varied significantly in their quantity and spatial coverage between surveys, with consequent implications for quantitative analysis.
Two potential reference sites north and south of Loch Gairloch (outside the MPA) were evaluated for on-going monitoring and comparison with sites in the Wester Ross MPA (Figure 9) which are protected from bottom-damaging activities. However, review of video data for the sites surveyed in these two study areas showed different maerl bed habitats recorded during each survey event (e.g. more or less exposed, deep vs shallow sites with very different substrate and biota), each surveyed only once. This precludes any useful temporal analysis, even for ‘proof of concept’ as any differences identified would more than likely be due to spatial and physical differences between the areas. Further sampling at these sites is required and is discussed later.
Transect locations over maerl habitat around the Summer Isles within the Wester Ross MPA were at sites separated by several miles and covered a range of depths, tidal conditions, and exposures where exposure values are shown for GIS layers available from the Scottish Association of Marine Science derived by methods detailed by Burrows et al. (2008) (Figure 9). Review of the video footage from these sites showed variation in the maerl beds surveyed in terms of the substrate type, depths and visible communities. Consequently, comparisons between samples to assess temporal community changes which might be related to changes in management measures would be confounded by high environmental variation across such a physically heterogeneous area. Furthermore, most transect data from 2010 were obtained using a different video technique with an oblique view and constant changes in the field of view with no laser scale, making robust semi-quantitative assessment impossible.
In the Summer Isles three transects MB81, WR41 and WR104 to the east of Glas-leac Mòr were surveyed in close proximity to one another in 2010, 2014 and 2018 respectively (Figure 10) and the data were explored in case it might be possible to make a temporal comparison of what could be the same maerl bed. On review of these transects MB81 was discounted on the basis that it began on rocky reef before dropping sharply over a ledge onto an area of dense maerl with sparse epibenthic taxa, in stark contrast to the more diverse shallower maerl bed(s) of transects WR41 and WR104.
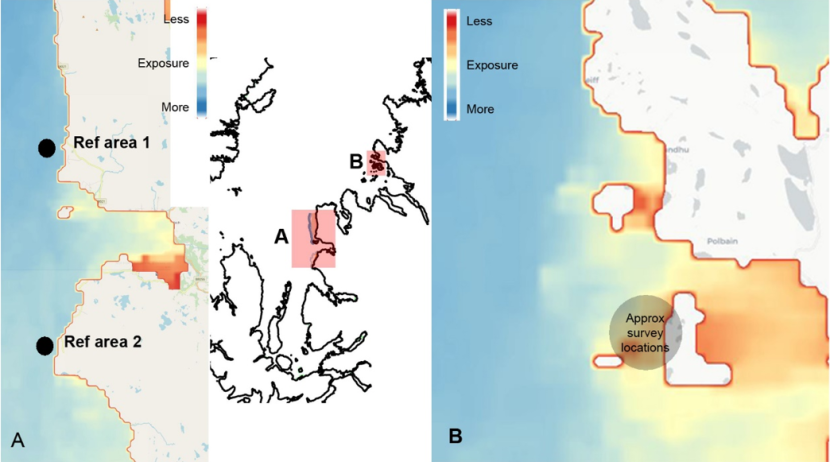
Figure 9. Comparison of the degree of exposure of sites at possible reference locations north and south of Loch Gairloch (A) and within the Wester Ross MPA (B). Exposure scale is derived from Burrows et al. (2008).
Click for a full description
(Contains information from the Scottish Association of Marine Science and OS data © Crown copyright [and database rights] 2021. Not to be used for navigation) Wave exposure all
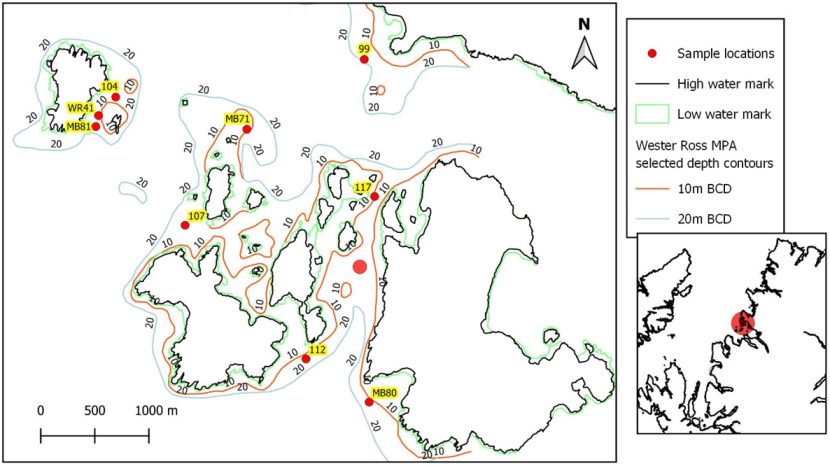
Figure 10. Complexity of the Summer Isles study area (Wester Ross MPA) in terms of depth and wave exposure (varying degrees of shelter afforded by land). (Contains OS data © Crown copyright [and database right] 2021. Not to be used for navigation).
Click for a full description
Wester Ross MPA complex
Location data confirmed that transects WR41 and WR104 ran almost end to end on the east side of the island of Glas-leac Mòr, collected some four years apart over more or less the same area of maerl habitat. However, with only a single transect available for comparison, statistical analysis using the same methods as for the Arran sites described above was impossible. The use of individual still images from each video transect as replicates was considered in an attempt to detect potential temporal change. However, such an approach calls into question whether the images recorded, each of which encompasses only a very small area of seabed (~0.15 m2), would provide a representative view of the wider habitat. In investigating the Arran data, the process of averaging data from all images along a transect to provide a single column of community data, ensured the species richness in that localised area (or community) was better represented in a single sample column. Using just single images where each image does not adequately capture a site’s species richness simply adds to the between-sample variation and could influence the end results if too much variation is related to the sampling methodology. A shade plot based on analysis of 51 images from both transects illustrates this latter point (Figure 11). The plot shows that the number of taxa per sample can vary by up to two-fold within either transect. Other relatively common taxa such as the tube-dwelling worm Chaetopterus sp. or the brown alga Desmarestia sp. are absent from several individual images (each covering a very small area of seabed) when they are clearly present throughout the broader habitat, evidenced by their occurrence in many other images and throughout the accompanying video footage.
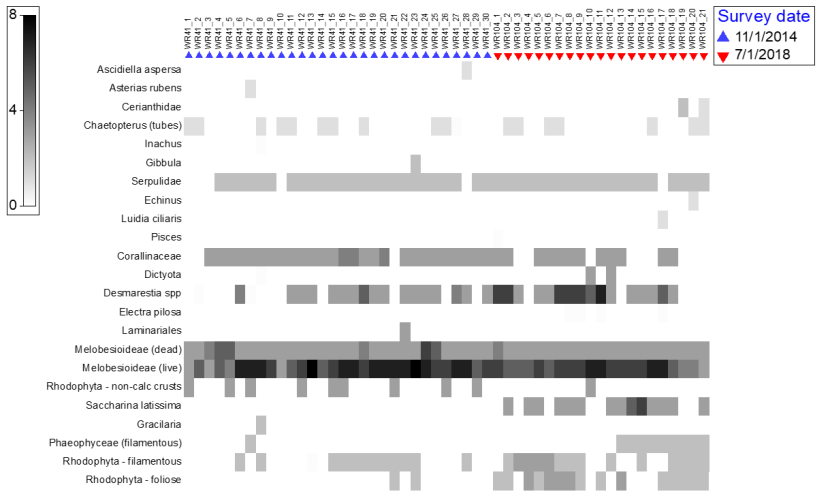
Figure 11. Shade plot showing converted SACFOR scores of taxa / taxon-groups recorded from individual still images (top horizontal axis) taken along transects WR41 (Nov 2014 ▲) and WR104 (Jul 2018 ▼) Summer Isles, Wester Ross MPA.
Click for a full description
(Grey shading scale on the left shows abundance scale conversion as per Strong & Johnson (2020); ‘P’ represented as 0.1).
Furthermore, images taken at more or less regular intervals along a single straight line transect cannot be considered to be randomly distributed throughout a site and are pseudo-replicates rather than true replicates. These are more likely to exhibit spatial autocorrelation with one another, meaning they are naturally more likely to show greater within-group similarities rather than between different time periods for example (Underwood, 1997) (see Section 5.1). The use of individual images as replicates from a single transect (that may or may not be representative of the wider maerl bed) risks conclusions being drawn regarding community changes that might not be occurring over the wider area i.e. the level of sample replication may not be appropriate to the spatial scales of the questions being asked of the data.
For these reasons, no statistical analyses of data from Wester Ross are presented here. However, qualitative observations of the images from the two transects do highlight some differences between the survey years and shade plots (such as the example above) could prove useful for illustrating the conclusions of any future condition assessments. Firstly algae such as Dictyota, Desmarestia, foliose and filamentous lifeforms and Saccharina latissima were noticeably more abundant in 2018 compared with 2014. This is clearly demonstrated in the shade plot above (Figure 11) and is very likely linked to the seasonal timing of the surveys – winter November 2014 and summer July 2018. Secondly, very obvious from the video playback, was the difference in the overall bed structure between the two survey events. In 2014 the maerl bed was very irregular, with a mosaic of short bands of live maerl arranged in haphazard directions across the seabed, whereas in 2018 the live maerl occurred in regular, parallel mega-ripples throughout the transect. This was easy to observe on a wide scale as the survey camera approached and left the seabed at the start and end of each transect; Figure 12 shows a screen shot from each survey. Without revisiting the location of each transect to reassess the bed type present it is not possible to explain why these differences in physical structure were observed and what processes, natural or anthropogenic, may have influenced them.
In summary, the available data from the Summer Isles maerl habitats cannot be used to test for ‘before / after’ effects of management measures because of the irregular seasonality of the data and the heterogeneity of maerl bed type over the survey area, coupled with the low number of replicate transects over any single maerl bed or in any one year. A discussion of future survey options and assessment techniques follows in the next sections.
Figure 12. Screenshots from survey transects WR41 in 2014 (top) and WR104 in 2018 (bottom) showing differences in the physical structure of the maerl bed.
Click for a full description
The dark areas in each image are mostly live maerl, distributed as an irregular mosaic in 2014 and as regular parallel mega-ripples in 2018. Images extracted from survey video.
Statistical power of detecting change at community level
The power of a statistical test to detect change is an important consideration in the design and execution of any experiment or monitoring programme. The collection of too few samples might mean that incorrect conclusions are reached if data cannot demonstrate significant differences when they are known to exist (Type II errors), whilst the collection of too many samples can be a waste of resources. Power analysis therefore is important in predicting future survey and analytical costs while ensuring that data collected are fit for purpose.
Power analysis of multivariate tests (specifically ANOSIM) on community composition data are not commonly employed owing to the problematic and complex nature of such a task. However, multivariate tests are sensitive to changes in community composition and this has been demonstrated in the examples from Arran discussed (Section 4.1) and other studies cited (Section 4.3) whereby, for the former at least, significant statistical differences existed even when significant ecological changes were not thought to have occurred.
Anderson & Walsh (2013) have demonstrated that the ANOSIM test is very sensitive to heterogeneity in the data with results indicating that ‘some differences’ exist: be that in terms of location or time (differences in means), dispersion (variances) or correlation (structure). Determination of the source(s) of these differences then requires further investigation of the data in terms of SIMPER and/or BEST analyses, MDS ordinations or tests on community diversity parameters to understand the nature of these differences and whether or not they are ‘real’ or ‘significant’ ecological changes. Examination of these further outputs will require expert knowledge of the sampling procedures used and the species involved, enabling a sensible and meaningful conclusion to be drawn. In turn, this reduces the likelihood of concluding perhaps incorrectly that differences in the data definitely translate into ecological relevance.
A benthic community may show significant annual fluctuations in taxon abundances and composition while maintaining the core taxa and ecological functions of those communities (e.g. Sebens, 1985a, cited in Hartnoll (1998), Sebens, 1985b). PERMANOVA tests are less sensitive to heterogeneity in the data but for relatively simple one-way analyses such as those relevant to the maerl bed habitats (i.e. temporal change within any given site) they will give very similar results to ANOSIM tests (Anderson et al., 2008). Therefore, we consider that the ANOSIM, SIMPER and BEST tests used here are appropriate, simple to run and have sufficient power to detect and explain inter-annual differences in community data. It is then expert judgement coupled with an understanding of the inherent variation in the data which will be critical to determining the ecological significance of any changes. Whether or not any changes are regarded as positive or negative from an ecological perspective would depend on the nature of any changes observed e.g. increases or decreases in species and or overall abundance, range changes of species, arrival of non-native taxa, changes in sediment composition etc.
With the knowledge that multivariate tests have the power to detect differences in community data but in the absence of a suitable technique to assess their power, another method was trialled to determine the number of images to analyse that could generate data representative of each transect surveyed. The statistical analyses presented previously were based on the analysis of 10-15 images per transect which represented between 1.8 m2 and 2.7 m2 of seabed. The data were then averaged and in the case of count data, standardised to represent 1 m2 before conversion to a single column of data representative of each transect; this was then cross-referenced with the complete video tow. Image analysis itself takes time, resources and consequently has financial costs. The question therefore arises: how many individual images need to be analysed to provide a transect sample representative of the benthic community surveyed. Taking the Arran Holy Isle data, raw quantitative image analysis data were available for each image for 2018 for the eight transects analysed (Section 4.1.2). For each transect either five or six images were chosen at random to ensure at least 1m2 of seabed was represented. Whether five or six images were used depended on the average visible area in each set of images per transect, calculated using the laser scales. These image data were then used to recalculate SACFOR scores for each transect before conversion using the methods described previously (Section 2.2.3; (Strong & Johnson, 2020). The data were then imported to PRIMER for multivariate analysis alongside the original data. An MDS plot showed the distribution of 2018 transect data generated from 5-6 images to be almost identical to that generated from 10-15 images with both sets of data remaining equally distinct from the 2014 data (Figure 13). An ANOSIM test comparing the two sets of 2018 data showed no significant difference between the data generated from either the full set or the randomly selected subsets of images (ANOSIM R=-0.06, P=0.766).
Figure 13. MDS plot of transect community composition data collected from maerl biotopes around Holy Isle off south Arran using different numbers of images. September 2014 (▲) and in 2018: 10-15 per transect (■) or 5-6 per transect (▲)(n = 18).
Click for a full description
There is one cluster for samples from 2014 where 10-15 images were collected per transect, and one cluster for samples from 2018 where 5-6 images were collected per transect.
During the process of recalculating the SACFOR scores for each transect based on a reduced number of images, it was apparent that several taxa/taxon-groups no longer featured in some transect samples. These variables were often present in low numbers / percentage cover; the process dictates that some loss of resolution from the data should be expected. Adding these variables back into the data set as ‘Present’ (0.1 on the SACFOR scale) as if they were noted from the playback of the entire video, made no discernible difference to the overall outcome (ANOSIM R=-0.064, P=0.779). However, several of these taxa could potentially be key in assigning biotopes to the habitats surveyed e.g. the gravel sea cucumber Neopentadactyla mixta or red filamentous algae ‘Rhodophyta – filamentous’. Losing such taxa from the analysis risks drawing incorrect conclusions from the data unless further measures to account for their presence when interpreting the results are taken. Table 2 lists the taxa/taxon groups and the transect samples from which they disappeared during this rationalisation exercise.
Table 2. Taxa/taxon groups and the transect numbers from which they were no longer recorded following reduction of image samples used to characterise a transect from 10-15 images down to 5-6 images.
| Species / species group | Transect number |
|---|---|
| Neopentadactyla mixta | 63, 65 |
| Bivalve siphons | 61 |
| Lanice conchilega | 66 |
| Pecten maximus | 60 |
| Porania pulvillus | 66 |
| Terebellidae | 66 |
| Gibbula sp. | 60 |
| Asterias rubens | 60 |
| Marthasterias glacialis | 56, 61, 65 |
| Live maerl | 56 |
| Membranipora membranacea | 63 |
| Bryozoa (turf) | 56 |
| Phaeophyceae (filamentous mat) | 64 |
| Rhodophyta – foliose | 60 |
| Ulva sp. | 56 |
Whilst the results suggest that analytical cost savings could be made through the analysis of fewer images, the effect of analysing a greater number of images (>15) has not been tested and may, or may not, generate a more accurate profile of the community composition. For this to be tested the survey camera would need to set down on the seabed a greater number of times per transect to generate more images for further statistical comparison; only then can more confidence be placed in the data gained from analysing fewer images. Inevitably, more samples will record more taxa/taxon groups but if these are few in number or low percentage cover then they will make lower contributions to overall sample similarities. Taxon accumulation curves for the images analysed along each transect suggest that along some transects the majority of taxa/taxon-groups might be recorded in <10 images with the curves becoming asymptotic thereafter (Figure 14). Data for other transects indicated continued accumulation of new taxa beyond 10-15 images. Accumulation curves vary between transects and will also likely vary between maerl beds, being dependent on the inherent taxon richness, density and their conspicuousness within any single bed. These results are discussed further in Section 5.1.3.
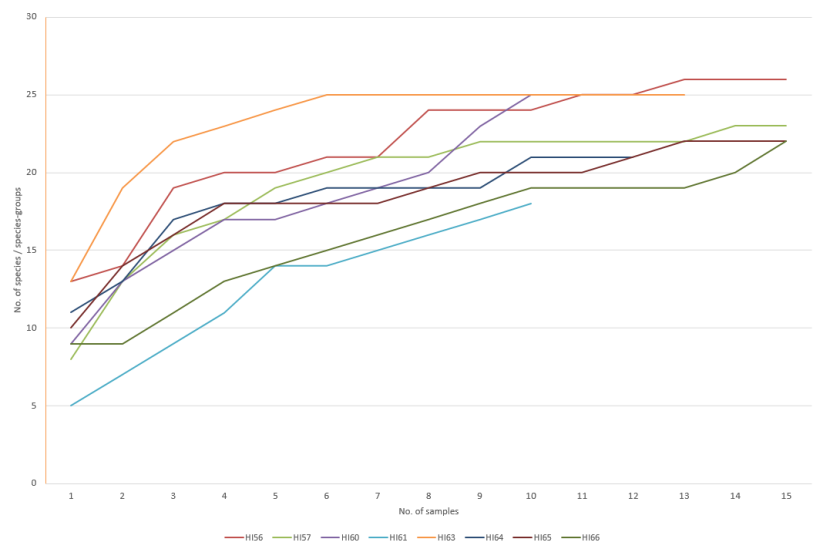
Figure 14. Species accumulation curves with increasing number of still images (samples) analysed per transect (HI##) for maerl habitats surveyed off Holy Isle, south Arran in 2018.
Click for a full description
Whilst the results suggest that analytical cost savings could be made through the analysis of fewer images, the effect of analysing a greater number of images (>15) has not been tested and may, or may not, generate a more accurate profile of the community composition. For this to be tested the survey camera would need to set down on the seabed a greater number of times per transect to generate more images for further statistical comparison; only then can more confidence be placed in the data gained from analysing fewer images. Inevitably, more samples will record more taxa/taxon groups but if these are few in number or low percentage cover then they will make lower contributions to overall sample similarities. Taxon accumulation curves for the images analysed along each transect suggest that along some transects the majority of taxa/taxon-groups might be recorded in <10 images with the curves becoming asymptotic thereafter.
Statistical power to detect change at species level
Again, it must be emphasised that the application of statistical tests to data which were not necessarily collected to answer specific questions can be fraught with difficulties; results should be interpreted with care and re-sampling might be required to test specific hypotheses. The data collected from the video and still images were originally intended to describe the overall biological communities present and may not be best-suited to target specific organisms or other parameters of interest. Sampling efficiency will vary from taxon to taxon depending on attributes such as behaviour, lifeform, conspicuousness etc. and from survey to survey depending on environmental conditions and the equipment used.
Despite this, the present data can be explored to determine which taxa or taxon-groups might yield useful information on their own with regard to population changes, and such data might be suited to univariate analysis. The results of the BVSTEP analysis (Section 4.1.2) on the Holy Isle data identified the main taxa and taxon-groups responsible for the overall temporal community patterns recorded. These included the anemone Cerianthus lloydii, and algae including live and dead maerl and filamentous red algae amongst others. Other species that might be of interest to monitor include the tube worm Chaetopterus sp. (relatively easily identifiable) and the gravel sea cucumber Neopentadactyla mixta, the latter of which characterises the maerl biotope SS.SMp.Mrl.Pcal.Nmix, ‘Phymatolithon calcareum maerl beds with Neopentadactyla mixta and other echinoderms in deeper infralittoral clean gravel or coarse sand’.
In order to know how many samples will be required to detect a significant change, it is necessary to determine what level of change would be considered significant. Section 5.2 discusses the types and level of inherent variation that can exist in the kind of ecological data collected here. Species data for Plymouth Sound and Estuaries SAC have been shown to exhibit variation of x2 or x10 over two consecutive years and longer time periods despite studies concluding that no significant ecological changes had occurred (Moore, 2000, Howson et al., 2005). The power analyses presented here for selected, taxon-groups and physical parameters examine the effect of varying the number of samples (transects) to detect changes of halving (-50%), doubling (+100%), tripling (+200%) or increasing by an order of magnitude (x10, or +1000%) the selected variables over either a five or ten year period. Power analyses were not completed on every variable owing to the time and resources available for the present exploratory study. The variables selected for power analysis and the reasons for their selection are provided (Table 3). These selected variables represent a range of abundances and lifeforms with different levels of variation and provide an overall indication of the number of sample replicates likely to be required to gain an understanding of the significance of the results.
Table 3. Variables selected for power analysis with justification for the selection.
| Variable | Reason for inclusion in exploratory power analysis |
|---|---|
| Anemone – Cerianthus lloydii | A common and abundant species found in several maerl beds, sometimes at high densities. Medium body size, easily and efficiently recorded from video analysis. Highly variable counts between transects. |
| Gravel sea cucumber – Neopentadactyla mixta | An uncommon species in the data analysed but occurs with increasing depth and can be common in patches. Medium body size, easily and efficiently recorded. Highly variable counts between transects in a bed – perhaps due to depth variation. |
| Ascididae – sea squirts cf. Ascidiella aspersa | Smaller and sometimes slightly more difficult to record than the two species above. Occurred in high numbers in the samples analysed with much lower variation between transects compared with N. mixta and C. lloydii |
| Tube worm – Chaetopterus sp. | A patchy and sometimes sparse species where it occurs in several maerl beds. Medium body size, tubes are distinct and efficiently recorded from video analysis. Variable counts between transects. |
| Rhodophyta – filamentous | Specific growth form of red algae, commonly occurring in many shallower transects albeit at low percentage cover. |
| Total algae | Broad scale assessment of total algal percentage cover. Occurred in all transects but varied considerably. |
| Dead maerl | Conspicuous physical structure facilitating quantification by percentage cover. Potential to vary widely throughout a survey area. |
| Live maerl | Conspicuous and easy to identify. Key to characterising each maerl bed being monitored. Often patchy and therefore highly variable. |
| Sand | Percent sand visible within a maerl bed could be an indicator of bed disturbance with consequences for the physical structure of the substrate and species diversity. Often varied with depth. |
As stated in Section 2.2.3, the univariate power analyses were completed using the MONITOR programme (Gibbs & Ene, 2010), outputs from which indicate the power to detect the specified levels of change over the specified time range based on the known or estimated population mean and standard deviation. In the exploratory power analysis presented here, the data used were those from analysis of images recorded from the Holy Isle maerl bed off south Arran in 2018. The power analyses were run on the default assumption of wanting a 90% chance of detecting a real change with the significance level of that change set at the standard 5% level. These parameters can be altered in future if there was reason to accept the findings of less powerful tests or to set significance thresholds at lower levels. Although there are no established conventions, it is common practice to seek power estimates exceeding 0.80 (Cohen, 1988) i.e. a monitoring program with power estimates in excess of 0.80 would detect trends, should they occur, >80% of the time. A default value of 0.9 desired power is used here to give confidence that any population changes are captured. “In a monitoring context [a significance (p) level of] 0.1 or 0.2 is perfectly reasonable depending on the seriousness of missing important trends versus the costs of exploring false detections. Justifying significance level is a critical part of designing monitoring programs” (Gibbs & Ene, 2010). Two monitoring designs were entered into the MONITOR interface depending on the hypotheses that might wish to be tested:
- Firstly, the power analyses were conducted on the basis of comparing abundance or percent cover values for a variable on a ‘before / after’ (management measures) to test a hypothesis that a two-fold (100%) increase or a ten-fold (1000%) increase might occur in those variables (Table 4).
- Secondly, data were analysed on the basis of wanting to test for changes of specified magnitude in a parameter over time, as if surveyed on an annual or biannual basis and to monitor for time trends (regression) in their abundance of percentage cover values over either five or ten years. These were examined on the basis of surveying either 8, 20 or 30 transects per maerl bed on each monitoring occasion.
Of the nine variables tested, four had standard deviation (SD) values either almost equal to or greater than the mean abundance/percentage cover values (C. lloydii, N. mixta, live maerl and % sand), whilst all the other variables showed high SD values relative to the mean, with the exception of ‘Ascididae’ (Table 4).
The first set of power analyses showed that unrealistically high numbers of transects would be required to detect a two-fold change in abundances (Table 4). A more realistic number of transects (~10) still had good power to detect changes of an order of magnitude or more in most variables although a high number of replicates would still be required to detect changes in less common and highly variable species e.g. N. mixta.
Table 4. Power analysis results (from MONITOR v11.0.0) showing the number of replicate transects required for selected variables to achieve a power of at least 0.9 at significance level p=0.05, for before / after comparisons e.g. to assess the efficacy of implementation of management measures.
| Variable | Population Estimated mean | Population Estimated stdev | No. of transects required to detect x2 increase (100% change) | No. of transects required to detect x10 increase (1000% change) |
|---|---|---|---|---|
| Anemone – Cerianthus lloydii (count) | 35.2 | 36.1 | 45 | 9 |
| Gravel sea cucumber – Neopentadactyla mixta (count) | 0.4 | 0.9 | 200 | 25 |
| Ascididae – sea squirts cf. Ascidiella aspersa (count) | 12.2 | 5.3 | 10 | 4 |
| Tube worm - Chaetopterus sp. (count) | 2.1 | 1.6 | 30 | 6 |
| Rhodophyta – filamentous (%) | 6.2 | 4.6 | 20 | 5 |
| Total algae (%) | 10.2 | 7.1 | 45 | 8 |
| Dead maerl (%) | 40.5 | 28.8 | 24 | 6 |
| Live maerl (%) | 0.9 | 1 | 55 | 10 |
| Sand (%) | 22.4 | 22.6 | 45 | 8 |
Table 5. Power analysis results (from MONITOR v11.0.0) showing the varying power (at significance level p=0.05) of different numbers of replicate transects to be surveyed annually to monitor time trends for selected variables. “-“ indicates no analysis completed, power would be <0.9.
| Variable | No.of transects | Change Decrease Half (50%) 5Yrs | Change Decrease Half (50%) 10 Yrs | Change Increase 2 fold (+100%) 5Yrs | Change Increase 2 fold (+100%) 10 Yrs | Change Increase 3 fold (+200%) 5 yrs | Change Increase 3 fold (+200%) 10 Yrs | Change 10 fold (+1000%) 5 Yrs | Change 10 fold (+1000%) 10 Yrs |
|---|---|---|---|---|---|---|---|---|---|
| Anemone – Cerianthus lloydii | 8 | <0.010 | <0.010 | <0.010 | <0.010 | <0.010 | 0.014 | 0.693 | 0.735 |
| Anemone – Cerianthus lloydii | 20 | 0.239 | 0.384 | 0.404 | 0.436 | 0.855 | 0.905 | 0.910 | 1 |
| Anemone – Cerianthus lloydii | 30 | 0.379 | 0.555 | 0.538 | 0.616 | 0.967 | 0.988 | 1 | 1 |
| Gravel sea cucumber – Neopentadactyla mixta | 8 | <0.010 | <0.010 | 0.073 | 0.095 | 0.119 | 0.16 | 0.586 | 0.565 |
| Gravel sea cucumber – Neopentadactyla mixta | 20 | - | - | - | - | - | - | - | 0.862 |
| Gravel sea cucumber – Neopentadactyla mixta | 30 | - | - | - | - | - | 0.371 | - | 0.962 |
| Ascidia – sea squirts cf. Ascidiella aspersa | 8 | 0.22 | 0.319 | 0.376 | 0.381 | 0.97 | 0.975 | 1 | 1 |
| Ascidia – sea squirts cf. Ascidiella aspersa | 20 | 0.986 | 1 | 0.998 | 1 | 1 | 1 | 1 | 1 |
| Ascidia – sea squirts cf. Ascidiella aspersa | 30 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Tube worm - Chaetopterus sp | 8 | 0.134 | 0.281 | 0.208 | 0.31 | 0.645 | 0.709 | 0.999 | 0.999 |
| Tube worm - Chaetopterus sp | 20 | 0.467 | 0.77 | 0.672 | 0.741 | 0.999 | 0.999 | 1 | 1 |
| Tube worm - Chaetopterus sp | 30 | 0.661 | 0.882 | 0.906 | 0.951 | 1 | 1 | 1 | 1 |
| Rhodophyta – filamentous | 8 | 0.223 | 0.358 | 0.372 | 0.379 | 0.77 | 0.795 | 1 | 1 |
| Rhodophyta – filamentous | 20 | 0.627 | 0.774 | 0.702 | 0.789 | 0.994 | 0.996 | 1 | 1 |
| Rhodophyta – filamentous | 30 | 0.795 | 0.886 | 0.878 | 0.994 | 1 | 1 | 1 | 1 |
| Total algae | 8 | 0.041 | - | 0.05 | - | 0.16 | 0.151 | 0.95 | 1 |
| Total algae | 20 | 0.607 | 0.793 | 0.78 | 0.866 | 0.999 | 0.999 | 1 | 1 |
| Total algae | 30 | 0.831 | 0.927 | 0.946 | 0.997 | 1 | 1 | 1 | 1 |
| Dead maerl | 8 | 0.176 | 0.287 | 0.28 | 0.289 | 0.718 | 0.787 | 0.994 | 1 |
| Dead maerl | 20 | 0.57 | 0.867 | 0.77 | 0.858 | 0.995 | 1 | 1 | 1 |
| Dead maerl | 30 | 0.863 | 0.974 | 0.94 | 0.971 | 1 | 1 | 1 | 1 |
| Live maerl | 8 | - | 0.163 | - | 0.171 | 0.402 | 0.439 | 0.96 | 0.96 |
| Live maerl | 20 | 0.246 | 0.337 | 0.319 | 0.33 | 0.806 | 0.864 | 0.999 | 1 |
| Live maerl | 30 | 0.329 | 0.453 | 0.488 | 0.561 | 0.954 | 0.971 | 1 | 1 |
| Sand | 8 | 0.081 | 0.13 | 0.116 | 0.117 | 0.323 | 0.392 | 0.938 | 0.979 |
| Sand | 20 | 0.264 | 0.457 | 0.405 | 0.46 | 0.904 | 0.925 | 1 | 1 |
| Sand | 30 | 0.51 | 0.666 | 0.605 | 0.719 | 0.985 | 0.999 | 1 | 1 |
The second set of power analyses was run assuming populations or physical variables might exhibit increasing or decreasing trends (regression) over time following the implementation of management measures. For each variable, the analyses were first run on the basis of using just eight transects since this was the number available to generate the test data. If the desired power of 0.9 was not obtained, the number of replicates was increased to 20 and then 30 to test for improvements in power values. The number of replicates was not increased above 30 as this was considered very unlikely to be feasible in terms of available survey resources. In most cases, except for C. lloydii and N. mixta, the power analysis indicated that eight replicate transects would be sufficient to detect changes of an order of magnitude (10-fold) over five monitoring occasions (Table 5). Power to detect changes of 3-fold or less differed between variables, again with lower powers generally being achieved for those with more variable data (higher SD). Again the power to detect trends in populations was lowest in variables which were less abundant and with the highest relative SD values e.g. N. mixta and live maerl. These results are discussed further in Section 5.3.
Discussion
Literature review
The review of available literature for previous maerl monitoring projects in the UK and Europe found that despite a long-standing interest in the protection of maerl, there are no published monitoring protocols designed to detect natural fluctuations within maerl associated communities; neither are there published statistically defendable studies illustrating impacts of or recovery from human disturbance or the success of management measures. There are a few reports that describe procedures designed for long term monitoring of maerl beds using the epibenthos rather than benthic infauna (Wilson et al., 2007, Peña & Bárbara, 2010), but to date results of repeat surveys (if any) are unavailable.
Review of the list of taxa recorded from the South Arran MPA on the Pladda and Holy Isle maerl beds in 2014 and 2018 showed that they are all widely distributed in other subtidal habitats around Scotland and not one of them is restricted to maerl beds. Review of other species lists from maerl beds showed that some taxa have attributes which are important in maintaining the structural integrity of maerl beds: for example some invertebrates with deep burrows or tubes (e.g. Cerianthus lloydii, Chaetopterus, Upogebia) are thought to be important in stabilising the maerl bed sediments. Some seaweed taxa with ramifying rhizoids which have been recorded from maerl beds can play a vital role in stabilising the maerl fragments by binding them together (e.g. some Cladophoraceae, Gelidiella calcicola, Gelidium maggsiae). It has been suggested that widely distributed seaweed taxa which commonly occur in maerl beds would be suitable targets for monitoring (Peña & Bárbara, 2010) (e.g. Cryptopleura ramosa, Dictyota dichotoma, Plocamium spp, Scinaia sp). Three of these taxa are included in the Pladda dataset and all are readily identified using high quality drop imagery.
A monitoring approach using the functional diversity of maerl communities to investigate change over time would be a valuable development. Methods have been proposed which allow for different representations of trait types (quantitative, semi-quantitative or ordinal) applied to either taxon abundances or simple presence / absence community data (Laliberté & Legendre, 2010). Studies on benthic infauna for example have demonstrated a link between the diversity of functional traits within the community and fisheries impacts (Bremner et al., 2003a), while Cadotte (2011) and references therein show that restoring or maintaining species diversity may not protect functional diversity adequately. Steneck & Dethier (1994) developed a very basic classification of functional traits for algae and this approach has now been greatly extended in studies on intertidal taxa (Mauffrey et al., 2020) with the intention of assessing impacts of climate change on these communities. An advantage of the functional traits approach is that it is not limited by the restricted geographical range of the individual taxa concerned and is therefore applicable over a wide geographical area: the same functional traits are represented by different (though possibly related) taxa. A library of functional traits for north east Atlantic taxa is available from MarLIN (2006) and the Arctic Traits Database, additional traits considered relevant to maerl bed functioning can be derived for additional taxa from a review of the available literature. The final selection of functional traits to be used for further study would be made taking account of traits which are relevant to the functioning of healthy maerl bed systems and which are likely to be impacted by the implementation of particular management measures, or the lack of them.
Monitoring programme sampling design and spatial autocorrelation
The following sections discuss several topics related to the sampling design and statistical analyses of the data presented in section 4:
- Monitoring programme sampling design and spatial autocorrelation
- Understanding of data ‘noise’ and the distinction between statistically significant and ecologically significant differences
- Statistical power
- Further considerations for data interpretation
The Scottish maerl beds surveyed to date all vary from one another in terms of extent, depth, exposure, physical structure and species richness. What defines an actual maerl bed is also likely to vary between monitoring areas (boxes) with some being continuous habitats and others more patchy in nature. By tailoring surveys and analyses to specific beds, the best data to monitor change in that particular maerl bed can be obtained in the most efficient manner.
Assuming the maerl bed monitoring programme is likely to continue using drop imagery techniques, some modifications to survey design can be made to ensure that the transects are independent of one another and randomly distributed within each maerl bed to be monitored. At present, transects are located within ‘monitoring boxes’ which extend well beyond the boundaries of maerl habitat. Given the likely limitations of field survey resources and if maerl bed communities are a continued monitoring priority, it must be ensured that sufficient randomly located transects are allocated within the known or at least the anticipated bounds of the maerl beds, rather than throughout the wider monitoring boxes. In the case of the south end of Holy Island and around Pladda, large areas of deep water habitat unsuitable for maerl growth could be excluded, for example. This approach will ensure sample replication is focused on maerl habitats providing robust statistical analyses.
Spatial autocorrelation occurs when samples obtained in close proximity to one another exhibit a tendency to be more similar. Whilst this is expected in biological systems where habitats support similar communities across their range, the occurrence of spatial autocorrelation can imply information redundancy i.e. inefficient sampling if transects are continually placed in close proximity to one another. Furthermore, if samples are taken in close proximity to one another (small spatial scale) and do not fully represent the broader area of interest, the data collected will not necessarily accurately reflect variation of the community over larger spatial scales (Fortin et al., 2016) due to either changes in environment or management measures which may confound data interpretation.
Around Arran, the number of replicate tows recorded as maerl biotope taken to date in each area of maerl (~8-10 per maerl habitat per year) provides a reasonable level of replication for the multivariate analyses undertaken, allowing each maerl bed to be monitored in isolation. By monitoring each maerl bed separately, variation resulting from different environmental conditions across beds e.g. tides, exposure, physical habitat, and even maerl bed type itself can be greatly reduced and temporal changes will more likely be detected by on-going monitoring. The definition of what is considered to be a maerl bed is the subject of a separate study, though it is not necessary to cover the entire bed in the monitoring programme so long as randomly allocated samples are representative of the bed and allow sound conclusions to be drawn.
Given the wide spatial areas over which the Arran maerl beds extend, there is scope to develop a stratified random sampling design for each area to generate independent samples for any given year. Our suggested approach, once the extent of each maerl bed is known or can be reasonably estimated using existing knowledge or modelling, is to divide the area up into sections (based on depth for example) and then randomly select “n” sample points (video transect start points) for each survey. If the maerl habitat is very patchy in the area that the camera drops at, randomly selected locations would only continue to collect data if maerl biotope was identified. Such drops would be made until “n” sample points classified as maerl biotope had been surveyed. Limits can be set on the distance between transects to avoid potential transect overlap. Each year a new set of randomly selected start points can be generated limiting any effects of bed-scale autocorrelation and increasing confidence that any temporal changes detected are occurring throughout the bed and not only in the few areas monitored. The example below for the Pladda maerl bed presents various options for achieving this depending on the balance of available resources and areas of interest.
- Survey method – drop video to give a video view along a transect >100 m2 (approximately 7-10 minutes of video on seabed) + high quality still images for analysis to generate abundance / percentage cover / SACFOR data.
- Define the spatial area of interest – for Pladda, maerl is present to the northwest and northeast of the island with the area bisected by a shallow, rocky reef which restricts access by the survey vessel. Therefore two polygons are generated in the GIS to define the survey area based on the known extent of the maerl bed.
- Create points to be surveyed using a GIS. Random points are generated inside polygons, ‘nearest neighbour’ limits being set to ensure transects do not overlap. The example image below shows 15 stations per depth band set with a minimum distance apart of 50 m (Figure 15), ten of these points (for example) are selected on the survey day taking into account the effects of wind and tide on the vessel to ensure the survey data collected remain within the area of the maerl bed. The next survey year a different set of survey points would be used thereby avoiding temporal non-independence (Underwood, 1997).
- The number of survey points over which to run transects in any given year will depend on the variable of interest. Ideally this would be derived from a power analysis although for multivariate analyses this is a complex and difficult task (see Section 4.4). It is more likely that the number of survey points would be determined based on resource availability with regard to survey duration and subsequent image analysis and reporting.
- As the 2018 data for Holy Isle illustrated, depth is likely to be an important environmental variable influencing the communities recorded. It might be better therefore, to focus efforts on either a single depth band or account for it as a known random factor in subsequent analyses. To mitigate for unpredictable survey conditions more random points than required for the final survey would be provided and then locations selected on the day such that all transects remain within the survey box.
Figure 15. Example of randomly generated transect start points within the Pladda maerl bed, Arran, stratified by depth (><10 m) within each survey box.
Click for a full description
(Contains public sector information, licensed under the Open Government Licence, from Marine Scotland and OS data © Crown copyright [and database right] 2021. Not to be used for navigation). Pladda samples
In Wester Ross, transects completed to date within the Summer Isles have varied in location and across maerl beds with distinct physical and biological differences. Pooling community data across these sites to represent maerl communities in the wider area will result in large inherent variability in the data and reduce the likelihood of detecting any meaningful community changes; this is particularly relevant if some sites were previously fished and others were not. High-resolution fishing activity data are not available to assess possible impacts for any individual maerl beds. Although some of the maerl beds within the Summer Isles appear limited in spatial extent, it might be possible to avoid the complications caused by broader scale environmental variation by increasing the number of randomly located transects made during each survey event over each maerl bed to at least 4-5 replicates per bed. Repeated monitoring over time followed by multivariate community analysis would then give an indication of any temporal changes in community composition of that bed. Ultimately, the available resources will play a key role in determining exactly which and how many maerl beds can be monitored in this way. However, if such an approach can be applied to multiple maerl beds, preferably selected based on knowledge of past or present exposure to fishing pressure, then more robust conclusions can be drawn with regard to changes in community quality.
For small maerl beds with restricted access for a survey vessel towing drop video, diver-based monitoring of fixed stations (pseudo-replicates) could be an alternative approach. This approach has potential to cause damage to slow-growing organisms by divers and survey equipment which could influence community data over time. Furthermore, this sort of approach only provides information for each fixed station, the results can only be applied to that site. Without replication of treatment at other sites and comparison against controls without management no wider conclusions can be drawn. If the study is confined to fixed point sampling (pseudo-replicates) within a bed the sampling regime will not provide independent samples either temporally or spatially. Conclusions derived for each sample can only be applied to that one sample location, they cannot be extrapolated to the wider study area of the local bed let alone to any other maerl bed.
Alternatively, gathering drop imagery instead of video transects would allow the random allocation of five or six sampling stations with 10-15 still images recorded at each. From these images a single measure of density / percent cover data per taxon can be generated for each station, thereby providing five or six random replicate samples per maerl bed per survey. For each new survey, sampling stations can be re-allocated at random throughout each bed.
If the subject of interest is a single variable, or selection of single variables then a different and more targeted sampling approach might be required to ensure sufficient replicates are available for meaningful statistical analyses to be carried out. The power of univariate data gathered using drop imagery is discussed in Section 5.3.
Determination of data ‘noise’ - statistically significant vs. ecologically significant differences
Analysis of the converted SACFOR data for Arran has shown significant dissimilarities between the data from 2014 and 2018 (Figure 4 and Figure 6). It must be decided whether or not this result represents significant ecological change, which is not possible at this point in the monitoring programme. As stated in Section 2.2.1, the analyses and results presented here are for proof of concept or testing analytical power only; the results should not be taken as direct evidence of real differences in maerl bed condition, community assemblage or species abundance between surveys at this stage. Consequently the results are discussed in a very general manner so as not to imply actual change has or has not occurred with regard to any specific variables.
Marine epibenthic communities do not exist in a stable state. Relative abundances of individual species fluctuate from year to year, a reflection of various factors including recruitment success, interspecific competition, food availability, changing abiotic variables or any of these factors in varying combinations in any given year. These factors can all create variability or ‘natural noise’ in multivariate community data. In addition to these natural factors ‘procedural noise’ might result from variations in environmental conditions at the time of survey, changes in survey equipment (technological improvements) or surveyors (varying levels of expertise), variation in analytical technique etc. This combination of natural and procedural noise in the community data will be reflected in the similarity matrices following analysis. This ‘noise’ in the data may be influencing the comparison of both the Holy Isle and Pladda data between 2014 and 2018 in the examples provided. Without including further data for surveys from multiple years it is not always possible to determine whether the differences in abundance indicated in the SIMPER analyses (Annex 6 and Annex 7) for taxa and taxon-groups in the maerl beds off Pladda and Holy Isle resulted from natural or procedural noise or changes in management practice. Taking the SIMPER analysis for Pladda as an example, it could be considered feasible for the abundance of ascidians to fluctuate by two orders of magnitude over four years or for filamentous red algae and Desmarestia sp. to increase by an order of magnitude if prevailing environmental conditions during those times favoured or disadvantaged those taxa (i.e. these might be real changes). However, the SIMPER results also suggest that the percent cover of dead maerl decreased by an order of magnitude whilst live maerl concurrently decreased by over an order of magnitude – where did the dead maerl go? Only more comparable annual survey data can enhance our understanding of the roles played by image quality and / or spatial heterogeneity in adding noise to the data. Further investigations are required on larger data sets before firm conclusions can be drawn regarding the impact of management measures on these benthic communities. The results of the analyses are inconclusive at this pilot study stage but the tests undertaken show that the multivariate converted SACFOR data are sensitive to changes in community abundance patterns.
Natural and procedural noise in community data can often result in statistically significant differences; hence interpretation of analytical results must be approached with care to discriminate between statistically significant differences in the data and ecologically significant changes in the habitats and communities being monitored. During surveys of rocky reef communities (percentage cover in quadrats) at three different sites in Plymouth Sound cSAC in 1998 and 1999, there were large differences (often greater than two-fold, some by an order of magnitude) in the abundances of characteristic species and considerable differences in the list of species present (>40% change); many of these differences were statistically significant and were greatest at tide-swept sites (Moore, 2000). With these differences observed over just two consecutive years (and similar in the present study), the potential for natural variability to be even greater over longer time periods should not be underestimated. Moore (2000) observed that some changes in the Plymouth Sound quadrat data were due to a combination of recording variability (procedural noise) and behavioural factors of the taxa (e.g. expansion or contraction of anemones and cryptic behaviour of mobile species), but most were due to real population changes (natural noise). Population changes could be manifested as changes in year-on-year percentage cover by slow-growing, long-lived species or could be due to a strong element of ‘chance’, subject to numerous environmental factors that affect whether settlement and growth of new individuals (both short and long-lived) will occur in the same place in subsequent years i.e. stochastic and patchy recruitment. Despite these population changes, communities were considered to be “essentially the same” in both years given that the biotopes remained the same and the species recorded were largely similar in each year; the study concluded that monitoring programmes need to consider these high levels of natural population fluctuations and set targets to reflect such levels (Moore, 2000).
Moore (2000) suggested that if both broad scale transect surveys (MNCR phase II) and quadrat surveys (cell frequency counts) identified changes in communities then the changes were more likely to be real rather than if only one survey method indicated that change had occurred. This is due to the different spatial scales that each technique encompasses (with associated different sampling efficiencies for individual taxa), therefore if both methods suggest change then greater confidence can be attributed to the conclusions. This approach was also taken by Howson et al. (2005) in subsequent surveys in Plymouth Sound SAC. The MNCR surveys provided a broader contextual understanding of the habitats over and above the smaller scale of the quadrat surveys and helped conclude that again, despite statistically significant differences being detected, real, ecologically significant changes had not occurred. In a further study of the quadrat and MNCR data from the Plymouth Sound SAC sites ranging from 1998 to 2019, Northen & Doggett (2020) analysed all data sets available for the survey sites. As previously reported, despite statistical analyses indicating significant differences in the data recorded between years, there was insufficient evidence to support any assertion that significant ecological change (improvement or degradation) had occurred at any of the sites. Many of the MDS ordinations showed non-directional interannual variation in the community data with data from 2019 being more similar to that from 1998/1999 compared with other years at some survey sites i.e. the MDS plots were reflecting noise inherent in the data rather than significant ecological change.
Similar results have been obtained by the present authors from in situ surveys of fixed quadrats using cell-frequency counts in Milford Haven within the Pembrokeshire Marine SAC (client confidential). In the on-going monitoring programme, multivariate analysis of eight consecutive years of data suggests significant differences between data between some years within each site. However, MNCR Phase II SACFOR data recorded annually across each survey site do not support any assertion that real and significant ecological changes have occurred. Univariate analysis of selected taxa or taxon-groups showed cyclical, multi-year patterns of recruitment and die-off that play a major role in influencing the results of the community analyses.
Hartnoll (1998) discussed several studies examining change in circalittoral benthic communities in temperate environments. Although many of these were focussed on rocky reef communities, the same principles surrounding community variation apply. Studies from the UK around Lundy and the Scilly Isles showed considerable stability in faunal turf communities given the longevity of many conspicuous taxa (Fowler and Pilley, 1992, cited in (Hartnoll, 1998). However, studies on the Swedish west coast demonstrated the potential for considerable natural variability in certain faunal turf communities, with some flipping from one stable state to another within the space of a single year and then remaining stable for up to a decade at a time (Lundälv, 1996, cited in Hartnoll (1998).
Given the apparent sensitivity of multivariate analyses to differences in community data within a maerl bed (see Section 4.1), there can be high confidence that any major differences in abundance or species presence or changes from one community type to another would be detected, the strength of which would also be reflected in the ANOSIM R value when compared with other more similar years. The Swedish study concluded that the more sheltered the environment, the more stable a community was likely to be; a finding in agreement with Moore (2000). A further series of long-term observations from the US east coast noted that, despite year-on-year variation in abundance of component species, a change in the “overall character” (equivalent to ‘biotope’) of the study sites was not observed. This finding corresponds well with both the Plymouth and Milford Haven studies cited above where the biotopes have not changed despite significant statistical variation in the community data (Sebens, 1985a, cited in Hartnoll (1998); Sebens, 1985b). The variation found in the Plymouth and Milford Haven studies could be a natural ‘stable-yet-dynamic’ characteristic of the communities being monitored.
The potential for maerl bed habitats to exhibit this ‘stable-yet-dynamic’ characteristic or ‘natural noise’ needs to be considered in order to determine whether or not statistically significant differences in temporal data within a maerl bed relate to ecologically significant change. Seasonal monitoring of epibenthic algal communities in three maerl beds in north western Spain over a 14 month period showed significant differences between sites and seasons (Peña & Bárbara, 2010). Furthermore, MDS plots of seasonal algal community samples on each maerl bed suggested temporal differences in the summer season algal composition between 2005 and 2006; pairwise statistical analyses were not presented to confirm if these differences were significant although they appeared very likely in at least two of the three surveys.
The analyses presented here (Section 4.1) do not allow the determination of the level of influence of natural cycles or noise in either the Holy Isle or Pladda maerl beds since only two time periods have been assessed alongside a concurrent change in management measures. Additional and comparable monitoring data across multiple years (potentially at least five annual sampling years) for the Scottish maerl beds examined here would be required to gain a better understanding of the noise inherent in the SACFOR community data.
Multiple reference locations will serve to increase confidence in conclusions regarding any management-related changes observed across the managed sites if similar changes are not observed in the reference locations. Southward (1995) makes the point that to determine the natural variability of a marine ecosystem, the monitoring period must reflect the expected lifespans of the dominant organisms and / or the organisms of interest and the time scales of the major environmental factors that influence them. In practical terms for the present study this may not be possible since, whilst certain short-lived species can vary considerably from year to year (e.g. the sea squirt Ciona intestinalis lives up to two years, (Jackson, 2008)). In terms of abundance and / or spatial distribution, others can live much longer (e.g. the sea squirt Ascidia mentula - up to eight years, (Svane & Lundälv, 1981)), while other taxa may out-live even multidecadal natural cycles such as the North Atlantic Oscillation (for example the soft coral Alcyonium digitatum can live up to three decades (Budd, 2008)). Maerl itself is particularly long-lived, with an estimated thallus life span of 20-100 years (Jackson, 2003). Whilst on-going monitoring (particularly relatively early-on in a monitoring programme) will not be able to account for this level of natural variation over different time-scales, it is important to consider such variables in determining the causes behind any changes observed in the data in addition to possible sources of procedural noise. Given likely resource constraints on monitoring programmes, detailed longer-term monitoring at fewer, carefully selected locations (based on factors such as maerl quality / type, known fishing intensity etc.) might provide more robust data than that spread more thinly over a greater number of locations.
Expert opinion, detailed knowledge of species ecology and careful interpretation of data from a variety of sampling regimes together with statistical evidence are critical in determining whether perceived and / or statistically significant changes in community data can be classed either as ecologically significant or the result of ‘noise’ inherent in the data. If additional sources of evidence from the maerl beds are available such as benthic trawl, core or grab samples across the same time period that can also be used to illustrate the changes (or stability) in the communities being investigated, they might add further confidence to the conclusions being drawn.
Unfortunately in reality, for a multitude of reasons, data rarely exist for several years prior to management changes or environmental impacts occurring within and around study sites, often meaning levels of inherent noise in the data are unknown from the start. This also gives very few replicate samples and/or time periods for before/after analytical comparisons. However, if these confounding factors are understood, explained and an argument based on compelling evidence can be made when interpreting data analyses then the chances of the final conclusions being accepted by all parties can be greatly improved (Underwood, 1997). Sampling design can be improved going forward to enable the most representative data for statistical analysis and expert interpretation to be collected (see Section 5.1).
Statistical power
The investigations of statistical power in Sections 4.3 and 4.4 investigated the sensitivity of multivariate analyses to changes in community composition before considering the number of still images required to adequately represent the communities being surveyed (Figure 14 and Figure 15). The results suggested that the analysis of just five or six images made no significant change to the community patterns observed whilst analysing 10 images often ensured most of the taxa present along a transect were recorded.
Typically, in situ surveys of marine habitats require a minimum area of 25 m2 (5x5 m) of any biotope to be present before it is assigned (Parry, 2019). Although increasing the number of images analysed to ~30 per transect (~4-5 m2 in the present example) will still not come near this value, reviewing the full transect video would provide confidence as to whether or not the images analysed adequately represented the broad scale community present. Ultimately, the minimum area for survey coverage suitable for multivariate community analysis would require further investigation during a future survey. To do this, 30 images from each transect surveyed per maerl bed should be quantitatively analysed. The effect of sample size per transect can then be investigated by randomly selecting ‘n’ images per transect to cover 1 m2, 2 m2, 3 m2 and 5 m2 and generating species accumulation curves. The data could also be subjected to multivariate analyses to test for significant differences; univariate analyses may be possible if sufficient power exists (see Section 4.4). The optimal number of images to analyse for each maerl bed can then be determined and justified. Even if the number of images selected for future analysis remains low, it is recommended to continue to record up to 30 still images per transect in future surveys. Then, if community shift is suspected, further images can always be drawn upon to add confidence to any conclusions reached.
Despite the need to verify the number of replicates required, the present indication is that analysis of fewer than 30 images can capture the principal taxa present within the Holy Isle maerl bed. Could these data still indicate broad scale community changes at a site over time? By analysing fewer images per transect, the number of taxa and taxon groups recorded in some transects was reduced (Table 2). It is inevitable that in analysing more images per transect the overall species count would increase i.e. the more you sample, the more you record (Clarke et al., 2014). This in itself could serve to reduce interannual variation in the data for any given maerl bed as less common species have a greater chance of being recorded each year, thereby reducing dissimilarities between years shown by multivariate analyses.
In a study of temperate reef community stability in Tasmania in relation to increasing ocean temperatures, Stuart-Smith et al. (2010) found that species most important for describing interannual changes were those with lower to mid-range abundances rather than the more common species. Therefore, reliance on more abundant taxa as indicators of ecosystem stability or change could be misleading. Moore (2000) also noted the danger of only recording those taxa present in high abundances given the lack of knowledge with regard to species-specific responses to either anthropogenic or natural impacts, and which taxa could be useful indicators of change. The analysis of fewer images per transect might therefore risk capturing only the more common indicator taxa present. If these taxa are less likely to exhibit variation in response to external factors then the process of analysing fewer images might unintentionally force the data to show community stability when in fact the rarer species might be undergoing significant change. Further investigation of sampling efficiency by means of species accumulation curves using up to 30 images per transect is recommended.
Species accumulation curves can indicate whether or not the number of images analysed per transect should be increased or decreased on a bed-by-bed basis. They can be a useful tool to optimise the number of images required per transect per maerl bed to ensure the data represent the communities in a manner capable of detecting change. If interest leans more toward changes in the abundance of individual taxa and involves univariate statistical analyses, then the number of images and/or transects required to detect meaningful change would differ again.
The power analyses undertaken on selected univariate elements of the community data (Section 4.4) demonstrated that the number of replicate transects required to detect changes, such as a population doubling or halving, was often high and influenced in large part by the high variability of the data. Samples with the highest Standard Deviation required >30 and up to 200 replicates to detect a population doubling. The number of replicates required was greatly reduced if testing for a population change of an order of magnitude or more was chosen, although the numbers required remained high for N. mixta at 25 transects (Table 4). These findings are similar to those of Moore (2000) who conducted power analyses on quadrat data for individual taxa in Plymouth Sound SAC; >30 to 100+ quadrats were often required to detect statistically significant changes in individual taxa. It must be borne in mind that the methods employed in the monitoring surveys here (and those in Plymouth Sound SAC) were designed to represent the communities as a whole and are not sampling techniques designed to target specific variables against which to test hypotheses. Multivariate analysis such as ANOSIM, the power of which is discussed above, provides a powerful and capable tool to detect community-level changes from year-to-year. Even if univariate data did suggest changes were occurring in any chosen variable, it may be advisable to design a targeted sampling survey specific to the taxon or taxon-group of interest to confirm the abundances and add confidence to any conclusions being drawn.
The efficiency of the sampling method must always be taken into account when making any judgements of whether or not change has occurred for any taxon considered in univariate form. For example, the multivariate SACFOR data for Holy Isle from 2014 and 2018 (Section 4.1.2) indicated significant differences between the data from the two years. If Cerianthidae (likely C. lloydii) is considered in isolation then it would appear to have increased from ‘common’ in 2014 to ‘abundant’ in 2018. This ‘increase’ however could be related to bias resulting from the greater apparent depth of some samples from 2018, or may be influenced by other species. The kelp Saccharina latissima was ‘superabundant’ in 2014 but only ‘occasional to frequent’ in 2018 and so might to an extent, have hidden many Cerianthidae from the camera’s field of view in 2014 resulting lower numbers being recorded. Factors such as this could explain some of the inherent noise in the data (see Section 5.2). Even if univariate tests can detect changes of an order of magnitude with confidence, statistically significant vs ecologically significant differences must be distinguished when determining what any changes in the data mean.
The univariate power analyses presented here provide a guide to the levels of replication that are likely to be required in order to detect significant differences and / or trends in the data collected using the present monitoring techniques. The power to detect trends in populations was lowest in variables which were less abundant and with the highest relative SD values (e.g. N. mixta and live maerl). Taxa or taxon-groups with higher statistical power to detect change (<8 transects required) were often conspicuous in nature and / or occurred in high abundances e.g. large ascidians, large tube worms (Chaetopterus sp.), broad scale algal groups (e.g. Rhodophyta – filamentous) and dead maerl. Any of these taxa, or similar, could be useful for on-going monitoring but would need to be selected on a bed-specific basis; taxa with shorter lifespans and rapid recruitment potential such as ascidians could indicate early stages of recovery from bottom-damaging activities whilst reductions in slower-growing, longer lived species could be better indicators of ‘negative’ impacts.
Ultimately selection of individual species for statistical analysis should be based on indications of any change from a multivariate community analysis, followed by assessment of their power to detect change, likely sampling efficiency using image data and finally, consideration of their usefulness in determining change e.g. growth / colonisation rates, susceptibility to disturbance etc. (see examples above and in Section 3.2.1). Consideration of these factors could help guide further analyses to assist in the approach to on-going monitoring and also in judging the validity of using taxon-specific data in isolation from the overall community data. In general, the univariate power analyses presented here suggest few of the individual taxa recorded from images under the current monitoring programme are likely to provide data with sufficient statistical power to reliably indicate change due to the high number of replicates required. It may be possible to identify some bed-specific taxa suitable for these analyses through investigation of the existing data for any given site and then design a specific sampling programme going forward, if that were considered appropriate. Any future efforts/resources should be prioritised at sites that have data based on stratified, random sampling, preferably with multiple ‘before management’ years against which to compare any post-management changes. This will provide the best chance of detecting change in maerl beds around Scotland attributable to management measures.
Further considerations for future monitoring and data interpretation
Converted SACFOR data as per Strong & Johnson (2020) show potential for monitoring community dynamics (see Sections 4.1.1 and 4.1.2) but as discussed above, future monitoring programs need careful experimental design in terms of transect replication and location. The number of images required to generate data representative of each transect also requires further evaluation. Both of these aspects of monitoring need to be developed on a bed-specific basis as a one-size fits all approach is unlikely to be efficient or effective in meeting the end goal of measuring change.
Owing to the paucity of pre-management data in the years prior to protection of these habitats, any statistical analyses may sit best alongside additional expert judgement in assessing whether or not observed changes in maerl habitats represent an improvement as a result of management changes. Reliance on statistics as the sole indicator of change whether due to new management measures or natural change with such limited pre-management data could to be met with criticism. That said, if maerl beds in reference locations exhibited consistently different directional community change (or not) compared to maerl beds that were protected from 2014 onward, then the analytical and data processing methods investigated above could provide compelling evidence for the efficacy of the management measures in use.
The equipment and procedures used in all future surveys should be standardised in so far as possible to keep them comparable with the high quality data obtained in more recent years and reduce procedural noise in the data collected in the future. Suggested protocols have been described in Section 3.3.
Image quality is a key parameter in studies such as this. Review of video data from the 2014 surveys illustrated the lower quality of these videos compared to those from more recent years which may have hindered the ability to distinguish sand and fine sand from other sediment types. This may have explained the differences in recorded substrate types between 2014 and 2018 and influenced the outcome of the BEST analyses (see Section 4.1.3) which seemed biased toward the strong influence of these substrates on the 2018 data and did not associate any abiotic variable directly with the data from 2014. Arguably video data are likely not the best method to generate precise sediment composition data and grab or core samples collected along each transect could provide better indications of substrate composition. However these would inevitably be limited to spot samples and may themselves be compromised in rockier areas. The recent and rapid improvement of high-definition video systems as used in the more recent surveys (e.g. 2018) may be a sufficient substitute and allow more detailed resolution of bed surface sediments along each transect, going forward.
Strong & Johnson (2020) also noted that the simplification of SACFOR data can make association with environmental parameters problematic. The authors found little correlation between biological and physical parameters in their multivariate analyses of simulated and real data. In the present example for Holy Isle data from 2014 and 2018, the BEST test identified sand and fine sand as having a correlation coefficient of 0.832 with the patterns observed in the data but did not identify transect depth which also appeared to influence the communities recorded in 2018 (Figure 8). Interpretation of possible environmental drivers alongside SACFOR data should be undertaken with due consideration of the available data resolution. Analysing the community data in its raw forms as either density or percent cover data (but not both concurrently) could clarify any associations that might exist if the BEST analysis with the converted SACFOR community data failed to yield any meaningful results in the final analysis of larger datasets.
The possibility of using grab samples above to generate more accurate sediment composition data could also be applied to gather faunal data. Whilst this method might generate data sets considered more ‘reliable’ given the rigorous laboratory analysis procedures required to enumerate all the species in a sample, it can be very costly. Furthermore the method can be quite destructive with potential implications for small areas of sensitive habitat - resources might be better placed to increase sample replication with remote sampling methods in the areas of greatest interest and retain comparability with existing data.
Conclusions
This project has drawn on existing knowledge about maerl beds and their associated communities to assess whether the monitoring design and available data formats (i.e. video and still images) are suitable for the application of statistical analysis techniques and / or other semi-qualitative methods using expert judgement. These assessments are intended to determine whether or not changes are occurring in maerl communities since the implementation of management measures.
- Multivariate analysis of converted SACFOR community composition data (as per Strong & Johnson (2020) appears sensitive to temporal differences in abundances of constituent taxa (see Section 4.1). The use of such converted data could help to improve data precision and reduce some elements of methodological variation which may be exacerbated if using empirical values. The converted SACFOR data coupled with standardised image analysis procedures can serve to reduce this potential element of data variation and help to improve the overall accuracy of assessments of community change.
- Analysis of additional community data for the maerl beds around Arran covering multiple years and taking into account factors such as depth and substrate type will enhance understanding of the variability in community data of this kind. In turn this can enhance confidence in any future conclusions regarding the ecological significance of any temporal differences observed.
- Environmental variables such as sediment composition need to be considered with due care as they may vary owing to the resolution of available images from which their values are derived (see Section 4.1.3). Furthermore association of abiotic variables with simplified SACFOR community data can be problematic and may not always show relationships (see Sections 4.1.3 and 5.2, and Strong & Johnson (2020).
- Key taxa and taxon-groups driving community patterns can be identified using the BVSTEP procedure; these can be monitored for change year-on year.
- The power of multivariate analyses to detect differences in community data was discussed and considered to be good based on the current survey design / level of replication (see Sections 4.3 and 5.3).
- The number of still images required for analysis to adequately represent the communities being monitored could be as low as 10 per transect but would benefit from further investigation during a future survey.
- The power of univariate analyses to detect changes in abundance of specific taxa is generally considered to be low with many taxa requiring upward of 20 transects to detect population increases or decrease over 5-10 year timescales (see Section 4.4). Univariate analyses will be best suited to conspicuous taxa or taxon-groups with high abundances and low standard deviations which often require at least eight transects to be completed per maerl bed to attain sufficient power to detect change. Any species data calculated as likely to have sufficient power for univariate analysis are likely to be bed-specific; potential examples from Holy Isle, Arran included large ascidians, large tube worms (Chaetopterus sp.), broad scale algal groups (e.g. Rhodophyta – filamentous) and dead or live maerl (see Section 5.3).
- Generally the use of univariate data from existing data sets is likely to be of little practical use in determining management impacts owing to the low power likely to be obtained from the low number of replicate tows conducted over each maerl bed.
- Examples have been provided for several potential ‘key’ indicator taxa associated with maerl (see Section 3.2.1), together with the anticipated variation in sampling effort that might be required to assess any significant change in these taxa with confidence (see Section 4.1.4).
- The use of various metrics related to the distribution of selected functional traits associated with taxa comprising maerl bed communities would be a valuable avenue to explore, investigating changes in the metrics of functional diversity in a community under different environmental drivers.
- Future monitoring programs need careful experimental design in terms of transect replication and location, and to take into account that data may need to be collected at different spatial scales to understand any significant ecological changes.
- The generation of random transects across each monitoring box for each survey event in the present monitoring programme may not always include sufficient areas of maerl to provide enough replicate samples of maerl habitat within any single bed, particularly if depth gradients result in community differences. For robust statistical analysis specifically of maerl habitats, sample designs need to be bed-specific and first trialled over a series of annual sampling programmes, so that adequate replicates are collected (see Section 5.1).
- Suggestions have been made to improve the standardisation of methods for data collection and analysis, thus reducing methodological variation in community data (see Sections 3.3 and 3.4).
- Where well-replicated historical transects exist over maerl habitats, the data can be re-analysed using the methods described here to provide data for multivariate community analyses. In some cases, univariate analyses can be undertaken if power analysis indicates such an approach is valuable. Careful consideration must be given to image quality and resolution from each survey and deemed as a potential factor if major differences are detected during analysis of biotic or abiotic variables (e.g. see Section 4.1).
- It is likely that confidence in significant ecological change (or not) will be achieved through robust statistical analysis to determine both fine and broad-scale changes confirmed by expert interpretation of the available evidence.
References
Allen, C., Axelsson, M., Dewey, S. & Wilson, J. 2014. Fal and Helford SAC Maerl Drop-down Video and Dive Survey 2013. A report to Natural England.
Álvarez-Fernández, I., Fernández, N., Sánchez-Carnero, N. & Freire, J. 2017. The management performance of marine protected areas in the North-east Atlantic Ocean. Marine Policy, 76, 159-168.
Anderson, M. J., Gorley, R. N. & Clarke, K. R. 2008. PERMANOVA+ for PRIMER: Guide to software and statistical methods, Plymouth: PRIMER-E.
Anderson, M. J. & Walsh, D. C. I. 2013. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecological Monographs, 83, 557-574.
Bárbara, I., Cremades, J. & Veiga, A. J. 2004. Floristic study of a maërl and gravel subtidal bed in the ‘Rîa de Arousa’ (Galicia, Spain). Botanica Complutensis, 28, 27-37.
Barberá, C., Mallol, S., Vergés, A., Cabanellas-Reboredo, M., Díaz, D. & Goñi, R. 2017. Maerl beds inside and outside a 25-year-old no‑take area. Marine Ecology Progress Series, 572, 77-90.
Bernard, G., Romero-Ramirez, A., Tauran, A., Pantalos, M., Deflandre, B., Grall, J. & Grémare, A. 2019. Declining maerl vitality and habitat complexity across a dredging gradient: Insights from in situ sediment profile imagery (SPI). Scientific Reports, 9, 16463.
Bessell, A., de Gregorio, S. & Weir, J. 2016. South Wight Maritime SAC - reef feature attribute survey. Report to Natural England by Fugro EMU. Report No: 160087. Natural England.
Blake, C. & Maggs, C. A. 2003. Comparative growth rates and internal banding periodicity of maerl species (Corallinales, Rhodophyta) from northern Europe. Phycologia, 42, 606-612.
Bremner, J., Frid, C. L. J. & Rogers, S. I. 2003a. Assessing Marine Ecosystem Health: The Long-Term Effects of Fishing on Functional Biodiversity in North Sea Benthos. Aquatic Ecosystem Health & Management, 6, 131-137.
Bremner, J., Rogers, S. I. & Frid, C. L. J. 2003b. Assessing functional diversity in marine benthic ecosystems: a comparison of approaches. Marine Ecology Progress Series, 254, 11-25.
Brodie, J., Williamson, C. J., Smale, D. A., Kamenos, N. A., Mieszkowska, N., Santos, R., Cunliffe, M., Steinke, M., Yesson, C., Anderson, K. M., Asnaghi, V., Brownlee, C., Burdett, H. L., Burrows, M. T., Collins, S., Donohue, P. J. C., Harvey, B., Foggo, A., Noisette, F., Nunes, J., Ragazzola, F., Raven, J. A., Schmidt, D. N., Suggett, D., Teichberg, M. & Hall-Spencer, J. M. 2014. The future of the northeast Atlantic benthic flora in a high CO2 world. Ecology and Evolution, 4, 2787-2798.
Budd, G. C. 2008. Alcyonium digitatum Dead man's fingers. In: Tyler-Walters H. and Hiscock K. eds. Marine Life Information Network: Biology and Sensitivity Key Information Reviews.
Bunker, F. S. D., Mercer, T. M., Howson, C. M., Moore, J. M., Diaz, P., Maggs, C. A. & Kamphausen, L. 2018. Site condition monitoring of maerl beds and seagrass beds in the Sound of Barra SAC 2015 – diving survey. Scottish Natural Heritage Research Report No. 924.
Burrows, M. T., Harvey, R. & Robb, L. 2008. Wave exposure indices from digital coastlines and the prediction of rocky shore community structure. Marine Ecology Progress Series, 353, 1-12.
Cabioch, J. 1966. Contribution à l’étude morphologique, anatomique et systématique de deux mélobésiécs: Lithothamnium calcareum (Pallas) Areschoug et Lithothamnium corallioides Crouan. Botanica Marina, 9, 33-53.
Cabioch, J. 1969. Les fonds de maerl de la Baie de Morlaix et leur peuplement végétal. Cahiers de Biologie Marine, 10, 139-161.
Cadotte, M. W. 2011. The new diversity: management gains through insights into the functional diversity of communities. Journal of Applied Ecology, 48, 1067-1069.
Clarke, K. R., Gorley, R. N., Somerfield, P. J. & Warwick, R. M. 2014. Change in marine communities: an approach to statistical analysis and interpretation, 3rd edition, Plymouth: PRIMER-E.
Coggan, R., Clements, A., White, J. & Golding, N. 2007. Recommended operating guidelines (ROG) for underwater video and photographic imaging techniques
Cohen, J. 1988. Statistical Power Analysis for the Behavioral Sciences, New York: Routledge.
Connor, D. W., Allen, J. H., Golding, N., Howell, K. L., Lieberknecht, L. M., Northen, K. O. & Reker, J. 2004. The Marine Habitat Classification for Britain and Ireland. Version 04.05.
Dales, R. P. 1957. Some quantitative aspects of feeding in sabellid and serpulid fan worms. Journal of the Marine Biological Association of the UK, 36, 309-316.
Fortin, M.-J., Dale, M. R. T. & Ver Hoef, J. M. 2016. Spatial Analysis in Ecology. In: N. Balakrishnan, T. C., B. Everitt, W. Piegorsch, F. Ruggeri and J.L. Teugels eds. Wiley StatsRef: Statistics Reference Online.
Garbary, D. 1976. Life-forms of algae and their distribution. Botanica Marina, 19, 97-106.
Gibbs, J. P. & Ene, E. 2010. Program Monitor: Estimating the statistical power of ecological monitoring programs. Version 11.0.0.
Graham, D. J. & Midgley, N. G. 2000. Graphical representation of particle shape using triangular diagrams: an Excel spreadsheet method. Earth Surface Processes and Landforms, 25, 1473-1477.
Hall-Spencer, J., White, N., Gillespie, E., Gillham, K. & Foggo, A. 2006. Impact of fish farms on maerl beds in strongly tidal areas. Marine Ecology Progress Series, 326, 1-9.
Hall-Spencer, J. M. & Atkinson, R. J. A. 1999. Upogebia deltaura (Crustacea: Thalassinidea) in Clyde Sea maerl beds, Scotland. Journal of the Marine Biological Association of the UK, 79, 871-880.
Hall-Spencer, J. M., Grall, J., Moore, P. G. & Atkinson, R. J. A. 2003. Bivalve fishing and maerl-bed conservation in France and the UK - retrospect and prospect. Aquatic Conservation Marine and Freshwater Ecosystems, 13, S33-S41.
Hall-Spencer, J. M., Kelly, J. & Maggs, C. A. 2010. Background document for Maerl beds. Biodiversity Series.
Hall-Spencer, J. M. & Moore, P. G. 2000. Scallop dredging has profound, long-term impacts on maerl habitats. ICES Journal of Marine Science, 57, 1407-1415.
Hartnoll, R. G. 1998. Volume VIII. Circalittoral faunal turf biotopes: an overview of dynamics and sensitivity characteristics for the conservation management of marine SACs
Hauton, C., Hall-Spencer, J. M. & Moore, P. G. 2003. An experimental study of the ecological impacts of hydraulic bivalve dredging on maerl. ICES Journal of Marine Science, 60, 381-392.
Hawes, W., O'Dell, J., Axelsson, M., Dewey, S. & Allen, C. 2014. St Austell Bay Maerl Survey - Acoustic and Video Analysis. A report to Natural England.
Hernandez-Kantun, J. J., Hall-Spencer, J. M., Grall, J., Adey, W., Rindi, F., Maggs, C. A., Bárbara, I. & Peña, V. 2017. North Atlantic Rhodolith Beds. In: Riosmena-Rodríguez, R., Nelson, W. & Aguirre, J. eds. Rhodolith/Maërl Beds: A Global Perspective. Cham: Springer International Publishing.
Hiscock, K. 1996. Marine Nature Conservation Review: rationale and methods, Peterborough: Joint Nature Conservation Committee.
Hiscock, K. 1998. Biological monitoring of marine Special Areas of Conservation: a review of methods for detecting change
JNCC report No. 284.
Hiscock, K., Southward, A., Tittley, I., Jory, A. & Hawkins, S. 2001. The impact of climate change on subtidal and intertidal benthic species in Scotland. Report to Scottish Natural Heritage.
Hitchin, R., Turner, J. A. & Verling, E. 2015. Epibiota remote monitoring from digital imagery: Operational guidelines
Howson, C., Bunker, F. & Mercer, T. 2004. Fal and Helford European Marine Site sublittoral monitoring 2002. Report to English Nature. English Nature.
Howson, C., Bunker, F. & Mercer, T. 2005. Plymouth Sound European Marine Site Sublittoral Monitoring 2003. Report to English Nature. Aquatic Survey and Monitoring Ltd.
Irving, R. A., Stanwell-Smith, D., Holt, R. H. F., Northern, K. O., Morris, E., Goudge, H., Lindenbaum, C., Stringwell, T., Camplin, M. & Sanderson, W. G. 2012. Across-Wales Diving Monitoring Project, 2007-2011: Standard Operating Procedures. Draft report for the Countryside Council for Wales by Sea-Scope Marine Environmental Consultants and Pelagial Marine Consultants.
Jackson, A. 2003. Lithothamnion glaciale Maerl. In: Tyler-Walters H. & Hiscock K. eds. Marine Life Information Network: Biology and Sensitivity Key Information Reviews.
Jackson, A. 2008. Ciona intestinalis A sea squirt. In: Tyler-Walters H. & Hiscock K. eds. Marine Life Information Network: Biology and Sensitivity Key Information Reviews.
JNCC. 2015. The Marine Habitat Classification for Britain and Ireland. Version 15.03.
Joshi, S., Duffy, G. P. & Brown, C. 2017a. Critical bed shear stress and threshold of motion of maerl biogenic gravel. Estuarine, Coastal and Shelf Science, 194, 128-142.
Joshi, S., Duffy, G. P. & Brown, C. 2017b. Mobility of maerl-siliciclastic mixtures: Impact of waves, currents and storm events. Estuarine, Coastal and Shelf Science, 189, 173-188.
Kaiser, M. J. 1996. Starfish damage as an indicator of trawling intensity. Marine Ecology Progress Series, 134, 303-307.
Kamenos, N. A., Moore, P. G. & Hall-Spencer, J. M. 2003. Substratum heterogeneity of dredged vs un-dredged maerl grounds. Journal of the Marine Biological Association of the UK, 83, 411-413.
Keegan, B. F. 1974. The macrofauna of maerl substrates on the west coast of Ireland. Cahiers de Biologie Marine, 15, 513-530.
Kevrekidis, T., Gouvis, N. & Koukouras, A. 1997. Population dynamics, reproduction and growth of Upogebia pusilla (Decapoda, Thalassinidea) in the Evros Delta (North Aegean Sea). Crustaceana, 70, 799-812.
Konnecker, G. & Keegan, B. F. 1973. In situ behavioural studies on echinoderm aggregations. Helgolander Wissenschaftliche Meeresuntersuchungen, 24, 157-162.
Laliberté, E. & Legendre, P. 2010. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 91, 299-305.
Maggs, C. A. 1984. A seasonal study of seaweed communities on sublittoral maerl (unattached coralline algae) in Galway, Ireland. Progress in Underwater Science, 9, 27-39.
Maggs, C. A. & Guiry, M. D. 1987. Gelidiella calcicola sp. nov. (Rhodophyta) from the British Isles and Northern France. British Phycological Journal, 22, 417-434.
Marine Scotland. 2016. Simple guide to fisheries management measure in Marine Protected Areas
Marine Scotland. 2017. Scottish Marine Protected Areas (MPA) Monitoring Strategy
Marine Scotland. 2019. Improving protection given to Priority Marine Features: Response to Scoping Consultation
MarLIN. 2006. BIOTIC - Biological Traits Information Catalogue. Marine Life Information Network. Marine Life Information Network.
Mason, N. W. H., Mouillot, D., Lee, W. G. & Wilson, J. B. 2005. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 111, 112-118.
Mauffrey, A. R. L., Cappelatti, L. & Griffin, J. N. 2020. Seaweed functional diversity revisited: Confronting traditional groups with quantitative traits. Journal of Ecology, 108, 2390-2405.
Melbourne, L. A., Hernández-Kantún, J. J., Russell, S. & Brodie, J. 2017. There is more to maerl than meets the eye: DNA barcoding reveals a new species in Britain, Lithothamnion erinaceum sp. nov. (Hapalidiales, Rhodophyta). European Journal of Phycology, 52, 166-178.
Mercer, T., Kamphausen, L., Moore, J., Bunker, F., Archer Thompson, J. & Howson, C. 2018. South Arran MPA diver survey of maerl beds, kelp and seaweed communities on sublittoral sediment, and seagrass beds 2014
Scottish Natural Heritage Research Report No. 882.
Mitchell, A. J. & Collins, K. J. 2004. Understanding the distribution of maerl, a calcareous seaweed, off Dorset, UK. In: Nishida, T., Kailola, P. J. & Hollinworth, C. E. eds. GIS/Spatial Analyses in Fishery and Aquatic Sciences (Vol. 2). . Fishery-Aquatic GIS Research Group, Saitama, Japan.
Moore, C. G. 2019. Biological analyses of underwater video from monitoring and research cruises in Lochs Ailort and Fyne, the Sounds of Barra and Mull, inner Moray Firth, off Wester Ross, Noss Head and Rattray Head, and around the Southern Trench in outer Moray Firth
Scottish Natural Heritage Research Report No. 1085.
Moore, C. G., Harries, D. B., Trigg, C., Porter, J. S. & Lyndon, A. R. 2011. The Distribution of Priority Marine Features and MPA search features within the Ullapool Approaches: a broadscale validation survey
Scottish Natural Heritage Commissioned Report No. 422.
Moore, J. 2000. Development of a monitoring programme and methods in Plymouth Sound cSAC: application of diver techniques – 1999. Report to English Nature. Cordah Ltd.
Moore, J., Bunker, F., van Rein, H. & Jones, J. 2015. Methodological trials: Recording subtidal epibiota in situ and in photographs, Portrush August 2013 and Sound of Mull August 2014
JNCC Report No. 561.
Moore, J., Van Rein, H., Sotheran, I., Mercer, T. & Ferguson, M. 2019. Optimisation of Benthic Image Analysis Approaches
JNCC Report No. 641.
Morris-Webb, E. S. & Stamp, T. S. 2015. Biological Analyses of underwater video footage from Arran, Loch Linnhe, Loch Shell and Loch Seaforth
NatureScot Commissioned Report No. 818.
Moss, B. & Lacey, A. 1963. The development of Halidrys siliquosa (L.) Lyngb. New Phytologist, 62.
Mouillot, D., Graham, N. A. J., Villéger, S., Mason, N. W. H. & Bellwood, D. R. 2013. A functional approach reveals community responses to disturbances. Trends in Ecology & Evolution, 28, 167-177.
Noble-James, T., Jesus, A. & McBreen, F. 2017. Monitoring Guidance for Marine Benthic Habitats. JNCC Report No. 598.
Northen, K. O. & Doggett, M. 2020. Condition assessment monitoring for reefs, Plymouth Sound and Estuaries Special Area of Conservation: subtidal reef communities survey July 2017 – 2019. Monitoring report II. Report to Natural England Menai Bridge: Marine Ecological Solutions.
O'Dell, J., Bulgakova, A., Amos, W. & Dewey, S. 2020. Biological analyses of seabed imagery from within and around Loch Alsh, Loch Carron, Wester Ross, Small Isles and South Arran Marine Protected Areas in 2018
Scottish Marine & Freshwater Science, 12(1).
Pardo, C., Guillemin, M.-L., Peña, V., Bárbara, I., Valero, M. & Barreiro, R. 2019. Local coastal configuration rather than latitudinal gradient shape clonal diversity and genetic structure of Phymatolithon calcareum maerl beds in North European Atlantic. Frontiers in Marine Science, 6, 149.
Parry, M. E. V. 2019. Guidance on Assigning Benthic Biotopes using EUNIS or the Marine Habitat Classification of Britain and Ireland
Revised 2019. JNCC Report NO. 546.
Peña, V. & Bárbara, I. 2008. Maërl community in the north-western Iberian Peninsula: a review of floristic studies and long-term changes. Aquatic Conservation: Marine and Freshwater Ecosystems, 18, 339-366.
Peña, V. & Bárbara, I. 2009. Distribution of the Galician maerl beds and their shape classes (Atlantic Iberian Peninsula): proposal of areas in future conservation actions. Cahiers de Biologie Marine, 50, 353-368.
Peña, V. & Bárbara, I. 2010. Seasonal patterns in the maërl community of shallow European Atlantic beds and their use as a baseline for monitoring studies. European Journal of Phycology, 45, 327-342.
Peña, V., Bárbara, I., Grall, J., Maggs, C. A. & Hall-Spencer, J. M. 2014. The diversity of seaweeds on maerl in the NE Atlantic. Marine Biodiversity, 44, 533-551.
Peña, V., Pardo, C., López, L., Carro, B., Hernandez-Kantun, J., Adey, W. H., Bárbara, I., Barreiro, R. & Le Gall, L. 2015. Phymatolithon lusitanicum sp. nov. (Hapalidiales, Rhodophyta): The third most abundant maerl-forming species in the Atlantic Iberian peninsula. Cryptogamie, Algologie, 36, 429-459.
Perrins, J., Bunker, F. & Bishop, G. 1995. A comparison of the maerl beds of the Fal Estuary between 1982 and 1992. Peterborough: English Nature.
REBENT. 2010. Macroalgues Subtidales: Algues calcifiées libres
Rogers, S. I., Ellis, J. R. & Dann, J. 2001. The association between arm damage of the common starfish, Asterias rubens, and fishing intensity determined from aerial observation. Sarsia, 86, 107-112.
Ropert, M. & Dauvin, J.-C. 2000. Renewal and accumulation of a Lanice conchilega (Pallas) population in the baie des Veys, western Bay of Seine. Oceanologica Acta, 23, 529-546.
Simon-Nutbrown, C., Hollingsworth, P. M., Fernandes, T. F., Kamphausen, L., Baxter, J. M. & Burdett, H. L. 2020. Species distribution modelling predicts significant declines in coralline algae populations under projected climate change with implications for conservation policy. Frontiers in Marine Science, 7, 758.
Smith, T. B. & Keegan, B. F. Seasonal torpor in Neopentadactyla mixta (Õstergren). In: Keegan, B. F. & O'Connor, B. D. S., eds. Proceedings of the Fifth International Echinoderm Conference, 1984 Galway. A.A. Balkema, 459.
Southward, A. J. 1995. The importance of long time-series in understanding the variability of natural systems. Helgoländer Meeresuntersuchungen, 49, 329-333.
Steneck, R. S. & Dethier, M. N. 1994. A functional group approach to the structure of algal-dominated communities. Oikos, 69, 476-498.
Strong, J. A. & Johnson, M. 2020. Converting SACFOR data for statistical analysis: validation, demonstration and further possibilities. Marine Biodiversity Records, 13, 2.
Stuart-Smith, R. D., Barrett, N. S., Stevenson, D. G. & Edgar, G. J. 2010. Stability in temperate reef communities over a decadal time scale despite concurrent ocean warming. Global Change Biology, 16, 122-134.
Svane, I. & Lundälv, T. 1981. Reproductive patterns and population dynamics of Ascidia mentula O.F. Müller on the Swedish west coast. Journal of Experimental Marine Biology and Ecology, 50, 163-182.
Turner, J. A., Hitchin, R., Verling, E. & Van Rein, H. 2016. Epibiota remote monitoring from digital imagery: Interpretation guidelines
Underwood, A. J. 1997. Experiments in Ecology, Cambridge: Cambridge University Press.
Wells, G. P. 1951. On the behaviour of Sabella. Proceedings of the Royal Society of London. Series B - Biological Sciences, 138, 278-299.
Wilson, S., Blake, C., Berges, J. A. & Maggs, C. A. 2004. Environmental tolerances of free-living coralline algae (maerl): implications for European marine conservation. Biological Conservation, 120, 279-289.
Wilson, S., P, J. M., Kelly, J., Clarkin, P. E. & Maggs, C. A. 2007. Assessment of extent and abundance of maerl beds and their associated biodiversity along the east Antrim coast. Report prepared by the Natural Heritage Research Partnership (NHRP) between Quercus, Queen’s University Belfast and the Northern Ireland Environment Agency (NIEA). Research and Development Series No. 13/05.







