NatureScot Research Report 1207 - Assessing the implications of wrasse fishing for marine sites and features
Published: 2025
Authors: Burns, N.M., Bailey, D.M. (University of Glasgow), and Hopkins, C.R. (University of Hull)
Cite as: Burns, N.M., Hopkins C.R. and Bailey D.M. 2025. Assessing the implications of wrasse fishing for marine sites and features. NatureScot Research Report 1207.
Contents
- Publication delay statement
- Keywords
- Background
- Main findings
- Acknowledgements
- Abbreviations
- Introduction
- Methods
-
Results
- Relative abundance of wrasse in the South Arran NC MPA
- Model prediction of all wrasse species
- Multivariate analysis of wrasse species relative abundance
- Habitat association
- Interaction with priority and protected features
- Biological comparative tables analysis
- Wrasse probability of presence habitat association analysis
- Consideration of wrasse monitoring methods
- Discussion
- Conclusions
- Annexes
Publication delay statement
This report was commissioned by NatureScot in 2019 (then known as “Scottish Natural Heritage (SNH)” and referred to as such throughout this report) with work by University of Glasgow commencing on 5th August 2019. The completed report was accepted by SNH following review on 19th March 2020. However, due to internal resourcing issues, the publication of this report was delayed until 2025. As such, this report is based on information available at the time of writing – up to March 2020.
Since the completion of this report, there have been some notable updates regarding the Scottish wrasse fishery and its management, noted below for context:
- Following a consultation from March to April 2020 led by the Marine Directorate of the Scottish Government (then known as “Marine Scotland” and referred to as such throughout this report), the voluntary fisheries management measures introduced in May 2018 became mandatory along with additional measures, including:
- an obligation for fishing vessels to accommodate fisheries observers if required (where practically appropriate),
- an obligation to install/use spatial monitoring equipment on fishing vessels if instructed by Marine Directorate,
- species-level reporting of the number of wrasse landed per fishing trip (to ICES statistical rectangle resolution)
- Marine Directorate are in the process of carrying out an assessment of the Scottish wrasse fishery, with advice provided by NatureScot in relation to protected sites and species.
Keywords
wrasse; Labridae; habitat association; salmon aquaculture; South Arran MPA; cleaner fish; PMF
Background
Concerns with chemical treatments for the control of sea lice affecting farmed salmon have prompted the Scottish salmon aquaculture industry to increasingly use cleaner fish - wrasse species (Labridae) and lumpsuckers (Cyclopterus lumpus) - as an alternative. Demand from the industry for cleaner fish has resulted in the development of a fishery using static traps targeting wrasse species. The extent of the demand for wrasse means that the fishery is currently being pursued in Scottish waters, the south west of England, Wales and Northern Ireland. The wrasse fishery in Scottish waters does not currently have any formal management measures. However, a voluntary code of practice was introduced in May 2018.
The wrasse species targeted are relatively slow growing, long-lived and exhibit strong site fidelity and territorial behaviour. This makes them particularly vulnerable to overexploitation. The fishery is data poor and our understanding of its effects are limited. Beyond the immediate impact of removing wrasse and reducing abundances, there is a need to consider the biodiversity implications of the fishery for protected sites and species. For example, to understand how closely linked wrasse are to rocky reef and kelp beds and the implications of the wrasse fishery for otters (Lutra lutra), which can become entangled or trapped in fishing gear of the type used in this fishery. Developing an understanding of the wrasse fishery’s interactions with the protected features of the Scottish Marine Protected Area (MPA) network and relevant Priority Marine Features (PMFs) will enable appropriate advice on this data poor fishery to be developed. This report develops a rationale to cover the potential inclusion of wrasse species in the monitoring of the Scottish MPA network and will enable Scottish Natural Heritage (SNH) to assess feasibility and develop an appropriate methodology.
In this study, a combination of methods is used to:
- explore wrasse abundance in a typical site on the Scottish West Coast,
- understand the associations of wrasse species with particular habitat types and,
- suggest the most appropriate methods to monitor changes in wrasse species abundances.
Archived baited camera survey data collected in the South Arran Nature Conservation Marine Protected Area (NC MPA) was analysed to provide relative abundance estimates of wrasse species within the NC MPA. This data was used to explore changes in wrasse abundance through time and understand the effects of using different sampling strategies. Using this data, statistical models were also built to explain the associations of wrasse species with particular habitat types. The present study also set out to review existing information in the scientific literature and compare species compositions with the physical characteristics of habitats from JNCC biological comparative tables to document habitat associations of wrasse species. Following this analysis, this study provides advice on developing appropriate monitoring methodology for wrasse species.
Main findings
- The assessment of habitat association indicated very strong associations between all wrasse species and shallow water, kelp covered, rocky reefs. Given the nature and strength of the association between these fish and this habitat type, wrasse can be considered typical species of rocky reef and kelp beds. Rocky reefs are a Habitats Directive Annex I habitat and kelp beds are a PMF.
- After reviewing all potential available data sources, no current programme of fish monitoring captures information about wrasse species in the habitats and localities where they are found. Fisheries monitoring programmes, although reporting occasional wrasse catches, take place in water too deep to properly monitor any of the UK wrasse species. Other surveys, like those previously conducted to monitor demersal fish species in transitional waters or Scottish MPA network monitoring programmes, also miss key wrasse habitat. Current monitoring programmes may indicate presence/ absence of wrasse but do not properly monitor wrasse populations and recruitment from one year to the next, and do not provide insight into the effects of the removal of wrasse on the wider ecosystem. Therefore, effective monitoring of wrasse requires development and use of specific methodology.
- To properly manage wrasse as a commercially fished species a more detailed understanding is required of their population structure and effects of the removal of wrasse on the wider ecosystem.
- Analysis of wrasse relative abundance in the South Arran NC MPA provided a case study to highlight important considerations for any monitoring programme. The spatial distribution and placement of individual sample points is critical to effectively monitor changes in wrasse abundance through time. In particular, given the close association of all wrasse species with rocky reef and kelp bed habitats, sampling and analysis must target these specific habitat types.
- Understanding the effects of wrasse territoriality and protogyny on abundance estimates and population dynamics is also essential.
- The analysis of relative abundance in the South Arran NC MPA also indicated that, to be effective, programmes designed to monitor changes in wrasse abundance over time must consider individual species rather than grouping all wrasse species in one taxon.
Acknowledgements
The authors thank the contribution of Natasha Walker-Milne and Dr Sophie Elliott, the Community of Arran Seabed Trust (COAST) and John Clarke at Marine Scotland Science (MSS) for providing the data used in the analysis in this report.
Abbreviations
Akaike Information Criterion (AIC)
Algae/boulder/cobble (ABC)
Algae/gravel/pebble (AGP)
Circalittoral habitat (CR)
Common Fisheries Policy (CFP)
Community of Arran Seabed Trust (COAST)
Confidence Interval (CI)
Convention on Biological Diversity (CBD)
Essential Fish Habitat (EFH)
European Nature Information System (EUNIS)
Generalised Additive Model (GAM)
Good Environmental Status (GES)
Gross tonnage (gt)
Infralittoral habitat (IR)
Inshore Fisheries and Conservation Authority (IFCA)
International Bottom Trawl Survey (IBTS)
International Council for the Exploration of the Sea (ICES)
JNCC (Joint Nature Conservation Committee)
Marine Protected Area (MPA)
Marine Scotland Science (MSS)
Nature Conservation Marine Protected Area (NC MPA)
Oslo Paris Convention (OSPAR Convention)
Priority Marine Feature (PMF)
Remotely Operated Vehicle (ROV)
Rigid Inflatable Boat (RIB)
Scottish Environmental Protection Agency (SEPA)
Scottish Natural Heritage (SNH)
Scottish Salmon Producers’ Organisation (SSPO)
Scottish West Coast Ground Fish Survey (SCOWCGFS)
Self-Contained Underwater Breathing Apparatus (SCUBA)
Special Area of Conservation (SAC)
Stereo Baited Remote Underwater Video (SBRUV)
Water Framework Directive (WFD)
Water Strategy Framework Directive (MSFD)
Introduction
Use of cleaner fish in the aquaculture industry
Sea lice are an increasingly important problem economically and environmentally in marine farmed salmon (Salmo salar) (Torrissen et al. 2013, Hamre et al. 2013, Murray 2017). Sea lice management on farmed Atlantic salmon quickly became a major problem soon after the first Norwegian fish farms were established (Heuch et al. 2005, Oppedal et al. 2011). Sea lice can negatively impact the commercial profitability of the industry by reducing the value of damaged fish and through declines in the health and welfare of the farmed fish (Jones et al. 2015). Sea lice is the common term adopted for two species of salmon ectoparasites (Lepeophtheirus salmonis and Caligus elongates). L. salmonis is a parasitic copepod which specifically affects salmonids (Heuch et al. 2005, Jones and Johnson 2015). C. elongates affects a much wider range of species of salmonids and non-salmonids. Sea lice infections in Atlantic salmon farms begin with C. elongates, with L. salmonis infestations occurring later (Todd 2007, Treasurer 2013, Leclercq et al. 2014a).
Traditional control methods using medications have become increasingly expensive as lice develop resistance to commonly used treatments (Hamre et al. 2013). Increased lice resistance to chemical treatments requires more frequent delousing and an associated increase in treatment costs. There have also been wider concerns over the environmental impacts of the chemical treatments on non-target wild species near salmon farms (Hamre et al. 2013), while elevated levels of lice around farms can increase parasitic burdens on wild fish (Hamre et al. 2013). These concerns are receiving greater attention from environmental monitoring agencies (e.g. Scottish Environmental Protection Agency (SEPA)), (SEPA 2019).
Solutions to these problems have been sought by using cleaner fish to remove lice from farmed salmon with some reported success (Rae 2002, Leclercq et al. 2014a). Species used to de-louse salmon include lumpfish (Cyclopterus lumpus) (Imsland et al. 2014) and several species of wrasse including ballan (Labrus bergylta) and goldsinny (Ctenolabrus rupestris), (Sesvik et al., 2013; Leclercq, Davie and Migaud, 2014b). Several other species are also used (Treasurer 2002). The UK and European aquaculture industry uses some farmed ballan wrasse and lumpfish (Brooker et al. 2018), but also relies heavily on wild caught ballan, goldsinny, corkwing (Symphodus melops), rock cook (Centolabrus exoletus) and cuckoo wrasse (Labrus mixtus), (Norwegian Directorate of Fisheries., 2013; Treasurer, 2013; Skiftesvik et al., 2014).
Wild caught wrasse were first used in Norway as a commercial treatment for lice in 1988 and around the UK coast between 1989 and 1990 (Bjordal 1988, Darwall et al. 1992a, Piccinetti et al. 2017). While medications have previously been relied on to treat lice infestations, increased lice resistance to chemicals has led to greater industry interest in the use of alternative measures. These include physical prevention and, mechanical and biological control. The effectiveness of biological control using cleaner fish has resulted in increased capture of wild wrasse. The increased exploitation of wild wrasse has raised concerns over depletion of these stocks (Ottesen et al. 2012). Wrasse have been used as cleaner fish in Norway (Bjordal 1991), Ireland (Darwall et al. 1992a) and Scotland (Treasurer 1993). Ballan, corkwing, goldsinny and rock cook wrasse are now extensively harvested as cleaner fish for aquaculture around the coasts of northern Europe (Skiftesvik et al. 2014). An approximate 3.5 million wild wrasse (mostly goldsinny) were caught in Norway in 1997 (Gjøsæter 2002a).
The commercial production of wrasse species in aquaculture is still developing and therefore there is a need for continued use of wild-caught wrasse (Piccinetti et al. 2017, Scottish Salmon Producers’ Organisation (SSPO) 2018). Of the wrasse species used by salmon farms, only corkwing and ballan wrasse reach a suitable size by age two to be used for delousing salmon in net pens (Skiftesvik et al. 2013). Ballan wrasse is the larger of these two species and has received the most attention, being the first wrasse species to be developed for intensive culture (Skiftesvik et al. 1996, Stone 1996, D’Arcy et al. 2012). The commercial production of juvenile ballan wrasse has been developed from Atlantic cod (Gadus morhua) farming enterprises but production does not meet the demand from salmon farms (Hamre et al. 2013, Piccinetti et al. 2017). The first farmed ballan wrasse were deployed for use in salmon pens in 2013 in Scotland, (Treasurer 2018). Prior to 2011, all cleaner fish used in salmon cages were sourced from the wild. In 2016, 46% of cleaner fish used in Norway were commercially produced. Of these 3.5% were ballan wrasse, the rest being lumpfish (Brooker et al. 2018). The cultivation of cleaner fish in the UK and elsewhere is the subject of ongoing development. However, at the time of writing, it is reasonable to assume that there will be continued demand for wrasse from the fishery for the foreseeable future.
The wrasse fishery
Of the six wrasse species found in the North Atlantic - ballan (Labrus bergylta), goldsinny (Ctenolabrus rupestris), corkwing (Symphodus melops), cuckoo (Labrus mixtus), rock cook (Centrolabrus exoletus) and scale-rayed (Acantholabrus palloni) - most live inshore on rocky reef habitat. Traditionally, these wrasse species have not been of any economic interest (Darwall et al. 1992b), but have been targeted recreationally by anglers. Extensive wild capture of ballan, corkwing, goldsinny and rock cook wrasse is now common in European and UK waters (Kvenseth 1996, Skiftesvik et al. 2014, 2015). While the fisheries targeting these species often treat them as a single group (i.e. “wrasse”), the different species adopt a variety of life histories which will require species-specific management and monitoring strategies.
Concerns of overharvesting of wild caught wrasse and overfishing of particular wrasse stocks have been highlighted in Ireland (e.g. Darwall et al. 1992b, Tully et al. 1996), with a reduction in wrasse numbers in small bays highlighted as a potential problem. The scientific community has raised concerns over high extraction rates in Norway (Skiftesvik et al. 2014, Halvorsen et al. 2017a).
During the winter months, the lice eating efficacy of wrasse is minimal as they become inactive at low temperatures (Deady et al. 1995, Treasurer 2002, 2013, Imsland et al. 2016). To supply cleaner fish demand, the wrasse fishing season (during the summer months) coincides with the spawning season for some wrasse species (e.g. goldsinny, rock cook, corkwing and ballan), which raises more concerns over the long-term sustainability of the fishery targeting these species (Skiftesvik et al. 2015). In Ireland, wild caught wrasse can be fished from June to September (Bolton-Warberg 2018). In Scotland the fishery is open from 1st May to 30th November (Marine Scotland 2018).
High mortality of cleaner fish in salmon farms results in very few cleaner fish surviving through a full salmon production cycle (The European Union Reference Laboratory 2016), meaning that cleaner fish stocks must be annually replaced with wild-caught fish. However, while farming cleaner fish is in its infancy, the sector is aiming to expand towards self-sufficiency using farmed fish (including ballan wrasse) in most countries (Brooker et al. 2018). Figures from Norway indicate that 46% of the 37.2 million cleaner fish (15.9 million lumpfish; 1.3 million ballan wrasse) deployed in 2016 were of farmed origin (Blanco Gonzalez and de Boer 2017). In the UK in 2016, 68% of the total cleaner fish deployed were farmed - 1.9 million lumpfish and 118 000 ballan wrasse (Munro and Wallace 2017).
Most UK salmon farms are located in Scotland and demand for wrasse is highest around the Scottish coast. However, local supply often does not meet demand. This lack of supply has resulted in an estimated one million wrasse being fished annually in English waters and transported to Scotland (Devon & Severn IFCA 2017, Riley et al. 2017).
Potential interactions with protected habitats and species
Wrasse species are recognised as an important component of coastal rocky reefs in temperate waters and form particular associations with algae covered reef (Quignard and Pras 1986, García-Rubies and Macpherson 1995). While wrasse species assemblages show seasonal variation in abundance, they are a key component of different rocky reef habitats throughout the year, making up a high proportion of resident reef species (Magill and Sayer 2002). For example, corkwing has been closely associated with small gaps in rocky reefs which it uses as refugia and feeds mostly on bedrock habitat (Rodrigues et al. 2015). Despite these close associations between wrasse and their habitat, only a few studies have aimed to understand these links (e.g. Treasurer, 1994; Tuya, Wernberg and Thomsen, 2009).
Wrasse likely play an important role in coastal ecosystems. They predate on planktivorous and benthic herbivores and are a food source for piscivorous predators (Moksnes et al. 2008, Baden et al. 2010). In several species of wrasse, habitat associations are strongly influenced by prey diversity living on and among the algae covered rocks (Bell and Harmelin-Vivien 1983, Hilldén 1984, Ruitton et al. 2000). The association between wrasse and the biological components of their habitat in some species like corkwing is particularly strong given their dependence on algae for nest building (Potts 1974a, Rodrigues et al. 2015). Wrasse likely play an important role in maintaining a balance in rocky and biogenic reef systems. For example, sea urchin species (Echinus esculentus and Psammechinus miliaris) feed on epifauna and damage serpulid reef worms (Poloczanska et al. 2004). Wrasse species feeding on small urchins will reduce the impact on both reef-forming worms and the algal species growing on the reef.
The European Habitats Directive (92/43/EEC) identifies the conservation status of natural habitats as: “…the sum of the influences acting on a natural habitat and its typical species that may affect its long-term natural distribution, structure and functions as well as the long-term survival of its typical species…”. To ensure this long-term survival, under Article 6 governments are required to employ conservation management like, “appropriate statutory, administrative or contractual measures” corresponding to the ecological requirements of the habitat. The Marine (Scotland) Act 2010 also requires action to manage species occupying specific habitat types. Under Section 82 (a) of the Marine (Scotland) Act 2010, the Act imposes an obligation on public authorities to exercise their functions “in the manner which it considers best furthers (i) the stated conservation objectives for the Nature Conservation Marine Protected Area (NC MPA)”. Under the Designation order for each NC MPA, protected features must either remain in a “favourable condition” or be “brought into such condition”. Favourable condition with respect to a marine habitat means “(a) its extent is stable or increasing; and (b) its structures and functions, its quality, and the composition of its characteristic biological communities are such as to ensure that it is in a condition which is healthy and not deteriorating” (e.g. The Scottish Government, 2014). Understanding the close links between wrasse and their habitat is particularly relevant to inform appropriate monitoring strategies and develop management and conservation measures which rely on spatial protection (e.g. MPAs) or ecosystem-based fishery management (Fitzpatrick et al. 2012, Greenstreet et al. 2012, Johnson et al. 2013).
Reefs are a habitat for which Special Areas of Conservation (SACs) are designated and kelp beds (of varying types) are a Scottish Priority Marine Feature (PMF). Kelp and seaweed communities on sublittoral sediment are also a designated feature in NC MPAs. SACs and NC MPAs are hereafter collectively referred to as the Scottish MPA Network. However, the description of the typical species and communities which occupy rocky reef and kelp bed habitats is vague. There are strong associations between wrasse and the kelp covered reefs found in Scottish waters, yet no detailed definition of a “typical” species exists. We suggest a potentially useful definition of a typical species would be: “a species which has a very high probability of being observed on a particular habitat type and a low probability of being observed in other habitats.”
It is also well understood that habitat and, specifically the characteristics of the substratum, govern the distribution of demersal and benthic fish (Anderson and Millar 2004, Stål et al. 2007, Damalas et al. 2010, Burns et al. 2019). The close link between specific habitats and the species of fish which occupy them has allowed the identification of ‘‘Essential Fish Habitats’’ (EFH). EFH are defined as waters and substrata which are necessary for fish spawning, feeding and growth (Benaka 1999). The link between fish and the habitats required for their survival has been identified as particularly important for management and conservation (Stål et al. 2007).
With respect to protected species, otters (Lutra lutra) are the most likely protected vertebrate to have an association with the wrasse fishery. Otters are an Annex IV and Annex II species under the Habitats Directive and therefore are species in need of strict protection and a species for which SACs can be designated. Otters are also a PMF. A large portion of the otter population in Scotland is associated with the coast and makes use of shallow marine habitats for foraging (Clavero et al. 2006, Findlay et al. 2015). Coastal otters forage in the intertidal and shallow subtidal zone, in depths of approximately 0 to 10 m (Heggberget 1993). Coastal otter diet can be comprised of marine species including crustaceans, cephalopods and small inshore fish. There is potential for interactions between the wrasse fishery and otters as both are likely to occur in the same rocky reef and algal habitat. The risk of otter mortality owing to entanglement in fish and crustacean traps was highlighted as a concern by the UK Marine SACs Project report in 2001 (UK Marine SACs Project, 2001).
Context for monitoring of the wrasse fishery
Marine fish in the seas around Scotland are currently sampled routinely in scientific surveys. A database of existing surveys is available from the UK Directory of Marine Observing Systems. The surveys around the Scottish coast are primarily for fisheries stock assessment conducted by Marine Scotland Science (MSS) funded by the Scottish Government. Limited survey effort is dedicated to assessing fish as indicators of marine habitat quality. The MSS surveys are undertaken as part of the Scottish West Coast Ground Fish Survey (SCOWCGFS).
The wrasse fishery in the UK is poorly documented and there is currently no fishery-independent monitoring of the fishery. UK landed wrasse are often identified under a generic wrasse code, resulting in a lack of species specific data (Faust et al., 2018). This lack of species-specific data makes assessing the effect of the wrasse fishery difficult. The wrasse fishery in Scottish waters has grown without formal management measures and a code of practice has only recently been established. Voluntary control measures for the Scottish fishery were introduced in May 2018. These measures include species-specific minimum landing sizes and reporting “records of numbers, linked to total weights of wrasse caught” on FISH1 forms (Marine Scotland, 2018). There are questions, however, about the use of FISH1 forms as a declaration of “landings” in this context, as it is unclear whether this includes transhipment to aquaculture facilities at sea. If fish being delivered direct to fish farms are not included as “landings”, this loophole should be closed. Further consultation is now underway (March to April 2020), led by Marine Scotland, with the intention of providing a mechanism for statutory underpinning of the code.
As a relatively newly harvested group of species, current scientific surveys were not designed to monitor wrasse species stock abundance. MSS bottom trawl surveys are conducted quarterly (primarily in Q1 and Q4), and are depth stratified in waters 20 to 400 m deep. One 30-minute trawl is conducted per ICES rectangle. In practice these trawls are seldom close to shore and trawl at least one nautical mile from shore. Survey sites are randomly selected from a number of known sites which are safe to trawl, meaning that the same area is often repeated from one year and quarter to the next. Importantly for wrasse species these trawls avoid rocky habitat to avoid damage to fishing gear. MSS do not routinely carry out monitoring of fish in shallow water (<30 m depth).
Several short studies to monitor fish at shallow depths have been conducted at Stonehaven in Aberdeenshire and in the Firth of Clyde. These studies have included comprehensive reviews (e.g. Bailey et al., 2011; Ryan and Bailey, 2012), often been part of undergraduate, Masters and PhD student projects (e.g. Elliott et al., 2016; Elliott et al., 2017b) and have employed a range of methods. SEPA have previously conducted surveys for demersal fish species in transitional waters (as defined by the Water Framework Directive (WFD)). These surveys were conducted using beam trawls, beach seines and fyke nets at three-yearly intervals. Although this monitoring programme is not currently operating, the approach required to sample wrasse species in shallow water on rocky reefs is very similar.
The legal obligations for monitoring marine species and habitats are relevant for the monitoring of wrasse species (Table 1). The existing surveys for fish monitoring use different methods and timescales making it hard to combine datasets across agencies. For example, the three-year sampling cycle used by SEPA greatly limits its usefulness to monitor wrasse species and inform on recruitment. The IBTS surveys do have some records of wrasse, however, this survey mostly conducts sampling too far offshore for the data to monitor wrasse populations. Consequently, there is no current monitoring activity that would provide for the requirements of site condition monitoring in the Scottish MPA network.
| Directive or treaty | Relevant law | Legal objectives | Timescales | Criteria and relevance to wrasse |
|---|---|---|---|---|
| Habitats Directive (92/43/EEC) | Conservation (Natural Habitats, &c.) Regulations 1994 (as amended) | Implementation of marine Special Areas of Conservation (SACs) contributing to the Natura 2000 network of protected areas. | In operation |
|
| Water Framework Directive (WFD) | Water Environment and Water Services (Scotland) Act 2003 | Good Ecological Status | To be achieved by 2027 | Fish in transitional waters within territorial waters (3 nautical miles of baseline). Reporting every 6 years |
| Marine Strategy Framework Directive (MSFD) | UK Marine Strategy Regulations 2010 | Good Environmental Status (GES) | To be achieved by 2020 | Positive trends in non-commercial species |
| Oslo Paris Convention (OSPAR) | Marine (Scotland) Act 2010 | Establish a network of marine protected areas which is both ecologically coherent and well-managed | To be achieved by 2016 |
|
| Convention on Biological Diversity (CBD) | Wildlife and Countryside act 1981 (as amended). Nature Conservation (Scotland) Act 2004 |
| To be achieved by 2020 | Coherent network of MPAs, biodiversity to be taken into account in inshore fisheries, improve monitoring |
Aims of a wrasse monitoring programme
The aims of a wrasse monitoring programme would be to:
- Inform sustainable catch limits for the fishery with regard to the legal obligations (outlined in Table 1) for sustainable fishing.
- Collect data on the status of wrasse populations in relevant protected sites with regard to meeting conservation objectives (Table 1).
To effectively monitor wrasse species in the context of the fishery and biodiversity monitoring obligations/needs, the abundance of individual wrasse species would need to be monitored across a representative range of sites consisting of routine sampling stations in Scottish coastal waters in depths of < 25 m.
- Over time these data will provide an index of relative abundance or density (depending on the approach adopted) that could be used to inform management decisions on catch or effort levels.
- In areas lacking appropriate baselines these data could allow Conservation Status (Habitats Directive) and Good Environmental Status (GES) (Marine Strategy Framework Directive, MSFD) to be assessed on a “trends based” approach (as advocated for individual species by Greenstreet et al., (2012).
- In Scottish MPA network locations these data could be used as an indicator of Site Condition (as a trend-based metric).
While a stock assessment approach which could lead to the calculation of catch limits would be ideal, there are some practical challenges to this approach. Any baited approaches (cameras, traps, lines) make estimation of the area sampled extremely difficult. Trawls and sleds will work in some but not all areas of interest and would have different amounts of environmental impact. Remotely Operated Vehicles (ROVs) and divers provide the only methods which can collect quantitative information over the ground of interest; noting that the use of lasers affects the behaviour of wrasse in a way which invalidates normal measures of density but could allow other approaches. The significant costs of SCUBA or ROV surveys over the number of samples necessary is also an issue.
Importantly the accuracy of estimating absolute abundance from measures of relative abundance is often unclear. Quantitative measures derived from sampling techniques like trawling, or SCUBA diver surveys can produce estimates of density (i.e. numbers of wrasse per m2). Baited sampling methods like Stereo Baited Remote Underwater Video (SBRUV) deployments do not allow the number of individuals per unit area to be calculated and rely on relative measures of abundance like maxN (the maximum number of individuals observed in the camera field of view in any one frame). However, both measures provide patterns of population trends which are proportional to the absolute abundance of the population. In addition, any stock assessment must be carried out at the appropriate spatial scale for the populations of which the stock is composed. The current uncertainty over the population structure of wrasse species in Scotland means that it would be difficult to identify different stocks and make estimates of spawning stock biomasses.
Methods
Wrasse abundance methodology
Study location
Located in the Firth of Clyde on the West coast of Scotland, the South Arran NC MPA was designated in 2014 and covers an area of 280 km2 (Figure 1). The designation of this area was intended to protect internationally important seagrass and maerl beds and other marine features like kelp and seaweed communities (SNH 2017). Four levels of protection are in place within the South Arran NC MPA, including the most protection afforded by any MPA in the Scottish network, the Lamlash Bay No Take Zone (designated in 2008) which covers 2.66 km2.
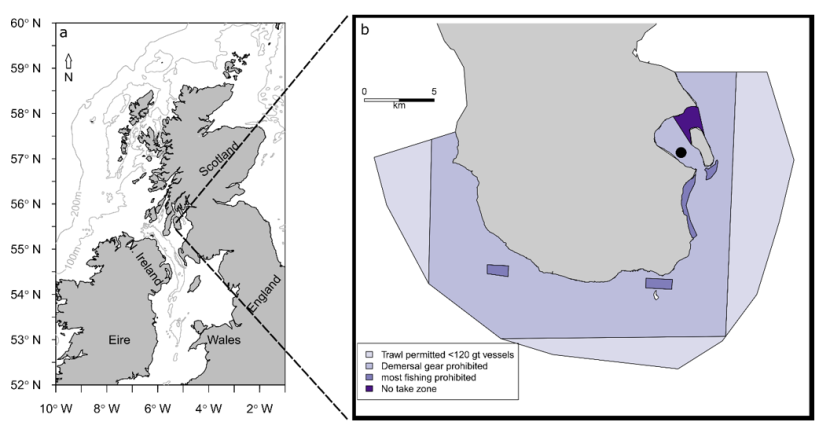
Figure 1. South Arran case study site used in the analysis of wrasse abundance.
Click for a full description
The South Arran NC MPA is located in the Firth of Clyde on the Scottish west coast (a). Four levels of management measures are in place in the NC MPA depicted in purple (from light to dark: trawling by vessels <120 gross tonnage permitted; mobile demersal gear prohibited; most fishing prohibited; no take zone) (b). The filled black circle shows the location of the salmon farm operated in the study area.
Biological data collection
Sampling was conducted with three SBRUV camera platforms described in full in Elliott et al. (2017a). Surveys were conducted in 2013, 2014, 2017 and 2018 (Figure 2). Data was collected in 2013 and 2014 by S. Elliott. Data collection in 2017 was carried out by MSS and data from 2018 was supplied by N. Walker-Milne. Surveys were also conducted in 2016 by the University of York. However, owing to a requirement for a different survey design where deployments were clustered (Annex 1, Figure A1), habitat occupied by wrasse was not sampled and consequently no wrasse were recorded in the 2016 surveys. No survey work was conducted in 2015.
SBRUV camera equipment used in the surveys consisted of a pair of Canon HF G25 video cameras encased in waterproof camera housings, mounted on a steel frame. Cameras were mounted at an angle of 15° to the seabed and facing in by 8° which provided coverage of the substratum and sufficient image overlap to allow length measurements to be made. Frames were freshly baited with locally caught mackerel for each station and deployments were carried out for 60 minutes, with recording time starting from when the frame landed on the seafloor. To avoid interference all deployments made in one day were positioned more than 500 m apart. Depths for the SBRUV deployments ranged from 5.8 m to 45.9 m. When recording results, an allowance of 3 minutes was made for settlement once the frame had reached the seabed to give fish present time to acclimatise and for resuspended sediments to settle. Videos were analysed using Event Measure™ software (licensed by SeaGIS 2013). Relative abundance was documented as the maximum number of individuals of each wrasse species to appear in the camera field of view at the same time (commonly referred to as maxN) (Merrett et al. 1994, Watson et al. 2005).
No formal data are available about wrasse fishing activity around Arran or in the MPA. However, it is thought that fishing occurred in this area from 2014 until 2018 when all salmon from the local farm (Figure 1) were harvested. Fishing for wrasse in this area has been reported to have been conducted by one fisher using fish traps.
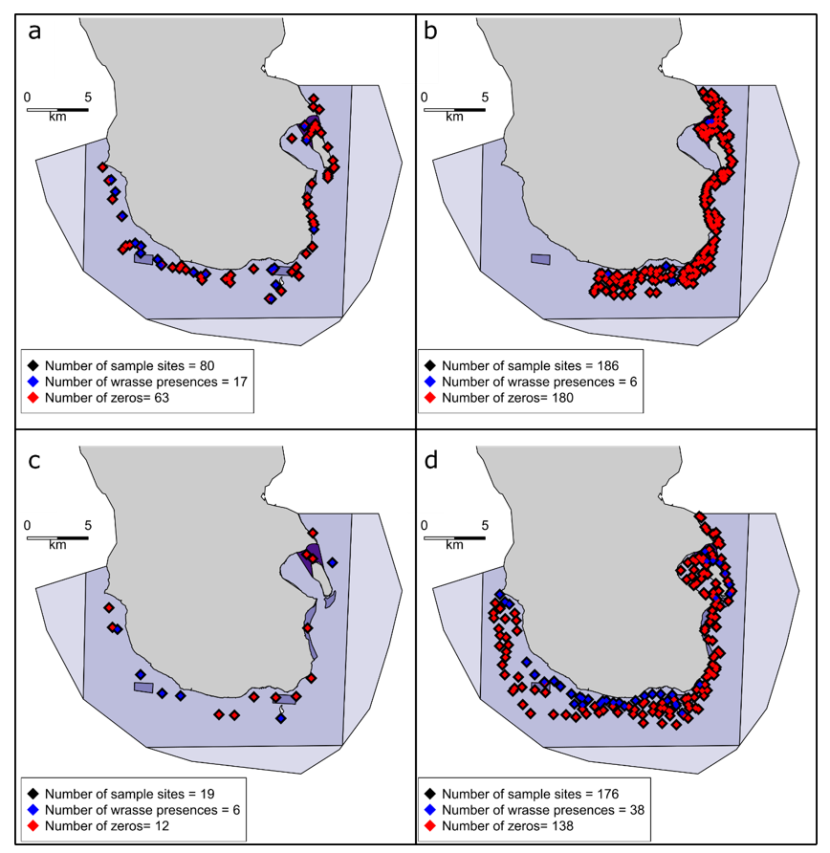
Figure 2. Stereo Baited Remote Underwater Video (SBRUV) deployment locations in the South Arran NC MPA in 2013 (a), 2014 (b), 2017 (c) and 2018 (d).
Click for a full description
Map showing SBRUV deployment locations within the South Arran Nature Conservation Marine Protected Area. The total number of deployments are given for each year: 80 in 2013, 186 in 2014, 19 in 2017, 176 in 2018. Deployments where wrasse were observed are coloured blue and those where no wrasse were observed are coloured red. The deployment sites for 2016 are shown in Annex 1, Figure A1. Four levels of management measures are in place in the NC MPA depicted in purple (from light to dark: trawling by vessels <120 gross tonnage permitted; mobile demersal gear prohibited; most fishing prohibited; no take zone).
Statistical analysis
A two-step statistical approach was adopted to analyse the abundance of wrasse in the South Arran NC MPA. All statistical analyses were carried out in R v 3.5.2 (R Core Team 2019). Initially the relative abundance of all species of wrasse were considered as one taxon (1). The second modelling step explored the relative abundance of each of the wrasse species using a multivariate approach (2).
- The relative abundance of all wrasse species was modelled as a function of depth, wave fetch, substratum type, the spatial variables latitude and longitude, and year to explore temporal variability. Generalised Additive Models (GAMs) were used to account for non-straight, linear relationships between wrasse abundance and the explanatory variables (Wood and Scheipl 2014). Model selection was conducted using a backwards stepwise model selection process with the Akaike Information Criterion (AIC) to select the optimal model.
- The relative abundance of each wrasse species was modelled in a generalised multivariate model using the mvabund package in R (Wang et al. 2013). Model selection and the variables tested were the same as described in (1).
Step one in the modelling approach was used to establish the general temporal, spatial and habitat trends which governed wrasse abundance. The second step was used to explore specific changes in abundance for each species between 2013 and 2018 in the South Arran NC MPA.
Habitat association methodology
Linking wrasse species with their essential habitat was carried out in three stages:
- A structured literature review
- Analysis of the Biological comparative tables (JNCC)
- Analysis of archived baited camera survey data (University of Glasgow 2013-2018)
Structured literature review of habitat association for wrasse
The review focused on five priority wrasse species - goldsinny (Ctenolabrus rupestris), cuckoo (Labrus mixtus),ballan (Labrus bergylta), corkwing (Symphodus melops, formerly Crenilabrus melops) and rock cook (Centrolabrus exoletus). The primary focus of the review was on habitat associations with the wrasse species listed above. The literature review followed the principles of a systematic review. Broad topic terms (Table 2, section 3.2.1) were used to conduct a comprehensive literature search of online databases of peer-reviewed articles. The search was carried out on the 21st August 2019 using Web of Science through the University of Glasgow portal. All years (1864 - 2019) were selected for the search which included all available databases. The search process was conducted systematically using broad topic terms in the first two searches. This was then refined with the inclusion of geographic terms and finally by searching for the species of interest by name (binomial nomenclature).
Each article and review was examined for reference to associations between habitat and any of the five wrasse species of interest. References were divided into two types: (1) those which were primary literature directly investigating habitat associations in wrasse, and (2) those which referenced wrasse species associations with specific habitat types. The links between specific habitat types and individual wrasse species were recorded and tabulated. Additional refined searches of geographic location and species were conducted. Each article found was examined for reference to associations between habitat and any of the wrasse species of interest.
Biological comparative tables analysis
The JNCC biological comparative tables were used to identify habitats and biotopes with which wrasse species are associated and depend upon. The percentage of core biotope records (above 20%) was extracted at EUNIS level 4 which best represents a biological community level (Elliott et al. 2018). By accounting for both abiotic and biotic variables in their classification, level 4 minimises environmental variation and allows for a better assessment of community associations within the broad habitat type (Davies et al. 2004).
Methods to predict the presence of wrasse given substratum type
The biological data for the following analysis were collected as described above in section 2.1. The statistical analysis was conducted as described in section 2.1.3 with the exception that binomial GAMs were used to model the probability of presence or absence of wrasse on specific substrata. Using a binomial distribution allowed the binary presence/ absence data to be modelled. Model selection considered year, latitude and longitude, fetch and depth and substratum type.
Monitoring
The recommendations in this report for a monitoring programme designed to effectively monitor wrasse species are based on an evaluation of current suitable methodologies. Three monitoring methods were selected for evaluation:
- SCUBA diver transects using stereo video camera.
- Stereo baited remote underwater video (SBRUV) camera deployments.
- Fyke netting and fish trapping
The advantages and disadvantages of each method, relative costs, and sample effort required were analysed. The above monitoring methods were selected using the expert opinion of the authors. Other monitoring options are available, but they have not been evaluated for this report. This is based on extensive experience in the study of fish distribution and behaviour, using a wide range of methods including: baited and unbaited stereo video methods, fixed photographic observatories, underwater vehicles, acoustic tracking, SCUBA diving and trawling. A list of these various techniques for monitoring was refined to the three chosen monitoring methods above following discussions around the suitability of different methods to: (1) monitor wrasse considering the life history and habitat requirements of wrasse species, and (2) achieve the aims of a wrasse monitoring programme (outlined in section 1.4.1).
Results
Relative abundance of wrasse in the South Arran NC MPA
During the years SBRUV studies have taken place (2013, 2014, 2016, 2017 and 2018), wrasse species have been encountered in all surveys apart from those in 2016. In 2016, SBRUV platforms were deployed in clusters to satisfy specific sampling aims and did not sample suitable wrasse habitat and were removed from the following analysis. Five species of wrasse were recorded occurring in the South Arran NC MPA study site. Ballan (Labrus bergylta), goldsinny (Ctenolabrus rupestris) and cuckoo wrasse (Labrus mixtus) were observed in every year considered in the following analysis (i.e. 2013, 2014, 2017 & 2018). Corkwing (Symphodus melops) were observed in 2013 and 2014, and rock cook wrasse (Ctenolabrus exoletus) were recorded in 2014 and 2018.
Model prediction of all wrasse species
The optimal Generalised Additive Model (GAM) predicting the relative abundance of all five wrasse species considered as one taxon (ballan, goldsinny, cuckoo, corkwing and rock cook) explained 55.7% of the deviance. Of the terms retained during model selection, substratum type was the most influential on wrasse relative abundance. Substratum consisted of five types of seabed, “Algae/boulder/cobble”, “algae/gravel/pebble”, “mud”, “sand” and “seagrass”.
Predicted relative abundance of all wrasse species in 2013, 2014, 2017 and 2018 showed a similar distribution across years (Figure 3). The area of coverage of sampling in 2014 (Figure 3b) was not evenly spatially distributed across the whole of the South Arran NC MPA. Consequently, the main areas of suitable wrasse habitat were not included in these surveys and the predicted relative abundance, although similarly distributed to other years, was lower.
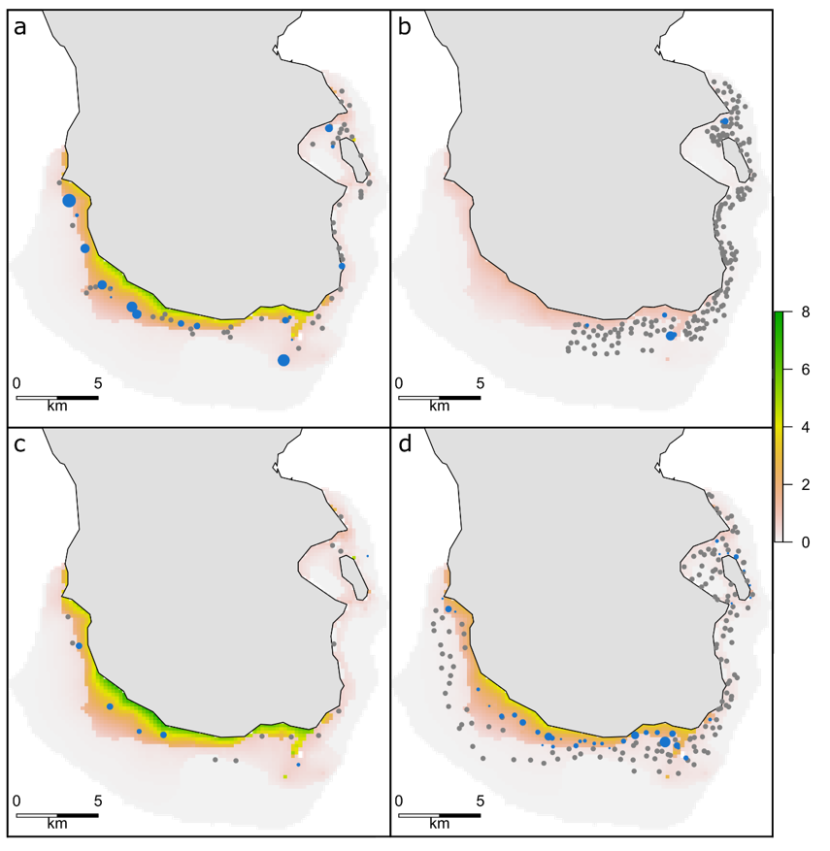
Figure 3. Predicted wrasse relative abundance from GAMs using SBRUV observations in 2013 (a), 2014 (b), 2017 (c) and 2018 (d).
Click for a full description
Maps show wrasse relative abundance, measured as predicted maxN, for each year. All years are scaled to the maximum maxN (7), with higher values in green and lower values in pink. Grey points show camera drops no wrasse were observed. Blue points show camera drops where wrasse (of any species) were observed and point size indicates the observed maxN.
The largest relative abundances of wrasse, measured as maxN, were observed in 2013 (Figure 3a). However, these observations were also accompanied by several SBRUV deployments which recorded zero wrasse. The fewer number of samples in 2017 were closer to shore and proportionally more often on rocky reef than those in 2013 or 2018. Wrasse were recorded more frequently in 2017, although often in fewer numbers resulting in higher predicted maxN (Figure 3c). Wrasse were consistently observed in SBRUV deployments and predicted by the GAMs to the south and west of Arran. The overall predicted relative abundance is displayed in Figure 4.
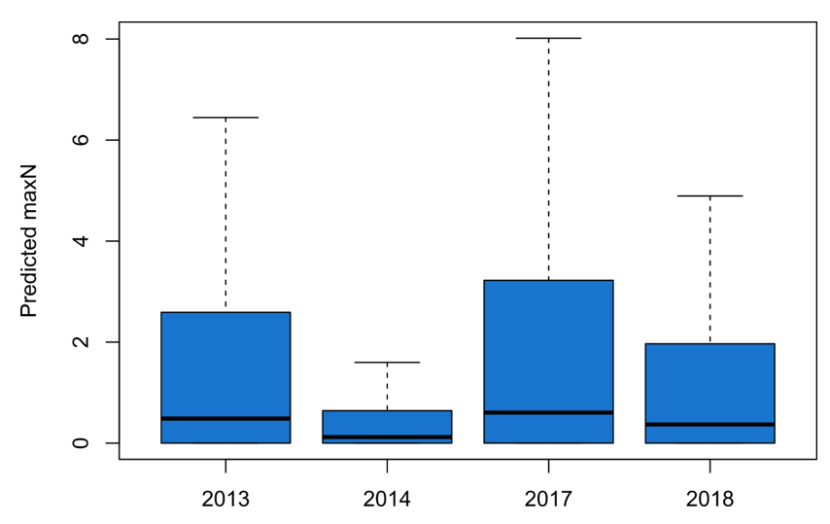
Figure 4. Predicted maxN across all wrasse species from GAMs using SBRUV observations.
Click for a full description
Boxplots of predicted maxN across all wrasse species. Thick horizontal lines represent the median in each survey year. Whiskers represent the highest predictions (highest quartile) across all deployments each year. Predicted maxN was highest and most variable in 2017, followed by 2013.
Multivariate analysis of wrasse species relative abundance
The multivariate analysis of wrasse relative abundance explored changes in abundance of individual species during the study period. Given the close association of wrasse to specific habitat types (see Model prediction of all wrasse species and Habitat association sections), relative abundance was modelled on suitable habitat only (i.e. ABC (Algae/boulder/cobble) and AGP (algae/gravel/pebble)). The optimal multivariate model to predict wrasse relative abundance included the variables “year” (p <0.001), “depth” (p <0.001), “substratum type” (p <0.001) and the interaction between “depth” and “substratum type” (p <0.01). These variables were tested in the multivariate model across all the observed wrasse species individually. The inclusion of these variables and their relative significance suggests that, overall the physical characteristics of the habitat (depth and substrata) affected the abundance of some or all wrasse species. The abundance of some or all wrasse species was also influenced by the year in which surveys were conducted.
To describe the effects of the above variables on individual species, only the three most commonly observed species were included in figures (goldsinny, ballan and cuckoo wrasse). Any predictions made for rock cook or corkwing would be based on minimal data and be less reliable. Substratum had the greatest influence on the wrasse species considered. Rocky reefs with larger boulder size were associated with higher relative abundance for goldsinny (p < 0.001) and cuckoo wrasse (p < 0.001) compared to habitats with smaller pebbles or gravel (Figure 5a and 5c). Ballan wrasse showed no statistically significant difference (p = 0.072) in relative abundance depending on the type of rocky reef (ABC or AGP) (Figure 5b).
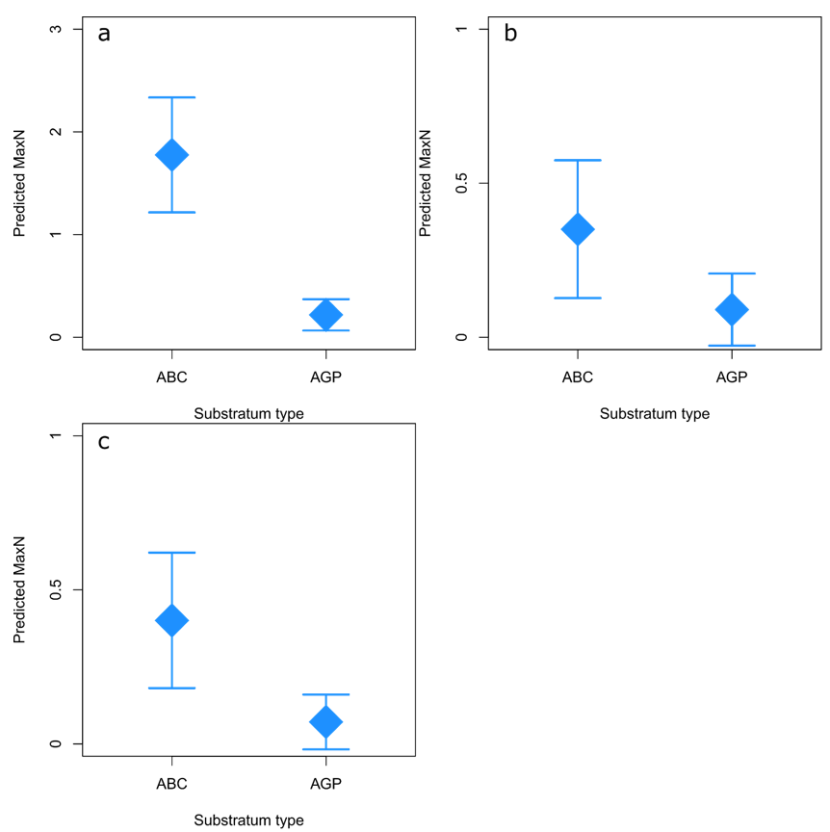
Figure 5. Predicted relative abundance (maxN) of goldsinny (C. rupestris) (a), ballan (L. bergylta) (b) and cuckoo (L. mixtus) (c) wrasse on ABC (Algae/boulder/cobble) and AGP (algae/gravel/pebble) substrata in 2013.
Click for a full description
Predicted relative abundance (measured as maxN) from GAMs using SBRUV observations at 10 m depth in 2013. 95% confidence intervals (CI) are shown. Predicted maxN was highest on ABC substratum for all species, although CIs overlap for ballan wrasse.
The variable “year”, like substratum type, only showed a significant effect on the abundance of goldsinny (Figure 6a, p <0.001) and cuckoo wrasse (Figure 6c, p = 0.05). In these two species the most obvious differences in abundance occur in 2014 (and 2017 in Figure 6a for goldsinny) compared with 2013 and 2018. The large confidence intervals for all species in 2017 are a result of the small sample number and the wide separation between sample points which make comparisons with other years difficult. While sampling points in 2014 were more similarly spaced apart than samples taken in 2013 and 2018, they did not cover the same area to the west of Arran (Figure 2). The resulting data contained many fewer observations of wrasse than years when the west of Arran was surveyed. These inconsistencies in sample coverage were likely the cause of any significant differences in abundance owing to the year in which surveys were conducted. Ballan wrasse showed no significant change in relative abundance over time (p = 0.057).
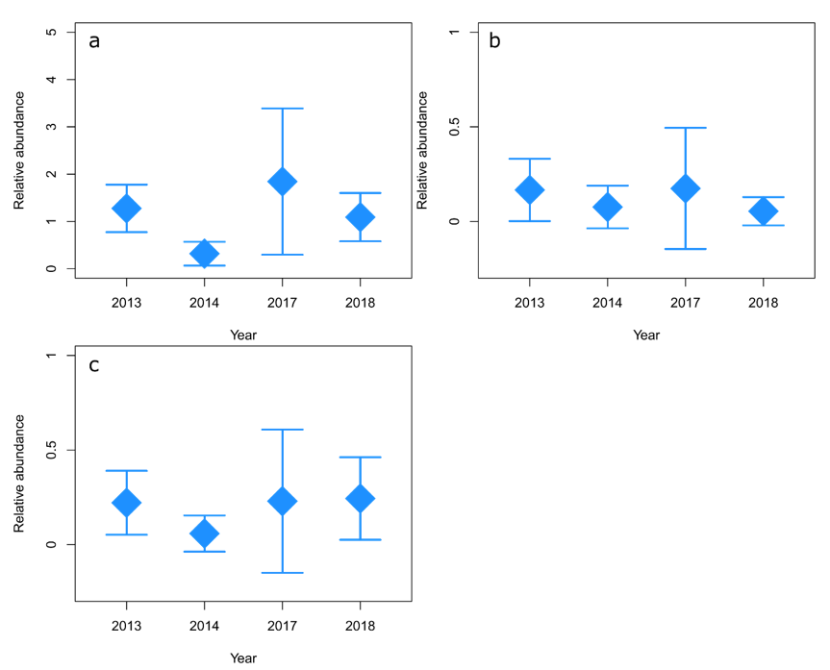
Figure 6. Predicted relative abundance (maxN) of goldsinny (C. rupestris) (a), ballan (L. bergylta) (b) and cuckoo (L. mixtus) (c) wrasse in 2013, 2014, 2017 and 2018.
Click for a full description
Predicted relative abundance (measured as maxN) from GAMs using SBRUV observations on ABC (Algae/boulder/cobble) at a depth of 15 m. 95% confidence intervals (CI) are shown.
Predicted goldsinny abundance was lowest in 2014 and highest (and most variable) in 2017. Predicted ballan wrasse abundance was similar across years, slightly higher in 2013 and 2017 (and more variable in 2017). Predicted cuckoo wrasse abundance was lowest in 2014 and similar in all other years, although more variable in 2017.
The multivariate analysis of wrasse species showed they were significantly more abundant at shallower depths (p <0.001). Figure 7 shows the significant effect of depth on the individual species. The effect of depth on the relative abundance of goldsinny (p < 0.001), ballan (p <0.05) and cuckoo wrasse (p < 0.001) are displayed for 2013 and 2018. The abundances of goldsinny and ballan wrasse were consistently higher at all depths in 2013 compared to 2018 (Figure 7a and 7b). However, there is a great deal of overlap in the 95% confidence intervals of these estimates. In 2018 ballan wrasse abundance was low across its depth range (Figure 7b). The greatest difference in ballan wrasse abundance when comparing 2013 with 2018 can be seen at the shallowest depths (Figure 7b). The difference in abundance between 2013 and 2018 was also greatest for goldsinny and cuckoo wrasse at shallower depths (Figure 7a and 7c), although this effect was less pronounced than in ballan wrasse.
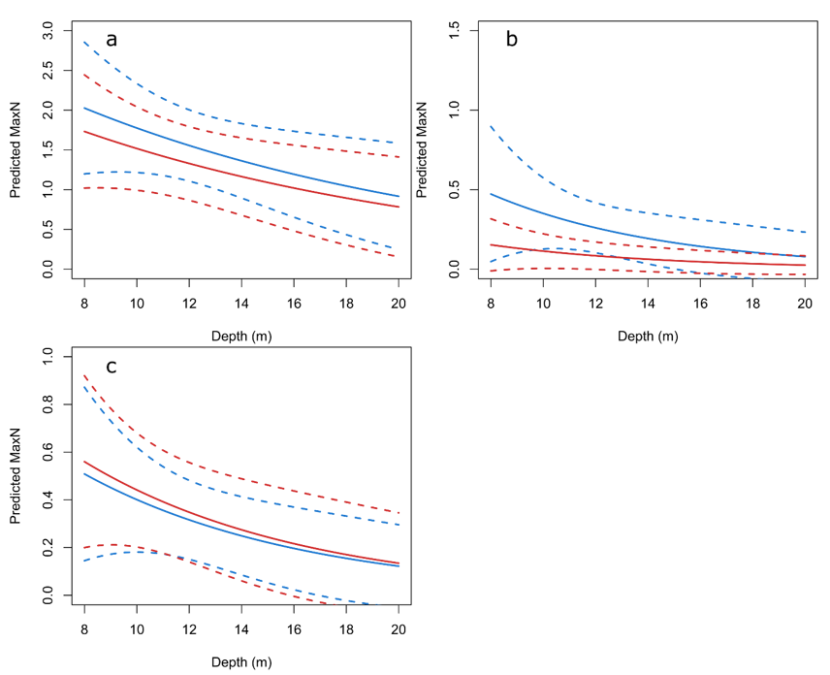
Figure 7. Predicted relative abundance (maxN) change with depth for goldsinny (C. rupestris) (a), ballan (L. bergylta) (b) and cuckoo (L. mixtus) (c) wrasse on ABC (Algae/boulder/cobble) substratum.
Click for a full description
Predicted relative abundance (measured as maxN) from GAMs using SBRUV observations on ABC (Algae/boulder/cobble) across a depth range from 8 to 20 m. Predictions from 2013 are shown as blue lines and predictions from 2018 are shown as red lines. The dashed lines show 95% confidence intervals (CI). Predicted maxN is highest at shallow depths for all species. The differences between years are also greater at shallow depths, especially for ballan wrasse.
Habitat association
Literature review
The literature review found a total of 1358 peer reviewed articles (Table 2); 155 were retained for analysis.
| Search terms | Number of results (Original articles, reviews) | Number of nonduplicate, relevant results retained for analysis | Year attracting most scientific interest (number of articles published) |
|---|---|---|---|
| wrasse + cleaner fish | 385 (346, 13) | 52 | 2018 (36) |
| wrasse + salmon farm | 121 (102, 8) | 21 | 2018 (17) |
| wrasse + UK | 71 (56, 3) | 8 | 2016 (5) |
| wrasse + Scotland | 65 (52, 3) | 4 | 2016 and 2014 (5) |
| Ctenolabrus rupestris | 191 (164, 6) | 15 | 2016 (8) |
| Labrus mixtus | 58 (50, 0) | 2 | 2016 & 2000 (4) |
| Labrus bergylta | 231 (196, 4) | 11 | 2018 (34) |
| Symphodus melops or Crenilabrus melops | 120 (103, 4) | 16 | 2016 (9) |
| Centrolabrus exoletus | 22 | 3 | 2018 (2) |
Details of the five wrasse species, ballan wrasse (Labrus bergylta), goldsinny (Ctenolabrus rupestris), cuckoo wrasse (Labrus mixtus), corkwing (Symphodus melops, formerly Crenilabrus melops) and rock cook (Centrolabrus exoletus) and their associated habitat were extracted from the articles retained for further analysis (Annex 2, Table A2).
Of the literature examined, 45% of the instances of wrasse species mentioned in reference to their habitat preference were in new primary research. The other 55% was made up of citations to other literature documenting habitat preferences. Across all the wrasse species considered, all of the literature discussing habitats occupied by wrasse described wrasse occupying shallow or coastal waters. In the literature examined, when describing prime wrasse habitat:
- 76% of studies described wrasse occupying rocky habitat.
- 51% described algae as an important component.
- 1% discussed seagrass as important for juvenile wrasse.
The relative exposure of sites was also described in 5% of studies, associating goldsinny and ballan wrasse with more exposed sites and corkwing with sheltered sites. Other types of habitat descriptions arose in 10% of the examined literature. These descriptions included patches of sand along with rocky substrata, biogenic reefs and wrasse abundance being higher over more complex habitat. Table 3 displays the percentage occurrence of individual wrasse species in the literature and the types of habitat most commonly described.
| - | Goldsinny (C. rupestris) | Ballan wrasse (L. bergylta) | Cuckoo wrasse (L. mixtus) | Corkwing (S. melops) | Rock cook (C. exoletus) |
|---|---|---|---|---|---|
| % coverage in literature | 40 | 22 | 8 | 22 | 8 |
| Rock | 92 | 88 | 86 | 87 | 86 |
| Algae | 62 | 94 | 57 | 67 | 57 |
| Other | 4 | 0 | 14 | 0 | 0 |
Rocky reef
Coastal rock is a foundation on which many biological habitats are based (Burrows et al. 2017). In the context of this report, the rock formations can be considered underwater rock or large boulders. An estimated 35% of the UK coastline is made up of rocky habitat (Magill and Sayer 2002). Under the Habitats Directive, “Reefs” (code 1170) are characterised by dominance (>50% coverage) of hard substrata (stones and bedrock), while depth distribution is not limited. The typical taxa of this habitat classification are Fucus vesiculosus, F. radicans, F. lumbricalis, all filamentous algae, epifaunal bivalves (Mytilus trossulus, Dreissena polymorpha), and barnacles (Amphibalanus improvisus). The coverage of these typical species must be ≥10 %.
Wrasse (Labridae) are found to associate strongly with rocky reef habitat (Darwall et al., 1992b). Different species of wrasse are some of the most abundant fish species on shallow rocky reefs and coastlines in Northern Europe (Halvorsen et al. 2017a). Goldsinny, corkwing, ballan and cuckoo wrasse have all been shown to associate with various rocky habitats.
Magill and Sayer (2002) found that in the overall fish assemblage on two inshore rocky subtidal sites on the west coast of Scotland, Labridae and Gobiidae were the dominant fish families. At one rocky reef site, goldsinny were the dominant fish species and classed as a resident and characteristic species of this habitat (Magill and Sayer 2002). Other periodic wrasse species, corkwing and rock cook, were recorded in the summer months (Magill and Sayer 2002). As a resident species, goldsinny may display seasonal variation in abundance but is a key species on rocky reef habitat throughout the year (Magill and Sayer 2002). The variation in seasonal abundance is due to wrasse remaining hidden among rocks during the winter, and so they do not appear in traditional surveys (Darwall et al. 1992b, Sayer et al. 1993). This need for refuge results in the need for complex rocky habitat and therefore a strong association between goldsinny and rocky reefs (Sayer et al. 1993). Goldsinny was also found to be the most abundant species on shallow rocky bottom stations in Sweden (Pihl et al. 1994).
Ballan wrasse display extreme site fidelity, sometimes to a single rock or a small group of rocks, especially during the spawning season (Mucientes et al. 2019). This strong site fidelity suggests that even small marine reserves could be effective for protecting ballan wrasse (Mucientes et al. 2019).
Kelp forest and algae
Kelps are large brown seaweeds (Order Laminariales) that grow in “beds” or “forests” forming one of the most widespread coastal habitats in the temperate and subpolar oceans (Smale et al. 2013). Kelps are extremely common along rocky coastlines throughout Scotland with different species dominant, varying according to environmental conditions; Laminaria hyperborean and Alaria esculenta are common on exposed coastlines and Saccarhina latissimia associated with sheltered conditions (Walker 1954, Smale et al. 2013).
Kelp beds and forests are important habitats for a wide variety of marine plants and animals (Dayton 1985). Several studies indicate that wrasse species are strongly associated with kelp and algae. Some wrasse species are strongly influenced by the diversity of prey species inhabiting the algal cover (Bell and Harmelin-Vivien 1983, Ruitton et al. 2000). Some genera, for example, Symphodus spp. are strongly influenced by their dependence on algae to build nests (Potts 1974b, Rodrigues et al. 2015).
Temperate wrasse species (e.g. ballan, goldsinny, and corkwing) are usually associated with rocky habitat covered with macroalgae (Costello 1991, Darwall et al. 1992b, Sayer and Treasurer 1996). Ballan wrasse inhabit both rocky reefs and kelp beds up to 60 m depth (Mucientes et al. 2019). Wrasse use algae to build nests, forage and for shelter (Ruitton et al. 2000).
There are potential implications for ecosystem management if there are large reductions in the wrasse species associated with kelp forests (Pita and Freire 2019). The trophic role of ballan wrasse is important in maintaining a healthy kelp forest ecosystem, as it predates on sea urchins, thereby reducing sea urchin overgrazing on kelp (Pita and Freire 2019).
Seagrass
Seagrasses (Zostera spp.), also known as eelgrasses, are flowering marine plants. Seagrasses often grow in dense beds or meadows that develop in intertidal and shallow subtidal areas sheltered from significant wave action, on sands and muds, down to 10 m. Seagrasses are important habitats in terms of delivery of ecosystem services and function, supporting biodiversity and resilience. In Scotland, seagrass beds are well developed compared to other areas of the UK (Hiscock et al. 2004, Burrows et al. 2017). Seagrasses are included on the OSPAR list of Threatened and/or Declining Species and Habitats and in Scotland, seagrass is listed as a Priority Marine Feature (PMF).
Labridae and Gobiidae have been found as the dominant fish families in UK seagrass habitats (Jackson et al. 2006). Often, the wrasse present in seagrass habitat are dominated by juvenile life history stages e.g. juvenile ballan and corkwing (Jackson et al. 2006). The cuckoo wrasse can be found in seagrass meadows in addition to rocky and kelp habitats (Matić-Skoko et al. 2013).
Interaction with priority and protected features
There are potential interactions of the wrasse fishery with several priority and protected features. Based on the results of the literature review, there is a close association between wrasse and several habitats that are listed as Scottish PMFs (Table 4). These include: Seagrass beds, Kelp and seaweed communities on sublittoral sediment, Kelp beds and Tide-swept algal communities. There is a potential for trophic level effects if wrasse are removed in large numbers from kelp habitats, as wrasse are important in maintaining healthy kelp ecosystems (Pita 2019). Several wrasse species also show a strong association with Reef habitat protected under the Habitats Directive (Table 4).
| Habitat | Protected Site (MPA or SAC) |
|---|---|
| Seagrass beds (PMF) | MPA:
|
| Kelp and seaweed communities on sublittoral sediment (PMF) | MPA:
|
| Kelp beds (PMF) | n/a |
| Tide-swept algal communities (PMF) | n/a |
| Rocky Reef (Annex I) | SAC:
SACs where rocky reefs are a qualifying feature, but not a primary reason for site designation:
|
While Table 4 lists areas within the Scottish MPA network which contain kelp and seaweed communities and reef habitat, these features also exist outwith MPAs and SACs. These habitats are also present in protected areas while not being listed as a reason for the designation. These areas which do not specifically cite the habitat type as a reason for the designation are not included in Table 4. Further, the restricted, shallow depth range over which wrasse occur (see also Section 3.2.1) means some of the listed protected areas (Table 4), while containing kelp and seaweed communities and reef habitat do not include suitable wrasse habitat.
The wrasse fishery also has the potential to interact with other PMF species which are associated with the same habitats as wrasse. Juvenile gadoids like cod (Gadus morhua) and whiting (Merlangius merlangus) occupy similar shallow water algal rocky habitats and seagrass at the same time as the wrasse fishery operates (Elliott et al. 2016). Otters (Lutra lutra) forage in shallow algal rocky habitats (Clavero et al. 2006, Findlay et al. 2015) which are also strongly associated with wrasse. There is therefore a potential interaction between the wrasse fishery and otters. If not properly modified, baited traps will attract otters, trapping and drowning them. Additional effects of the wrasse fishery on otters may also include limiting the availability of prey species if fish abundance is reduced through disturbance from noise and human activity. Assuming the wrasse fishery could reduce prey availability for otters, there are also a number of coastal SACs designated for otters that could be potentially at risk from an unmanaged wrasse fishery. These SACs include: Yell Sound Coast, Loch Etive Woods, Loch Moidart & Loch Shiel Woods, Moine Mhor and South Uist Machair.
Overall, the wrasse fishery may interact with some designated sites in the Scottish MPA network more than others. Some sites may have little interaction with the fishery, however, without a current assessment of the fishery footprint, it is difficult to know which sites are heavily or minimally impacted.
Biological comparative tables analysis
A species is considered to be present in a biotope in the JNCC biological comparative tables if it occurs in more than 20% of the records. At all levels of EUNIS classification, wrasse species were present in 59 biotope types of infralittoral habitat (IR) on the JNCC biological comparative tables. In circalittoral habitats (CR), wrasse species occurred in 79 records of the JNCC biological comparative tables (further details in Annex 3, Table A3).
Amalgamating these occurrences to EUNIS level 4, which includes biotope complexes like “Kelp with cushion fauna and/or foliose red seaweeds (IR.HIR.KFaR)” and “Kelp and seaweed communities in tide-swept sheltered conditions (IR.MIR.KT)” best represents biological communities. Figure 8 displays the amalgamated occurrences of four wrasse species - ballan, goldsinny, cuckoo and corkwing. Goldsinny was the most widespread occurring in 12 biotope types followed by ballan in 11 biotopes, while cuckoo and corkwing were only observed in 5 biotopes. Only shallow water rocky habitats had occurrences of wrasse and most occurrences were on biotopes characterised by the presence of macroalgae. However, faunal communities like sponges, molluscs and echinoderms must provide suitable habitat, with relatively high occurrences of wrasse also being observed in these biotopes.
The common characteristics of the biotopes where wrasse were present were the presence of a rocky substratum and some level of habitat complexity provided by biotic communities. Biotopes where wrasse occurrences were not recorded were characterised by softer substrata, less habitat complexity and the absence of other habitat-forming biota.
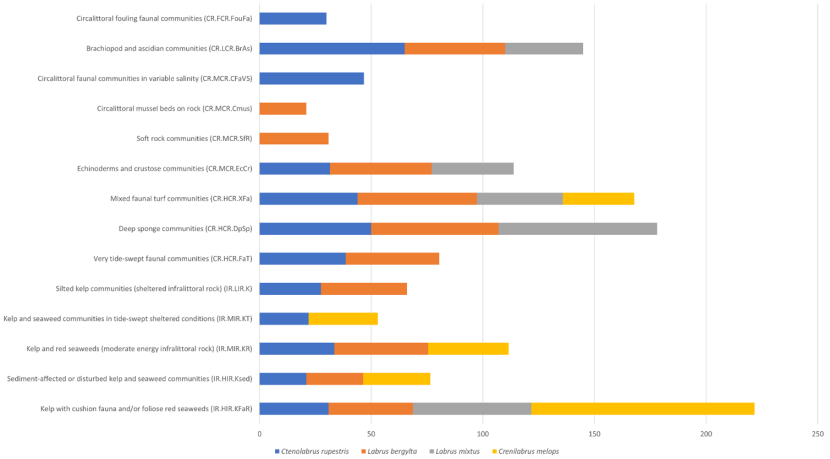
Figure 8. Number of biotope records where wrasse species are considered to be present according to JNCC Biological Comparative tables.
Click for a full description
Number of individual records for each biotope where each wrasse species occurred in more than 20% of the biotope records. The biotope with the highest number of records of wrasse presence was “Kelp with cushion fauna and/or foliose red seaweeds (IR.HIR.KFaR)”. Goldsinny were present in the highest number of different biotopes (12), followed by ballan wrasse (11). Cuckoo and corkwing wrasse were each present in five biotopes.
Wrasse probability of presence habitat association analysis
The optimal binomial Generalised Additive Model (GAM) contained the variables: “depth”, “substratum type” and “year”. The GAM showed that wrasse were more likely to be present in shallower depths. The most important variable in the GAM was substratum type. The presence of wrasse was highly associated with algae covered hard substrata, particularly ABC (Algae/boulder/cobble) during all years of the study (2013, 2014, 2017 and 2018) (Figure 10). ABC (Algae/boulder/cobble) and AGP (algae/gravel/pebble), shaded in green in Figure 10b to the south and southwest of Arran, show the highest probabilities of presence (Figure 10a) for all wrasse species.

Figure 9. Images of cuckoo wrasse (Labrus mixtus) and typical algae/boulder/cobble (ABC) habitat.
Click for a full description
Wrasse attracted to SBRUV deployment during the 2018 surveys in the South Arran NC MPA. One male cuckoo wrasse is shown in picture (a). Pictures (b) and (c) show a male (bottom right) and female above and to the left. Picture (d) shows typical ABC habitat with Laminaria hyperborea (photographed at St Kilda by Richard Shucksmith, SNH).
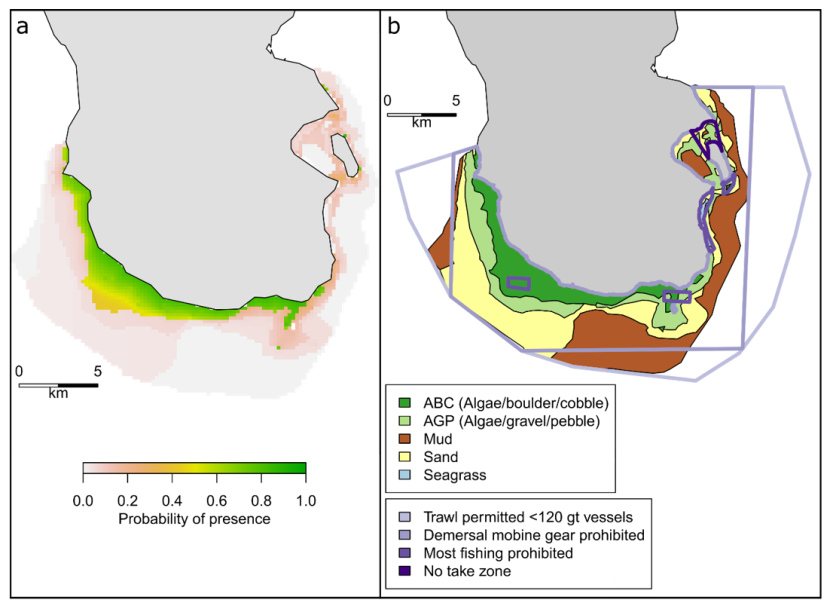
Figure 10. Probability of presence across all wrasse species for all years combined (a) and predicted substratum types (b) within the South Arran NC MPA.
Click for a full description
Heatmap of probability of presence across all wrasse species in the South Arran NC MPA from 2013, 2014, 2017 and 2018 SBRUV observations combined (a). Areas with a higher probability of presence are shown in green. Map of modelled substrata in the South Arran NC MPA from Elliott et al. (2017a) (b). Purple lines show the four levels of fisheries management measures in the NC MPA.
The area where the highest probability of wrasse presence was predicted (a) overlaps with the algae/boulder/cobble (ABC) habitat (b).
Figure 11 displays the probability of wrasse from the binomial GAM being present on each type of substratum. Having a median probability of 0.52, ABC (Algae/boulder/cobble) is the most likely substratum to be occupied by wrasse. AGP (algae/gravel/pebble), the only other hard substratum modelled, also showed a higher likelihood of harbouring wrasse than the soft substrata modelled (median probability = 0.04) but to a much lesser extent than ABC. Soft substrata (mud, sand, seagrass) displayed zero or little likelihood of wrasse being present. Wrasse were observed occurring over sand (median modelled probability of occurrence = 0.01). However, on these occasions the wrasse observed were moving between habitats composed of hard substrata.
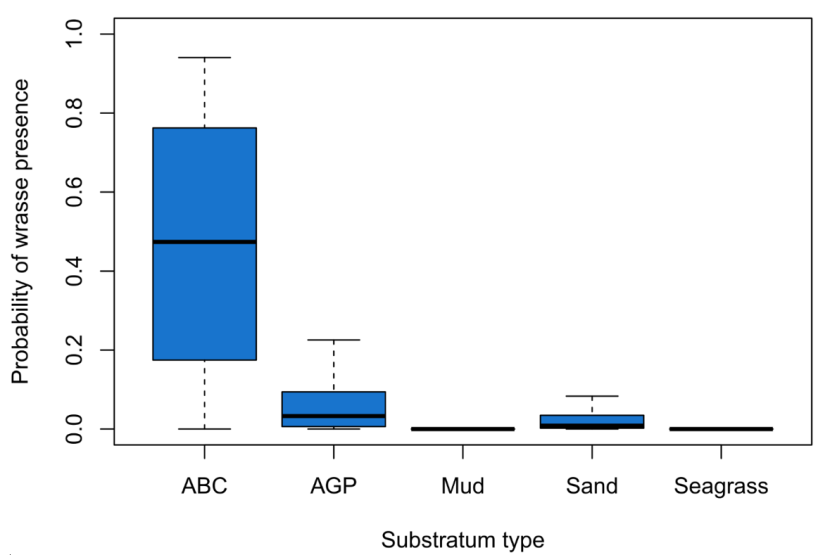
Figure 11. Probability of wrasse presence across all species for different substratum types.
Click for a full description
Thick horizontal lines represent the median probability of wrasse presence and whiskers represent the upper and lower quartiles.
The highest probability of wrasse presence was on algae/boulder/cobble (ABC) habitat (median probability = 0.52), followed by algae/gravel/pebble (AGP) habitat (median probability = 0.04). The probability of presence was very low on sand (median probability = 0.01) and no wrasse were observed on mud or seagrass.
Consideration of wrasse monitoring methods
There are three suggested monitoring options described in the following section with an analysis of the advantages and disadvantages of each option. Other monitoring options exist but in the view of the authors, the three methods we describe offer the best balance of costs and benefits. The same set of sampling sites and survey timings are proposed for the three methods.
- SCUBA diver transects using stereo video camera.
- Stereo baited remote underwater video (SBRUV) camera deployments.
- Fyke netting and fish trapping
SCUBA diver transects
Method
SCUBA diver transects can be conducted as described in Elliott et al. (2016). Briefly, sampling locations should include algae covered rocky substrata to ensure wrasse habitat is being sampled. Transects should also be located to stratify sampling for abiotic variables like wave fetch, depth and possible salinity or temperature gradients. 100 m x 4 m or 50 m x 4 m belt transects could be used depending on site specific constraints. Belt transects are a standard and accurate method for assessing fish abundance (Kimmel 1985, Hunter and Sayer 2009).A leaded line can be used in place of a measuring tape to make the operation of laying the transect line faster. Transects should be laid at constant depth following the depth contour. Each dive day can consist of two dives and 6 transects (three transects per dive). For example, dive one could consist of two transects at 20 m depth and one at 10 m, dive two could then consist of one 20 m deep transect and two at 10 m depth. After laying the transect line a settlement period should be allowed for any disturbance caused by the divers to dissipate (Dickens et al. 2011). The divers should swim at a constant speed (ideally 10 m/min) along the transect line at about 1 m above the seabed with stereo cameras held at such an angle to capture fish in front of the field of view and the substratum. These methods should be usable for citizen scientists who are experienced temperate water divers, but they would require further training.

Figure 12. SCUBA diver conducting a transect survey using a stereo-video camera system.
Click for a full description
Only one of the two cameras in waterproof housing is visible (a). Another similarly positioned camera is at the other end of the frame. Additional buoyancy may be required for heavy frame designs (b). A leaded line (c) can be laid to measure the transect length and aid diver navigation.
Human resources required
A minimum of two qualified divers is needed for the scientific dive team (e.g. Benjamin and Mackintosh, 2016) and an additional Dive Supervisor (Health and Safety Executive 2014). A boat skipper is also required. The costings below assume a standing research dive team and therefore do not include the costs of training and dive specific equipment.
Physical resources required
Portable stereo video camera system with lighting. SCUBA equipment. A boat would be required for sites inaccessible by shore. A weighted survey line for measuring the length of the transect.
Advantages and disadvantages
| Advantages | Disadvantages |
|---|---|
| Density measures can be calculated i.e. counts per m2 | Depth limited to the range of scientific diving capabilities |
| Additional observations or experimental manipulations are possible with a diver in the water | High level of scientific diver training required and maintaining a regular dive team can be difficult |
| Fish lengths can be calculated from stereo video camera footage | Analysis of the resultant stereo video camera footage is time consuming |
| A permanent record of the data is created which can be re-analysed, re-categorised and archived (if video is used) | Low number of dives possible per diver day |
| The seabed type/benthos is recorded | Relatively expensive |
Estimated costs
| Resource | Cost |
|---|---|
| Divers | Min 2 divers, 1 safety cover + boat crew |
| Camera equipment (no lighting) | £1,300.00 - £2,500.00 |
| Stereo video calibration hardware and software | £5,319.18* |
| Boat | £500.00 - £1,000.00 per day |
*The authors at the University of Glasgow and MSS have current licences for this software and own the required hardware.
SBRUV camera deployments
Method
Surveys using Stereo Baited Remote Underwater Videos (SBRUV) cameras can be deployed from small vessels like Rigid Inflatable Boats (RIBs) but larger (> 7 m) hard boats are more suitable. Ideally the vessel used would have a large deck area for SBRUV equipment handling and a winch. Commercial creeling vessels are ideally suited to surveys using SBRUV camera equipment.
A stratified random sampling approach should be taken to SBRUV camera deployments. Sampling should be stratified by relevant environmental variables like depth, wave fetch and substrata. However, given the close association of wrasse with hard substrata like algae covered rocks these areas should be prioritised. Using more than one SBRUV camera frame will maximise sampling efficiency.
SBRUV camera systems can be set-up as described in Harvey and Shortis (1995, 1998). Ideally the system will use a pair of high-definition cameras in waterproof housings (e.g. SeaGIS, 2013). The cameras should be angled inward to maximise both the field of view and overlap. Underwater torches can be mounted on the camera frames to provide better illumination and aid species identification. A strobe can also be used at the beginning of filming to synchronise the stereo-video images.
Camera systems should be deployed for one hour. This length of deployment time maximises the bait soak time while allowing for multiple deployments in a day (Unsworth et al. 2014). SBRUV deployments should be made a suitable distance apart to avoid interference between one bate plume and another. This distance will depend on local conditions, namely tidal flow and local currents. Deployments should also be made at least three hours after sunrise and before sunset to avoid diurnal variation in fish behaviour (Keats and Steele 1992, Bertelli and Unsworth 2014).
The most appropriate method of estimating relative abundance using SBRUV systems (and un-baited camera systems) records the maximum number of individuals of the same species appearing in a frame at the same time (maxN) (Priede et al. 1994, Cappo et al. 2005, Watson et al. 2005). Using the maxN method rather than simply counting individuals throughout the recording period avoids repeatedly counting the same individual re-entering the field of view. To use the stereo capability of an SBRUV system effectively and be able to measure the length of fish, videos must be analysed with specialist software (e.g. Event Measure software).
The SBRUV method described here would also be a suitable methodology for citizen scientists/fishers to conduct. Training would be required, and equipment costs may be prohibitive for an individual. Fishers targeting wrasse are in an ideal position to deploy SBRUV equipment and could play an active role in monitoring their target species.

Figure 13. SBRUV camera frame with bait box (i), strobe (ii), video camera and housing (iii), and operational data logger for temperature and environmental variables (iv).
Click for a full description
-
Human resources required
Two people are needed for the SBRUV team for deploying and operating the camera system. A boat skipper is also required.
Physical resources required
SBRUV system and frame with lighting. A boat would be required to deploy the equipment. The capital expense for this survey method is high. However, this is offset by the number of samples which can be collected over the life of the equipment even assuming short-term three-year depreciation.
Advantages and disadvantages
| Advantages | Disadvantages |
|---|---|
| Existing surveys in the South Arran NC MPA have used this method which allows for comparisons with existing data | Relative abundance measures only can be calculated |
| A low level of expertise is required to carry out the data collection | Analysis of the resultant stereo video camera footage is time consuming (3 hours analysis per 1 hour survey footage) |
| Fish lengths can be calculated from stereo video camera footage | Potential for equipment loss |
| A permanent record of the data is created which can be re-analysed, re-categorised and archived | - |
| The seabed type/benthos is recorded | - |
| Deployment and recovery possible from small vessels. | - |
Estimated costs
| Resource (assuming 3 SBRUV platforms) | Cost |
|---|---|
| Survey team | 1 staff + boat crew |
| Camera equipment, lights & frame | £9,967.74 |
| Stereo video calibration hardware and software | £5,319.18* |
| Boat | £500.00-£1,000.00 per day |
*The authors at the University of Glasgow and MSS have current licences for this software and own the required hardware.
Fyke netting and fish trapping
Method
Fyke nets (Figure 14) are cylindrical shaped nets held open with metal rings. The conical shaped partitions with one-way entries trap fish (Van Der Veer et al. 1992). Single or multiple leader nets direct fish into the trap body. Multiple fyke nets can be linked together in fleets to sample a larger area (e.g. Treasurer, 1994). Otter boards should be attached to fyke nets to stop otters entering the trap body and drowning. Given the high wave exposure and strong tidal flow often found in prime wrasse habitat, fyke nets should be firmly attached to stakes or heavily weighted (Figure 14). However, fyke nets may not be suitable for use in areas with very strong currents. In conditions with strong wave action or fast flowing currents the leader nets will likely collapse, reducing their efficiency.
As with the previously discussed methods, sampling sites should be selected to encompass prime wrasse habitat and should be stratified to account for relevant environmental variables like depth. Fyke nets can be set at slack water and low tide to aid deployment and left to fish for 24 hours. In very shallow water sampling areas, deployment sites may be accessible from the shore by wading. However, to sample across the depth range where wrasse occur, use of a boat will be required. An initial visual site survey is recommended to assess its suitability. Sites with very steep slopes, large boulders or other entanglement hazards should be avoided. The fyke net should be deployed with leader nets perpendicular to the shore as fish often move parallel to the shore. With weights attached at the closed end of the net it can be lowered into the water until it reaches the seabed. The rest of the net can be deployed, playing out the net as the boat reverses. SCUBA divers may be required to hammer stakes into the substratum or secure the net if it will be left in high energy conditions. A shot line and buoy attached to the leader net(s) are used for recovery. Once recovered, fish are removed from the end pot by untying the end of the net.
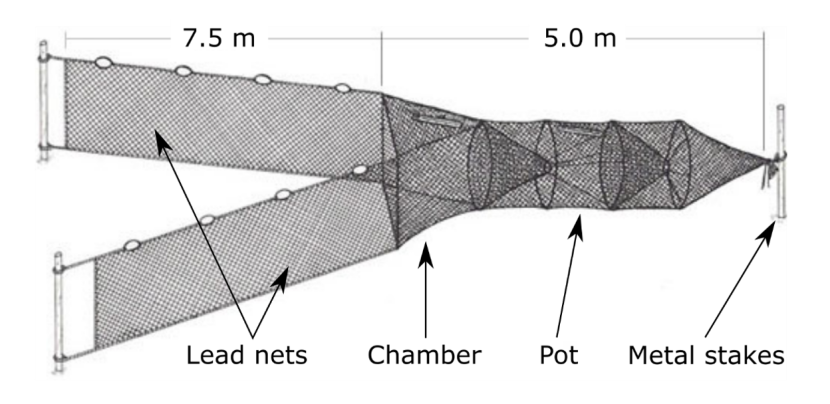
Figure 14. Diagram of an example fyke net.
Click for a full description
The example shown includes 7.5 m long lead nets, as well as the chamber, pot, and metal stake beyond the pot (collectively measuring 5 m). Several designs exist but most comprise of the labelled components. Multiple or single leader nets can be used depending on the site and floats mean they remain vertical in the water column. Weights or metal stakes (in fast currents and areas of strong wave action) are used to hold the net in place. Diagram adapted from fyke net supplier Fipec Industries (2019).
Fish traps used in fish surveys can be traps used commercially and can be deployed from the shore for shallow sample sites or from boats in deeper water. For wrasse in particular, modified Nephrops creels (57 cm x 40 cm x 30 cm, entry aperture of 40 mm and 12 mm mesh size) have been used successfully (e.g. Treasurer, 1994). A thorough understanding of the biology and behaviour of target species is required because traps are highly selective. For example, the use of bait may bias results in favour of species attracted to a particular bait type. Territoriality may also affect trap selectivity, with species like goldsinny not moving through the territories of other conspecifics to enter a trap. While trap selectivity will vary between species, for any one species sampled at a given location and time, the relative abundances generated from trapping can be used to monitor annual population fluctuations.
Traps are more robust than fyke nets and can be used to sample higher energy areas and over rough ground. Traps can be deployed individually with a marker buoy for each trap or more commonly deployed as a fleet of several traps. Fleets of traps should be deployed similarly to fyke nets with the first trap lowered to the seabed by the ground rope and other traps deployed as the boat reverses. In shallow, tidal areas, traps can be deployed from the shore.
Human resources required
Two people are needed for the data collection team for deploying the fishing equipment and recording the data. A boat skipper is also required.
Physical resources required
Fyke nets or fish traps.
A boat would be required to deploy the traps in deeper water.
Weights (10 kg per pair of fyke nets).
Advantages and disadvantages
| Advantages | Disadvantages |
|---|---|
| Identical fishing gear to commercial fishing gear could be used which allows for comparisons with commercial fishing data | Only relative abundance measures can be calculated (i.e. not quantitative) |
| A low training cost is required to carry out the data collection | No data on the seabed type/ benthos is collected |
| Specimens can be collected for analysis | Fyke nets can be difficult to deploy correctly |
| Equipment is relatively cheap providing a cost-effective technique | - |
| Data collection is fast | - |
Estimated costs
| Resource | Cost |
|---|---|
| Survey team | 2 staff + boat crew |
| Fyke nets and fish traps | £135.00 (per fyke net), £60.00 (per fish trap) * |
| Boat | £500.00-£1,000.00 per day |
*Net and trap costs based on average price from three suppliers, January 2020.
Discussion
The findings of the SBRUV surveys in the South Arran NC MPA, and the findings related to wrasse habitat association are discussed below in relation to each other. The implications of these results for wrasse fishing and protected features are discussed.
Wrasse abundance
South Arran NC MPA
This is the first analysis of fishery-independent relative wrasse abundance data from Scotland. Any insight into wrasse abundance has the potential to be valuable in informing both the conservation of the species and the management of the fishery. The results describing wrasse relative abundance demonstrate the importance of consistent sampling methodology. While changes in abundance are observed over time, any true change in abundance which resulted from fishing activity is masked by the inconsistencies in sampling methodology. The data analysed here have been used opportunistically and the methodologies used were designed for several different studies. For example, in 2016 SBRUV platforms were deployed in clusters to satisfy specific sampling aims and did not sample suitable wrasse habitat. In 2017 surveys conducted by MSS did not have sufficient sample numbers to properly sample the area required and in 2014 an area of suitable wrasse habitat (see section 3.2.3 for more detail) was omitted. Given these constraints, a definitive assessment of changes in wrasse abundance over time is not possible for the South Arran NC MPA.
Nonetheless, the results are helpful in informing the development of future approaches to monitoring wrasse and do highlight the importance of understanding the ecology of any species which is to be surveyed. Section 3.2 of this report aims to address this knowledge gap. However, from the results here it is clear that habitat type, depth and the spatial distribution of sampling points must be carefully defined to properly sample wrasse.
The wrasse species found in the survey - ballan wrasse (Labrus bergylta), rock cook wrasse (Ctenolabrus exoletus), cuckoo wrasse (Labrus mixtus), goldsinny (Ctenolabrus rupestris), and corkwing (Symphodus melops) - are among those expected in relation to their natural range and the recorded habitat types.
Of the species that were found, corkwing and rock cook were recorded infrequently. The small numbers of corkwing and rock cook observed is indicative of the type of sampling method used (SBRUV). This method requires large sample sizes to generate meaningful data. These species (corkwing and rock cook) may be present in the South Arran NC MPA in small numbers. Alternatively, these species may be less attracted to the bait used and therefore not represented in baited camera surveys. The presence of more abundant species in the survey such as goldsinny is likely the result of actual higher abundance, but this species may also be more attracted to the camera drop and the bait used.
Fluctuations in wrasse numbers were observed across the years of the surveys. Sampling in 2014 showed reduced abundances for all species. This is likely the result of reduced sampling effort on prime wrasse habitat (rocky reef). Sampling in 2017 also had a reduced number of SBRUV deployments and detected higher relative abundances than 2014, but with greatly reduced confidence. The sampling protocol used in the 2017 surveys conducted by MSS consisted of lower sampling effort and clusters of camera drops (non-even distribution). This means that limited conclusions can be drawn from the years 2014 and 2017.
When comparing the relative abundance of all wrasse species on all substrata from 2013 to 2018, no change in relative abundance is detectable. However, when comparing the relative abundance of individual wrasse species from 2013 and 2018 on prime wrasse habitat, some evidence for reduced abundance is observed for goldsinny and ballan wrasse. The overall more abundant goldsinny showed a marginal decrease in relative abundance between 2013 and 2018. In shallower water ballan wrasse were also detected less frequently in 2018 compared to 2013. While a great deal of overlap existed in the confidence intervals for these abundance estimates between years (Figure 7), a precautionary approach should account for the potential for declines of these species. Therefore, future work examining changes in wrasse abundance through time must: (i) be species specific, while accounting for correlations between species resulting from their similar ecological requirements, and (ii) consider the strong habitat association shown by all species.
Wider implications
The South Arran NC MPA case study site (280 km2) used here provides a localised analysis of wrasse abundance subjected to minimal fishing pressure and includes a 2.66 km2 no-take zone (all fishing prohibited). Other wrasse fishing areas are likely to be more heavily fished and lack any form of fishing prohibition. In other areas with heavier fishing pressure, declines in wrasse abundance may be more pronounced. Additionally, the influence of fishing will likely extend beyond simply removing individuals from the population. Given that many wrasse species remain hidden among rocks during the winter, all fishing pressure occurs during warmer months which coincide with wrasse breeding. Fishing using traps during this time is likely to selectively target larger fish, and in corkwing, particularly males (Darwall et al. 1992b). The removal of dominant males is likely to affect population structure, may destabilize social structure (i.e. territories and dominance hierarchies), and removal of nest-guarding male corkwing would reduce egg survival (Darwall et al. 1992b, Halvorsen et al. 2017b). Therefore, fishing pressure will remove individuals from the population and reduce resilience by affecting the potential for future recruitment. Adopting a precautionary approach, it should be assumed that wrasse fishing will affect wrasse abundance. Additionally, the influence of wrasse fishing on individual species may not be consistent (Halvorsen et al. 2017b). Therefore, any approaches to management, including recording of landings, should be species-specific rather than grouping all wrasse species under one “wrasse” taxon.
Wrasse fishery and protected features
Habitat association
The results from the probability of presence analysis show a very strong habitat preference among wrasse species for hard substrata. The association of wrasse with these rocky reefs is strengthened as reefs are also frequently associated with kelp which some wrasse species like corkwing use as nesting material. Wrasse are also more likely to be observed on rocky reef with larger boulder size rather than smaller cobbles. Presumably larger boulder sizes allow more usable gaps for wrasse to hide and overwinter in. The overlap of high probability of wrasse presence areas on the map in Figure 10 with the ABC substratum type explains why the positioning of sample sites is critically important to properly survey any wrasse species. The habitat association between wrasse and rocky reef is such that they are much more likely to be found here than on any other type of substratum and, that observing wrasse on substrata which are not rocky reefs is very unlikely.
Wrasse habitat association is very clear from the probability of presence model. All wrasse species included in the model were most likely to be found on hard substrata habitats, particularly coarser habitat classified as “Algae/boulder/cobble”. Wrasse were also likely present on the only other hard substratum habitat modelled, “Algae/gravel/pebble”. The model predictions of presence show that wrasse are much more likely to be present in areas of rocky reef habitat than any other habitat type. These model predictions are further supported by a review of the wider scientific literature which shows wrasse species to be strongly associated with rocky reef habitat and kelp (e.g. Magill and Sayer, 2002; Skiftesvik et al., 2015; Halvorsen, Larsen, et al., 2017). Analysis of the biotope records (JNCC Biological Comparative Table - Figure 8) where wrasse were observed further supports the strong association of wrasse with rocky reef and kelp habitat.
The probability of presence model also shows that where rocky reef is absent, wrasse are unlikely to be present. The other substrata considered in the model include soft substrata like mud, sand and sand including seagrass. We therefore consider wrasse to be typical species of rocky reef habitat. However, based on observations from other scientific studies, it is likely that seagrass habitat may be important for juvenile life stages (Jackson et al. 2006).
Interactions with protected habitats and species
The fishery for wrasse species will have multiple interactions with protected habitats and the associated communities, and where overfishing results in depleted wrasse populations, negative impacts are likely. Reducing the abundance of any individual wrasse species will influence algae covered rocky reefs by: (i) reducing numbers of key characteristic species (wrasse), (ii) changing benthic habitat and fish community structure and (iii) altering the trophic structure of food webs of communities reliant on the feature. Although it is recognised that the use of static gears (in this case fish traps or modified creels) have a lower potential for abrasion than mobile gears (e.g. trawling or dredging), they can have impacts on rocky reef habitats (Eno et al. 2001, Shester and Micheli 2011).
Damage to algal communities is particularly impactful for wrasse and other taxa which use macroalgae for refuge, nest building and foraging. Beyond the direct removal of wrasse and impacts to habitat features, entrapment of non-target species is also of concern. In particular, the otter (Lutra lutra) is likely to be exposed to this fishery where traps are set in waters less than 10 m depth. However, there is the potential to mitigate this risk through modification to the fish traps to prevent otters from entering in the first place. Otter guards are required, for example, in the eel fishery in England and Wales (under byelaws) (Environment Agency 2017). The specification for fish and shellfish traps in England and Wales (for eel, crayfish, etc.) is, “Traps must have a rigid ring guard (no more than 95mm wide inside) fitted to the funnel. The trap must be made of one of the following: tightly stretched flexible netting with a mesh size no more than 75mm when wet knot to knot, or 300mm round the edge.” Therefore, it is possible that mitigating the risk of otter entrapment in wrasse fishing traps could be relatively straightforward.
Otter guards are fitted to the relevant access aperture on a fish trap, and while narrow enough to prevent the entry of otters, they are unlikely to affect the ability of wrasse to enter the trap and this could be readily assessed. However, currently there is a lack of any description of the types of fishing gear used, the intensity or the distribution of fishing effort in the wrasse fishery. While the use of ‘otter guards’ are referred in the industry code (Marine Scotland 2018), there are no specifications or guidance for their use in bespoke fish traps or modified crustacean creels. Otters are classed as European Protected Species (EPS) under the Conservation (Natural Habitats, &c.) Regulations (1994). Therefore, otters are protected within and outside protected areas. To aid fishers in avoiding the entrapment of this protected species, specifications of traps that prevent otter entry should be part of the management of the fishery.
Wrasse monitoring
Adoption of any of the suggested monitoring strategies in this report will provide adequate data to identify wrasse species present at sample locations and (depending on the method adopted) provide either density or relative abundance measures at the time of survey. Ideally any monitoring strategy adopted would be consistent and supply long-term data from several locations, cost-efficient and practical to implement and operate. Importantly, the method of data collection must be consistent between geographic locations and through time.
The wrasse abundance data collected in this report in the South Arran NC MPA shows that even slight changes in methodology (e.g. the clustering of sample sites in 2016 or the differing spatial coverage in 2014) may misrepresent trends in abundance. The data collected in the South Arran NC MPA identified algal covered rocky reefs to be the most likely habitat to harbour all wrasse species. This report provides supporting evidence that this habitat type should be targeted in survey designs. The spatial distribution of sampling effort should reflect this strong habitat association focusing on areas on and around rocky reef habitat.
In the short term, continuing the already established monitoring programme in the South Arran NC MPA using SBRUV deployments should be considered. There would be merit in continuing to collect data in this area as this would provide a longer-term dataset to enable further comparisons over time.
Immediate priority should be given to developing data sets which allow wrasse abundance to be assessed in several representative locations around the Scottish coast. These additional representative locations should include protected and unprotected sites. The inclusion of unexploited areas (i.e. where wrasse fishing is excluded) would be valuable in a monitoring context as it would allow differences between natural and anthropogenic changes to wrasse populations to be determined. Using the existing SBRUV deployment methodology would allow new monitoring sites to be compared to existing data and could be implemented quickly. Longer term strategies could include SCUBA diver transects to establish the relationship between the relative abundancies recorded from SBRUV deployments and actual fish densities.
Knowledge gaps, priority research areas and recommendations
The intentions of the Scottish Government outlined in the “Future of Fisheries Management in Scotland: National Discussion Paper” are towards high standards in fisheries management (The Scottish Government, 2019). This includes documented fishing with measures to prevent overfishing and impacts on the wider environment. These intentions reflect existing commitments in the Common Fisheries Policy (CFP) to adopt an ecosystem approach to fisheries management. Based on the Scottish Government commitment to deliver a sustainable, evidence-based approach to the management of Scottish fisheries based on high quality scientific data (The Scottish Government 2019), our recommendation is to continue to collect data on the wrasse fishery which should be used to inform management decisions.
Given the close association of wrasse species with Scottish Priority Marine Features (PMFs) such as “tide-swept algal communities”, “kelp and seaweed communities on sublittoral sediment” and “kelp beds”, several key habitats occupied by wrasse are already within designated NC MPAs. There is also a close association with wrasse species and features listed under the Habitats Directive, like “Reefs” (code 1170), which are represented within Special Areas of Conservation (SAC) sites. This close association with algae covered rocky reefs limits the spatial distribution of wrasse and makes spatial management measures an obvious choice to manage this fishery.
Rocky reef habitat within protected areas is, in several cases, already managed to restrict fishing using mobile gears like trawl and dredge. Taking a precautionary approach to managing the wrasse fishery should include consideration of extending Scottish MPA network management measures to include fish trapping/creeling for wrasse on rocky reef features within existing protected areas. Spatial measures are appropriate for the wrasse fishery given the potential for local depletion of stocks (Halvorsen et al. 2017a). The current Scottish MPA network contains a variety of sites that are relevant to the wrasse fishery i.e. sites that are potentially vulnerable and sensitive to the pressures associated with wrasse fishing. It will be important to prioritise spatial management in these MPAs. This would prevent adverse impacts on the MPAs but also support the sustainability of the fishery. In this way the sustainability of wrasse fishing to supply the salmon farming industry could be improved using the existing Scottish MPA network.
All sites within the Scottish MPA network have requirements for monitoring to indicate site or feature condition, and that there should not be a deterioration in condition. Information will therefore be required on habitat extent, its structure and function; and, also on the status of the various species the habitat typically supports (Marine Scotland, 2017). Given the close association of wrasse species with algae covered rocky reef habitat, and our recommendation that they be considered typical species, the condition of wrasse species will likely influence wider site and habitat feature condition. Therefore, monitoring and management of wrasse fishing and their associated habitats will be necessary in the assessment of site condition. For “Reef” features and otters under the Habitats Directive, governments are responsible for their “Conservation Status” across all waters under their control, not just those reefs and otters inside SACs. Additionally, development and use of the marine environment must consider the national status of PMFs.
The information on wrasse abundance and wrasse habitat association collected in the South Arran NC MPA provides some preliminary indications of the implications for the monitoring and management of protected sites and features and the wrasse fishery. Monitoring and further research effort should be targeted at rocky reef habitats to improve baseline data for wrasse species. A wider distribution of survey sites to include rocky reef habitat on the west coast of Scotland is likely to further improve understanding of wrasse abundance.
Recommendations:
- Availability of accurate data: The estimates of wrasse abundance were based on a small amount of robust data from baited camera surveys in the South Arran NC MPA. Accurate data should be collected from representative sites round the Scottish coast. Ideally these sites would include fished and non-fished sites at a range of latitudes and with a range of wave exposures. Importantly, data collection should also focus on the spatial footprint of the wrasse fishery.
- Habitat association and Priority Marine Features: There is a strong association between wrasse species and algae covered rocky reef habitat and an association with habitat Priority Marine Features (e.g. tide-swept algal communities, kelp and seaweed communities on sublittoral sediment, kelp beds). The authors conclude that wrasse are also typical species of features protected by the Habitats Directive. As such, impacts to these species are not consistent with the duty to protect these features. Impacts on wrasse within protected areas could affect site condition and impacts on rocky reef in or outwith protected areas could affect the Conservation Status of reef habitats. It will therefore be important to manage the impacts on wrasse as they could have wider implications on the status of protected sites and features.
- Monitoring: The sustainability of the wrasse fishery would benefit from a monitoring programme which will establish a baseline and be robust enough to detect changes in wrasse abundance over time. The monitoring programme will require on-going review and consultation with experts should be sought to ensure the sampling design is fit for purpose. As part of the site and feature condition monitoring requirement for the Scottish MPA network, monitoring the impacts on wrasse within protected areas should also be a priority. An effective monitoring programme must include fisheries independent surveys and observers to understand the relationship between catches, landings and discards as well as the reported landings data from fishers.
Future research priorities:
- Understand abundance and population structure of wrasse species. An important research priority will be to understand how changes in wrasse abundance and population affect ecosystem function.
- Implications of a wrasse fishery for protected sites and features. To achieve good site and feature condition within protected sites of the Scottish MPA network, it will be important to further research any interactions between the wrasse fishery and protected sites and features, including the effects of removing wrasse on ecosystem function and site condition.
- Understand the effects of wrasse fishing on wrasse breeding and recruitment. In temperate waters, wrasse are active in the spring, summer and autumn, resulting in fishing coinciding with breeding and recruitment. To properly manage the fishery, understanding the wider effects of extraction during these key times is essential.
- Confirmation of specifications for otter exclusion devices applicable to wrasse fishing. Otters are likely to be exposed to this fishery in shallow waters. To mitigate the risk of otter entrapment, an assessment of modifications to the fish traps is required. Including provision for on-board observers in the management of the fishery would be valuable in providing fishery-independent data on otter entrapment risk and efficacy of otter guard specification.
Conclusions
This report aims to provide a starting point for understanding the wider impacts of a wrasse fishery on protected sites and features and a methodology for the assessment and monitoring of the wild-capture wrasse fishery in Scotland. It is clear that our assessments of wrasse abundance are based on robust but limited data from one location on the west coast of Scotland. While these data are limited, the authors believe that enough information exists to draw conclusions about the importance of rocky reef and kelp habitat to wrasse. Data on habitat association and wrasse species are drawn both from analysis of baited camera data and existing literature. These data have also provided an insight into wrasse abundance at that site and into the issues and challenges of developing methodology to support the monitoring necessary for a sustainable fishery and to ensure the protection of relevant protected areas. The proposed monitoring framework provides a tool to aid assessment of wrasse abundance in Scottish waters.
Annexes
Annex 1: South Arran sample sites 2016
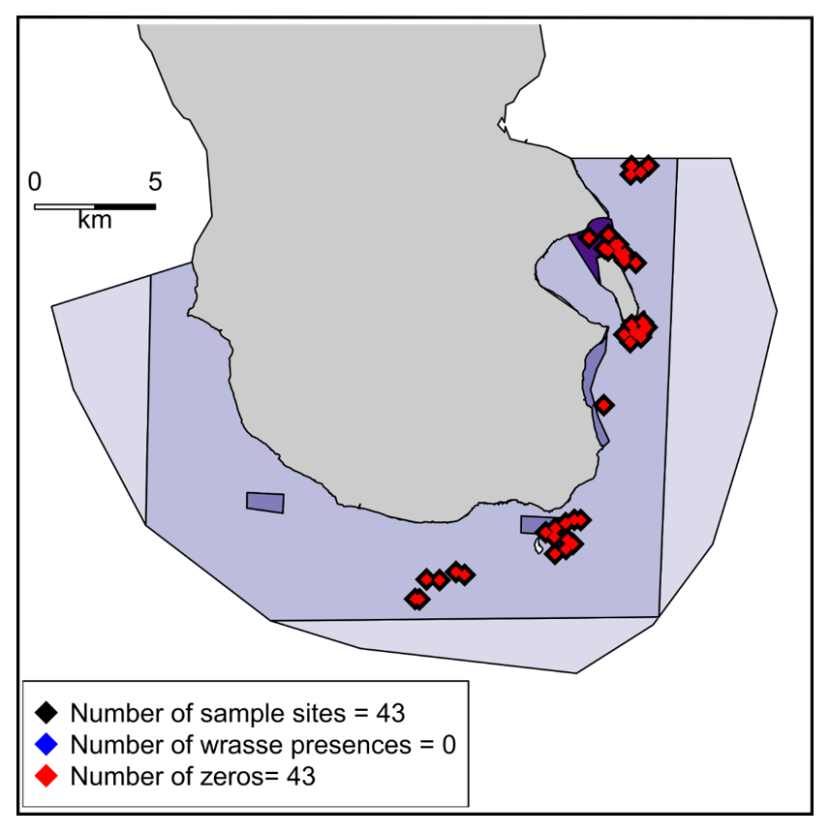
Figure A1. Stereo Baited Remote Underwater Video (SBRUV) deployment locations in the South Arran NC MPA in 2016.
Click for a full description
Map showing SBRUV deployment locations in 2016. The total number of deployments (46) is given, however no wrasse were observed in any deployments. Four levels of management measures are in place in the NC MPA depicted in purple (from light to dark: trawling by vessels <120 gross tonnage permitted; mobile demersal gear prohibited; most fishing prohibited; no take zone).
Annex 2: Summary of wrasse habitat association literature
| Habitat type | Wrasse species | Type of sampling/analysis (sample size) | Geographic range | Reference |
|---|---|---|---|---|
| Rocky shores. | L. bergylta | Angling (341) | Southern Isle of Mann | Dipper et al. 1977 |
| Kelp covered rocky bottom. | C. rupestris | Unbaited fyke nets & wrasse pots (3906) | Skagerrak | Halvorsen et al. 2017a |
| Kelp forests. | L. bergylta | Spear fishing (36) | Galicia, NW Spain | Pita and Freire 2019 |
| Rocky reefs and patches of sandy bottom. | L. bergylta | Photo ID (5) | Galicia, NW Spain | Mcientesu et al. 2019 |
| Rock and algal cover. | C. rupestris S. melops | Fishery using shrimp pots (26,820) | Ireland (NW) | Darwall et al. 1992b |
| Weed-covered rock pools. Rocky reef. | S. melops | Hand nets, push nets & angling (801) | Galway Bay, Ireland | Deady and Fives 1995a |
| Sheltered rocky areas (close to shore). | L. bergylta | Angling & trammel nets (293) | Galway Bay | Deady and Fives 1995b |
| Stones and rock with denser macroalgae cover and patches of sandy bottom. | C. rupestris | Beach seine (436) | Norwegian Skagerrak coast | Fjøsne and Gjøsœter 1996 |
| Rock in substratum increased number of fish. Vegetation is highly preferred compared to barren bottom. | C. rupestris | Beach seine (323) | Norwegian Skagerrak coast | Gjøsæter 2002a |
| Rocky bottom with dense macroalgae cover. | C. rupestris | Pots in commercial fishery (> 351,000) | Norwegian Skagerrak coast | Gjøsæter 2002b |
| Abundance increased with habitat complexity | C. rupestris C. melops | SCUBA visual census (234 surveys) | Scotland | Hunter and Sayer 2009 |
| Rocky bottom. Turbulent water (C. rupestris). Shallow protected areas (S. melops). Larger fish found in more exposed areas. | C. rupestris S. melops C. exoletus L. bergylta | Fyke net (55 stations caught 14, 384 wrasse) | Southwestern Norway | Skiftesvik et al. 2015 |
| Rocky bottoms covered by Laminaria and Fucus | C. rupestris | SCUBA (412 hours of observations) | Northwest Sweden | Hilldén 1981 |
| Rocky reef | C. rupestris L. bergylta C. exoletus S. melops L. mixtus | SCUBA transects | Scotland | Magill and Sayer 2002 |
| Non macroalgae covered artificial reef. | C. rupestris | SCUBA point count | NW Sweden, Skagerrak | Wilhelmsson et al. 2006 |
| Kelp covered and rocky bottom. | C. rupestris S. melops | Fyke nets & wrasse pots | Southern Norway | Halvorsen et al. 2017b |
| Inshore, rocky algal areas. | All species particularly C. rupestris C. exoletus | Creel and fyke net (1546) | West coast of Scotland | Treasurer 1994 |
| Equally among rock, cobbles and large pebbles. | C. rupestris | SCUBA quadrat (164 sampling stations) | North Sea | Wehkamp and Fischer 2013 |
| Rocky with refuges. | C. rupestris | SCUBA survey (21 sites) | Scottish west coast | Sayer et al. 1993 |
| Algae covered reef. | S. melops | SCUBA diving visual surveys (144 fish) | Portugal | Rodrigues et al. 2015 |
| Rocky-bottom habitats (rather than soft sedement) | C. rupestris S. melops | gill-net and SCUBA transect (243 sample sites) | West coast Sweden | (Stål et al. 2007) |
| Rock boulder scree with algal cover | C. rupestris | SCUBA – 12 tagged fish | West coast Scotland | (Sayer 1999) |
| Serpula vermicularis reefs fringing shores (6 -10 m, max 14 m) | L. bergylta C. exoletus C. rupestris C. melops | underwater television (4 weeks, 1 site) | West coast of Scotland (Loch Creran) | (Poloczanska et al. 2004) |
| vegetated or rocky substrate | C. rupestris | un-baited fyke nets and two-chamber pots (935) | Norwegian Skagerrak coast | (Olsen et al. 2019) |
| Rocky reefs. | L. bergylta | Citation of Quignard et al, 1986 | - | Muncaster et al. 2010* |
| Coastal rocky bottoms. | L. bergylta | Citation of Rodríguez and Vázquez, 1994 | - | Pita and Freire 2011* |
| Rocky reefs. | L. bergylta | Citation of Quignard et al, 1994 | - | Brooker et al. 2018* |
| rocky reefs and kelp beds. | L. bergylta | Citation of Figueiredo et al., 2005 | - | Villegas-Ríos et al. 2013* |
| Rocky reefs and kelp beds. | L. bergylta | Citation of Figueiredo et al., 2005 | - | Mcientesu et al. 2019* |
| Rocky habitats. | All species | Citation of Darwall et al., 1992b; Sayer et al., 1994 | UK | Deady and Fives 1995a* |
| Coastal waters in areas with rock and algal cover. | All species | Citation of Wheeler, 1978; Quignard and Pras, 1986. | Irish waters | Deady et al. 1995* |
| Rocky shores down to depths of 50 m. | C. rupestris | Citation of Quignard and Pras, 1986; Sayer et al., 1996a | - | Skiftesvik et al. 2015* |
| High densities in areas of turbulent water. | - | Citation of Hillden, 1978; Sayer et al., 1993; Gjøsæter, 2002 | - | Skiftesvik et al. 2015* |
| Suitable refuge is factor limiting as this species remains in refugia for long periods. | - | Citation of Sayer et al., 1993; Sayer, 1999 | - | Skiftesvik et al. 2015* |
| Rocky reefs covered by algae or in seagrass meadows. | L. mixtus | Citation of Muus and Nielsen 1999 | - | Matić-Skoko et al. 2013* |
| Rocky, algal inshore areas. | All species | Citation of Costello 1991 | UK | Treasurer 1994* |
| Rocky shores, often close to refuge holes Caught rarely over sand | C. rupestris | Citation of Costello, 1991; Sayer et al., 1993 & Wheeler, 1969; Sayer & Treasurer, 1996 | - | Sayer 1999* |
| Predators on Serpula vermicularis biogenic reefs | C. rupestris C. melops. | Citation of Bosence 1979 | West coast Scotland and West coast Ireland | Poloczanska et al. 2004* |
| high algal cover, e.g. kelp forests and eel-grass beds | C. melops. | Citation of Quignard & Pras 1986; Lythgoe & Lythgoe 1991 | - | Skiftesvik et al. 2014* |
| sheltered or exposed rocky shores, mudflats and kelp forests | C. rupestris C. melops | Citation of Thangstad 1999 | - | Skiftesvik et al. 2014* |
| shallow, rocky coastal habitats | C. melops | Citation of Quignard and Pras, 1986; Darwall et al., 1992b; Sayer and Treasurer, 1996 | - | Halvorsen et al. 2017b* |
| nearshore-demersal, most abundant in shallow coastal rocky reefs and kelp beds. Overnight in rock crevices. | L. bergylta | Citation of Dipper et al., 1977; Costello, 1991 | - | Leclercq et al. 2018* |
| rocky- or vegetated substrates with access to refuges | C. rupestris | Citation of (Costello, 1991; Sayer et al., 1993; Gjøsæter, 2002a | - | Olsen et al. 2019* |
Annex 3: Biological comparative table
| Wrasse species name | Symphodus melops | Ctenolabrus rupestris | Labrus bergylta | Labrus mixtus | Total |
|---|---|---|---|---|---|
| Biotope Code (EUNIS level) | - | - | - | - | - |
| IR.HIR.KFaR.LhypFa (5) | - | - | 26 | - | 26 |
| IR.HIR.KFaR.LhypPar (5) | 100 | - | 100 | 100 | 300 |
| IR.HIR.KFaR.LhypR (5) | - | - | 24 | - | 24 |
| IR.HIR.KFaR.LhypR.Ft (6) | - | - | 23 | - | 23 |
| IR.HIR.KFaR.LhypR.Pk (6) | - | - | 23 | - | 23 |
| IR.HIR.KFaR.LhypR.Loch (6) | - | - | 38 | - | 38 |
| IR.HIR.KFaR.FoR (5) | - | 26 | 33 | 26 | 85 |
| IR.HIR.KFaR.FoR.Dic (6) | - | 36 | 43 | 33 | 112 |
| IR.HIR.KFaR.LhypRVt (5) | - | - | 29 | - | 29 |
| IR.HIR.KSed.LsacSac (5) | - | - | 29 | - | 29 |
| IR.HIR.KSed.LsacChoR (5) | 30 | - | - | - | 30 |
| IR.HIR.KSed.XKScrR (5) | - | - | 22 | - | 22 |
| IR.HIR.KSed.XKHal (5) | 36 | 21 | - | - | 57 |
| IR.MIR.KR.LhypT (5) | - | - | 33 | - | 33 |
| IR.MIR.KR.LhypT.Ft (6) | - | - | 26 | - | 26 |
| IR.MIR.KR.LhypT.Pk (6) | - | 23 | 46 | - | 69 |
| IR.MIR.KR.HiaSw (5) | 31 | 44 | 63 | - | 138 |
| IR.MIR.KT.FilRVS (5) | - | 22 | - | - | 22 |
| IR.LIR.K.LhypLoch (5) | - | - | 44 | - | 44 |
| IR.LIR.K.LhypLsac.Pk (6) | - | 22 | - | - | 22 |
| IR.LIR.K.Sar (5) | - | 33 | 33 | - | 66 |
| CR (2) | - | 23 | 23 | - | 46 |
| CR.HCR (3) | - | 36 | 43 | 29 | 108 |
| CR.HCR.FaT (4) | - | 28 | 41 | - | 69 |
| CR.HCR.FaT.BalTub (5) | - | - | 25 | - | 25 |
| CR.HCR.FaT.CTub (5) | - | 33 | 48 | - | 81 |
| CR.HCR.FaT.CTub.CuSp (6) | - | 55 | 55 | - | 110 |
| CR.HCR.FaT.CTub.Adig (6) | - | - | 40 | - | 40 |
| CR.HCR.DpSp (4) | - | 50 | 57 | 71 | 178 |
| CR.HCR.DpSp.PhaAxi (5) | - | 50 | 57 | 71 | 178 |
| CR.HCR.XFa (4) | - | 35 | 42 | 26 | 103 |
| CR.HCR.XFa.ByErSp (5) | - | 51 | 65 | 36 | 152 |
| CR.HCR.XFa.ByErSp.Eun (6) | - | 55 | 60 | 65 | 180 |
| CR.HCR.XFa.ByErSp.DysAct (6) | - | 53 | 67 | 22 | 142 |
| CR.HCR.XFa.ByErSp.Sag (6) | - | 46 | 66 | 37 | 149 |
| CR.HCR.XFa.CvirCri (5) | 32 | 68 | 79 | 63 | 242 |
| CR.HCR.XFa.SwiLgAs (5) | - | 30 | 40 | - | 70 |
| CR.HCR.XFa.FluCoAs (5) | - | 25 | - | - | 25 |
| CR.HCR.XFa.FluCoAs.SmAs (6) | - | 45 | 25 | - | 70 |
| CR.HCR.XFa.FluCoAs.X (6) | - | 40 | - | - | 40 |
| CR.HCR.XFa.SpNemAdia (5) | - | - | - | 27 | 27 |
| CR.HCR.XFa.SubCriTf (5) | - | 50 | 50 | - | 100 |
| CR.HCR.XFa.SpAnVt (5) | - | 29 | 41 | 31 | 101 |
| CR.MCR.EcCr.CarSwi (5) | - | 27 | - | 32 | 59 |
| CR.MCR.EcCr.CarSwi.Aglo (6) | - | 29 | 29 | 29 | 87 |
| CR.MCR.EcCr.CarSwi.LgAs (6) | - | 27 | - | 33 | 60 |
| CR.MCR.EcCr.CarSp (5) | - | 43 | 51 | 49 | 143 |
| CR.MCR.EcCr.CarSp.Bri (6) | - | 33 | - | - | 33 |
| CR.MCR.EcCr.CarSp.PenPcom (6) | - | 44 | 57 | 56 | 157 |
| CR.MCR.EcCr.FaAlCr.Sec (6) | - | 23 | - | - | 23 |
| CR.MCR.EcCr.FaAlCr.Car (6) | - | 26 | - | 21 | 47 |
| CR.MCR.SfR.Pid (5) | - | - | 31 | - | 31 |
| CR.MCR.CMus.Mdis (5) | - | - | 21 | - | 21 |
| CR.MCR.CFaVS (4) | - | 42 | - | - | 42 |
| CR.MCR.CFaVS.CuSpH (5) | - | 48 | - | - | 48 |
| CR.MCR.CFaVS.CuSpH.As (6) | - | 52 | - | - | 52 |
| CR.MCR.CFaVS.CuSpH.VS (6) | - | 45 | - | - | 45 |
| CR.LCR.BrAs.LgAsSp (5) | 25 | 65 | 45 | 35 | 170 |
| CR.FCR (3) | - | 22 | - | - | 22 |
| CR.FCR.FouFa (4) | - | 30 | - | - | 30 |
| CR.FCR.FouFa.AdigMsen (5) | - | 31 | - | - | 31 |
| CR.FCR.FouFa.Aasp (5) | - | 29 | - | - | 29 |
References
Anderson, M.J., and Millar, R.B. 2004. Spatial variation and effects of habitat on temperate reef fish assemblages in northeastern New Zealand. Journal of Experimental Marine Biology and Ecology, 305, 191–221.
Baden, S., Boström, C., Tobiasson, S., Arponen, H. and Moksnes, P.O. 2010. Relative importance of trophic interactions and nutrient enrichment in seagrass ecosystems: A broad-scale field experiment in the Baltic-Skagerrak area. Limnology and Oceanography, 55(3), 1435–1448.
Bailey, N., Bailey, D., Bellini, L., Fernandes, P., Fox, C., Heymans, S., Holmes, S., Howe, J., Hughes, S, Magill, S., McIntyre, F., McKee, D., Ryan, M., Smith, I., Tyldsley, G., Watret, R. and Turrell, W. 2011. The West of Scotland Marine Ecosystem : A Review of Scientific Knowledge. Marine Scotland Science Report.
Bell, J. D., and Harmelin-Vivien, M. 1983. Fish fauna of French Mediterranean Posidonia oceanica seagrass meadows. II: feeding habits. Tethys, 11, 1–14.
Benaka, L.R. 1999. Fish Habitat: Essential Fish Habitat and Rehabilitation. American Fisheries Society, Symposium 22.
Benjamin, J., and Mackintosh, R. 2016. Regulating Scientific Diving and Underwater Archaeology: Legal and historical considerations. International Journal of Nautical Archaeology, 45(1), 153–169.
Bertelli, C.M., and Unsworth, R.K.F. 2014. Protecting the hand that feeds us: Seagrass (Zostera marina) serves as commercial juvenile fish habitat. Marine Pollution Bulletin.
Bjordal, Å. 1988. Cleaning symbiosis between wrasse (Labridae) and lice infested salmon (Salmo salar) in mariculture. International Council for the Exploration of the Sea, Mariculture Committee.
Bjordal, Å. 1991. Wrasse as cleaner fish for farmed salmon. Prog. Underwater Sci, 16, 17–28.
Blanco Gonzalez, E., and de Boer, F. 2017. The development of the Norwegian wrasse fishery and the use of wrasses as cleaner fish in the salmon aquaculture industry. Springer Tokyo.
Brooker, A. J., Papadopoulou, A., Gutierrez, C., Rey, S., Davie, A. and Migaud, H. 2018. Sustainable production and use of cleaner fish for the biological control of sea lice: Recent advances and current challenges. Veterinary Record, 183, 383.
Burns, N. M., Bailey, D.M. and Wright, P. J. 2019. A method to improve fishing selectivity through age targeted fishing using life stage distribution modelling. PLoS ONE, 14(4), e0214459.
Burrows, M.T., Hughes, D. J., Austin, W.E.N., Smeaton, C., Hicks, N., Howe, J.A., Allen, C., Taylor, P., and Vare, L.L. 2017. Assessment of Blue Carbon Resources in Scotland’s Inshore Marine Protected Area Network. Scottish Natural Heritage Commissioned Report 957.
Cappo, M., De’ath, G., Boyle, S., Aumend, J., Olbrich, R., Hoedt, F., Perna, C. and Brunskill, G. 2005. Development of a robust classifier of freshwater residence in barramundi (Lates calcarifer) life histories using elemental ratios in scales and boosted regression trees. Marine and Freshwater Research, 56, 713–723.
Clavero, M., Prenda, J. and Delibes, M. 2006. Seasonal use of coastal resources by otters: Comparing sandy and rocky stretches. Estuarine, Coastal and Shelf Science, 66, 387–394.
Costello, M. J. 1991. Review of the biology of wrasse (Labridae: Pisces) in Northern Europe. Progress in Underwater Science, 16, 29–51.
D’Arcy, J., Dunaevskaya, E., Treasurer, J.W., Ottesen, O., Maguire, J., Zhuravleva, N., Karlsen, A., Rebours, C. and Fitzgerald, R.D. 2012. Embryonic development in ballan wrasse Labrus bergylta. Journal of Fish Biology, 81(3), 1101–1110.
Damalas, D., Maravelias, C.D., Katsanevakis, S., Karageorgis, A.P. and Papaconstantinou, C. 2010. Seasonal abundance of non-commercial demersal fish in the eastern Mediterranean Sea in relation to hydrographic and sediment characteristics. Estuarine, Coastal and Shelf Science, 89(1), 107–118.
Darwall, W., Costello, M.J. and Lysaght, S. 1992a. Wrasse – how well do they work? Aquaculture Ireland, 50, 26–29.
Darwall, W.R.T., Costello, M. J., Donnelly, R., and Lysaght, S. 1992b. Implications of life‐history strategies for a new wrasse fishery. Journal of Fish Biology, 41, 111–123.
Davies, C.E., Moss, D. and Hill, M.O. 2004. EUNIS Habitat Classification Revised 2004
Dayton, P.K. 1985. Ecology of kelp communities. Annual review of ecology and systematics, 16, 215–245.
Deady, S., and Fives, J.M. 1995a. The diet of corkwing wrasse, Crenilabrlis Melops, in Galway Bay, Ireland, and in Dinard, France. Journal of the Marine Biological Association of the United Kingdom, 75, 635–649.
Deady, S., and Fives, J.M. 1995b. Diet of ballan wrasse, Labrus Bergylta, and some comparisons with the diet of corkwing wrasse, Crenilabrus Melops. Journal of the Marine Biological Association of the United Kingdom, 75, 651–665.
Deady, S., Varian, S.J.A. and Fives, J.M. 1995. The use of cleaner-fish to control sea lice on two Irish salmon (Salmo salar) farms with particular reference to wrasse behaviour in salmon cages. Aquaculture, 131, 73–90.
Devon & Severn IFCA, 2017. Management of the Live Wrasse Fishery. Potting Permit Byelaw. Management of the ‘live’ wrasse pot fishery. Devon & Severn Inshore Fisheries and Conservation Authority, 1–44.
Dickens, L.C., Goatley, C.H.R., Tanner, J.K. and Bellwood, D.R. 2011. Quantifying relative diver effects in underwater visual censuses. PLoS ONE, 6(4), e18965.
Dipper, F.A., Bridges, C.R., and Menz, A. 1977. Age, growth and feeding in the ballan wrasse Labrus bergylta Ascanius 1767. Journal of Fish Biology, 11, 105–120.
Elliott, S.A.M., Ahti, P.A., Heath, M.R., Turrell, W.R. and Bailey, D.M. 2016. An assessment of juvenile Atlantic cod Gadus morhua distribution and growth using diver operated stereo-video surveys. Journal of Fish Biology, 89(2), 1190-1207.
Elliott, S.A.M., Guérin, L., Pesch, R., Schmitt, P., Meakins, B., Vina-Herbon, C., González-Irusta, J.M., de la Torriente, A. and Serrano, A. 2018. Integrating benthic habitat indicators: Working towards an ecosystem approach. Marine Policy, 90, 88–94.
Elliott, S.A.M., Sabatino, A.D., Heath, M.R., Turrell, W.R. and Bailey, D.M. 2017a. Landscape effects on demersal fish revealed by field observations and predictive seabed modelling. PLoS ONE, 12(12), e0189011.
Elliott, S.A.M., Turrell, W.R., Heath, M.R. and Bailey, D.M. 2017b. Juvenile gadoid habitat and ontogenetic shift observations using stereo-video baited cameras. Marine Ecology Progress Series, 568, 123–135.
Eno, N.C., MacDonald, D.S., Kinnear, J.A.M., Amos, S.C., Chapman, C. J., Clark, R.A., Bunker, F.S.P.D. and Munro, C. 2001. Effects of crustacean traps on benthic fauna. ICES Journal of Marine Science, 58(1), 11-20.
Environment Agency, 2017. Permission to trap crayfish, eels, elvers, salmon and sea trout
Faust, E., Halvorsen, K.T., Andersen, P., Knutsen, H. and André, C. 2018. Cleaner fish escape salmon farms and hybridize with local wrasse populations. Royal Society Open Science, 5, 171752.
Findlay, M., Alexander, L. and Macleod, C. 2015. Site condition monitoring for otters (Lutra lutra) in 2011-12. Scottish Natural Heritage Commissioned Report 521.
Fipec Industries, 2019. Fipec Industries
Fitzpatrick, B.M., Harvey, E.S., Heyward, A.J., Twiggs, E.J. andColquhoun, J. 2012. Habitat specialization in tropical continental shelf demersal fish assemblages. PLoS ONE, 7(6), e39634.
Fjøsne, K., and Gjøsœter, J. 1996. Dietary composition and the potential of food competition between 0- group cod (Gadus morhua L.) and some other fish species in the littoral zone. ICES Journal of Marine Science, 53, 757–770.
García-Rubies, A., and Macpherson, E. 1995. Substrate use and temporal pattern of recruitment in juvenile fishes of the Mediterranean littoral. Marine Biology, 124, 35–42.
Gjøsæter, J. 2002a. Fishery for goldsinny wrasse (Ctenolabrus rupestris) (Labridae) with pots along the Norwegian Skagerrak coast. Sarsia, 87, 83–89.
Gjøsæter, J. 2002b. Distribution and density of goldsinny wrasse (Ctenolabrus rupestris) (Labridae) in the Risør and Arendal areas along the Norwegian Skagerrak coast. Sarsia, 87, 75–81.
Greenstreet, S.P.R., Rossberg, A.G., Fox, C.J., Le Quesne, W.J.F., Blasdale, T., Boulcott, P., Mitchell, I., Millar, C., and Moffat, C.F. 2012. Demersal fish biodiversity: Species-level indicators and trends-based targets for the Marine Strategy Framework Directive. ICES Journal of Marine Science, 69(10), 1789–1801.
Halvorsen, K.T.,Larsen, T., Sørdalen, T.K., Vøllestad, L.A., Knutsen, H. and Olsen, E.M. 2017a. Impact of harvesting cleaner fish for salmonid aquaculture assessed from replicated coastal marine protected areas. Marine Biology Research, 13, 359–369.
Halvorsen, K.T., Sørdalen, T.K., Vøllestad, L.A., Skiftesvik, A.B., Espeland, S.H. and Olsen, E.M. 2017b. Sex-and size-selective harvesting of corkwing wrasse (Symphodus melops)-a cleaner fish used in salmonid aquaculture. ICES Journal of Marine Science, 74, 660–669.
Hamre, K., Nordgreen,A., Grøtan, E. and Breck, O. 2013. A holistic approach to development of diets for ballan wrasse (Labrus berggylta) - A new species in aquaculture. PeerJ, 2013:e99.
Harvey, E.S., and Shortis, M.R. 1998. Calibration Stability of an Underwater Stereo-Video System: Implications for Measurement Accuracy and Precision. Marine Technology Society Journal, 32(2), 1–26.
Harvey, E., and Shortis, M. 1995. A system for stereo-video measurement of sub-tidal organisms. Marine Technology Society Journal, 29(4),10–22.
Health and Safety Executive, 2014. Scientific and archaeological diving projects. Pages 1–29. UK.
Heggberget, T. M. 1993. Marine-feeding otters (Lutra lutra) in Norway: Seasonal variation in prey and reproductive timing. Journal of the Marine Biological Association of the United Kingdom, 73, 297–312.
Heuch, P.A., Bjørn, P.A., Finstad, B., Holst, J.C., Asplin, L. and Nilsen, F. 2005. A review of the Norwegian “National Action Plan Against Salmon Lice on Salmonids”: The effect on wild salmonids. Aquaculture, 246, 79–92.
Hilldén, N.O. 1984. Behavioural ecology of the labrid fishes (Teleostei: labridae) at Tjärnöon the Swedish west coast. University of Stockholm.
Hilldén, N.O. 1981. Territoriality and reproductive behaviour in the goldsinny, Ctenolabrus rupestris L. Behavioural Processes, 6, 207–221.
Hiscock, K., Southward, A., Tittley, I. and Hawkins. S. 2004. Effects of changing temperature on benthic marine life in Britain and Ireland. Aquatic Conservation: Marine and Freshwater Ecosystems, 14, 333–362.
Hunter, W.R., and Sayer, M.D.J. 2009. The comparative effects of habitat complexity on faunal assemblages of northern temperate artificial and natural reefs. ICES Journal of Marine Science, 66, 691–698.
Imsland, A.K., Reynolds, G. Eliassen, Hangstad, T.A., Foss, A., Vikingstad, E. and Elvegård, T.A. 2014. The use of lumpfish (Cyclopterus lumpus L.) to control sea lice (Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon (Salmo salar L.). Aquaculture, 424–425, 18–23.
Imsland, A. K., P. Reynolds, P., Eliassen, G., Hangstad, T.A., Jónsdóttir, Ó.D.B., T. A. Elvegård, Ó.D.B., Lemmens, S.C.A., Rydland, R. and Nytrø, A.V. 2016. Investigation of behavioural interactions between lumpfish (Cyclopterus lumpus) and goldsinny wrasse (Ctenolabrus rupestris) under controlled conditions. Aquaculture International, 24, 1509–1521.
Jackson, E.L., Attrill, M.J. and Jones, M.B. 2006. Habitat characteristics and spatial arrangement affecting the diversity of fish and decapod assemblages of seagrass (Zostera marina) beds around the coast of Jersey (English Channel). Estuarine, Coastal and Shelf Science, 68, 421–432.
JNCC, 2018. The Marine Habitat Classification for Britain and Ireland
Johnson, A.F., Jenkins, S.R, Hiddink, J.G. and Hinz, H. 2013. Linking temperate demersal fish species to habitat: Scales, patterns and future directions. Fish and Fisheries, 14(3), 256–280.
Jones, A.C., Mead, A., Kaiser, M.J., Austen, M. C.V., Adrian, A.W., Auchterlonie, N.A., Black, K.D., Blow, L.R., Bury, C., Brown, J.H., Burnell, G.M., Connolly, E., Dingwall, A., Derrick, S., Eno, N.C., Gautier, D.J.H., Green, K.A., Gubbins, M., Hart, P.R., ... Sutherland, W.J. 2015. Prioritization of knowledge needs for sustainable aquaculture: A national and global perspective. Fish and Fisheries, 16(4), 668-683.
Jones, S., and Johnson, S. 2015. Sea lice monitoring and non-chemical measures. Page DFO Can. Sci. Advis. Sec. Sci. Advis. Rep.
Keats, D.W., and Steele, D.H. 1992. Diurnal feeding of juvenile cod (Gadus morhua) which migrate into shallow water at night in eastern Newfoundland. Journal of Northwest Atlantic Fishery Science, 13, 7-14.
Kimmel, J. J. 1985. A new species-time method for visual assessment of fishes and its comparison with established methods. Environmental Biology of Fishes, 12, 23–32.
Kvenseth, P.G. 1996. Large-scale use of wrasse to control sea lice and net fouling in salmon farms in Norway. Fishing News Books Ltd.
Leclercq, E., Davie, A. and Migaud, H. 2014a. Delousing efficiency of farmed ballan wrasse (Labrus bergylta) against Lepeophtheirus salmonis infecting Atlantic salmon (Salmo salar) post-smolts. Pest Management Science, 70, 1274–1282.
Leclercq, E., Davie, A. and Migaud, H. 2014b. The physiological response of farmed ballan wrasse (Labrus bergylta) exposed to an acute stressor. Aquaculture, 434, 1–4.
Leclercq, E., Zerafa, B., Brooker, A.J., Davie, A. and Migaud, H. 2018. Application of passive-acoustic telemetry to explore the behaviour of ballan wrasse (Labrus bergylta) and lumpfish (Cyclopterus lumpus) in commercial Scottish salmon sea-pens. Aquaculture, 495, 1–12.
Magill, S.H., and Sayer, M.D.J. 2002. Seasonal and interannual variation in fish assemblages of northern temperate rocky subtidal habitats. Journal of Fish Biology, 61, 1198–1216.
Marine Scotland, 2017. Scottish Marine Protected Areas (MPA) Monitoring Strategy.
Marine Scotland, 2018. Voluntary control measures for the live capture of Scottish wild wrasse for salmon farms.
Matić-Skoko, S., Bojanić Varezić, D., Šiljić, J., Tutman, P. and Pallaoro, A. 2013. The cuckoo wrasse, Labrus mixtus (Pisces: Labridae): biological indices for life history and conservation. Scientia Marina, 77, 595–605.
Merrett, N.R., Bagley, P. M., Smith, A., and Creasey, S. 1994. Scavenging Deep Demersal Fishes of the Porcupine Seabight, North-East Atlantic: Observations by Baited Camera, Trap and Trawl. Journal of the Marine Biological Association of the United Kingdom, 74(3), 481–498.
Moksnes, P.O., Gullström, M., Tryman, K. and Baden, S. 2008. Trophic cascades in a temperate seagrass community. Oikos, 117(5), 763–777
Mucientes, G., Irisarri, J. and Villegas-Ríos, D. 2019. Interannual fine-scale site fidelity of male ballan wrasse Labrus bergylta revealed by photo-identification and tagging. Journal of Fish Biology, 95, 1151–1155.
Muncaster, S., Andersson, E., Kjesbu, O.S., Taranger, G.L., Skiftesvik, A.B. and Norberg, B. 2010. The reproductive cycle of female Ballan wrasse Labrus bergylta in high latitude, temperate waters. Journal of Fish Biology, 77, 494–511.
Munro, L., and Wallace, I. 2017. Scottish Fish Farm Production Survey 2016. Edinburgh.
Murray, A.G. 2017. A model of the process of spillover and adaption leading to potential emergence of disease in salmon held with cleaner fish used to control lice. Aquaculture, 473, 283–290.
NatureScot, n.d. Marine Protected Areas (MPAs)
Norwegian Directorate of Fisheries, 2009. Key figures from Norwegian aquaculture industry.
Olsen, E.M., Halvorsen, K.T., Larsen, T. and Kuparinen, A. 2019. Potential for managing life history diversity in a commercially exploited intermediate predator, the goldsinny wrasse (Ctenolabrus rupestris). ICES Journal of Marine Science, 76, 410–417.
Oppedal, F., Dempster, T. and Stien, L.H. 2011. Environmental drivers of Atlantic salmon behaviour in sea-cages: A review. Aquaculture, 311, 1–18.
Ottesen, O.H., Dunaevskaya, E. and Arcy, J.D. 2012. Development of Labrus bergylta (Ascanius 1767) larvae from hatching to metamorphosis. Journal of Aquaculture Research and Development, 3, 1–4.
Piccinetti, C.C., Grasso, L., Maradonna, F., Radaelli, G., Ballarin, C., Chemello, G., Evjemo, J.O., Carnevali, O., and Olivotto, I. 2017. Growth and stress factors in ballan wrasse (Labrus bergylta) larval development. Aquaculture Research, 48, 2567–2580.
Pihl, L., Wennhage, H. and Nilsson, S. 1994. Fish assemblage structure in relation to macrophytes and filamentous epiphytes in shallow non-tidal rocky- and soft-bottom habitats. Environmental Biology of Fishes, 39, 271–288.
Pita, P., and Freire, J. 2011. Movements of three large coastal predatory fishes in the northeast Atlantic: a preliminary telemetry study. Scientia Marina, 75, 759–770.
Pita, P., and Freire, J. 2019. Trophic ecology of an Atlantic kelp forest fish assemblage (NW Spain) targeted by recreational fishers and implications for coastal management. Journal of the Marine Biological Association of the United Kingdom, 99, 19–29.
Poloczanska, E.S., Hughes, D.J. and Burrows, M.T. 2004. Underwater television observations of Serpula vermicularis (L.) reefs and associated mobile fauna in Loch Creran, Scotland. Estuarine, Coastal and Shelf Science, 61, 425–435.
Potts, G.W. 1974. The colouration and its behavioural significance in the corkwing wrasse, Crenilabrus melops. Journal of the Marine Biological Association of the United Kingdom, 54, 925–938.
Priede, I.G., Bagley, P.M., Smith, A. and Creasey, S. 1994. Scavenging deep demersal fishes of the Porcupine Seabight, north-east Atlantic: observations by baited camera, trap and trawl. Journal of the Marine Biological Association of the United Kingdom, 74(3), 481–498.
Quignard, J., and Pras, A. 1986. Labridae. pp 919–942, In: P.J.P. Whitehead, M.L. Bauchot, J.C. Hureau, J. Nielsen, and E. Tortonese, eds. Fishes of the North-eastern Atlantic and the Mediterranean. UNESCO, Paris.
R Core Team, 2019. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rae, G.H. 2002. Sea louse control in Scotland, past and present. Pest Management Science, 58, 515–520.
Riley, A., Jeffery, K., Cochrane-Dyet, T., White, P. and Ellis, J. 2017. Northern European wrasse – summary of commercial use, fisheries and implications for management.
Rodrigues, D., Horta e Costa, B., Cabral, H.N., and Gonçalves, E.J. 2015. Habitat associations and behavioural patterns of Symphodus spp. (Pisces: Labridae). Acta Ethologica, 18, 269–282.
Ruitton, S., P. Francour, and C. F. Boudouresque. 2000. Relationships between algae, benthic herbivorous invertebrates and fishes in rocky sublittoral communities of a temperate sea (Mediterranean). Estuarine, Coastal and Shelf Science, 50(2), 217–230.
Ryan, M.R., and Bailey, D.M. 2012. Trawling and Dredging in the Clyde Sea Area : History, Impacts and Prospects for Recovery University of Glasgow.
Sayer, M.D.J. 1999. Duration of refuge residence by goldsinny, Ctenolabrus rupestris. Journal of the Marine Biological Association of the United Kingdom, 79, 571–572.
Sayer, M.D.J., Gibson, R.N. and Atkinson, R.J.A. 1993. Distribution and density of populations of goldsinny wrasse (Ctenolabrus rupestris) on the west coast of Scotland. Journal of Fish Biology, 43, 157–167.
Sayer, M., and Treasurer, J. 1996. North European wrasse: identification, distribution and habitat. pp 3–12. In: M. Sayer, J. Treasurer, and M. Costello, eds. Wrasse: biology and use in aquaculture. Blackwell Science Inc, Oxford, UK.
SeaGIS, 2013. SeaGIS
SEPA, 2019. Protection of the Marine Environment Discharges from Marine Pen Fish Farms: A Strengthened Regulatory Framework.
Shester, G.G., and Micheli, F. 2011. Conservation challenges for small-scale fisheries: Bycatch and habitat impacts of traps and gillnets. Biological Conservation, 144(5), 1673–1681.
Skiftesvik, A.B., Bjelland, R.M., Durif, C.M.F., Johansen, I.S., and Browman, H.I. 2013. Delousing of Atlantic salmon (Salmo salar) by cultured vs. wild ballan wrasse (Labrus bergylta). Aquaculture, 402–403, 113–118.
Skiftesvik, A.B., Blom, G., Agnalt, A.L., Durif, C.M.F., Browman, H.I., Bjelland, R.M., Harkestad, L.S., Farestveit, E., Paulsen, O.I., Fauske, M., Havelin, T., Johnsen, K., and Mortensen, S. 2014. Wrasse (Labridae) as cleaner fish in salmonid aquaculture - The Hardangerfjord as a case study. Marine Biology Research, 10, 289–300.
Skiftesvik, A.B., Boxaspen, K., and Parsons, A. 1996. Preliminary breeding trials of wrasse in an intensive system. pp 136–141. In: M. D. J. Sayer, J. W. Treasurer, and M. J. Costello, eds. Wrasse: Biology and Use in Aquaculture. Blackwell.
Skiftesvik, A.B., Durif, C.M.F., Bjelland, R.M., and Browman, H.I. 2015. Distribution and habitat preferences of five species of wrasse (family Labridae) in a Norwegian fjord. ICES Journal of Marine Science, 72, 890–899.
Smale, D.A., Burrows, M.T., Moore, P., O’Connor, N., and Hawkins, S.J. 2013. Threats and knowledge gaps for ecosystem services provided by kelp forests: A northeast Atlantic perspective. Ecology and Evolution, 3(11), 4016–4038.
SSPO, 2018. Sustainable use of wild wrasse to help salmon farmers manage sea lice
Stål, J., Pihl, L., and Wennhage, H. 2007. Food utilisation by coastal fish assemblages in rocky and soft bottoms on the Swedish west coast: Inference for identification of essential fish habitats. Estuarine, Coastal and Shelf Science, 71, 93–607.
Stone, J. 1996. Preliminary trials on the culture of goldsinny and corkwing wrasse. pp142–170. In: M.D.J. Sayer, J.W. Treasurer, and M.J. Costello, eds. Wrasse biology and aquaculture applications. Blackwell.
The Conservation (Natural Habitats, &c.) Regulations 1994
The European Union Reference Laboratory, 2016. EURL fish and crustacean diseases
The Scottish Government, 2014. South Arran Nature Conservation Marine Protected Area Order.
The Scottish Government, 2019. Future of Fisheries Management in Scotland: National Discussion Paper.
Todd, C.D. 2007. The copepod parasite (Lepeophtheirus salmonis (Krøyer), Caligus elongates Nordmann) interactions between wild and farmed Atlantic salmon (Salmo salar L.) and wild sea trout (Salmo trutta L.): A mini review. Journal of Plankton Research, 29. i61–i71.
Torrissen, O., Jones, S., Asche, F., Guttormsen, A., Skilbrei, O.T., Nilsen, F., Horsberg, T.E. and Jackson, D. 2013. Salmon lice - impact on wild salmonids and salmon aquaculture. Journal of Fish Diseases, 36, 171–194.
Treasurer, J. 2013. Use of wrasse in sea lice control. Ardtoe Marine Laboratory.
Treasurer, J.W. 1993. Management of sea lice (Caligidae) with wrasse (Labridae) on Atlantic salmon (Salmo salar) farms. pp 335–345. In: G.A. Boxshall and D. Defaye, eds. Pathogens of Wild and Farmed Salmonids: Sea Lice. Ellis Horwood, Chichester.
Treasurer, J.W. 1994. The distribution, age and growth of wrasse (Labridae) in inshore waters of west Scotland. Journal of Fish Biology, 44, 905–918.
Treasurer, J.W. 2002. A review of potential pathogens of sea lice and the application of cleaner fish in biological control. Pest Management Science, 58, 546–558.
Treasurer, J.W. 2018. Cleaner fish biology and aquaculture applications. M5 Publishing, Sheffield.
Tully, O., Daly, P.,Lysaght, S., Deady, S., and Varian, S.J.A. 1996. Use of cleaner-wrasse (Centrolabrus exoletus (L.) and Ctenolabrus rupestris (L.)) to control infestations of Caligus elongatus Nordmann on farmed Atlantic salmon. Aquaculture, 142, 11–24.
Tuya, F., Wernberg, T., and Thomsen, M.S. 2009. Habitat structure affect abundances of labrid fishes across temperate reefs in south-western Australia. Environmental Biology of Fishes, 86, 311–319.
UK Marine SACs Project, 2001. UK Marine Special Areas of Conservation
Unsworth, R.K.F., Peters, J.R., McCloskey, R.M., and Hinder, S.L. 2014. Optimising stereo baited underwater video for sampling fish and invertebrates in temperate coastal habitats. Estuarine, Coastal and Shelf Science, 150, 281–287.
Van Der Veer, H.W.,Witte, J.I., Beumkes, H.A., Dapper, R., Jongejan, W.P., and Van Der Meer, J. 1992. Intertidal fish traps as a tool to study long-term trends in juvenile flatfish populations. Netherlands Journal of Sea Research, 29, 119–126.
Villegas-Ríos, D., Alós, J., March, D., Palmer, M., Mucientes, G., and Saborido-Rey, F. 2013. Home range and diel behavior of the ballan wrasse, Labrus bergylta, determined by acoustic telemetry. Journal of Sea Research, 80, 61–71.
Walker, F.T. 1954. Distribution of laminariaceae around Scotland. ICES Journal of Marine Science, 20(2), 160–166.
Wang, Y., Naumann, U., Wright, S.T., and Warton, D.I. 2013. Package ‘ mvabund ’: Statistical methods for analysing multivariate abundance data.
Watson, D.L., Harvey, E.S., Anderson, M.J., and Kendrick, G.A. 2005. A comparison of temperate reef fish assemblages recorded by three underwater stereo-video techniques. Marine Biology, 148, 415–425.
Wehkamp, S., and Fischer, P. 2013. Impact of hard-bottom substrata on the small-scale distribution of fish and decapods in shallow subtidal temperate waters. Helgoland Marine Research, 67, 59–72.
Wilhelmsson, D., Yahya, S.A.S., and Öhman, M.C. 2006. Effects of high-relief structures on cold temperate fish assemblages: A field experiment. Marine Biology Research, 2, 136–147.
Wood, S., and Scheipl, F. 2014. gamm4: Generalized additive mixed models using mgcv and lme4.
Last updated:





