
Genetic Scorecard Indicator - Juniper
Juniper (Juniperus communis)

IUCN Category:
- Great Britain: Least concern (indicated above) CR for ssp hemisphaerica.
- Europe: Least concern
- Global: Least concern

Genetic Health Status:
- Scottish Risk: Moderate (indicated above)
- UK Risk: Moderate
- Scottish Mitigation status: Partially effective
- UK Mitigation status: Partially effective
Background
Juniperus communis (juniper) is a widespread dioecious gymnosperm with a range of forms, from prostrate shrub to small tree. It is wind-pollinated, with bird-dispersed seed. In the UK, there are three subspecies – communis, nana, and hemisphaerica – the latter of which is limited to a few sites on maritime cliffs in south of England; the others are found elsewhere in the global range (Thomas et al., 2007). The UK population has two primary distribution centres, in Scotland and on the chalk downs of southern England, with significant populations in northern England and Wales. It had a more widespread distribution in the past, but genetic and survey data suggest that populations have been reduced, probably through burning and grazing (McVean & Ratcliffe, 1962, Preston et al., 2002). Baker et al (2025 a,b) note that juniper trees and juniper scrub habitat are “almost a habitat in [their] own right” (Wilkins & Duckworth, 2011), hosting dozens of species of lichens, bryophytes, plants and animals, many of which are specialists and depend on juniper entirely (Binder and Ellis, 2008; Ellis and Coppins, 2009). Juniper scrub is also an important component of forest successional transitions, aiding tree recruitment and being associated with other bare-ground plants (Wilkins and Duckworth, 2011).
The species has been extensively studied using genetic markers; AFLP (Central Europe, Michalczyk et al., 2010; NW Europe / Belgium, Vanden-Broeck et al., 2011); allozymes (Netherlands, Oostermeijer and De Knegt, 2004); nuclear (nSSR) and chloroplast microsatellites (Ireland, Provan et al., 2008); nuclear microsatellites and chloroplast SNP (Germany, Reim et al., 2016); nSSRs (Eurasia, Hantemirova & Bessonova, 2023); chloroplast sequence (Eurasia, Hantemirova et al, 2017). The genetic studies in mainland Europe found varying degrees of fragmentation/genetic isolation, depending on the sampling range. In the UK, however, studies conducted using nSSRs & SNPs (Baker et al., 2025 a,b); AFLPs (Van Der Merwe et al., 2000); nSSRs (Wales, Reynolds, 2022) have consistently shown significant within population diversity levels and among-population structure. Within Great Britain, there is notable genetic differentiation among regional populations from the Scottish Highlands, northern England and southern England, and varying levels of among-population differentiation within each.
Significant differentiation has been observed among regions within the UK (Fst 0.02-0.23), although much lower levels were noted (0.06) among populations within Scotland, even over substantial geographic distances.
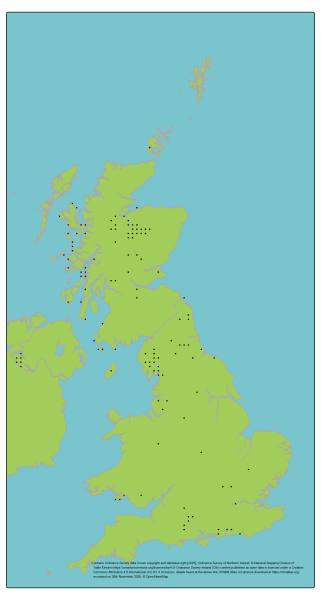
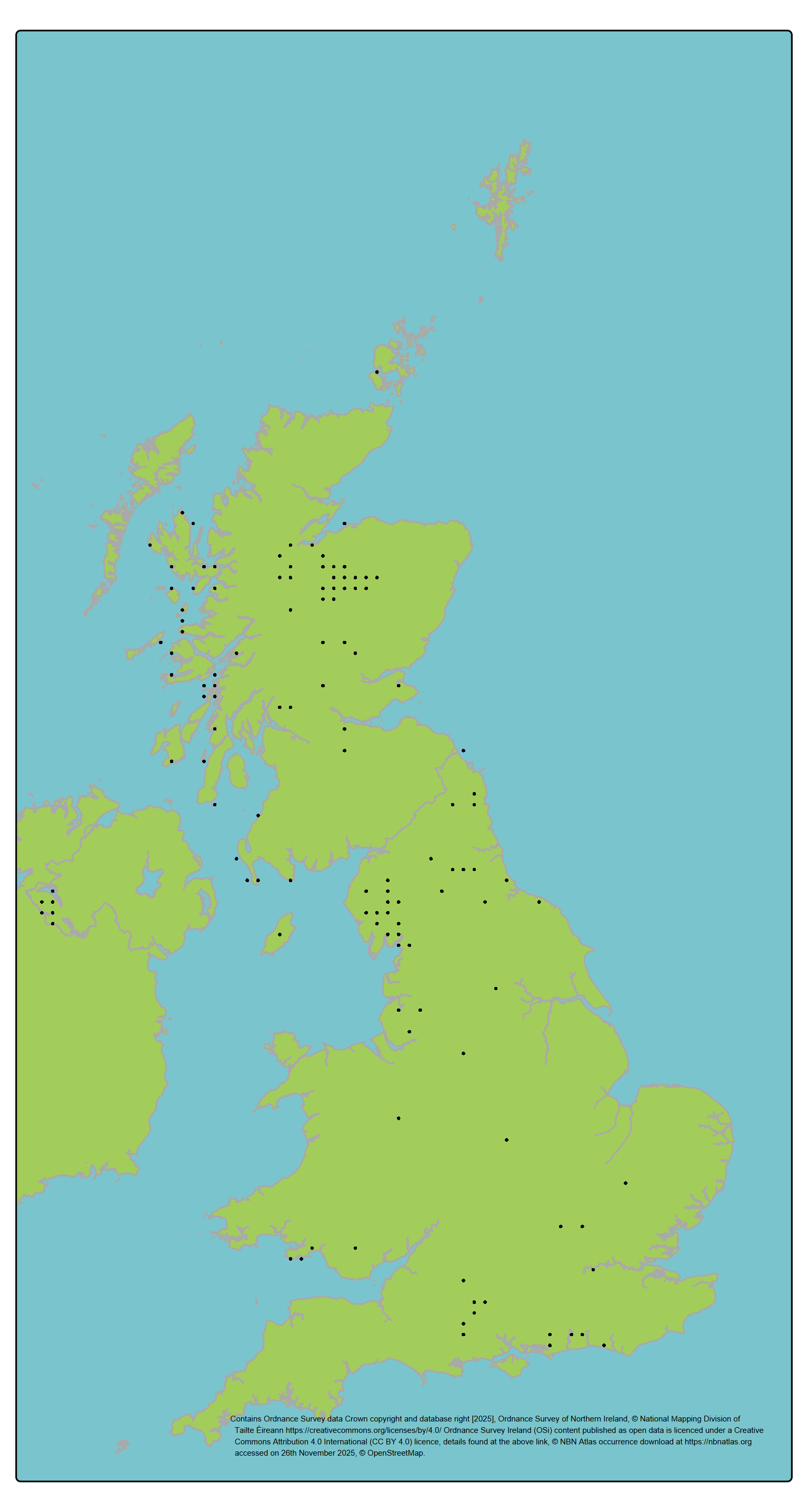
View a larger version of the distribution map for the Juniper.
Current Threats
The species continues to see population erosion, fragmentation and decline. Many populations are aging and developing biased sex ratios. There is widespread lack of significant regeneration for a variety of reasons, although regeneration is plentiful at some sites. The species faces a major threat from the invasive pathogen Phytophthora austrocedri (Mitchell et al., 2023), which appears to have been introduced multiple times. The pathogen is likely to have a major impact although recent work shows the presence of heritable resistance to the pathogen (Green et al., 2021; Crowson et al., 2025) so an adaptive response is possible. Care must be taken to limit the use of clonal propagation in restoration projects, which is often adopted due to the difficulty of propagating from seed, and to minimise planting of stock reared off-site.
Contribution of Scottish/UK population to total species diversity
J. communis is globally widespread in the northern hemisphere, with the widest distribution of any conifer species. The Scottish population is a small part of the global distribution but may occupy distinctive northern oceanic environments relative to the majority of the continental range. Populations in different regions (e.g. Scottish Highlands, Southern Uplands, Lake District, Southern England) are moderately differentiated (Baker et al., 2025; Van Der Merwe, 2000). Subspecies nana and hemisphaerica are present in specific areas; both are also present in other parts of the range, and the latter is genetically distinctive using molecular markers. Trial data suggests a degree of divergent local adaptation among the UK populations.
Genetic risks
Diversity loss: population declines
Populations face ongoing reduction and fragmentation, increasing age and sex ratio shifts, and lack of regeneration. Genetic studies have found evidence for inbreeding in several populations (Baker et al., 2025). These ongoing pressures and selection linked to variation in susceptibility to P. austrocedri will impose genetic bottlenecks and there is a risk of genetic diversity loss.
Global Biodiversity Framework Indicators
Population definitions:
Here, populations were defined based on management units. The four GB Regions of Provenance (Herbert et al., 1999) were selected to represent the major population groupings / management units (treating Northern Ireland as an additional region) as they broadly reflect the environmental variation across which the species is distributed and are likely to capture major patterns of genetic structure.
An estimate of Ne is made below, using the rule of thumb 0.1 x census size. This should be considered highly uncertain: tree species generally maintain large population sizes with overlapping generations that stabilise the gene pool over time. However, a wide range of factors including, in this case, dioecy, variation in reproductive success among individuals, and demographic bottlenecks (e.g. in response to arrival of a novel disease) can impact population genetic diversity, and consequently estimates of Ne.
Ne500: The proportion of populations that have an effective population size of more than 500.
- Proportion of populations with Ne > 500 in Scotland = 2/2
- Proportion of populations with Ne > 500 in UK = 5/5
PM: Proportion of populations that existed in 2000 that still exist in 2025.
- Proportion of populations maintained in Scotland = 2/2
- Proportion of populations maintained in UK = 5/5
Diversity loss: functional variation
Functional variation
Common garden trials have shown genetic variation in phenotypic traits suggesting a degree of divergent local adaptation among populations (Baker et al., 2025 a,b). Population erosion and lack of regeneration may be particularly high in some regions, especially the English downs, posing a risk of loss of within species adaptive genetic diversity.
Divergent lineages
As regional groupings within the UK (notably Scottish Highlands, Lake District, Southern England) show genetic differentiation (Baker et al., 2025 a,b), they merit recognition as distinct genetic units for conservation and management. Threats relevant to the species level are relevant to each unit; English units may face a higher degree of risk of loss.
Hybridisation/Introgression
There is no evidence of risk from hybridisation/introgression. Subspecies within the UK have maintained themselves historically despite overlapping ranges,in the case of sspp. communis/nana, and likely in the face of gene flow among populations of all three subspecies.
Low turnover - constraints on adaptive opportunities
Variable / poor reproductive success and limited recruitment is a key issue for most populations. Although genetic diversity is present, severely limited recruitment will impede adaptive change without intervention.
Cumulative Risk Summary
Overall Genetic Health Status
Scotland
- Risk: Moderate
- Mitigation: Partially effective
Great Britain/UK
- Risk: Moderate
- Mitigation: Partially effective
Overall Genetic Health status explanation
Substantial populations of the species persist in a patchy distribution across the UK, with some larger sites regenerating effectively. Active restoration and management efforts are happening in places (e.g. Trees for Life (2025)) However, in many sites population decline continues and regeneration is severely limited. The threat posed by the invasive pathogen Phytophthora austrocedri is substantial.
In situ genetic threat level
In situ genetic threat level
- In situ Risk for Scotland: Moderate
- In situ Risk for UK: Moderate
Major limitations on regeneration, which have persisted for some time, pose a threat to many populations; this will be exacerbated by both the threat from P. austrocedri and limited gene flow among populations.
Confidence in in situ threat level
- Confidence score for Scotland: High
- Confidence score for UK: High
Substantial research on both the host and pathogen, using DNA and trait-based data, underpin conclusions.
Ex situ representation
Seed collections in the Millenium Seed Bank (MSB) cover (a) 46 of 1062 occupied 10-km squares (4%); (b) an EOO of 341,316 km² out of 528,704 km² occupied (65%); and (c) 5 out of 5 Regions of Provenance (100%). For ssp hemisphaerica, there is a collection representing 1 of 1 occupied 10-km squares (100%) for this sub-species.
Current conservation actions
Key required actions are; 1) protect and expand populations – restore where sites have been lost; 2) manage grazing & burning; make interventions to promote natural regeneration; 3) limit opportunities for translocation of Phytophthora austrocedri among sites (restrict translocations of plants, observe biosecurity measures on sites); 4) ensure substantial and viable ex situ seed collections are maintained.
| Ex situ | Translocation | Habitat management | Legal protection of habitat or species | Regulation of exploitation | Control of INNS/pests/pathogens |
|---|---|---|---|---|---|
| X | - | X | X | - | X |
Population assessment/monitoring
Population
Demographic
N pops assessed/monitored in Scotland = 2/2
N pops assessed/monitored in UK = 5/5
Genetic
N pops assessed/monitored in Scotland = 2/2
N pops assessed/monitored in UK = 5/5
Further research
New research results are due in 2025/26 on seed treatments for improved germination, genetic diversity across the global range, on genetic variation for susceptibility to P. austrocedri, and on the population genetics of the pathogen. A revision of the scorecard to include this work would be merited. Field/lab work to assess the sex ratios, mating system and environmental cofactors of regeneration in a wide sample of populations would be merited. Comprehensive survey of a representative set of populations, sampling across genetic groups, with a view to systematic monitoring should be initiated. Declaration of Genetic Conservation Units within the EUFORGEN framework has been initiated and should be expanded for the species.
References
Baker, J. et al. (2025a). Evidence of genetic isolation and differentiation among historically fragmented British populations of common juniper, Juniperus communis L. BioRxiv.
Baker, J. et al. (2025b). Local genetic adaptations among remnant populations of British Common Juniper, Juniperus communis, Indicated by a Common Garden Trial. Ecology and Evolution, 15(3), e71049.
Binder, M. D., & Ellis, C. J. (2008). Conservation of the rare British lichen Vulpicida pinastri : changing climate, habitat loss and strategies for mitigation. The Lichenologist, 40(1), 63–79.
Crowson et al., (2025) Vulnerable juniper populations show adaptive potential in the face of a highly damaging invasive tree pathogen. Abstract for the BSPP Plant Pathology 2025 and ECPP 2025 Conference.
Ellis, C. J., & Coppins, B. J. (2009). Quantifying the role of multiple landscape-scale drivers controlling epiphyte composition and richness in a conservation priority habitat (juniper scrub). Biological Conservation, 142(7), 1291–1301.
Green, S., James, E.R., Clark, D. et al. (2021) Evidence for natural resistance in Juniperus communis to Phytophthora austrocedri. Journal of Plant Pathololgy 103, 55–59
Hantemirova, E. V., & Bessonova, V. A. (2023). Genetic Diversity of Juniperus communis L. in Eurasia and Alaska Inferred from Nuclear Microsatellite Markers. Russian Journal of Genetics, 59(3), 271–280.
Hantemirova, E. V. et al. (2017). A new Eurasian phylogeographical paradigm? Limited contribution of southern populations to the recolonization of high latitude populations in Juniperus communis L. (Cupressaceae). Journal of Biogeography, 44(2), 271–282.
McVean, D. N., & Ratcliffe, D. A. (1962). Plant communities of the Scottish High- lands. A study of Scottish mountain, moorland and forest vegetation. London: HMSO
Merwe, M. V. et a;. (2000). Spatial and temporal aspects of the genetic structure of Juniperus communis populations. Molecular Ecology, 9(4), 379–386.
Michalczyk, I. M. et al (2010). Genetic support for perglacial survival of Juniperus communis L. in Central Europe. The Holocene, 20(6), 887–894.
Mitchell, R. J., Green, S., & Hollingsworth, P. M. (2023). Plant health, biosecurity, and conservation translocations. In M. J. Gaywood, J. G. Ewen, P. M. Hollingsworth, & A. Moehrenschlager (Eds.), Conservation Translocations (pp. 241-270). Cambridge University Press.
Oostermeijer, J. G. B., & De Knegt, B. (2004). Genetic population structure of the wind-pollinated, dioecious shrub Juniperus communis in fragmented Dutch heathlands. Plant Species Biology, 19(3), 175–184.
Preston, C. D., Pearman, D. A., & Dines, T. D. (2002). New atlas of the British and Irish flora. An atlas of the vascular plants of Britain, Ireland, the Isle of Man and the Channel Islands. Oxford University Press.
Provan, J. et al. (2008). Restricted gene flow in fragmented populations of a wind-pollinated tree. Conservation Genetics (Print), 9(6), 1521–1532
Reim, S. et al. (2016). Genetic structure and diversity in Juniperus communis populations in Saxony, Germany. Biodiversity Research and Conservation, 42(1), 9–18.
Reynolds, F. (2022). The current status of Juniper (Juniperus communis) in Wales: an assessment of site and genetic management. [Master thesis]. Aberystwyth University.
Thomas, P. A. et al. (2007). Biological Flora of the British Isles: Juniperus communis L. Journal of Ecology, 95(6), 1404–1440.
Trees for Life (2025) Unique Highland juniper forest given special European status in UK first
Vanden-Broeck, A. et al. (2011). Genetic structure and seed-mediated dispersal rates of an endangered shrub in a fragmented landscape: a case study for Juniperus communis in northwestern Europe. BMC Genetics, 12, 73.
Wilkins, T., & Duckworth, J. C. (2011). Breaking new ground for juniper: a management handbook for lowland England. Plantlife.
Assessor: Stephen Cavers, UK Centre for Ecology & Hydrology
Reviewers:
- Richard Ennos, University of Edinburgh
- James Baker, UK Centre for Ecology & Hydrology
- Sarah Green, Forest Research
- Peter Hollingsworth, Royal Botanic Garden Edinburgh
Last updated:








{kind=link}