NatureScot Research Report 1365 - Elasmobranch distributions and interactions with fisheries
Year of publication: 2024
Authors: Régnier, T., Gibb, F.M., Clarke, J., Langton, R., Kinnear, S. and Boulcott, P. (Scottish Government Marine Directorate, Marine Laboratory Aberdeen)
Cite as: Régnier, T., Gibb, F.M., Clarke, J., Langton, R., Kinnear, S. and Boulcott, P. 2024. Elasmobranch distributions and interactions with fisheries. NatureScot Research Report 1365.
Contents
- Keywords
- Background
- Main findings
- Acknowledgements
- Abbreviations
- 1. Introduction
- 2. Methods
- 3. Results
- 4. Discussion
- 5. Conclusions
- 6. References
- Annexe A: Survey gear importance in the Random Forest models
Keywords
elasmobranch distributions; fisheries interactions; species distribution modelling; predicted bycatch hotspots, fisheries management measures, marine protected areas; conservation
Background
Elasmobranch (sharks, skates and rays) are ancient, diverse and ecologically important, but are one of the most at-risk vertebrates, mainly as a result of unsustainable harvesting. Knowledge of their biology, ecology or assessment of their status is often impaired by data deficiency which form an important barrier to the development of a dedicated elasmobranch action plan and the implementation of appropriate management actions. In particular, designing effective spatial management measures such as Marine Protected Areas (MPAs) requires knowledge of habitat use and distribution of the target species as well as identifying areas of high vulnerability where such measures are most needed.
This aim of this study was to provide the key evidence necessary for the future development of a conservation and management strategy for elasmobranchs in Scottish waters, through the compilation of elasmobranch survey data and fisheries distribution data.
Elasmobranch data from multiple survey sources were combined to estimate the distributions of nine elasmobranch species in Scottish waters: flapper skate Dipturus intermedius, cuckoo ray Leucoraja naevus, spotted ray Raja montagui, starry ray Amblyraja radiata, thornback ray Raja clavata, spurdog Squalus acanthias, blackmouth dogfish Galeus melastomus, lesser spotted dogfish Scyliorhinus canicula, and starry smooth-hound Mustelus asterias. Species Distribution Models (SDMs) were developed to provide important information regarding their habitat requirements, through the identification of key environmental drivers of these distributions, and the importance of the various fishing gears deployed in the surveys for the detection of species occurrence and variation in their respective abundances.
Elasmobranch distributions predicted by the SDMs were then analysed with regard to the distributions of the Nephrops and scallop fisheries in order to identify areas of importance for elasmobranchs (species richness and diversity) and predicted bycatch hotspots. This combined information is necessary to identify areas where elasmobranchs would benefit from the implementation of fisheries management measures or candidate areas to be considered in the development of an elasmobranch action plan.
Main findings
- Recent modelling methods which combine multiple data sources can provide the required evidence to inform future management decisions and help sustainability and resilience of elasmobranch species, even those considered as data deficient.
- Seven out of nine elasmobranch species considered show preferences for depths of 100 m or deeper, which explains their predominance on the west of Scotland, where deep trenches can be found close inshore and within sea lochs.
- Species with either a preference for shallow depths (cuckoo ray) or no particular depth preference (spotted ray) were commonly found in the North Sea, to the east of Scotland.
- Elasmobranch seabed preferences ranged from soft to coarse substrates and most species preferred waters characterised by low bottom primary productivity and high salinity (except the starry smooth-hound and the starry ray).
- Skates and rays were more often retained in the dredge compared to squaliform species, and all species were caught in large numbers in bottom trawl surveys.
- Overlap in the distributions of the considered elasmobranchs and the scallop and Nephrops fisheries revealed seven areas where conservation measures may have an impact: south of Jura, Kintyre, NE Arran, south of Barra, Skye to Red Point, Rattray Head and to the east of Montrose.
- Most of the identified important areas are either overlapping or contiguous with the existing Scottish MPA network.
- Future implementation of fishery management measures within the MPA network and extensions to some of the managed areas could offer protection to this threatened group of fish.
Acknowledgements
This work was funded by NatureScot under project 118019 aiming at: Developing elasmobranch conservation policy and promoting elasmobranchs.
Abbreviations
Akaike information criterion (AIC)
Baited remote underwater video systems (BRUVs)
Bottom trawl (BT)
Database of trawl surveys (DATRAS)
European Union (EU)
French southern Atlantic bottom trawl survey (EVHOE)
Gaussian random field generalized linear mixed effects model (GLMM)
GOV (Grande Ouverture Verticale)
Integrated mapping for the sustainable development of Ireland's marine resource (INFOMAR)
International council for the exploration of the sea (ICES)
International union for conservation of nature (IUCN)
Inverse distance weighing (IDW)
Irish groundfish survey (IE-IGFS)
Loch Sunart to the Sound of Jura marine protected area (LStSJ MPA)
Marine protected areas (MPAs)
North Sea international bottom trawl survey (NS-IBTS)
Oslo-Paris conventions for the protection of the marine environment of the North-East Atlantic (OSPAR)
Priority marine features (PMFs)
PSA (particle size analysis)
Root mean square error (RMSE)
Scottish biodiversity strategy (SBS)
Scottish Government’s Marine Directorate (MD)
Scottish west coast IBTS (SWC-IBTS)
Shannon diversity (H)
Special Area of Conservation (SAC)
Species distribution models (SDMs)
Swept-area ratio (SAR)
Total allowable catch (TAC)
United Kingdom biodiversity action plan (UKBAP)
1. Introduction
The ability to identify sites of marine biodiversity value and adopt relevant spatial management measures is critical for preserving diversity and essential ecological processes. Identification of such sites and the implementation of efficient, informed spatial management measures requires knowledge of both the distributions of the marine resources targeted and of the threats they face.
Elasmobranchs (sharks, rays and skates) form one of the most ancient, diverse and widely distributed vertebrate clade (Stein et al., 2018) and can be found in a variety of habitats, from freshwater to the deepest parts of our oceans and seas. Elasmobranchs are associated with the provision of a wide range of benefits to society, often occupying a top or near-top position in marine food webs helping to maintain the structure and stability of marine ecosystems (Polovina et al., 2009) and, as such, are often seen as an indicator of health in marine ecosystems. Elasmobranchs have an important economic value, indirectly through their role in maintaining the structure of marine ecosystems and the stability of fisheries, but also directly via targeted fisheries or through their recreational value: elasmobranchs are often a prized target for recreational angling and a key driver of eco-tourism.
While elasmobranchs provide clear societal benefits, they are in global decline, with over one third of its species threatened with extinction (Dulvy et al., 2021). The main causes for this decline are overfishing, habitat loss and degradation, climate change, and pollution. Overfishing is considered to be by far the largest threat, with all threatened elasmobranchs affected in some way by fishing (Dulvy et al., 2021). Species sensitivity to overfishing is related to their distribution, gear selectivity (Piet et al., 2009; Walker et al., 2017), and life-history traits. In particular, the typical large size, slow growth, late maturity and low fecundity of elasmobranchs make them particularly vulnerable to fishing pressure (Pope et al., 2000; Beddington and Kirkwood, 2005) and the associated slow rate of population growth (Myers and Worm, 2005) makes recovery post-depletion a slow and uncertain process. Aside from overfishing, other human activities have resulted in the destruction, degradation or disturbance of elasmobranch habitat. Feeding and nursery areas, such as estuaries, sea grass beds and mangroves, are particularly impacted by pollution and increases in coastal development. Climate change has also impacted elasmobranchs, with the effects of ocean warming and acidification found on locomotion (Vilmar and Di Santo, 2022) and physiological rates (Santos et al., 2021). These threats are often misunderstood and unaccounted for and likely lead to a greater sensitivity of elasmobranchs species in some areas where pressures overlap. For example, elasmobranchs at the margin of their distribution area are predicted to show an increased sensitivity to over-exploitation resulting from the interaction between climate change effects and fishing pressure (Rindorf et al., 2020).
Targeted fisheries exist in certain regions of the globe, however, most elasmobranchs are caught as bycatch in mixed fisheries (Baum and Myers, 2004; Clarke et al., 2015; Stevens et al., 2000). The gear used in mixed fisheries, often bottom trawls, is generally unselective, resulting in both target and non-target species being caught (Pauly and Murphy, 1982; Alverson et al., 1994). As a result, large-bodied elasmobranchs, are particularly sensitive to mixed fisheries, and these are considered to be a major driver in their global decline (Brander, 1981; Walker and Hislop, 1998; Dulvy et al., 2000; Ford et al., 2015). Bentho-pelagic species, such as many sharks (squaliform sharks, catsharks and hound sharks), and bentho-demersal species, like skates and rays, are expected to be retained by bottom trawls and benthic gear such as beam trawls or dredges are likely to retain skates and rays and have the potential to damage the eggs of most elasmobranchs.
Fisheries management measures generally take two forms: (i) fishing effort management and (ii) catch management. Fishing effort management aims to reduce fishing pressure and can be implemented through restrictions on the number and size of vessels operating in an area, the amount of time and/or the area where a fishery can operate, or a restriction on the type and amount of gear that can be deployed (e.g. number of static gears that can be deployed). In practice, fishing effort management is often implemented through fishing closures (spatially and seasonally) and gear restriction (e.g. mesh size). Catch management aims at limiting the amount of fish harvested by a fishery and is implemented through setting a Total Allowable Catch (TAC), minimum landing size or landing bans. The impact of fisheries management measures on elasmobranchs is often difficult to assess due to a lack of resources, ineffective or partial implementation and enforcement of measures, but particularly as a result of gaps in knowledge of basic elasmobranch life-history.
Important knowledge gaps include basic understanding of species biology and ecology, population status, scale and intensity of threats, and the effectiveness of management measures (Jorgensen et al., 2022). Elasmobranchs have particular life-histories, often characterised by low fecundity, slow growth and late maturity. As a result, population growth rates are lower than most teleost fish (Myers and Worm, 2005). Filling these knowledge gaps is necessary to assess the population status of elasmobranchs and to take informed decisions regarding their conservation and recovery. Knowledge of elasmobranch distributions is also vital to the successful implementation of spatial measures aiming at protecting target species throughout their life-cycle or at those key developmental stages (e.g. egg stage, nursery areas) most sensitive to the identified threats. However, a lack of data resulting from misidentification and misreporting of catch data (Lack and Sant, 2011) make the estimation of these variables difficult to obtain. As elasmobranchs are caught in mixed fisheries which do not target them specifically, they are often discarded or recorded as bycatch or unidentified elasmobranchs (Barker and Schluessel, 2005). Estimating the population status, distributions and exploitation patterns is therefore difficult (Lack and Sant, 2011). As a result, most elasmobranchs in UK waters are listed as data-deficient (ICES, 2012a). The necessity to fill these knowledge gaps in order to implement effective management measures therefore requires a flexible approach taking advantage of all potential data sources available. A key priority is to build the evidence base necessary to inform decision-making in the future development of efficient management and conservation measures for elasmobranchs. This evidence base requires an assessment of elasmobranch diversity, habitats and distributions, as well as an assessment of threats (e.g. vulnerability to fishing gear) and their distributions (e.g. bycatch hotspots).
Scottish waters extend from the North Sea to the East Atlantic, encompassing a large diversity of habitats, from the shallow areas of the North Sea to the Rockall trough, the continental shelf and the deep sea lochs to the west of Scotland. This range of habitat hosts a diversity of elasmobranchs (Table 1, see also Scotland’s Marine Atlas), of which two thirds are classified as at least “near threatened” by the IUCN. Most of the species classified as “least concern” are deep sea sharks for which no population status can be estimated. While not a specific target of fisheries around the UK, elasmobranchs are mainly caught as accidental bycatch in mixed demersal fisheries, longlines and inshore with tangle nets. Over the period 2007-2010, 60% of skates and rays landings were associated with bottom-trawling (Silva et al., 2012). Since 2009, commercial fisheries are required to report elasmobranch landings by species as opposed to family (pre-2009, e.g. skates and rays were reported as a single group). However, misidentification and misreporting is still an issue. For example, species of the common skate complex (Dipturus intermedius and Dipturus flossada, the flapper and blue skate respectively) have long been reported as a single species (Dipturus batis) but recently identified as two species (Griffiths et al., 2010; Iglésias et al., 2010). As a consequence, detailed knowledge of elasmobranch distributions and vulnerability to fisheries in Scottish waters is lacking.
Table 1. Elasmobranchs found in Scottish waters, information on conservation status, management and classification as Priority Marine Features in Scotland.
| Common name | Latin Name | Status | Priority Marine Feature |
|---|---|---|---|
| Angelshark | Squatina squatina | Critically endangered. Prohibition on retention in all ICES areas. | No |
| Arctic skate | Amblyraja hyperborea | Least concern. No target fishery. | No |
| Basking shark | Cetorhinus maximus | Endangered. Prohibition on retention in all ICES areas. | Yes |
| Birdbeak dogfish | Deania calcea | Endangered. No target fishery but taken as bycatch. | No |
| Black dogfish | Centrophorus fabridi | Least concern. No target fishery but taken as bycatch. | No |
| Black skate | Dipturus nidarosiensis | Threatened no current target fishery. | No |
| Blackmouth dogfish | Galeus melastomus | Least concern. Zero Total Allowable Catch (TAC) in EU waters. | No |
| Blonde ray | Raja brachyuran | Near threatened no known target fishery. | No |
| Blue shark | Prionace glauca | Near threatened. No target fishery but taken as bycatch. | Yes |
| Bluntnose sixgill | Hexanchus griseus | Near threatened. No target fishery but caught as bycatch in deep water trawls. | No |
| Common skate | Dipturus batis complex | Critically endangered. Prohibition on retention in all ICES areas. | Yes |
| Common stingray | Dasyatis pastinaca | Near threatened. No target fishery. | No |
| Cuckoo ray | Leucoraja naevus | Least concern. No target fishery. | No |
| Demon catsharks | Apristurus spp. | Least concern. No target fishery but taken as bycatch. | No |
| Frilled shark | Chlamydoselachus anguineus | Least concern. No target fishery but taken as bycatch. | No |
| Great lantern shark | Etmopterus princeps | Least concern. No target fishery but taken as bycatch. | No |
| Greenland shark | Somniosus microcephalus | Near threatened. No target fishery but taken as bycatch. | No |
| Kitefin shark | Diarias licha | Least concern. No target fishery but taken as bycatch. | No |
| Knifetooth dogfish | Scymnoden ringens | Near threatened. No target fishery but taken as bycatch. | No |
| Leafscale gulper shark | Centrophorus squamosus | Endangered. No target fishery. | Yes |
| Long-nose skate | Dipturus oxyrinchus | Near threatened. No target fishery. | No |
| Longnose velvet dogfish | Centroselachus crepidater | Least concern. No target fishery but taken as bycatch. | No |
| Nursehound | Scyllorhinus stellaris | Near threatened. Some target fishery. | No |
| Porbeagle shark | Lamna nasus | Critically Endangered. Zero TAC in EU waters. | Yes |
| Portuguese dogfish | Centroscymnus coelolepis | Endangered. Zero TAC in EU waters. | Yes |
| Sandy ray | Leucoraja circularis | Endangered. Managed under a common TAC for skates and rays. | Yes |
| Shagreen ray | Leucoraja fullonica | Near threatened. No target fishery. | No |
| Lesser spotted dogfish | Scyllorhinus canicula | Least concern. No target fishery but larger specimens kept for human consumption. | No |
| Spurdog | Squalus acanthias | Endangered. TAC set by ICES. | Yes |
| Spotted ray | Raja montagul | Endangered. No current target fishery. | No |
| Starry ray | Amblyraja radiata | Least concern. No target fishery. | No |
| Starry smooth-hound | Mustelus asterias | Least concern. Unimportant bycatch in multispecies trawls. | No |
| Thornback ray | Raja davata | Near threatened. Target fishery within Europe. | No |
| Thresher shark | Alopias vulpinus | Near threatened. No target fishery but taken as bycatch. | No |
| Торе | Galeorhinus galeus | Vulnerable. No target fishery in Scotland, but target fishery within England and Wales.. | No |
| Velvet belly laternshark | Etmopterus spinax | Near threatened. No target fishery but taken as bycatch. | No |
In addition to catch management advice provided by ICES and EU legislation, in Scottish waters, a number of elasmobranchs benefit from additional protection. The Sea Fishing (Prohibition on the Removal of Shark Fins) (Scotland) Amendment Order 2009 was put in place to ban the shark finning practice and in 2012 the Scottish Government introduced legislation to offer extra protection to sharks, skates and rays in Scottish waters (covering 21 elasmobranch species). A number of elasmobranchs are classified as Priority Marine Features (PMFs) in Scotland. PMFs are species or habitats present on existing conservation lists (e.g. Natura, Wildlife and Countryside Act 1981, UKBAP, SBS, OSPAR) with a significant presence in Scottish waters and with key functional roles. The designation of current and future management measures in Scottish waters aim to conserve or recover PMFs. In addition, several elasmobranch species are Scottish Marine Protected Area (MPA) Network Features. Four MPAs have been designated with elasmobranchs as a feature, Loch Sunart to the Sound of Jura and Red Rocks and Longay for flapper skate, Sea of Hebrides for basking shark and West of Scotland for the gulper shark and Portuguese dogfish (Figure 1).
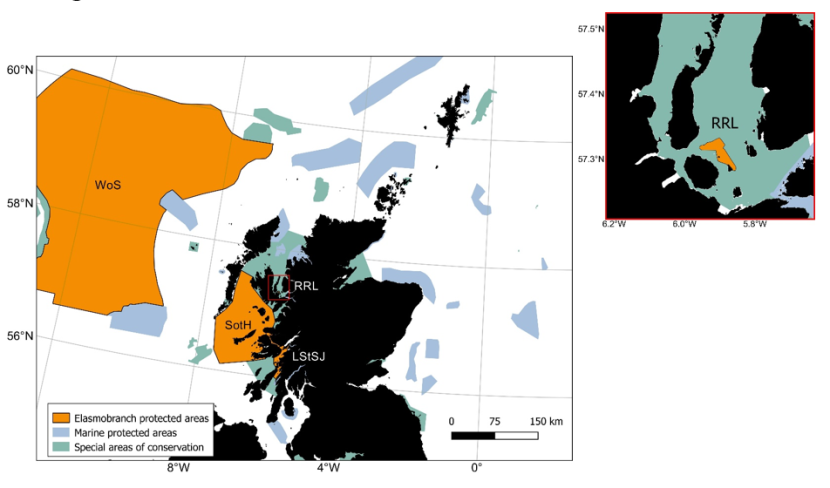
Figure 1. Scottish Marine Protected Areas network with sites where elasmobranchs are a protected key features represented in orange.
Click for a full description
Map of Scottish waters with inshore and offshore Marine Protected Areas (MPAs) visible as shaded polygons. Four MPAs with specific protection for elasmobranchs are shaded in a different colour:
- WoS – West of Scotland
- SotH – Sea of the Hebrides
- LStSJ – Loch Sunart to the Sound of Jura
- RRL – Red Rocks and Longay
A small inset to the right of the main figure shows a close-up view of the Red Rocks and Longay MPA which is hard to make out on the main figure. Special areas Areas of conservation Conservation are shaded in another colour distinct from the MPAs.
As highlighted in the “2020 Challenge for Scotland’s Biodiversity”, the Scottish government aims to establish a coherent network of Marine Protected Areas, promote sustainable use and conservation and collate information on the location of PMFs and their sensitivities, and make this information available to support their protection. In addition, the “Biodiversity strategy to 2045” lists a number of key objectives such as the delivery of fisheries management measures for existing MPAs, the implementation of fisheries measures for PMFs outside of MPAs, and the development of an ecosystem based approach to the management of sea fisheries to avoid adverse impacts on non-target species. With regard to elasmobranchs in Scottish waters, the delivery of such objectives requires a solid evidence base documenting distributions and sensitivities to potentially harmful human activities. The objectives of the present report are therefore to consolidate the evidence base by assessing elasmobranch distributions in Scottish waters and to comment on their sensitivities to various fishing gear and candidate fisheries. For this purpose, and for species with sufficient distribution data, data originating from various sources were combined to produce Species Distribution Models (SDMs, Figure 2A). In addition to estimating the likely distributions of elasmobranchs in Scottish waters, the SDMs provide a statistical framework to identify and account for environmental drivers responsible for these distributions as well as sensitivity to the capture methods (i.e. fishing gear) used for the collection of data. The overlap between the estimated elasmobranch distributions and the distributions of candidate fisheries can then be used (Figure 2B) to identify areas where elasmobranchs are more vulnerable to these activities. Finally, this information can then be used to inform decision making in the future development of conservation or management measures for elasmobranchs in Scottish waters (Figure 2C).
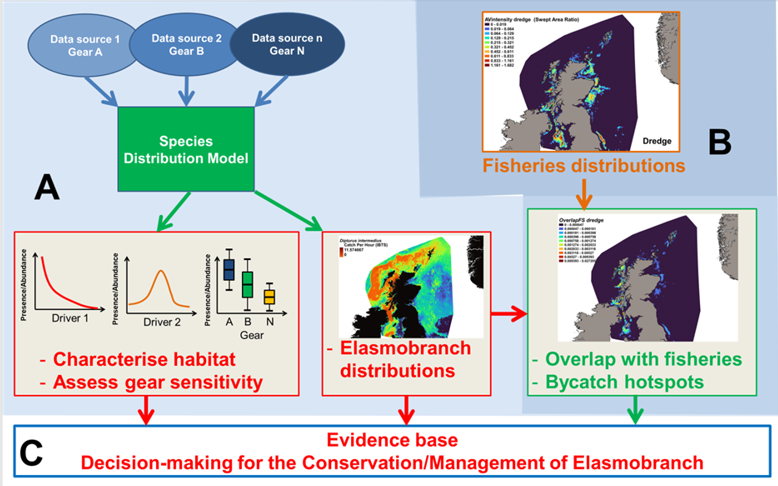
Figure 2. Objectives of this report.
Click for a full description
Arrows from Sections A and B point into Section C.
Section A – Species data from various sources feed into a Species Distribution Model (SDM) which will inform habitat characterisation, assess gear sensitivity and allow mapping of elasmobranch distributions.
Section B – Fisheries distribution data will be analysed along with elasmobranch distributions to define areas of overlap and predict bycatch hotspots.
Section C – Outputs from the project will provide the evidence base for effective management and conservation of elasmobranchs in Scottish waters.
2. Methods
2.1 Distribution data
Distribution information for elasmobranch species in Scottish waters was collated from fisheries independent survey data, comprising the ICES Database of Trawl Surveys (DATRAS) and the Scottish Government’s Marine Directorate (MD) research surveys; Table 2.
Table 2. Sources of data for Elasmobranchs.
| Survey | Description | Temporal coverage | N. surveys | N. recorded elasmobranch species (including complexes) |
|---|---|---|---|---|
| ICES DATRAS | Database of Trawl Surveys | 1965-2022 | 288 | 42 |
| MD FSS | Fisheries Survey System | 2013-2022 | 49 | 36 |
| MD RVR | Research Vessel Records (pre-cursor to MD FSS) | 1925-1999 | 227 | 31 |
| MD sBRUV | Stereo baited remote underwater video surveys | 2017-2019, 2021-2022 | 11 | 9 |
| MD Scallop | Inshore scallop surveys | 1993-2022 | 91 | 10 |
| MD Inshore | Charter and Inshore surveys | 2001, 2003-2005, 2009-10 | 9 | 1 |
The ICES groundfish surveys comprised data from various European countries, where research vessels were equipped with a variety of bottom gears (including beam and otter trawls) with various rigging configurations and catch efficiencies (ICES, 2022a). Marine Directorate surveys used various platforms (research and charted fishing vessels) and gears (trawls, dredges and BRUVs – baited remote underwater video systems). Data from each survey recorded both chronological data (e.g. location, date, depth, gear) along with catch data, including total numbers caught per species and species length frequencies. The collated information comprised 675 individual surveys, within which 9 out of 37 recorded elasmobranch species were deemed in significant number to estimate distributions (Table 3).
Table 3. Elasmobranch species selected for the present report based on data availability (PMF species in bold).
| Grouping | Common name | Species | Priority Marine Feature | N. records |
|---|---|---|---|---|
| Skate and Ray species | Flapper skate | Dipturus intermedius | Yes | 5,339 |
| - | Cuckoo ray | Leucoraja naevus | No | 40,774 |
| - | Spotted ray | Raja montagui | No | 47,838 |
| - | Starry ray | Amblyraja radiata | No | 89,548 |
| - | Thornback ray | Raja clavata | No | 58,445 |
| Squaliform species | Spurdog | Squalus acanthias | Yes | 206,739 |
| - | Blackmouth dogfish | Galeus melastomus | No | 50,669 |
| - | Lesser spotted dogfish | Scyliorhinus canicula | No | 601,801 |
| - | Starry smooth-hound | Mustelus asterias | No | 15,022 |
For DATRAS and MD surveys (except the camera-based BRUV surveys), abundance was calculated as the numbers of fish caught per individual haul, standardised to catches per one hour of fishing and locations assigned to the midpoints of the tows. Within the BRUV survey series, laboratory-based video analysis confirmed the presence/absence of six identifiable elasmobranch species (Dipturus intermedius, Galeus melastomus, Raja clavata, Scyliorhinus canicular, Scyliorhinus stellaris and Squalus acanthias) or species complexes (e.g. Scyliorhinus), with abundance defined as the maximum number (MaxN) of each species captured within one frame over a period of one hour. Point distributions were plotted using QGIS (version 2.14) to determine species extent to inform the modelling inputs. Hauls with zero elasmobranch catches were not considered true absences unless the entire catch was sampled routinely during the survey series.
Haul locations of each of the 14 individual gears deployed by the fishery-independent surveys from 1980 – 2022 are indicated by heatmaps (Figure 3). Dredge data were collected from the MD scallop survey, which sampled 3 main regions; the west coast of Scotland, Scottish east coast and Shetland. Highest activity was focussed within inshore regions, particularly around the Shetland coast, Moray Firth and to the NW of Skye. GOV trawl data were collected from the NS-IBTS, EVHOE, IE-IGFS and Scottish west coast IBTS surveys and thus comprised the most robust, consistent and spatially/temporally comprehensive data source across the North Sea and west coast of Scotland. Tow intensity for the Scottish west coast IBTS surveys was highest to the north of Ireland and off the NW coast of Lewis. Within the North Sea, the gridded survey design resulted in high numbers of deployments from the coast out into deeper waters of the central and northern North Sea. Data within the Irish Sea were collected by otter trawl, as part of the ICES groundfish surveys, and from 4 m beam trawl surveys, with tows concentrated inshore. Data from 8 m beam trawl surveys were collected exclusively from the North Sea, extending from the Moray Firth to deeper more offshore regions of the central North Sea. Data from the Aberdeen gear, an 180 ft trawl, were collated mainly from North Sea surveys, particularly in the Moray Firth, off the NE coast of Scotland, around Orkney/Shetland and into the deeper central North Sea, but with a few hauls to the west coast of Scotland to the north of the Outer Hebrides. The BT237 (similar to the Jackson trawl) was used for MD gear trial surveys, with data collected from the south of Shetland and Moray Firth. Similarly, MI trawl data was collected from MD gear trials in the Moray Firth. The BT184 was a deepwater trawl used off the continental shelf on the west coast of Scotland, with data collected primarily from west of the Outer Hebrides. Data from the BT196 deepwater trawl were collected from tows off the west coast continental shelf, south of the Outer Hebrides, around Orkney and Shetland and into the NW North Sea. Data collected from deepwater surveys using the Jackson trawl were from the west coast continental shelf, south of the Outer Hebrides and in the central North Sea. Data collected by the Dutch herring trawl surveys were exclusively from the North Sea (particulary around Shetland, east of Orkney, the Moray Firth, east of the Firth of Forth and out into the central North Sea) with no activity conducted on the west coast. Data from the 180 ft herring trawl were collated from surveys within the central and western North Sea, particularly to the SE of Shetland. For areas where fishing was not permitted or unsuitable for trawling, distribution data was collected from BRUVs surveys, deployed exclusively on the west of Scotland, including near inshore locations within the Minch, Sound of Sleat (SE Skye), Sound of Mull, Firth of Lorn, Sound of Jura and the Firth of Clyde.
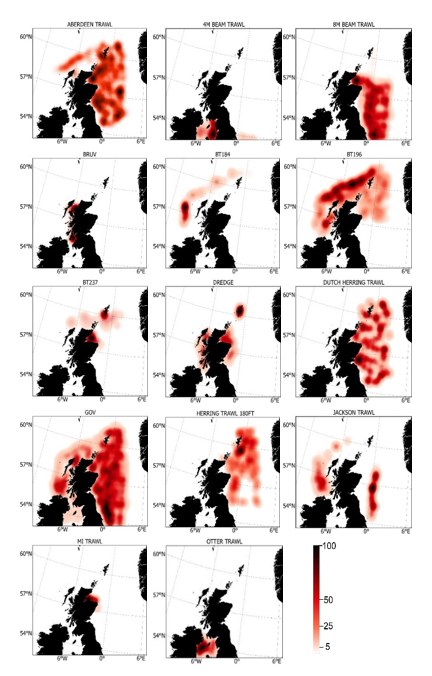
Figure 3. Heatmaps showing intensity of fisheries independent survey sampling for each individual gear type between 1980 and 2022. Range indicates distribution percentiles.
Click for a full description
Fishing intensity of each gear is represented by individual heatmaps with a colour ramp of distribution percentiles from 0-100. Heatmaps are for 14 gear types, including Aberdeen Trawl, 4 m Beam Trawl, 8 m Beam Trawl, BRUV, BT184, BT196, BT237, Dredge, Butch Herring Trawl, GOV, Herring Trawl 180 ft, Jackson Trawl, MI Trawl and Otter Trawl. Trawl distributions are widespread both inshore and offshore, whereas dredge distributions are concentrated inshore.
2.2 Efficiency of survey gears
Survey gear dimensions are shown in Table 4. Whilst survey gear catchability was not estimated within this study, it is clear from individual gear dimensions, particularly gear wing spread (Table 4), that the dredge is capable of sampling a much smaller area as compared to the much larger trawl gears (Table 4) and thus would be expected to catch fewer fish. Gear avoidance of the dredge is also likely higher than in trawl fishing, with the dredge limited to catching less mobile species, particularly those which reside on the actual substrate. While catches using scientific dredges may not be directly comparable to those of commercial scallop fishers (the gear having smaller rings and more teeth), given the large size of the species considered it is unlikely this difference would affect overall catchability. The longer tow durations (just short of one hour) of the Aberdeen, and larger headline height of the BT184 and BT196 trawls may increase elasmobranch catchability, not least due to the increased swept area but also through possible swimming fatigue.
Table 4. Survey gear dimensions (average values from valid tows).
| Gear | Haul duration (mins) | Wing spread (m) | Swept area (km2) | No. valid tows (area) | Net opening (m) | Volume fished (km3) | No. valid tows (vol) |
|---|---|---|---|---|---|---|---|
| Aberdeen Trawl | 58.02 | 16.00a | - | 1512 | 3.00a | - | - |
| Beam Trawl 4M | 28.84 | 4.00 | 0.01432 | 7234 | - | - | - |
| Beam Trawl 8M – Single | 29.29 | 8.00 | 0.03062 | 3370 | - | - | - |
| Beam Trawl 8M – Double | 29.19 | 8.00 | 0.05924 | 1170 | - | - | - |
| BT184 | 45.99 | 26.45 | 0.12042 | 213 | 4.00b | 0.00048c | - |
| BT196 | 57.93 | 27.10 | 0.17329 | 341 | 6.00b | 0.00104c | - |
| BT237 | 18.59 | 22.56 | 0.04117 | 85 | 5.50b | 0.00023c | - |
| Dredge | 30.79 | 9.00 | 0.06208 | 6538 | - | - | - |
| Dutch Herring Trawl | 36.79 | - | - | 1125 | - | - | - |
| GOV | 30.40 | 19.58 | 0.07055 | 16326 | 4.56 | 0.00034 | 15517 |
| GOV (gear trials) | 15.00 | 20.45 | 0.03438 | 69 | 5.00 b | 0.00017c | - |
| Herring Trawl 180ft | 44.16 | - | - | 149 | - | - | - |
| Jackson Trawl | 56.21 | 29.86 | 0.15847 | 609 | 5.42 | 0.00086 | 599 |
| MI Trawl | 17.00 | 17.71 | 0.03294 | 13 | 4.00 b | 0.00013c | - |
| Otter Trawl | 34.91 | 15.72 | 0.05220 | 103 | 2.85 | 0.00015 | 103 |
a from Neat and Campbell, 2011
b pers comm from Rob Kynoch (Gear expert, MD)
c estimated
2.3 Distribution models
Only fisheries independent surveys containing at least one verified occurrence of a given species were kept for a given species model for the time period 1980-2022. Species with offshore distributions or with very localised distribution records (e.g. Rockall, Faroe) were discarded (9 species out of 37 retained for the analyses). The data used in the Species Distribution Models are presented in Figure 4.
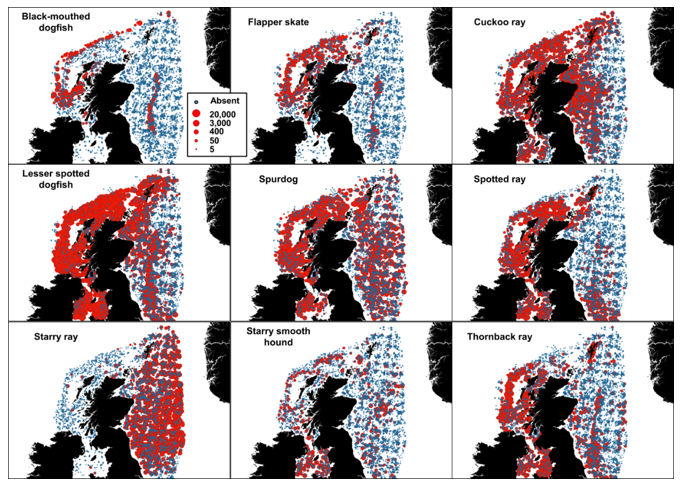
Figure 4. Fishery-independent data used in the Species Distribution Models. The abundance indicated is in number caught per hour.
Click for a full description
Presence absence maps for black-mouthed dogfish, flapper skate, cuckoo ray, lesser spotted dogfish, spurdog, spotted ray, starry ray, starry smooth hound and thornback ray are shown in individual panes. Absence records are indicated by different symbology from presence. Presence is indicated by dots of varying size - representing a range in abundance from 5 – 20,000 catch per hour. There is good data coverage for all species around Scotland, but presence - in terms of abundance and geographic distribution - varies between species. The blackmouth dogfish, flapper skate and starry smooth hound have fewer presence data than the other species. Records of blackmouth dogfish, flapper skate, thornback ray and spotted ray presence are higher on the west coast, whereas starry ray presence records are predominantly found on the east coast. Presence records of data rich species such as the lesser spotted dogfish, spurdog and cuckoo ray are ubiquitous across all regions.
Environmental covariates
A number of environmental covariates were used to fit the SDMs. Bathymetry was obtained from EMODnet and slope was derived from bathymetry (See Langton et al., 2020, Figures 5). Chlorophyll A, current velocity, light intensity, primary productivity, salinity and temperature at the level of the seabed (Figures 5-6) were obtained from BIO-ORACLE (Assis et al., 2017). Percentage gravel, sand and silt were spatially interpolated using records of sediment PSA from the British Geological Survey, Marine Recorder and INFOMAR. Spatial interpolation was achieved using Kriging, Inverse Distance Weighing (IDW) and thin plate regression splines and the output were combined in an ensemble model using cross-validated Root Mean Square Error (RMSE) as a weighting factor (Figure 7). Interpolations were done in R 4.2.2 using the packages “gstat” (Kriging and IDW, Gräler et al., 2016) and “mgcv” (thin plate regression, Wood, 2003).
A polygon corresponding to the area covered by all environmental variables was then used to clip fish distribution data and as the spatial extent used for predicting elasmobranch distributions. The prediction data frame was constructed from a grid (1 km x 1 km) adapted to the spatial extent described above and covariates were resampled to this resolution using a nearest neighbour approach. Due to high collinearity, not all variables were included simultaneously in the models. In particular, chlorophyll A was not used due to a high covariation with both salinity and primary productivity, and % silt was removed as it can be deduced by combining % sand and % gravel.
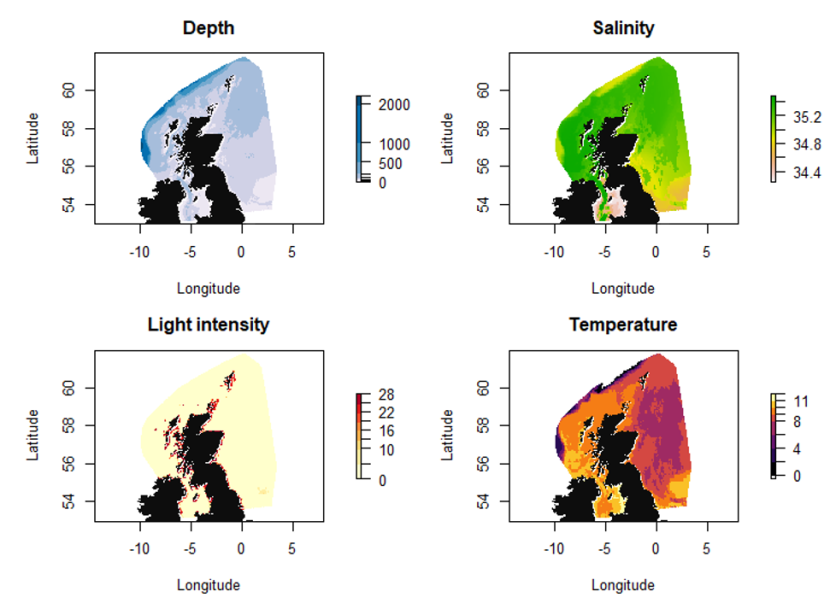
Figure 5. Covariates depth, salinity, light intensity and temperature used in the various SDMs.
Click for a full description
Variation in water depth is indicated by a colour gradient representing a range from 0 - 2000 m. Depths are deepest off the continental shelf to the north west of Scotland. Shallower water is found in inshore waters around the coastline, and within the central and southern North Sea.
Variation in salinity is indicated by a colour gradient representing a range from 34.4 - 35.2 ppt. Salinity is highest to the north and west of Scotland, and lowest in the central-southern North Sea.
Variation in light intensity is indicated by a colour gradient representing a range from 0-28 intensity per meter squared per year. Light intensity is highest around the Scottish coast, and low within offshore areas.
Variation in sea temperature is indicated by a colour gradient representing a range from 0 - 11 °C. Temperatures are highest to the north and west of Scotland and lowest temperatures are seen within offshore areas of the North Sea and off the continental shelf.
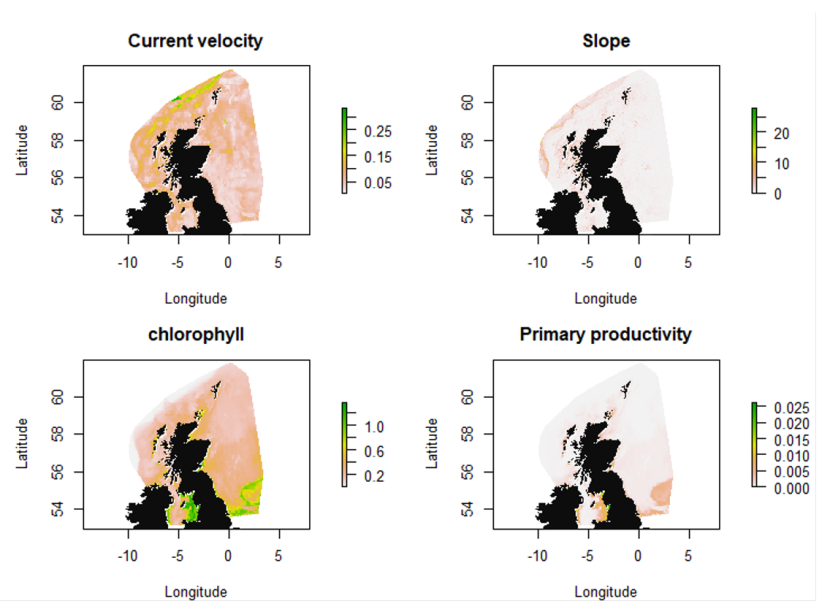
Figure 6. Covariates current velocity, slope, chlorophyll and primary productivity used in the various SDMs.
Click for a full description
Variation in current velocity is indicated by a colour gradient representing a range from 0.05- 0.25 m.s-1. Highest velocities are along the continental shelf to the north and north-west of Scotland.
Variation in slope is indicated by a colour gradient representing a range from 0 - 20 m. Steepest slopes are found off the continental shelf to the north and north-west of Scotland, and off the islands of the west coast.
Variation in chlorophyll is indicated by a colour gradient representing a range from 0.2 – 1.0 mg.m-3, and is highest in coastal areas, particularly in the Irish Sea and southern North Sea.
Primary productivity is shown by a colour gradient representing a range from 0 – 0.025 g.m-3.day-1. Productivity is highest in coastal areas, particularly to the west of the Irish Sea.
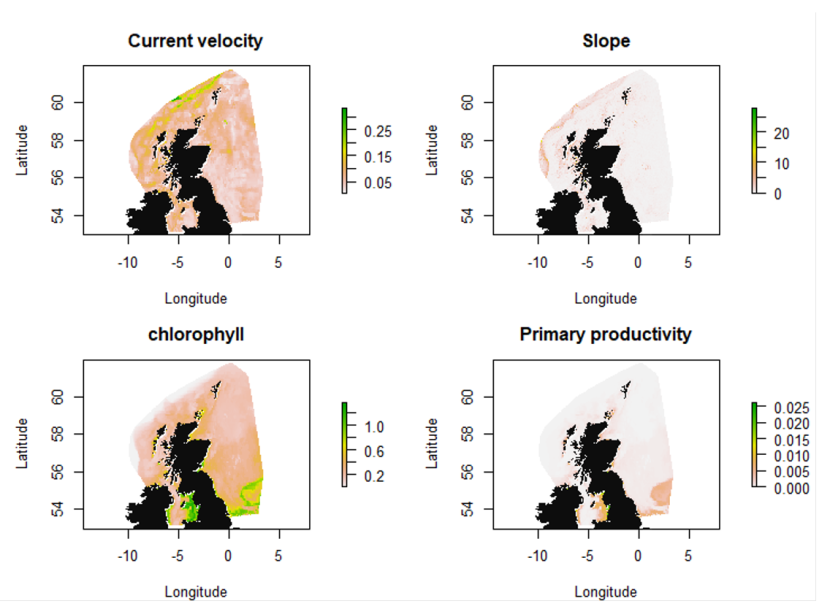
Figure 7. Covariates percentage gravel, sand and silt used in the various SDMs.
Click for a full description
Variation in percentage gravel is indicated by a colour gradient representing a range from 0 - 70. Sediments with the highest gravel content have a patchy distribution, found mainly in offshore waters to the north-west of Orkney and to the west of Scotland, particularly off the tip of Kintryre and to the north of Ireland.
Variation in percentage sand is indicated by a colour gradient representing a range from 0 - 80. Sandy sediments predominate in both in the North Sea and offshore west coast areas, except for a small offshore region of the Scottish east coast, in inshore waters of the west coast and off the continental shelf.
Variation in percentage silt is indicated by a colour gradient representing a range from 0 - 80. Silty sediments show a patchy distribution, predominating in an offshore region off the north-east coast of Scotland, within inshore regions of the west coast and off the continental shelf.
Species Distribution Modelling
Two separate Species Distribution Models (SDM) were fitted to the data and their predictions were then combined in an ensemble model. The explanatory variables for both models were the retained environmental variables described above and the gear type.
The first model was fitted using a spatial Gaussian random field generalized linear mixed effects model (GLMM) with the R package sdmTMB (Anderson et al., 2022). The GLMM took the form of a hurdle model, with in a first part, presence absence data modelled with a binomial distribution (and logit link) and a spatial random field and in the second part, abundance data were modelled using a Gamma distribution (and log link) and a spatial random field. For both the binomial and gamma parts, the full models included all variables with second order polynomial terms. Covariate selection was based on deviance explained, AIC and cross-validated Root Mean Square Error (RMSE).
The second model was fitted using Random Forest regression (Liaw and Wiener, 2002). Variable selection on the Random Forest model was based on the percentage increase in Mean Square Error (MSE). All variables with a positive percentage increase in MSE were retained in the model.
For both models, predictions were made as number of fish caught per hour in a standard IBTS survey (with GOV trawl as fishing gear). Five-fold cross-validation was used to calculate RMSE for each split and model and the predictions of each model were then combined in an ensemble model using average RMSE (for the 5 splits) for each SDM (GLMM or Random Forest) such as:
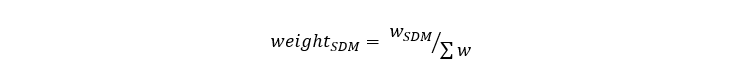
Equation 1
Click for a full description
Relative weight given to a SDM is the weight of the SDM divided by the sum of weights of both SDMs (GLMM and Random Forest).
Where w is the sum of average RMSE for the GLMM and Random Forest models and:
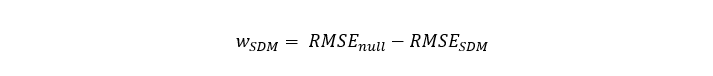
Equation 2
Click for a full description
A given SDM weight is the difference between the Root Mean Square Error (RMSE, from cross-validation) of the null model and the SDM considered.
Prediction for the cell at longitude I and latitude j on the prediction grid was then calculated as:
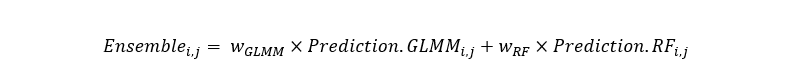
Equation 3
Click for a full description
Predicted abundance at latitude i and longitude j on the prediction grid is the sum of the products of relative weight (Equation 1) from the GLMM by prediction from the GLMM for latitude i and longitude j and relative weight (Equation 1) from the Random Forest by prediction from the Random Forest model for latitude i and longitude j.
2.4 Predicting bycatch hotspots
Information on the distribution and intensity of fishing activity using mobile bottom contacting gears (namely the otter trawl and dredge) in Scottish waters was obtained from ICES, which has produced spatial data layers on fishing intensity/pressure within the OSPAR Maritime Area annually from 2009. These layers were requested by OSPAR to assess benthic impact, underpinning Indicator BH3. Data were collated from relevant vessel monitoring (VMS) and fisher’s logbook data, and further based on the fishing pressure estimated by gear métiers (Table 5), following the approach of Eigaard et al. (2016), to the spatial resolution of single c-squares (0.05° x 0.05°, ~ 15 km2).
Table 5. Glossary of ICES métier groupings, based on Eigaard et al., 2016.
| High-level métier | BENTHIS métier | Description |
|---|---|---|
| Total | - | All métier groups combined |
| Dredge | DRB_MOL | Dredge for scallops and mussels |
| Otter Trawl | OT_CRU | Otter trawl for Nephrops or shrimp |
| - | OT_MIX_CRU | Otter trawl for mixture of species with focus on shrimp |
| - | OT_MIX_CRU_DMF | Otter trawl for Nephrops and mixed fish |
In this study, swept-area ratio (SAR) was taken as the measure of fishing intensity, defined as the swept-area divided by the surface area of the grid cell. Anonymity of individual vessel activity was assured by the classification into categories for c-squares with data from two vessels or fewer. The layers of fishing distributions were clipped to the same spatial extent as for the elasmobranch distribution boundaries, and data averaged for the period 2009-2020 for each c-square in Q-GIS.
Thus, average fishing intensity at the c-square level for the candidate fisheries (Nephrops and scallop) were calculated for the period 2009-2020. Relative fishing intensity was then calculated following the method described in Roe et al. (2014) as:
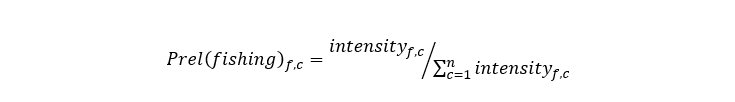
Equation 4
Click for a full description
Relative fishing intensity for fishery f in c-square c is the fishing intensity for fishery f in c-square c divided by the total fishing intensity for fishery f across all c-squares.
Where intensityf,c is the fishing intensity for fishery f in c-square c.
Predicted distributions for each elasmobranch species were then expressed at the c-square level by averaging the predictions of all cells falling within a given c-square (15 prediction cells contained within a c-square on average). Relative abundances were then calculated as:
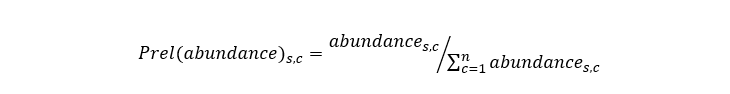
Equation 5
Click for a full description
Relative predicted abundances for species s in c-square c is the abundance of species s in c-square c divided by the total abundance for species s across all c-squares.
Where abundances,c is the abundance of species s in c-square c.
Finally, the relative overlap between fishery f and species s in c-square c was calculated as:
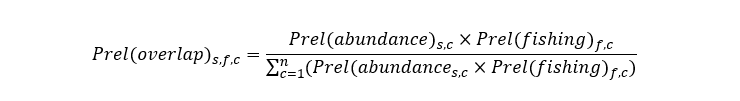
Equation 6
Click for a full description
The relative overlap of fishery f and species s in c-square c is the product of the relative abundance of species s in c-square c and the relative intensity of fishery f in c-square c divided by the sum of the products of the relative abundance of species s and the relative intensity of fishery f across all c-squares.
The distribution of predicted bycatch hotspots (i.e. overlap) was then represented on a map and qualitatively assessed with regard to both species and fisheries distributions.
2.5 Species richness and diversity indices
Using the predicted abundance measures for all considered species, measures of elasmobranch species richness and diversity were calculated for each cell of the prediction grid covering the study area. Species richness was defined as the total number of elasmobranch species present in a standard 1h IBTS tow in a given cell. A threshold value of 0.5 fish caught per hour was used to identify a given species as present (0.5 or above) or absent (<0.5). Elasmobranch diversity was estimated using the Shannon Diversity Index calculated as:
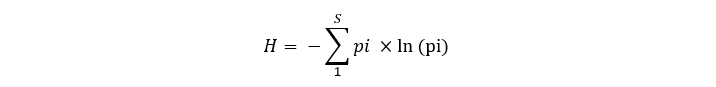
Equation 7
Click for a full description
Diversity Index is minus the sum of the proportion of individuals that belong to species i (number of individuals of species i divided by the total number of individuals of all species s) multiplied by the natural log of this proportion.
Where S is the total number of species (9) and pi is the ratio of the abundance of species i on the total elasmobranch abundance.
As both measures of richness and diversity calculated here can be biased when the models predict low abundances (non-zero) for a number of species (high richness) and high abundances for one or a few species (low diversity), areas of both high elasmobranch richness and diversity were also characterised. Such areas were identified by extracting the areas corresponding to the intersection between a layer of high elasmobranch richness (with only richness values > 3 species were considered) and a layer of high elasmobranch diversity (H values > 2 were considered). This analysis was done using the software QGIS 3.20.2.
3. Results
Variable importance in the Random Forest model for each species is given in Table 6, and relative weights given to the individual SDMs in the ensemble model are given in Table 7.
Table 6. Variable importance measured as the percentage increase in mean square error (when the variable is omitted) in the Random Forest models for each species. The 3 most important variables are indicated in bold.
| Covariate | Flapper skate | Cuckoo ray | Spotted ray | Starry ray | Thorn-back ray | Spurdog | Black mouth dogfish | Lesser spotted dogfish | Starry smooth- hound |
|---|---|---|---|---|---|---|---|---|---|
| Depth | 0.102 | 0.359 | 0.591 | 0.335 | 0.274 | 0.690 | 0.157 | 1.164 | 0.170 |
| % Gravel | 0.076 | 0.315 | 0.464 | 0.422 | 0.211 | 0.548 | 0.08 | 1.169 | 0.136 |
| % Sand | 0.080 | 0.195 | 0.394 | 0.357 | 0.269 | 0.382 | 0.057 | 0.822 | 0.135 |
| Salinity | 0.177 | 0.305 | 0.725 | 0.385 | 0.436 | 0.818 | 0.164 | 1.700 | 0.110 |
| Current velocity | 0.061 | 0.124 | 0.196 | 0.185 | 0.156 | 0.300 | 0.039 | 0.647 | 0.042 |
| Light intensity | 0.062 | 0.278 | 0.319 | 0.417 | 0.197 | 0.448 | 0.072 | 0.773 | 0.110 |
| Temperature | 0.178 | 0.271 | 0.952 | 0.631 | 0.614 | 0.978 | 0.204 | 2.488 | 0.135 |
| Primary productivity | 0.100 | 0.334 | 0.504 | 0.357 | 0.333 | 0.761 | 0.126 | 1.085 | 0.127 |
| Fishing gear | 0.022 | 0.304 | 0.145 | 0.203 | 0.247 | 0.078 | 0.028 | 0.802 | 0.082 |
Table 7. Weight given to the individual SDMs in the ensemble approach.
| Common name | Latin name | GLMM weight | RF weight |
|---|---|---|---|
| Flapper skate | Dipturus intermedius | 0.508 | 0.492 |
| Cuckoo ray | Leucoraja naevus | 0.541 | 0.459 |
| Spotted ray | Raja montagui | 0.542 | 0.458 |
| Starry ray | Amblyraja radiata | 0.487 | 0.513 |
| Thornback ray | Raja clavata | 0.526 | 0.474 |
| Spurdog | Squalus acanthias | 0.437 | 0.563 |
| Black mouth dogfish | Galeus melastomus | 0.493 | 0.507 |
| Lesser spotted dogfish | Scyliorhinus canicula | 0.565 | 0.435 |
| Starry smooth-hound | Mustelus asterias | 0.591 | 0.409 |
3.1 Flapper skate
3.1.1 Species overview
The flapper skate (Dipturus intermedius) is one of the largest and most endangered elasmobranch species in the North-East Atlantic, its former distribution is now highly contracted, only occasionally found in the northern North Sea and in a number of relic inshore populations off the north-west coast of Scotland (Brander, 1981; Walker and Hislop 1998; Daan et al., 2005; Neat et al., 2015; Benjamins et al., 2018). Model predictions suggest that environmental suitability for the flapper skate appears to be driven by both depth and distance from the coast, with a high probability of presence seen in inshore waters around the Outer Hebrides, inner Minches and west coast sea lochs but also around offshore islands and seamounts (Pinto, 2015). They are considered largely benthic, occurring on soft, sandy sediments and coarser shell/gravel seabeds (Benjamins et al., 2018). Based on evidence from tagging studies, the probability of its occurrence is highest between 100 - 400 m depth (Pinto et al., 2016), with a preference for deep trenches, with localised core depth ranges between 50 -180 m (Neat et al., 2015; Thorburn et al., 2018a). Such studies have evidenced limited dispersal rates, with site fidelity apparent in mature individuals (Neat et al., 2015). Primary productivity and temperature appear additional environmental drivers of distribution (Bache-Jeffreys et al., 2021) and the species appears negatively influenced by strong tidal currents, based on anecdotal observations by anglers (Benjamins et al., 2018).
The species has a relatively low fecundity, a long gestational period (Du Buit, 1977; Benjamins et al., 2021) and is thought to use egg nurseries (Dodd et al., 2022). These traits, combined with a large juvenile and adult body size and coastal preference, leave the species highly vulnerable to anthropogenic pressures, particularly commercial exploitation. Flapper skate are taken as incidental bycatch in demersal (particularly bottom trawl and dredge) and net fisheries and are a direct target for recreational anglers. Resting behaviour, where they settle on the seabed for prolonged periods (Pinto et al., 2016; Wearmouth and Sims, 2009), makes them significantly more vulnerable to dredging activity. Gear modifications designed to mitigate bycatch (such as removal of the tickler chain from trawls and dredges, Kynoch et al., 2015) may be more effective during spring when active swimming is less prevalent (Thorburn et al., 2022).
The species was mistakenly synonymised with the smaller blue skate, Dipturus flossada, and regarded as a single species (common skate Dipturus batis) over much of the 20th century until genetic evidence presented in 2009 forced a taxonomic revision (Iglésias et al., 2010). However, the complex of the two distinct species remains for reported landings. This confusion has challenged accurate species-specific assessment and, in the absence of reliable survey and catch data, leaves ICES unable to provide current advice on the status of the stock. The most recent ICES advice for North Sea follows earlier advice, since 2008, that the species complex remains depleted (ICES, 2023a).
A precautionary approach to management has been taken for the common skate complex. EU wide fishing regulations have prohibited landings since 2009 and anglers are subject to catch-and-release directives. Discard and discard survival rates remain unquantified, a common complication for managers in species with a zero TAC. Since 2006, the common skate complex is considered to be ‘Critically Endangered’ both globally and in European waters by the IUCN (Ellis et al., 2021), is listed under The Sharks, Skates and Rays (Prohibition of Fishing, Trans-shipment and Landing) (Scotland) Order 2012 and is on the Priority Marine Feature list in Scotland. The “common skate” complex is listed under Annex V of OSPAR regulations (Fowler, 2010), with a UK Biodiversity Action plan also in place (BAP, 2011). Two protected areas in Scotland offer regional spatial management specifically for the flapper skate, the Loch Sunart to the Sound of Jura Marine Protected Area (LStSJ MPA) (designated in 2014) and the Red Rocks and Longay MPA within the Sound of Skye (permanently designated in 2023). While catch rates of species in the complex have increased in fisher-independent surveys since the prohibition (ICES, 2020; Rindorf et al., 2020), these tentative positive trends were not thought to be related to species-specific management measures (Rindorf et al., 2020) given that fishery pressures remain high, with no quantitative evidence to suggest stock recovery (Garbett et al. 2020; Rindorf et al. 2020). However, a more recent study (Régnier et al., 2024) provided empirical evidence of a substantial increase in skate abundance, especially for young juveniles, within the LStSJ MPA following the introduction of fishing restrictions. As the scale of this trend was not mirrored in the wider west of Scotland area, it appears to demonstrate a local effect of restrictions on fishing pressure.
3.1.2 Distribution model
Survey data incorporated into the distribution model for flapper skate are shown in Table 8 and Figure 4.
Table 8. Sources of abundance data for flapper skate Dipturus intermedius. Total number of records used in SDM is 5,338.
| Datasource | Years | No. surveys | No. records |
|---|---|---|---|
| ICES DATRAS | 2010-2022 | 43 | 3,951 |
| MD FSS | 2013-2022 | 28 | 1,352 |
| MD BRUV | 2019, 2022 | 3 | 14 |
| MD Scallop | 2019-2022 | 7 | 21 |
The binomial part of the GLMM included a linear effect of temperature and primary productivity and second order polynomial terms for depth and % sand, % gravel as well as effects of gear type (Figure 8). The probability of presence was maximised at both low and high % sand, intermediate % gravel and depths between 300-500 m, however the confidence intervals for higher depth and % gravel were extremely large. Presence increased with temperature and decreased with primary productivity. High probabilities of presence were associated with the dredge.
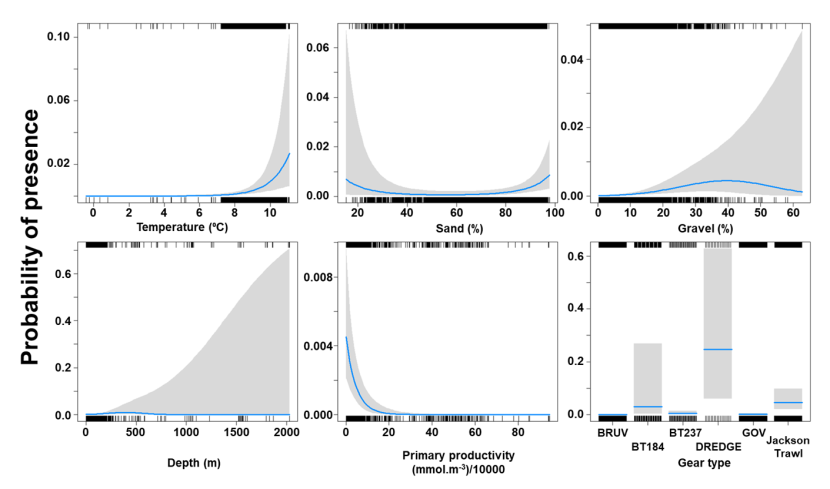
Figure 8. Predicted relationships between variables retained in the binomial part of the GLMM and probability of presence of Dipturus intermedius.
Click for a full description
Five individual line charts of the relationship between the probability of flapper skate presence – ranging from 0 to 0.6 – and environmental variables, including:
- temperature on a scale from 0 to 10 degrees Celsius
- percentage sand on a scale from 20 to 100
- percentage gravel on a scale from 0 to 60
- depth on a scale from 0 to 2000 m
- primary productivity on a scale from 0 to 80 mmol per meter cubed divided by 10000
A single histogram depicts the relationship between the probability of flapper skate presence – ranging from 0 to 0.6 - and six gear types, including: BRUV, BT184, BT237, Dredge, GOV and Jackson Trawl.
Refer to Section 3.1.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included a linear effect of the light intensity and second order polynomial terms for depth as well as gear effects. Abundance was maximised at high bottom light intensities and depths around 200 m (Figure 9). The largest abundances were associated with bottom trawl BT184 (Figure 9).
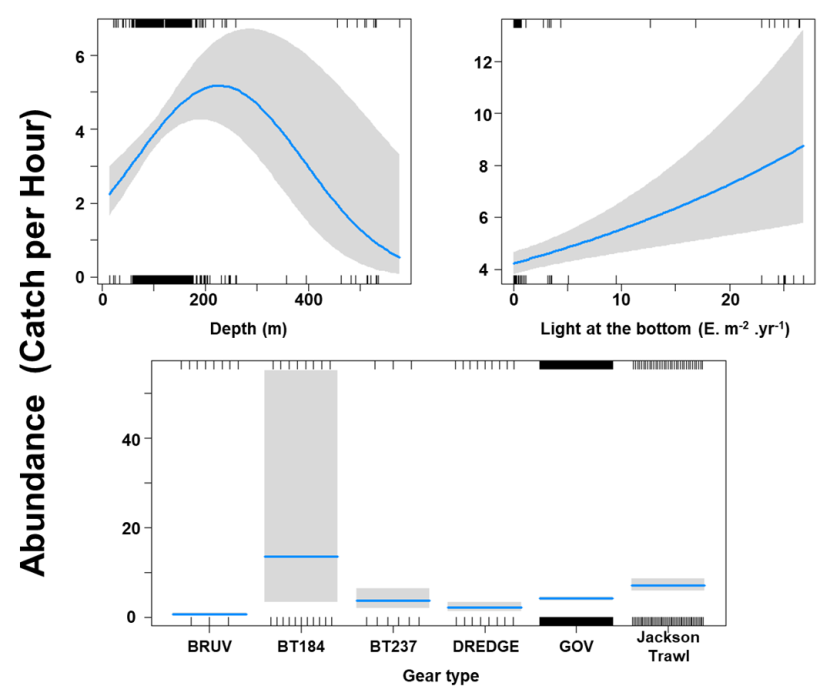
Figure 9. Predicted relationships between variables retained in the Gamma part of the GLMM and probability of abundance of Dipturus intermedius (catch per unit hour).
Click for a full description
Two individual line charts of the relationship between flapper skate abundance – catch per hour ranging from 0 to 12 – and two environmental variables, including:
- depth on a scale from 0 to just over 500 m.
- light intensity at the sea bottom on a scale from 0 to just over 25 intensity per meter squared per year.
A single histogram depicts the relationship between flapper skate abundance – catch per hour ranging from 0 to 50 - and six gear types, including: BRUV, BT184, BT237, Dredge, GOV and Jackson Trawl.
Refer to Section 3.1.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, salinity and depth (Table 6). Partial dependence of the model on survey gear type is provided in Annex A1 and shows that Jackson trawl and dredge are the two most important gear influencing flapper skate abundance.
The predicted distribution of Dipturus intermedius from the ensemble model is presented in Figure 10 and the relative weights given to the models in Table 7. D. intermedius is predicted along the Scottish shelf edge from Northern Ireland to the Northern Isles but also shows a strong presence in the deep inshore waters of the west coast of Scotland. In particular, the deep trenches found in the Minch, the inner Sound of Skye, around the Small Isles, the Sound of Jura and the Firth of Lorn and in the Northern channel represent predicted areas of high abundance for this species. D. intermedius is also found in the deeper part of the Clyde estuary. While predicted on the east coast of Scotland, D. intermedius show no significant high abundance areas in this region (Figure 10).
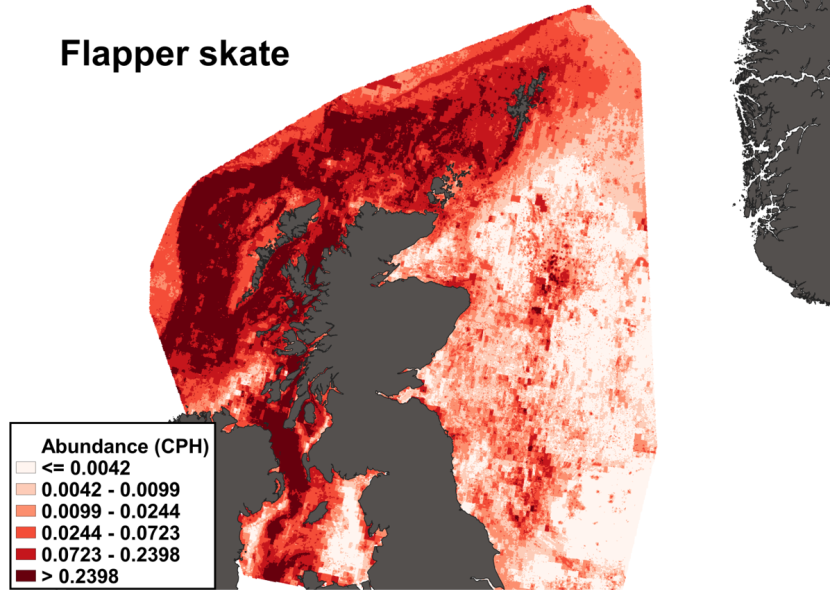
Figure 10. The predicted catch per unit hour of Dipturus intermedius in a standard IBTS survey from the ensemble SDM. Variation in abundance is represented by a colour ramp.
Click for a full description
A heatmap of predicted flapper skate abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.0042 to greater than 0.2398. Refer to Section 3.1.2 in the main body of text for full description of the distribution.
3.1.3 Predicted bycatch hotspots Nephrops
The estimated overlap between D. intermedius predicted distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas in the Minch (particularly leading to the Inner Sound of Skye) and in the Clyde show the highest levels of overlap between this fishery and D. intermedius distribution (Figure 11).
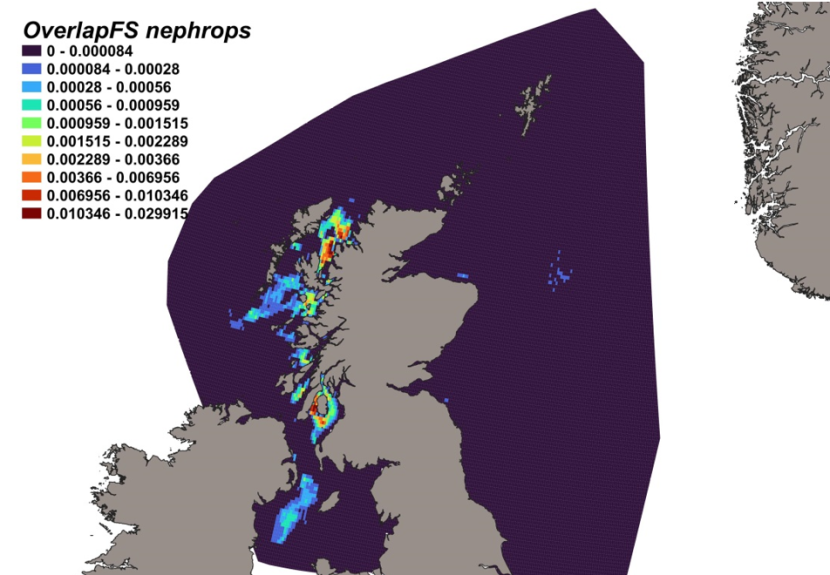
Figure 11. Predicted bycatch hotspots for Dipturus intermedius in the Nephrops fishery.
Click for a full description
A heap of the overlap between the distributions of flapper skate and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.029915. Refer to Section 3.1.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.1.4 Predicted bycatch hotspots Scallops
The calculated overlap between D. intermedius predicted distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots were forecast in the Firth of Lorn, the Sound of Jura and to the south of the Kintyre peninsula (Figure 12).
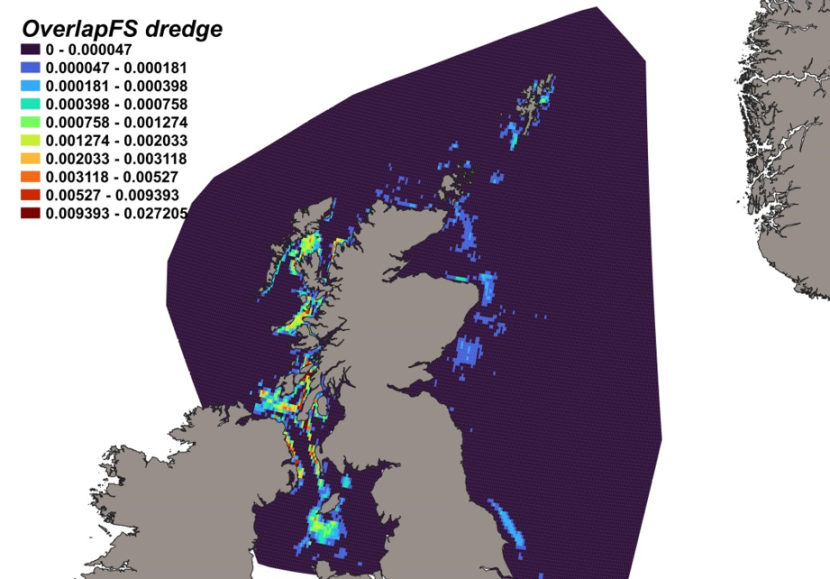
Figure 12. Predicted bycatch hotspots for Dipturus intermedius in the scallop fishery.
Click for a full description
A heatmap of the overlap between the distributions of flapper skate and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.027205. Refer to Section 3.1.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.2 Cuckoo ray
3.2.1 Species overview
The cuckoo ray, Leucoraja naevus, is a medium sized demersal species, widely distributed in the North-East Atlantic, including the North, Irish and Celtic Seas, and Mediterranean, typically offshore on the continental shelf and slope. It is associated with depths between 20-500 m (most commonly encountered around 200 m), and on coarse sediments, soft sands and muddy seabeds (Ebert and Stehmann, 2013; Moriarty, 2021). Previous environmental modelling of their distribution indicated suitable habitat along the coastlines of the Celtic Seas, and less suitable habitat in the Irish and North Seas, with potentially suitable habitats in offshore deeper waters (Moriarty, 2021). Temperature has been shown to explain more than 50 % of the variance in habitat suitability in this species (Coulon et al., 2024).
A relatively short-lived species, the cuckoo ray matures between 4-7 years old (Gallagher et al., 2005; Walker, 1999), spawning all year round with an estimated maximum fecundity of 60-90 eggs per year (Walker, 1999; Maia et al., 2012). While not directly targeted, the species is often caught as bycatch in mixed demersal commercial fisheries for roundfish and flatfish through much of its range, and discard levels, while unquantified, are thought to be high (ICES, 2018).
Biomass indices suggest an increase in stock size in the Greater North Sea region between 2004 – 2012, followed by marked inter-annual variability and evidence of decreasing stock size since 2017 until 2022 (ICES, 2023b). Relative biomass in the western stock has shown a steady increase since 2005 (ICES, 2022b).
As with other data-poor species, North-East Atlantic stocks are managed subject to a maximum sustainable yield approach (ICES, 2023). Further protections were introduced for the declining western stock which extends from the north-west of Scotland, Ireland, Irish and Celtic Seas and into the English Channel. This stock has seen a 50 % decline in abundance indicators, based on survey index trend, since the 1990s, providing evidence of its over exploitation. This localised decline has resulted in the species being designated as ‘Vulnerable’ in the IUCN red list in Ireland (under category A2bd; Clarke et al., 2016), but remains assessed as ‘Least Concern’ globally. ICES has advised maximum landings of less than 79 and 8,064 tonnes for 2024 in the North Sea and Western stocks respectively (ICES, 2022b; 2023b).
3.2.2 Distribution model
Survey data incorporated into the distribution model for cuckoo ray are shown in Table 9 and Figure 4.
Table 9. Sources of abundance data for cuckoo ray Leucoraja naevus. Total number of records used in SDM is 35,995.
| Datasource | Years | No. surveys | No. records |
|---|---|---|---|
| ICES DATRAS | 1980-2022 | 178 | 29,589 |
| MD FSS | 2013-2022 | 13 | 1,231 |
| MD RVR | 1980-1999 | 21 | 665 |
| MD SCALLOP | 1994-2022 | 85 | 4,510 |
The binomial part of the GLMM included linear effects of primary productivity, salinity, depth, temperature, % gravel and sand as well as effects of gear type (Figure 13). The probability of presence increased with salinity, temperature, % gravel and sand but decreased with depth and primary productivity. High probabilities of presence were associated with the dredge, BT196 and Jackson trawl (Figure 13).
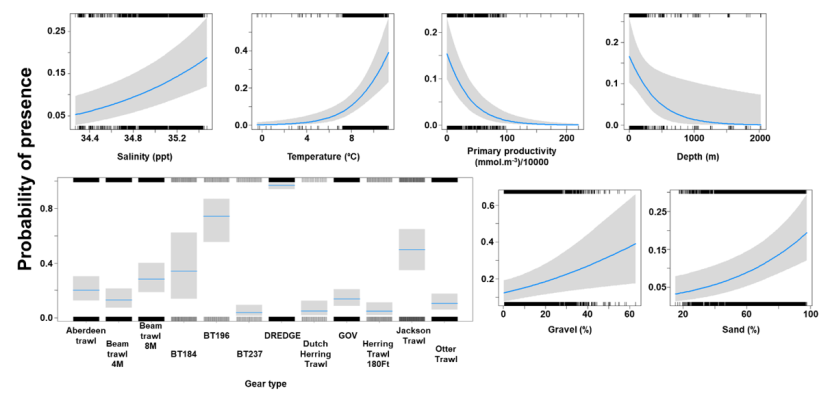
Figure 13. Model plot for the Binomial part of the GLMM for Leucoraja naevus.
Click for a full description
Six individual line charts of the relationship between the probability of cuckoo ray presence – ranging from 0 to 0.6 – and environmental variables, including:
- salinity on a scale from 34.4 to 35.4 ppt
- temperature on a scale from 0 to 10 degrees Celsius
- primary productivity on a scale from 0 to 200 mmol per meter cubed - divided by 10000
- depth on a scale from 0 to 2000 m
- percentage gravel on a scale from 0 to 60
- percentage sand on a scale from 20 to 100
A single histogram depicts the relationship between the probability of cuckoo ray presence – ranging from 0 to 0.8 - and 12 gear types, including Aberdeen Trawl, Beam Trawl 4m, Beam Trawl 8m, BT184, BT196, BT237, Dredge, Dutch Herring Trawl ,GOV, Herring Trawl 180ft, Jackson Trawl, Otter Trawl.
Refer to Section 3.2.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included linear effects of % sand and primary productivity and a second order polynomial term for % gravel as well as gear effects. Abundance decreased with an increase in primary productivity and was maximised over substrates composed of gravel at 30 % and high proportions of sand (Figure 14). The largest abundances were associated with the beam trawl (8 m), BT196 and Jackson trawl (Figure 14).
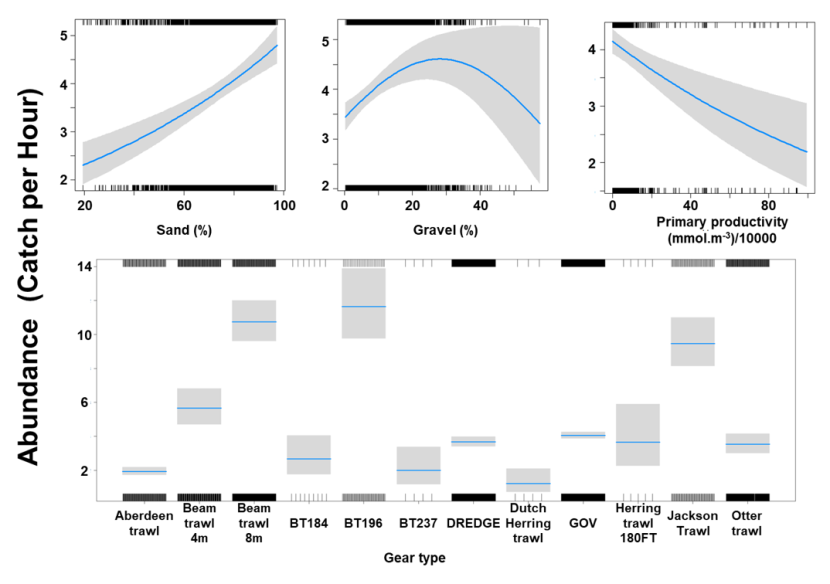
Figure 14. Model plot for the Gamma part of the GLMM for Leucoraja naevus.
Click for a full description
Three individual line charts of the relationship between cuckoo ray abundance – catch per hour ranging from 0 to 5 – and environmental variables, including:
- percentage sand on a scale from 20 to 100
- percentage gravel on a scale from 0 to 60
- primary productivity on a scale from 0 to 100 mmol per meter cubed - divided by 10000
A single histogram of the relationship between cuckoo ray abundance – catch per hour ranging from 0 to 14 - and 12 gear types, including: Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8m, BT184, BT196, BT237, Dredge, Dutch Herring Trawl, GOV, Herring Trawl 180ft, Jackson Trawl, Otter Trawl.
Refer to Section 3.2.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were depth, primary productivity and % gravel (Table 6). Partial dependence of the model on survey gear type is provided in Annex A2 and shows that BT196 and dredge are the two most important gear influencing cuckoo ray abundance.
The predicted distribution of Leucoraja naevus from the ensemble model is presented in Figure 15 and the relative weights given to the models in Table 7. L. naevus is predicted around Scotland in shallow waters, west of the Outer Hebrides, in the Sea of Hebrides, in the south of the Sound of Jura, in the northern part of the North Channel, in the Solway Firth and along the north and east coast of Scotland and south of Shetland. (Figure 15).
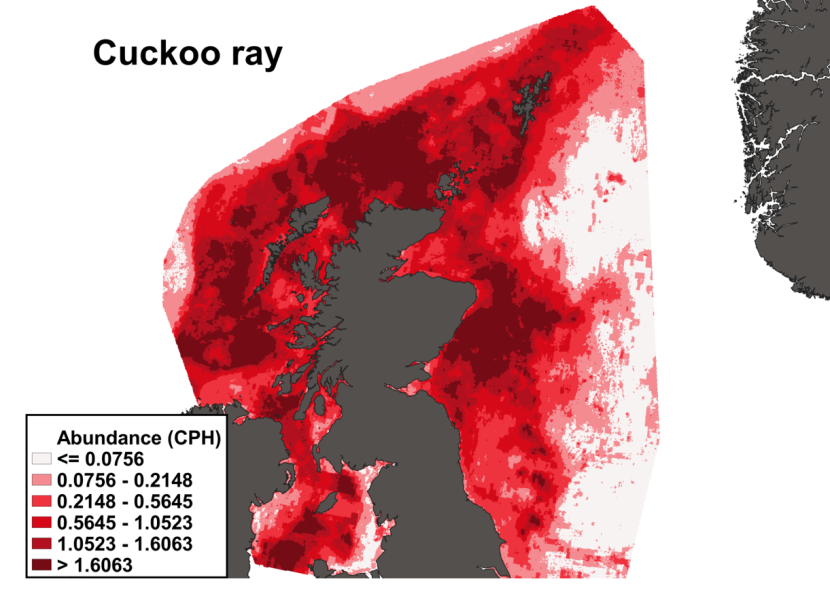
Figure 15. Predicted distribution of Leucoraja naevus from the ensemble SDM.
Click for a full description
A heatmap of predicted cuckoo ray abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.0756 to greater than 1.6063. Refer to Section 3.2.2 in the main body of text for full description of the distribution.
3.2.3 Predicted bycatch hotspots Nephrops
The estimated overlap between L. naevus predicted distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas to the north of the Minch, around the Small Isles, in the Sound of Jura and the Clyde show the highest predicted risk of bycatch to the west of Scotland while the Firth of Forth show the highest level of overlap between this fishery and predicted L. naevus distribution on the east coast (Figure 16).
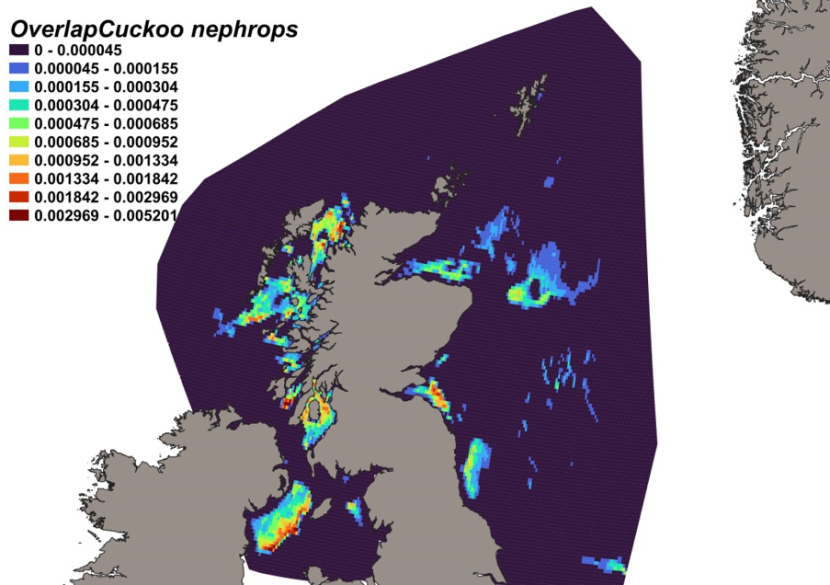
Figure 16. Predicted bycatch hotspots for Leucoraja naevus in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of cuckoo ray and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.005201. Refer to Section 3.2.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.2.4 Predicted bycatch hotspots Scallops
The calculated overlap between L. naevus distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots on the west of Scotland were predicted in the Sound of Jura, near Coll and Tiree and in the Solway Firth. The most significant predicted risk of bycatch is however on the east of Scotland on Turbot Bank and southerly towards the Firth of Forth (Figure 17).
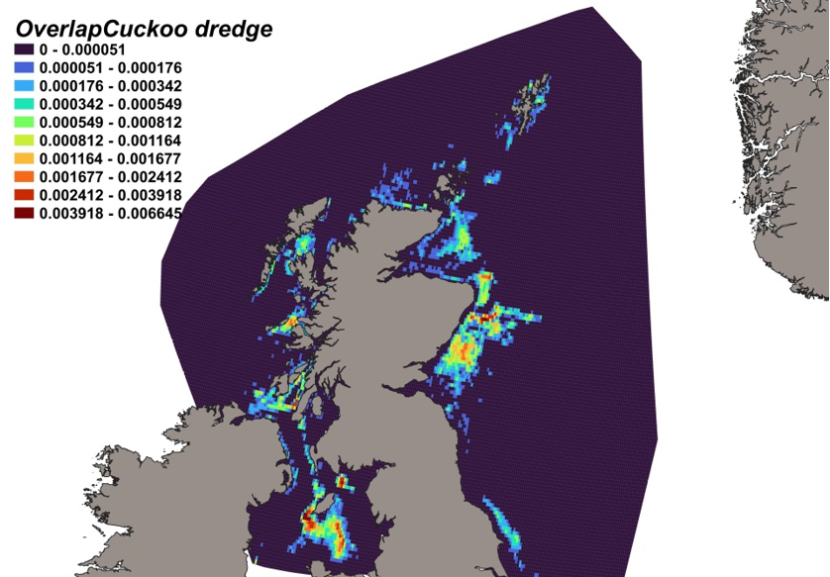
Figure 17. Predicted bycatch hotspots for Leucoraja naevus in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of cuckoo ray and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.006645. Refer to Section 3.2.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.3 Spotted ray
3.3.1 Species overview
The spotted ray, Raja montagui, one of the smallest species in the Rajidae family, is widespread in the North-East Atlantic and Mediterranean Sea and tends to occur on sandy and sandy-gravel substrates (Serena, 2005) in inshore waters and shelf seas, at preferred temperatures of 7 – 16 °C and depths from 8 – 283 m (Ellis et al., 2005a), but most commonly up to 120 m. Significant predictors of occurrence include depth, substrate type (fine sand, mud, gravel, pebbles), bed sheer stress, sea surface temperature and salinity (Martin et al., 2012). Data relating to life history and population structure is poor and spawning and nursery grounds are yet to be fully delineated, but age associated habitat use is suggested by the presence of juveniles in inshore areas and adults being more common further offshore. Catch data is also uncertain, as the species has been misidentified as both R. brachyura and R. clavata (Ellis et al., 2007). Like other elasmobranchs, the spotty ray has limited commercial value but is a common part of bycatch in commercial fishing, notably by beam and bottom trawl fisheries and more recently by seine fisheries. They are, however, a target for recreational fishers (usually catch and release). The spotted ray is likely to be less sensitive to overexploitation than other ray species, with traits such as higher fecundity (with 60-70 egg capsules deposited over one spawning season), low length at first maturity, higher growth rate and smaller overall size possibly offering some resilience to fishing impacts.
Indeed, in OSPAR regions II, III and IV, stock-size indicators show an increasing trend, with Region II (primarily to the western North Sea) showing a more pronounced increase above the long-term average. The biomass of spotted ray in the northern part of the North Sea has substantially increased since the early 2000s. Given the lack of knowledge on population structuring, ICES still advises a maximum sustainable yield approach to landings, with a maximum of 36 tonnes to be landed in each of the years 2023 and 2024 for the west of Scotland/west and south-west Ireland area (ICES, 2023c), and ≤ 1517 and ≤ 1415 for 2024 and 2025 respectively for the North Sea, Skagerrak, Kattegat and eastern English Channel area (ICES, 2023d). The conservation status of the spotted ray is currently listed as ‘Least Concern’ on the IUCN and European Red Lists (Nieto et al., 2015).
3.3.2 Distribution model
Survey data incorporated into the distribution model for the spotted ray are shown in Table 10 and Figure 4.
Table 10. Sources of abundance data for spotted ray Raja montagui. Total number of records used in SDM is 47,758.
| Datasource | Years | No. surveys | No. records per survey |
|---|---|---|---|
| ICES DATRAS | 1980-2022 | 178 | 45,858 |
| MD FSS | 2013, 2015, 2018, 2020-2022 | 9 | 1,045 |
| MD RVR | 1990-1993, 1998-1999 | 10 | 258 |
| MD SCALLOP | 1994-2022 | 54 | 597 |
The binomial part of the GLMM included linear effects of light intensity, salinity, primary productivity, temperature and % sand and second order polynomial terms for depth and % gravel as well as effects of gear type (Figure 18). The probability of presence was maximised at a depth of 100 m and substrates composed of gravel at around 30 %. Probability of presence increased with salinity, temperature, light intensity and % sand but decreased with primary productivity. High probabilities of presence were associated with the dredge (Figure 18).
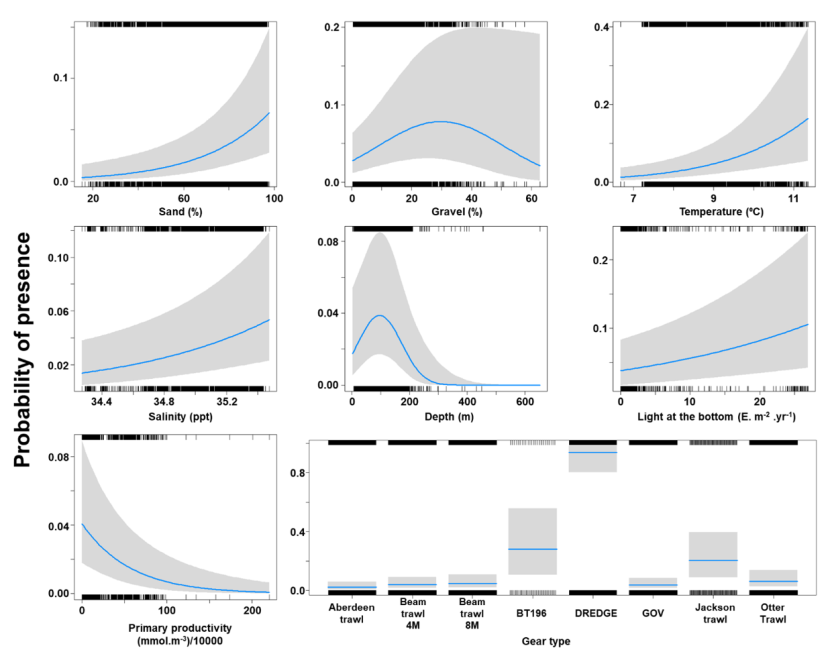
Figure 18. Model plot for the Binomial part of the GLMM for Raja montagui.
Click for a full description
Seven individual line charts of the relationship between the probability of spotted ray presence – ranging from 0 to 0.4 – and environmental variables, including:
- percentage sand on a scale from 20 to 100
- percentage gravel on a scale from 0 to 60
- temperature on a scale from 7 to 11 degrees Celsius
- salinity on a scale from 34.4 to 35.4 ppt
- depth on a scale from 0 to 600 m
- light intensity at the sea bottom on a scale from 0 to 25 intensity per meter squared per year.
- primary productivity on a scale from 0 to 200 mmol per meter cubed - divided by 10000
A single histogram depicts the relationship between the probability of spotted ray presence – ranging from 0 to 1 - and eight gear types, including: Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8 m, BT196, Dredge, GOV, Jackson Trawl, Otter Trawl.
Refer to Section 3.3.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included linear effects of % sand and gravel and temperature and second order polynomial terms for depth, salinity and primary productivity as well as gear effects. Abundance was maximised at a depth of 100 m and intermediate salinity levels. Abundance increased with temperature and % sand and gravel but decreased with primary productivity (Figure 19). The largest abundances were associated with the beam trawl (8 m), BT196, otter trawl and Jackson trawl (Figure 19).
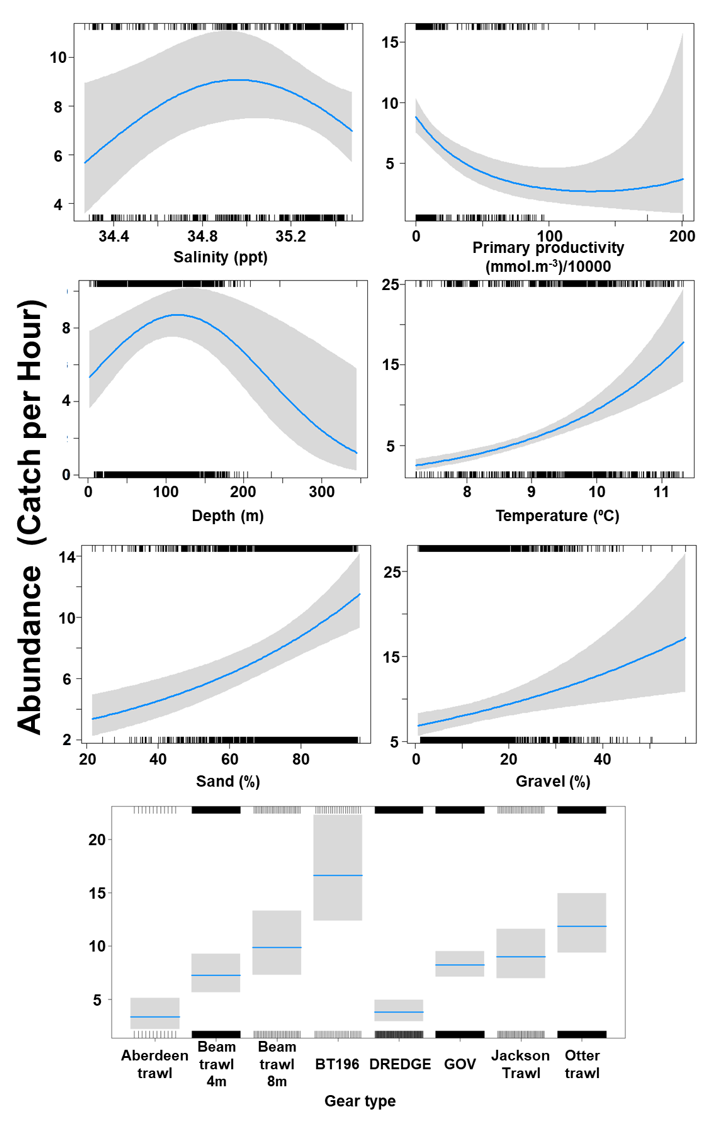
Figure 19. Model plot for the Gamma part of the GLMM for Raja montagui.
Click for a full description
Six individual line charts of the relationship between spotted ray abundance – catch per hour ranging from 0 to 25 – and environmental variables, including:
- salinity on a scale from 34.4 to 35.4 ppt
- primary productivity on a scale from 0 to 200 mmol per meter cubed - divided by 10000
- depth on a scale from 0 to 300 m
- temperature on a scale from 7 to 11 degrees Celsius
- percentage sand on a scale from 20 to 100
- percentage gravel on a scale from 0 to 60
A single histogram of the relationship between spotted ray abundance – catch per hour ranging from 0 to 20 - and eight gear types, including: Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8 m, BT196, Dredge, GOV, Jackson Trawl, Otter Trawl.
Refer to Section 3.3.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, salinity and depth (Table 6). Partial dependence of the model on survey gear type is provided in Annex A3 and shows that BT196 and dredge are the two most important gear influencing spotted ray abundance.
The predicted distribution of Raja montagui from the ensemble model is presented in Figure 20 and the relative weights given to the models in Table 7. R. montagui is predicted on the west of Scotland, from the Irish Sea to Shetland. The distribution is centred on depths around 100 m but R. montagui is predicted inshore in the Sound of Jura, around the Small Isles, in the Minch and on the North coast. Very low abundances are predicted on the east coast of Scotland (Figure 20).
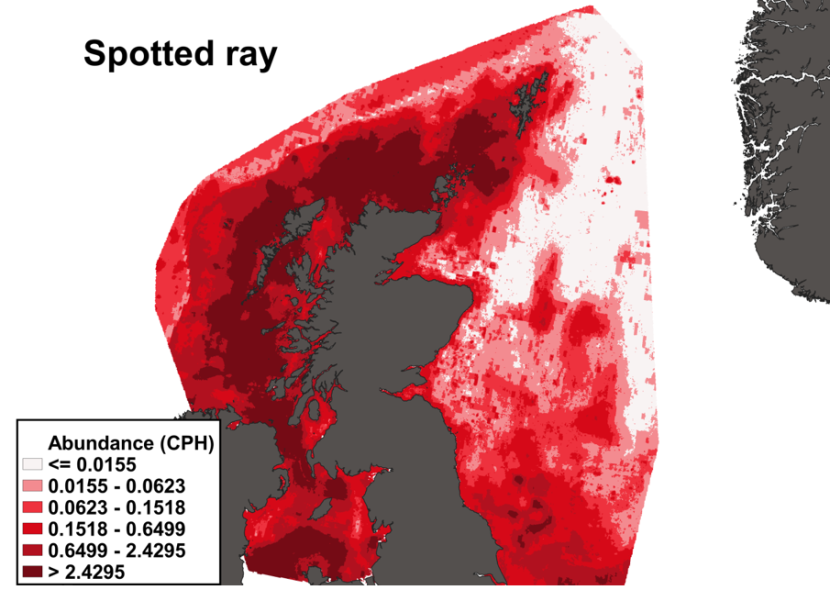
Figure 20. Predicted distribution of Raja montagui from the ensemble SDM.
Click for a full description
A heatmap of predicted spotted ray abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.0155 to greater than 2.4295. Refer to Section 3.3.2 in the main body of text for full description of the distribution.
3.3.3 Predicted bycatch hotspots Nephrops
The estimated overlap between R. montagui distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas in the Sea of Hebrides and the Sound of Jura show the highest predicted risk of bycatch (Figure 21).
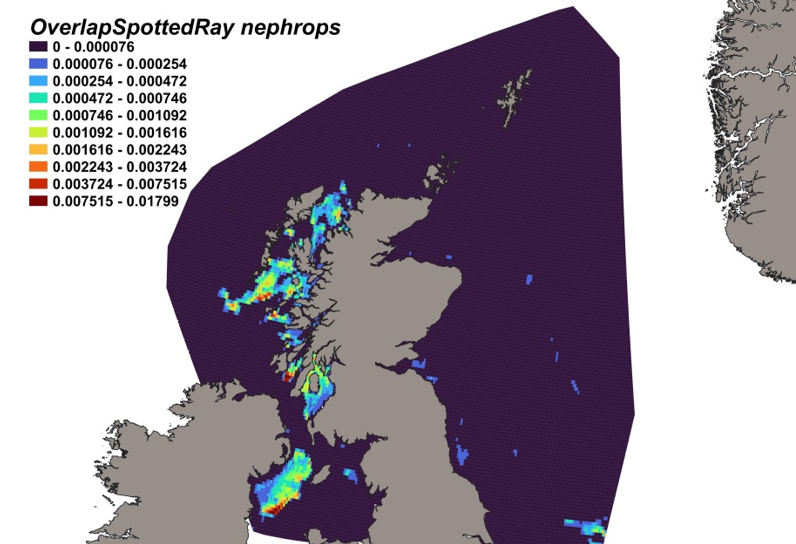
Figure 21. Predicted bycatch hotspots for Raja montagui in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of spotted ray and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.01799. Refer to Section 3.3.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.3.4 Predicted bycatch hotspots Scallops
The calculated overlap between R. montagui distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots on the west of Scotland were predicted in the Solway Firth and the Sound of Jura (Figure 22).
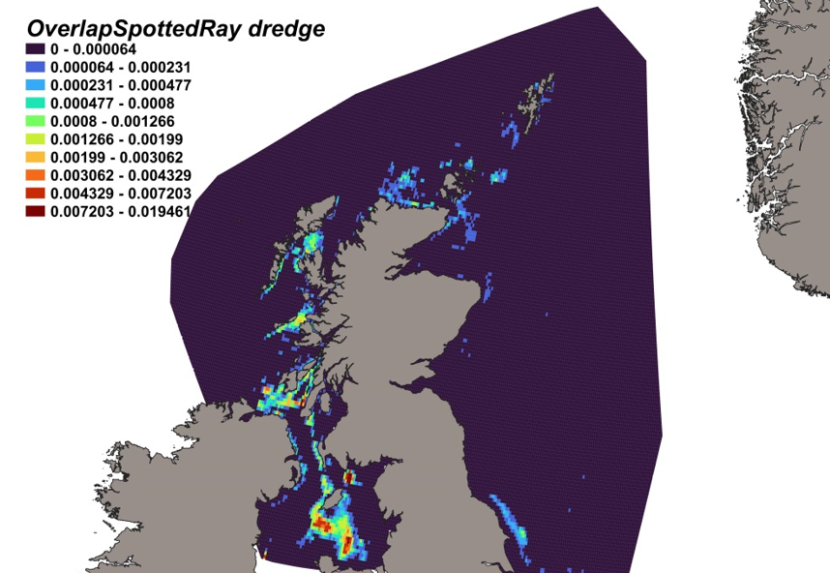
Figure 22. Predicted bycatch hotspots for Raja montagui in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of spotted ray and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.019461. Refer to Section 3.3.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.4 Starry ray
3.4.1 Species overview
The starry ray, or thorny skate, Amblyraja radiata, is a small-bodied elasmobranch that is widespread throughout much of the North-East Atlantic Ocean over wide depth ranges up to 1200 m but found more commonly up to 100 m (McEachran, 2002; Stehmann and Bȕrkel, 1984). Despite being associated with a wide variety of substrates including sand, fragmented shell, gravel, pebbles and soft muds, the species shows a preference for complex, structured habitats with coarser sediments (Scott, 1982; Sosebee et al., 2016). The species has a narrower thermal range than other skate species (Hogan et al., 2013), with a preference for cooler temperatures, but can still tolerate a range in temperatures from -1.4 to 14 °C (McEachran and Muisck, 1975). Abundant catches have been associated with salinities between 32 and 35 practical salinity units (COSEWIC, 2012). This species is not commercially targeted and is only occasionally landed as incidental bycatch in demersal trawl fisheries, but usually discarded (Walker and Heessen, 1996). Its small length at maturity may offer some resilience to fishing pressures. Due to these factors, European stocks are either stable or have increased. Central North Sea stocks showed a marked increase in abundance between from 1980 to a peak in 1992. More recent surveys (1991–2011) indicated a decline in the North Sea, but this is believed to be due to a change in survey gear (Ellis et al., 2005b) rather than an actual change in abundance. Globally, A. radiata has most recently (2019) been assessed as ‘Vulnerable’ under criteria A2bcd in The IUCN Red List of Threatened Species.
3.4.2 Distribution model
Survey data incorporated into the distribution model for the starry ray are shown in Table 11 and Figure 4.
Table 11. Sources of abundance data for starry ray Amblyraja radiata. Total number of records used in SDM is 82,961.
| Datasource | Years | No. surveys | No. records |
|---|---|---|---|
| ICES DATRAS | 1980-2022 | 273 | 80,079 |
| MD FSS | 2013-2022 | 43 | 1,464 |
| MD RVR | 1980-1999 | 35 | 1,342 |
| MD Scallop | 1993-2022 | 28 | 76 |
The binomial part of the GLMM included linear effects of primary productivity, salinity, and % gravel and second order polynomial terms for % sand and temperature as well as effects of gear type (Figure 23). The probability of presence increased with % sand and primary productivity but decreased with salinity, temperature and % gravel. High probabilities of presence were associated with the dredge, Aberdeen and beam trawls (Figure 23).
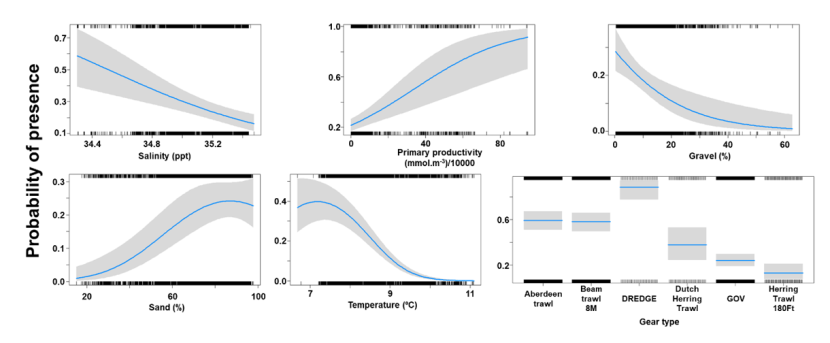
Figure 23. Model plot for the Binomial part of the GLMM for Amblyraja radiata.
Click for a full description
Five individual line charts of the relationship between the probability of starry ray presence – ranging from 0 to 1 – and environmental variables, including:
- salinity on a scale from 34.4 to 35.4 ppt
- primary productivity on a scale from 0 to 90 mmol per meter cubed - divided by 10000
- percentage gravel on a scale from 0 to 60
- percentage sand on a scale from 20 to 100
- temperature on a scale from 7 to 11 degrees Celsius
A single histogram depicts the relationship between the probability of starry ray presence – ranging from 0 to 1 - and six gear types including Aberdeen Trawl, Beam Trawl 8 m, Dredge, Dutch Herring Trawl, GOV, Herring Trawl 180 ft.
Refer to Section 3.4.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included linear effects of % sand and a second order polynomial term for salinity as well as gear effects. Abundance was maximised at intermediate salinities and over substrates with high percentage sand (Figure 24). The largest abundances were associated with the beam trawl (8m); Figure 24.
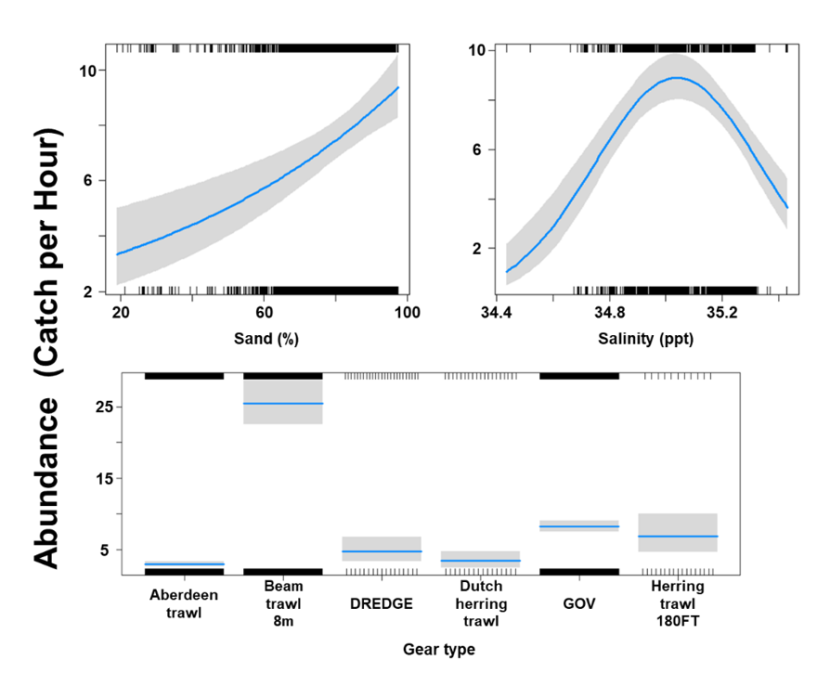
Figure 24. Model plot for the Gamma part of the GLMM for Amblyraja radiata.
Click for a full description
Two individual line charts of the relationship between starry ray abundance – catch per hour ranging from 0 to 10 – and environmental variables, including percentage sand on a scale from 20 to 100 and salinity on a scale from 34.4 to 35.4 ppt.
A single histogram of the relationship between starry ray abundance – catch per hour ranging from 0 to 30 - and six gear types, including: Aberdeen Trawl, Beam Trawl 8 m, Dredge, Dutch Herring Trawl, GOV, Herring Trawl 180 ft.
Refer to Section 3.4.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, % gravel and light intensity (Table 6). Partial dependence of the model on survey gear type is provided in Annex A4 and shows that beam trawl (8 m) and dredge are the two most important gear influencing starry ray abundance.
The predicted distribution of Amblyraja radiata from the ensemble model is presented in Figure 25 and the relative weights given to the models in Table 7. A. radiata is predicted mostly to the east of Scotland in the North Sea in offshore water but some high abundance areas are predicted in the Moray Firth and the Tay estuary (Figure 25).
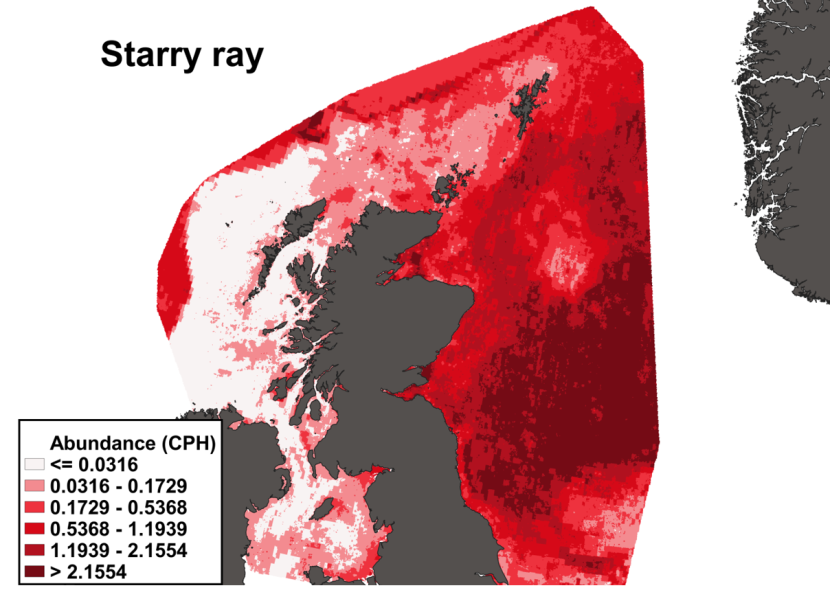
Figure 25. Predicted distribution of Amblyraja radiata from the ensemble SDM.
Click for a full description
A heatmap of predicted starry ray abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.0316 to greater than 2.1554. Refer to Section 3.4.2 in the main body of text for full description of the distribution.
3.4.3 Predicted bycatch hotspots Nephrops
The estimated overlap between A. radiata distribution and the distribution of the Nephrops fishery revealed a few potential bycatch hotspots. Bycatch hotspots were predicted in the Firth of Forth and offshore in the North Sea (Figure 26).
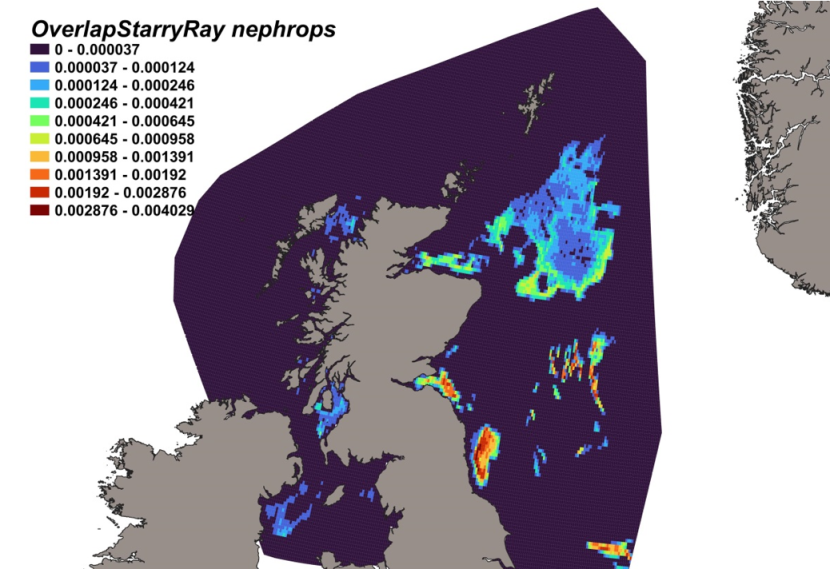
Figure 26. Predicted bycatch hotspots for Amblyraja radiata in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of starry ray and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.004029. Refer to Section 3.4.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.4.4 Predicted bycatch hotspots Scallops
The calculated overlap between Amblyraja radiata predicted distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots are on the east of Scotland from Rattray head to Turbot Bank, extending south to the Forth and Tay region (Figure 27).
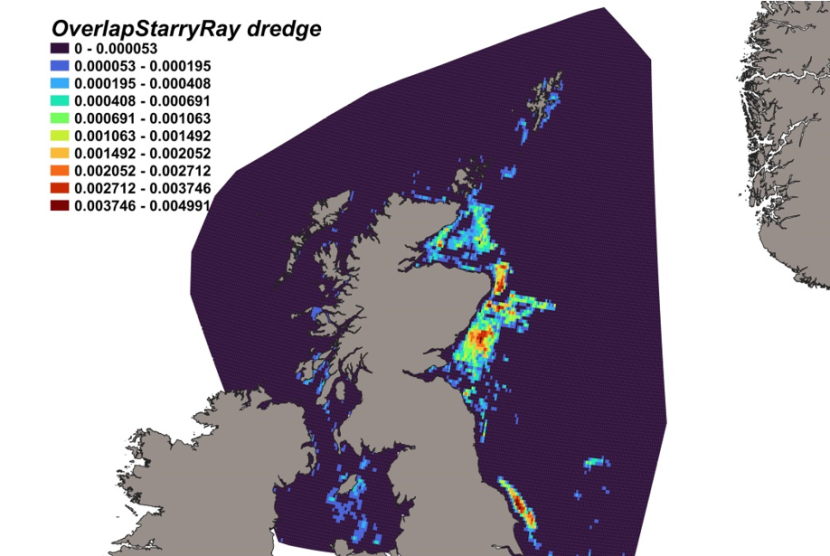
Figure 27. Predicted bycatch hotspots for Amblyraja radiata in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of starry ray and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.004991. Refer to Section 3.4.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.5 Thornback ray
3.5.1 Species overview
The thornback ray (Raja clavata) is a small-bodied demersal skate, widely distributed in the North-East Atlantic and adjacent seas, common all around the coast of Britain and Ireland with a greater abundance in the eastern English Channel and southern North Sea (Ellis et al., 2008; Elliot et al., 2020). The species inhabits mud, sand, shingle and gravel substrates across a wide bathymetric range, although is most abundant in 10-60 m water depth off coastal areas (Ebert and Stehmann, 2013; Ellis, 2016), using shallow waters (< 30 m) particularly during the juvenile stage (Ebert and Stehmann, 2013; McAllister et al., 2023). They tolerate a broad salinity range and probability of presence has been shown to be related to both sea bottom temperature and season (Papadopoulo et al., 2023). Primarily a non-migratory species, R. clavata is known to make small-scale movements to breed and lay eggs in shallower inshore waters during the spring before moving into deeper waters in winter. Their egg laying rate appears sensitive to temperature, with maximum rates observed in warmer conditions (Holden et al., 1971). The life history parameters of this species are relatively well known and are characterised by a relatively fast growth rate, later maturity, low fecundity (although higher relative to other elasmobranch species) and low natural mortality (Gallagher et al., 2005; Whittamore and McCarthy, 2005; Kadri et al., 2014).
On account of these traits, and its abundance, R. clavata is particularly vulnerable to overexploitation when fishing pressure is high (Dulvy and Reynolds, 2002; Abella and Serena, 2005). The thornback ray is a well-known and commercially valuable species, accessible to different fishing métiers such as net fisheries (gill, entangling and trammel nets) and demersal trawl fisheries, particularly as bycatch. Prior to 2009, thornback ray landings were recorded within the generic grouping of “skates and rays”, but species-specific data from fishery-independent surveys in the North Sea evidence a markedly downward trend in long term abundance since the start of the 20th Century (Heessen, 2003) and the species’ range has significantly contracted, with local depletions seen in some areas (ICES, 2007). Despite this susceptibility to fishing pressure, the observed reduction in landings and commercial catches may be more associated with an increase in discards than a decline in the abundance of the stock. Indeed, ICES biomass indices and stock size indicators are increasing in OPSAR Regions II (Greater North Sea) and III (Celtic Seas). However, due to data uncertainties and incomplete knowledge of gear-specific discard survival rates, ICES has adopted a precautionary approach to landings advice, regulated through a Group-TAC (introduced in 1999 for the North Sea). This advice includes all skate and ray species, except R. undulata and those listed as prohibited. Annual landings for the thornback ray are to be no more than 96 and 2446 tonnes for the west coast of Scotland and North Sea, respectively in 2023 and 2024 (ICES, 2022c,d). R. clavata has been included in the OSPAR List of Threatened/Declining Species and Habitats since 2008 (OSPAR Agreement 2008-6) and has most recently been assessed for The IUCN Red List of Threatened Species as ‘Near Threatened’, though this is currently under review.
3.5.2 Distribution model
Survey data incorporated into the distribution model for thornback ray are shown in Table 12 and Figure 4.
Table 12. Sources of abundance data for thornback ray Raja clavata. Total number of records used in SDM is 53,657.
| Datasource | Years | No. surveys | No. records per survey |
|---|---|---|---|
| ICES DATRAS | 1980-2022 | 206 | 52,305 |
| MD FSS | 2013-2022 | 20 | 627 |
| MD RVR | 1983-1999 | 18 | 259 |
| MD BRUV | 2021-2022 | 2 | 8 |
| MD SCALLOP | 1994-2022 | 60 | 458 |
The binomial part of the GLMM included a linear effect of primary productivity and second order polynomial terms for depth, % gravel and temperature as well as effects of gear type (Figure 28). The probability of presence was maximised at both low and high temperatures (but wide confidence interval for low temperatures), over substrates composed at 15% of gravel and at a depth around 200 m. Presence was higher in areas characterised by low bottom primary productivity. High probabilities of presence were associated with the dredge.
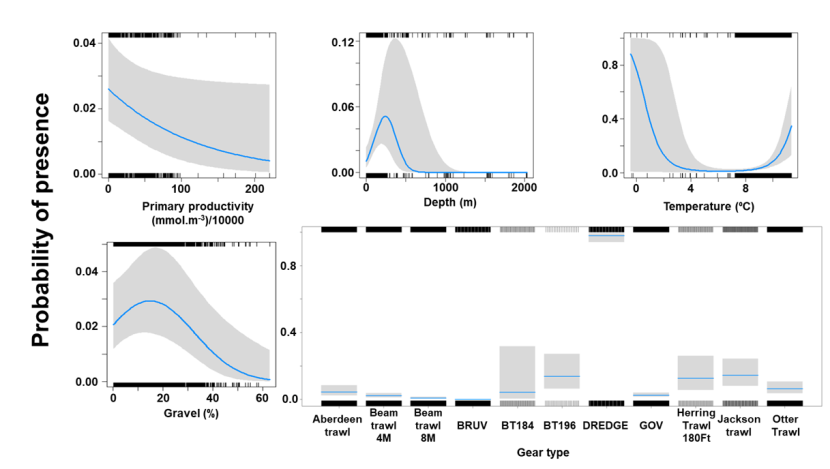
Figure 28. Model plot for the Binomial part of the GLMM for Raja clavata.
Click for a full description
Four individual line charts of the relationship between the probability of thornback ray presence – ranging from 0 to 1 – and environmental variables, including:
- primary productivity on a scale from 0 to 200 mmol per meter cubed - divided by 10000
- depth on a scale from 0 to 2000 m
- temperature on a scale from 0 to 12 degrees Celsius
- percentage gravel on a scale from 0 to 60
A single histogram of the relationship between the probability of thornback ray presence – ranging from 0 to 1 - and eleven gear types, including: Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8 m, BRUV, BT184, BT196, Dredge, GOV, Herring Trawl 180 ft, Jackson Trawl, Otter Trawl.
Refer to Section 3.5.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included second order polynomial terms for % sand, primary productivity and current velocity as well as gear effects. Abundance was maximised at high current velocities, intermediate levels of primary productivity and over substrates composed of sand at 75% (Figure 29). The largest abundances were associated with the otter trawl (Figure 29).
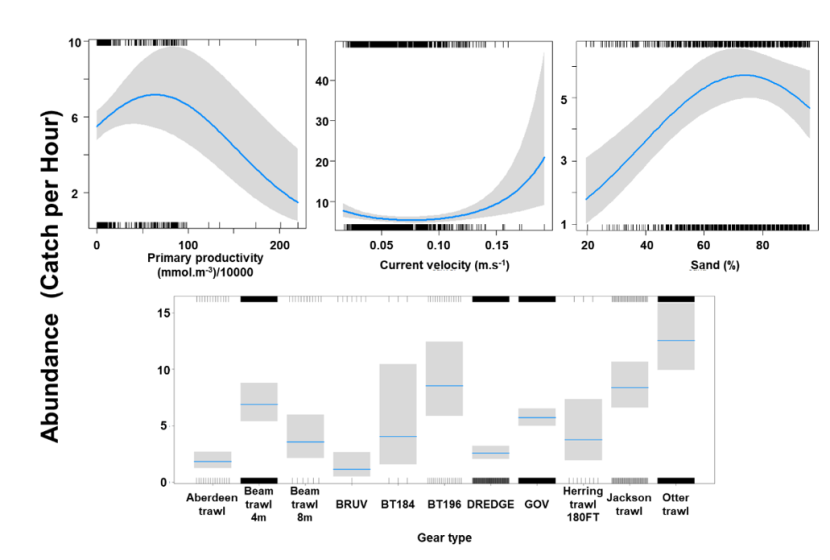
Figure 29. Model plot for the Gamma part of the GLMM for Raja clavata.
Click for a full description
Three individual line charts of the relationship between thornback ray abundance – catch per hour ranging from 0 to 50 – and environmental variables, including
- primary productivity on a scale from 0 to 200 mmol per meter cubed - divided by 10000
- current velocity on a scale from 0 to 0.2 meters per second
- percentage sand on a scale from 20 to 100
A single histogram of the relationship between thornback ray abundance – catch per hour ranging from 0 to 15 - and eleven gear types, including: Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8 m, BRUV, BT184, BT196, Dredge, GOV, Herring Trawl 180ft, Jackson Trawl, Otter Trawl.
Refer to Section 3.5.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, salinity and primary productivity (Table 6). Partial dependence of the model on survey gear type is provided in Annex A5 and shows that the dredge and Jackson trawl are the two most important gear influencing thornback ray abundance.
The predicted distribution of Raja clavata from the ensemble model is presented in Figure 30 and the relative weights given to the models in Table 7. R. clavata is predicted principally on the west coast of Scotland in both inshore and offshore waters, extending to the shelf edge. In particular, R. clavata is predicted as abundant in waters to the west of the outer Hebrides, in the Minch, the inner Sound of Skye, around the Small Isles, the Sound of Jura and the Firth of Lorn. While predicted as present on the east coast of Scotland, the most noticeable areas of high abundance are predicted offshore (Figure 30).
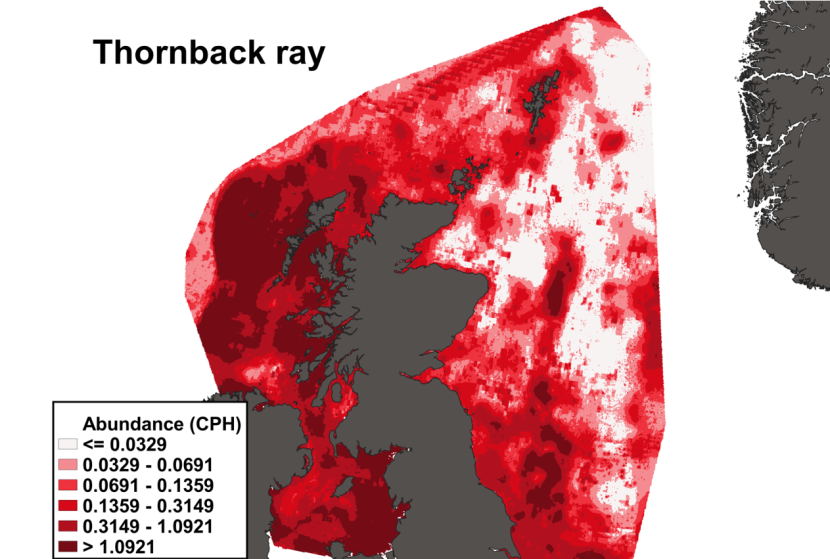
Figure 30. Predicted distribution of Raja clavata from the ensemble SDM.
Click for a full description
A heatmap of predicted thornback ray abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.0329 to greater than 1.0921. Refer to Section 3.5.2 in the main body of text for full description of the distribution.
3.5.3 Predicted bycatch hotspots Nephrops
The estimated overlap between R. clavata predicted distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas in the Minch, around the Small Isles, in the Firth of Lorn and the Sound of Jura show the highest levels of overlap between this fishery and predicted R. clavata distribution (Figure 31).
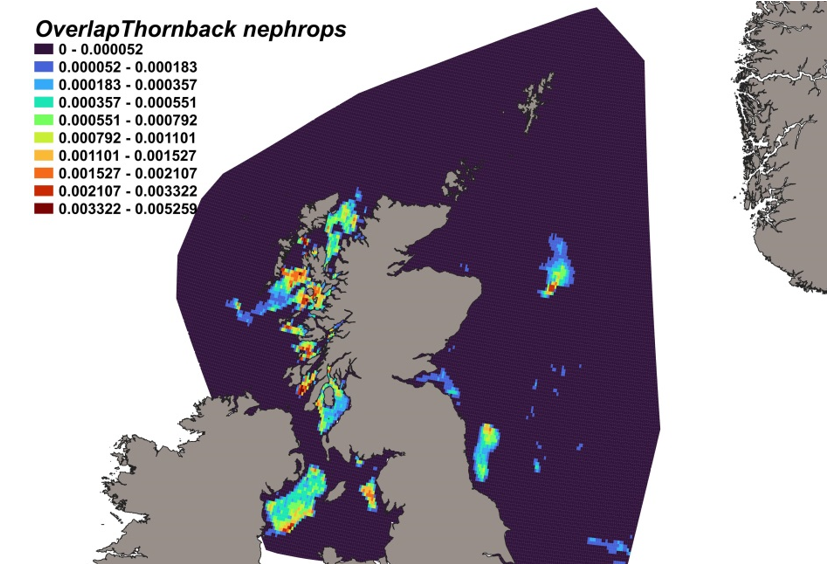
Figure 31. Predicted bycatch hotspots for Raja clavata in the Nephrops fishery.
Click for a full description
Overlap between the distributions of thornback ray and the Nephrops fishery is depicted by a heatmap, with a colour ramp representing the degree of overlap ranging from 0 to 0.005259. Refer to Section 3.5.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.5.4 Predicted bycatch hotspots Scallops
The calculated overlap between predicted R. clavata distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots were situated in the Sound of Jura and the Minch (Figure 32).
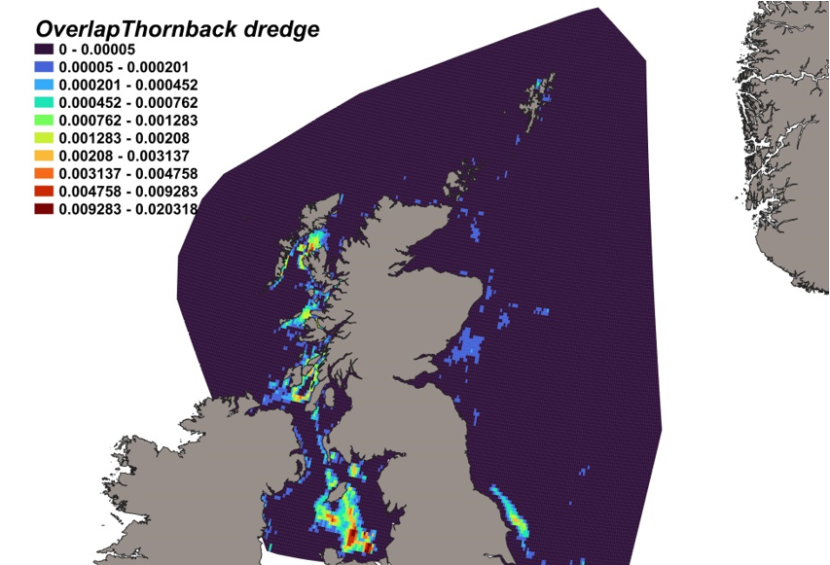
Figure 32. Predicted bycatch hotspots for Raja clavata in the scallop fishery.
Click for a full description
Overlap between the distributions of thornback ray and the scallop fishery is depicted by a heatmap, with a colour ramp representing the degree of overlap ranging from 0 to 0.020318. Refer to Section 3.5.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.6 Spurdog
3.6.1 Species overview
The spurdog, Squalus acanthias, is a globally distributed elasmobranch species found widely throughout British and Irish offshore and inshore habitats, remaining predominately epibenthic at depths < 900 m (Compagno et al., 2005; Dell’Apa et al., 2015). Whilst they tolerate a variety of depths and temperatures, the species shows a preference for waters up to 200 m (Campana et al., 2008) and favour water temperatures in the range of 7 – 15 °C (Compagno, 1984). In general, the species shows a preference for full salinity waters (Burgess, 2002; Stehlik, 2007), but large females are abundant in more brackish, nearshore and more productive (higher concentrations of chl-a) environments, presumably to enhance embryo growth in such warmer waters (Jones and Ugland, 2001), while adult males are more commonly found in deeper, offshore and colder waters with lower chl-a concentrations (Dell’Apa et al., 2016; Shepherd et al., 2002; Methratta and Link, 2007; Sagarese et al., 2014). Adults spend a significant proportion of their time feeding on the bottom, over a variety of substrates, but do so consistently over transitional sand-mud sediments and rocky outcrops (Gottschall et al., 2000; Stehlik, 2007). They have also been found to feed on fish schooling around plankton concentrated by tidal currents (Zamon, 2003). Sprudog is a slow-growing, late-maturing, long-lived species with low reproductive capacity due to its long gestational period (up to 24 months). Based on tagging studies, spurdog appear to be a highly mobile, wide ranging species (Gauld and Macdonald, 1982; Templeman, 1984), with evidence of both migratory (McFarlane and King 2003) and site associated groups (Carlson et al., 2014; Thorburn et al., 2015), possibly due to aggregations differing by size and sex (Shepherd et al., 2002; Dell’Apa et al., 2014). The high dispersal potential of the species is reflected in the lack of natal philopatry and regional genetic differentiation found to date (Hauser et al., 2007), the latter indicating that few barriers to gene flow exist. However, evidence of smaller, genetically distinct groups suggests local spatial management of key habitats may be beneficial to population survival (Thorburn et al., 2018b).
Whilst movements in coastal waters around the UK are poorly understood, the spurdog’s distribution is known to overlap with intensive fishing activities. Spurdog have supported a valuable commercial fishery in the NE Atlantic (mainly on fishing grounds in the North Sea, west of Scotland and Celtic Sea) since the early part of the 20th century and were principally exploited for human consumption and liver oil. The species has been targeted historically by gill-net and long-line fisheries but more recently they have been taken as by-catch in otter trawl and other bottom trawl multi-species fisheries. Landings peaked at over 60,000 tonnes in the early 1960s but have since declined. Discard survival rates are unknown but are likely to be variable. In addition to these fisheries pressure, the species may also be under threat from the consequences of habitat degradation, discarded mobile fishing gears, pollution and coastal development given the reliance on seabed habitats by this species and its prey (ASMFC, 2008; Fordham et al., 2016).
As a consequence of its sensitive life history traits, shoaling behaviour and susceptibility to incidental capture, the spurdogs’ vulnerability to overexploitation by commercial fisheries has been reflected in the steady decline in total biomass and recruitment to the mid-2000s in the North Sea (Greenstreet and Hall, 1996; Heessen and Daan, 1996), with the NE Atlantic biomass in 2010 estimated to be around 20% of historic levels (ICES, 2019). Evidence of such vulnerability and has led to the inclusion of the species to numerous conservation measures since 2000. From 2011, spurdog was a prohibited species for commercial fishing in EU, UK, and Norwegian waters, with the exception of bycatch within some approved avoidance programmes (UK/EU waters) and the landing of dead bycatch (Norway) (ICES, 2019). A recent ICES assessment (ICES, 2023) suggests that the stock has stabilised over the last decade, recovering sufficiently to permit a precautionary re-opening of the fishery. Management measures included a small UK Total Allowable Catch (TAC) (2,781 tonnes in the North Sea and 4,825 tonnes in Western waters for 2023) and the prohibition of landings of mature spurdog (> 100 cm TL), the latter providing protection for the breeding stock (ICES, 2023). However, as the stock remains at a low level compared to historic levels, effective management remains essential to ensure that the fishery expands gradually and that the sustainability of the stock is assured over the long term. Spurdog is currently listed as Vulnerable (criteria A2bd) by the IUCN Red List of Threatened Species due to population decreases (Finucci et al., 2020), is a Priority Species under the UK Post-2010 Biodiversity Framework, has been included on the OSPAR list of Threatened and/or Declining Species and Habitats since 2008, and remains a Priority Marine Feature in Scotland.
3.6.2 Distribution model
Survey data incorporated into the distribution model for spurdog are shown in Table 13 and Figure 4.
Table 13. Sources of abundance data for spurdog Squalus acanthias. Total number of records used in SDM is 109,447.
| Datasource | Years | N. surveys | N. records per survey |
|---|---|---|---|
| ICES DATRAS | 1980-2022 | 179 | 101,994 |
| MS FSS | 2013-2022 | 25 | 2,530 |
| MD RVR | 1980-1999 | 29 | 4,824 |
| MD BRUV | 2021-2022 | 2 | 99 |
The binomial part of the GLMM included a linear effect of salinity, second order polynomial terms for depth and light intensity and effects of gear type (Figure 33). The probability of presence was maximised at a depth of 160 m, at both low and high levels of light intensity and increased with salinity. High probabilities of presence were associated with bottom trawls gear and particularly the BT196, BT184 and Aberdeen trawl.
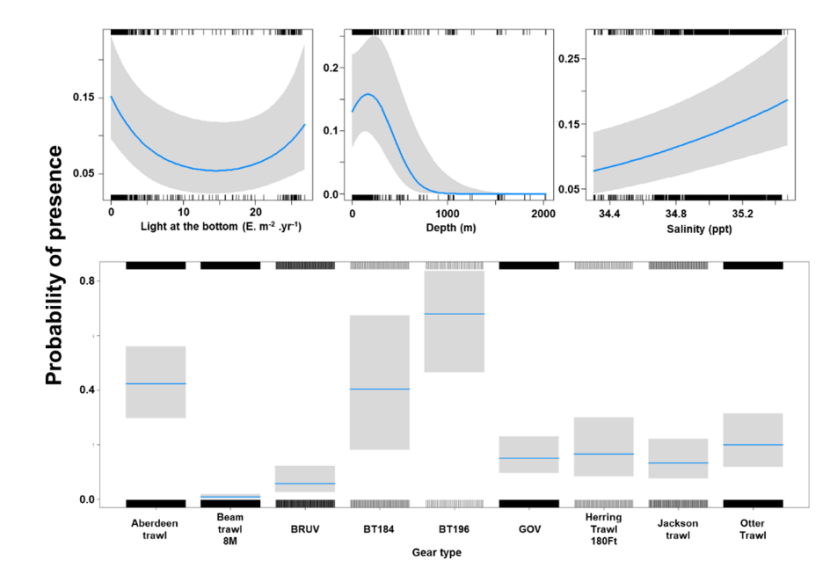
Figure 33. Model plot for the Binomial part of the GLMM for Squalus acanthias.
Click for a full description
Three individual line charts of the relationship between the probability of spurdog presence – ranging from 0 to 0.25 – and environmental variables, including:
- light intensity at the sea bottom on a scale from 0 to just over 25 intensity per meter squared per year
- depth on a scale from 0 to 2000 m
- salinity on a scale from 34.4 to 35.4 ppt
A single histogram depicts the relationship between the probability of spurdog presence – ranging from 0 to 0.8 - and nine gear types, including: Aberdeen Trawl, Beam Trawl 4 m, BRUV, BT184, BT196, GOV, Herring Trawl 180 ft, Jackson Trawl, Otter Trawl.
Refer to Section 3.6.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included second order polynomial terms for the percentage of sand and gravel, linear effects of salinity and temperature and gear effects. Abundance was maximised over sediments characterised by a low sand fraction and proportions of gravel around 20% (Figure 34). Abundance increased with both salinity and temperature and the largest abundances were associated with the otter trawl (Figure 34).
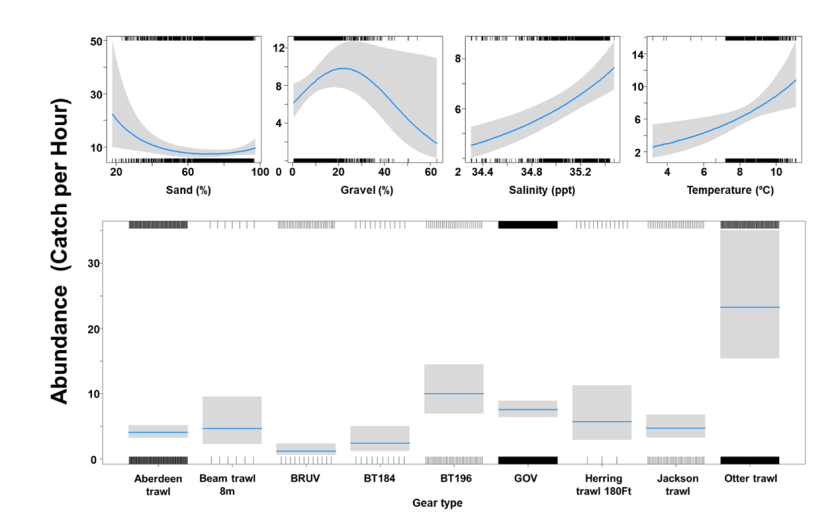
Figure 34. Model plot for the Gamma part of the GLMM for Squalus acanthias.
Click for a full description
Four individual line charts of the relationship between spurdog abundance – catch per hour ranging from 0 to 50 – and environmental variables, including:
- percentage sand on a scale from 20 to 100
- percentage gravel on a scale from 0 to 60
- salinity on a scale from 34.4 to 35.4 ppt
- temperature on a scale from 4 to 12 degrees Celsius
A single histogram of the relationship between spurdog abundance – catch per hour ranging from 0 to 35 - and nine gear types, including: Aberdeen Trawl, Beam Trawl 4 m, BRUV, BT184, BT196, GOV, Herring Trawl 180 ft, Jackson Trawl, Otter Trawl.
Refer to Section 3.6.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, salinity and primary productivity (Table 6). Partial dependence of the model on survey gear type is provided in Annex A6 and shows that BT196 and Aberdeen trawl are the two most important gear influencing spurdog abundance.
The predicted distribution of Squalus acanthias from the ensemble model is presented in Figure 35 and the relative weights given to the models in Table 7. S. acanthias is mostly predicted on the west of Scotland, the Northern Channel and the Irish Sea, at depths in the region of 150-200 m characterised with high salinity. While relatively low abundances are predicted in the Clyde, high abundances are predicted in the Minch, Sea of Hebrides, in and around the LStSJ MPA, North Lewis and to the west of Orkney. While S. acanthias is predicted to be rare in inshore waters on the east coast of Scotland, aggregations are predicted in deeper offshore waters in the North Sea (Figure 3).
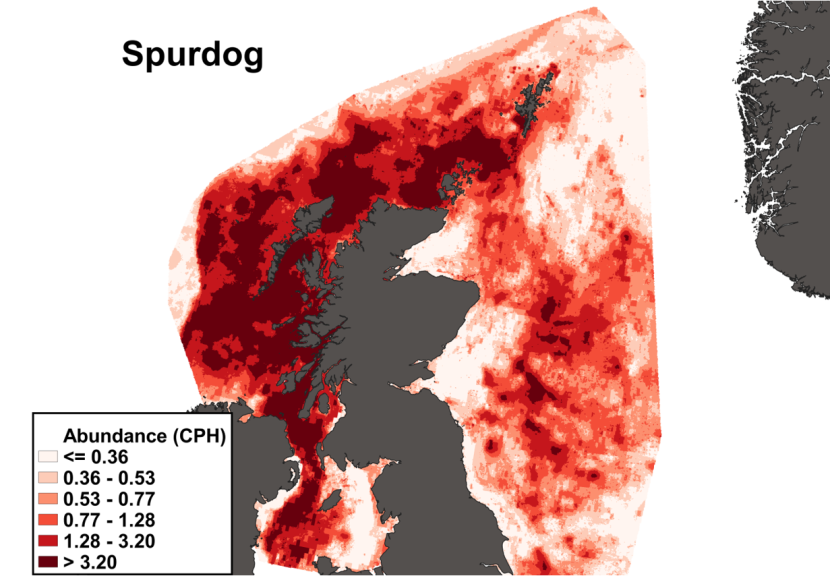
Figure 35. Predicted distribution of Squalus acanthias from the ensemble SDM.
Click for a full description
A heatmap of predicted spurdog abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.36 to greater than 3.2. Refer to Section 3.6.2 in the main body of text for full description of the distribution.
3.6.3 Predicted bycatch hotspots Nephrops
The calculated overlap between S. acanthias distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas to the north-east of Lewis, in the Sea of Hebrides (South of Barra) and in the Sound of Jura show the highest levels of overlap between this fishery and S. acanthias predicted distribution (Figure 36).
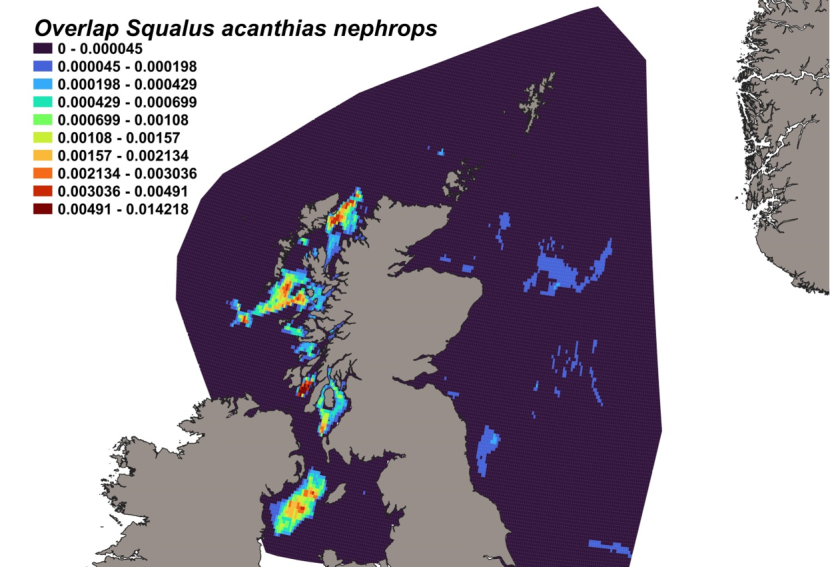
Figure 36. Predicted bycatch hotspots for Squalus acanthias in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of spurdog and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.014218. Refer to Section 3.6.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.6.4 Predicted bycatch hotspots Scallops
The calculated overlap between S. acanthias distribution and the distribution of the scallop fishery revealed a number of potential bycatch hotspots. In particular, areas in the Sound of Jura, south of the tip of the Kintyre peninsula, to the south of Coll and Tiree, to the north west of Skye and to the north of Lewis show the highest levels of overlap between this fishery and S. acanthias predicted distribution (Figure 37).
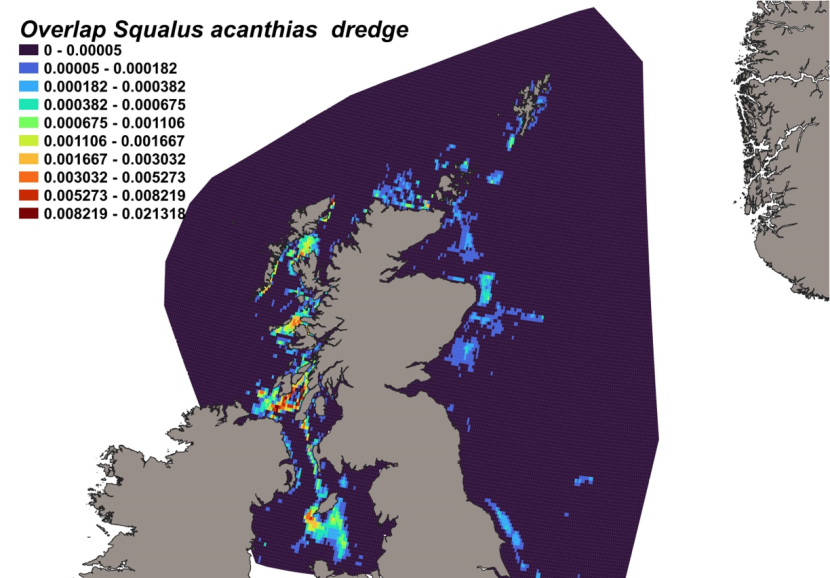
Figure 37. Predicted bycatch hotspots for Squalus acanthias in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of spurdog and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.021318. Refer to Section 3.6.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.7 Blackmouth dogfish
3.7.1 Species overview
The blackmouth dogfish, Galeus melastomus, is a small bottom dwelling squaliform species, distributed widely across the eastern Atlantic Ocean and Mediterranean Sea. It is common throughout a wide bathymetric range, on the outer continental shelf and upper slopes from 55 – 1873 m, but commonly found between 300 – 800 m (D’Iglio et al., 2021). It is generally associated with a wide variety of sediments, from hard substrata to muddy sand sediments, mainly in deep waters with a high seabed gradient (Pennino et al., 2013). Knowledge of the biology of the species is limited, with maturity, age and growth remaining unvalidated. An oviparous species, it spawns all year round and is known to play an important role in the deep-water food web, providing a trophic link between upper and lower levels (D’iglio et al., 2021). Older fish are more commonly found in inshore, warmer waters than their juvenile counterparts. The species is of limited interest to fisheries, caught as by-catch in offshore and deepwater trawls, longlines and traps and largely discarded. Discard mortality is unknown, but likely to be high based on severe injuries obtained when individuals are captured (Serena et al., 2009).
Fisheries independent data indicate increasing biomass in the west of Scotland, southern Celtic Seas and English Channel to about 2012, after which abundance has fluctuated without trend (ICES, 2023e). The lack of reliable catch data means that ICES has not provided catch scenarios for this species. Being one of the most abundant elasmobranchs across its range, there is no evidence of population decline and the species is assessed as ‘Least Concern’ on the IUCN red list. It has protection by the general management arrangements in place since 2010 for deep-water sharks across the North-East Atlantic.
3.7.2 Distribution model
Survey data incorporated into the distribution model for the blackmouth dogfish are shown in Table 14 and Figure 4.
Table 14. Sources of abundance data for blackmouth dogfish Galeus melastomus. Total number of records used in SDM is 45,271.
| Datasource | Years | No. surveys | No. records |
|---|---|---|---|
| ICES DATRAS | 1985-2022 | 123 | 39,880 |
| MS FSS | 2013-2022 | 26 | 4,755 |
| MD RVR | 1983-1999 | 13 | 632 |
| MD BRUV | 2021 | 1 | 4 |
The binomial part of the GLMM included a linear effect of temperature and a second order polynomial term for depth as well as effects of gear type (Figure 38). The probability of presence was maximised at depths between 400-700 m and increased with temperature. High probabilities of presence were associated with the Aberdeen trawl, BT196 and the Jackson trawl.
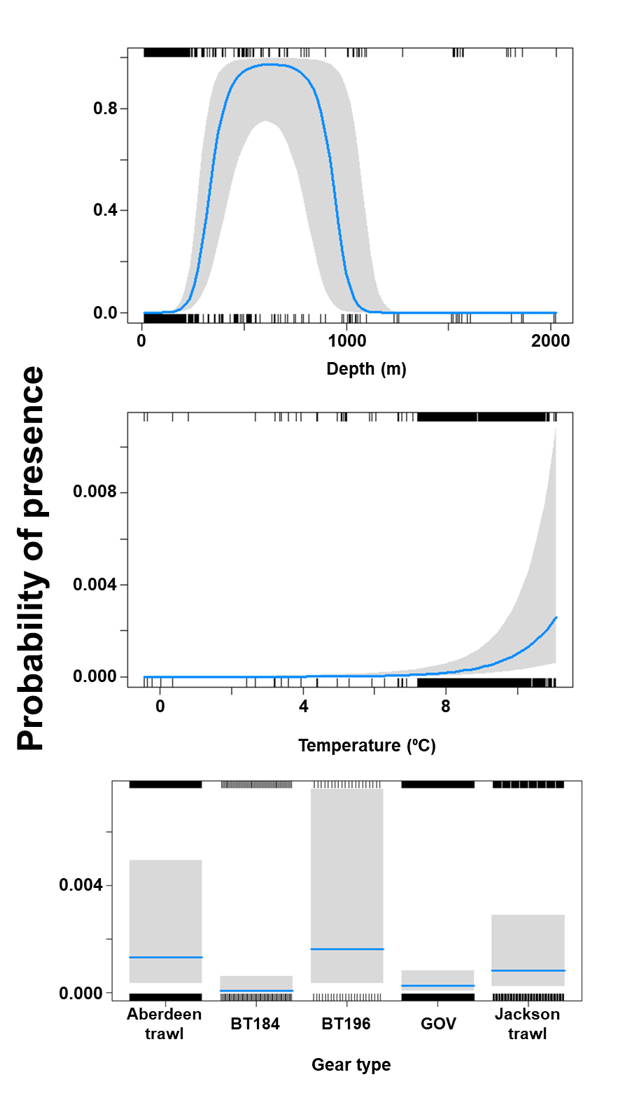
Figure 38. Model plot for the Binomial part of the GLMM for Galeus melastomus.
Click for a full description
Two line charts of the relationship between the probability of blackmouth dogfish presence – ranging from 0 to 1 – and environmental variables, including depth on a scale from 0 to 2000 m and temperature on a scale from 0 to 10 degrees Celsius.
A single histogram depicts the relationship between the probability of blackmouth dogfish presence – ranging from 0 to 0.006 - and five gear types, including Aberdeen Trawl, BT184, BT196, GOV, Jackson Trawl.
Refer to Section 3.7.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included a linear effect of the proportion of gravel and second order polynomial terms for depth and current velocity as well as gear effects. Abundance was maximised over sediments characterised by a low proportion of gravel, a depth around 450 m and both low and high current velocities (Figure 39). However, the confidence interval around the effect of high velocity current is large. The largest abundances were associated with bottom trawls BT184 and BT196 (Figure 39).
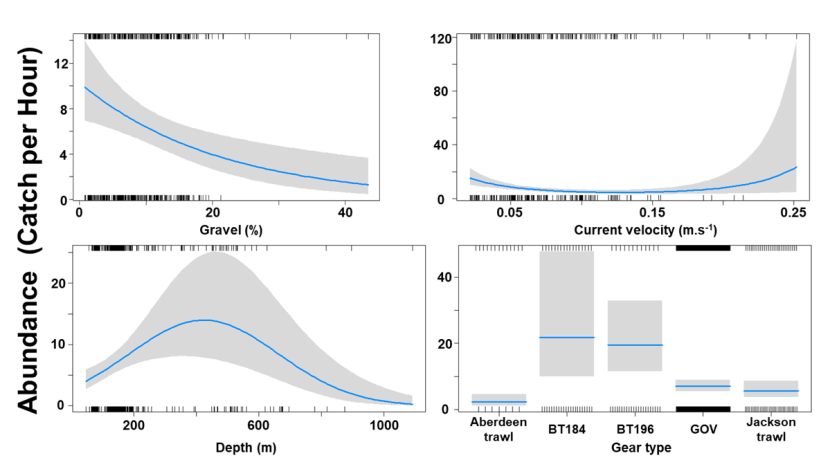
Figure 39. Model plot for the Gamma part of the GLMM for Galeus melastomus.
Click for a full description
Three individual line charts of the relationship between blackmouth dogfish abundance – catch per hour ranging from 0 to 120 – and environmental variables, including:
- percentage gravel on a scale from 0 to 40
- current velocity on a scale from 0 to 0.25 meters per second
- depth on a scale from 0 to 1000 m
A single histogram of the relationship between blackmouth dogfish abundance – catch per hour ranging from 0 to 50 - and five gear types, including Aberdeen Trawl, BT184, BT196, GOV, Jackson Trawl.
Refer to Section 3.7.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, salinity and depth (Table 6). Partial dependence of the model on survey gear type is provided in Annex A7 and shows that BT196 and Jackson trawl are the two most important gear influencing black-mouth dogfish abundance.
The predicted distribution of Galeus melastomus from the ensemble model is presented in Figure 40 and the relative weights given to the models in Table 7. G. melastomus is distributed principally at high depth along the Scottish shelf edge but also show a strong presence in the deep inshore waters of the east coast of Scotland. In particular, the deep trenches found in the Minch, the inner Sound of Skye, around the Small Isles, the Sound of Jura and the Firth of Lorn and in the Northern Channel represent predicted abundance hotspots for this species. G. melastomus is also predicted to be found in the deeper part of the Clyde estuary and in deeper offshore waters on the east coast of Scotland (Figure 40).
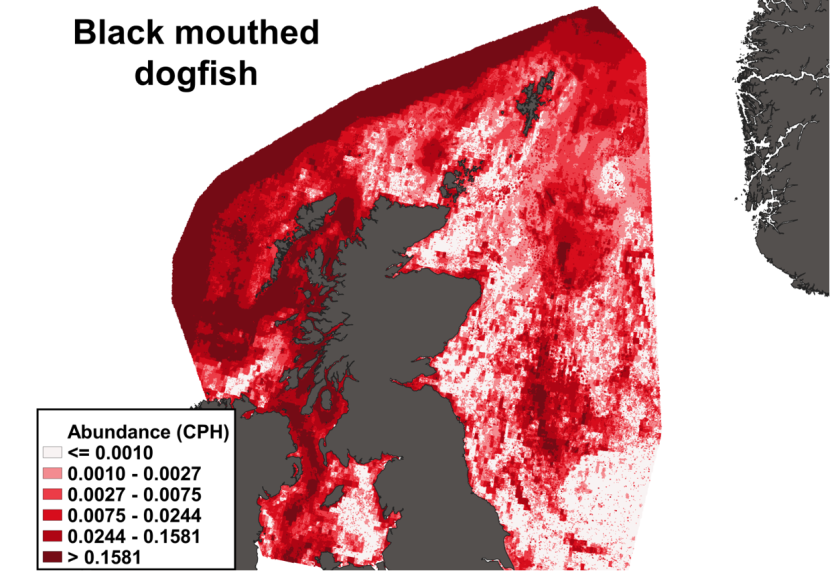
Figure 40. Predicted distribution of Galeus melastomus from the ensemble SDM.
Click for a full description
A heatmap of predicted blackmouth dogfish abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.0010 to greater than 0.1581. Refer to Section 3.7.2 in the main body of text for full description of the distribution.
3.7.3 Predicted bycatch hotspots Nephrops
The estimated overlap between G. melastomus distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas in the Sound of Jura, the sea of Hebrides and in the Clyde show the highest levels of predicted overlap between this fishery and G. melastomus distribution (Figure 41).
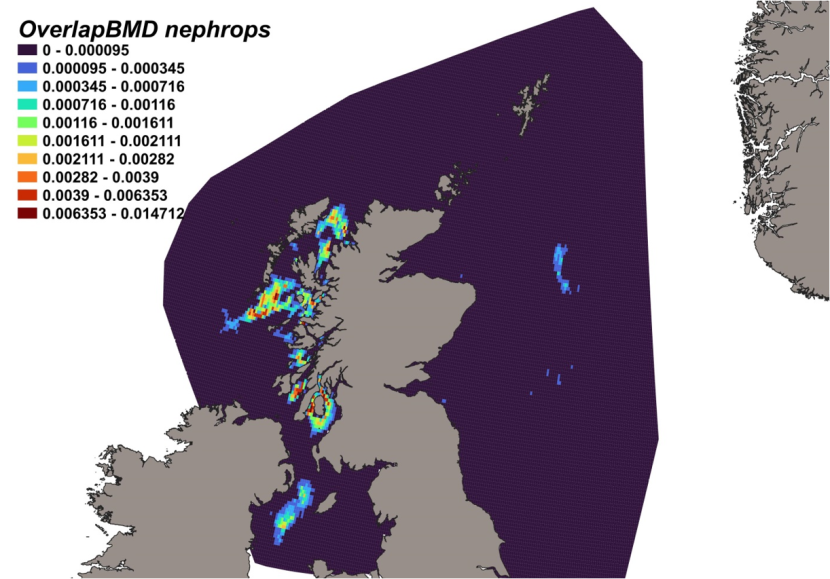
Figure 41. Predicted bycatch hotspots for Galeus melastomus in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of blackmouth dogfish and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.014712. Refer to Section 3.7.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.7.4 Predicted bycatch hotspots Scallops
The calculated overlap between G. melastomus distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots were predicted in the Sound of Mull and the Sound of Jura (Figure 42).
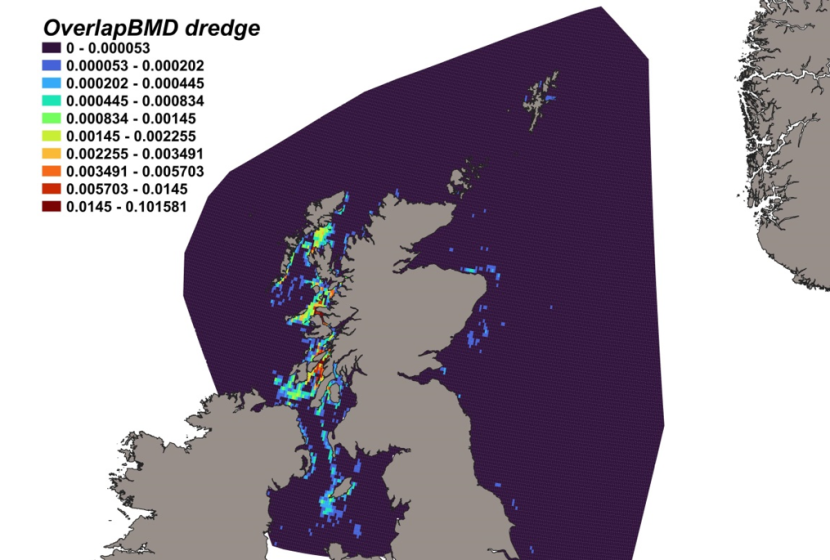
Figure 42. Predicted bycatch hotspots for Galeus melastomus in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of blackmouth dogfish and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.101581. Refer to Section 3.7.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.8 Lesser spotted dogfish
3.8.1 Species overview
The lesser spotted dogfish, Scyliorhinus canicula, is a small, temperate, demersal squaliform species that is considered among the most abundant elasmobranch species in Europe. The species is widely distributed within the North-East Atlantic Ocean and Mediterranean Sea, from the shallow sublittoral to the continental slope at depths mainly around 110 m but up to 800 m (Compagno, 1984; Mytilineou et al., 2005), and is associated with sandy, gravelly or muddy seabeds (Wheeler, 1978; Compagno, 1984). S. canicula is known to aggregate by sex and, less frequently, by size, with adults showing a high degree of site fidelity (Rodríguez-Cabello et al., 2004). Variables such as diel activity, seasonal cycles and bottom sea temperature have been shown to significantly influence spatial behaviour (Papadopoulo et al., 2023). Evidence supports geographical variation in life history across the species range, particularly in reproductive traits such as size and maturity and egg-laying rates. Spawning is continuous throughout the year, with an average gestational period ranging between 5 – 9 months (Ellis and Shackley, 1997). Despite being a non-commercially exploited elasmobranch, S. canicular is caught as incidental by-catch in trawl, gillnet and long line fisheries (Fowler et al., 2005; Rodríguez-Cabello et al., 2004), is targeted by recreational fishers (Baeta et al., 2010), and has a growing importance in crab and whelk fisheries as bait. It appears to have high resilience to commercial fishing due to its high survival rate (e.g. 90 - 100%; Rodríguez-Cabello et al., 2005; Sanchez et al., 2000) after discard. Scientific survey catch data suggest that the population is stable or increasing throughout its range despite some local depletions. ICES recently advised a maximum sustainable yield approach to management, with a maximum landing of 3,984 tonnes for the west of Scotland, Irish Sea and southern Celtic Seas, and 2,680 tonnes for the North Sea, Skagerrak, Kattegat and English Channel in each of the years 2024 and 2025 (ICES, 2023f,g). The species is assessed as ‘Least Concern’ on the IUCN red list.
3.8.2 Distribution model
Survey data incorporated into the distribution model for the lesser spotted dogfish are shown in Table 15 and Figure 4.
Table 15. Sources of abundance data for lesser spotted dogfish Scyliorhinus canicula. Total number of records used in SDM is 592,724.
| Datasource | Years | No. surveys | No. records |
|---|---|---|---|
| ICES DATRAS | 1980-2022 | 195 | 578,347 |
| MS FSS | 2013, 2015, 2017-2022 | 18 | 8,000 |
| MD RVR | 1980-1999 | 23 | 5,988 |
| MD BRUV | 2017-2019, 2021-2022 | 8 | 117 |
| MD SCALLOP | 1994-2022 | 70 | 272 |
The binomial part of the GLMM included linear effects of salinity and bottom temperature, second order polynomial terms for depth and proportion of sand and effects of gear type (Figure 43). The probability of presence was maximised at a depth of 150 m over a substrate composed of 75% sand and increased with salinity and bottom temperature. High probabilities of presence were associated with BT184, dredge and otter trawl.
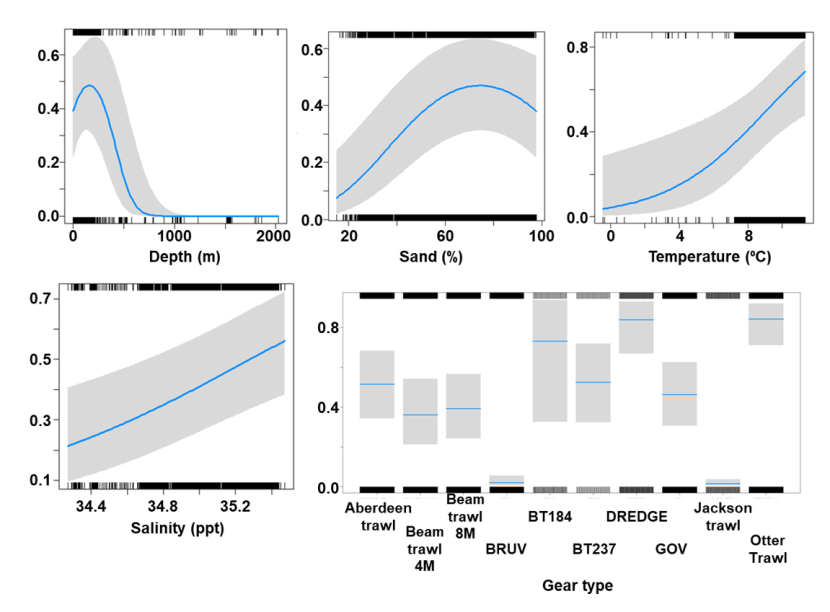
Figure 43. Model plot for the Binomial part of the GLMM for Scyliorhinus canicula.
Click for a full description
Four individual line charts of the relationship between the probability of lesser spotted dogfish presence – ranging from 0 to 0.8 – and environmental variables, including:
- depth on a scale from 0 to 2000 m
- percentage sand on a scale from 20 to 100
- temperature on a scale from 0 to 10 degrees Celsius
- salinity on a scale from 34.4 to 35.4 ppt
A single histogram depicts the relationship between the probability of lesser spotted dogfish presence – ranging from 0 to 0.8 - and ten gear types, including Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8 m, BRUV, BT184, BT237, Dredge, GOV, Jackson Trawl, Otter Trawl.
Refer to Section 3.8.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included second order polynomial terms for percentage of gravel, temperature, primary productivity and temperature and gear effects. Abundance was maximised over sediments characterised proportions of gravel around 25%, a depth around 180 m, high temperatures and low primary productivity (Figure 44). The largest abundances were associated with the otter trawl (Figure 44).

Figure 44. Model plot for the Gamma part of the GLMM for Scyliorhinus canicula.
Click for a full description
Four individual line charts of the relationship between lesser spotted dogfish abundance – catch per hour ranging from 0 to 140 – and environmental variables, including:
- temperature on a scale from 0 to 12 degrees Celsius
- depth on a scale from 0 to 700 m
- percentage gravel on a scale from 0 to 60
- primary productivity on a scale from 0 to 200 mmol per meter cubed - divided by 10000
A single histogram of the relationship between lesser spotted dogfish abundance – catch per hour ranging from 0 to 60 - and ten gear types, including Aberdeen Trawl, Beam Trawl 4 m, Beam Trawl 8 m, BRUV, BT184, BT237, Dredge, GOV, Jackson Trawl, Otter Trawl.
Refer to Section 3.8.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were temperature, salinity and % gravel (Table 6). Partial dependence of the model on survey gear type is provided in Annex A8 and shows that otter trawl and BT237 are the two most important gear influencing lesser-spotted dogfish abundance.
The predicted distribution of Scyliorhinus canicula from the ensemble model is presented in Figure 45 and the relative weights given to the models in Table 7. S. canicula is mostly distributed on the west and north of Scotland, the Northern Channel and the Irish Sea, at depths in the region of 150-200 m. While relatively low abundances are predicted in the Clyde, high abundances are predicted in Sea of Hebrides, the Sound of Jura, North Lewis and along the continental shelf. On the east coast of Scotland the most noticeable aggregation is predicted in the deeper offshore waters (Figure 45).
Figure 45. Predicted distribution of Scyliorhinus canicula from the ensemble SDM.
Click for a full description
A heatmap of predicted lesser spotted dogfish abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.1870 to greater than 24.8668. Refer to Section 3.8.2 in the main body of text for full description of the distribution.
3.8.3 Predicted bycatch hotspots Nephrops
The estimated overlap between S. canicula distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas in the Minch, to the east of Lewis, around the Small Isles, in the Sound of Jura and in the Clyde show the highest levels of predicted overlap between this fishery and S. canicula distribution (Figure 46).
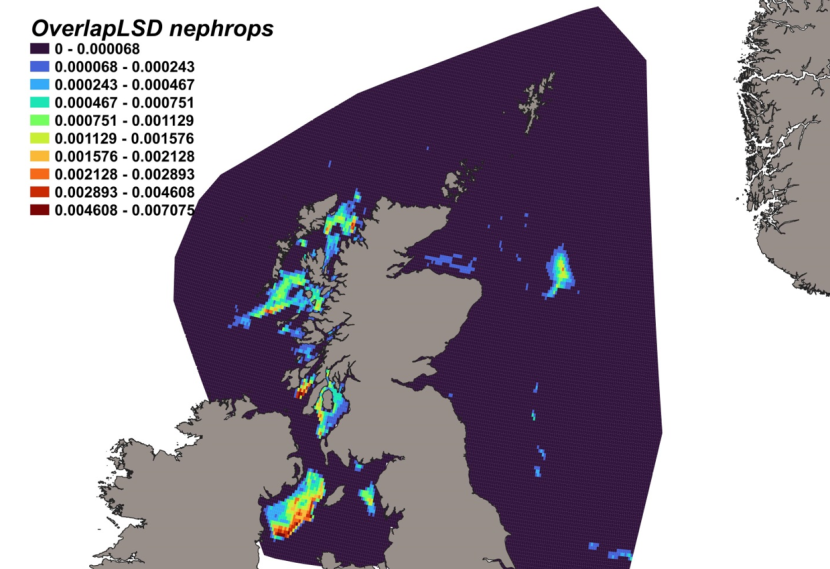
Figure 46. Predicted bycatch hotspots for Scyliorhinus canicula in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of lesser spotted dogfish and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.007075. Refer to Section 3.8.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.8.4 Predicted bycatch hotspots Scallops
The calculated overlap between S. canicula distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots were predicted in the Solway Firth, the North channel and the Sound of Jura (Figure 47).
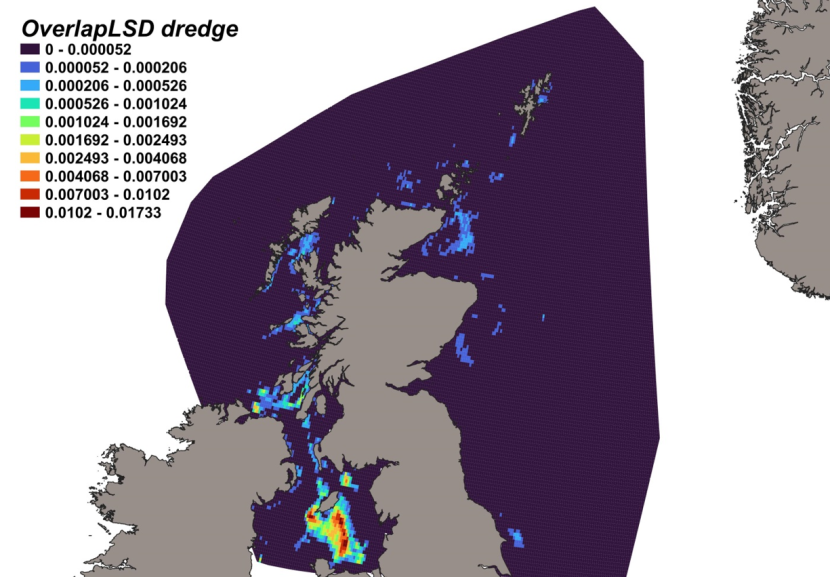
Figure 47. Predicted bycatch hotspots for Scyliorhinus canicula in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of lesser spotted dogfish and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.01733. Refer to Section 3.8.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.9 Starry smooth-hound
3.9.1 Species overview
The starry smooth-hound, Mustelus asterias, is a medium-sized squaliform species that is widely distributed throughout the North-East Atlantic, south-east Atlantic and Mediterranean Seas, and is common within UK waters on sand and shingle substrates in shallower waters up to 200 m (Ebert and Stehmann, 2013; Weigmann, 2016). Evidence from mark-recapture tagging studies indicate that they occur at a maximum depth of 118 m and temperatures between 7 – 22 °C. Proximity to the seabed is consistent throughout the year, with seasonal movements from cooler, deeper waters in the winter to warmer, shallow waters in the summer (Griffiths et al., 2020). The species shows a preference for coarse, mixed sediments which are usually found in inshore coastal areas (Sguotti et al., 2016). They were historically found to the south and west of the British Isles, but their range has expanded to such an extent that they are now considered the most frequently encountered smooth-hound species in the NE Atlantic (Farrell et al., 2009; ICES, 2017).
Knowledge of M. asterias ecology, behaviour and population structuring is limited, but NE Atlantic populations are thought to be slower growing, later maturing, with a slower reproductive rate than their Mediterranean counterparts (Farrell et al., 2010), potentially leaving them more susceptible to overexploitation. Contrary to stocks in the Mediterranean, which are targeted by commercial fisheries for human consumption, stocks in the NE Atlantic are caught as bycatch in demersal trawl, gillnet and longline fisheries and are largely discarded due to their limited market value (ICES, 2012b). Given its preference for shallower waters, M. asterias is also targeted by recreational fisheries. Issues with phenotypic-based identification have prevented the collation of reliable species-specific landings, rather aggregated as ‘Mustelus spp’, but the NE Atlantic stock consists primarily of starry smooth-hound (ICES, 2023g). The resulting uncertainties in stock data estimation means management advice follows a maximum sustainable yield approach, with landings capped at 5,329 tonnes in each of the years 2024 and 2025 (ICES, 2023g). Fishery-independent survey data suggest that the abundance of Mustelus spp. has nominally increased in recent years in the Celtic and Irish Seas, southern North Sea and English Channel (ICES, 2023h). However, as a result of regional declines, particularly the local extinctions within many parts of the Mediterranean Sea, it is estimated that the stock has undergone an overall population reduction of 20-29 % since the early 1980s due to increasing levels of exploitation. Thus, M. asterias has most recently been assessed as ‘Near Threatened’ under criteria A2d (but close to meeting ‘Vulnerable A2d’) for The IUCN Red List of Threatened Species in 2020 (Jabado et al., 2021).
3.9.2 Distribution model
Survey data incorporated into the distribution model for the starry smooth-hound are shown in Table 16 and Figure 4.
Table 16. Sources of abundance data for starry smooth-hound Mustelus asterias. Total number of records used in SDM is 15,021.
| Datasource | Years | No. surveys | No. records |
|---|---|---|---|
| ICES DATRAS | 1989-2022 | 123 | 6,314 |
| MD FSS | 2007-2021 | 29 | 8,707 |
The binomial part of the GLMM included a linear effect of current velocity and second order polynomial terms for depth, proportion of sand and gravel, and salinity as well as effects of gear type (Figure 48). The probability of presence was maximised at a depth around 100 m over a coarse substrate composed of high proportions of sand and gravel, increased with current velocity and in low salinity waters. High probabilities of presence were associated with the use of an otter trawl.
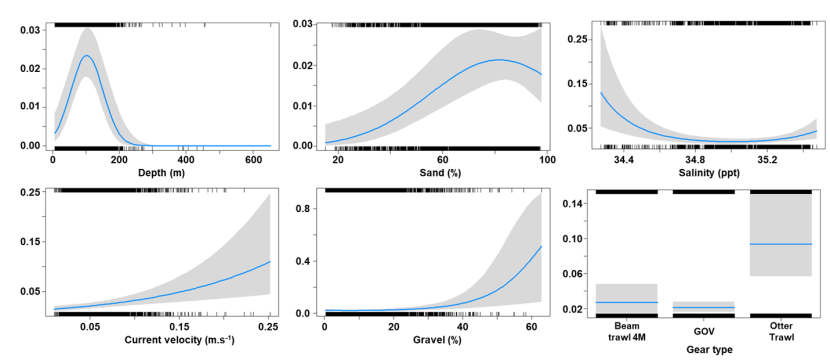
Figure 48. Model plot for the Binomial part of the GLMM for Mustelus asterias.
Click for a full description
Five individual line charts of the relationship between the probability of starry smooth-hound presence – ranging from 0 to 0.8 – and environmental variables, including:
- depth on a scale from 0 to 600 m
- percentage sand on a scale from 20 to 100
- salinity on a scale from 34.4 to 35.4 ppt
- current velocity on a scale from 0 to 0.25 meters per second
- percentage gravel on a scale from 0 to 60
A single histogram depicts the relationship between the probability of starry smooth-hound presence – ranging from 0 to 0.14 - and three gear types including Beam Trawl 4 m, GOV, Otter Trawl.
Refer to Section 3.9.2 in the main body of text for full descriptions of the relationships.
The Gamma (abundance) part of the GLMM model included a linear effect of the proportion of gravel and a second order polynomial term for primary productivity as well as gear effects. Abundance was maximised over sediments characterised by a high proportion of gravel and high primary productivity (Figure 49). The largest abundances were associated with the otter trawl (Figure 49).
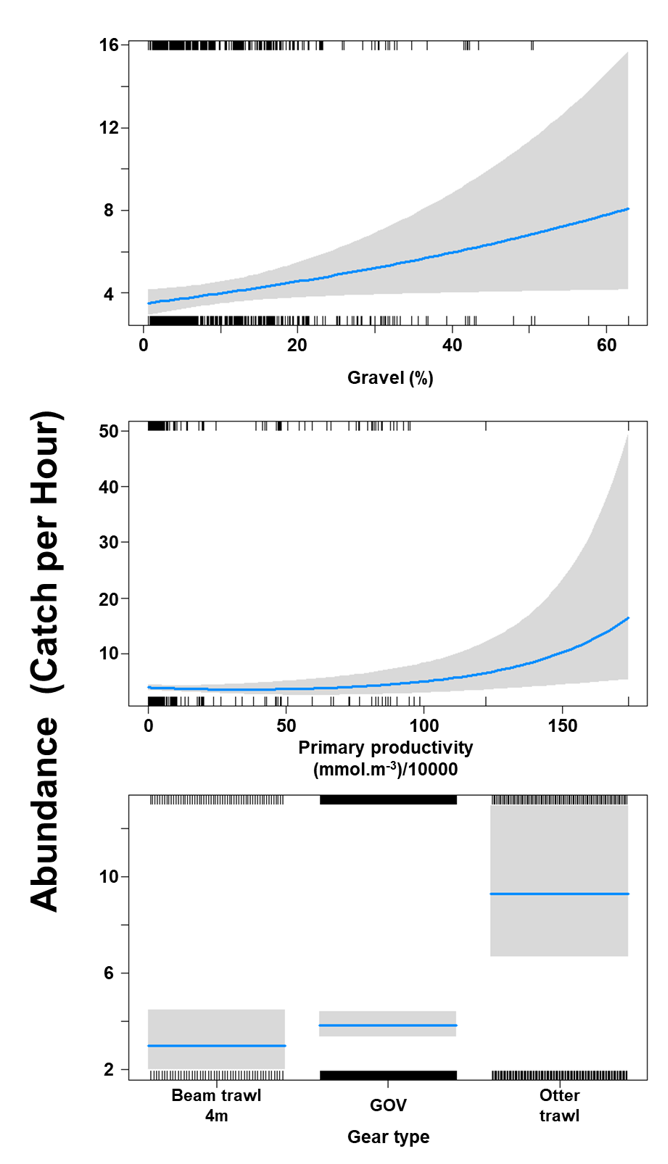
Figure 49. Model plot for the Gamma part of the GLMM for Mustelus asterias.
Click for a full description
Two individual line charts of the relationship between starry smooth-hound abundance – catch per hour ranging from 0 to 50 – and environmental variables, including:
- percentage gravel on a scale from 0 to 60
- primary productivity on a scale from 0 to 175 mmol per meter cubed - divided by 10000
A single histogram of the relationship between starry smooth-hound abundance – catch per hour ranging from 0 to 12 - and three gear types including Beam Trawl 4 m, GOV, Otter Trawl.
Refer to Section 3.9.2 in the main body of text for full descriptions of the relationships.
The three most important variables in the Random Forest model were depth, % gravel and % sand (Table 6). Partial dependence of the model on survey gear type is provided in Annex A9 and shows that otter trawl is the two most important gear influencing starry smooth-hound abundance.
The predicted distribution of Mustelus asterias from the ensemble model is presented in Figure 50 and the relative weights given to the models in Table 7. M. asterias is distributed around Scotland, along the shelf edge and around the Northern Isles with aggregations inshore in the Sound of Jura and the Firth of Lorn on the west of Scotland, in the Northern Channel and the Irish Sea. Mustelus asterias is also predicted in the deeper part of the Clyde estuary and the Solway Firth. On the east coast of Scotland, M. asterias is predicted in the deeper offshore waters (Figure 50).
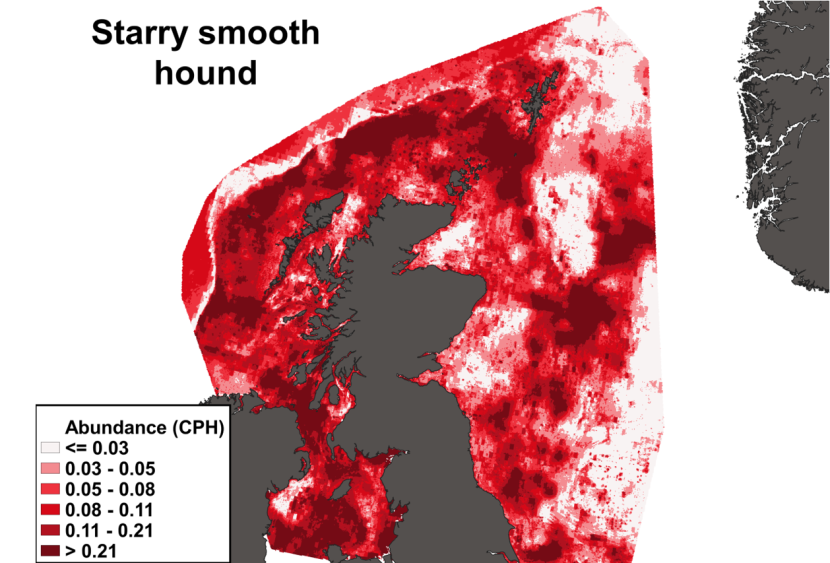
Figure 50. Predicted distribution of Mustelus asterias from the ensemble SDM.
Click for a full description
A heatmap of predicted starry smooth-hound abundance, with a colour ramp representing catch per hour ranging from less than or equal to 0.03 to greater than 0.21. Refer to Section 3.9.2 in the main body of text for full description of the distribution.
3.9.3 Predicted bycatch hotspots Nephrops
The estimated overlap between M. asterias distribution and the distribution of the Nephrops fishery revealed a number of potential bycatch hotspots. Areas in the Sound of Jura, the Firth of Lorn and in the Clyde show the highest levels of predicted overlap between this fishery and M. asterias distribution (Figure 51).
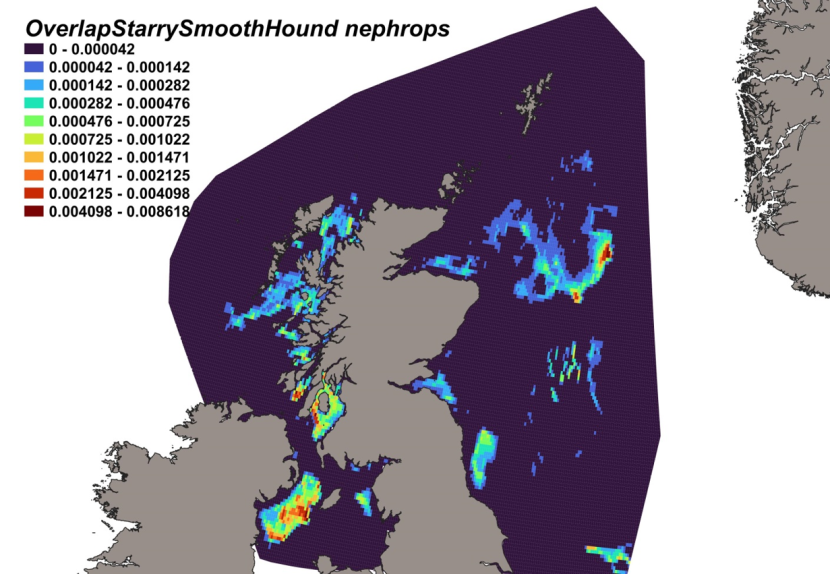
Figure 51. Predicted bycatch hotspots for Mustelus asterias in the Nephrops fishery.
Click for a full description
A heatmap of the overlap between distributions of starry smooth-hound and the Nephrops fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.008618. Refer to Section 3.9.3 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.9.4 Predicted bycatch hotspots Scallops
The overlap between M. asterias predicted distribution and the distribution of the scallop fishery revealed that the most likely bycatch hotspots were predicted in the Solway Firth, the Sound of Jura, to the south of the Mull of Kintyre and to the south of Coll and Tiree (Figure 52).
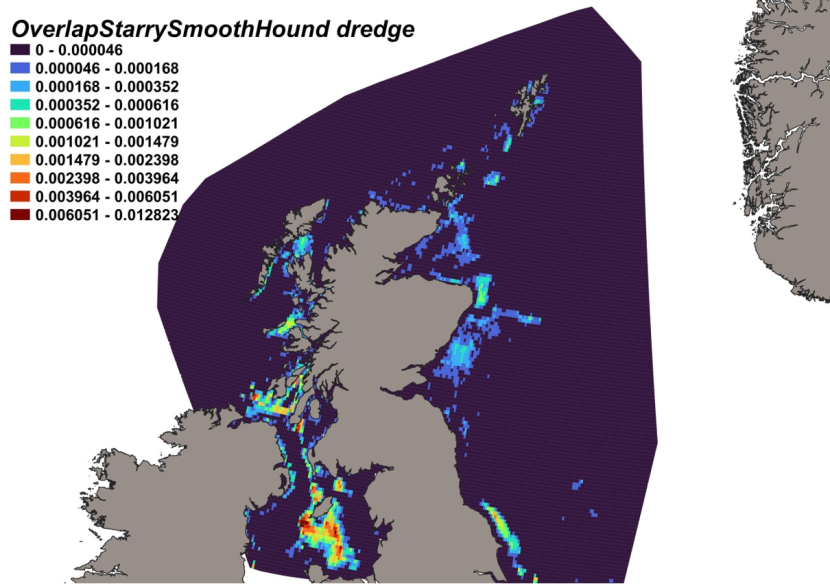
Figure 52. Predicted bycatch hotspots for Mustelus asterias in the scallop fishery.
Click for a full description
A heatmap of the overlap between distributions of starry smooth-hound and the scallop fishery, with a colour ramp representing the degree of overlap ranging from 0 to 0.012823. Refer to Section 3.9.4 in the main body of text for full description of the distribution of potential bycatch hotspots.
3.10 Elasmobranch diversity and species richness
Measures of elasmobranch species richness and diversity (Shannon Diversity Index; H) are shown in Figure 53.
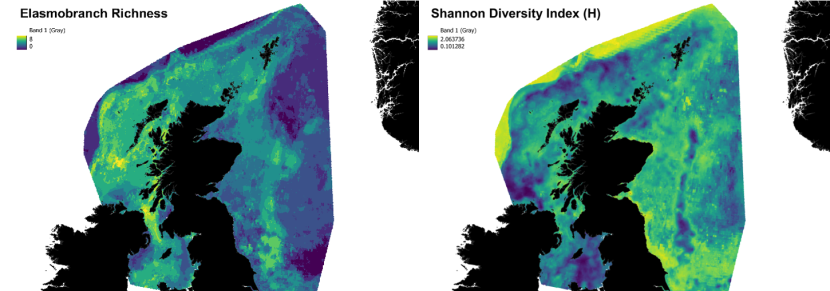
Figure 53. Maps showing measures of elasmobranch species richness (left) and diversity (right) around Scotland. Within the colour ramp, high values of each measure are indicated in yellow, and low values in dark purple.
Click for a full description
Elasmobranch richness is on a scale from 0 to 8 species, and the Shannon Diversity Index is on a scale from 0.1 to 2.06. Refer to Section 3.10 in the main body of text for full description of the variation in richness and diversity.
Elasmobranch richness ranged from 0 to 8 species and diversity from 0.1 to 2.06. A high diversity index (1.75-2.06) would suggest a stable habitat with high species richness and evenness, and a low diversity index (0-1) would indicate habitats with either fewer species or an elasmobranch community dominated by one species.
Patches of high elasmobranch species richness (6-8 species) were predicted off the SW tip of the Kintyre peninsula, within the Sound of Jura, the inner Sound of Skye and within the Hebrides Sea to the south of Barra (Figure 53 left ). Moderate levels of species richness (3-5 species) were widespread across the west coast of Scotland, particularly within the Inner and Outer Hebrides and along the shelf edge. Habitats with low species richness (0-2 species) were predicted off the continental shelf and in patches to the east of Lewis, east of Rum, to the south-east of Eigg and the Firth of Clyde area. Species richness within the North Sea was, overall, much lower, with more discrete patches evident between and around Orkney and the south of Shetland and within deeper waters off the east coast. Relatively low species richness was predicted within the Moray Firth, off the Scottish east coastline and in deeper waters within northern and central areas of the North Sea.
Conversely, high species diversity was more widespread within the North Sea than the west coast, particularly evident at the mouth of the Firth of Forth, and along the SE Scotland coastline (Figure 53 right). Higher diversity values were also apparent in deeper waters off the continental shelf. Within the west coast of Scotland, patches of relatively high elasmobranch diversity were predicted to the south of Arran, off the southern tip of the Kintyre peninsula and around the islands of Colonsay, Mull, Coll and Tiree and into the North Minch. Relatively high diversity was also seen off the Outer Hebrides, particularly to the south of Barra. Habitats with low elasmobranch diversity indices were predicted west and south-east of the Kintyre peninsula, south of the island of Eigg (off Mallaig), and off the north-east tip of Lewis. Lower diversity indices were often the result of dominance by one or a few species in the considered area, while low richness areas could achieve higher diversity indices with most elasmobranch species considered predicted to be present at similarly low abundances (often below the 0.5 fish caught per hour threshold used to determine richness).
Areas of particular interest are characterised by both moderate/high species richness (3-8 species) and high diversity index (Figure 54). Such areas were predicted in the deep trench in the Clyde , in two areas inshore and off the western coast of Islay, in the Sound of Jura, the Firth of Lorn and the Sound of Mull. Similar rich and diverse sites can also be found further north, to the south-east of Barra, near Loch Dunvegan to the west of Skye, in the Sound of Skye around Rona and near Gruinard Bay. To the east of Scotland, similar areas of high richness and diversity were predicted inshore near Rattray head, and in the Forth and Tay region (Figure 54).
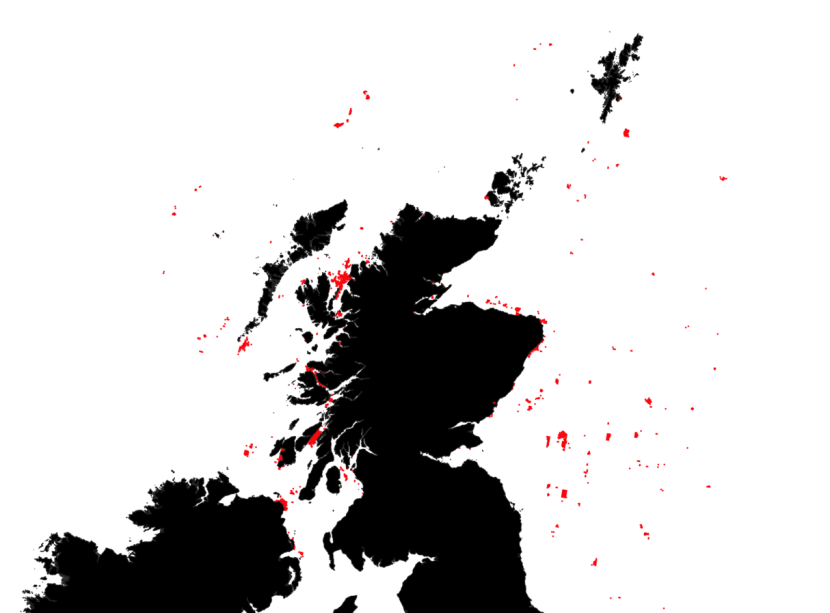
Figure 54. Areas characterised by both a high species richness and diversity are indicated in red.
Click for a full description
Areas characterised by high species richness - between 3 – 8 species - and a high diversity index are shown in patchy locations of the North Sea and west coast of Scotland. Refer to Section 3.10 in the main body of text for full description of the variation in distribution of such areas.
4. Discussion
4.1 Ecological drivers of elasmobranch distributions
Using fishery-independent survey data and ensemble modelling, combining two SDMs, the present study enabled the characterisation of the habitat of nine elasmobranch species present in Scottish waters. While the ecological drivers of the species distributions were often in agreement with previous observations and recorded habitat associations, the approach highlighted the importance of drivers not previously considered and, by accounting for key environmental covariates, enabled the prediction of habitat suitability for the considered species.
Ecological drivers of flapper skate distributions include high gravel substrates, low primary productivity and depths of 200 m. The preference for coarser shell or gravel seabeds has been previously documented (Benjamins et al., 2018), however sediment type was not found to be a key driver of flapper skate distribution in an earlier study (Pinto et al., 2016). The preferred depth predicted by the model conforms with the known high probability of occurrence of the species between 100 – 400 m, only slightly deeper than the core depth ranges of 50-180 m reported by Neat et al. (2015) and Thorburn et al. (2018a) and confirms the outputs of an earlier flapper skate distribution model (Pinto et al., 2016). The areas characterised by the highest primary productivity in the considered region are to the south in the Irish Sea and the North Sea where the flapper skate is rarely observed, which explains the association with low productivity in the models. While the only environmental driver identified in common between the present approach and an earlier model is depth (Pinto et al., 2016), the predicted distribution of flapper skate shows very good agreement between the two studies.
Modelled distributions of the cuckoo ray were driven by high salinity, high temperature, low primary production, shallow waters, 30% gravel and high sand. Preference for such coarse, soft sand and muddy substrates is well supported in the literature (Ebert and Stehmann, 2013; Moriarty, 2021). Temperature, and by close association, salinity, has previously been found to explain habitat preference in this species (Coulon et al., 2024). The preference for combined high salinity and low productivity supports previous observations indicating that the cuckoo ray is distributed along the continental shelf but with low abundances in inshore waters (Ellis et al., 2005a) where salinity is lower and productivity higher.
The distribution models for spotted ray highlight the importance depth, low primary productivity, intermediate salinity, higher temperature and light intensity and substrates with high sand and gravel content. The estimated optimum depth (100 m) is consistent with the known preferred depth range (up to 120 m). The predicted seabed preference (coarse sand) is also consistent with earlier findings (sandy and sandy-gravels; Serena, 2005). Given the importance of light intensity and primary productivity to the modelled species distributions, these variables should be considered in future studies. Bed sheer stress has been evidenced as a significant predictor of spotted ray however, bottom current velocity, a variable related to bed sheer stress, was not found to be a significant driver of spotted ray distribution in this study.
Existing knowledge of starry ray habitat preferences show that the species is associated with cooler temperatures and oceanic levels of salinity (Hogan et al., 2013; COSEWIC, 2012). This is partly confirmed by the SDMs within this study, which found that probability of presence was highest at low temperatures and low salinity levels. Therefore, a high probability of occurrence is predicted in the colder North Sea. While a wide sediment preference for this species has been observed, with complex, coarse substrates identified in some studies (Scott, 1982; Sosebee et al., 2016) and sand, gravel, broken shell, pebbles, to soft mud identified in others (reviewed in Packer et al., 2003), the present SDMs identify a preference for substrates with high sand and low gravel contents. High primary productivity was an important driver of starry ray distribution, which may be associated with the species’ known preference for more inshore water depths up to 100 m.
The thornback ray is known to be associated with mud, sand and gravelly substrates throughout its range (Ebert and Stehmann, 2013). This is in part, confirmed by the modelled habitat drivers within this study, where sand and coarse sand (70 % sand and 15 % gravel) were identified as important. The estimated preferred depth of 250 m was deeper than the published preferred water depth range, from 10-60 m (Ebert and Stehmann, 2013; Ellis, 2016, McAllister et al., 2023), but the species is known to inhabit a wide bathymetric range, and depth occupancy can vary by life stage (juveniles use shallower waters; Ebert and Stehmann, 2013; McAllister et al., 2023), which was not accounted for within this study. High current velocity and low/intermediate levels of primary productivity were shown to be additional important drivers of modelled distributions. Whilst high and low salinity levels are tolerated by this species, it is not thought to be a significant driver of presence, which is mirrored by the modelling results of this study.
Estimated drivers of spurdog distributions included high temperature, high salinity, a preferred depth of 160 m and muddy rather than sandy substrates. These parameters fit well with existing knowledge of the species habitat preferences (McMillan and Morse, 1999; Burgess, 2002; Stehlik, 2007), where occurrence coincided with highly saline, warm (up to 15 °C) waters at depths up to 200m (Campana et al., 2008). The species preference for soft substrates is also documented in the literature (McMillan and Morse, 1999).
The blackmouth dogfish was predicted to occur at depths between 400 – 800 m, which is consistent with existing evidence of a depth preference between 300 – 800 m (D’Iglio et al., 2021). The species is known to prefer soft substrates, which is supported by the importance of low percentage gravel habitats from the modelled outputs, but also shows an association during spawning times with Lophelia reefs (which require hard substrates for attachment) which can provide ideal egg-laying habitat (Navas et al., 2014). The blackmouth dogfish was found to prefer the higher end of the temperature range observed in the study. This temperature effect is likely associated with this species being mainly found to the west of Scotland, where the warmer North Atlantic current predominates, as opposed to the colder waters dominating the part of the North Sea considered in this study. However, such results must be interpreted with caution, as older fish are known to prefer warmer waters, and species-age-structures were not considered in the distribution models.
The modelled predicted depth of the lesser spotted dogfish (150 m) is consistent with the depth at which the species is commonly associated with (110 m, but up to 800 m; Compagno, 1984; Mytilineou et al., 2005). Sand, gravel and muddy substrates are known to be suitable habitat for the species (Wheeler, 1978; Compagno, 1984). The present study narrows down the relative importance of habitat types and indicates a preference for coarser sand (75 % sand, 20 % gravel). While bottom sea temperature has been shown to significantly influence spatial behaviour, this study also shows that high temperature and high salinity are drivers of lesser spotted dogfish distributions, evidenced by predicted high abundances in offshore, warmer waters off the west coast of Scotland.
The preference for depth around 100 m and low salinities identified in the present study for the starry smooth-hound are consistent with the species’ known preference for shallower, inshore waters up to 200 m (Ebert and Stehmann, 2013; Weigmann, 2016). High primary productivity and high current velocity were also significantly important variables in determining occurrence, which is likely related to the species presence along the northern shelf edge and between the Northern Isles. Known habitat preference for coarse, mixed sediments (Sguotti et al., 2016) were also captured well by the models, where high sand and high gravel substrates were found to be important to the species.
In summary, of the five modelled skate and ray species distributions studied, four were driven by the presence of high sand substrates, and areas of low productivity. All four of the squaliform species distributions were driven by depth, and, in three of the four species, by high water temperature and substrates containing lower percentages of gravel. However, there are a number of caveats that should be considered prior to interpretation of the modelling results. Survey field sampling is a biased snapshot of species presence/absence and abundance and has inherent restrictions caused by sampling design, operational reach and temporal (both year and season) coverage. Model limitations also extend to the biological realism of data inputs, as data on the age, maturity, size structure and behaviour of each species were not accounted for in this study. Moreover, elasmobranchs associate with different habitats during certain life-stages and with activity (e.g. feeding, spawning, seasonal movements), and this was not captured by the model. Non-inclusion of such temporal, behavioural and biological factors may confound model outcomes, and limit their application to species-specific spatial management measures.
4.2 Survey gear efficiency
The vulnerability of a species to certain fishing gears can be affected by many factors, including the efficiency of the gear type, seabed type, species size and body shape, species behaviour, fisher experience and effort, environmental conditions and the proximity of the gear to the seabed. Given these variables, estimating the degree of species vulnerability to certain gears is difficult to predict, however, results from the two modelling approaches applied to survey data, may allow us to infer which gears have more impact on the elasmobranchs under study.
Predicted presence of the flapper skate, derived from the GLMM model, was highest with the dredge, but highest abundances were associated with bottom trawls like the BT184 and Jackson Trawls along the shelf edge and in deeper troughs of the North Sea. For the flapper skate and all the ray species (thornback, cuckoo, starry and spotted rays), the dredge had the largest effect on the probability of occurrence. Similarly, in the Random Forest model, without formally separating occurrence and abundance, the dredge consistently appeared in the two most significant gear affecting skate and ray distributions while it only had a marginal effect in one of the squaliform elasmobranch (i.e. lesser-spotted dogfish). In the GLMM models, bottom trawls were found to be the most effective type of gear at detecting variations in skate and ray abundances and in both models were identified as the most significant type of gear for squaliform elasmobranch. Substrate-dwelling species which have depressed body morphologies associated with limited manoeuvrability (Parson et al., 2011) therefore appear particularly vulnerable to the dredge as compared to bentho-pelagic squaliforms. Bottom trawls were more quantitative and reflected variations in species abundances. The estimated effects of particular bottom trawl types were related to the overlap between species distributions and survey deployment areas. For example, otter trawls and 4 m beam trawls were only deployed within the Irish Sea, and sensitivity to these gears was only seen in the species which had a high presence in that area (thornback and spotted rays, spurdog, starry smooth-hound and lesser spotted dogfish). The starry ray (mostly distributed within the North Sea) was sensitive to the Aberdeen, 8 m beam and herring trawls which were primarily deployed only in the North Sea, but these gears have little impact on most of the west coast species due to the limited overlap between their distributions and the gear deployment in the surveys.
Results from both modelling approaches suggest that spurdog is often captured in bottom trawls, particularly by the BT196. Catchability was highest in the otter trawl in the Irish Sea, where there was high overlap between high species abundances and gear deployment. Other squaliform species, including the lesser spotted dogfish and starry smooth-hound were similarly affected by bottom gears, with both modelling approaches indicating high catchability with the otter trawl in both species. While the dredge detected lesser spotted dogfish presence in the GLMM model, it had a negligible effect on species abundance and only a marginal effect was quantified in the Random Forest model. Similar squaliform species are known to be highly manoeuvrable with maximum escape performance (Domenici et al., 2004; Seamone et al., 2014), which may enable gear avoidance, particularly for small ground gears like the dredge and beam trawls. Both high probability of occurrence and abundances of blackmouth dogfish were associated with the BT196 trawl gear, and catchability in the Jackson trawl was high in areas where deployment of the gear was intensive (deep regions off the west coast of Scotland and North Sea). Sensitivity to the Aberdeen trawl was high due to the overlap between the species distribution and survey distributions, with presence high along survey paths off the shelf edge and deeper areas of the North Sea. High survey intensity in inshore waters off the east coast of Scotland was not reflected in species abundances, fitting with the species preference for deeper waters.
In fishery-independent surveys, all skate and ray species were often captured in the dredge but variations in their abundances were better captured by bottom trawls as a result of larger gear dimensions. The squaliform species were vulnerable to a wide range of bottom trawls, being mostly present in the BT196, Aberdeen, BT184 and otter trawls, but showed a greater capacity at avoiding the dredge compared to skates and rays. Based on these results and where their modelled distributions overlap with the respective fisheries, skate and rays may be more vulnerable to scallop dredging while all elasmobranch are likely to be caught in large numbers in the Nephrops fishery.
4.3 Species diversity and richness patches
Patches characterised by both a high elasmobranch species richness and diversity are distributed throughout the region, occurring both to the west of Scotland and within the North Sea. The locations of patches are often characterised by depth > 50 m or in close proximity of deeper areas. Patches identified within the North-East Atlantic are primarily located inshore, within the Sound of Raasay/Inner Sound (east of Skye), tip of Barra, the Sound of Mull, Sound of Jura, Loch Indaal/Laggan Bay (Isle of Islay) and off Fair Head to the north-east of Northern Ireland. More offshore patches include those to the far east of Islay and patches along the continental shelf. These patches to the west of Scotland are often characterised by substrates with low sand content while areas in the North Sea have high sand contents. Patches within the North Sea are mainly offshore in the deeper troughs to the east of Scotland and south-east of Shetland but can be found inshore along the NE Buchan and Moray coasts.
These predicted patches of high elasmobranch diversity and species richness are subjected to variable levels of fishing pressure, with the patches associated with the deep trench in the Clyde and around Rona exposed to moderate to high level of Nephrops fishing effort. The patches identified in the Sound of Mull and in and around the Loch Sunart to the Sound of Jura MPA can be subjected to intermediate to high levels of scallop dredging. Similarly, some of the patches off the east coast of Scotland are subject to scallop dredging.
4.4 Predicted bycatch hotspots
The measure of the overlap between the modelled distribution of elasmobranch species and Nephrops and scallop fisheries have highlighted a number of potential bycatch “hotspots” around the Scottish coast. The Sound of Jura was shown to be the most important area of overlap across all species and both the Nephrops and the scallop fisheries. Important bycatch hotspots for the scallop fishery were also predicted in the Minch, between Mull and Coll and south of the Kintyre peninsula on the west of Scotland and around Rattray head and offshore in the Forth and Tay region to the east of Scotland. Likely bycatch hotspots in the Nephrops fishery were predicted around the Small Isles and Inner Hebrides to the west, and in the Firth of Forth to the east of Scotland. In particular, bycatch of squaliform species appeared most likely within the Sound of Jura, Small Isles and off Lewis, with skate and ray predicted bycatch hotspots within the Sound of Jura, Minch, Firth of Forth and Small Isles.
4.5 Future management
Twenty one elasmobranch species (eight of which are PMFs) in Scottish waters are currently offered protection under Scottish Government legislation introduced in 2012. Four MPAs have been designated with elasmobranch as a listed feature, Loch Sunart to the Sound of Jura and Red Rocks and Longay for flapper skate, Sea of Hebrides for basking shark and West of Scotland for the gulper shark and Portuguese dogfish. However, the knowledge gaps regarding elasmobranch distributions and vulnerabilities to fishing gear and fisheries have hindered the consideration of such spatial management measures for other elasmobranch species in Scottish waters. The present study provides data that helps to reduce these evidence gaps.
Many of the areas of high elasmobranch diversity/richness predicted in this study fall within the existing MPA network (e.g. areas near the Sound of Jura (Loch Sunart to the Sound of Jura MPA), SE Barra (Sea of the Hebrides MPA), Gruinard Bay (Wester Ross MPA), Rattray Head (Southern Trench MPA), and therefore benefit from fishery management measures, where relevant measures are in place, or may benefit from the future implementations of such measures. Others are situated outside the current MPA network (deep trenches within the Clyde, inshore and off the western coast of the Isle of Islay, near Loch Dunvegan to the west of Skye, in the Sound of Skye around Rona and the Forth and Tay region). By considering both patches of predicted high elasmobranch richness and diversity and potential bycatch hotspots, areas of conservation importance for elasmobranch have been identified (Figure 55) as an important first step in the building of the evidence base necessary for the future management of marine resources and the development of an elasmobranch action plan for Scottish waters.
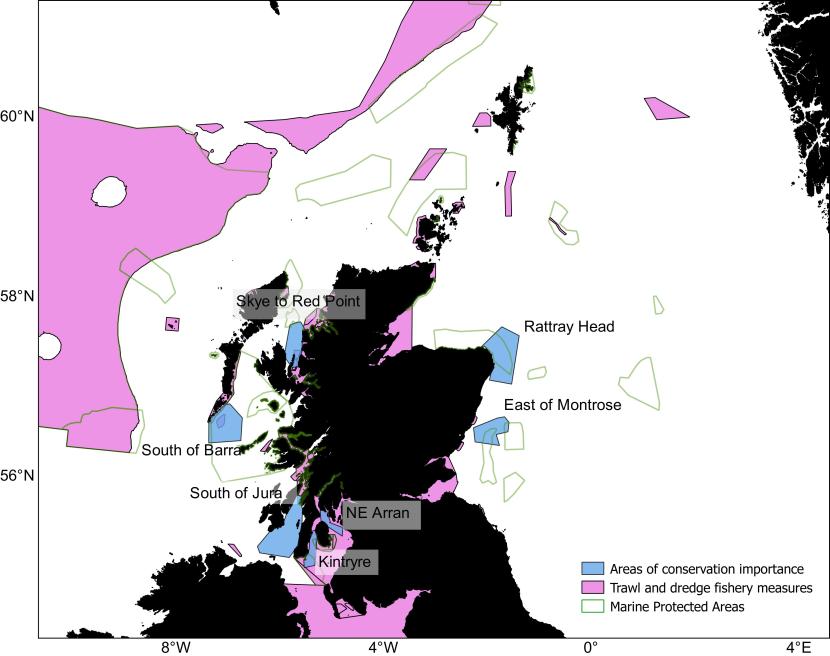
Figure 55. Identified areas of conservation importance overlaid on areas with existing mobile gear restrictions and/or prohibitions including areas with specific protection for scallop and Nephrops species and Marine Protected Areas.
Click for a full description
Polygons representing areas of conservation importance are shaded and labelled. West coast areas are labelled Skye to Red Point, South of Barra, South of Jura, Kintyre, NE Arran. North Sea areas are labelled East of Montrose and Rattray Head.
The map also includes the polygon outline of existing Marine Protected Areas and shaded polygons representing areas with fishing restrictions. Refer to Section 4.5 in the main body of text for fuller description of the overlap between these areas and the areas of conservation importance.
One area of conservation importance identified through this process is Skye to Red Point. This area is particularly important to flapper skate, thornback ray, blackmouth dogfish and, to a lesser extent, to spurdog and lesser-spotted dogfish. In particular, for flapper skate (a PMF), this area is an important bycatch hotspot for the Nephrops fishery. While this area is situated primarily outside of the existing MPA network, it encompasses a deep trench, likely used by flapper skate to reach the Red Rocks and Longay MPA (located slightly to the south), which was designated in 2021 to specifically protect a flapper skate egg nursery (Dodd et al., 2022). While a very small portion of this important area has existing fishing prohibitions in place (Loch Torridon and Northern Inner Sound; Figure 55) it would constitute a candidate for future considerations regarding the expansion of fisheries management measures – more specifically those which minimise the destructive effects of bottom trawl fishing to flapper skate as well as other elasmobranch bycatch (e.g. gear restrictions or modifications such as the removal of the tickler chains from bottom gears).
A second area of conservation importance was identified within the Sound of Jura (i.e. South of Jura, Figure 55). This area is important to the two of the PMFs considered (flapper skate and spurdog) as well as rays (thornback ray, spotted ray, cuckoo ray) and squaliform species (stary smooth-hound, lesser-spotted dogfish and blackmouth dogfish). Both the scallop and Nephrops fisheries are active in the area and, therefore, potential bycatch hotspots in these fisheries have been predicted for all the above-mentioned species in this area. This area of conservation importance has protection already in place in the form of the LStSJ MPA to the north, within which fishing is prohibited, however, the south of the sound of Jura could benefit from future measures offering protection to these elasmobranchs from dredging and bottom trawls.
An area to the south of Barra was also identified as important for elasmobranch species diversity and richness and was assessed as a bycatch hotspot for a variety of elasmobranch in the Nephrops fishery. This area is particularly important for flapper skate, spotted ray, cuckoo ray, spurdog, lesser-spotted dogfish and blackmouth dogfish and of marginal importance to thornback ray and starry smooth-hound. This area is within the Sea of Hebrides MPA and is further protected by fishing restrictions within the East Mingulay SAC (Figure 55).
An area in the Kilbrannan Sound, to the west of Arran in the Clyde, was identified as an area of conservation importance (i.e. Kintyre, Figure 55). This area is important to flapper skate, starry smooth-hound and blackmouth dogfish and, to a lesser extent, to thornback ray, spotted ray, spurdog and lesser-spotted dogfish. This area was identified as a bycatch hotspot in the Nephrops fishery and the southern end of the Kintyre peninsula was identified as a bycatch hotspot for the scallop fishery for many of these species. This area is partly covered by the South Arran MPA to the north-east and overlaps with the Clyde Sea Sill MPA to the south, which also has fisheries management measures in place for cod such as seasonal closures and vessel size restrictions (Figure 55). Still within the Clyde estuary, an area of conservation importance for flapper skate, spurdog and blackmouth dogfish was identified in the deep trench of the Sound of Bute (North East Arran, Figure 55). This area was identified as a bycatch hotspot for these species in the Nephrops fishery.
While most species considered show both higher levels of presence and abundance on the west of Scotland, a few areas on the east coast of Scotland were identified as being important. An area around Rattray Head (Figure 55) is of particular importance to cuckoo ray and on the margin of the distribution of other elasmobranchs, including flapper skate, starry ray, spotted ray, spurdog and starry smooth-hound. The area is marked by scallop dredging activity and is therefore identified as a bycatch hotspot for these species. This area overlaps with the Southern Trench MPA, hence the future implementation of management measures may provide protection to elasmobranchs.
Similarly, another area predicted in the Forth and Tay region (East of Montrose, Figure 55) was identified as particularly important to cuckoo ray and on the margin of the distribution of other elasmobranchs. This area was predicted as a bycatch hotspot in the scallop fishery and overlaps with the Firth of Forth Banks complex MPA. The implementation of dedicated fishery measures in these MPA would therefore likely benefit species such as the starry ray, cuckoo ray and starry smooth-hound.
5. Conclusions
Using elasmobranch survey data compiled from multiple sources as well as fisheries distribution data, this study provides an important contribution to the evidence necessary for the future development of an action plan for the conservation management of elasmobranchs in Scottish waters. Ensemble Species Distribution Models were developed for nine elasmobranch species (including two classified as Priority Marine Features), enabling the estimation of (1) their respective distributions in Scottish waters, (2) a characterisation of their habitat requirements through the identification of key environmental drivers of these distributions and (3) the importance of the various fishing gear, deployed in the surveys, for the detection of species occurrence and variation in their respective abundances. Predicted distributions for the considered species were then used to identify (4) areas of importance for elasmobranch (species richness and diversity) and (5) potential bycatch hotspots in the scallop and Nephrops fisheries. This combined information is necessary to identify areas where elasmobranchs would benefit from the implementation of fisheries measures or candidate areas to be considered in the development of an elasmobranch action plan.
An important lesson learned from this approach is that, while elasmobranch are often considered as data deficient due to the lack of a single standardised source of data, multiple data sources, varying in quality and spatial extent, can be combined using recent modelling methods to provide the required evidence to inform future management decisions and help sustainability and resilience of commercial fisheries.
The distribution models in this study revealed that seven out of the nine elasmobranch species considered show preferences for depths of 100 m or deeper, which explains their predominance on the west of Scotland, where deep trenches can be found close inshore within sea lochs. The two species with either a preference for shallow depths (cuckoo ray) or no particular depth preference (spotted ray) were commonly found in the North Sea, to the east of Scotland. Seabed preferences ranged from soft to coarse substrates and most species preferred waters characterised by low bottom primary productivity and high salinity (except the starry smooth-hound and the starry ray).
Skates and rays were more often retained in the dredge compared to squaliform species and all species were caught in large numbers in bottom trawl surveys. Overlap in the distributions of the considered elasmobranchs and the scallop and Nephrops fisheries revealed a number of areas where conservation measures may have impact, of which the most significant are summarised in Figure 55. Such areas include the Sound of Jura, extending to the south of the Isle of Islay (South of Jura, Figure 55), areas east of the Kintyre peninsula and north-east of Arran in the Clyde (Kintyre and NE Arran respectively; Figure 55), an area to the south-east of Barra (South of Barra; Figure 55), and the deep channel leading from Raasay, just north of Rona in the inner sound of Skye (Skye to Red Point; Figure 55). On the east of Scotland, areas near the Rattray Head headland (Rattray Head; Figure 55) and east of Montrose in the Forth and Tay region (East of Montrose; Figure 55) were identified. This study predicts areas of high species richness and diversity for elasmobranchs in Scotland and improves our understanding of the potential interactions that elasmobranch species have with relevant fisheries. This information will make a significant contribution to informing and supporting future conservation actions and the management of sustainable fisheries.
6. References
Abella A.J., and Serena, F. 2005. Comparison of elasmobranch catches from research trawl surveys and commercial landings at port of Viareggio, Italy, in the last decade e-Journal of Northwest Atlantic Fishery Science, 35, 345-356.
Alverson, D.L., Freeberg, M.H., Pope, J.G. and Murawski, S.A. 1994. A global assessment of fisheries bycatch and discards FAO Fisheries Technical Paper. No. 339. Rome, FAO. 233p.
Anderson, S.C., Ward, E.J., English, P.A. and Barnett, L.A.K. 2022. sdmTMB: an R package for fast, flexible, and user-friendly generalized linear mixed effects models with spatial and spatiotemporal random fields bioRxiv 2022.03.24.485545.
ASMFC, 2008. Interstate Fishery Management Plan for Atlantic Coastal Sharks Management Report No. 46 of the Atlantic States Marine Fisheries Commission. 172p.
Assis, J., Tyberghein, L., Bosh, S., Verbruggen, H., Serrão, E.A. and De Clerck, O. 2017. Bio-ORACLE v2.0: Extending marine data layers for bioclimatic modelling Global Ecology and Biogeography, 27(3), 277-284.
Bache-Jeffreys, M., de Moraes, B.L.C., Ball, R.E., Menezes, G., Pálsson, J., Pampoulie, C., Stevens, J.R. and Griffiths, A.M. 2021. Resolving the spatial distributions of Dipturus intermedius and Dipturus batis—the two taxa formerly known as the ‘common skate’ Environmental Biology of Fishes, 104, 923-936 (2021).
Baeta, F., Batista, M., Maia, A., Costa, M.J. and Cabral, H. 2010. Elasmobranch bycatch in a trammel net fishery in the Portuguese west coast Fisheries Research 102(1-2), 123-129.
BAP, 2011. Biodiversity Action Plan (BAP) (Issue December 2011).
Barker. M.J. and Schluessel, V. 2005. Managing global shark fisheries: suggestions for prioritizing management strategies Aquatic Conservation: Marine and Freshwater Ecosystems, 15(4), 325-347.
Baum, J.K and Myers, R.A. 2004. Shifting baselines and the decline of pelagic sharks in the Gulf of Mexico Ecology Letters, 7(2), 135-145.
Beddington J.R. and Kirkwood G.P. 2005. The estimation of potential yield and stock status using life-history parameters Philosophical Transactions of the Royal Society B: Biological Sciences, 360(1453), 163-170.
Benjamins, S., Dodd, J., Thorburn, J., Milway, V.A., Campbell, R. and Bailey, D.M. 2018. Evaluating the potential of photo-identification as a monitoring tool for flapper skate (Dipturus intermedius) Aquatic Conservation: Marine and Freshwater Ecosystems, 28(6), 1360-1373.
Benjamins, S., Cole, G., Naylor, A., Thorburn, J.A. and Dodd, J. 2021. First confirmed complete incubation of a flapper skate (Dipturus intermedius) egg in captivity Journal of Fish Biology, 99(3), 1150-1154.
Brander, K. 1981. Disappearance of common skate Raia batis from Irish Sea Nature 290, 48-49.
Burgess, G.H. 2002. Spiny dogfishes: family Squalidae. In: Collette, B.B., Klein-MacPhee, G., eds. Bigelow and Schroeder’s Fishes of the Gulf of Maine. 3rd Edition. Washington, DC: Smithsonian Institution Press. p. 48-57.
Campana, S.E., Gibbson, A.J.F. Marks, L., Joyce, W., Rulifson, R.A. and Dadswell, M.J. 2008. Stock structure, life history, fishery and abundance indices for spiny dogfish (Squalus acanthias) in Atlantic Canada Fisheries and Oceans Canada, Science. Research Document 2007/89. Ottawa, Canada. 136p.
Carlson, A.E., Hoffmayer, E.R., Tribuzio, C.A. and Sulikowski, J.A. 2014. The use of satellite tags to redefine movement patterns of spiny dogfish (Squalus acanthias) along the U.S. east coast: Implications for fisheries management PLoS ONE, 9(7), e103384.
Clarke, T., Espinoza, M. & Ahrens, R. and Wehrtmann, I. 2015. Elasmobranch bycatch associated with the shrimp trawl fishery off the Pacific coast of Costa Rica, Central America Fishery Bulletin, 114(1), 1-17.
Clarke, M., Farrell, E.D., Roche, W., Murray, T.E., Foster, S. and Marnell, F. 2016. Ireland Red List No. 11: Cartilaginous fish [sharks, skates, rays and chimaeras] National Parks and Wildlife Service, Department of Arts, Heritage, Regional, Rural and Gaeltacht Affairs, Dublin, Ireland. 142p.
Compagno, L.J.V. 1984. Squaliformes-Dogfish sharksIn: Fischer, W., Nauen, C., eds. FAO species catalogue, Volume 4. Sharks of the world: An annotated and illustrated catalogue of shark species known to date. Part I. hexanchiformes to lamniformes. Vol. 125. FAO Fisheries Synopsis, Rome. pp. 24-129.
Compagno, L., Dando, M. and Fowler, S. 2005. Princeton field guides: sharks of the world Princeton University Press, Princeton, NJ. 496 p.
COSEWIC, 2012. COSEWIC assessment and status report on the Thorny Skate, Amblyraja radiata, in Canada Committee on the Status of Endangered Wildlife in Canada. Ottawa. ix + 75 pp.
Coulon, N., Elliott, S., Teichert, N., Auber, A., McLean, M., Barreau, T., Feunteun, E. and Carpentier, A. 2024. Northeast Atlantic elasmobranch community on the move: Functional reorganization in response to climate change Global Change Biology, 30(1), e17157.
Daan, N., Heessen, H.J.L. and ter Hofstede, R. 2005. North Sea Elasmobranchs: distribution, abundance and biodiversity ICES CM 2005/N:06. 15 pp.
D’Iglio, C., Albano, M., Tiralongo, F., Famulari, S., Rinelli, P., Savoca, S., Spanò, N. and Capillo, G. 2021. Biological and ecological aspects of the blackmouth catshark (Galeus melastomus Rafinesque, 1810) in the southern Tyrrhenian Sea Journal of Marine Science and Engineering, 9(9), 967.
Dell’Apa, A., J. Cudney-Burch, D. G. Kimmel, and R. A. Rulifson. 2014. Sexual segregation of spiny dogfish in fishery-dependent surveys in Cape Cod, Massachusetts: potential management benefitsTransactions of the American Fisheries Society, 143(4), 833-844.
Dell’Apa, A., Bangley, C.W. and Rulifson, R.A. 2015. Who let the dogfish out? A review of management and socio-economic aspects of spiny dogfish fisheries Reviews in Fish Biology and Fisheries, 25, 273-295.
Dell'Apa, A., Grazia, P.M. and Bonzek, C.F. 2016. Modeling the habitat distribution of spiny dogfish (Squalus acanthias), by sex, in coastal waters of the northeastern United States Fishery Bulletin, 115, 89-100.
Dodd, J., Baxter, J.M., Donnan, D.W., James, B.D., Lavender, E., McSorley, C.A., Mogg, A.O.M. and Thorburn, J.A. 2022. First report of an egg nursery for the Critically Endangered flapper skate Dipturus intermedia (Rajiformes: Rajidae) Aquatic Conservation: Marine and Freshwater Ecosystems, 32(10), 1647-1659.
Domenici, P., Standen, E.M. and Levine, R.P. 2004. Escape manoeuvres in the spiny dogfish (Squalus acanthias) Journal of Experimental Biology, 207(13), 2339-2349.
Du Buit, M.H. 1977. Age et croissance de Raja batis et de Raja naevus en Mer Celtique ICES Journal of Marine Science, 37(3), 261-265.
Dulvy, N.K. and Reynolds, J.D. 2002. Predicting extinction vulnerability in skatesConservation Biology, 16(2), 440-450.
Dulvy, N.K., Metcalfe, J.D., Glanville, J., Pawson, M.G. and Reynolds, J.D. 2000. Fishery stability, local extinctions, and shifts in community structure in skates Conservation Biology, 14(1), 283-293.
Dulvy, N.K., Pacoureau, N., Rigby, C.L., Pollom, R.A., Jabado, R.W., Ebert, D.A., Finucci et al. 2021. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis Current biology, 31(21), 4773-4787.e8.
Ebert, D.A. and Stehmann, M.F.W. 2013. Sharks, batoids, and chimaeras of the North Atlantic FAO Species Catalogue for Fishery Purposes No. 7. FAO, Rome. 523 p.
Eigaard, O., Bastardie, F., Breen, M., Dinesen, G., Hintzen, N., Laffargue, P., Nielsen J. et al. 2016. Estimating seabed pressure from demersal trawls, seines, and dredges based on gear design and dimensions ICES Journal of Marine Science, 73(Supplement 1), i27-i43.
Elliott, S.A.M., Carpentier, A., Feunteun, E. and Trancart, T. 2020. Distribution and life history trait models indicate vulnerability of skates Progress in Oceanography, 181, 102256.
Ellis, J. 2016. Raja clavata. The IUCN Red List of Threatened Species 2016, e.T39399A103110667.
Ellis, J.R., Cruz-Martinez, A., Rackham, B.D. and Rogers, S.I. 2005a. The distribution of chondrichthyan fishes around the British Isles and implications for conservationJournal of Northwest Atlantic Fisheries Science, 35, 195-213.
Ellis, J.R., Dulvy, N.K., Jennings, S., Parker-Humphreys, M. and Rogers, S.I. 2005b. Assessing the status of demersal elasmobranchs in UK waters: A review Journal of the Marine Biological Association of the United Kingdom, 85, 1025-1047.
Ellis, J., Ungaro, N., Serena, F., Dulvy, N.,Tinti, F., Bertozzi, M., Pasolini, P., Mancusi, C. and Noarbartolo di Sciara, G. 2007. Raja montagui The IUCN Red List of Threatened Species 2007, e.T63146A12623141.
Ellis, J., Burt, G.J., Cox, L.P.N., Kulka, D.W. and Payne, A.I.L 2008. The status and management of thornback ray Raja clavata in the south-western North Sea ICES CM 2008/K:13, 45pp. International Council for the Exploration of the Sea, Copenhagen.
Ellis, J, McCully, S.R., Sims, D., Walls, R.H.L., Cheok, J., Derrick, D. and Dulvy, N.K. 2021. Dipturus intermedius The IUCN Red List of Threatened Species 2021. IUCN Redlist.
Ellis, J.R. and Shackley, S.E. 1997. The reproductive biology of Scyliorhinus canicula in the Bristol Channel, UK Journal of Fish Biology, 51(2), 361-372.
Farrell, E.D., Clarke, M.W. and Mariani, S. 2009. A simple genetic identification method for Northeast Atlantic smoothhound sharks (Mustelus spp.) ICES Journal of Marine Science, 66(3), 561-565.
Farrell, E.D., Mariani, S. and Clarke, M.W. 2010. Reproductive biology of the starry smoothhound shark Mustelus asterias: Geographic variation and implications for sustainable exploitation Journal of Fish Biology, 77(7), 1505-1525.
Finucci, B., Cheok, J., Chiaramonte, G.E., Cotton, C.F., Dulvy, N.K., Kulka, D.W. et al. 2020. Squalus acanthias The IUCN red list of threatened species 2020, e.T91209505A124551959
Ford, R.B., Galland, A., Clark, M.R., Crozier, P., Duffy, C.A.J., Dunn, M.R., Francis, M.P. and Wells, R. 2015. Qualitative (Level 1) Risk Assessment of the impact of commercial fishing on New Zealand Chondrichthyans New Zealand Aquatic Environment and Biodiversity Report No. 157. 111p
Fordham, S., Fowler, S.L., Coelho, R.P., Goldman, K. and Francis, M. P. 2016. Squalus acanthias The IUCN Red List of Threatened Species 2016, e.T91209505A2898271.
Fowler, S.L., Cavanagh, R.D., Camhi, M., Burgess, G.H., Cailliet, G.M., Fordham, S.V., Simpfendorfer, C.A. and Musick, J.A. 2005. Sharks, rays and chimaeras: the status of the Chondrichthyan fishes IUCN Species Survival Commission Shark Specialist Group. IUCN, Gland, Switzerland, and Cambridge, UK.
Fowler, S. 2010. OSPAR commission: Background document for common skate Dipturus batis Biodiveristy Series. 19 p.
Gallagher, M.J., Nolan, C.P. and Jeal, F. 2005. Age, growth and maturity of the commercial ray species from the Irish Sea Journal of Northwest Atlantic Fisheries Science, 35, 47-66.
Garbett, A., Phillips, N.D., Houghton, J.D., Prodöhl, P., Thorburn, J., Loca, S.L., et al. 2020. The critically endangered flapper skate (Dipturus intermedius): Recommendations from the first flapper skate working group meeting Marine Policy, 124, 104367.
Gauld, J.A. and Macdonald, W.S. 1982. The results of tagging experiments on spurdogs, Squalus acanthias L., around Scotland ICES CM Doc 1982/H:51. International Council for the Exploration of the Sea, Copenhagen.
Gottschall, K., Johnson, M.W. and Simpson, D.G. 2000. The distribution and size composition of finfish, American Lobster, and long-finned squid in Long Island Sound based on the Connecticut Fisheries Division Bottom Trawl Survey, 1984- 1994 NOAA Tech. Rep. NMFS 148. 195 p.
Gräler, B., Pebesma, E., Heuvelink, G. 2016. Spatio-Temporal Interpolation using gstat The R Journal, 8, 204-218.
Greenstreet, S.P.R. and Hall, S.J. 1996. Fishing and the ground-fish assemblage structure in the north-western North Sea: an analysis of long-term and spatial trends Journal of Animal Ecology, 65(5), 577-598.
Griffiths, A.M., Sims, D.W., Cotterell, S.P., El Nagar, A., Ellis, J.R., Lynghammar, A. et al. 2010. Molecular markers reveal spatially segregated cryptic species in a critically endangered fish, the common skate (Dipturus batis) Proceedings of the Royal Society B: Biological Sciences, 277(1687), 1497–1503.
Griffiths, C.A., Wright, S.R., Silva, J.F., Ellis, J.R., Righton, D.A. and McCully Phillips, S.R. 2020. Horizontal and vertical movements of starry smooth-hound Mustelus asterias in the northeast Atlantic PLoS One, 15, e0239480.
Hauser, L., Franks, G. H., Vega, N. and Gallucci, V. 2007. Potential for sustainable expansion of the dogfish (Squalus acanthias) fishery in the northeast Pacific NOAA Report.
Heessen, H.J.L. ed. 2003. Development of elasmobranch assessments DELASS. Final report of DG Fish Study Contract 99/055, 605 pp.
Heessen, H.J.L. and Daan, N. 1996. Long-term trends in ten non-target North Sea fish speciesICES Journal of Marine Science, 53(6), 1063-1078.
Hogan, F., Cadrin, S. and Haygood, A. 2013. Fishery management complexes: An impediment of aid to sustainable harvest? A discussion based on the Northeast Skate complexNorth American Journal of Fisheries Management, 33(2), 406-421.
Holden, M.J., Rout, D.W. and Humphreys, C.N. 1971. The rate of egg laying by three species of ray ICES Journal of Marine Science, 33(3), 335-339.
ICES, 2007. Report of the Working Group of the Elasmobranch Fishes (WGEF). 22-28 June 2007, Galway, Ireland.
ICES, 2012a. ICES Implementation of Advice for Data-limited Stocks in 2012 in its 2012 Advice ICES CM 2012/ACOM 68. 42 pp.
ICES, 2012b. Report of the Working Group on Elasmobranch Fishes (WGEF), Lisbon, Portugal, 19-26 June 2007. ICES CM 2012/ACOM:19. International Council for the Exploration of the Sea (ICES), Denmark.
ICES, 2017. Smooth-hounds in the Northeast Atlantic. ICES Report of the Working Group of the Elasmobranch Fishes (WGEF). 31 May - 7 June 2017, Lisbon, Portugal.
ICES, 2018. Cuckoo ray (Leucoraja naevus) in subareas 6 and 7 and divisions 8.a–b and 8.d (West of Scotland, southern Celtic Seas, and western English Channel, Bay of Biscay) ICES Advice on fishing opportunities, catch, and effort Bay of Biscay and the Iberian Coast, Celtic Seas, Greater North Sea and Oceanic Northeast Atlantic ecoregions - rjn.27.678abd.
ICES, 2019. Working Group on Elasmobranch Fishes (WGEF) ICES Scientific Reports, 1:25. 964 pp.
ICES, 2020. Working Group on Elasmobranch Fishes (WGEF) ICES Scientific Reports 2: 77. 789 pp.
ICES, 2022a. ICES Survey Protocols ICES Publications. Collection.
ICES, 2022b. Cuckoo ray (Leucoraja naevus) in subareas 6 and 7, and in divisions 8.a–b and 8.d (West of Scotland, southern Celtic Seas, and western English Channel, Bay of Biscay) In: Report of the ICES Advisory Committee, 2022. ICES Advice 2022 – rjn.27.678abd.
ICES, 2022c. Thornback ray (Raja clavata) in Subarea 6 (West of Scotland) In: Report of the ICES Advisory Committee, 2022. ICES Advice 2022 – rjc.27.6.
ICES, 2022d. Thornback ray (Raja clavata) in Subarea 4 and in divisions 3.a and 7.d (North Sea, Skagerrak, Kattegat, and eastern English Channel) In: Report of the ICES Advisory Committee, 2023. ICES Advice 2023 – rjc.27.3a47d.
ICES, 2023. Working Group on Elasmobranch Fishes (WGEF) ICES Scientific Reports. 5:92. 837 pp.
ICES, 2023a. Common skate complex (Blue skate (Dipturus batis) and flapper skate (Dipturus intermedius) in Subarea 4 and Division 3.a (North Sea, Skagerrak and Kattegat) In Report of the ICES Advisory Committee, 2023. ICES Advice 2023 – rjb.27.3a4.
ICES, 2023b. Cuckoo ray (Leucoraja naevus) in Subarea 4 and Division 3.a (North Sea, Skagerrak, and Kattegat) ICES Advice 2023 – rjn.27.3a4.
ICES, 2023c. Spotted ray (Raja montagui) in Subarea 6 and divisions 7.b and 7.j (West of Scotland, west and southwest of Ireland) ICES Advice 2022 – rjm.27.67bj.
ICES, 2023d. Spotted ray (Raja montagui) in Subarea 4 and in divisions 3.a and 7.d (North Sea, Skagerrak, Kattegat, and eastern English Channel) ICES Advice 2023 –rjm.27.3a47d.
ICES, 2023e. Blackmouth dogfish (Galeus melastomus) in subareas 6 and 7 (West of Scotland, southern Celtic Seas, and English Channel) ICES Advice 2023 –sho.27.67.
ICES, 2023f. Lesser-spotted dogfish (Scyliorhinus canicula) in Subarea 6 and divisions 7.a–c and 7.e–j (West of Scotland, Irish Sea, southern Celtic Seas) ICES Advice 2023 – syc.27.67a-ce-j.
ICES, 2023g. Lesser-spotted dogfish (Scyliorhinus canicula) in Subarea 4 and in divisions 3.a and 7.d (North Sea, Skagerrak and Kattegat, eastern English Channel) ICES Advice 2023 – syc.27.3a47d.
ICES, 2023h. Smooth-hound (Mustelus spp.) in subareas 1–10, 12, and 14 (Northeast Atlantic and adjacent waters) ICES Advice 2023 – sdv.27.nea.
Iglésias, S.P., Toulhoat, L. and Sellos, D.Y. 2010. Taxonomic confusion and market mislabelling of threatened skates: Important consequences for their conservation status Aquatic Conservation: Marine and Freshwater Ecosystems 20(3), 319–333.
Jabado, R.W., Ellis, J.R., McCully, S., Dulvy, N.K., Farrell, E.D., Mancusi, C., and Derrick, D. 2021. Mustelus asterias. The IUCN Red List of Threatened Species 2021, e.T39357A124405496.
Jones, T.S. and K.I. Ugland. 2001. Reproduction of female spiny dogfish Squalus acanthias in the Oslofjord Fisheries Bulletin, 99(4), 685-690.
Jorgensen, S.J., Micheli F., White T.D. Van Houtan K.S., Alfaro-Shigueto J., Andrzejaczek S. et al. 2022. Emergent research and priorities for shark and ray conservation Endangered Species Research, 47, 171–203.
Kadri, H., Marouani, S., Saïdi, B., Bradai, M.N., Bouaïn, A., and Morize, E. 2014. Age, growth, sexual maturity and reproduction of the thornback ray, Raja clavata (L.) of the Gulf of Gabès (south-central Mediterranean Sea) Marine Biology Research, 10(4), 416-425.
Kynoch, R. J., Fryer, R. J., and Neat, F. C. 2015. A simple technical measure to reduce bycatch and discard of skates and sharks in mixed-species bottom-trawl fisheries. ICES Journal of Marine Science, 72(6).
Lack, M. and Sant, G. 2011. The future of sharks: a review of action and inaction TRAFFIC report. TRAFFIC International and the Pew Environment Group, Cambridge.
Langton, R., Stirling, D.A., Boulcott, P. and Wright, P.J. 2020. Are MPAs effective in removing fishing pressure? Biological Conservation, 247, 108511.
Liaw, A. and Wiener, M. 2002. Classification and Regression by Random Forest R News, 2, 18-22.
Maia, C., Erzini, K., Serra-Pereira, B. and Figueiredo, I. 2012. Reproductive biology of cuckooray Leucoraja naevus Journal of Fish Biology, 81(4), 1285–1296.
Martin, C.S., Vaz S., Ellis, J.R., Lauria, V., Coppin, F. and Carpentier, A.2012. Modelled distributions of ten demersal elasmobranchs of the eastern English Channel in relation to the environment Journal of Experimental Marine Biology and Ecology, 418–419, 91–103.
McAllister, M., Fraser, S. and Henry, L.-A. 2023. Population ecology and juvenile density hotspots of thornback ray (Raja clavata) around the Shetland Islands, ScotlandJournal of Fish Biology, 104(3), 576-589.
McEachran, J.D. 2002. Skates. Family Rajidae. In: Collette, B.B., Klein-MacPhee, G., eds. Bigelow and Schroeder’s fishes of the Gulf of Maine. 3rd Edition. Washington, DC: Smithsonian Institution Press.
McEachran, J.D. and Musick, J.A. 1975. Distribution and relative abundance of seven species of skates (Pisces:Rajidae) which occur between Nova Scotia and Cape HatterasFishery Bulletin, 73(1), 110-136.
McFarlane, G.A. and King, J.R. 2003. Migration patterns of spiny dogfish in the North Pacific Ocean —Squalus acanthias Fisheries Bulletin, 101, 358−367.
McMillan, D. and Morse, W. 1999. Essential fish habitat source document: Spiny dogfish, Squalus acanthias, life history and habitat characteristics In: ed., NOAA Technical Memorandum NMFS-NE:150. 19 p.
Methratta, E.T. and Link, J.S. 2007. Ontogenetic variation in habitat association for four groundfish species in the Gulf of Maine–Georges Bank regionMarine Ecology Progress Series, 338, 169–181.
Moriarty, M. 2021. Changes in species distribution for temperate marine fish: historic analysis and future projections on the western European margin Thesis submitted for the degree of Doctor of Philosophy, Ulster University. Ulster University, Ulster.
Myers, R.A. and Worm, B. 2005. Extinction, survival or recovery of large predatory fishesPhilosophical Transactions of the Royal Society B: Biological Sciences, 360(1453), 13–20.
Mytilineou, C., Politou, C.-Y., Papaconstantinou, C., Kavadas, S., D'Onghia G. and Sion, L. 2005. Deep-water fish fauna in the Eastern Ionian Sea Belgian Journal of Zoology, 135(2), 229–233.
Navas, J.M., Miller P.I., Henry, L.-A., Hennige, SJ, Roberts, J.M. 2014. Ecohydrodynamics of cold-water coral reefs: A case study of the Mingulay reef complex (Western Scotland) PLoS ONE, 9(5), e98218.
Neat, F., Pinto, C., Burrett, I., Cowie, L., Travis, J., Thorburn, J. et al. 2015. Site fidelity, survival and conservation options for the threatened flapper skate (Dipturus cf. intermedia) Aquatic Conservation: Marine and Freshwater Ecosystems, 25(1), 6–20.
Nieto, A., Ralph, G.M., Comeros-Raynal, M.T., Kemp, J., García Criado, M., Allen, D.J.et al. 2015. European Red List of marine fishes Luxembourg: Publications Office of the European Union.
Packer, D.B., Zetlin, C.A. and Vitaliano, J.J. 2003. Essential fish habitat source document. Thorny skate, Amblyraja radiata, life history and habitat characteristics NOAA technical memorandum NMFS-NE; 178. 50 pp.
Papadopoulo, K., Villegas‐Ríos, D., Mucientes, G., Hillinger, A. and Alonso‐Fernández, A. 2023. Drivers of behaviour and spatial ecology of the small spotted catshark (Scyliorhinus canicular) Aquatic Conservation: Marine and Freshwater Ecosystems, 33(5), 443–457.
Parson, J.M., Fish, F.E. and Nicastro, A.J. 2011. Turning performance of batoids: Limitations of a rigid body Journal of Experimental Marine Biology and Ecology, 402(1–2), 12–18.
Pauly, D. and Murphy, G.I. eds. 1982. Theory and management of tropical fisheries ICLARM Conference Proceedings 9, 360 p. International Center for Living Aquatic Resources Management, Manila, Philippines and Division of Fisheries Research, Commonwealth Scientific and Industrial Research Organisation, Cronulla, Australia.
Pennino, G.M., Muñoz, F. Conesa, D. López-Quίlez, A. and Bellido, J.M. 2013. Modeling sensitive elasmobranch habitats Journal of Sea Research, 83, 209–218.
Piet, G.J., Van Hal, R., Greenstreet, S.P.R. 2009. Modelling the direct impact of bottom trawling on the North Sea fish community to derive estimates of fishing mortality for non-target fish species ICES Journal of Marine Science, 66(9), 1985–1998.
Pinto, C. 2015. Integration of multiple sparse and limited datasets helps to inform spatial conservation for an endangered marine species A thesis presented for the degree in Doctor of Philosophy in Ecology at the University of Aberdeen. University of Aberdeen, Aberdeen.
Pinto, C., Thorburn, J.A., Neat, F., Wright, P.J., Wright, S., Scott, B.E. et al. 2016. Using individual tracking data to validate the predictions of species distribution models Diversity and Distributions, 22(6), 682–693.
Polovina, J.J., Abecassis, M., Howell, E.A. and Woodworth, P.A. 2009. Increases in the relative abundance of mid-trophic level fishes concurrent with declines in apex predators in the subtropical North Pacific, 1996–2006 Fishery Bulletin, 107(4), 523–531.
Pope, J.G., Macdonald, D.S., Daan, N., Reynolds, J.D. and Jennings, S. 2000. Gauging the impact of fishing mortality on non-target species ICES Journal of Marine Science, 57(3), 689–696.
Régnier, T., Dodd, J., Benjamins, S., Gibb, F.M. and Wright, P.J. 2024. Spatial management measures benefit the critically endangered flapper skate, Dipturus intermedius Aquatic Conservation: Marine and Freshwater Ecosystems, 34(4), e4150.
Rindorf, A., Gislason, H., Burns, F., Ellis, J. R. and Reid, D. 2020. Are fish sensitive to trawling recovering in the Northeast Atlantic? Journal of Applied Ecology, 57(10), 1936–1947.
Rodrı́guez-Cabello, C., Sánchez, F., Fernández, A. and Olaso, I. 2004. Is the lesser spotted dogfish (Scyliorhinus canicula) population from the Cantabrian Sea a unique stock? Fisheries Research, 69(1), 57–71.
Rodríguez-Cabello, C., Fernandez, A., Olaso, I. and Sanchez, F. 2005. Survival of small-spotted catshark (Scyliorhinus canicula) discarded by trawlers in the Cantabrian SeaJournal of the Marine Association of the United Kingdom, 85(5), 1145–1150.
Roe, J.H., Morreale, S.J., Paladino, F.V., Shillinger G.L., Benson S.R., Eckert S.A., Bailey H., et al. 2014. Predicting bycatch hotspots for endangered leatherback turtles on longlines in the Pacific Ocean Proceedings of the Royal Society B: Biological Sciences, 281, 28120132559.
Sagarese, S. R., M. G. Frisk, T. J. Miller, K. A. Sosebee, J. A. Musick, and P. J. Rago. 2014. Influence of environmental, spatial, and ontogenetic variables on habitat selection and management of spiny dogfish in the Northeast (US) shelf large marine ecosystem Canadian Journal of Fisheries and Aquatic Sciences, 71(4), 567–580.
Sanchez, P., Demestre, V., Ramon, M. and Kaiser, M.J. 2000. The impact of otter trawling on mud communities in the northwestern Mediterranean ICES Journal of Marine Science, 57(5), 1352–1358.
Santos, C.P., Sampaio, E., Pereira, B.P.., Pegado, M.R., Borges, F.O., Wheeler, C.R., Bouyoucos, I.A., Rummer, J.L., Frazão Santos, C. and Rosa, R. 2021. Elasmobranch responses to experimental warming, acidification, and oxygen loss—a meta-analysis Frontiers in Marine Science, 8, 735377.
Scott, J.S. 1982. Selection of bottom type by groundfishes of the Scotian Shelf Canadian Journal of Fisheries and Aquatic Sciences, 39(7), 943–947.
Seamone, S., Blaine, T. and Higham, T.E. 2014. Sharks modulate their escape behavior in response to predator size, speed and approach orientation Zoology, 117(6), 377–382.
Serena, F. 2005. Field identification guide to the sharks and rays of the Mediterranean and Black Sea FAO Species Identification Guide for Fishery Purposes. Rome, FAO. 2005. 97 p.
Serena, F., Mancusi, C., Morey, G. and Ellis, J.R. 2009. Dasyatis pastinaca. In: IUCN 2013. IUCN Red List of Threatened Species Version 2013.1.
Sguotti, C., Lynam, C.P., Garcia-Carreras, B., Ellis, J.R. and Engelhard, G.H. 2016. Distribution of skates and sharks in the North Sea: 112 years of change Global Change Biology, 22(8), 2729-2743.
Shepherd, T., Page, F. and B. Macdonald, B. 2002. Length and sex-specific associations between spiny dogfish (Squalus acanthias) and hydrographic variables in the Bay of Fundy and Scotian Shelf Fisheries Oceanography, 11(2), 78–89.
Silva, J.F. Ellis, J.R. and Catchpole T.L. 2012. Species composition of skates (Rajidae) in commercial fisheries around the British Isles and their discarding patterns Journal of Fish Biology, 80(5), 1678–1703.
Sosebee, K.A., Miller, A., O’Brien, L. McElroy, D. and Sherman, S. 2016. Update of thorny skate commercial and survey data NOAA National Marine Fisheries Service Center Reference Document 16-08. 137 pp.
Stehlik, L.L. 2007. Essential Fish Habitat Source Document: Spiny Dogfish, Squalus acanthias, Life History and Habitat Characteristics 2nd ed. December 2007. NOAA Technical Memorandum NMFS-NE-203. US Department of Commerce, Massachusetts. 52pp.
Stehmann, M. and Bȕrkel, D.L. 1984. Rajidae In: Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese, E. eds. Fishes of the North-eastern Atlantic and Mediterranean. Volume 1. UNESCO, Paris. pp. 163–196
Stein, R.W., Mull, C.G., Kuhn, T.S., Aschliman, N.C., Davidson, L.N., Joy, J.B., Smith, G.J., Dulvy, N.K. and Mooers, A.O. 2018. Global priorities for conserving the evolutionary history of sharks, rays and chimaeras Nature Ecology and Evolution, 2, 288–298.
Stevens, J.D., Bonfil, R., Dulvy, N.K. and Walker, P.A. 2000. The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems ICES Journal of Marine Science, 57(3), 476–494.
Templeman, W. 1984. Migrations of spiny dogfish, Squalus acanthias, and recapture success from tagging in the Newfoundland area, 1963–65. Journal of Northwest Atlantic Fisheries Science, 5, 47–53.
Thorburn, J., Neat, F., Bailey, D.M., Noble, L.R. and Jones, C.S. 2015. Winter residency and site association in the Critically Endangered North East Atlantic spurdog Squalus acanthiasMarine Ecology Progress Series, 526, 113–124.
Thorburn, J., Dodd, J. and Neat, F. 2018a. Spatial ecology of flapper skate (Dipturus intermedius - Dipturus batis complex) and spurdog (Squalus acanthias) in relation to the Loch Sunart to the Sound of Jura Marine Protected Area and Loch Etive SNH Research Report No. 1011. NatureScot, UK.
Thorburn, J., Jones, R., Neat, F., Pinto, C., Bendall, V., Hetherington, S., Bailey, D. M., Leslie, N. and Jones, C. 2018b. Spatial versus temporal structure: Implications of inter-haul variation and relatedness in the North-east Atlantic spurdog Squalus acanthias Aquatic Conservation: Marine and Freshwater Ecosystems, 28(5), 1167–1180.
Thorburn, J., Lavender, E., Cole, G., Smout, S., and James, M. 2022. Movement Ecology of Flapper Skate report.
Vilmar, M. and Di Santo, V. 2022. Swimming performance of sharks and rays under climate change Reviews of Fish Biology and Fisheries, 32, 765–781.
Walker, P.A. 1999. Fleeting images: dynamics of North Sea ray populations PhD Thesis, University of Amsterdam, the Netherlands, 145 pp.
Walker P.A. and Heessen H.J.L. 1996. Long-term changes in ray populations in the North SeaICES Journal of Marine Science, 53(6), 1085–1093.
Walker, P.A. and Hislop, J.R.G. 1998. Sensitive skates or resilient rays? Spatial and temporal shifts in ray species composition in the central and north-western North Sea between 1930 and the present day ICES Journal of Marine Science, 55(3), 392–402.
Walker, N. D., Maxwell, D. L., Le Quesne, W. J. F., and Jennings, S. 2017. Estimating efficiency of survey and commercial trawl gears from comparisons of catch-ratios ICES Journal of Marine Science, 74(5), 1448–1457.
Wearmouth, V.J. and Sims, D.W. 2009. Movement and behaviour patterns of the critically endangered common skate Dipturus batis revealed by electronic tagging Journal of Experimental Marine Biology and Ecology, 380(1–2), 77–87.
Weigmann, S. 2016. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity Journal of Fish Biology, 88(3), 837–1037.
Wheeler, A. 1978. Key to the fishes of northern Europe. Frederick Warne, London.
Whittamore, J.M. and McCarthy, I.D. 2005. The population biology of the thornback ray, raja clavata in Caernarfon Bay, North Wales Journal of the Marine Biological Association of the United Kingdom, 85(5), 1089–1094.
Wood, S.N. 2003. Thin-plate regression splines Journal of the Royal Statistical Society (B), 65(1), 95–114.
Zamon, J.E. 2003. Mixed species aggregations feeding upon herring and sandlance schools in a nearshore archipelago depend on flooding tidal currentsMarine Ecology Progress Series, 261, 243–255.
Annexe A: Survey gear importance in the Random Forest models
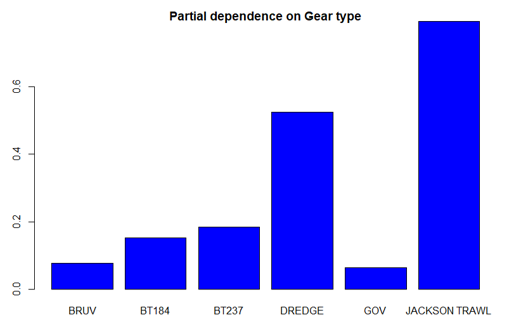
Figure A1. Partial dependence of the Random Forest model on survey gear for the flapper skate.
Click for a full description
A bar chart shows that Jackson trawl and dredge are the two most important fishing gear types influencing flapper skate abundance.
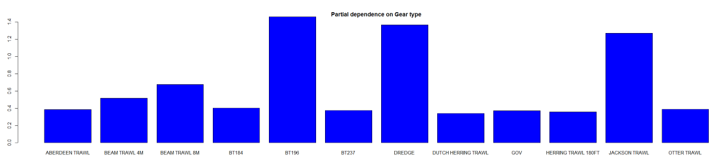
Figure A2. Partial dependence of the Random Forest model on survey gear for the Cuckoo Ray.
Click for a full description
A bar chart shows that BT196 and dredge are the two most important fishing gear types influencing cuckoo ray abundance.
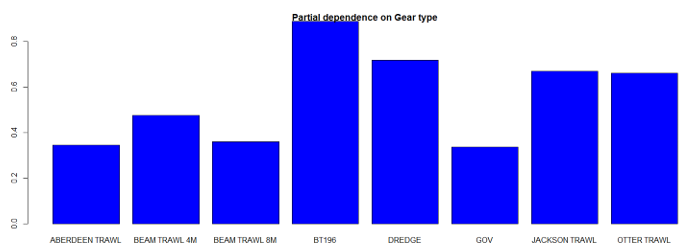
Figure A3. Partial dependence of the Random Forest model on survey gear for the Spotted Ray.
Click for a full description
A bar chart shows that BT196 and dredge are the two most important fishing gear types influencing spotted ray abundance.
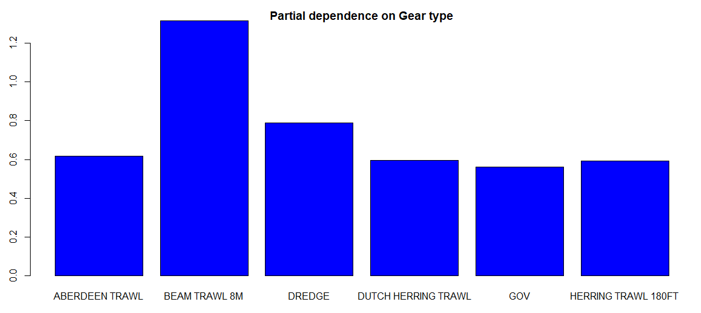
Figure A4. Partial dependence of the Random Forest model on survey gear for the Starry Ray.
Click for a full description
A bar chart shows that beam trawl (8m) and dredge are the two most important fishing gear types influencing starry ray abundance.
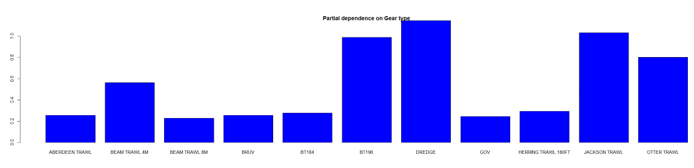
Figure A5. Partial dependence of the Random Forest model on survey gear for the Thornback Ray.
Click for a full description
A bar chart shows that the dredge and Jackson trawl are the two most important fishing gear types influencing thornback ray abundance.
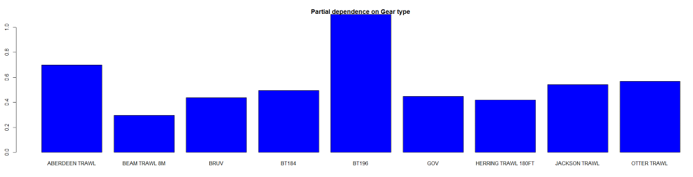
Figure A6. Partial dependence of the Random Forest model on survey gear for the Spurdog.
Click for a full description
A bar chart shows that BT196 and Aberdeen trawl are the two most important fishing gear types influencing spurdog abundance.
Figure A7. Partial dependence of the Random Forest model on survey gear for the Blackmouth dogfish.
Click for a full description
A bar chart shows that BT196 and Jackson trawl are the two most important fishing gear types influencing black-mouth dogfish abundance.
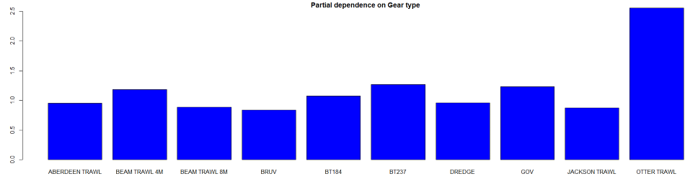
Figure A8. Partial dependence of the Random Forest model on survey gear for the Lesser spotted dogfish.
Click for a full description
A bar chart shows that otter trawl and BT237 are the two most important fishing gear influencing lesser-spotted dogfish abundance.
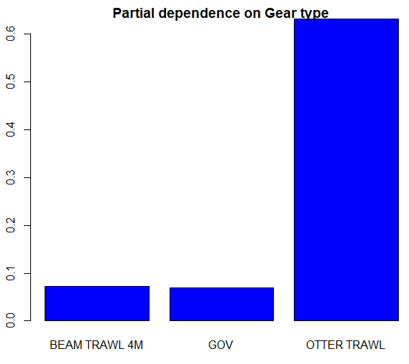
Fugure A9. Partial dependence of the Random Forest model on survey gear for the Starry smooth-hound.
Click for a full description
A bar chart shows that otter trawl is the most important fishing gear type influencing starry smooth-hound abundance.





