NatureScot Research Report 876 - Preliminary assessment completed in 2015 of the feasibility of maintaining, limiting or eradicating feral pigs in Scotland
Year of publication: 2022
Authors: Massei G. and Ward A.
Cite as: Massei G. and Ward A. 2022. Preliminary assessment completed in 2015 of the feasibility of maintaining, limiting or eradicating feral pigs in Scotland. NatureScot Research Report 876.
Note: this interim report was undertaken to inform the development of Scottish Government policy on feral pigs in Scotland. It was completed in 2015. Since this report was published, the number and distribution of feral pigs in Scotland has changed, as has the evidence for different methods of managing feral pig populations and the risk of diseases such as African Swine Fever. The non native species risk assessment for feral pigs was updated in 2016.
Keywords
wild boar; distribution; population management; population dynamics; impact; risk assessment; feral pig
Background
In the UK, wild boar were hunted to extinction by 1300 AD but became re-established in England, Scotland and Wales in the recent decades as a result of escapes from wild boar farms and illegal releases of hybrids between wild boar and domestic pigs.
The overall objective of this project is to review practical, humane and cost-effective options for managing populations of feral pigs in Scotland. The results will provide NatureScot with science-based evidence to develop informed policies for the management of feral pigs in Scotland.
Main findings
The distribution, size and dynamics of feral pig populations in the UK:
- self-sustaining populations of feral pigs occur in England, Scotland and Ireland
- in Scotland there are at least two breeding populations of free-living feral pigs and sightings of free-living animals in several other locations throughout Scotland.
- estimates suggest 50-100 animals per population, with more than 100 animals at Lochaber but very little information is available on the number of pigs in each population.
- at least 1300 feral pigs are kept in captivity across many sites and private estates in Scotland.
Farms with captive animals are not required by law to equip pigs with unique identification marks such as ear tags unless the animals are moved away from the premises. As a consequence, illegal releases or escapes cannot be traced back to the original source.
The information required to model feral pig population dynamics in Scotland:
- The key population parameters that must be collected are natality, mortality and an estimate of the size of the population
- natality can be estimated as a combination of litter size and proportion of females that reproduce in a year
- mortality can be estimated by marking new-born animals and assessing their survival throughout subsequent years or from hunting statistics.
Four methods to detect feral pig presence are proposed:
- large-scale mapping of feral pig sightings,
- bait stations with camera traps,
- camera grids and activity signs on transects, and
- putative attractants
Four methods are proposed to assess feral pig population trends or estimate density:
- Passive Activity Index based on camera trap grids and activity signs on transects,
- density estimates based on camera trap grids,
- distance sampling through thermal imaging and
- road traffic accidents.
We drafted a risk assessment of feral pigs in Scotland using the GB Non-Native Species Secretariat’s template. This assessment concludes that the current social, economic and environmental impact of feral pigs in Scotland is likely to be localised and minor because their number is still low and their distribution is limited to a few areas. Feral pigs may increase significantly in numbers and range, and then they could have a major social and economic impact and more conflicts could be expected if action is not taken.
To eradicate populations of feral pigs or control their numbers, a combination of trapping and shooting is most likely to be successful in Scotland.
- We modelled the effort required to maintain population size or eradicate a population of feral pigs in Scotland and suggest that eradication or population stabilization through culling appears to be feasible in Scotland, given current best estimates of the species’ distribution and abundance.
- Trapping is predicted to be less expensive than shooting although the majority of feral pig eradication campaigns use a combination of shooting and trapping.
- Costs are likely to increase with declining culling efficiency as population density decreases.
We make several recommendations to address the information gaps regarding the number and distribution of feral pigs in Scotland:
- establish a central record of distribution and numbers of free-living and captive feral pigs,
- test different methods of assessing presence and density of feral pigs in Scotland,
- collect data about natality and mortality and on factors that may affect natality and mortality such as age, gender, body mass as well as climate, availability of natural resources and supplementary food, diseases and population management,
- use this data to model feral pig population dynamics,
- coordinate stakeholders’ roles and responsibilities and train stakeholders in methods to estimate local population size and presence of feral pigs,
- field-test different population management options through an adaptive management framework where lessons learned are proactively used to inform and optimise mitigation of human-feral pig conflicts.
Acknowledgements
We are grateful to Steve Campbell (SASA), Bruce Sewell (Forest Enterprise Scotland), Ron Rose and Colin Lavin (Forestry Commission Scotland), Janet Talling (APHA), Charlie Wilson (Natural England), Morag Milne and Stan Whitaker (NatureScot), Niall Moore and Olaf Booy for commenting and for providing advice on various aspects of the report.
Introduction
Wild boar and feral pigs (Sus scrofa) are among the most widely distributed mammals in the world either as a native or as an introduced species. They are characterised by the highest reproductive rate among ungulates, with annual increases in population which may exceed 200%. Worldwide, recreational hunting is rarely able to control the growth of wild boar and feral pig populations that are increasing in numbers and range. This species’ environmental and economic impact include damage to crops and livestock, vehicle collisions, transmission of diseases to wildlife, livestock and people and reduction in plant and animal abundance and richness. Current trends of human and wild boar population growth and landscape development indicate that the impact of wild boar and feral pigs is likely to increase in the near future.
In the UK, wild boar were hunted to extinction by 1300 AD but became re-established in England, Scotland and Wales in the recent decades as a result of escapes from wild boar farms and illegal releases of hybrids between wild boar and domestic pigs. Boar can interbreed with domestic pigs and the genetics of the established feral populations may come from a mix of both wild boar and domestic pigs. Scottish Natural Heritage therefore refers to these animals as ‘feral pigs’.
In Scotland, as well as in the rest of the UK, feral pig populations still appear to be isolated from each other. In the absence of adequate control, feral pig populations will likely increase in the UK. Scotland is in the favourable position of being able to develop a pro-active approach to feral pig population management before potential conflicts escalate.
The overall objective of this project is to review practical, humane and cost-effective options for managing populations of feral pigs in Scotland. The results will provide NatureScot with science-based evidence to develop informed policies for the management of feral pigs in Scotland.
The specific aims of the project are:
- To review available data and knowledge gaps on the distribution, size and dynamics of feral pig populations in Scotland and to identify methods that could be used to address these gaps.
- To identify key information required to inform a model that will describe feral pig population dynamics in Scotland and anticipate population responses to management.
- To draft a risk assessment of feral pigs in Scotland using the GB Non-Native Species Secretariat’s template
- To review strengths and weaknesses of practical options for maintaining, limiting, or eradicating feral pigs and to estimate the amount of effort required to deliver each option or combinations of options in Scotland.
Objective 1 - Review of available data and knowledge gaps on the distribution, size and dynamics of feral pig populations in Scotland and identification of methods that could be used to address these gaps
Objective 1 comprised the following sections:
- An overview of wild boar ecology and potential environmental impact, based on published peer-reviewed papers.
- A review of population trends of wild boar in mainland Europe and of feral pigs in the US, where non-native feral pigs have significantly increased in the last 20 years. The review was based on published literature, internal reports made available to the Animal Plant and Health Agency (APHA) by various organisations and on data made available by European and UK colleagues. The review highlighted factors affecting the expansion of wild boar and feral pig populations worldwide, with particular reference to Europe.
- A review of the current data on distribution of feral pig populations in the UK in the last decades. This was carried out by drawing on published papers and reports on distribution and potential spread of this species across Great Britain (e.g. Wilson, 2005 and 2014; Moore & Wilson 2005; Defra, 2008; McDevitt et al., 2013) and in particular across Scotland. Data from Scotland were based on the report by Campbell & Hartley (2010) on locations of feral pigs in Scotland and on the SNH (2011) preliminary assessment of feral pig populations that was updated at the beginning of this project. The review also identified gaps in the knowledge of the ecology, behaviour and distribution of feral pigs in Scotland. Methods are proposed to address these gaps in the short and medium term (3-5 years).
Wild boar ecology and environmental impact
Wild boar and feral pigs (Sus scrofa) are among the most widely distributed large mammals in the world. The natural range of this species is Eurasia (Sjarmidi & Gerard, 1988) but wild boar and feral pigs have been introduced to the US, South America, Australia, New Zealand and Africa (Choquentot et al., 1996; Engeman et al., 2001; Mayer & Brisbin, 2009). Throughout this section, the species will be referred to as “wild boar” or “feral pigs” where appropriate.
Wild boar occur in a wide range of habitats which include woodlands, marshlands, Alpine grasslands, agricultural environments and suburban areas. Provided that high-energy food (such as acorns and crops), shelter and fresh water are available, this species can live virtually everywhere (Schley & Roper, 2003; Massei & Genov, 2004; Rosvold & Andersen, 2008; Barrios-Garcia & Ballari, 2012). Snow cover that persists for several weeks prevents wild boar from rooting for food and hampers their movements and can thus limit the distribution of this species. However, wild boar populations occur in countries characterised by harsh winters, such as Sweden and Norway, where widely spread supplementary feeding increases the winter survival of this species (Rosvold & Andersen, 2008; Thurfjell et al., 2013).
Wild boar live in social groups composed of females and piglets; adult males are solitary, whilst juvenile males form temporary groups (Gerard & Campan, 1988). During the mating season, that may extend for a few months between September and January, males join a group of females and fight other males to gain access to the sows. Among ungulates, wild boar are characterised by the highest annual reproductive rate that may exceed 200% (Bieber & Ruf, 2005; Fonseca et al., 2011; Keuling et al., 2013). Females can reach sexual maturity in their first year of age (Gethoffer et al., 2007) and may give birth in any month (Maillard & Fournier, 2004), although most births occur in spring. After a gestation of circa 114 days females build a nest and give birth to a litter of 3-7 piglets (e.g. Massei et al.,1996; Nahlik & Sandor, 2003; Fonseca et al., 2004; Gethoffer et al., 2007). Females produce one litter, and occasionally two litters per year: the number of litters, the litter size and the proportion of females that reproduce in a year are strongly affected by the availability of energy-rich food such as acorns (e.g. Groot-Bruinderink et al., 1994; Massei et al., 1996).
The main causes of natural mortality for this species are starvation due to extreme weather conditions (Okarma et al., 1995; Massei et al., 1997), diseases (e.g. Rossi et al., 2011) and, to a lesser extent, predation by wolf (Canis lupus) (Jędrzejewski et al., 1992; Nores et al., 2008). Hunting, followed by road traffic accidents, make the greatest contribution to wild boar mortality (e.g. Toigo et al., 2008; Gamelon et al., 2011; Keuling et al., 2013; Morelle et al., 2013; Prevot & Licoppe, 2013).
The environmental impact of wild boar and feral pigs on conservation and economic interests includes spread of diseases to wildlife, livestock and people, vehicle collisions, damage to crops and amenities, predation on and competition with native species, changes in the species composition of plant and animal communities and in the cycling of soil nutrients (e.g. Welander, 2000; Hone, 2002; Schley & Roper, 2003; Massei & Genov, 2004; Bueno et al., 2010; Barrios-Garcia & Ballari, 2012). The following section will focus on the environmental impact of wild boar as the other types of impact are briefly described in the Risk Assessment under Objective 3.
Wild boar are omnivores that feed on plants, vertebrates and invertebrates, eggs and crops (Schley & Roper, 2003; Massei & Genov, 2004). As opportunistic feeders, wild boar change their diet according to the season and to the relative availability of different food sources (Genov, 1981; Massei et al., 1996; Schley & Roper, 2003). The environmental impact derives from direct consumption of native plants and animals, effects of rooting on plant and animal species and on soil processes, seed dispersal and competition for food resources with other wildlife.
Wild boar in Western Europe feed on around 400 species of plants, animals and fungi (Schley & Roper, 2003). In particular, wild boar obtain a large proportion of their diet from rooting for roots, bulbs, fern rhizomes, invertebrate larvae and earthworms (Kotanen, 1995; Baubet et al., 2003; Schley & Roper, 2003). The bulk of the diet consists of plant material such as mast, roots, tubers, wild fruits and crops. Energy-rich plant food, like acorns and pine nuts as well as agricultural crops such as maize, wheat, oats and potatoes are particularly important as wild boar are not ruminants like other ungulates (Mackin, 1970; Andrzejewski & Jezierski, 1978; Groot Bruinderink et al., 1994; Massei et al., 1996; Schley & Roper, 2003).
Most crops are consumed when available (Mackin, 1970; Genov, 1981; Schley & Roper, 2003). Animals are regular components in the diet of the wild boar but occur in relatively small proportion compared to vegetable matter (Schley & Roper, 2003). Among animal foods, insects, earthworms, birds and mammals are eaten most consistently but the diet also include amphibians, reptiles, gastropods and myriapods as well as carcasses of larger animals and eggs of ground-nesting birds (Genov, 1981; Schley & Roper, 2003; Barrios-Garcia & Ballari, 2012). Wild boar may exploit mast hoards collected by small mammals and stored underground (Focardi et al., 2000) and may compete for food with other ungulates as well as with omnivores such as the Eurasian badger (Meles meles) whose diet overlaps with that of wild boar (Schley & Roper, 2003; Massei & Genov, 2004; Barrios-Garcia & Ballari, 2012).
Very few of the many studies conducted on the diet of wild boar and feral pigs have quantified the impact of wild boar on plant and animal populations. One of these studies, conducted in the Great Smoky Mountains National Park in the US, showed that rooting at a specific site may occur as many as 3 to 7 times per growing season and up to 80% of the woodland can be rooted every year (Howe et al., 1981). This study found that the five forbs species preferred by wild boar decreased in abundance from 30% of the area which was unoccupied by wild boar to 1% in sites where wild boar had been present for 20 years. In the latter, the abundance of soil macroinvertebrates declined from 2.2 x 106/ ha to 0.5 x 106/ ha. Another study conducted in the same area showed that the depth and weight of forest litter were at least 60% lower in intensively rooted sites compared to site without wild boar (Singer et al., 1984). This study also highlighted the indirect impact of wild boar rooting on ground-dwelling small mammals and found that repeated rooting decreases the food available to insectivores and destroys the habitat of surface-dwelling rodents. Mammals such as the red-backed vole (Clethrionomys gapperi) and short-tailed shrew (Blarina brevicauda) associated with habitats such as the leaf-litter were nearly eliminated from intensively rooted sites. In contrast, the density of semi-arboreal small mammals, whose habitat was not affected by rooting, did not change (Singer et al., 1984). Other studies conducted in the rain forest in Hawaii found that the removal of feral pigs resulted in the abundance of springtails (Collembola spp.) to increase by 3.5 times and in the total biomass of microarthropods to increase by 2.5 times (Vtorov, 1993). Conversely, studies carried out in Australia suggest that direct feral pig predation on snake-necked turtle (Chelodina rugosa) resulted in strong compensatory response such as juvenile turtle recruitment, early onset of maturity and fast post-maturity growth. These responses decreased significantly the impact that feral pigs have on the populations of turtles (Fordham et al., 2007).
Most studies report rooting as the major cause of impact on plant communities. The spatial and temporal patterns of rooting depend on a number of factors that include availability of different food sources, climate conditions, habitat and soil type. Although most studies indicate that wild boar are associated with decreased plant biomass, conflicting evidence exists concerning the effect of rooting on plant species richness and composition. For instance in Sweden and in Malaysia, the number of plant species increased following wild boar rooting (Welander, 2000; Ickles et al., 2001). Similarly, in the Spanish Pyrenees alpine grasslands, wild boar rooting created large gaps that increased plant community heterogeneity and maintained high levels of plant diversity (Bueno et al., 2010). In California species richness decreased in the first year after disturbance but increased thereafter, exceeding the number of species found in undisturbed areas (Kotanen, 1995). In England, Sims et al., (2014) found that rooting by feral wild boar significantly reduced the percentage cover and density of bluebells (Hyacinthoides non-scripta) by up to 95 and 60 %, respectively. However, excluding wild boar enabled substantial recovery in percentage cover and the density of bluebells within 2 years.
Several studies found that wild boar and feral pigs promote seed dispersal either by ingesting fruits with seeds that are then found intact in faeces or by transporting seeds through the coat and hooves. For instance in Australia and in Hawaii, feral pigs act as vectors for the dispersal of seeds of non-native invasive plant species: when the fruits are eaten the seeds are not digested and appear viable in faeces (Grice, 1996; Lynes & Campbell, 2000). In Germany Heinken et al., (2006) found that soil seed banks near trees used by wild boar as rubbing posts are richer in plant species than control trees and suggest wild boar has a role in the dispersal of forest plant species.
Contrasting results exist on the consequences of rooting on soil processes and cycling of minerals, with some studies suggesting that rooting has a significant impact on these processes, whilst others finding little evidence of impact (reviewed in Barrios-Garcia & Ballar in 2012). For instance, no effect of rooting on soil pH, organic matter, nitrogen content and regeneration of broadleaved and conifer species was observed in the Netherlands, with the exception of oak (Quercus spp.) and beech (Fagus sylvestris) regeneration that was negatively correlated with wild boar rooting (Groot Bruinderink & Hazebroek, 1996). Other studies in the US (Singer et al., 1984; Siemann et al., 2009) found that feral pig rooting accelerated nitrogen mineralization rates and lowered the carbon:nitrogen ratio. In Switzerland, rooting by wild boars resulted in significant increases in mineral soil C and N concentrations and microbial biomass, which could lead to improved growth conditions for plants. However, total plant cover and sapling counts were reduced on rooted plots, possibly due to mechanical disturbance or due to reduced nitrogen: the study concluded that simple characterizations of wild boar rooting as beneficial or detrimental to forest ecosystems should be avoided (Wirthner et al., 2012).
In summary, the environmental impact of wild boar depends on a large number of factors which include whether the species is native to the area, local wild boar density, vulnerability of plant and animal species, population management and availability of alternative food resources such as crops and supplementary feeding. When wild boar or feral pigs appear in a new area or, as in the case of Scotland, in an area where they had not been present for several centuries, all these factors should be carefully considered to evaluate the consequences of the presence of this species on the environment.
Worldwide population trends of wild boar and feral pigs
In the last few decades wild boar and feral pig populations have increased dramatically worldwide. For instance, in the US the number of states where wild pigs were recorded increased from 17 in 1982 to 40 in 2012 (The Wildlife Society, 2012; USDA 2013 in Christie et al., 2014). In parallel, a number of states recorded a significant increase in the spread of wild pigs: for instance the number of counties where this species occurred in California increased from 9 counties in the mid ‘60s to 31 counties in 1983-1985, 36 counties in 1992-1994 and 47 counties in 2006-2007 (Sweitzer & McCann, 2007, in Christie et al., 2014). In many states, such as Mississippi, Alabama, Georgia, Florida, Oklahoma, Texas, Tennessee and South Carolina wild pigs are now present in > 90% of the counties (The Wildlife Society, 2012). In the United States as well as in Australia, feral pig expansion was attributed to deliberate releases from hunters, range expansion as population numbers increased, escapes from farms, milder winters, and increased forage availability associated with agricultural development (Waithman et al., 1999; Hutton et al., 2006; USDA, 2013).
In Europe, many native wild boar populations had gone extinct or existed at very low densities at the beginning of the 20th century (Apollonio et al., 2010). A review on wild boar population trends, based on hunting statistics, found that a simultaneous sharp increase of wild boar numbers throughout Europe between the ‘60s and the mid ‘70s was followed by an apparent stabilization of numbers in the following decade (Saez-Royuela & Tellería, 1986). Wild boar increases in numbers were attributed to a combination of the species’ life-history traits such as high reproductive output and dispersal potential and to other changes that included lack of large predators, reforestation, deliberate releases for sport hunting, supplementary feeding, habitat alteration and mild winters that improved survival (e.g. Genov, 1981; Erkinaro et al., 1982; Geisser & Reyer, 2005; Gethöffer et al., 2007; Jerina et al., 2014).
Three decades later, many of these factors are still operating and current trends of landscape development indicate that human-wild boar conflicts such as crop damage and vehicle collisions are increasing (e.g. Apollonio et al., 2010; Massei et al., 2011). Trends in numbers and distribution of wild boar in Europe are exemplified by data from France: in 1983-1986 the species occurred in 17% of the regional departments, and only 6% of the departments shot > 3000 animals per year; in 2003-2006 wild boar presence had spread to 85% departments, with 56% of these shooting > 3000 animals per year (Pfaff & Saint Andrieux, 2007). In parallel, the apparent decline of hunter numbers observed in several western countries (e.g. Riley et al., 2003) suggest that the relative importance of hunting, as the main cause of wild boar mortality, is decreasing.
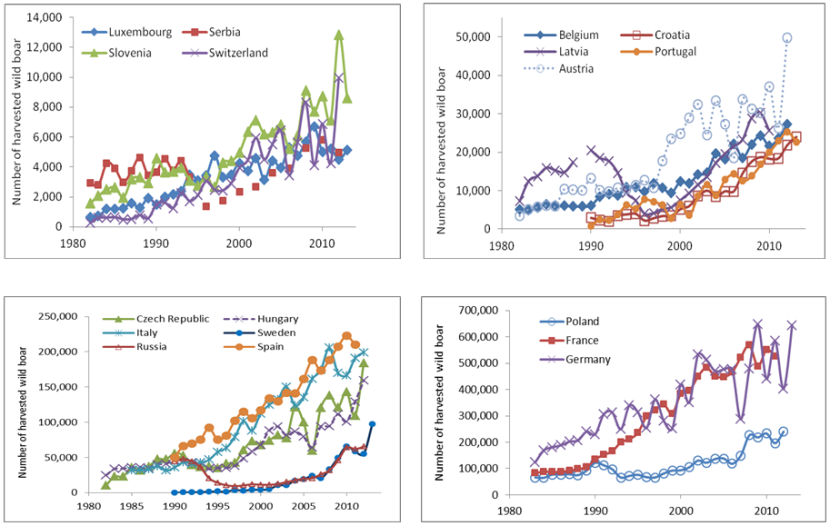
Figure 1.1. Wild boar hunting bags in European countries.
Click for a full description
4 line graphs of wild boar hunting bags in European countries from 1980 to 2010.
Massei et al., (2014) analysed trends in wild boar hunting bags in 18 European countries as well as trends in numbers of hunters in the last three decades. The results of this study showed that wild boar numbers continued to grow throughout Europe between 1982 and 2013 (Figure 1.1).
In 2012, a minimum of 2.5 million wild boar were harvested across the 18 countries, against circa 864,000 harvested in 1992 (when for the first time data became available for all 18 countries). At present, countries such as Germany and France harvest 450,000-600,000 wild boar per year. The results also showed that in the last 30 years, the ratio between animals harvested in one year and animals harvested the previous year has consistently exceeded one. In the same period, the number of hunters has been comparatively stable or even declining in most European countries. Assuming the hunting bag statistics reflect the actual number of animals present, and that the number of hunters is a reasonable indicator for the mortality due to hunting, these findings suggest that recreational hunting per se is currently not enough to limit wild boar population growth and that the economic and environmental impact will continue to increase.
Distribution of feral pig populations in the UK
In the UK, native wild boar were hunted to extinction by 1300 AD (Yalden, 1999). Since the 1990s, free-living feral pig populations have become established in several parts of the UK as a result of escapes from farms and illegal introductions (Wilson, 2014). Genetic analysis has demonstrated that the ‘wild boar’ found in the Forest of Dean, in western England, are hybrids between domestic pigs and wild boar, the latter believed to have been imported from western European populations (Frantz et al., 2012; Wilson, 2014). In Ireland wild boar sightings in the wild were first officially recorded in April 2009 on the Invasive Species Ireland website. By October 2012 there had been 27 recorded sightings reported on the National Biodiversity Data Centre website. Microsatellite analysis revealed that almost all the Irish individuals belong to the domestic pig genetic cluster, with only three individuals being classified as hybrids between wild boar and domestic pig (McDevitt et al., 2013).
In England, the distribution of feral wild boar populations was monitored using 5 x 5 km grid squares. The results of the last two decades indicate that, on average, one to two escapes or release incidents occurred in England per year, each involving from one to more than 50 animals (Wilson, 2014) (Figure 1.2). The records are also consistent with wild boar having spread in the last decade. For instance, the number of 5 x 5 km squares where wild boar were recorded in Kent/East Sussex rose from 7 in 2002 to 10 in 2010; in parallel, this number rose from 4 to 9 in West Dorset and from 5 to 8 in Gloucestershire (Wilson, 2014). At least four separate breeding populations now occur in Kent/East Sussex, West Dorset, Forest of Dean (Gloucestershire) and Devon/Somerset, each comprising at least a few tens of feral wild boar with the exception of the Forest of Dean. In the Forest of Dean Gill (2014) estimated that there were between 325 and 885 animals wild boar in 2013 and between 506 and 1325 animals in 2014. Approximately 130 animals were culled in 2013-2014 (Gill, 2014).
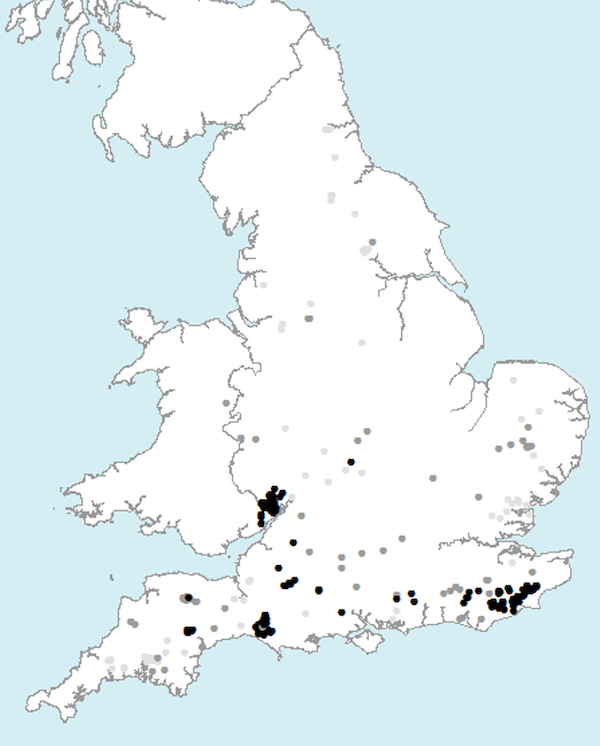
Figure 1.2. Distribution of reports of free-ranging wild boar in England from 1994 to 2014.
Click for a full description
Black dots indicate animals still present in 2014, pale dots show areas where animals are believed to be no longer present. Source: C. Wilson, 2014.
In Scotland, recorded sightings of feral pigs by Forest Enterprise Scotland staff are currently sent to Science Advice for Scottish Agriculture (SASA) in a standard format (B. Sewell, pers. comm.). As feral pigs do not naturally occur in Scotland, the species has a ‘formerly native’ status, and for the purposes of the Wildlife and Countryside Act 1981 (section 14) this means that feral pigs and their hybrids are considered not native (SNH, 2014). A report by SASA (Campbell & Hartley, 2010) suggested that in 2010 at least 1,300 wild boar or feral pigs were kept in farms across 11 sites.
Information provided in July 2014 (Campbell, personal communication) indicates that there are at least two populations of feral pigs breeding in Scotland (Figure 1.3). The first is located in Dumfries (originally two separate groups, one around New Abbey/Dalbeattie and one in Carsphairn Forest, about 40 miles from each other and likely now to be a single population). The second known breeding population is in Lochaber around Glen Dessary and Invergarry and possibly extends much further north and east. Three other populations have been recorded, and these animals may be breeding although this has not been confirmed. The first is in the area around Cawdor (SE of Inverness), and several escapes and reports have been recorded near Blairgowrie (N of Perth) and near Drumlean (Aberfoyle).
Many other sightings or reports of feral pigs shot have been collected across several parts of Scotland. The number of animals in each breeding population is estimated to be between 50 and 100 and the Lochaber population may exceed 100 (C. Lavin and S. Campbell, pers. comm.) but no formal density estimate exists. The Cawdor population is probably fewer animals and the number of animals in the Blairgowrie and Drumlean populations are likely to be fewer still.
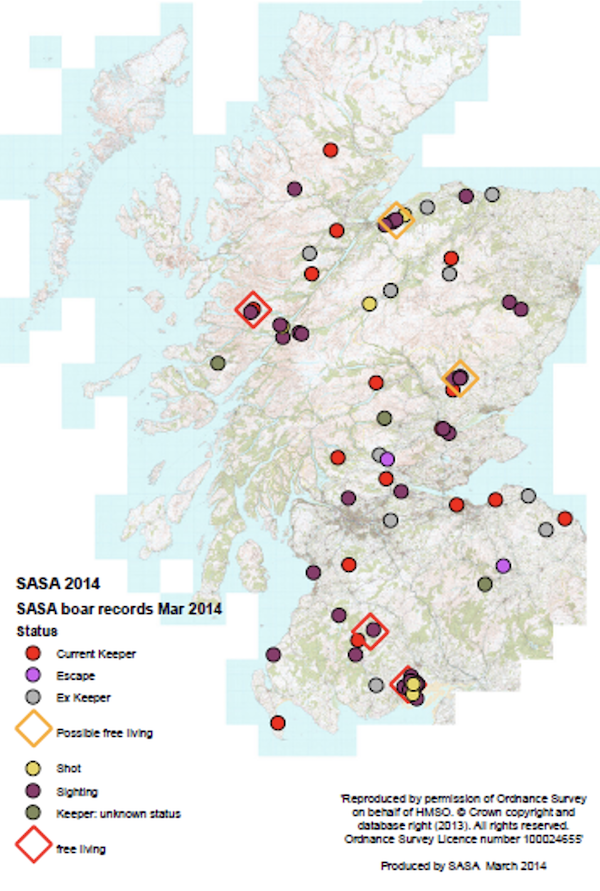
Figure 1.3. Distribution of reports of feral pigs in Scotland up to March 2014. Red and orange diamonds indicate free-living populations (confirmed and possible). Red dots (keeper) indicate current farms. Source: S. Campbell, 2014.
Shooting occurs in several populations of feral pigs in Scotland, with more than 50 animals harvested per year in the Lochaber population. In parallel, there is anecdotal evidence of illegal releases or escapes, direct evidence of supplementary feeding and indication that some landowners are reportedly instructing their stalkers not to shoot so as to encourage numbers to build.
Wild boar on farms fall under the same regulations that govern pig farms. Both Defra and the Scottish Government do not require that animals are marked whilst kept on the holding premises. However new rules for the identification and registration of pigs came into force on 1 October 2011. These changes were effected by European law (Council Directive 2008/71/EC) and intended to improve traceability and thus manage a disease outbreak more effectively. These new rules are implemented and enforced by the Pigs (Records, Identification and Movement) (Scotland) Order 2011. Under these rules, farmers are required to apply unique identifiers such as ear tags, tattoos, slap mark or temporary paint marks to each animal when the animal is moved from the premises. Pigs that are not moved do not require identifiers. As a consequence, the source of feral pigs that escape from farms or that are intentionally released into the wild cannot be identified.
In summary, the only information on the presence of feral pigs in Scotland appears to be the distribution map based on data collated ad hoc by SASA. The main gaps in knowledge on the feral pigs in Scotland are information on numbers and distribution at a local scale, reproductive output such as number of females reproducing per year and litter size and mortality (including mortality due to hunting). In addition, very little information is available on the current number of farms and animals kept in captivity, on the methods farms use to prevent escapes, on the extent, type and timing of supplementary feeding and on the management of feral pig populations by estates that encourage sport hunting. Furthermore, current controls clearly do not prevent escapes, which cannot be traced back to the original farm or estate. These gaps in our knowledge could be addressed by taking a range of actions which are listed in Table 1.
Table 1. Actions that could help to address the gaps in our knowledge about the distribution of feral pigs in Scotland
| Knowledge gaps | Proposed actions |
|---|---|
| Population size unknown |
|
| Population dynamics and distribution at local level unknown |
|
| Local management on private estates and farms unknown |
|
Objective 2 - Key information required to inform a model that will describe feral pig population dynamics in Scotland and anticipate population responses to management
Objective 2 comprised the following sections:
- A review of information required to model feral pig population dynamics in Scotland and anticipate population responses to management. The review was based on analogous models developed to describe population trends of wild boar and feral pigs and population responses to management.
- A review of methods used to estimate feral pig population trends. Cost-effective methods for the reliable detection of feral pig presence and for estimating relative abundance of feral pigs are essential to evaluate the impact of population control options on numbers of feral pigs. This part, based on a study that APHA (formerly AHVLA) conducted for Defra, summarised these methods and described their advantages and disadvantages.
Review of information required to model feral pig population dynamics in Scotland and anticipate population responses to management
Population models focus on understanding the processes that influence population dynamics through births, deaths, immigration and emigration. These models suggest how demographic changes are influenced by external factors such as environmental fluctuations and by population management actions. Quantitative models of population dynamics can thus be used in wildlife management as general predictive tools for assessing a population’s risk of extinction or potential for growth. By employing different types of analyses (such as elasticity, sensitivity, and perturbation analyses) and simulating different contexts, modellers can assess the impact of population control methods, suggest which variables are most critical for prioritising data collection and ultimately optimising allocation of resources.
The fundamental parameters affecting population dynamics are natality, mortality, immigration and emigration. These parameters are in turn influenced by individual factors such as age, gender, body mass and by external factors such as climate, availability of natural resources and supplementary food, diseases, predation and competition as well as management. A number of these factors have been investigated for populations of wild boar and feral pigs and are summarised in Table 2.
As feral pig populations in Scotland are still isolated, the impact of immigration and emigration on population dynamics can be regarded as negligible; thus the review focussed only on mortality and natality. Data on wild boar reproduction in individual populations can be investigated through the following methods:
- number of corpora lutea, embryos or foetuses per female and proportion of pregnant or lactating females per year;
- number of piglets/litter per female and proportion of females with piglets
- ratio between number of piglets and number of adult females.
The presence of corpora lutea in an ovary indicates that the animal is sexually mature and cycling; the number of corpora lutea corresponds to the number of eggs ovulated (Gethöffer et al., 2007). This variable, which is usually obtained from harvested animals, is particularly important when establishing the proportion of females under one year of age that are sexually mature.
The number of foetuses or embryos provides an indication of pre-natal litter size. This variable, which is usually obtained from harvested animals, can be used to assess the maximum reproductive potential of a feral pig population. Due to pre-natal mortality, the actual litter size is generally lower than the number of foetuses or embryos.
Due to infant mortality, particularly in the early months of life, litter size also varies between birth and weaning (3-4 months of age) and between weaning and one year of age (Toigo et al., 2008). The resulting ratio between number of piglets and number of adult females throughout the year can be used to calculate juvenile mortality and recruitment to the population.
| Aim of the study | Method | Results | References |
|---|---|---|---|
| Determine the impact of different environmental conditions on litter size, % of females reproducing and survival rate of wild boar | Model based on data provided in the literature | In good environmental conditions 50% of females of all age classes produce 4.5-6.8 piglets/litter, annual survival rates is 0.52-0.71% and population growth is 1.63 In poor environmental conditions 30-90% of females produce 3.5-6.3 piglets/litter, annual survival rates is 0.25- 0.58% and population growth is 0.85 | Bieber & Ruf, 2005 |
| Assess how the intensity of hunting affects wild boar population growth rate | Model based on field data collected on wild boar populations under different levels of hunting pressure | High hunting pressure results in a higher proportion of females reproducing and in larger litter size | Servanty et al., 2009 |
| Determine the impact of food availability and climate on wild boar reproductive output | Model based on long-term field data collected on wild boar reproduction, climate and mast production | Females must reach a body mass of 27-33 kg before breeding. The proportion of females in oestrus in November (0 -90%) varied in relation to food resources and climate | Servanty et al., 2011 |
| Assess impact of natural mortality and hunting mortality on wild boar population dynamics | Model based on survival rate of marked wild boar of all ages estimated by Capture-Mark-Recapture | Overall annual mortality is > 50% for all sex and age classes, mostly due to hunting Natural mortality rate is similar for males and females (~ 15%) Natural female mortality varies annually independently of mast production | Toigo et al., 2008 |
| Predict effect of geographic region, hunting and mast years on wild boar population growth | Model based on field data collected on wild boar populations throughout Europe | Hunting pressure and production of mast but not geographic region affect wild boar population growth | Holland et al., 2009 |
The population growth rate results from the combination of natality (i.e. the proportion of females giving birth multiplied by average litter size) and mortality.
Data on mortality in wild boar population can be obtained by:
- Marking new-born animals (for instance with ear tags) or animals of known age and assessing the survival of these animals throughout the years by counting the numbers found dead or culled;
- Hunting statistics.
Age in wild boar can be determined from patterns of tooth eruption and replacement up to 24-36 months (Matschke, 1967; Genov et al., 1992). Hunting statistics, ideally presented as number of wild boar culled by sex and age, can be used to calculate relative mortality due to hunting (compared to natural mortality and mortality due to vehicle collisions) and age-class specific survival (e.g. Okarma et al., 1995; Servanty et al., 2011, reviewed in Bieber & Ruf, 2005).
Applying ear tags (Massei et al., 1997; Toigo et al., 2008) is relatively simple and allows managers to assess survival and dispersal as well as to apply methods such as capture-mark-recapture to estimate population size.
It is important to stress that for wild boar both litter size and proportion of females reproducing may change dramatically, within a population, between years (Table 2). For instance, in a Mediterranean area, 90% of females produced on average 4.5 piglets/ litter following a large mast production; one year later only 20% of the females had piglets, with a mean litter size of 2.4 (Massei et al., 1996). Other studies highlighted how the proportion of females giving birth can be extremely high if supplementary feeding or crops are available: this was the case in Estonia where 95% of 1-2 year old females and 86% of older females were found pregnant with an average of 4.8 and 5.2 embryos respectively (Veeroja & Mannil, 2014).
In summary two sets of data are required as the minimum number of parameters to model the population dynamics of feral pig populations in Scotland: natality and mortality, ideally age-specific. In addition, data on diet (collected through analysis of faeces or stomach contents), availability of food resources such as crops, mast and supplementary feeding as well as data on climate (precipitations and monthly temperatures) can also be collected as these factors may affect feral pig population dynamics. Multi-year data series can thus be used to evaluate the variation of population parameters and to predict local population trends. Based on these data, the responses of feral pig populations to different management options can be assessed.
Review of methods to estimate wild pig population dynamics and presence
Estimating local feral pig abundance is crucial to quantify the effect of interventions aimed at mitigating human-feral pig conflicts. In addition, detecting presence of feral pigs in new areas is important to monitor the distribution and spread of this species. The following information summarises the results of a study funded by Defra to develop surveillance methods to monitor density and range expansion of wild boar in woodlands in England.
Wild boar presence
Four methods were developed to detect wild boar presence in an area and to monitor the species’ range expansion: (i) large-scale mapping of wild boar sightings, (ii) bait stations with camera traps, (iii) camera grids and activity signs on transects and (iv) use of putative attractants. In addition, modelling was used to determine the effort required to detect feral pigs at low density.
Large-scale mapping of sightings were provided by Natural England and obtained by collating ad hoc reports from the public, other agencies and the media, including sightings, reports of damage or rooting activity and reports of illegal releases and escapes. The location of each report was recorded to the nearest 1km UK Ordnance Survey national grid square and data were presented as number of 5 x 5 km squares where wild boar presence had been recorded. This method was found useful to monitor changes in wild boar distribution through the decades and to record the long-term expansion of the species at national level.
The method of single bait stations with motion-activated camera traps (Reconyx HC Hyperfire 500, Reconyx Inc. 3828 Creekside Lane Holmen, WI, US www.reconyx.com) was developed as a tool to confirm the presence of wild boar in an area. A single bait station and two camera traps were placed in small woodlands (50-150 ha each): to maximise the likelihood of detecting wild boar presence, bait stations were placed either on sites that were most likely to be visited by wild boar (such as mature oak or chestnut woods) or where wild boar had been sighted previously. The bait used was maize (circa 7-8 kg per bait station), replaced after one week. To avoid bait consumption by non-target species, maize was placed in pipes with holes that wild boar could shake to obtain food. When placing camera traps in each wood, 1-2 hours were spent walking on tracks in the wood and recording ad hoc wild boar activity signs (rooting or trails). At the end of week 1 and week 2 the presence of wild boar was confirmed where camera traps had taken at least one picture of this species.
The method of camera grids and activity signs on transects was developed to determine the minimum effort, in terms of number of transects and camera traps to be employed to detect the presence of wild boar in a new area. Camera grids were also used to calculate the density of wild boar in an area following the procedure developed by Rowcliffe et al., (2008) (see next section).
In each wood, forest rides and tracks (hereafter referred to as forest tracks) were mapped using Ordnance SurveyTM MastermapTM data series and ArcMap 9.3 GIS software (ESRI, California). On the tracks, 200 m x 1m transects were located to obtain a density of 10 transects/100 hectares. On each transect, the number of wild boar trails and rooting areas were recorded. As previous research had shown that trails and rooting are more difficult to detect in summer than in winter, this method was applied only in winter.
In parallel, motion-activated camera traps were placed in each woodland on a grid and at a density of 16/100 hectares. The number of camera nights was defined as the number of camera traps multiplied by the number of nights each camera was left in the woodland. As a minimum of 250 camera nights per site, based on 20 or more camera traps per site is recommended by the literature on density estimation using camera traps (Rovero & Marshall, 2009), 30-47 evenly distributed camera traps were placed in each of the five woodlands surveyed for the study and left in situ for nine nights. Monitoring was carried out in winter, in parallel to the activity signs survey. If the number of camera traps available was not sufficient to survey a woodland, camera traps were initially positioned in the northern part of the woodland, left 9 days and then moved to the central part and to the southern part of the woodland to complete a survey in 18-27 days. As fully randomized placement could result in cameras being positioned in areas of no visibility, cameras were positioned in areas of relatively higher visibility within 25 m of the grid points. The number of wild boar visits per camera per 9 days was then calculated for each wood. One visit was defined as one or more photos of wild boar until there was a lapse of at least 10 minutes between consecutive photos: photos of wild boar taken more than 10 minutes apart were counted as a new independent visit as preliminary observations with ear-tagged animals indicated the same group rarely return to the same area within 10 minutes. Bootstrapping (Efron, 2000) was used to derive the probability of detecting wild boar presence in a wood and hence the survey effort (in number of transects or camera traps) required to detect wild boar presence in woods with different densities of wild boar. Bootstrapping was carried out by randomly selecting a set number of transects/camera traps (from the original transects/cameras data set for each wood) and assessing whether wild boar had been detected in those. This process was reiterated 10,000 times and the probability of detection of wild boar was derived as the proportion of those iterations where wild boar had been detected.
The results suggested that the number of transects/100 ha required to detect wild boar presence with 90% or more confidence varied between 1 and 7 transects/ 100 ha. However, in one out of ten surveys, the maximum number of transects surveyed per 100 ha (i.e. 10 transects/100 ha) resulted in an 88.6% probability of detecting wild boar. The minimum number of camera traps/100 ha required to detect wild boar presence with 90% or more confidence varied between 2 and 9 (Figure 2.1). If the wood that had the minimum density of wild boar recorded during the study was excluded, the minimum number of camera traps/100 ha required to detect wild boar presence with 90% or more confidence varied between 2 and 4 (Figure 2.1). However, the wood with the lower density suggests that nine traps (i.e. more traps) would be needed.
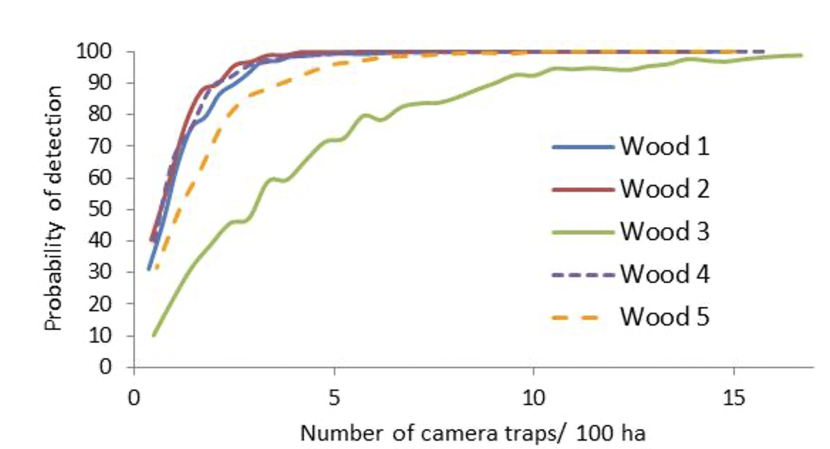
Figure 2.1. Detectability functions for wild boar derived from pictures recorded by camera traps against the number of camera traps used in each wood during a winter survey.
Click for a full description
A line graph showing the probability of detection (y-axis) against the number of camera traps per 100 ha (x-axis) for 5 different woods (numbered 1 to 5).
The consistency of wild boar detection by relatively small numbers of camera traps, compared to the more variable results obtained with activity signs on transects suggested that camera traps should be used instead of activity signs to detect wild boar presence in new areas.
A putative site attractant, based on birch wood tar, was found effective in changing boar behaviour: a pilot trial with the compound applied to stakes confirmed that wild boar spent more time rubbing against treated than non-treated stakes. Wild boar were observed sniffing the compound and rubbing against the stake.
When the attractant was applied to trees, this behavioural change was supported: wild boar left signs on trees treated with this compound more often than control trees. The number of activity signs such as rooting around the tree, tusk marks, hair and mud left on the tree was significantly higher on treated than on control trees. This effect of the compound persisted for at least 4 weeks without re-treating the trees. These results suggested that this compound could be used to improve the probability of detecting the presence of wild boar in woodland and might be tested to improve the efficiency of trapping, although the latter would need to be confirmed by further study. We suggest that bait stations with camera traps, and the birch tar compound offer options for confirming the presence of wild boar in an area.
We modelled the effort required to detect a single wild boar in a large wood (55 km2): the results indicated that approximately 20 camera traps per 100 ha should be deployed to have a 90% probability of detecting a single wild boar.
Wild boar density
Four methods were proposed to assess wild boar population trends or estimate density: (i) Passive Activity Index (PAI) based on camera trap grids and activity signs on transects, (ii) density estimates based on camera trap grids, (iii) distance sampling through thermal imaging and (iv) monitoring of Road Traffic Accidents (RTA). In addition, a simulation of density assessed through camera traps and distance sampling was carried out to assess the precision and accuracy of these methods.
A density index for feral pig abundance, called “PAI” (Passive Activity Index) was calculated using data collected through camera trap grids and activity signs surveys described in the previous section. Mixed models were used to compare the activity index between years within each wood and to derive an activity index for each wood. The mixed models were fitted using the restricted maximum likelihood and had year as a fixed effect. The proportion of transects with rooting signs and with trails, and the number of trails per transect was calculated for each wood and each of the two years when this survey was carried out.
The predicted number of boar trails or signs of rooting xij for year i and transect j was calculated as follows:
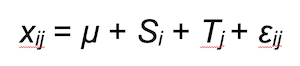
where Si is a fixed effect for year, Tj is a random effect for transects and εij accounts for the residual variability.
Following Engeman et al., (2001, 2002 and 2013) a Passive Activity Index (PAI) for each winter i was then derived as:
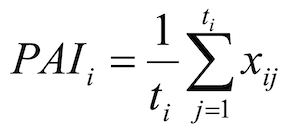
where ti is the number of transects within year i.
This was done for each wood separately. Bootstrapping (Efron, 2000) was used to estimate the uncertainty associated with the activity estimates by re-sampling with replication 10,000 times the data from boar trails or signs of rooting on transects at random and by estimating the corresponding wild boar index. Then, for each wood and each season, a mean activity index and a standard error were obtained from the bootstrapped data.
The same method was applied to the data obtained from camera grids, as follows:
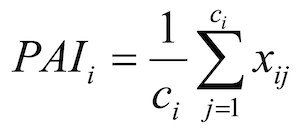
where ci is the number of cameras within winter i and xij is the predicted number of visits for camera trap j in winter i and can be written as:
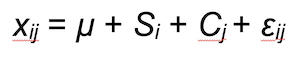
where Si is a fixed effect for winter and Cj is a random effect for cameras and εij accounts for the residual variability.
The results of the application of these methods indicated that PAIs based on camera traps or on activity signs on transect are a reasonably quick, inexpensive method to generate a feral pig population density index. This index can be used to compare trends between years or relative densities between sites.
The main limitation of using trails as an index of population trends is that several animals can use the same trail, thus resulting in an underestimate of number of individual trails. On the other hand, the area covered by rooting activity can be extremely variable in size (ranging from about 50 cm2 to many tens of square metres) and rooting per se is strongly dependent on food availability. The overall conclusion is that both wild boar trails and signs of rooting could be used as the simplest way to monitor wild boar population trends by calculating the proportion of transects where rooting or trails were recorded.
Density estimates based on camera trap grids were obtained following the method developed by Rowcliffe et al., (2008). A density estimator D was calculated for each wood and each survey separately, based on the number of wild boar visits recorded by camera traps per 9 days as follows:
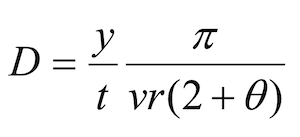
where y/t = number of visits y per unit time t
r and θ= radius and angle of the camera’s detection area, expressed in radians
v = speed of movements.
D was then multiplied by group size (estimated independently of the camera trapping surveys) to obtain the number of wild boar per km2 in each woodland (Rowcliffe et al., 2008). Independent estimates of group size were obtained by using bait stations and camera traps. To minimize potential double counting, individual groups or animals were identified by a number of features including ratio of females to piglets and physical traits such as body size and coat colour. The speed of movements, obtained from wild boar (n=7) equipped with GPS collars (Quy et al., 2014) was (v) = 0.274 (SD 0.052) km/hr. Group size in each year was 2.50 and 3.74. Bootstrapping (Efron, 2000) was used to estimate the uncertainty associated with the density estimates by re-sampling with replication the camera trap data 10,000 times at random and by estimating the corresponding wild boar density. Then, for each wood and each season, a mean density and standard error were obtained from the bootstrapped data. Bootstrapping was also used to obtain the 95% confidence intervals around density estimates and to determine how increasing the number of camera traps reduced the variation around the estimated mean.
The relationship between the number of camera traps employed to estimate wild boar densities and the 95% confidence intervals around each estimated density suggested that, in most instances, between 6 and 15 camera traps/100 ha would be sufficient to calculate wild boar density. The results indicated that increasing the sampling effort, in terms of number of camera traps, would only marginally increase the precision of the density estimate, at least for wild boar densities similar to those recorded in this study. For field applications, a minimum number of 15 camera traps/100 ha, left in place for 9-10 days, is thus recommended to assess density of wild boar with reasonable precision in small woodlands (approximately 150 ha). For populations that exist at very low density (such as recently established populations) or for woods smaller than 150 hectares, we suggest that at least 20 camera traps, left in place for 13-14 days are used, as recommended by Rovero & Marshall (2009).
Distance sampling through thermal imaging was employed to assess wild boar density in each wood (Buckland et al., 2001). This method is based on observations of the distribution of animals in relation to transects. Observations of feral pigs were made from a vehicle and on foot on paths and roads that traversed the study site. A thermal imager was used to detect warm-bodied animals at night which typically have surface temperatures several degrees above ambient. When a group of animals was found, species, group size, distance from the observer and angle between the group and the transect were recorded.
The perpendicular distances between the transect and each group detected were used to estimate the detection function, by fitting a curve to the frequency distribution of detection distances using the ‘Distance’ software. Mean density (D) is then estimated from the following equation:
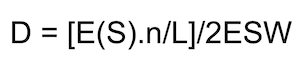
where E(S) is the mean group size; n/L is the number of groups encountered per unit transect length and ESW is the effective strip width, the definite integral of the detection function between the transect (0) and (w), the maximum perpendicular distance of animals detected.
To obtain density estimates using distance sampling, it is recommended that 50+ observations (of groups, not individuals) are used to fit a detection function (Buckland et al., 2001). As detectability of ungulates varies with vegetation density, fields and forest were sampled separately and detection functions and densities estimated for each in turn.
The results suggested that where feral pigs are present at a relatively low density, as it is possibly the case for Scotland, the number of animals calculated through this method may have a very large coefficient of variation, i.e. the estimate is not very precise. Even for higher densities the method may produce fairly large confidence intervals: for instance, in the Forest of Dean, a mean density of 8.7 wild boar/km2 (95% Confidence Interval = 5.3- 14.4) was estimated through distance sampling in March 2013 and a mean density of 12.3 wild boar (with 95% Confidence Interval = 7.6-19.9) was estimated with the same method in March 2014 (Gill, 2014).
The monitoring of RTAs was carried out by collating information collected by local rangers and Forestry Commission staff and by comparing these numbers, used as a proxy for changes in population size, and traffic flow. Data on the latter were obtained from Local Authorities and expressed as number of vehicles per day recorded at several locations within the study site. The results showed that the number of RTAs increased and that traffic flow did not change. We concluded that the trend in the number of RTAs, if changes in traffic flow can be discounted, has potential as a quick index to monitor feral boar population trends. This approach has been used in several European countries such as Sweden (Liberg et al., 2010), the Netherlands (Van Wieren & Groot-Bruinderink, 2010), Switzerland (Imesh-Bebie’ et al., 2010), and Spain (Rosell et al., 2013).
The simulation model of density assessed through camera traps and distance sampling found that both methods may underestimate the true density of wild boar by 18-30%. The model suggested that the camera trap estimates were relatively precise and not affected by population size although they were highly sensitive to group size. Density estimates based on distance sampling had wider confidence intervals but accuracy was unaffected by group size or population size. The agreement between these findings and the results on densities obtained by using camera traps and distance sampling provides reassurance about the reliability of estimates produced by these two methods, although the relationship between estimated and actual densities could only be confirmed with populations of known densities. If eradication of a feral pig population was planned and implemented, using the above methods to estimate density prior to eradication would provide an opportunity to determine the accuracy of these estimates. Camera trap surveys have the advantage that they can be used throughout the year whilst surveys based on distance sampling should be conducted only in winter, when there is minimum vegetation cover and thus maximum visibility of feral pigs.
Based on the above results, we provided a decision tree that stakeholders could use when assessing the presence of wild boar in an area (Figure 2.2).
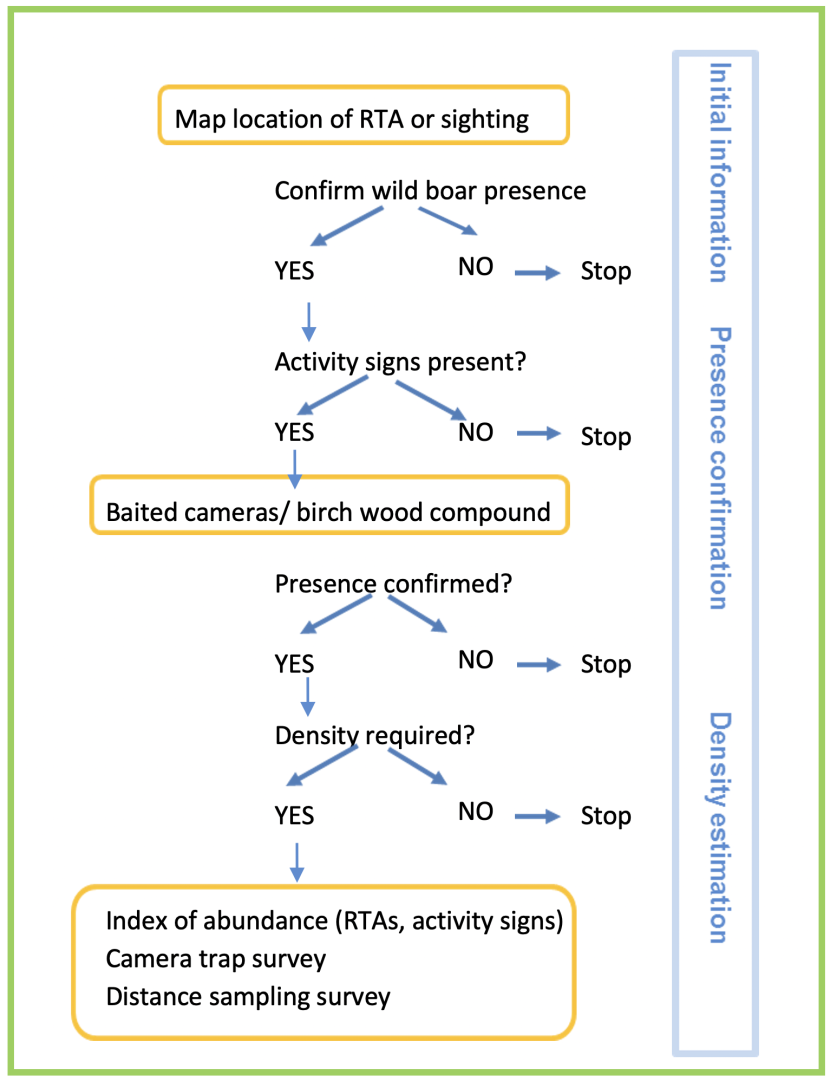
Figure 2.2. Decision tree of methods to monitor presence and local abundance of wild boar.
Click for a full description
The decision tree assumes that an RTA or a sighting of feral pig occurred in an area where the species had not been previously recorded.
Objective 3 - Risk assessment of feral pigs in Scotland
GB non-native organism risk assessment scheme
Name of organism: Feral pig (Sus scrofa)
Author: Giovanna Massei
Risk Assessment Area: Scotland
Draft: 10 September 2014
| Stage 1. Organism Information | RESPONSE | COMMENT |
|---|---|---|
| 1. Identify the organism. Is it clearly a single taxonomic entity and can it be adequately distinguished from other entities of the same rank? | Feral pig | Pigs, feral pigs and wild boar belong to the same species Sus scrofa. In areas where the species is not native, such as the USA or Australia, these animals are often referred to as feral or wild pigs, hogs or swine (Engeman et al., 2001; Choquenot et al., 1996). Genetic analysis has been often used to determine the degree of inbreeding between wild boar and domestic pig. Animals farmed in England are believed to have been imported from western European wild boar populations (Wilson, 2014), although some of these have wild boar/domestic pig ancestry (Frantz et al., 2012). Throughout the Risk Assessment the species will be referred to as “wild boar” or “feral pigs” where appropriate. |
| 2. If not a single taxonomic entity, can it be redefined? (if necessary use the response box to re-define the organism and carry on) | Partially | Defra’s Action Plan (2008) refers to animals escaped from farms in the UK as “feral wild boar”. The Action Plan sets out the Government’s position that free roaming wild boar are feral wild animals: as such they do not belong to anyone, and the responsibility for managing feral boar rests with the relevant landowner or land manager. Further information on legislation relating to feral wild boar can be found on the Deer Initiative’s Best Practice Guide ‘Wild Boar Legislation’. The SG and NatureScot refer to this species as “feral pigs”. As a former native species, the wild boar is not included on the list of non-native species listed on the GB Non-Native Species Information Portal. In Scotland it is considered to be a Non-Native Species under the Wildlife and Countryside Act 1981 (Scottish version as amended by the Wildlife and Natural Environment (Scotland) Act 2011 because former natives are considered to be outwith their native range. According to section 14P (3) and also 3.8 and 3.12 of the Code of Practice on Non-Native Species “animals and plants that were once native in a location but have become extinct are considered to be “former natives”. For the purposes of the 1981 Act former natives are considered to be outwith their native range and it is therefore an offence to release a former native without a licence”. |
| 3. Does a relevant earlier risk assessment exist? (give details of any previous risk assessment) | Yes | The first qualitative risk assessment (RA), commissioned by Defra (Goulding et al., 1998) highlighted that there were two breeding populations of feral boar in England, one in Kent and East Sussex and one in Dorset. Based on an initial population of 100 animals, modelling suggested that the number of boar would increase to 169 in 5 years, with a minimum of 108 animals and a maximum of 326. A 15- year projection gave an average population size of 485. The RA also mentioned that wild boar could be associated with different types of impact, ranging from crop damage to animal health and public safety. A subsequent RA, covering England and Wales was commissioned by Defra as part of the action plan for feral wild boar in England (2008). The RA was mainly focussed on likelihood and impacts of transmission of diseases between freer-ranging wild boar, humans and livestock in England. The RA concluded that the risk of exotic disease incursion was low; the impact should these incursions occur was medium and the likelihood of zoonotic diseases was low. The document also pointed out that population increase would affect these conclusions and modify the disease risk. The RA on the environmental and economic impact concluded that at moderate densities, environmental impact was likely to be minor, whilst economic impacts, such as agricultural damage, were likely to become significant in the longer term particularly if the populations spread and increased substantially. |
| 4. If there is an earlier risk assessment is it still entirely valid, or only partly valid? | Partially | Some conclusions of the risk assessment published as part of the 2008 Defra’s Action Plan were based on much smaller numbers of feral wild boar and on very conservative predictions on population growth. It is possible these conclusions will be revised as in the last few years at least one wild boar population (in the Forest of Dean) has increased to circa 800 animals and other breeding populations have appeared in Scotland. |
| 5. Where is the organism native? | Eurasia | The natural range of wild boar extends from Western Europe and the Mediterranean basin to Eastern Russia, Japan and Southeast Asia (Sjarmidi & Gerard, 1988). In Europe, wild boar recently recolonized Sweden, Finland, Estonia and Norway (Erkinaro et al., 1982; Rosvold & Andersen, 2008; Veeroja & Männil, 2014). In Denmark the species has become re-established following farm escapes (Andersen & Holthe, 2010). |
| 6. What is the global distribution of the organism (excluding Great Britain)? | Eurasia, USA, Australia, New Zealand, several countries in South America and Africa | Outside their native range, wild boar and feral pigs have been introduced to the US, South America, Australia, New Zealand and in many African countries where the species now occurs as wild boar, feral pig or as a mixture of wild boar and feral pig (Choquentot et al., 1996; Mayer & Brisbin, 2009). |
| 7. What is the distribution of the organism in Great Britain? | Isolated, breeding populations occur in England, Wales, Scotland and N. Ireland | In the UK, wild boar are native but they were hunted to extinction by 1300 AD (Yalden, 1999). Since the 1990s, free-living feral pig populations have become established in several areas of the UK as a result of escapes from farms and of illegal introductions (Goulding et al., 2003; Hartley, 2010; Wilson, 2005; Wilson, 2014). Using a suite of molecular markers, Frantz et al., (2012) demonstrated that the ‘wild boar’ found in the Forest of Dean, in western England, were a genetic mixture of wild boar and domestic pigs. In Ireland wild boar sightings in the wild were first officially recorded in April 2009 on the Invasive Species Ireland. By October 2012 there had been 27 recorded sightings of the species in Ireland (Figure S1; National Biodiversity Data Centre. Microsatellite analysis revealed that almost all the Irish individuals belonged to the domestic pig genetic cluster and only a few individuals were classified as hybrids between wild boar and domestic pig (McDevitt et al., 2013). In Scotland, Campbell & Hartley (2010) suggested that at least 1,300 wild boar were kept in farms across 11 sites. Information provided in July 2014 (Campbell, personal communication) indicates that there are at least two populations of feral pigs breeding in Scotland, one in Dumfries (originally two separate groups, one around New Abbey/Dalbeattie and one in Carsphairn Forest, about 40 miles from each other and likely now to be a single population) and another in Lochaber around Glen Dessary and Invergarry. Two other free-living populations have been recorded as possibly self-sustaining, one around Cawdor (SE of Inverness), and another near Blairgowrie (N of Perth). Many other sightings or reports of feral pigs shot have been collected across many parts of Scotland. The number of animals in each breeding population is estimated to be between 50 and 100 and the Lochaber population may exceed 100 (C. Lavin and S. Campbell, pers. comm.) but no formal density estimate exists. |
| 8. Is the organism known to be invasive (i.e. to threaten organisms, habitats or ecosystems) anywhere in the world? | Yes | The IUCN (World Conservation Union) lists this species among the 100 worst alien invasives. The impact of feral pigs on conservation and economic interests includes spread of diseases to wildlife, livestock and people, vehicle collisions, damage to crops and amenities, predation on native species, reduction in plant and animal abundance and richness (e.g. Welander, 2000; Hone, 2002; Schley & Roper, 2003; Massei & Genov, 2004; Bueno et al., 2010; Barrios-Garcia & Ballari, 2012). |
| Stage 2. Screening Questions | - | - |
| 9. Has this risk assessment been requested by the GB Programme Board? (If uncertain check with the Non-native Species Secretariat) | Risk assessment requested by SNH If yes, go to section B (detailed assessment) | - |
SECTION B – Detailed assessment
PROBABILITY OF ENTRY
| QUESTION | RESPONSE | CONFIDENCE | COMMENT |
|---|---|---|---|
| 1.1. How many active pathways are relevant to the potential entry of this organism? | few | high | Active pathways relevant to entry of feral pigs in Scotland are 1. Escapes from farms and 2. Illegal releases. These pathways have also caused the re-establishment of wild boar in several European countries where the species had become extinct in the last century (Sáez-Royuela & Tellería, 1986; Pfaff & Saint Andrieux, 2007; Apollonio et al., 2010). The same pathways are mentioned for the US where the number of states reporting the presence of feral pigs rose from 23 in 1988, to 39 in 2004 (Hutton et al., 2006; Centner & Shuman, 2014). |
| 1.2. List relevant pathways through which the organism could enter. Where possible give detail about the specific origins and end points of the pathways. | Escapes from farms and illegal releases. | high | Several papers and reports on feral pigs in GB (e.g. (Goulding et al., 2003; Defra, 2008; Hartley, 2010; Wilson, 2003; Wilson, 2005; McDevitt et al., 2013; Wilson, 2014) as well as anecdotal evidence from rangers, hunters and groups operating in areas where feral pigs occur suggest the presence of this species in Scotland can be assigned to both pathways, although it is often difficult to establish whether the escape from farms is accidental or provoked. |
| Pathway name: | Escapes from farms | - | - |
| 1.3. Is entry along this pathway intentional (e.g. the organism is imported for trade) or accidental (the organism is a contaminant of imported goods)? | Mostly accidental | high | In Scotland, farmed wild boar can be kept in captivity under licence from Local Authority under the Dangerous Wild Animals Act 1976 (Modification) Scotland Order 2008. The Act specifies conditions in the licence to ensure the animals are confined in a way that prevents their escape. The requirement for a DWA licence applies to all Suidae, including farmed wild boar and hybrids where at least one parent is wild boar (SNH 2014). Feral pigs in Scotland are likely to have mixed wild boar and domestic pig ancestry and they are regarded as Non-Native Species under section 14C of the Wildlife and Countryside Act 1981. In Scotland, there is a presumption of ‘no-release’ for any species ‘outwith their native range’. Section 14 of the Wildlife and Countryside 1981 Act makes it an offence to release an animal, allow it to escape, or cause it to be outwith the control of any person, at a place outwith its native range. Feral pigs are domesticated animals and are therefore outwith their native range. For the purposes of the 1981 Act former natives, like wild boar, are considered to be outwith their native range and the same offences, therefore, apply. |
| 1.4. How likely is it that large numbers of the organism will travel along this pathway from the point(s) of origin over the course of one year? | unlikely | medium | By collating reports of escapes or releases of feral pigs and by assessing the presence of animals in the wild, Wilson (2014) concluded that between 1989 and 2009 an average of 1-2 escape/release per year occurred in England, involving from one to 140 individuals. Similar reports, in terms of a few animals sighted in the wild as a result of farm escapes or illegal releases, exist for Scotland (Campbell & Hartley, 2010). |
| 1.5. How likely is the organism to survive during passage along the pathway (excluding management practices that would kill the organism)? | very likely | very high | Feral pigs escaped from farms are likely to find food and shelter in the vicinity of the farm or in neighbouring woodlands where they can survive and reproduce. |
| 1.6. How likely is the organism to survive existing management practices during passage along the pathway? | likely | high | The only significant factors affecting the mortality of feral pigs are hunting and vehicle collisions. Hunting aimed at eradicating pigs from an area might prevent these animals from establishing in that area, particularly if only a few pigs have recently been introduced. Vehicle collisions are more likely to occur following recent animal introductions into an area (Massei et al., 2010). |
| 1.7. How likely is the organism to enter Scotland undetected? | NA | NA | - |
| 1.8. How likely is the organism to arrive during the months of the year most appropriate for establishment? | NA | NA | - |
| 1.9. How likely is the organism to be able to transfer from the pathway to a suitable habitat or host? | likely | high | Wilson (2014) suggested that feral pigs are likely to escape from farms and establish breeding populations in the wild. |
| 1.10. Estimate the overall likelihood of entry into Scotland based on this pathway? | very likely | very high | In Scotland, accidental escapes may occur from farms as well as from estates that offer wild boar hunting and that have poor animal containment facilities. Reports in England and Scotland (Campbell & Hartley, 2010; Wilson, 2014) suggest that in the last decades animals have regularly escaped from farms. |
| Pathway name: | Illegal releases | - | - |
1.3. Is entry along this pathway intentional (e.g. the organism is imported for trade) or accidental (the organism is a contaminant of imported goods)? If intentional, only answer questions 1.4, 1.9, 1.10, 1.11 | Intentional | high | Wilson (2014) suggested that between 1989 and 2009 an average of 1-2 escape/release per year occurred in England. Similar reports, in terms of a few animals sighted in the wild as a result of farm escapes or illegal releases, exist for Scotland (Campbell & Hartley, 2010). Although in many instances it is not possible to separate farm escapes from illegal releases, this pathway of entry is very common in other countries (e.g. Choquenot et al., 1996, for Australia, Centner & Shuman, 2014 for the US). |
| 1.4. How likely is it that large numbers of the organism will travel along this pathway from the point(s) of origin over the course of one year? | unlikely | medium | By collating reports of escapes or releases of feral pigs and by assessing the presence of animals in the wild, Wilson (2014) concluded that between 1989 and 2009 an average of 1-2 escape/release per year occurred in England, involving from one to 140 individuals. Similar reports, in terms of a few animals sighted in the wild as a result of farm escapes or illegal releases, exist for Scotland (Campbell & Hartley, 2010). |
| 1.5. How likely is the organism to survive during passage along the pathway (excluding management practices that would kill the organism)? | NA | NA | - |
| 1.6. How likely is the organism to survive existing management practices during passage along the pathway? | NA | NA | - |
| 1.7. How likely is the organism to enter Scotland undetected? | NA | NA | - |
| 1.8. How likely is the organism to arrive during the months of the year most appropriate for establishment? | NA | NA | - |
| 1.9. How likely is the organism to be able to transfer from the pathway to a suitable habitat or host? | likely | high | Several studies (e.g. Choquenot et al., 1996; Campbell & Hartley, 2010; Centner & Shuman, 2014; Wilson 2014) suggested that feral pigs are likely to be illegally released and establish breeding populations in the wild. Wild boar and wild pigs can adapt to a very wide spectrum of environmental conditions, which include woodlands, marshlands, agri-environment and suburban areas. Provided that high-energy food (such as acorns, crops, animal food), shelter and water for wallowing are available, feral pigs can live virtually everywhere (Schley & Roper, 2003; Massei & Genov, 2004; Rosvold & Andersen, 2008; Barrios-Garcia & Ballari 2012). |
| 1.10. Estimate the overall likelihood of entry into Scotland based on this pathway? | very likely | very high | Experiences in the US and in continental Europe suggest that illegal releases of feral pigs are likely to increase in parallel with hunters’ interest for a new game species (Centner & Shuman, 2014). In Scotland, releases might also occur as the result of lobby groups that advocate the return of wild boar as a former native species. Population modelling suggested that in Scotland a release of as few as five animals would be sufficient to establish a viable population of feral pigs (Leaper et al., 1999). Experience of escape/release incidents and the establishment of feral populations in England tend to confirm the predictions of these models (Wilson, 2014). |
| End of pathway assessment, repeat as necessary. | - | - | - |
| 1.11. Estimate the overall likelihood of entry into Scotland based on all pathways (comment on the key issues that lead to this conclusion). | very likely | very high | Illegal releases and escape from farms are likely to continue. If the number of free-living populations increases it is likely that the number of illegal releases and escape from farms will increase. In Scotland, holding feral pigs in captivity without adequate containment is likely to lead to further escapes. Farmers are required by the Council Directive 2008/71/EC to identify pigs with an ear tag, tattoo or slap mark before animals are moved between holdings but not whilst pigs are held on the farm. The lack of permanent identification means it is more difficult to attribute escaped animals to a particular farm. Another practice that is growing in Scotland is recreational hunting of feral pigs, encouraged by private estates or through hunting clubs. This practice might lead to an increase of feral pigs in farms and to an increase of illegal releases, as already occurred in the US and in several other continental European countries such as Sweden and The Netherlands (Van Wieren & Groot-Bruinderink, 2010; Liberg et al., 2010; USDA-APHIS, 2013). |
PROBABILITY OF ESTABLISHMENT
Important instructions:
For organisms which are already well established in Scotland, only completed questions 1.15 and 1.21 as recommended by the Non-native Species Secretariat.
| QUESTION | RESPONSE | CONFIDENCE | COMMENT |
|---|---|---|---|
| 1.12. How likely is it that the organism will be able to establish in Scotland based on the similarity between climatic conditions in Scotland and the organism’s current distribution? | very likely | very high | Free-living, isolated populations of feral pigs already occur in Scotland (Campbell & Hartley, 2010). The widespread range of wild boar and feral pigs through different habitat types, latitudes and climatic regions shows that new free-living populations of this species can easily establish in Scotland. Wild boar and wild pigs can adapt to a very wide spectrum of environmental conditions, which include woodlands, marshlands, agri-environment and suburban areas. Provided that high-energy food (such as acorns, crops, animal food), shelter and water for wallowing are available, feral pigs can live virtually everywhere (Schley & Roper, 2003; Massei & Genov, 2004; Rosvold & Andersen, 2008; Barrios-Garcia & Ballari, 2012). |
| 1.13. How likely is it that the organism will be able to establish in Scotland based on the similarity between other abiotic conditions in Scotland and the organism’s current distribution? | very likely | very high | Two main factors limit the establishment of wild boar in an area: food availability and presence of snow cover for several consecutive weeks (Jędrzejewski et al., 1992; Jędrzejewska et al., 1997). Availability of high-energy food is crucial for this species as wild boar are unable to digest plants as efficiently as ruminants. Snow cover prevents wild boar from rooting for food and hampers their movements. However, wild boar populations are increasing in countries characterised by harsher winters than Scotland. Examples include Sweden, where widely spread supplementary feeding, used by hunters to increase local densities of wild boar and reduce crop damage, have increased the spread of this species (Thurfjell et al., 2013). In Norway, wild boar that in the past were limited to the broad leaved deciduous forests have recently settled in areas with harsher climate, thanks to availability of crops and supplementary feeding that have allowed this species to find food throughout the year (Rosvold & Andersen, 2008). In Germany, wild boar originally restricted to large, deciduous forests, now occur also in poorer habitats like spruce forests and in areas characterised by snowy winters (Wotschikowsky, 2010). |
| 1.14. How likely is it that the organism will become established in protected conditions (in which the environment is artificially maintained, such as wildlife parks, glasshouses, aquaculture facilities, terraria, zoological gardens) in Scotland? | NA | NA | - |
| 1.15. How widespread are habitats or species necessary for the survival, development and multiplication of the organism in Scotland? | moderately widespread | high | Wild boar and wild pigs can adapt to a very wide spectrum of environmental conditions. In Scotland, increased reforestation, particularly where focussed on deciduous trees, is likely to increase the habitat available for breeding populations of feral pigs. |
| 1.16. If the organism requires another species for critical stages in its life cycle then how likely is the organism to become associated with such species in Scotland? | NA | NA | - |
| 1.17. How likely is it that establishment will occur despite competition from existing species in Scotland? | very likely | very high | Wild pigs are omnivores that feed opportunistically on wild fruits (acorns, beech mast, berries, etc.), roots and tubers, vertebrates (including carcasses), invertebrates (including earthworms), eggs and crops (Schley & Roper, 2003; Massei & Genov, 2004). As the species’ reproductive potential is higher than that of any other ungulate or medium- or large-size mammal occurring in Scotland, it is likely that feral pigs will outcompete any species with similar feeding habits. These include deer and ungulates feeding on beach mast and acorns as well as other generalists such as badgers. |
| 1.18. How likely is it that establishment will occur despite predators, parasites or pathogens already present in Scotland? | very likely | very high | In Europe, the only significant predator of wild boar is the wolf (Canis lupus) (Jędrzejewski et al., 1992). This predator is not found in the wild in Scotland. Diseases and pathogens, such as for instance Classical Swine Fever or African Swine Fever could significantly affect mortality of feral pig populations (Defra 2008, Defra 2014) but they are unlikely to prevent establishment and also unlikely to be affecting simultaneously several isolated populations. |
| 1.19. How likely is the organism to establish despite existing management practices in Scotland? | very likely | very high | Feral pigs are already established in some parts of Scotland. In Europe, hunting makes the greatest contribution to boar mortality (Nores et al., 2008; Toigo et al., 2008; Keuling et al., 2013). Thus culling and trapping to achieve eradication are the only management options that may prevent establishment of new populations in Scotland. In a few instances, animals escaped from farms in Scotland and in England have been shot, thus preventing the establishment of a population in an area (Campbell & Hartley, 2010; Wilson, 2014). In the US, feral pig populations were prevented from becoming established only in those states that reacted swiftly to the presence of this species by making recreational hunting for pigs illegal, by promoting electronic identification of animals kept in captivity or by precluding import, transport or release of this animals (Centner & Shuman, 2014). |
| 1.20. How likely are management practices in Scotland to facilitate establishment? | not likely | medium | At present, there are no management practices in Scotland that would facilitate establishment of feral pigs. |
| 1.21. How likely is it that biological properties of the organism would allow it to survive eradication campaigns in Scotland? | unlikely | high | Successful eradication campaigns of isolated feral pig populations have occurred in the US (Cruz et al., 2005; McCann & Garcelon, 2008; Massei et al., 2011). As populations or feral pigs are still isolated in Scotland, the eradication of these populations should be possible through a combination of culling and trapping. Wild boar and feral pigs respond to intense hunting pressure by becoming more nocturnal and in some instances by moving away from the area for distances up to several tens of km (Andrzejewski & Jezierski, 1978; Bouldoire & Vassant, 1989). The species’ high reproductive rate also means that eradication campaigns should be carried out in the shortest possible time, ideally in less than a year, to avoid the replacement of animals through recruitment. |
| 1.22. How likely are the biological characteristics of the organism to facilitate its establishment? | very high | high | Among ungulates, wild boar are characterised by the highest annual reproductive rate that may exceed 200% (Bieber & Ruf, 2005; Fonseca et al., 2011; Keuling et al., 2013). Females can reach sexual maturity in their first year of age (Gethoffer et al., 2007) and produce one litter of 3-7 piglets, and occasionally two litters per year. The number of litters, the litter size and the proportion of females that reproduce in a year are strongly affected by the availability of energy-rich food such as acorns (e.g. Groot-Bruinderink et al., 1994; Massei et al., 1996; Nahlik & Sandor, 2003, Fonseca et al., 2004). |
| 1.23. How likely is the capacity to spread of the organism to facilitate its establishment? | very high | high | Wild boar can move over large distances. In the UK, the maximum distance tracked from site of capture was 20 km (Moore, 2004). Radio-tracking studies found that in a single night, boars can travel between 2 and 15 km and that the direct distance between resting places varies between 0 and 7 km (Spitz, 1986). During seasonal migrations wild boar can move over longer distances, from 12 km to 250 km (Andrzejewski & Jezierski, 1978; Singer et al., 1981). A French study showed that between 18 and 55% of wild boar trapped and ear-tagged in six areas where hunting occurred had been shot by hunters at more than 10 km from the initial trapping site (Cargnelutti et al., 1992). |
| 1.24. How likely is the adaptability of the organism to facilitate its establishment? | very high | high | Wild boar and wild pigs can adapt to a very wide spectrum of environmental conditions, which include woodlands, marshlands, agri-environment and suburban areas. Provided that high-energy food (such as acorns, crops, animal food), shelter and water for wallowing are available, feral pigs can live virtually everywhere (Schley & Roper, 2003; Massei & Genov, 2004; Rosvold & Andersen, 2008; Barrios-Garcia & Ballari, 2012). |
| 1.25. How likely is it that the organism could establish despite low genetic diversity in the founder population? | very high | high | Several reports (Campbell & Hartley, 2010; Wilson, 2014) suggested feral pigs current populations derived from relatively few individuals. It is thus very likely that this species will establish irrespective of potentially low genetic variability. |
| 1.26. Based on the history of invasion by this organism elsewhere in the world, how likely is to establish in Scotland? (If possible, specify the instances in the comments box.) | very high | high | Feral pigs are already established in Scotland as escapes from farms or illegal releases. |
1.27. If the organism does not establish, then how likely is it that transient populations will continue to occur? Subnote: Red-eared Terrapin, a species which cannot re-produce in GB but is established because of continual release, is an example of a transient species. | N/A | N/A | - |
| 1.28. Estimate the overall likelihood of establishment (mention any key issues in the comment box). | very high | high | Feral pigs are already established in Scotland. Based on the history of invasion of this species in other countries (e.g. Erkinaro et al., 1982; Andersen & Holthe, 2010; Centner & Shuman, 2014), establishment of new populations in Scotland is very likely. |
PROBABILITY OF SPREAD
Important notes:
Spread is defined as the expansion of the geographical distribution of a pest within an area.
| QUESTION | RESPONSE | CONFIDENCE | COMMENT |
|---|---|---|---|
| 2.1. How important is the expected spread of this organism in Scotland by natural means? (Please list and comment on the mechanisms for natural spread.) | moderate | medium | The spread of this organism in Scotland by natural means is expected to be moderate if hunting pressure is high and as long as small populations exist in relatively localised areas. Where local populations are present in confined areas of preferred habitat, dispersal may be slow until high densities occur. For instance, in the South of England, free-living populations of feral pigs subjected to relatively high hunting pressure have existed for about 20 years and experienced moderate levels of increase in range and spread (Wilson, 2014). |
| 2.2. How important is the expected spread of this organism in Scotland by human assistance? (Please list and comment on the mechanisms for human-assisted spread.) | major | high | Deliberate releases or increase of local populations for sport hunting, and supplementary feeding provided by hunters are likely to increase the establishment and spread of feral pigs in Scotland. All these factors have played a significant role in the spread of wild boar and feral pigs in mainland Europe and in the US (Saez-Royuela & Telleria, 1986; Centner & Shuman, 2014; Massei et al., 2014). |
| 2.3. Within Scotland, how difficult would it be to contain the organism? | moderate | medium | As feral pig populations are still isolated, eradication or containment through sustained culling and trapping is still achievable. If populations expand, the cost of eradication is likely to increase dramatically, as shown in island eradications carried out in the US and summarized by McCann & Garcelon (2008). In the UK, an example of dramatic population growth is found in the Forest of Dean (Gloucestershire) where in early 2013 a comprehensive thermal imaging survey for feral wild boar carried out on the public forest estate indicated that 535 feral pigs occurred in the area. A follow-up survey in early 2014, estimated that 819 feral pigs occurred in the forest, despite a cull of around 130 animals between 2013 and 2014, indicating a significant growth of the number of pigs in one year (Forest Research, 2014). For geographically isolated populations, McCann & Garcelon (2008) suggested that an intensive eradication program should be preferred to sustained control as only a high intensity program can achieve eradication in a short period. In addition, the high cost of a short but intensive eradication program is likely to be less than that of sustained control over a period of several years and a short, well-managed program is less likely to be exposed to factors that can undermine its success. These factors include reproduction that causes the pig population to increase, pigs learning to avoid control, public opposition, legal challenges arising in the course of the project, increased lack of staff motivation, and funders’ fatigue which may result in lack of sustained funding to complete the program (Morrison et al., 2007; Parkes et al., 2010). |
| 2.4. Based on the answers to questions on the potential for establishment and spread in Scotland, define the area endangered by the organism. | Throughout Scotland, mixed woodlands, pastures adjacent to woodlands (including conifer plantations) and/or crops | medium | The area endangered by feral pigs will depend, to an extent, on the density and distribution range of animals. The latter in turn will depend on whether local populations are supplemented with food, whether crops and natural food are available throughout the year to sustain high densities of feral pigs and whether hunting has a significant impact on population growth (Massei & Genov, 2004; Bieber & Ruf, 2005; Barrios-Garcia & Ballari, 2012). In Europe, increasing numbers of wild boar sightings were reported in urban and suburban areas, for instance in Berlin, Barcelona, Vilnius and Budapest (e.g. Náhlik in Massei et al., 2014; Cahill et al., 2003; Jansen et al., 2007). It is also possible feral pigs in Scotland will colonise these areas. |
| 2.5. What proportion (%) of the area/habitat suitable for establishment (i.e. those parts of Scotland were the species could establish), if any, has already been colonised by the organism? | 0-10 | medium | Likely to be less than 1%. |
| 2.6. What proportion (%) of the area/habitat suitable for establishment, if any, do you expect to have been invaded by the organism five years from now (including any current presence)? | 0-10 | low | It is very difficult, at present, to answer this point. The answer is based on the population trends observed in the Forest of Dean in England, where feral boar occupy an area of about 50 km2. In the last four years this population increased from 200-250 feral boar estimated in 2010 to 500 in 2013 and up to 819 animals in 2014 but their spread to surrounding areas was not as conspicuous as their numerical increase. For other populations in mainland Europe, the spread per decade, in numbers and range, was more pronounced: for instance, in France in 1983-1986, 85% of the regional departments shot 1500-3000 wild boar, and only 15% of the departments shot > 3000 animals per year; in 1993-1996 45% of the departments shot 1500-3000 wild boar, and 55% shot > 3000 animals per year; in 2003-2006 14% of the departments shot 1500-3000 wild boar, and the remaining 86% shot > 3000 animals per year (Pfaff & Saint Andrieux,2007). In Sweden, the natural spread of wild boar since the species recolomised the country in the late ‘80s was calculated at 3-4 km/year but the expansion was speeded up by new illegal releases (Liberg et al., 2010). |
| 2.7. What other timeframe (in years) would be appropriate to estimate any significant further spread of the organism in Scotland? (Please comment on why this timeframe is chosen.) | 10 | medium | If a rapid increase in spread occurs, such as those recorded by Wilson (2014) in England, significant change could take place undetected for several years and this could limit the choice of management responses. |
| 2.8. In this timeframe what proportion (%) of the endangered area/habitat (including any currently occupied areas/habitats) is likely to have been invaded by this organism? | 10-20% | low | Please see considerations at point 2.6. The confidence of this statement is low as spread is likely to depend on factors such as type of habitat where feral pigs already occur and surrounding habitat, population size and population management (including level of hunting pressure), new releases etc. that are difficult to quantify or predict with available information. |
| 2.9. Estimate the overall potential for future spread for this organism in Scotland (using the comment box to indicate any key issues). | moderate | medium | Based on the information above, and on the fact that much of the woodland and agricultural land, as well as urban and suburban areas are suitable for establishment, feral pigs are likely to spread. |
PROBABILITY OF IMPACT
| QUESTION | RESPONSE | CONFIDENCE | COMMENTS |
|---|---|---|---|
| 2.10. How great is the economic loss caused by the organism within its existing geographic range excluding Scotland, including the cost of any current management? | major | very high | Wild boar and feral pigs cause major economic losses due to spread of diseases to livestock and people, vehicle collisions and damage to crops and amenities. As wild boar and pigs numbers increase, these losses grow (Massei et al., 2014). Examples include compensation for crop damage caused by wild boar in France rose from about €2.5 million in 1973 to €21 million in 2005 and €32.5 million in 2008 (Guibert et al., 2008; Maillard et al., 2010). In Luxembourg, compensation for crop damage caused by wild boar increased from about €100,000 in 1971 to €900,000 in 2004 (Schley et al., 2008) and in Slovenia from €292,000 in 2005 to €575,000 in 2013 (ZGS, 2014, in Massei et al., 2014). The number of wild boar-vehicle collisions in Sweden rose from about 50 per year in the early 2000 to about 1000 in 2005 and over 4000 in 2012 (Liberg et al., 2010). In the Netherlands this number rose from 142 in 1995 to 320 in 2003 (Van Wieren & Groot-Bruinderink, 2010), and in Switzerland, in the same period, this number increased from 212 to 412 (Imesh-Bebie’ et al., 2010). In Germany, out of the 227,000 traffic accidents with deer and wild boar in 2005, 13700 involved wild boar (Wotschikowsky, 2010). In Catalonia (North-eastern Spain) the number of accidents involving animals increased by 41.6% between 2007 and 2011, with wild boar responsible for 85% of the accidents (Rosell et al., 2013). In the US feral pigs cause significant economic loss: in California pig damage to crops in 2006 was estimated at $ 1.2 million, whilst damage to natural areas in 2005-2007 was estimated at $ 11.3 million (Sweitzer 2007 in Christie et al., 2014). In 2013, the USDA Wildlife Services estimated $ 28 million damage caused by feral pigs to crops, rangeland and developed land in California (Christie et al., 2014). Throughout the US, feral pig damage to crops in 2002 was valued at $800 million per year (Pimentel et al., 2002). This estimate did not consider livestock predation, disease transmission, or environmental impact. In Australia, annual crop damage by feral swine exceeded >$100 million (Choquenot et al., 1996). PLEASE NOTE: the role of feral boar in disease transmission and its related economic loss is covered under point 2.24. |
| 2.11. How great is the economic cost of the organism currently in Scotland excluding management costs? | minor | medium | Anecdotal evidence suggests that in Scotland the economic cost of feral pigs is still small, due to relatively low local densities of animals (Campbell & Hartley, 2010). Although the economic cost of this species in Scotland has not been quantified, impact so far include isolated incidents of disturbance to grasslands and woodlands through rooting activity, damage to gardens, vehicle collisions and pigs feeding from pheasant feeders (Campbell & Hartley, 2010). This cost must be in the order of £10,000s. |
| 2.12. How great is the economic cost of the organism likely to be in the future in Scotland excluding management costs? | moderate | medium | If feral pigs increase in numbers and spread, following for example the trend observed in the Forest of Dean as well as in the US and mainland Europe, the economic cost will grow for Scotland. The growing population of feral boar in the Forest of Dean has led to significant impacts on the resident community and on visitors to the area. Impacts include rooting of amenity grasslands, woodlands and road side verges, perceived risks of attacks by walkers, attacks on dogs, road traffic accidents and several instances of horse riders thrown off their horse when scared by a feral pig encounter. Damage to grasslands in the villages includes play areas, caravan parks, sports pitches, golf courses and private gardens (Forest Research, 2014). In 2013, for the first time the number of road traffic accidents involving feral wild boar in the Forest of Dean has surpassed the number of deer-vehicle collisions (Forest Research, 2014). Social impact will also depend on perceived risks of feral pigs’ attacks on humans and on risk of pig attacks on dogs. In Scotland, there have been reports of recently escaped feral pigs visiting gardens and rooting on a golf course near Aberfoyle, although there are no estimates for the financial value of this damage. |
| 2.13. How great are the economic costs associated with managing this organism currently in Scotland? | minor
| high
| At present there are no reports on economic costs associated with the management of feral pigs in Scotland. |
| 2.14. How great are the economic costs associated with managing this organism likely to be in the future in Scotland? | moderate | medium | The cost associated with managing this species in Scotland will depend on the location, number, local densities and distribution range of feral pigs. The higher the number of feral pigs, local densities and species range, the higher the cost. In addition, the cost will depend on the efficacy of methods used to manage this species. Typically, both culling and trapping are relatively inexpensive (in terms of man hour per pig) when densities are high but become progressively more expensive when numbers of animals decline (Parkes et al., 2010). Although it is not possible to establish a cost with current information, this could be moderate (between £100k and £1m) if feral pigs increase and must be quickly contained. In case of a sudden disease outbreak such as Classical Swine Fever or Foot-and Mouth Disease, the cost could exceed £1m. |
| 2.15. How important is environmental harm caused by the organism within its existing geographic range excluding Scotland? | major | high | Wild boar are very opportunistic feeders that adapt their diet according to the season and to the relative availability of different food sources (Genov 1981, Massei et al., 1996; Schley & Roper, 2003; Baubet et al., 2003). The environmental impact can derive from direct consumption of native plants and animals and from disturbance to the ground that affects ground-dwelling vertebrates (Massei & Genov, 2004; Barrios-Garcia & Ballari, 2012). Wild boar feed on around 400 species of plants, animals and fungi in Western Europe (Schley & Roper, 2003). The bulk of the food consists of plant material such as mast, roots, tubers, wild fruits and crops. Energy-rich plant food, like acorns and beechmast or agricultural crops, is particularly important as wild boar are not ruminants like other ungulates (Andrzejewski & Jezierski, 1978; Groot Bruinderink et al., 1994; Schley & Roper, 2003). Most crops are consumed when available (Genov, 1981; Schley & Roper, 2003). Animals are regular components in the diet of the wild boar but occur in relatively small quantity compared to plants (Schley & Roper, 2003). Insects and earthworms are the most important; however most small animals including birds, mice and invertebrates as well as carcasses of larger animals and eggs of ground-nesting birds are consumed opportunistically (Genov, 1981; Schley & Roper, 2003; Barrios-Garcia & Ballari, 2012). In addition, wild boar may exploit mast hoards collected by small mammals and stored underground (Focardi et al., 2000). Contrasting results exist on the consequences of rooting on soil processes and cycling of minerals, with some studies suggesting wild boar and feral pigs have a significant impact on these processes, whilst other studies finding little evidence of impact (reviewed in Barrios-Garcia & Ballari, 2012). No effect of rooting on soil pH, organic matter, nitrogen content and regeneration of broadleaved and conifer species was observed in the Netherlands, with the exception of oak and beech regeneration that was negatively correlated with wild boar rooting (Groot Bruinderink & Hazebroek, 1996). |
| 2.16. How important is the impact of the organism on biodiversity (e.g. decline in native species, changes in native species communities, hybridisation) currently in Scotland? | minimal
| low
| At present there are no reports on the impact of feral pigs on biodiversity in Scotland. |
| 2.17. How important is the impact of the organism on biodiversity likely to be in the future in Scotland? | moderate | medium | Impact on biodiversity in Scotland will depend on feral pig local densities and location, proximity to fragile ecosystems that could be affected by extensive rooting, availability of food sources that might maintain high densities. As the species feed opportunistically, it is also possible that feral pigs will impact on eggs of ground-nesting birds such as grouse and other galliformes. |
| 2.18. How important is alteration of ecosystem function (e.g. habitat change, nutrient cycling, trophic interactions) caused by the organism currently in Scotland? | minimal | low | At present there are no reports on alteration of ecosystem function due to feral pigs in Scotland. |
| 2.19. How important is alteration of ecosystem function (e.g. habitat change, nutrient cycling, trophic interactions) caused by the organism likely to be in Scotland in the future? | moderate | medium | Wild boar are described as “ecosystem engineers” because their rooting behaviour has marked ecosystem-level effects. Rooting affects above and below ground components of plant and animal communities but also indirectly affects other organisms by physically changing habitat characteristics and modifying resource availability. For instance, wild boar rooting affects soil mineralisation, plant growth and abundance of invertebrates and ground-dwelling mammals (reviewed in Massei & Genov, 2004; Barrios-Garcia & Ballari, 2012). It is likely that alteration of ecosystems will occur in Scotland if the density of feral pigs is relatively high. |
| 2.20. How important is decline in conservation status (e.g. sites of nature conservation value, WFD classification) caused by the organism currently in Scotland? | minimal | medium | At present there are no reports on decline in conservation status caused by feral pigs in Scotland. |
| 2.21. How important is decline in conservation status (e.g. sites of nature conservation value, WFD classification) caused by the organism likely to be in the future in Scotland? | moderate | medium | It is possible that feral pigs will have an impact on areas of conservation value such as Sites of Special Scientific Interest and Natura 2000 sites. The decline in conservation status of these areas will depend on local wild boar density and on measures taken to mitigate this impact. |
| 2.22. How important is it that genetic traits of the organism could be carried to other species, modifying their genetic nature and making their economic, environmental or social effects more serious? | moderate | medium | If feral pigs come in contact with rare breeds of pigs held in outdoors farm, they are likely to interbreed, with potential loss of genetic traits in the rare breed. Accidental entry of wild boar in pig farms have occurred in England (Massei, pers. comm.). The most recent accident, recorded in January 2015 in England involved a wild boar that entered an outdoor farm and killed a Gloucester Old Spot pig |
2.23. How important is social, human health or other harm (not directly included in economic and environmental categories) caused by the organism within its existing geographic range?
| major
| high
| Wild boar and feral pigs have a significant impact on human economic interests and on conservation throughout their range. The main impacts are related to transmission of diseases, road traffic accidents, crop damage and effects on plant and animal communities. In addition, this species may cause damage to sites of archaeological importance: for instance in the US 42% of these sites containing artefacts in a protected area of around 40000 ha showed signs of rooting by feral pigs |
| 2.24. How important is the impact of the organism as food, a host, a symbiont or a vector for other damaging organisms (e.g. diseases)? | major | very high | Wild boar can act as reservoir or as vector for numerous diseases that can be transmitted to other wildlife species, livestock and people (e.g. Meng et al., 2009). A qualitative risk assessment of the role of the feral wild boar in the likelihood of incursion and on the impact of effective disease control of selected exotic diseases in England was carried out in 2010 (Hartley, 2010). The assessment concluded that the greatest risks of exotic disease incursion into the UK were associated with disease entering through the consumption of infected pork meat or meat products by either wild boar or domestic swine. The diseases identified as highest risk were classic swine fever (CSF, a virus), foot and mouth disease (FMD, a virus) and Trichinella sp (a parasitic roundworm). This risk assessment was based on relatively low densities of feral wild boar (200 animals in Kent/East Sussex and 30-50 boar in the Forest of Dean). The assessment acknowledged that if populations did increase or if an outbreak of disease was not contained and entered wild boar populations in England, there would be potential for a reservoir of disease to be established for CSF and FMD. The Defra (2008) Action Plan reports that a CSF outbreak in GB in 2000 cost more than £20 million. Outbreaks of notifiable diseases can result in the banning of export of pig products and disruption to the home pig industry. In 2008 an outbreak of Aujeszky’s disease, on a single farm cost at least £500K. If spread occurs this figure could possibly reach £1 million. The 2001 outbreak of FMD cost the UK over £8 billion. It is theoretically possible for wild boar to act as a reservoir for Bovine Tuberculosis (bTB, caused by a bacterium, Mycobacterium bovis) (Defra 2005). For instance, at the high densities experienced in southern Spain, epidemiological, pathological and microbiological evidence is consistent with wild boar perpetuating TB infection independently within their populations, and thus posing transmission risks to other species, including livestock (Naranjo et al. (2008)). However, evidence from other countries is conflicting (Machackova et al., 2003; Corner, 2006). In England, M. bovis was first isolated in free-living wild boar in 2010 (Foyle et al., 2010). While Scotland remains bTB free, there is considerable uncertainty about the role feral pigs might play in introducing or perpetuating the disease in Scotland. |
| 2.25. How important might other impacts not already covered by previous questions be resulting from introduction of the organism? (specify in the comment box) | N/A | N/A | - |
| 2.26. How important are the expected impacts of the organism despite any natural control by other organisms, such as predators, parasites or pathogens that may already be present in Scotland? | major | high | A recent review on wild boar population trends in Europe (Massei et al., 2014) showed that wild boar numbers are growing significantly. This trend was first reported in the 1980s (Saez-Royuela & Tellería, 1986). The review by Massei et al., (2014) suggested that natural mortality, such as predation, is not sufficient to reduce population growth and that predators have a very modest impact on wild boar mortality. Conversely, recreational hunting, which is the most important cause of mortality of wild boar throughout Europe, has declined. In localised areas, wild boar numbers can decrease due to diseases but in these instances human intervention usually ensures the disease is eradicated, particularly when the pathogen can be transmitted to livestock. At present there are no predators or pathogens in Scotland that could reduce the impact of feral pigs. |
| 2.27. Indicate any parts of Scotland where economic, environmental and social impacts are particularly likely to occur | Any area where feral pigs occur or are encouraged to increase | medium | The highest economic, environmental and social impacts will occur where feral pig populations exist in or near fragile ecosystems that could be affected by extensive rooting, in areas where wild boar could prey on ground-nesting birds or prevent plant regeneration. Social impact will also occur if feral pigs live in proximity of villages and towns or in areas with frequent visitors (such as camping sites, forests where pigs may encounter dog walkers, golf courses etc.). Disease outbreak will have major economic impact in areas where other wild ungulates and livestock occur. |
RISK SUMMARIES
| - | RESPONSE | CONFIDENCE | COMMENT |
|---|---|---|---|
| Summarise Entry | very likely | very high | Entry here is regarded as “additional entry” as free-living populations of feral pigs are already present in Scotland. It is very likely that illegal releases and escapes from farms will continue, thus increasing the spread of feral pigs across Scotland. |
| Summarise Establishment | very likely | very high | Feral pigs can adapt to live in many environments and are increasingly occurring in urban and suburban areas across Europe. Establishment could derive from a few founder animals. The high annual reproductive rate of feral pigs, which may exceed 200% is another important factor that promotes the fast growth of small populations. |
| Summarise Spread | moderate | medium | Spread will depend on hunting pressure (culling and trapping to eradicate), numbers of animals released, improvement of controls and implementation of containment and tagging of farmed pigs. |
| Summarise Impact | major | high | The environmental, social and economic impact of feral pigs ranges from the ecological impact on plant and animal communities, on soil and cycling of nutrients through to disease transmission, crop damage, road traffic accidents and damage to properties, infrastructures and attacks on dogs and potentially some livestock. |
| Conclusion of the risk assessment | high | high | The main conclusions are based on trends in impact and population numbers of feral pigs and wild boar in Europe, USA and Australia. The feral pigs already present in Scotland are likely to increase in numbers and spread. In parallel, their economic, social and environmental impact will grow unless measures are taken to control entry, establishment and spread of this species in the wild. |
ADDITIONAL QUESTIONS - CLIMATE CHANGE
| - | RESPONSE | CONFIDENCE | COMMENT |
|---|---|---|---|
| 3.1. What aspects of climate change, if any, are most likely to affect the risk assessment for this organism? | Milder winters | medium | Milder winters will decrease winter mortality due to lack of food. More food available will increase breeding success and decrease the mortality of juveniles thus increasing recruitment of new animals in a population. |
| 3.2. What is the likely timeframe for such changes? | 5-10 years | medium | Changes are likely to occur and be detected within a 5 to 10 year timeframe. |
| 3.3. What aspects of the risk assessment are most likely to change as a result of climate change? | Probability of establishment and impact | medium | Higher breeding success and local densities as well as increased probability of establishment might increase local impact of feral pigs. |
ADDITIONAL QUESTIONS - RESEARCH
| - | RESPONSE | CONFIDENCE | COMMENT |
|---|---|---|---|
| 4.1. If there is any research that would significantly strengthen confidence in the risk assessment please summarise this here. | yes | high | To strengthen the conclusions of the RA, particularly in areas where the confidence in the assessment is medium or low, research should focus on the following areas:
The research recommended above would give stakeholders a sound basis for managing the risks derived from the presence of feral pigs in Scotland. To manage these risks, research should focus on the following areas:
|
Objective 4 - Review of practical options for maintaining, limiting, or eradicating feral pigs and estimate of the effort required to deliver these options in Scotland
Objective 4 comprised the following sections:
- An overview of lethal and non-lethal methods used for maintaining, limiting, or eradicating feral pigs.
- A simulation model to quantify the likely amount of effort required to control and/or eradicate discrete feral pig populations in Scotland, under a variety of starting abundance scenarios.
- Conclusions.
An overview of lethal and non-lethal methods used for maintaining, limiting, or eradicating feral pigs
This section was based on an updated version of the review conducted by Massei et al., (2011) that summarized the advantages and disadvantages of lethal and nonlethal methods to control populations and impact of feral pigs. For each method, advantages and disadvantages should be regarded as relative to those of other control options.
Lethal methods
Trapping and killing
Wild boar and feral pigs are relatively easy to trap provided the availability of natural food is low. Trapping can remove large numbers of animals in a short time and traps are widely used to control this species (Table 3). Many types of trap designs are available, ranging from those that capture single animals or small groups to corral traps for larger groups (e.g. Saunders et al., 1993; Caley, 1994; Choquenot et al., 1996; Sweitzer et al., 1997; West et al., 2009). The majority of traps are made of mesh frames with drop gates and side-hinges or top-hinged spring-gates that boar must push to gain access to the food placed inside the trap. Corrals have similar gates but are larger and may have a funnelled entrance to guide animals toward box traps that are used to remove the pigs. A special type of corral, the BoarBuster™ (http://www.noble.org/ag/wildlife/boarbuster/), is a suspended corral trap that can be observed and dropped remotely from anywhere provided Internet service is available. The automated trap has been designed to send text or email messages upon motion activation and streams live video through a designated Web server. This trap technology allows the user to observe and activate the traps via smartphone or computer. As the corral is suspended, animals may enter or leave from all directions, eliminating trap-wary behaviour associated with conventional trap gates. The user-activated trigger eliminates the risk of non-target animals being captured. The corral design allows for captured pigs to be loaded out through an integrated door.
Drop-nets are also used to capture groups of feral pigs or boar: originally employed with an operator that would release the net once a group of boar was under the net (Jullien et al., 1987), recent models are remotely controlled nets suspended over bait used to attract animals to the area. Gaskamp et al., (2012) compared the effectiveness and efficiency of an 18.3 x 18.3 m drop-net and a traditional corral trap for trapping feral pigs in Oklahoma. The results showed captures of single groups of up to 27 and 15 pigs with drop-nets and corral traps, respectively, corresponding to 86 and 49% of the pigs removed using drop-nets and corral traps, respectively. Catch per unit effort was 1.9 and 2.3 hours per pig for drop-nets and corral traps, respectively. Feral hogs did not appear to exhibit trap shyness around drop-nets, which often allowed the managers to capture entire sounders in a single drop. Use of drop-nets also eliminated capture of non-target species.
The food most frequently used as bait to attract feral pigs to traps is maize, fermented wheat or other crops but also other baits such as various vegetables, fruit, blood, fish, animal parts, or carcasses are employed (Choquenot et al., 1996; Cruz et al., 2005; Twigg et al.,2005). However, in the UK, the Animal By-Products Regulations (Environment Agency 2008) prevents feeding pigs with food of animal origin, thus other food attractive to feral pigs, such as maize or peanuts should be used.
Traps need to be checked at least once per day, so that animals are dispatched and non-target species released. Large traps that allow the whole social group to be captured are likely to have little impact on social behaviour. The latter is particularly important as social perturbation may lead to increased contact rates, with the potential risk of increasing disease transmission and may encourage long-distance movements, thus, extending the impact to adjacent areas (Sodeikat & Pohlmeyer ,2002).
The maintenance and checking of traps can be expensive in staff time and can be applied over only relatively small areas. However, traps can be moved and redeployed to other areas, and trapping can be fitted around other routine control activities. Traps are difficult to transport and use on high slopes or rough terrain; conversely, they can be easily deployed to remove animals from residential areas. Compared to poisoning, traps have the advantage that the number of animals captured is known and carcasses can be safely removed. However, traps can be easily damaged by people opposed to culling.
Trapping, often combined with shooting, has been employed in many feral pig eradication projects (e.g. McIlroy, 1983; McCann & Garcelon, 2008). Trapping success depends on a variety of factors, including topography, time of year, type of trap used, number and density of traps deployed, trap location, number of nights each trap is used, type of bait used, duration of pre-feeding before the traps are set and the possibility that animals become trap-wary (Hone 1983; Choquenot et al., 1996; West et al., 2009; Parkes et al., 2010).
All these factors must be evaluated on a case-by-case context and will certainly need to be considered for Scotland if trapping is selected as one of the options to control feral pigs.
Snares
Snares have been used extensively in several parts of the world to kill feral pigs (e.g. Anderson & Stone, 1993; Kessler, 2002). Snares are sometimes used in conjunction with other lethal methods of feral pig control and often to mitigate local human-pig conflicts rather than to control populations (Table 3). The use of snares is regulated in many parts of the world and Scotland has the most detailed and comprehensive legislation on snaring in the UK. Under the Wildlife and Countryside Act 1981, as amended by the Wildlife and Natural Environment (Scotland) Act 2011, it is illegal to set in position or otherwise use any self-locking snare (or a snare of any other type specified in an order made by the Scottish Ministers) or any snare likely to cause bodily injury to any wild animal included in Schedule 6, such as badger, red squirrel and hedgehog. Under the Deer (Scotland) Act 1996 it is an offence to use any trap or snare for the purpose of taking or killing any deer. In Scotland, snaring for the taking of feral pigs may be feasible but it would have to be highly regulated and is not recommended because there are uncertainties about its impacts on animal welfare and potential bodily injuries to both target and non-target species.
Shooting
Aerial shooting from helicopters is relatively common in countries such as the United States and Australia, which have vast, uninhabited areas of sparse vegetation where it is relatively easy to locate groups of animals. This method can achieve quick decreases in feral pig abundance over large areas. As aerial shooting of wild boar is not carried out in Europe, where ground shooting is the main method used by hunters to control wild boar numbers, the review will focus on ground shooting.
Throughout Europe recreational hunting is carried out by shooting wild boar from the ground or from high seats at bait stations. Hunters may hunt alone, in small teams, or in large groups to carry out drive hunts in which animals are driven towards a line of hunters by people walking along a front to flush pigs from cover (Table 3). In Europe, the use of hunting dogs to flush wild boar out is widespread, particularly in areas with dense vegetation (Geisser & Reyer, 2004). Shooting is particularly efficient in areas with high densities of feral pigs or boar, as many animals can be culled in relatively short periods. Shooting may allow targeting specific age or sex classes and provide hunters with the additional incentive of meat and trophies. However, wild boar and feral pigs may learn to avoid hunters by becoming more active during the night and by avoiding areas where hunting occurs. The impact of shooting on spatial behaviour may depend on the level of human disturbance that animals have experienced and on the type and frequency of hunting. This is probably why some studies found that the use of dogs whilst hunting caused animals to increase home range sizes (Calenge et al., 2003; Sodeikat & Polheimer, 2002) whilst other studies (McIlroy & Saillard, 1989; Keuling et al., 2008) found no effect of shooting on spatial behaviour of boar and feral pigs.
One disadvantage of the use of dogs whilst hunting is that poorly-trained dogs may pursue and kill other animals, thus causing disturbance to local wildlife (Massei & Toso, 1993; Cruz et al., 2005). Other disadvantages include social disturbance and animal welfare issues. Social disturbance has the potential to increase contact rate with conspecifics with negative consequences for disease transmission. Animal welfare issues concern animals that are injured but not killed and dogs that can be severely injured by boar. Controlled shooting by experienced staff can overcome this problem, and dogs trained in flushing but not attacking feral pigs are less likely to be injured.
Shooting has been employed in a large number of projects aimed at eradicating or controlling feral pig populations. Several of these projects found this method effective, but time consuming (Coblentz & Baber, 1987; Kessler 2002; Geisser & Reyer, 2004; Cruz et al., 2005) particularly in the final stages of the eradication campaign when the density of animals was very low. Cruz et al., (2005) found that in 2000 the effort required to remove each pig from an island was 450 times greater than in 1998. However, opportunistic shooting over bait sites was particularly useful as a secondary technique to reduce feral pig numbers after trapping (Cruz et al., 2005).
Targeting a particular sex or age class through shooting could improve hunting efficiency. For instance, reducing juvenile survival has the largest effect on population growth rate, and increasing hunting pressure on adult females, particularly in years of low food availability, appears to be the most effective approach to population control (Sweitzer et al., 2000; Bieber & Ruf, 2005). However, compensatory responses to culling, such as increased immigration and reproduction, can limit the success of hunting (Hanson et al., 2009; Gamelon et al., 2011; Servanty et al., 2011). When shooting is used to prevent population growth, Häggmark Svensson et al., (2014) calculated that even removing 35% of the population is not sufficient (Figure 4.1) and Keuling et al., (2013) suggested that at least 65% of the starting population should be removed.
Shooting has been also been employed to control disease outbreaks, such as classic swine fever. A cost analysis model showed that, by implementing flexible hunting strategies that vary according to the density of pigs and disease prevalence, managers can minimize the cost of hunting and the sanitary costs associated with the infection over a specific period of time (Bolzoni & De Leo, 2007). These results can be used to design cost-effective contingency plans to control feral pig populations in case of disease outbreak.
Figure 4.1. Calculated and predicted total wild boar population in Sweden under different hunting pressures. Source: Häggmark Svensson et al., 2014.
Although in many European countries hunting caused the local extinction of wild boar in the late 19th century (Saez-Royuela & Telleria, 1986), present recreational hunting appears unable to control wild boar populations, as evidenced from current trends in wild boar and feral pig numbers in Europe, Australia, and the United States (Choquenot et al., 1996; Hutton et al., 2006). In Hawaii, Reeser and Harry (2005) showed that volunteer hunting or public hunting failed to remove feral pigs at the required rate, while professional hunters were more successful. In France, Toigo et al., (2008) found that between 1984 and 2004 the number of wild boar culled in the study area by recreational hunters rose from 200 to 1,000 and that the propensity of hunters to target adult males instead of females and piglets reduced the effectiveness of population control. In the US, the New York Department for Environmental Conservation precludes people from owning live feral pigs and precludes the hunting and trapping of feral pigs unless authorised by appropriate officials (Centner & Shuman, 2014). This prohibition discourages individuals from illegally releasing feral pigs in new areas and prevents recreational hunters from dispersing feral pigs. Conversely, recreational hunting offers the opportunity for hunters to be directly involved in participatory management of a sustainable resource. In this capacity, hunters may also volunteer precious skills and free labour that can benefit the often tight budgets of projects aimed at mitigating feral pig impact.
Table 3. Lethal methods to manage human-feral pig conflicts (modified from Massei et al., 2011).
| Method | Advantages | Disadvantages |
|---|---|---|
Trapping and euthanasia
|
|
|
| Snares |
|
|
| Ground shooting (with or without dogs) |
|
|
| Judas pigs |
|
|
Poisoning
Poisoning can achieve a rapid reduction in the number of feral pigs on a large scale and at moderate costs and has been used extensively to control feral pigs in the Americas and in Australia (e.g. Hone, 1983; Coblentz & Baber, 1987; McIlroy & Saillard, 1989; Cruz et al., 2005; Cowled et al., 2008) (Table 3). However, there is significant public opposition to the use of poisons for wildlife management in the UK, mainly due to welfare and safety concerns regarding target and non-target species. As there are no toxicants registered for use on feral pigs or wild boar in Europe, this method cannot currently be implemented in Scotland.
Judas pigs
Judas pigs are animals that are trapped, equipped with a radio-collar, and released so that they re-join other pigs. The whole group can then be located and shot by hunters (Table 3). When tested in Australia, the best results of this method were achieved by releasing sows captured in the same area where they had been trapped (McIlroy & Gifford, 1997). Out of the 15 Judas pigs released, 12 established contact with ≤12 other animals; pigs released in the same site of capture re-joined their group within 1 week. This method can be employed to locate the last few trap-shy feral pigs once the population density has been drastically reduced through trapping or shooting (Parkes et al., 2010). The main advantage of using Judas pigs is quick detection of animals; using this technique, Wilcox et al., (2004) showed that remaining pigs were detected within 1 hour compared to 4.1 hours to locate pigs without telemetry when the population was at its maximum density, and almost 60 hours when the density was very low. On Santa Cruz Island, only 9% of the 5,036 pigs removed to achieve eradication were dispatched as a result of their association with Judas pigs (Parkes et al., 2010). To reduce the cost and time required to trap the last few animals, pigs captured and kept in captivity at the beginning of a population control programme can be used as Judas pigs (McIlroy & Gifford, 1997). McCann & Garcelon (2008) suggested that all Judas pigs are surgically sterilised before release; Parkes et al., (2010) sterilized all males prior to release, and induced oestrus in the females to enhance their attractiveness to males, showing that these females were significantly better than the males at attracting other pigs.
Nonlethal methods
Fertility control
Chemical sterilization has been increasingly advocated as alternative to lethal control to reduce overabundant wildlife populations (Fagerstone et al., 2002; Massei & Cowan, 2014). Recently developed immunocontraceptives that can induce infertility for several years after administration of a single dose have reawakened interest in this method to control wild boar and feral pigs. Immunocontraceptives act by causing the production of antibodies against hormones or proteins essential for reproduction (Miller et al., 2008). These compounds have been formulated as single-shot vaccines, capable of inducing long-term infertility after a single injection. For instance, the Gonadotropin-Releasing-Hormone (GnRH) vaccine stimulates the production of antibodies against GnRH, which is, in turn, responsible for the production of sex hormones that lead to ovulation and spermatogenesis. Animals injected with this vaccine can be rendered infertile for 1 to 6 years, depending on species, age, gender and formulation (Killian et al., 2008; Miller et al., 2008; Massei et al., 2012; Massei & Cowan, 2014). Injectable GnRH vaccines have been tested extensively on many wildlife species, including feral pigs and wild boar. In most species, these contraceptives have been found to be safe and effective for many years without side effects on the animals’ behaviour, welfare, and physiology (Killian et al., 2006; Massei et al., 2008; Massei et al., 2012). Immunocontraceptives also have been proposed as a possible means of decreasing transmission of several wildlife diseases by reducing the abundance of new-born, susceptible animals within the population (Killian et al., 2007).
Fertility control has a high level of public acceptance and could be used to decrease numbers of feral pigs, particularly for isolated populations where immigration is nil and emigration negligible. However, managing feral pig populations by using injectable contraceptives could be more expensive than trapping and lethal control, as the costs of contraceptives will add to those of trapping. This method is thus more likely to be used for small scale, specific contexts where lethal control is not feasible or desirable. Examples of the latter are urban areas or national parks where shooting is not allowed or contexts where lethal control could affect contact rates and spread of diseases. Compared to trapping or shooting, fertility control is relatively slow in decreasing local abundance because the benefits of immunocontraception can be accrued after several years. On the other hand, fertility control can be used to keep the density of feral pigs at a set level once lethal control has been applied (Cowan & Massei, 2008). More research is ongoing to develop oral contraceptives to widen the spectrum of contexts where fertility control could be applied. If oral, nonspecies-specific contraceptives become available, the possibility of affecting non-target species must be addressed. Pig-specific feeders have been designed and evaluated for bait uptake by target and non-target species. For instance, the Boar Operated System (BOS™) is an effective device developed to deliver contraceptives and other pharmaceuticals to wild boar and feral pigs (Massei et al., 2010).
Fencing
Fencing is used in three different contexts: (1) as a preventive measure, to reduce feral pig impact into economically or conservation sensitive areas, such as nesting grounds, threatened habitats, wildlife refuges, farms and agricultural fields; (2) as a reactive measure to protect an area from feral pig impact once local eradication has been achieved; (3) to partition an area, typically a large island, into smaller units and to facilitate eradication from each unit (Table 4).
Many types of fencing, simple or electrified, are available and often consist of woven wire mesh 65 to 80-cm-high with strands of barbed wire strung along the top, bottom, and above the woven wire to create a fence of 110 to 120 cm in height; the fence often is buried to a depth of 40 to 60 cm to prevent pigs from forcing their way through it (Hone & Atkinson, 1983; State of Hawaii, 2007; McCann & Garcelon, 2008). Fences can be fitted with one-way gates to allow animals to exit an area but not to re-enter it.
Several electric fencing designs are available to exclude feral pigs; these usually consist of 2 to 3 strands of electrified fencing spaced 15-30cm apart. In Australia, different fence designs have been tested to protect crops and lambing paddocks (reviewed in Hone & Atkinson, 1983). In California, electric fencing has been used to prevent feral pigs from entering irrigated summer pastures (Barrett, 1978). In France, steel-wire electric fencing was used extensively to prevent damage to valuable crops over relatively small areas (Vassant & Boisaubert, 1984; Vassant, 1994), although Geisser & Reyer (2004) noted that this method may cause a shift in damage to adjacent, non-fenced fields. The general conclusions from many studies are that fence design affects the effectiveness of the method and that electrification significantly reduces the number of feral pigs crossing the fences, although the cost of maintenance is high for the electric fencing (Hone & Atkinson, 1983; Reidy et al.,2008). To prevent overgrown vegetation from damaging the fence or interrupting the circuits and to maintain the functionality of the fence, herbicides or manual clearance of vegetation must be used regularly (Littauer, 1993). Recently developed polywire electric fencing that uses conductive wires incorporated into ribbons or ropes is now available. Compared to fixed-steel wire electric fencing, the new model has the advantage that the fence can be easily set up, removed, and reused so that it can be employed temporarily. Using portable polywire, electric fencing, Reidy et al., (2008) found that 2 strands at 20 and 45 cm from the ground excluded 75% of feral pigs from bait stations in Texas. In Switzerland, Geisser & Reyer (2004) found that the 2-strand electric fencing, locally used to protect crops, was not as effective as shooting to decrease damage to crops by wild boar. In Slovenia, a combination of polywire-polytape electric fencing reduced damage to maize fields by 100% but also caused an increase in damage to neighbouring arable fields (Vidrih & Trdan, 2008). If permanent fencing is used, the main disadvantage is the initial setup costs and subsequent maintenance costs. In some areas, such as the Hawaiian rainforest, wire fences erected to exclude pigs from sensitive areas required monthly inspections and had to be replaced every 5 to 15 years (Katahira et al., 1983). In addition, fences had to be repaired following storms.
Repellents
A large number of olfactory, acoustic, and gustatory repellents have been developed to decrease the impact of wildlife on human activities (Conover, 2002). The overall conclusions of the many studies carried out to identify deterrents for wild boar suggested that animals habituate to all types of repellents within a few days (e.g. Vassant & Boisaubert, 1984; Cai et al., 2008; Vilardell et al., 2008; Schlageter & Haag-Wackernagel, 2012). Thus, the evidence so far suggests that repellents are not effective in reducing the impact of feral pigs and wild boar.
Table 4. Non-lethal methods to manage human-wild pig conflicts (modified from Massei et al., 2011).
| Method | Advantages | Disadvantages |
|---|---|---|
| Fertility control (injectable contraceptives) |
|
|
| Fencing |
|
|
| Diversionary feeding |
|
|
| Translocation |
|
|
Diversionary feeding
Diversionary feeding, also referred to as “supplementary feeding”, is often carried out by hunters to concentrate wild boar on a site and optimize culling effort in an area and to decrease crop damage (Geisser & Reyer, 2004). To remain effective, supplementary food must be available continuously, which makes this method expensive in terms of staff and resources; however, these costs can be absorbed by hunter groups and volunteers (Vassant et al., 1987) (Table 4). The effectiveness of this technique in reducing crop damage is controversial. While some studies reported that diversionary feeding was successful (Andrzejewski & Jezierski, 1978; Vassant, 1994; Calenge et al., 2004), others found limited or no effect on crop damage (Hahn & Eisfeld, 1998; Geisser & Reyer, 2004). In France, Vassant et al., (1987) used maize, distributed every day in the forest from late June till August and concluded that, although this method was effective to reduce crop damage by wild boar, its actual cost was similar to that of replacing crop losses.
In Switzerland, Geisser & Reyer (2004) found that in September and October, when maize and wheat are ready to harvest and particularly vulnerable to damage, wild boar hardly visited the feeding stations where supplementary food was provided, irrespective of the type of food these stations offered. In another French site, the use of maize as dissuasive food to protect valuable vineyards resulted in a 60% reduction in both the proportion of damaged vineyards and the level of damage, with net financial benefits for the farmers (Calenge et al., 2004). Although in mainland Europe supplementary feeding is still widely used by hunters, there is increasing concern that this practice could enhance reproductive success and survival of wild boar and, thus, contribute to long-term increase in damage to crops (Andrzejewski & Jezierski, 1978; Geisser & Reyer, 2004; Schley et al., 2008; Van Wieren & Groot-Bruinderink, 2010). However, if the amount of diversionary feeding provided is small compared to the availability of natural food and the feeding is localized in time and space and used to decrease damage to localized, valuable crops, the effect of this method on feral pig population dynamics might be regarded as negligible (Calenge et al., 2004; Conover, 2002).
Translocation
Translocation of problem animals is increasingly advocated to mitigate human–wildlife conflicts, often as a result of public pressure rather than motivated by scientific or economic evidence (Bradley et al., 2005; Athreya et al., 2010; Fernando et al., 2012). A recent review of translocation of problem animals found that, despite belief that this method was humane; translocations may have a detrimental impact on survival rates and lead to extreme dispersal movements (Massei et al., 2010). Translocated animals may resume the nuisance behaviour at the release site and for some animals translocation may result in malnutrition, dehydration, decreased immunocompetence and predation. More importantly, translocations have the potential to spread diseases (Kock et al., 2010, Fernando et al., 2012) (Table 4).
Worldwide, transport and release of feral pigs by hunting clubs is the most important factor explaining the marked increase in distribution of this species (e.g., Gipson et al., 1998; Spencer & Hampton, 2005). As translocations may encourage irresponsible releases many countries have made translocating feral pigs illegal (Hutton et al., 2006; Centner & Shuman, 2014). However, enforcing the law banning translocations is particularly challenging as feral pigs are relatively easy to trap and relocate to a different site.
A simulation model to quantify the likely amount of effort required to control and/or eradicate discrete wild boar populations in Scotland, under a variety of starting abundance scenarios
The aim of this part of the study was to assess culling efficiency and estimate the number of feral pigs that could be removed by trapping and shooting for given levels of effort. We used data from available records on trapping and shooting to estimate culling rates and to derive estimates of the relative costs of control. We then applied these culling rates to populations of different sizes to estimate the effort and costs required for eradication or maintenance of the population at the current level.
Methods
Data collection
The National Wildlife Management Centre (NWMC) undertook regular feral pig trapping in two woods within Forestry Commission England’s (FCE) Forest of Dean District for research purposes. Information on numbers, age class and sex of feral pigs caught and trapping effort (in trap nights) from January 2008 to April 2010 was extracted from the trapping database.
Information on numbers, age class and sex of feral pigs shot, and shooting effort expended were received from the FCE Forest of Dean District, which is the only FCE District within which feral pigs are specifically targeted for shooting by FCE staff (i.e. they are actively pursued and not simply shot opportunistically). Similar records were not available from Forestry Commission Scotland (FCS). Rough estimates, based on expert opinion, of the number of feral pigs in each population currently present in Scotland were received from Science and Advice for Scottish Agriculture (SASA) and FCS. Each population was treated as a closed unit, with no immigration or emigration. No formal measures of uncertainty were available for estimates of population size. We ran simulations for populations ranging from 10 to 200 individuals, thus model outputs are likely to include estimates of effort and cost that are indicative of those likely to be appropriate for all current populations in Scotland.
Simulation models
The rate of population change imposed by control effort was estimated for trapping and shooting separately. We assumed that trapping effort (measured as the number of trap nights) was fixed over time (which it generally was), and that shooting removed a constant proportion of the population throughout the campaign. Thus the rate of change (λ) was modelled as:
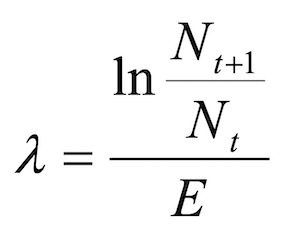
Where:
Nt+1= Number of pigs remaining at the end of the culling campaign
Nt = Number of pigs at the start of the culling campaign
E = Effort (trap nights for trapping and man hours for shooting
We estimated the number of feral pigs that could be trapped or shot using NWMC’s trapping data and the FCE’s cull records, across a range of input population sizes and efforts to simulate variation in effort and the effects of time of year.
We calculated the proportion of the feral pig population that was shot by each ranger by comparing cull records from 2009 with estimates of total pig abundance in the Forest of Dean from the same year. We assumed that the proportion culled for a given effort was the same between populations of feral pigs throughout Scotland. However, we modelled culling rate with a uniform distribution, with the lower and upper limits described by the two rangers’ culling rates. Scenarios were run for a range of population sizes derived from the information provided under section 1c: these populations varied from a few individuals to 200 animals.
Starting abundance (in each Scottish population) was multiplied by the culling rate (the proportion of the total population culled per man hour) and effort (man hours) for effort ranging from zero to 4,250 man hour in 250 man hour increments. Simulations were run 2000 times each in Crystal Ball software (Decisioneering Inc. Denver, USA). Outputs were the median and 95th percentile range of the estimate of the cumulative number of feral pigs culled for each increment of effort. These were plotted as curves to illustrate the relationship between effort and numbers of pigs culled.
We undertook a similar analysis using the trapping data for 2009, replacing the Forest of Dean feral pig abundance estimates with those estimated for Scotland. Trapping rate data were modelled with a triangular distribution since they could be broken down into seasonal rates and summarised across the year, giving three rates with which to work (highest, lowest and most likely). Effort (trap.nights) was varied from 0 to 200 in 10 trap.night increments.
We applied the number of feral pigs that are required to be culled to achieve each of the objectives (eradication or stabilisation) to these curves to derive an estimate of the effort required to achieve each objective for each scenario. We then plotted the starting number of feral pigs against the effort required to yield the requisite number of carcasses. Eradication was taken as the effort at which the number of animals culled equalled the number in the starting population, which assumes that the number of births was the same as the number of non-cull deaths. Population stabilisation was assumed as achieved by removal of 65% of the starting population (Keuling et al., 2013).
Costs of control
We estimated the cost per pig culled and total cost of each campaign by multiplying the effort expended with shooting and trapping feral pigs by the cost per hour (in terms of labour) and adding the setup costs (including equipment purchase and installation) and transport costs. For all calculations we assumed a daily labour cost of £240, which equates to approximately £30 per hour for an eight-hour working day.
Sensitivity analysis
In order to quantify the likely effects on culling effort required, of changing conditions under which feral pigs exist within Scotland, we ran simulations with varying culling efficiency. The underlying assumption was that culling efficiency (defined as number of pigs removed per unit of effort) whether by trapping or shooting) is likely to decline with density, so that varying culling efficiency would indicate the effects of changes in local abundance and/or distribution, thereby implying changes in effort required should feral pigs disperse in response to a culling campaign. We ran simulations, for both trapping and shooting independently, for culling efficiencies of 10% and 50% of the values measured in the field.
Results
Trapping records
From January 2008 to March 2010 26 adult females, 16 adult males and 88 juvenile feral pigs were trapped in the Forest of Dean woods over 633 trap nights involving ten to 14 traps per night (Figure 4.2). This resulted in a simple average of 0.21 feral pigs per trap night. In addition, 13 adult females were re-trapped. However, it was not possible to distinguish adult males and juveniles trapped for the first time from re-captures since these were not marked before release. We assumed that all male and piglet captures represented first-time captures, which may have over-estimated trapping rates.
Trapping effort varied over time (Figure 4.2). Neither the number of adults nor juveniles captured correlated with effort (Pearson’s correlation, p>0.01), but the total number of captures did (r = 0.502, n = 17, p = 0.04). The summer peaks in piglet captures are likely to result from the peak time of year for births (early spring), although piglets may be born throughout the year.
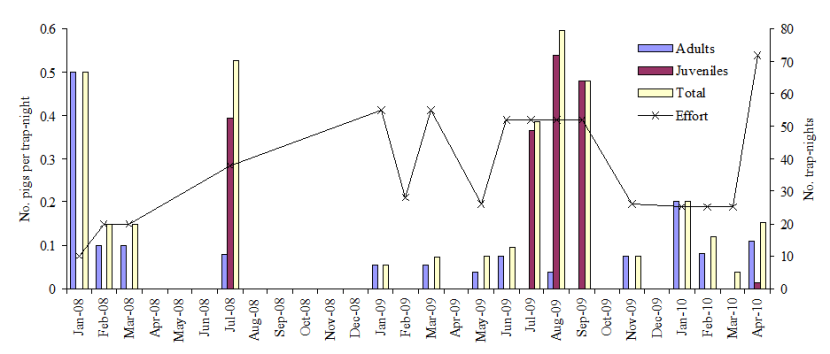
Figure 4.2. Feral pig trapping results.
Click for a full description
The line represents trapping effort, and the bars number of pigs caught per trap night. The Total column combines all captures, including re-captures of females.
The total number of captures was significantly higher during the summer months than at any other time of year (Poisson count regression, with ‘effort’ and ‘season’ as explanatory variables χ2 = 135.8, d.f. = 6, p<0.001), but the number of adults trapped per trap night was fairly constant. The poor trapping results during 2008 may be explained by a high degree of trap-interference. Relationships between NWMC staff and members of the public using the study site were actively improved during late 2008, and the frequency of trap interference subsequently reduced. In addition, concomitant tracking of feral pigs using GPS tags allowed more effective targeting of trapping towards areas more heavily used by feral pigs. In previous studies monitoring of bait consumption at bait points or along bait trails has advised trap placement (Saunders et al., 1993).
FCE cull data
Juvenile feral pigs were not represented in the FCE cull records used. Consequently, estimates of effort derived from FCE data are likely to over-estimate the effort required to shoot sufficient numbers of feral pigs to achieve either of the two objectives since juveniles will need to be included. Two FCE rangers currently undertake the vast majority of the feral pig culling work for FCE, with one further ranger opportunistically shooting feral pigs during deer culling duties. From 1st January to 31st December 2009 one ranger (“A” in Figure 4.3) spent approximately 616 hours targeting feral pigs. This involved establishing bait points (0.5 hours per day for eight days), and shooting from a high seat (three hours at dawn and at dusk per day for 14 days). This was repeated every month for seven months and yielded 13 males and 20 females. This equates to one pig shot for every 18 hours and 40 minutes of effort. In addition, this ranger retrieved two males killed by collisions with road traffic.
The other ranger (“B” in Figure 4.3) spent approximately 960 hours targeting feral pigs. This involved 8.5 hours per week pre-baiting and shooting from high seats and 12 hours per week of stalking throughout the 12-month period. This yielded 13 males and 10 females, and three males and two females were retrieved that had been killed in collisions with road traffic. This culling rate equates to one pig for every 41 hours and 44 minutes of effort.
A further ranger (“C” in Figure 4.3), during deer culling duties, opportunistically shot two female feral pigs, three males and retrieved a male road casualty. However, since feral pigs were not specifically targeted by this ranger, it was not possible to estimate the effort expended culling them.
Figure 4.3. Number of feral pigs shot, by season, during 2009 by FCE rangers A, B and C.
Culling rates were not constant throughout the year (Figure 4.3). More pigs were shot during the summer than at any other time of the year, although ranger B shot slightly more pigs in autumn than at any other time of year. Fewest pigs were shot during the winter. This is likely to be largely due to population density being lowest during the winter and highest during the spring and summer. In addition, less effort tends to be devoted to deer culling during the summer months.
Costs of control
Trapping
Single-capture feral pig box traps used by NWMC cost £500 to manufacture, and it has taken 8 man.days to place 13 traps (i.e. to survey for suitable locations, to move traps on site and build them takes four people two days for 13 traps). These traps required pre-baiting with maize, taking 3 sacks of 25kg each per day, costing £10 per sack, for 7 days, taking 3 hours per day, including local travel time, and with £5 in local travel costs. Thus, establishment costs were:
£500 + ((£240 x 8)/13) + £1 + ((£30 x 3 x 7)/13) + (5/13) = £698 per trap
Trap setting then took 40 minutes per trap per night, including local travel time, and the following morning trap checking took 10 minutes per trap, costing £25 in labour and £0.40 in local travel costs. Staff engaged in trapping needed local accommodation and expenses, estimated at £76 per person per day for two people.
The culling of trapped feral pigs should be fairly quick, if shooting is the preferred method. However, carcasses will need to be extracted, probably by quadbike and sled moving to a 4-wheel drive vehicle equipped with an internal winch to lift the carcass into the vehicle. We have not included these additional costs, nor costs for rifle purchase, maintenance and ammunition in estimates of trapping costs.
The total costs of trapping for 2009 are summarised in Table 5. These figures exclude costs of culling and extraction and travel to the locality. However, they include accommodation costs and expenses associated with over-night trips. These would not accrue if staff local to the area were used to trap feral pigs. This effort resulted in the capture of nine males, six females and 72 piglets, at a cost of £287 per pig, from a breeding population of approximately 21 adults.
Shooting
Rifles and peripheral equipment suitable for shooting feral pigs can be purchased for approximately £1000. Each shooter may require approximately 100 rounds of ammunition per year to allow for culling, zeroing and practice, costing approximately £100. FCE rangers spent 1,576 hours culling feral pigs during 2009, at a labour cost of £30 per hour. Local travel is also likely to have cost approximately £5 per day.
Assuming that new rifles were purchased specifically for culling feral pigs, FCE may have spent £2,200 on rifles and ammunition, £985 on local travel, and £42,280 on labour to cull 56 pigs. The total cost may have been approximately £45,465.
Table 5. Costs of trapping with single-capture box traps over 398 trap nights in two FCE woodlands during 2009.
| Item | Unit cost (£) | Number of units | Total cost (£) |
|---|---|---|---|
| Trap establishment | 698 | 14 | 9,772 |
| Trap setting and checking | 25.40 | 14 x 30 nights | 10,668 |
| Accommodation and expenses | 76 | 2 x 30 | 4,560 |
| Total | - | - | 25,000 |
Simulation models
As described above, cull rates (by shooting) varied from one feral pig every 18 hours 40 minutes to one every 41 hours 44 minutes of effort. These equate to 0.0129% to 0.0288% of the population culled per hour of effort with an average of 0.0191%. Applying these values to estimates of feral pig abundance yielded a sigmoidal curve of effort versus cull numbers (Figure 4.4).
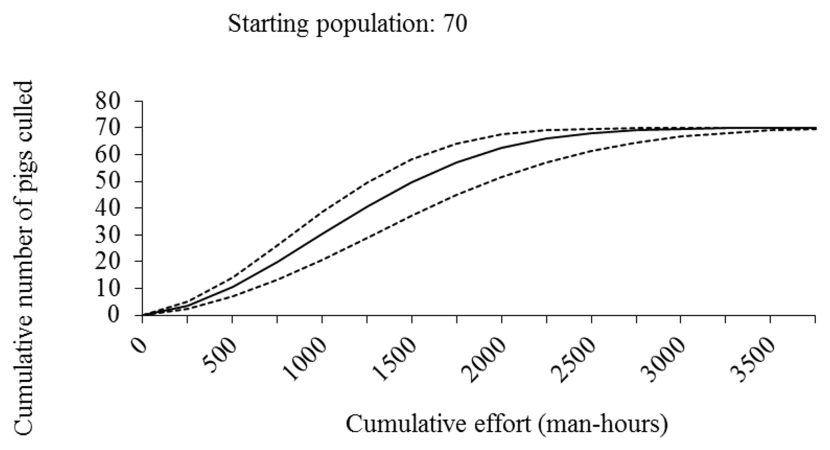
Figure 4.4 Change in the total number of feral pigs culled with increasing shooting for a population of 70 feral pigs.
Repeating these simulations across the range of starting pig abundances for each site resulted in similarly-shaped curves, which varied in location between the axes.
Trapping rates varied from 0.183% of the population removed per trap night during the autumn, to 0.427% during the summer, with an average rate across the year of 0.251%. Applying these values to the abundance estimates yielded a sigmoidal curve of effort versus numbers trapped (Figure 4.5).
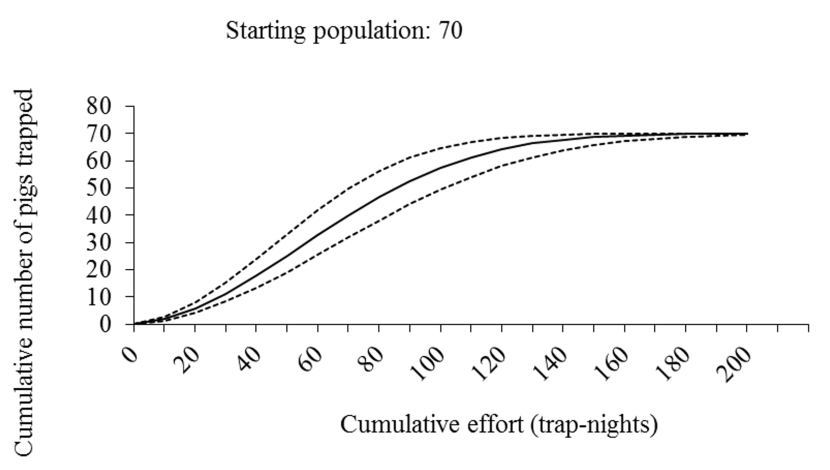
Figure 4.5 Change in the total number of feral pigs culled with increasing trapping effort for a population of 70 feral pigs.
The effort required to cull sufficient numbers of pigs varied between scenarios. Predictably, the greatest effort was required to eradicate the largest population (Table 6) and the least effort to maintain a population at its current size. However, it is important to bear in mind that this relates to a single year of culling. If successful, eradication only needs to be done once, whereas the effort to stabilise a population needs to be invested every year in perpetuity. Due to the assumption that a fixed proportion of the population was removed for each unit of effort, the effort required to maintain a population at its current size was the same, regardless of population size. To remove 65% of a population, which should keep the population static, between 1150 and 1750 man hours of shooting (median: 1375 man hours) or 65 to 90 trap nights of trapping (median: 80 trap nights) would be required each year.
Table 6. Effort required to eradicate populations of feral pigs, using shooting (in man.hours) and trapping with single-capture box traps (in trap.nights), for scenarios likely to be experienced in Scotland.
| Population size | Shooting - Median | Shooting - Lower 95th percentile | Shooting - Upper 95th percentile | Trapping - Median | Trapping - Lower 95th percentile | Trapping - Upper 95th percentile |
|---|---|---|---|---|---|---|
| 10 | 2500 | 3000 | 2000 | 130 | 160 | 110 |
| 15 | 2500 | 3250 | 2250 | 140 | 170 | 120 |
| 30 | 2750 | 3500 | 2250 | 160 | 190 | 130 |
| 50 | 3000 | 3750 | 2500 | 170 | 200 | 140 |
| 60 | 3000 | 3750 | 2500 | 170 | 200 | 140 |
| 70 | 3000 | 3750 | 2500 | 170 | 200 | 140 |
| 100 | 3250 | 3750 | 2750 | 180 | 210 | 150 |
| 125 | 3250 | 3750 | 2750 | 180 | 210 | 150 |
| 150 | 3250 | 4000 | 2750 | 180 | 220 | 150 |
| 200 | 3250 | 4000 | 2750 | 190 | 220 | 150 |
The effort required to shoot or trap a population to extinction increased with population size, but did not follow a straight line (Table 6). In the example given in Table 6, for a population of 100 feral pigs, eradication through shooting is likely to require between 2750 and 3750 man.hours, hence worst-case planning would be based on the lower 95th percentile of 3750 man.hours.
Costs of control
The cull required for each scenario could be achieved by trapping by varying the number of traps and the number of nights of trapping, and by shooting by varying the number of rangers and number of hours spent shooting. Assuming the cost structures detailed above, but including accommodation and expenses for shooting as well as trapping, for comparative purposes, these relationships can be described by surface plots (Figures 4.6 and 4.7). Four areas on the surface are defined by a curved boundary and variable shading. For trapping these correspond to combinations that cost up to £20,000, between £20,001 and £40,000, between £40,001 and £60,000 and between £60,001 and £80,000 (Figure 4.6). For shooting these correspond to combinations that cost up to £20,000, from £ 20,001 to £30,000, from £30,001 to £40,000 and from £40,001 to £50,000) (Figure 4.7).
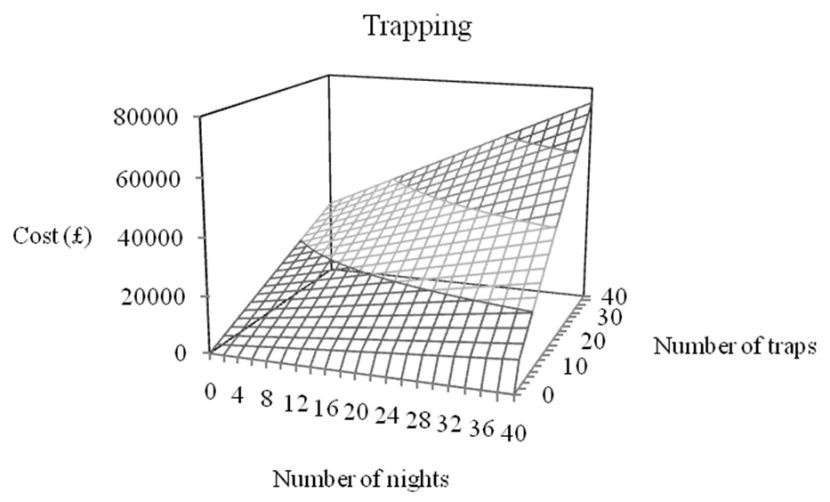
Figure 4.6 The costs of trapping with variation in effort.
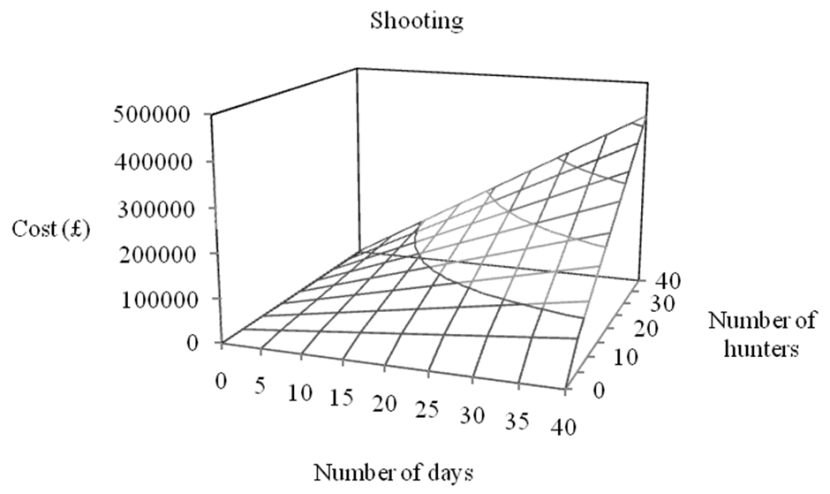
Figure 4.7. The costs of shooting with variations in effort.
Figure 4.6 suggests that trapping could be more cost-effective using fewer traps over more nights than more traps over fewer nights, largely due to the enhanced establishment costs associated with purchasing large numbers of traps. The reverse may be true for shooting; Figure 4.7 suggests that shooting could be marginally more cost-effective using more rangers over shorter periods of time than fewer rangers over longer periods.
Using the cost matrices constructed to derive these curves we estimated the optimal costs (cheapest combination) required to achieve target culls for each scenario. The results presented in Table 7 are based on the lower 95th percentile limit of the estimate of effort (the greatest estimate of effort required) which represents budgeting for the worst-case scenario. Budgeting according to the median or upper 95th percentiles poses a greater risk of providing inadequate resources to achieve the required cull.
Sensitivity analysis
To facilitate planning against the most challenging conditions, we report estimates of effort for the lower 95th percentile limit, which represent the highest likely costs.
Predictably, as culling efficiency (modelled as the shooting and trapping rate) decreased, the effort required to cull sufficient feral pigs increased (Table 8). Under all scenarios both trapping and shooting required approximately three times the effort when culling efficiency was 10% of that measured in the field and approximately 1.5 times higher when culling efficiency was 50% of that measured.
Discussion
The key implication of Tables 5 and 6 is that eradication or population stabilisation of feral pigs through culling appear to be feasible in Scotland, given current best estimates of this species’ distribution and abundance. When considering each method in isolation, trapping with single-capture box traps is predicted to offer lower cost than shooting. However, the comparison of shooting with trapping may not have been entirely valid in this simulation, since shot animals were killed, whereas trapped animals were released alive. We partially controlled for the potential biases this could introduce in favour of trapping by only using data from the first time of capture for adult females. Unfortunately, it was not possible to identify, and therefore control for re-trapped males and piglets since these were not marked during the field study. This could have resulted in our estimates of trapping efficiency being biased high if the recapture rate was higher than the rate of trap avoidance. While both trap avoidance and attraction have been inferred for individuals of other species (e.g. in badgers: Tuyttens et al., 1999), we were not able to quantify the rates of these for the feral pig population studied.
Table 7. Estimated costs of delivering a cull, by shooting or trapping, likely to be sufficient to eradicate populations of feral pigs in scenarios likely to be experienced in Scotland.
| Population size | Shooting - Number of hunters | Shooting - Number of nights | Shooting - Cost (£) | Trapping - Number of nights | Trapping - Number of traps | Trapping - Cost (£) | Total cost (£) (combined approach) |
|---|---|---|---|---|---|---|---|
| 10 | 11 | 34 | 100,814 | 27 | 6 | 12,406 | 113,220 |
| 15 | 11 | 37 | 109,127 | 28 | 6 | 12,711 | 121,838 |
| 30 | 13 | 33 | 118,674 | 32 | 6 | 13,929 | 131,994 |
| 50 | 14 | 33 | 124,098 | 20 | 8 | 14,108 | 138,206 |
| 60 | 14 | 33 | 124,098 | 20 | 8 | 14,108 | 138,206 |
| 70 | 14 | 33 | 124,098 | 25 | 8 | 14,108 | 138,206 |
| 100 | 14 | 33 | 124,098 | 35 | 6 | 14,842 | 138,940 |
| 125 | 14 | 33 | 124,098 | 35 | 6 | 14,842 | 138,940 |
| 150 | 14 | 36 | 134,616 | 32 | 7 | 15,440 | 150,056 |
| 200 | 14 | 36 | 134,616 | 32 | 7 | 15,440 | 150,056 |
Table 8. Maximum effort (expressed as lower 95th percentile limit) likely to be required to eradicate populations of feral pigs, using shooting (in man hours) alone and trapping (in trap nights) alone, for culling rates (efficiency) at 10% and 50% of those measured in the field, for scenarios likely to be experienced in Scotland.
| Population size | Shooting - 10% efficiency | Shooting - 50% efficiency | Trapping - 10% efficiency | Trapping - 50% efficiency |
|---|---|---|---|---|
| 10 | 10,000 | 4,500 | 530 | 230 |
| 15 | 11,000 | 4,750 | 560 | 250 |
| 30 | 11,500 | 5,250 | 620 | 270 |
| 50 | 12,750 | 5,500 | 650 | 280 |
| 60 | 12,750 | 5,500 | 670 | 290 |
| 70 | 13,250 | 5,750 | 680 | 290 |
| 100 | 13,250 | 5,750 | 700 | 310 |
| 125 | 13,250 | 5,750 | 700 | 310 |
| 150 | 13,500 | 5,750 | 700 | 310 |
| 200 | 13,750 | 6,000 | 750 | 330 |
Among feral pig eradication campaigns, the majority used shooting and trapping together (simultaneously or sequentially) to deliver the cull, and considered this combined approach to be essential to maximise the chances of eradication (Saunders & Bryant, 1988; Lombardo & Faulkner, 2000; McCann & Garcelon, 2008). Moreover, shooting can be made more cost-effective if effort is varied in response to changes in feral pig density as the cull progresses (Bolzoni & De Leo, 2007). To achieve population eradication a combined approach is more likely to be successful than a single approach, with costs likely to approximate the sum of shooting and trapping for each scenario detailed in Table 7.
From our results, efficient culling of small boar populations (up to 200 animals) should employ several shooters for short-duration, intensive shooting campaigns and approximately four to 16 traps over several nights of trapping. Moreover, trapping success is likely to be highest during the summer, due to the presence of piglets, although reasonable numbers of adults may also be caught during the winter, but trapping efficiency is likely to be very low in autumn, particularly during years of high beech mast production. FCE rangers appear to have had greatest shooting success in the summer too, but this may be somewhat confounded by their focus on deer culling duties at other times of the year. We cannot conclude how shooting should be deployed on the ground in Scotland, since published studies on eradication campaigns were on either small, off-shore islands (Lombardo & Faulkner, 2000; Schuyler et al., 2002 [in McCann & Garcelon, 2008]; Parkes et al., 2010) or fenced populations of feral pigs (Barrett et al., 1988; McCann & Garcelon, 2008), which are different from the situation in Scotland. Some authors have noted altered behaviour and enhanced dispersal of feral pigs in response to culling campaigns (Saunders & Bryant, 1988; Dexter, 1996), and this possibility should be considered when planning population control or eradication campaigns in Scotland.
NWMC staff improved their trapping techniques as their studies progressed. The results reported here are for the use of single-capture box traps, with which we have had considerable success. However, multi-capture corral traps have recently been trialled, which may be more expensive to purchase and install, but which can capture several animals (including entire maternal groups) at a time, with potential efficiency savings. As costs and efficiencies become apparent for this and other types of trap it may be helpful to quantify the contribution that they could make to the management of feral pigs in Scotland.
Sensitivity analysis
Our assessments of effort by trapping and shooting were based on measurements of the use of these methods on one population and for one year in English woodlands. Feral pig population abundance varies with time, and so the feasibility of population eradication or the number that need to be removed to stabilise the population is also likely to vary. For a population of a given size, more effort is likely to be required to cull a sufficient number of individuals if they are more widely dispersed (i.e. at lower density), than if they are aggregated. Indeed, our simulations of reducing culling efficiency (Table 8) imply substantially elevated required effort for lower culling efficiencies. Shooting and trapping rates varied between individual rangers and times of year, but culling efficiency is also likely to vary with feral pig population density, which can change with local feral pig abundance and dispersion. We consider it likely that the feasibility of eradication will reduce if feral pigs disperse, for whatever reason, to exist at densities substantially lower than those on which we based our calculations. However, at the time when data on this cull were collected, estimated feral pig densities within the Forest of Dean were among the lowest for any population in England (1.08km-2), and may be comparable to some populations in Scotland. Consequently, our estimates of shooting effort were based on data collected under some of the most difficult culling conditions. Trapping data were collected in an English woodland, which had feral pigs present at 5 km-2, which was at the time the highest density recorded in England (Gill & Brandt, 2010). However, this is much lower than densities experienced in continental Europe, where densities of up to 66 animals km-2 have been estimated in an Italian area (Franzetti et al., 2012). Consequently, our estimates of trapping effort required are also likely to represent very difficult conditions. Table 8 predicts that the effort required to cull sufficient feral pigs will increase with declining culling efficiency. However, since the current densities of feral pigs in Scotland are likely to be low, it is likely that while the effort required might multiply with declining density, it may remain within the same order of magnitude of the estimates presented here.
Clearly, these simulations indicate that costs are likely to increase with declining culling efficiency, but they do not indicate the point at which eradication or control is likely to become unfeasible. In practical terms a decision is required to define the cost threshold beyond which control will not be undertaken. Application of this threshold to our simulation models would allow identification of the minimum culling efficiency required. In turn, and following information on culling rates from a trial cull, this would indicate whether it is feasible to cull sufficient feral pigs to achieve eradication or control given the resources available.
Our analyses also reveal priorities for future research to enhance confidence in the predictions made using the approaches presented here. All our analyses rely on assumptions which caused uncertainty regarding the reliability of some outputs. In addition, the empirical data summarised are limited, which enhances data variability and decreases the precision with which we are able to make predictions. In order to derive more robust estimates of effort required and associated costs for scenarios likely to be experienced in Scotland, we recommend the research highlighted below to fill knowledge gaps:
- Feral pig abundance in Scotland was not measured. Robust estimates of abundance collected for each discrete feral pig population are needed to obtain reliable estimates of the effort required to reduce population size or eradicate local populations.
- Estimates of effort required for different densities of feral pigs would help inform control efforts at the outset and during their progression, as density declines. This effort is likely to be affected by a wider distribution of feral pigs if animals respond to culling by dispersing. Estimation of the effort required under such conditions could help identify the likely higher costs entailed. Moreover, spatially explicit models describing sampling effort and feral pig responses could inform the design of efficient and effective management strategies to control dispersal and hence reduce effort required. Whenever trapping or shooting is carried out in Scotland, data on effort (expressed as man hours and other costs) should be collected and analysed to inform feral pig management.
- It is uncertain whether the proportion of a population that is culled remains constant with a fixed-effort cull. This hypothesis underpins sustainable harvesting theory, but it has not been robustly demonstrated for populations of wild ungulates. A proper assessment of whether this is the case, for both trapping and shooting, or measurement of how the proportion culled changes as the cull progresses during eradication campaigns, would greatly facilitate the rapid assessment of the resources required to deliver a cull of a known proportion of a population.
- Data on culling efficiency were based on culls of adult feral pigs only. However, particularly during eradication campaigns, juvenile pigs must also be targeted. Data collection in this area will indicate whether a sufficient proportion of juveniles is targeted and ultimately result in a reduction of the effort and costs associated with shooting feral pigs.
- The fairly low variation in estimated shooting effort required, and hence costs, were largely due to having only two estimates of shooting effort with which to build a data distribution. These data were suitable for application to the Forest of Dean situation, from where the data were derived, but may be unrealistic for Scottish populations. Consequently, empirical estimates of effort and numbers of feral pigs culled are required to better describe variation in this crucial measure.
- We modelled Scottish populations as discrete populations. This assumption is probably true for most since there are considerable distances between them. However, the range of each population may not be adequately defined. It would be beneficial to define the status of populations with respect to each other (i.e. whether they are independent or part of the same population) and to map the extent of each populations’ range to develop a clear definition of a control or eradication zone for each population and to facilitate development of management plans.
- We have not considered the reaction of the public or landowners to these alternatives. The reaction of stakeholders could have large and unknown effects on the efficiency, practicality and cost of these options.
When deciding whether to eradicate or control feral pig populations, managers may wish to consider an adaptive management framework (AMF, Gosling & Baker, 1989; Kaji et al., 2010; Parkes & Barron, 2012). AMF is a systematic, rigorous approach for deliberately learning from management actions with the intent to improve subsequent management policy or practice. The AMF assumes natural resource management policies and management actions are not static but adjusted based on the combination of new scientific and socio-economic information in order to improve management by learning from the ecosystems being affected. Following the AMF, good estimates of population density, size and range could be derived in advance of culling in order to estimate the resources likely to be needed and how to most efficiently deploy them. This could be followed by culling, with information on numbers culled per unit of effort used to repeatedly re-estimate starting population size, the proportion of the population removed per unit of effort, and hence to refine estimates of effort required. The principle behind this approach is that as the cull progresses, more and better quality data are collected, yielding more accurate estimates to inform management decisions.
Conclusions
It is often assumed that reducing the density of pigs reduces their actual or potential impacts, although the results of management actions are rarely monitored (Reddiex et al., 2007), and density-damage relationships rarely quantified (e.g. Hone, 1980; Choquenot et al., 1997).
This review highlighted that, given the current distribution and likely numbers of feral pigs in Scotland, three options are available to managers and stakeholders:
- Do nothing
- Prevent growth and spread of feral pig populations
- Eradication.
The first option, if adopted, carries the risk of populations growing in parallel with environmental and economic impact. Parts of the US provides a good example of what may happen on a smaller scale such as may be experienced in Scotland under this policy. In states like Texas, where action on feral pig control was taken well after population establishment, eradication is no more regarded as an option as pigs have spread and multiplied beyond control. In other states, prompt action led to eradication whilst populations were still isolated and manageable (Centner & Shuman, 2014). In general, the longer feral pigs have been established, the wider their distribution and numbers, which makes eradication very difficult to achieve, and is often opposed by recreational hunters. Although the income from hunting tourism can generate local revenue, this benefit must be weighed against environmental and economic costs of feral pig impact.
The second option, although technically possible, requires constant effort to maintain population growth as close to zero as possible. As wild boar and feral pigs have the highest reproductive output amongst ungulates, preventing population growth is challenging and often impossible to achieve through recreational hunting only. A multitude of examples from mainland Europe suggests that recreational hunters rarely control wild boar numbers sufficiently to prevent population growth.
However, where eradication is unfeasible or is opposed by local groups, sustained control may be an option to reduce the impact of feral pig populations. Sustained control includes methods to provide short-term solutions (e.g., to reduce crop damage or vehicle collisions) and long-term management to mitigate or prevent the occurrence of conflicts for several years. If a reduction of feral pig density is expected to mitigate the conflict, the feasibility, sustainability, cost, humaneness and social acceptance of different control methods should be evaluated. In case any of these issues is expected to be controversial, for instance if strong public opposition arises toward some of the proposed methods or if adequate funding is not available to implement a population reduction programme, the control strategy should be re-evaluated.
This review indicated that coordinated culling can quickly reduce the size of a population. When hunters are co-ordinated, shooting may substantially reduce feral pig numbers (Geisser & Reyer, 2004; Parkes et al., 2010). Based on these considerations, several authors (Geisser & Reyer, 2004; Massei et al., 2011; Keuling et al., 2013) recommended the development and introduction of integrated harvest models among local hunting teams to maximize the success of population control. We suggest these models could include: (1) integrating shooting with other methods, such as trapping or fencing; (2) employing reliable methods to estimate feral pig density before, during and after control; (3) monitoring of the impact of different hunting pressures on population size and impact; and (4) coordinating planning and effort with other stakeholders to mutually agree goals to achieve participatory management of feral pig populations.
Many stakeholders believe that the ultimate aim of feral pig control should be eradication. This view, however, is not shared by all stakeholders. In addition, different stakeholders may choose or prefer different methods of control, although the method chosen is not necessarily based on evidence. Although eradication of feral pigs is difficult and expensive, it has been achieved, largely on small islands. Feral pigs have now been eradicated from at least 25 islands with areas from 5 to 600 km2 (Kessler, 2002; McCann & Garcelon, 2005).
Sites with newly established, geographically isolated populations can be regarded as ecological islands and this is likely still the case for all the populations of feral pigs occurring in Scotland. If eradication is the ultimate aim, efforts in these areas should be focused toward eradication before population range and numbers increase, although shooting could cause pigs to move considerable distances (Leaper et al., 1999) and may ultimately affect the success of a local eradication.
Control of feral pigs may also require that managers alter techniques in response to changing animal densities, animal behaviour, and environmental conditions. For instance, trapping does not always remove older, more experienced pigs, ground shooting may preferentially remove solitary boars, and trapping may preferentially remove females (Choquenot et al., 1993; Saunders et al., 1993; Mitchell, 1998). The vast majority of successful eradication programmes employed an integrated management approach where several control options were carried out at the same time or in sequence. This ensured that animals that could not be targeted by one method could still be removed by adopting complementary control techniques.
For islands and geographically isolated populations, an intensive eradication programme is likely to be the most cost – effective option for the following reasons: (1) a high intensity programme can achieve eradication in a short period; (2) fewer pigs need to be culled as populations are not given the time to reproduce or to learn to avoid control; (3) the high cost of an intensive eradication programme is likely to be less than that of sustained control over a period of several years; and (4) a short, well-managed programme is likely to receive less public opposition (McCann & Garcelon, 2008).
In addition, the longer an eradication project runs, the more it is exposed to factors that can undermine its success (Morrison, 2007; Parkes et al., 2010). These factors include new releases of animals in areas already cleared of feral pigs, reproduction that causes the pig population to increase, public opposition from some stakeholders, legal challenges arising in the course of the project, increased lack of staff motivation, and funders’ fatigue which may result in lack of sustained funding to complete the programme.
Post-eradication monitoring should be included in any eradication programme to confirm achievement of the objectives or to respond if eradication has not been achieved. Monitoring is critical to determine the effectiveness of the methods used to decrease feral pig population size or impact. The greatest challenge for managers of eradication programmes is deciding whether the inability to detect pigs indicates that the species has been eliminated. Stopping the monitoring too soon risks declaring eradication incorrectly, but monitoring for too long results in a waste of resources (Morrison et al., 2007; Ramsey et al., 2009). Several methods are available to monitor the effects of population control on feral pig numbers. These include the indices of abundance and density estimation methods described under Section 2b as well as other methods based on monitoring bait uptake at baiting stations. Indices of abundance based on bait consumption tend to overestimate population reduction because they do not include animals that do not feed on the bait.
Besides quantifying the impact of population control, monitoring has the advantage that managers and stakeholders can see the long- and short-term consequences of control, for instance the decrease in activity signs, such as soil disturbance, or the increase in species previously negatively affected by the presence of feral pigs. Ramsey et al., (2009) developed models to estimate the degree of confidence in the success of eradication programmes when monitoring failed to detect any more pigs. These models allowed managers to determine the relationship between detection probability and searching effort through aerial or ground hunting and could be used to explain to managers the risks inherent in decisions that must be taken before declaring an eradication complete. Applying a similar approach, Morrison et al., (2007) reduced the time for eradication and post-eradication monitoring of pigs on Santa Cruz Island from an initial estimate of 6-11 years to approximately 2 years.
With few exceptions, very little research has been conducted to determine what proportion of a feral pig population should be targeted to decrease population size, despite the requirement in several countries for management plans to be submitted to the authorities before the opening of the hunting season. A number of studies (summarised by Keuling et al., 2013) suggested that less than 65% of the summer feral pig population must be harvested to decrease population size and that this proportion should include at least 80% of the piglets and target older females.
In summary, whilst Scotland could opt for any of the options highlighted above, we recommend that the effort, the costs and the result of any action aimed at controlling the impact of feral pigs are documented and quantified to inform and optimise future control plans.
Appropriate population monitoring and evaluation of control are crucial to optimise the mitigation of human-feral pig conflicts. Research could assist in training staff in population monitoring and in identifying aspects that need to be addressed for population management to be effective. The latter include implementing and monitoring effectiveness of trapping and shooting at different population densities, areas and times of the year, analysing feral pig spatial and temporal behaviour before, during and after culling and in response to different methods of culling, testing whether some of the methods illustrated above (such as electric fencing) could be effective in reducing impacts of feral pigs, and identifying optimal integrated management approaches.
References
Andersen, R. & Holthe, V. 2010. Ungulates and their management in Denmark. In: Apollonio, M., Andersen, R. & Putman, R. (eds) European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press, pp. 71-85.
Anderson, S.J. & Stone, C.P. 1993. Snaring to control feral hogs (Sus scrofa) in a remote Hawaiian rain forest. Biological Conservation, 63, 195-202.
Andrzejewski, R. & Jezierski, W. 1978. Management of a wild boar population and its effects on commercial land. Acta Theriologica, 23, 309-339.
Apollonio, M., Andersen, R. & Putman, R. 2010. European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press.
Athreya, V., Odden, M., Linnell, J.D.C. & Karanth, K.U. 2010. Translocation as a tool for mitigating conflict with leopards in human-dominated landscapes of India. Conservation Biology, 25, 133-141.
Barrett, R.H., Goatcher, B.L., Gogan, P.J. & Fitzhugh, E.L. 1988. Removing feral pigs from Annadel State Park. Transations of the Western Section of the Wildlife Society, 24, 47-52.
Barrett, R.H. 1978. The feral hog on the Dye Creek Ranch, California. Hilgardia, 46, 283-355.
Barrios-Garcia, M. & Ballari, S. 2012. Impact of wild boar Sus scrofa in its introduced and native range: A review. Biological Invasions, 14, 2283-2300.
Baubet, E., Ropert-Coudert, Y. & Brandt, S. 2003. Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L.). Wildlife Research, 30, 179-186.
Bieber, C. & Ruf, T. 2005. Population dynamics in wild boar Sus scrofa: ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. Journal of Applied Ecology, 42, 1203-1213.
Bolzoni, L. & De Leo, G. 2007. A cost analysis of alternative culling strategies for the eradication of classical swine fever in wildlife. Environment and Development Economics, 12, 653-671.
Bouldoire, J.-L. & Vassant, J. 1989. Le sanglier. Paris: Hatier.
Bradley, E.H., Pletscher, D.H., Bangs, E.E., Kunkel, K.E., Smith, D.W., Mack, C.M., Meier, T.J., Fontaine, J.A., Neemeyer, C.C. & Jimenez, M.D. 2005. Evaluating wolf translocation as a non-lethal method to reduce livestock conflicts in the northwestern United States. Conservation Biology, 19, 1498-1508.
Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake, J.L., Borchers, D.L. & Thomas, L. 2001. Introduction to Distance Sampling: Estimating Abundance of Biological Populations. Oxford: Oxford University Press.
Bueno, C.G., Barrio, I.C., Garcia-Gonzalez, R., Alados, C.L. & Gomez-Garcia, D. 2010. Does wild boar rooting affect livestock grazing areas in alpine grasslands? European Journal of Wildlife Research, 56, 765-770.
Cahill, S., Limona, F. & Garcia, J. 2003. Spacing and nocturnal activity of wild boar Sus scrofa in a Mediterranean metropolitan park. Wildlife Biology, 9, 13-33.
Cai, J., Jiang, Z., Zeng, Y., Li, C. & Bravery, B.D. 2008. Factors affecting crop damage by wild boar and methods of mitigation in a giant panda reserve. European Journal of Wildlife Research, 54, 723-728.
Calenge, C., Maillard, D., Fournier, P. & Fouque. C. 2004. Efficiency of spreading maize in the garrigues to reduce wild boar damage to Mediterranean vineyards. European Journal of Wildlife Research, 50, 112-120.
Calenge, C., Maillard, D., Vassant, J. & Brandt, S. 2003. Summer and hunting season home ranges of wild boar (Sus scrofa) in two habitats in France. Game & Wildlife Science, 19, 281-301.
Caley P. 1994. Factors affecting the success rate of traps for catching feral hogs in a tropical habitat. Wildlife Research, 21, 287-292.
Campbell, S. & Hartley, G. 2010. Locations of Captive Wild Boar, Wild Boar Hybrids and feral pigs in Scotland. Report to Scottish Government Animal Health and Welfare Division.
Cargnelutti, B., Spitz, F. & Valet, G. 1992. Analysis of the dispersion of wild boar (Sus scrofa) in southern France. In: Ongulés/Ungulates 91. Spitz., F., Janeau, G., Gonzalez, G.& Aulagnier, S. (eds) Proceedings of the International Symposium ‘Ongulés/Ungulates 91’. Toulouse: SFEPM-IRGM, pp. 423-425.
Centner, T. J. & Shuman, R. M. 2014. Governmental Provisions to Manage and Eradicate Feral Swine in Areas of the United States. Ambio, 44, 121-130.
Choquenot, D., McIlroy, J. & Korn, T.1996. Managing vertebrate pests: feral pigs. Canberra: Australian Government Publishing Service.
Christie, J., DeMarco, E., Hiroyasu, E., Kreger, A. & Ludington, M. 2014. Wild Pig Management at Tejon Tanch. Bren School Group Project.
Coblentz, B.E. & Baber, D.W. 1987. Biology and control of feral pigs on Isla Santiago, Galapagos, Ecuador. Journal of Applied Ecology, 24, 403-418.
Conover, M.R. 2002. Resolving human–wildlife conflicts: the science of wildlife damage management. Boca Raton: Lewis Publishers.
Corner, L.A. 2006. The role of wild animal populations in the epidemiology of tuberculosis in domestic animals: How to assess the risk. Veterinary Microbiology, 112, 303-312.
Cowan, D.P. & Massei, G. 2008. Wildlife contraception, individuals and populations: how much fertility control is enough? In: Timm, R.M. & Madon, M.B. (eds) Proceedings of the 23rd Vertebrate Pest Conference. Davis: University of California, pp. 220-228.
Cowled, B.D., Lapidge, S.J. & Elsworth, P. 2008. Additional toxins for feral hog (Sus scrofa) control: identifying and testing Achilles’ heels. Wildlife Research, 35, 651-662.
Cruz, F., Josh, J.C., Campbell, K. & Carron, V. 2005. Conservation action in the Galàpagos: feral hog (Sus scrofa) eradication from Santiago Island. Biological Conservation, 121, 473-478.
Defra, 2008. Feral wild boar in England: An action plan. London: Department for Environment, Food and Rural Affairs.
Defra, 2014. Disease Control Strategy for African and Classical Swine Fever in Great Britain.
Dexter, N. 1996. The Effect of an Intensive Shooting Exercise from a Helicopter on the Behaviour of Surviving Feral Pigs. Wildlife Research, 23, 435-441.
Efron, B. 2000. The Bootstrap and Modern Statistics. Journal of the American Statistical Association, 95, 1293-1296.
Engeman, R.M., Constantin, B.U., Nelson, M., Woolard, J. & Bourassa, J. 2001. Monitoring changes in feral swine population and spatial distribution of activity. Environmental Conservation, 28, 235-240.
Engeman, R.M., Pipas, M.J., Gruver, K.S., Bourassa, J., & Allen, L. 2002. Plot placement when using a passive tracking index to simultaneously monitor multiple species of animals. Wildlife Research, 29, 85-90.
Engeman, R.M., Massei, G., Sage, M., & Gentle, M.N. 2013. Monitoring wild pig populations: a review of methods. Environmental Science and Pollution Research, 20, 8077-8091.
Environment Agency, 2008. Animal By-Products Regulations. Chester: C-Tech Innovations LTD.
Erkinaro, E., Heikura, K., Lindgren, E., Pulliainen, E. & Sulkava, S. 1982. Occurrence and spread of the wild boar (Sus scrofa) in eastern Fennoscandia. Memoranda Societatis pro Fauna et Flora Fennica, 58, 39-47.
Fagertsone, K.A., Coffey, M.A., Curtis, P.D., Dolbeer, R.A., Killian, G.J., Miller, L.A. & Wilmot, L.M. 2002. Wildlife fertility control: technical review. Bethesda: The Wildlife Society.
Fernando, P., Leimgruber, P., Prasad, T. & Pastorini, J. 2012. Problem-elephant translocation: translocating the problem and the elephant? PloS ONE, 7, e50917.
Focardi, S., Capizzi, D. & Monetti, D. 2000. Competition for acorns among wild boar (Sus scrofa) and small mammals in a Mediterranean woodland. Journal of Zoology, 250, 329-334.
Fonseca, C., Alves da Silva, A., Alves, J., Vingada, J. & Soares, A.M.V.M. 2011. Reproductive performance of wild boar females in Portugal. European Journal of Wildlife Research, 57, 363-371.
Fonseca, C., Santos, P., Monzón, A., Bento, P., Alves da Silva, A., Silverio, A., Soares, A.M.V.M. & Petrucci-Fonseca, E. 2004. Reproduction in the wild boar (Sus scrofa Linneaus 1758) populations of Portugal. Galemys, 16, 53-65.
Fordham, D.A., Georges, A. & Brook, B.W. 2007. Demographic response of snake‐necked turtles correlates with indigenous harvest and feral pig predation in tropical northern Australia. Journal of Animal Ecology, 76, 1231-1243.
Forest Research, 2014. Wild boar and deer in the Forest of Dean 2014.
Foyle, K.L., Delahay, R.J. & Massei, G. 2010. Isolation of Mycobacterium bovis from a feral wild boar (Sus scrofa) in the UK. Veterinary Record, 166, 663-664.
Frantz, A.C., Massei, G., & Burke, T. 2012. Genetic evidence for past hybridisation between domestic pigs and English wild boars. Conservation Genetics, 13, 1355-1364.
Franzetti, B., Ronchi, F., Marini, F., Scacco, M., Calmanti, R., Calabrese, A., Aragno, P., Montanaro, P. & Focardi, S. 2012. Nocturnal line transect sampling of wild boar (Sus scrofa) in a Mediterranean forest: long-term comparison with capture–mark–resight population estimates. European Journal of Wildlife Research, 58, 385-402.
Gamelon, M., Besnard, A., Gaillard, J.M., Servanty, S., Baubet, E., Brandt, S. & Gimenez, O. 2011. High hunting pressure selects for earlier birth date: wild boar as a case study. Evolution, 65, 3100-3112.
Gaskamp, J.A., Roberts, S., Gee, K.L., Silvy, N.J. & Campbell, T.A. 2012. Efficacy of Drop-nets versus Corral Traps for Feral Hog Capture. 2012 International Wild Pig Conference San Antonio, Texas.
Geisser, H. & Reyer, H.U. 2005. The influence of food and temperature on population density of wild boar Sus scrofa in the Thurgau (Switzerland). Journal of Zoology, 267, 89-96.
Genov, P.V. 1981. Significance of natural biocenoses and agrocenoses as the source of food for wild boar (Sus scrofa L.). Ekologia polska, 29, 117-136.
Genov, P.V., Massei, G., Barbalova, Z. & Kostova, V. 1992. Aging Wild boar (Sus scrofa L.) by teeth. In: Spitz., F., Janeau, G., Gonzalez, G.& Aulagnier, S. (eds) Proceedings of the International Symposium ‘Ongulés/Ungulates 91’. Toulouse: SFEPM-IRGM, pp. 399-402.
Gerard, J.F. & Campan, R.1988. Variabilité éco-éthologique chez le sanglier européen: comparaison des travaux français. Cahiers d’Ethologie Appliquée, 8, 63-130.
Gethöffer, F., Sodeikat, G. & Pohlmeyer, K. 2007. Reproductive parameters of wild boar (Sus scrofa) in three different parts of Germany. European Journal of Wildlife Research, 53, 287-297.
Gill, R. & Brandt, G. 2010. Thermal imaging to estimate wild boar density. Wrecclesham: Forest Research.
Gill, R. 2014. Wild Boar and Deer in the Forest of Dean. Forest Research, Forestry Commission.
Gipson, P.S., Hlavachick, B. & Berger, T. 1998. Range expansion of feral hogs across the central United States. Wildlife Society Bulletin, 26, 279-286.
Gosling, L.M. & Baker, S.J. 1989. The eradication of muskrats and coypus from Britain. Biological Journal of the Linnean Society, 38, 39-51.
Goulding, M.J. & Smith, G.C. 1998. Current Status and Potential Impact of Wild Boar (Sus scrofa) in the English Countryside: A Risk Assessment. Report to Conservation Management Division C MAFF.
Goulding, M.J., Roper, T.J., Smith, G.C. & Baker, S.J. 2003. Presence of free-living wild boar in southern England. Wildlife Biology, 9, 15-20.
Grice, A.C. 1996. Seed production, dispersal and germination in Cryptostegia grandiflora and Ziziphus mauritiana, two invasive shrubs in tropical woodlands of northern Australia. Australian Journal of Ecology, 21, 324-331.
Groot Bruinderink, G.W.T.A. & Hazebroek, E. 1996. Wild boar (Sus scrofa scrofa L.) rooting and forest regeneration on podzolic soils in the Netherlands. Forest Ecology and Management, 88, 71-80.
Groot Bruinderink, G.W.T.A, Hazebroek, E. & van der Voot, H. 1994. Diet and condition of wild boar, Sus scrofa scrofa, without supplementary feeding. Journal of Zoology London, 233, 631-648.
Guibert, B. 2008. Bilan national de l’indemnisation des degats agricoles de sangliers. In: Klein, F., Guibert, B. & Baubet, E., Modalites de gestion du sanglier, Actes du colloque Reims. Paris: F.N.C. -O.N.C.F.S., pp.73-78.
Häggmark Svensson, T., Gren, I.M., Andersson, H., Jansson, G. & Jägerbrand, A. 2014. Costs of traffic accidents with wild boar populations in Sweden. Working Paper Series 2014:05. Uppsala: Swedish University of Agricultural Sciences.
Hahn, N. & Eisfeld, D. 1998. Diet and habitat use of wild boar (Sus scrofa) in SW-Germany. Gibier Faune Sauvage, 15, 595-606.
Hanson, L.B., Mitchell, M.S., Grand, J.B., Jolley, D.B., Sparklin, B.D. & Ditchkoff, S.S. 2009. Effect of experimental manipulation on survival and recruitment of feral pigs. Wildlife Research, 36, 185-191.
Hartley, M. 2010. Qualitative risk assessment of the role of the feral wild boar (Sus scrofa) in the likelihood of incursion and the impacts of effective disease control of selected exotic diseases in England. European Journal of Wildlife Research, 56, 401-410.
Heinken, T., Schmidt, M., Von Oheimb, G., Kriebitzsch, W.U. & Ellenberg, H. 2006. Soil seed banks near rubbing trees indicate dispersal of plant species into forests by wild boar. Basic Applied Ecology, 7, 31-44.
Holland, E.P., Burrow, J.F., Dytham, C., & Aegerter, J.N. 2009. Modelling with uncertainty: Introducing a probabilistic framework to predict animal population dynamics. Ecological Modelling, 220, 1203-1217.
Hone, J. & Atkinson, B. 1983. Evaluation of fencing to control feral hog movement. Australian Wildlife Research, 10, 499-505.
Hone, J. 1983. A short-term evaluation of feral pig eradication at Willandra in Western New South Wales. Australian Wildlife Research, 10, 269-275.
Hone, J. 2002. Feral pigs in Namadgi National park, Australia: dynamics, impacts and management. Biological Conservation, 105, 231-242.
Hone, J. 1980. Effect of feral pig rooting on introduced and native pasture in North-Eastern New South Wales. Journal of the Australian Institute of Agricultural Science, 46, 130-132.
Howe, T., Singer, J.S. & Ackerman, B.B. 1981. Forage relationships of European wild boar invading northern hardwood forest. Journal of Wildlife Management, 45, 748-754.
Hutton, T., DeLiberto, T., Owen, S. & Morrison, B. 2006. Disease risks associated with increasing feral swine numbers and distribution in the United States. Midwest Association of Fish and Wildlife Agencies.
Ickes, K., Dewalt, S.J. & Appanah, S. 2001. Effects of native pigs (Sus scrofa) on woody understorey vegetation in a Malaysian lowland rain forest. Journal of Tropical Ecology, 17, 191-206.
Imesh-Bebie’, N., Gander, H. & Schnidrig-Petrig, R. 2010. In: Apollonio, M., Andersen, R. & Putman, R. (eds) European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press, pp. 357-391.
Jansen, A., Luge, E., Guerra, B., Wittschen, P., Gruber, A.D., Loddenkemper, C., Schneider, T., Lierz, M., Ehlert, D., Appel, B., Stark, K., & Nöckler. K. 2007. Leptospirosis in urban wild boars, Berlin, Germany. Emerging Infectious Diseases, 13, 739-742.
Jędrzejewska, B., Jędrzejewski, W., Bunevich, A.N., M iłkowski, L. & Krasiński, Z.A. 1997. Factors shaping population densities and increase rates of ungulates in Białowieża Primeval Forest (Poland and Belarus) in the 19th and 20th centuries. Acta Theriologica, 42, 399-451.
Jędrzejewski, W., Jędrzejewska, B., Okarma, H., & Ruprecht. A.L. 1992. Wolf predation and snow cover as mortality factors in the ungulate community of the Bialowieża National Park, Poland. Oecologia, 1, 27-36.
Jerina, K., Pokorny, B. & Stergar, M. 2014. First evidence of long-distance dispersal of adult female wild boar (Sus scrofa) with piglets. European Journal of Wildlife Research, 60, 367-370.
Jullien, J.M., Vassant, J., Delorme, D. & Brandt, S. 1987. Une technique de capture de groupes de sangliers particulierement efficace: le filet tombant. Gibier Faune Sauvage, 4, 203-208.
Kaji, K., Saitoh, T., Uno, H., Matsuda, H. & Yamamura, K. 2010. Adaptive management of sika deer populations in Hokkaido, Japan: theory and practice. Population Ecology, 52, 373-387.
Katahira, L.K., Finnegan, P. & Stone, C.P. 1993. Eradicating feral hogs in montane mesic habitat at Hawaii Volcanoes National Park. Wildlife Society Bulletin, 21, 269–274.
Kessler, C.C. 2002. Eradication of feral goats and pigs and consequences for other biota on Sarigan Island, Commonwealth of the Northern Mariana Islands. In: Veitch, C.R. & Clout, M.N. (eds) Turning the tide: the eradication of invasive species. Gland: IUCN - The World Conservation Union, pp 132-149.
Keuling, O., Baubet, E., Duscher, A., Ebert, C., Fischer, C., Monaco, A., Podgórski, T., Prevot, C., Ronnenberg, K., Sodeikat, G., Stier, N. & Thurfjell. H. 2013. Mortality rates of wild boar Sus scrofa L. in central Europe. European Journal of Wildlife Research, 59, 805-814.
Keuling, O., Stier, N., & Roth, M. 2008. How does hunting influence activity and spatial usage in wild boar Sus scrofa L.? European Journal of Wildlife Research, 54, 729-737.
Killian, G., Eisemann, J.D., Wagner, D., Werner, J., Shaw, D., Engeman, R. & Miller, L.A. 2006. Safety and toxicity evaluation of GonaCon immuno-contraceptive vaccine in white-tailed deer. In: Tirnm, M. & O'Brien, J. M. (eds). Proceedings of the 22nd Vertebrate Pest Conference. Davis: University of California, pp. 82-87.
Killian, G., Fagerstone, K.A., Kreeger, T., Miller, L.A. & Rhyan, J. 2007. Management strategies for addressing wildlife disease transmission: the case for fertility control. In: Nolte, D.L., Arjo, D.H. & Stalman, W.M. (eds) Proceedings of the 12th Wildlife Damage Management Conference, Lincoln: University of Nebraska, pp. 265-271.
Killian, G., Thain, D., Diehl, N.K., Rhyan, J. & Miller, L.A. 2008. Four-year contraception rates of mares treated with single- injection porcine zona pellucida and GnRH vaccines and intrauterine devices. Wildlife Research, 35, 531-539.
Kock, R.A., Woodford, M.H. & Rossiter, P.B. 2010. Disease risks associated with the translocation of wildlife. Revue scientifique et technique (International Office of Epizootics), 29, 329-350.
Kotanen, P.M. 1995. Responses of vegetation to a changing regime of disturbance: effects of feral pigs in a California coastal prairie. Ecography, 18, 190-199.
Leaper, R., Massei, G., Gorman, M.L. & Aspinall, R. 1999. The feasibility of reintroducing wild boar to Scotland. Mammal Review, 29, 239-259.
Liberg, O., Bergstrom, R., Kindberg, J. & Von Essen, H. 2010. Ungulates and their management in Sweden. In: Apollonio, M., Andersen, R. & Putman, R. (eds) European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press, pp. 37-70.
Littauer, G.A. 1993. Control techniques for feral hogs. In: Hanselka, C.W. & Cadenhead, J.F. (eds) Feral swine: a compendium for resource managers. Kerrville: Texas Agriculture Extension Service, pp. 39-148.
Lombardo, C.A. & Faulkner, K.R. 2000. Eradication of feral pigs, (Sus scrofa) from Santa Rosa Island, Channel Islands National Park, California. In: Browne, D.R., Mitchell, K. L. & Chaney, H.W. (eds) Proceedings of 5th California islands symposium. Washington D.C.: U.S. Department of the Interior, pp. 300-306.
Lynes, B.C. & Campbell, S.D. 2000. Germination and viability of mesquite (Prosopis pallida) seed following ingestion and excretion by feral pigs (Sus scrofa). Tropical Grasslands, 34, 125-128.
Machackova, M., Matlova, L., Lamka, J., Smolik, J., Melicharek, I., Hanzlikova, M., Docekal, J., Cvetnic, Z., Nagy, G., Lipiec, M., Ocepek, M. & Pavlik. I. 2003. Wild boar (Sus scrofa) as a possible vector of mycobacterial infections: review of literature and critical analysis of data from Central Europe between 1983 to 2001. Veterinary Medicine, 48, 51-65.
Mackin, R. 1970. Dynamics of Damage by Wild Boar to different agricultural crops. Acta Theriologica, 27, 447-458.
Maillard, D. & Fournier, P. 2004. Timing and synchrony of births in the wild boar (Sus scrofa Linnaeus, 1758) in a Mediterranean habitat: the effect of food availability. Galemys, 16, 67-74.
Maillard, D., Gaillard, J.M., Hewison, M,, Ballon, P., Duncan, P., Loison, A., Toigo, C., Baubet, E., Bonenfant, C., Garel, M., & Saint-Andrieux, C. 2010. Ungulates and their management in France. In: Apollonio, M., Andersen, R. & Putman, R. (eds) European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press, pp. 441-474.
Massei, G. & Toso, S. 1993. Biology and management of wild boar. Bologna: Istituto Nazionale per la Fauna Selvatica.
Massei, G., Genov, P.V. & Staines, B.W. 1996. Diet, food availability and reproduction of wild boar in a Mediterranean coastal area. Acta Theriologica, 41, 307-320.
Massei, G., Genov, P.V., Staines, B.W. & Gorman, M.L. 1997. Mortality of wild boar in a Mediterranean area in relation to sex and age. Journal of Zoology London, 242, 394-400.
Massei, G. & Genov, P.V. 2004. The environmental impact of wild boar. Galemys, 16, 135-145.
Massei, G., Cowan,D.P, Coats,J., Gladwell,F., Lane,J.E. & Miller,L.A. 2008. Effect of the GnRH vaccine GonaConTM on the fertility, physiology and behaviour of wild boar. Wildlife Research, 35, 1-8.
Massei, G., Coats, J., Quy, R., Storer, K., & Cowan, D.P. 2010. The BOS (Boar-Operated-System): a novel method to deliver baits to wild boar. Journal of Wildlife Management, 74, 333-336.
Massei, G., Quy, R., Gurney, J. & Cowan, D.P. 2010. Can translocations be used to manage human-wildlife conflicts? Wildlife Research, 37, 428-439.
Massei, G., Cowan, D.P., Coats,J., Bellamy, F., Quy, R., Brash, M. & Miller, L.A. 2012. Long-term effects of immunocontraception on wild boar fertility, physiology and behaviour. Wildlife Research, 39, 378-385.
Massei, G. & Cowan, P. 2014. Fertility control to mitigate human-wildlife conflicts: a review. Wildlife Research, 41, 1-21.
Massei, G., Kindberg, J., Licoppe, A., Šprem, N., Kamler, J., Baubet, E., Hohmann, U., Monaco, A., Ozoliņš, J., Cellina, S., Podgórski, T., Fonseca, C., Markov, N., Pokorny, B., Rosell, C., & Náhlik, A. 2014. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Management Science DOI 10.1002/ps.3965.
Massei, G., Roy, S. & Bunting, R. 2011. Too many hogs? A review of methods to mitigate impact by wild boar and feral pigs. Human-Wildlife Interactions, 5, 79-99.
Matschke, G.H. 1967. Aging European wild hogs by dentition. Journal of Wildlife Management, 31, 109-113.
Mayer, J.J. & Brisbin, I.L. 2009. Wild pigs: biology, damage, control techniques and management. Aiken: Savannah River National Laboratory.
McCann, B. E., & Garcelon, D.K. 2008. Eradication of feral hogs from Pinnacles National Monument. Journal of Wildlife Management, 72, 1287-1295.
McDevitt, A.D., Carden, R.F, Coscia, I. & Frantz, A.C. 2013. Are wild boars roaming Ireland once more? European Journal of Wildlife Research, 59, 761-764.
McIlroy, J.C. 1983. The sensitivity of Feral Pigs (Sus scrofa) to 1080 and its implications for poisoning campaigns. Australian Wildlife Research, 10, 139-148.
McIlroy, J.C. & Gifford, E.J. 1997. The ‘Judas’ pig technique: a method that could enhance control programmes against feral hogs, Sus scrofa. Wildlife Research, 24, 483-491.
McIlroy, J.C. & Saillard, R.J. 1989. The effect of hunting with dogs on the numbers and movements of feral pigs, Sus scrofa, and the subsequent success of poisoning exercises in Namadgi National Park ACT. Australian Wildlife Research, 16, 353-363.
Meng, X.J., Lindsay, D.S. & Sriranganathan, N. 2009. Wild boars as sources for infectious diseases in livestock and humans. Philosophical Transactions of the Royal Society B Biological Sciences, 364, 2697-2707.
Miller, L.A., Gionfriddo, J., Fagerstone, K.A., Rhyan, J. & Killian, G. 2008. The single-shot GnRH imunocontraceptive vaccine (GonaConTM) in white-tailed deer: comparison of several GnRH preparations. American Journal of Reproductive Immunology, 60, 214-223.
Mitchell, J. 1998. The effectiveness of aerial baiting for control of feral pigs (Sus scrofa) in North Queensland. Wildlife Research, 25, 297-303.
Moore, N. 2004. The ecology and management of wild boar in Southern England. Central Science Laboratory, Defra final project report, VC0325.
Moore, N. & Wilson, C.J. 2005. Feral wild boar in England: Implications of future management options. London: Defra.
Morelle, K., Lehaire, F. & Lejeune, P. 2013. Spatio-temporal patterns of wildlife-vehicle collisions in a region with a high-density road network. Nature Conservation, 5, 53-73.
Morrison, S.A., Macdonald, N., Walker, K., Lozier, L. & Shaw. M.R. 2007. Facing the dilemma at eradication’s end: uncertainty of absences and the Lazarus effect. Frontiers in Ecology and the Environment, 5, 271-276.
Náhlik, A. & Sándor, G. 2003. Birth rate and offspring survival in a free-ranging wild boar Sus scrofa population. Wildlife Biology, 9, 37-42.
Naranjo, V., Gortazar, C., Vicente, J. & de la Fuente, J. 2008. Evidence of the role of European wild boar as a reservoir of Mycobacterium tuberculosis complex. Veterinary microbiology, 127, 1-9.
Nores, C., Llaneza, L. & Álvarez, A. 2008. Wild boar Sus scrofa mortality by hunting and wolf Canis lupus predation: an example in northern Spain. Wildlife Biology, 14, 44-51.
Okarma, H., Jedrzejewska, B., Jederzejewski, W., Krasinski, Z.A. & Milkowski, L. 1995. The roles of predation, snow cover, acorn crop, and man-related factors on ungulate mortality in Bialowieza Primeval Forest, Poland. Acta Theriologica, 40, 197-217.
Parkes, J. & Barron, M. 2012. Data collection requirements to allow analysis and assessment of ungulate control. Whāngārei: Landcare Research Report.
Parkes, J.P., Ramsey, D.S., Macdonald, N., Walker, K., McKnight, S., Cohen, B.S. & Morrison, S.A. 2010. Rapid eradication of feral hogs (Sus scrofa) from Santa Cruz Island, California. Biological Conservation, 143, 634-41.
Pfaff, E., & Saint Andrieux, C. 2007. Le développement du sanglier en France. In: Colloque sur les modalités de gestion du sanglier, Reims: Fédération Nationale des Chasseurs, pp. 3-11.
Pimentel, D., Lach, L., Zuniga, R. & Morrison, D. 2002. Environmental and economic costs associated with nonindigenous species in the United States. In: Pimentel, D. (ed) Biological Invasions: Economic and environmental costs of alien plant, animal, and microbe species. Boca Raton: CRC Press, pp. 285-303.
Prevot, C. & Licoppe, A. 2013. Comparing red deer (Cervus elaphus L.) and wild boar (Sus scrofa L.) dispersal patterns in southern Belgium. European Journal of Wildlife Research, 59, 795-803.
Quy,R. J., Massei, G., Lambert, M.S., Coats, J., Miller, L.A. & Cowan,D.P. 2014. Effects of a GnRH vaccine on the movement and activity of free-living wild boar (Sus scrofa). Wildlife Research, 41, 185-193.
Ramsey, D. S., Parkes, J., & Morrison, S. A. 2009. Quantifying eradication success: the removal of feral pigs from Santa Cruz Island, California. Conservation Biology, 23, 449-459.
Reeser, D. & Harry, B. 2005. Controlling ungulate populations in native ecosystems in Hawaii. Hawaii Conservation Alliance: 1-7.
Reddiex, B., Forsyth, D.M., McDonald-Madden, E., Einoder, L.D., Griffioen, P.A., Chick, R.R., & Robley, A.J. 2007. Control of pest mammals for biodiversity protection in Australia. I. Patterns of control and monitoring. Wildlife Research, 33, 691-709.
Reidy, M.M., Campbell, T.A. & Hewitt, D.G. 2008. Evaluation of electric fencing to inhibit feral pig movements. Journal of Wildlife Management, 72, 1012-1018.
Riley, S.J., Decker, D.J,, Enck, J.W., Curtis, P,D, & Lauber. T.B. 2003. Deer populations up, hunter populations down: implications of interdependence of deer and hunter population dynamics on management. Ecoscience, 10, 455-461.
Rosell, C., Fernández- Bou, M., Camps, F., Boronat, C., Navàs, F., Martínez, M. & Sorolla, A. 2013. Animal-Vehicle Collisions: A New Cooperative Strategy is Needed to Reduce the Conflict. Proc. ICOET 2013 International Conference on Ecology and Transportation. Scottsdale, Arizona.
Rossi, S., Toigo, C., Hars, J., Pol, F., Hamann, J.L., Depner, K. & Le Potier, M.-F. 2011. New insights on the management of wildlife diseases using multi-state recapture models: the case of classical swine fever in wild boar. PloS ONE, 6, e24257.
Rosvold, J. & Andersen, R. 2008. Wild boar in Norway – is climate a limiting factor? – NTNU Vitenskapsmuseet Zoologisk rapport, 1, 1-23.
Rovero, F. & Marshall, A.R. 2009. Camera trapping photographic rate as an index of density in forest ungulates. Journal of Applied Ecology, 46, 1011-1017.
Rowcliffe, J.M., Field, J., Turvey, S.T. & Carbone, C. 2008. Estimating animal density using camera traps without the need for individual recognition. Journal of Applied Ecology, 45, 1228-1236.
Sáez-Royuela, C. & Tellería, J.L. 1986. The increased population of the Wild Boar (Sus scrofa L.) in Europe. Mammal Review, 16, 97-101.
Saunders, G., Kay, B. & Nicol, H. 1993. Factors affecting bait uptake and trapping success for feral pigs (Sus scrofa) in Koscuiska National Park. Wildlife Research, 20, 653-665.
Saunders, G. & Bryant, H. 1988. The evaluation of a feral pig eradication program during a simulated exotic disease outbreak. Australian Wildlife Research, 15, 73-81.
Schlageter, A. & Haag-Wackernagel, D. 2012. A gustatory repellent for protection of agricultural land from wild boar damage: an investigation on effectiveness. Journal of Agricultural Science, 4, 61-68.
Schley, L. & Roper, T.J. 2003. Diet of wild boar Sus scrofa in Western Europe, with particular reference to consumption of agricultural crops. Mammal Review, 33, 43-56.
Schley, L., Dufrêne, M., Krier, A. & Frantz, A.C. 2008. Patterns of crop damage by wild boar (Sus scrofa) in Luxembourg over a 10-year period. European Journal of Wildlife Research, 54, 589-599.
Servanty, S., Gaillard, J.-M., Ronchi, F., Focardi, S., Baubet, É. & Gimenez, O. 2011. Influence of harvesting pressure on demographic tactics: implications for wildlife management. Journal of Applied Ecology, 48, 835-843.
Servanty, S., Gaillard, J.-M., Toïgo, C., Brandt, S. & Baubet, E. 2009. Pulsed resources and climate-induced variation in the reproductive traits of wild boar under high hunting pressure. Journal of Animal Ecology, 78, 1278-1290.
Siemann, E., Carrillo, J.A., Gabler, C.A., Zipp, R. & Rogers, W.E. 2009. Experimental test of the impacts of feral hogs on forest dynamics and processes in the southeastern US. Forest Ecology and Management, 258, 546-553.
Sims, N.K., John, E.A. & Stewart, A.J. 2014. Short-term response and recovery of bluebells (Hyacinthoides non-scripta) after rooting by wild boar (Sus scrofa). Plant Ecology, 215, 1409-1416.
Singer, F.J., Otto, D.K., Tipton, A.R. & Hable, C.P. 1981. Home ranges, movements, and habitat use of the European Wild Boar in Tennessee. Journal of Wildlife Management, 45, 343–353.
Singer, F.J., Swank, W.T., & Clebsch, E.E. 1984. Effects of wild pig rooting in a deciduous forest. Journal of Wildlife Management, 48, 464-473.
Sjarmidi, A. & Gerard, J. 1988. Autour de la systematique et la distribution des suidés. Monitore Zoologico Italiano, 22, 415-448.
SNH, 2014. Wild boar in Scotland: Scottish Government’s draft position statement. SNH.
Sodeikat, V. & Pohlmeyer, R. 2002. Temporary home range modifications of wild boar family groups (Sus scrofa L.) caused by drive hunts in Lower Saxony (Germany). Zeitschrift fur. Jagdwissenschaft, 8, 161-166.
Spencer, P.B.S. & Hampton, J.O. 2005. Illegal translocation and genetic structure of feral hogs in western Australia. Journal of Wildlife Management, 69, 377-384.
Spitz, F. 1986. Current state of knowledge of wild boar biology. Pig News and Information, 7 171-175.
State of Hawaii, 2007. Review on methods and approaches for control of non-native ungulates in Hawaii. State of Hawaii Department of Land and Natural Resources Technical Report 07-01. Honolulu: Hawaii.
Sweitzer, R.A., Gonzales, B.J., Gardner, I. A., Vuren, D.V., Wauthman, J.D. & Boyce, W.M. 1997. A modified panel trap and immobilization technique for capturing multiple feral hogs. Wildlife Society Bulletin, 25, 699-705.
Sweitzer, R.A., Van Vuren, D., Gardner, I.A., Boyce, W.M. & Waithman, J.D. 2000. Estimating sizes of wild pig populations in the North and Central Coast regions of California. Journal of Wildlife Management, 64, 531-543.
The Wildlife Society, 2012. Annual State Summary Report – Wild Hog Working Group.
Thurfjell, H., Spong, G. & Ericsson, G. 2013. Effects of hunting on wild boar Sus scrofa behavior. Wildlife Biology, 19, 87-93.
Toıgo, C., Servanty, S., Gaillard, J.-M., Brandt, S. & Baubet, E. 2008. Disentangling natural from hunting mortality in an intensively hunted wild boar population. Journal of Wildlife Management, 72, 1532-1539.
Tuyttens, F.A.M., Macdonald, D.W., Delahay, R., Rogers, L.M., Mallinson, P.J., Donnelly, C.A. & Newman, C. 1999. Differences in trappability of European badgers Meles meles in three populations in England. Journal of Applied Ecology, 36, 1051-1062.
Twigg, L.E., Lowe, T.J., Martin, G.R. & Everett, M. 2005. Feral hogs in north-western Australia: basic biology, bait consumption, and the efficacy of 1080 baits. Wildlife Research, 32, 281-296.
USDA-APHIS, 2012. Feral Swine Management Report. Wildlife Services New York, pp. 24.
Van Wieren, S. E. & Groot-Bruinderink, W.T.A. 2010. Ungulates and their management in the Netherlands. In: Apollonio, M., Andersen, R. & Putman, R. (eds) European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press, pp. 265-183.
Vassant, J. & Boisaubert, B. 1984. Bilan des expérimentations enterprises en Haut-Marne pour réduire les dégâts de sangliers (Sus scrofa) à l’encontre des cultures agricoles. In: Symposium International sur le sanglier. Spitz, F. & Pépin, D. (eds). Toulouse SFEPM-IRGM, pp. 187-199.
Vassant, J., Jullien, J.M. & Brandt, S. 1987. Réduction des dégâts de sangliers sur blé et avoine en été. Etude de l’efficacité de l’épandage de maïs grain en forêt. Bulletin mensuel de l'Office national de la chasse, 113, 23-34.
Vassant, J. 1994. Les techniques de prévention des dégâts de sanglier. Office National de la Chasse, Paris, France. Bulletin mensuel de l'Office national de la chasse, 191, 90-93.
Veeroja, R & Männil, P. 2014. Population development and reproduction of wild boar (Sus scrofa) in Estonia. Wildlife Biology in Practice, 10, 17-21.
Vidrih, M. & Trdan, S. 2008. Evaluation of different designs of temporary electric fence systems for the protection of maize against wild boar (Sus scrofa L., Mammalia, Suidae). Acta Agriculturae Slovenica, 91, 343-349.
Vilardell, A., Capalleras, X., Budó, J., Molist, F. & Pons, P. 2008. Test of the efficacy of two chemical repellents in the control of Hermann’s tortoise nest predation. European Journal of Wildlife Research, 54, 745-748.
Vtorov, I.P. 1993. Feral pig removal: effects on soil microarthropods in a Hawaiian rain forest. Journal of Wildlife Management, 57, 875-880.
Waithman, J.D., Sweitzer, R.A., Van Vuren, D., Drew, J.D., Brinkhaus, A.J., Gardner, I.A. & Boyce, W.M. 1999. Range expansion, population sizes, and management of feral hogs in California. Journal of Wildlife Management, 63, 298-308.
Welander, J. 2000. Spatial and temporal dynamics of wild boar (Sus scrofa) rooting in a mosaic landscape. Journal of Zoology London, 252, 263-271.
West, B.C., Cooper, A.L. & Armstrong, J.B. 2009. Managing feral hogs: a technical guide. Human–Wildlife Interactions Monograph 1. Starkville: Berryman Institute.
Wilcox, J.T., Aschehoug, E.T., Scott ,C.A. & Van Vuren, D.H. 2004. A test of the Judas technique as a method for eradicating feral hogs. Transactions of the Western Section of the Wildlife Society, 40, 120-126.
Wilson, C. J. 2003. Distribution and status of feral wild boar in Dorset, southern England. Mammal Review, 33, 302-307.
Wilson, C. J. 2005. Feral wild boar in England: status, impact and management. London: Defra.
Wilson, C.J. 2014. The Establishment and Distribution of Feral Wild Boar (Sus scrofa L.) in England. Wildlife Biology in Practice, 10, 1-6.
Wirthner, S., Schütz, M., Page-Dumroese, D.S., Busse, M.D., Kirchner, J.W. & Risch, A.C. 2012. Do changes in soil properties after rooting by wild boars (Sus scrofa) affect understory vegetation in Swiss hardwood forests? Canadian Journal of Forest Research, 42, 585-592.
Wotschikowsky, U. 2010. Ungulates and their management in Germany. In: Apollonio, M., Andersen, R. & Putman, R. (eds) European Ungulates and their Management in the 21st Century. Cambridge: Cambridge University Press, pp. 201-222.
Yalden, D. 1999. The History of British Mammals. London: Poyser/Academic Press.
Appendix 1 - Further information
Further Information
- Natural England, advice on wild boar
- Wild boar action plan
- General advice on animal health
- Legislation on line
- Firearms guidance
- Wild Game Guide
- Wild Boar website includes the following guidelines:
- Guidance for land managers on the impacts of wild boar and their management
- Guidance on welfare such as minimum recommended firearm calibres
- Guidance on best practice and safe shooting
- Guidance on carcass handling including meat for human consumption and waste disposal
- Advice to aid hunters, gamekeepers and stalkers in disease identification
- Public awareness of wild boar including safety advice
- Advice on dealing with wounded wild boar
- Advice for keepers of wild boar and Local Authorities to minimise the risk of further escapes
Disclaimer: Scottish Natural Heritage (SNH) has changed its name to NatureScot as of the 24th August 2020.
At the time of publishing, this document may still refer to Scottish Natural Heritage (SNH) and include the original branding. It may also contain broken links to the old domain.
If you have any issues accessing this document please contact us via our feedback form.






