NatureScot Research Report 1346 - Loch Creran Special Area of Conservation (SAC) review of activities and pressures
Year of publication: 2024
Authors: Hughes, D.J., Ross, O., Carey, R., McNeill, C., and O’Dell, A. (SAMS Enterprise)
Cite as: Hughes, D.J., Ross, O., Carey, R., McNeill, C., and O’Dell, A. 2024. Loch Creran Special Area of Conservation (SAC) review of activities and pressures. NatureScot Research Report 1346.
Contents
- Keywords
- Background
- Executive summary
- Key knowledge gaps
- Glossary and key terms
- Abbreviations
- Literature review: Serpulid reef declines and regeneration, Loch Creran and beyond
-
A timeline of environmental change and human activities in Loch Creran, 1980-2022
- Introduction
- West Scotland climatic data
- Physical characteristics of Loch Creran
- Loch Creran phytoplankton
- Human activities
- Summary
-
Screening assessment
- Introduction
- West Scotland climatic data
- Phytoplankton
- Human activities
- Assessment of activities, pressures and knowledge gaps
Keywords
Loch Creran; Serpulid reef; Decline; Regeneration; Activities; Pressures; Knowledge gaps
Background
The Loch Creran Special Area of Conservation (SAC) was established in 2005 under the EC Habitats Directive (92.43 EEC) due to its reefs, which include biogenic reefs constructed by Serpula vermicularis tubeworms and horse mussels. Loch Creran contains the largest extent of serpulid reefs in the world however, there have been reports in recent years detailing a significant decline.
This review summarises available information on the dynamics of serpulid polychaete reefs, covering observed case studies of reef decline and regeneration and the underlying processes of recruitment, growth and mortality, with a main focus on S. vermicularis in Loch Creran. This is followed by an examination of the current available data on environmental changes and human activities between 1980 and 2022. A screening assessment was used to assess the relevance and quality of the data and whether changes in and/or around the Loch coincide with observed changes in serpulid reef condition. The final phase of this review assesses the findings of the report and attempts to provide a greater understanding of whether the activities may be contributing to the observed decline of serpulid reefs and identify any knowledge gaps which currently exist in understanding the decline.
The SAC in Loch Creran was designated on the 17th of March 2005. To protect (avoid deterioration of) the designated feature ‘Reefs’ it was determined that the extent, distribution, structure and function, and processes that support the habitat be maintained in the long term. The SAC designation also aims to maintain the distribution of typical species within the habitat, viability of typical species as components of the habitat and to ensure no significant disturbance of typical species within the habitat occurs. The latest assessed condition for the Loch Creran SAC remains ‘Unfavourable Declining’ as of 1st of August 2018.
Executive summary
- A detailed review of the literature reveals that the serpulid reefs of Loch Creran were in a healthy condition until at least 2005, with visible deterioration in the condition of certain localised reefs apparent from 2011. The most recent 2019 survey shows the decline has extended throughout most of the Loch, leading to a ~20% loss of serpulid reef habitat.
- To try and understand the cause(s) of decline of the reefs in Loch Creran, relevant, available data were gathered and assessed. These included environmental, physical, biological, chemical and human activity data (including some personal communications, where relevant) which were considered important due to their potential to affect the status of the SAC.
- A screening assessment of 15 factors was carried out to determine if temporal changes in reef condition could be detected from available data and any plausible causal link(s) could be made between any parameter(s) and the decline of the serpulid reefs.
- A detailed timeline of environmental and human activitiy changes in Loch Creran over the period of 1980-2022 is hindered by the incompleteness of the available data. Data for some potentially important parameters are only semi-quantitative and are based on a small number of measurements taken at widely separated intervals.
- Available data are inadequate to determine the cause of the decline of reefs in Loch Creran. However, it was not possible to rule out certain factors that were investigated or their cumulative impacts. Because the data relate to past processes, they cannot now be recovered. Consequently, it is unlikely that the process of reef decline since (approximately) 2011 can be resolved or understood in any greater detail than it is now.
- The hypothesis that the observed reef decline is a natural process, reflecting a decadal-scale cycle of development and senescence, is not disproven by any of the available evidence.
- There is no convincing support for any hypothesis attributing reef decline to human activities in the Loch, but in the absence of robust evidence the possibility that such a link exists cannot be ruled out.
Key knowledge gaps
- There is a lack of information surrounding the biology and ecology of the species S. vermicularis. For example, the recruitment and reproductive patterns, food preference and dietary requirements (i.e., optimum density/type of phytoplankton) and broader environmental conditions needed to maintain S. vermicularis reefs are largely unknown. Where possible, comparisons with other species within the Family Serpulidae may be inferred but species-specific information could be improved.
- A nationwide analysis of S. vermicularis is presented which suggests that decline is occurring across other regions in Scotland. Improved data on the state and occurrence of these other aggregations are needed, to show if the pattern of decline is unique to Loch Creran or is occurring on a nationwide scale. An improved understanding of these dynamics will allow testing of certain hypotheses such as those that suggest decline and regeneration occur on regular temporal scales as part of the natural cycles of this species, or if there is some causal factor such as anoxia (as documented in an Irish case study of S. vermicularis decline).
- The latest condition assessment of Loch Creran’s SAC was in 2018. We note that the ‘unfavourable declining’ status of S. vermicularis in Loch Creran requires updating. The programme of six-year intervals between surveys may be insufficient in the circumstances.
- Recommendations for future actions or studies that will improve our knowledge and understanding of the biology and ecology of S. vermicularis are included.
Glossary and key terms
Reefs – Implies an ecosystem, or a ridge or shoal of rock lying beneath the surface of a natural body of water. The feature designation for Loch Creran SAC is listed as ‘Reefs’ (as identified in Annex 1 of the Habitats Directive). Annex I reefs are defined as rocky areas or concretions made by marine animals that arise from the surrounding seafloor. The Annex 1 definition of reef will in many instances incorporate serpulid ‘aggregations’ as defined below.
Aggregations – A measure of the degree to which individuals of the same type are spatially clumped. While the designated feature for Loch Creran relates to the Annex 1 habitat, ‘Reefs’, elsewhere, the presence of S. vermicularis are defined as ‘Aggregations’ (for example the Loch Sunart Marine Protected Area (MPA)). Aggregations, therefore, do not always result in the formation of reefs but where they are dense enough, aggregations of S. vermicularis may also be defined as reefs.
Pressure – Force acting upon the marine environment, for example smothering of seabed habitats and species, climate change or other forces that might cause change.
Impact – Here we use impact as a marked effect or an observed environmental change, generally which is caused by a pressure as defined above.
Unfavourable Declining – The condition of the SAC feature (S. vermicularis) is assessed by monitoring and assigned to one of seven categories set by NatureScot (formerly SNH, 2010, see also Moore et al., 2020):
- Favourable Maintained – the attribute targets set for the natural features have been met, and the natural feature is likely to be secure on the site under present conditions.
- Favourable Recovered – the condition of the natural feature has recovered from a previous unfavourable condition, and attribute targets are now being met.
- Unfavourable Recovering – one or more of the attribute targets have not been met on the site, but management measures are in place to improve the condition.
- Unfavourable No Change – one or more of the attribute targets have not been met, and recovery is unlikely under the present management or other activity on the site.
- Unfavourable Declining – one or more of the attribute targets have not been met, evidence suggests that condition will worsen unless remedial action is taken.
- Partially Destroyed – something has happened on the site which has removed part of the natural features, there is no prospect of restoring the destroyed area.
- Totally Destroyed – the natural feature is no longer present, there is no prospect of restoring it.
Abbreviations
- AIS – Automatic Identification System
- AZ – Azamethiphos
- BCD – Buoyancy Control Device
- CAR – Controlled Activities Regulations
- NMPi – National Marine Plan interactive
- SACFOR – Superabundant, Abundant, Common, Frequent, Occasional, Rare
- SEPA – Scottish Environment Protection Agency
- SPRI – Scottish Pollutant Release Inventory
- BOD – Biochemical Oxygen Demand
- BSAC – British Sub Aqua Club
- CES – Crown Estate Scotland
- CMEP – Crustacean Mortality Expert Panel
- DIN – Dissolved Inorganic Nitrogen
- DO – Dissolved Oxygen
- DSP – Diarrhetic Shellfish Poisoning
- EC – European Commission
- EMBZ – Emamectin Benzoate
- EQS – Environmental Quality Standard
- EU – European Union
- FLS – Forestry and Land Scotland
- FSA – Food Standards Agency
- FSS – Food Standards Scotland
- HAB – Harmful Algal Bloom
- JNCC – Joint Nature Conservation Committee
- LC50 – Lethal Concentration
- LD50 – Lethal Dose
- MMO – Marine Management Organisation
- MNCR – Marine Nature Conservation Review
- MPA – Marine Protected Area
- MRC – Marine Resource Centre
- NAUI – National Association of Underwater Instructors
- NFI – National Forestry Inventory
- NOAA – National Oceanic and Atmospheric Administration
- OC – Official Control
- PADI – Professional Association of Diving Instructors
- PCI – Phytoplankton Community Index
- PEPFAA – Prevention of Environmental Pollution from Agricultural Activity
- PSP – Paralytic Shellfish Poisoning
- PSU – Practical Salinity Unit
- SAA – Sub Aqua Association
- SAC – Special Areas of Conservation
- SAMS – Scottish Association for Marine Science
- SAMS Enterprise – SAMS Applied Marine Science Enterprise Ltd
- SCM – Site Condition Monitoring
- SNH – Scottish Natural Heritage
- SS – Suspended Solids
- SSAC – Scottish Sub Aqua Club
- SSF – Scottish Sea Farms
- SSWD – Sea Star Wasting Disease
- SSWS – Sea Star Wasting Syndrome
- TOC – Total Organic Carbon
Literature review: Serpulid reef declines and regeneration, Loch Creran and beyond
Introduction
This review summarises available information on the dynamics of serpulid polychaete reefs, covering observed case studies of reef decline and regeneration and the underlying processes of recruitment, growth and mortality. The main focus of the review is Serpula vermicularis in Loch Creran and the small number of other localities where reef formation by this species has been recorded. Data has been collected from peer-reviewed scientific literature and reports to relevant agencies and supplemented by unpublished personal notes by one of the authors (D.J. Hughes). Relevant information about other reef-building serpulids is summarised where relevant.
Serpula vermicularis
Loch Creran
Status of reefs in Loch Creran
The first record of serpulid reefs in Loch Creran dates from 1887 (Smith, 1887). The significance of this early report is uncertain as the next observations were made over a century later and it is not known whether reefs were continuously present in the Loch throughout the intervening period. The first scientific observations were made in 1989 when Loch Creran was visited as part of the Marine Nature Conservation Review (MNCR), a major field survey of the habitats and biological communities of Scotland’s sea lochs (Connor et al., 1990, Howson et al., 1994). The first detailed mapping of reef distribution and abundance was conducted in November/December 1994 by Heriot-Watt University, Edinburgh (Moore 1996; Moore et al., 1998). During 50 transects surveyed by scuba divers, reefs were recorded around the Loch periphery in the depth range of 1 – 14 m, with occurrence peaking at 6 – 10 m. Reef abundance was greatest along the south shore of the main Loch basin, in a band extending roughly from Sgeir Caillich to Barcaldine, with lighter coverage along the north shore. East of the Creagan Narrows, the upper Loch basin was also fringed with reefs, although abundance was generally low. Mapping studies continued through the later 1990s and into the 2000s, using sidescan sonar and video transects to refine monitoring methods and provide further details of reef distribution. This work also documented areas of localised damage caused by boat moorings and towed fishing gear (Moore et al., 2003, 2009).
In March 2005 Loch Creran was designated a Marine SAC on the basis of its reefs (rocky reefs and biogenic reefs of S. vermicularis and horse mussels, Modiolus modiolus). The SAC designation was followed by a mandatory Site Condition Monitoring (SCM) survey to establish a baseline for future assessment of reef condition and allow detection of any changes in the status of key features. The survey was carried out by Heriot-Watt University, using techniques developed in their previous work, and involved 110 diver transects in July 2005, supplemented by side-scan sonar and video ground-truthing in September 2005 (Moore et al., 2006). Results were broadly similar to those obtained in 1994, with observed differences mostly attributable to the increased sampling density. Reefs were recorded along 66% of the diver transects, with seabed coverage in the range 3 – 17% at the four sites studied in most detail. The total area of the reef band around the Loch periphery was estimated at 108 hectares (ha). Comparisons with previous data showed no significant reduction in reef area over the period 2000 – 2005, and there was some evidence of reef regeneration in the vicinity of the former Barcaldine alginate factory (see the section Reef regeneration in Loch Creran). Tube occupancy was also recorded during this survey, with an overall high occupancy rate across all sites at greater than 50% (Moore et al., 2020). The survey confirmed that dredging, boat moorings, and seabed disturbance around salmon and mussel farms remained a concern, but outside these areas of localised damage, Loch Creran’s serpulid reefs appeared to be in a healthy condition in the mid-2000s, an assessment supported by video monitoring of reefs in July/August 2003 (Poloczanska et al., 2004) and by a transplantation experiment which ran from July 2004 to July 2005 (Hughes et al., 2008). Both of these projects were conducted in the vicinity of the former Sea Life Centre, a site chosen because of the large size and dense coverage of the local reefs.
Loch Creran was designated a MPA in 2014, with the same extent as the existing Marine SAC. MPA status was awarded to afford protection to the Loch’s flame shell (Limaria hians) beds which were not covered by the existing SAC designation. Observations by divers from Heriot-Watt University in 2013 - 2014 indicated a marked deterioration in the state of serpulid reefs at several localities (Moore et al., 2020). Tube occupancy had significantly decreased from observations during the previous survey to between 1 and 10% tube occupation, compared with greater than 50% during the 2005 survey. A second SCM survey was therefore carried out between June 2017 and January 2019 to quantify the extent of these changes. Forty transects were surveyed by divers, with detailed comparisons made at four sites previously visited in 2005. The results confirmed a significant deterioration in reef condition, expressed as widespread fragmentation, reduction in canopy height and lower frequency of tube occupation. The areal loss of habitat was estimated as ~20% (Moore et al., 2020). Severe degradation was documented in all the principal areas formerly supporting a rich growth of reefs (Sea Life Centre Bay, Rubha Mór, South Shian, South Creagan Bay) with the exception of Rubha Riabhach Bay, where reef habitat condition was similar to that recorded in 2005. Using the SAC Site Attribute criteria for reefs, the status of serpulid reef habitat in Loch Creran was categorised as “Unfavourably Declining” (Moore et al., 2020). Moore et al., (2020) noted that widespread deterioration throughout much of the Loch suggested a process driven by natural collapse of reef structures rather than by anthropogenic physical disturbance, which is localised and clearly recognizable (Moore et al., 2003, 2009).
Although reef degradation was first noted by Heriot-Watt divers in 2013, Moore et al., (2020) suggest that the process may have been underway for several years. Some supporting evidence is provided by the following observations (Table 1), made by one of the authors of this report (D. J. Hughes) on recreational dives between March 2008 and September 2017. The dive site was on the south shore of the Loch (approximate position 56.513 N, 05.373 W), a locality used for the experiments described in Hughes et al., (2008) and Hughes (2011), and corresponding roughly to Area F shown in Figure. 2 of Moore et al., (2020). Observations of reef condition are reproduced exactly as recorded in the logbook shortly after each dive. These are casual notes from recreational dives but made by an experienced reef researcher very familiar with the locality. Observations suggest that reef condition at this site was visibly deteriorating from July 2011, and possibly as early as March 2008.
| Date | Observations |
|---|---|
| 16 March 2008 | Swam around for a bit over the reefs. Seemed to be a lot of colonies that had fallen over, but hard to be sure if there were really more than usual. |
| 30 July 2011 | Serpula reefs looked pretty broken up and overgrown. |
| 26 May 2013 | Lots of algal fouling, all looked a bit overgrown. |
| 24 August 2014 | Serpulid aggregations looked a bit shabby and fragmented. |
| 6 April 2015 | Scrappy serpulid colonies |
| 19 September 2015 | Reefs quite broken up and overgrown, not in great shape. |
| 2 September 2017 | Tubeworm reefs in poor shape – collapsed and heavily overgrown, but still with living worms visible. |
| 17 September 2017 | Tubeworm reefs looking very decrepit. |
Schuler (2021) provides an update on the situation described by Moore et al., (2020). Observations along ten transects surveyed in May 2017 and February 2020 showed a decline in free-standing reefs from ~50% to ~24%, along with a shift towards smaller size classes of reef. Synchronous collapse of reefs was recorded along all ten transects. Tube occupancy by living worms also declined between the two surveys, although this was found to be independent of reef collapse and fragmentation, i.e., the number of living worms was reduced in both free-standing and fallen reefs. The process is well-illustrated by the photographs in Schuler’s (2021) report, which leave no doubt that the condition of serpulid reefs of Loch Creran continues to deteriorate.
Recruitment, growth and mortality in Loch Creran
The status and condition of serpulid reefs in Loch Creran is determined by the rates of the underlying demographic processes of recruitment, growth and mortality. Unfortunately, the data on all three factors are limited and derived from short-term experiments rather from than a systematic monitoring programme.
Chapman et al., (2007) measured the recruitment of S. vermicularis onto a variety of substrata deployed in Loch Creran. Larval settlement was recorded from mid-June to mid-October, peaking in late August/early September. Recruitment was higher on slate substrate than on scallop shell. When settling on scallop shells, larvae preferred vertically oriented shells or the underside of shells placed horizontally. Recruitment to the upper surface of horizontal shells was very low. This preference suggests that a response to light may be involved in substrate selection. Cook et al., (2021) deployed tiles at six localities in Loch Creran at different times of year to measure spatial and temporal variation in recruitment of S. vermicularis. Tiles deployed in July showed the highest settlement. Tiles placed at sites without reefs attracted on average 3.6 times more recruits than those placed among existing reefs, a finding interpreted as suggesting that larval supply away from existing reefs is not a limiting factor in reef development in Loch Creran.
Chapman et al., (2007) found no evidence of enhanced recruitment onto tubes of S. vermicularis, whether occupied or unoccupied. In a five-year field experiment, Hughes (2011) did record recruitment of juvenile worms onto reef fragments attached to panels placed horizontally on the Loch seabed. Settlement was much higher on fragments enclosed within plastic mesh cages (intended to exclude grazing urchins) than on fragments open to the environment. The difference in recruitment between caged and open fragments was highest on panels left in situ for three years. However, fragments left for five years showed no effect, with few recruits surviving on either treatment. Moore et al., (2020) report the results of another five-year field experiment using experimental substrata originally deployed in March 2012 (Cook, 2016). Successful settlement and growth leading to incipient reef formation was recorded on both cobbles and scallop shells when these substrata were enclosed in large mesh bags, whereas individual boulders or cobbles/shells deployed in open piles showed little or no serpulid growth. Reviewing the available data on substratum preference and recruitment, Moore et al., (2020) concluded that dead, broken reef material provides a poor surface for colonisation. Reef growth on cobbles and shells in mesh bags may indicate that pressures arising from benthic predators, such as urchins and sea stars, can have a significant effect on recruitment. Bagged cobbles/shells will offer a matrix of complex, rugose surfaces shielded from larger predators, and this may enhance the survival of vulnerable early recruits.
Hughes et al., (2008) recorded a mean tube extension rate of 3.3 cm y-1 for S. vermicularis monitored for one year in Loch Creran, with individual extension rates as high as 8.1 cm y-1. The mean rate agrees with an estimate of ~3.0 cm y-1 by Moore et al., (2020). Occupied tubes deployed in Loch Creran for one year gave a confirmed annual mortality rate of 5.3%, although this should be considered a minimum estimate as some tubes were not recovered at the end of the experiment (Hughes et al., 2008). Little is known of individual lifespan in serpulid worms but species comparable in size to S. vermicularis, such as Spirobranchus polycerus, Ficopomatus enigmaticus or Spirobranchus giganteus, may live for several decades (Kupriyanova et al., 2001). In New Zealand, the reef-building serpulids Galeolaria hystrix and Spirobranchus cariniferus were estimated to live for 10 -12 years (Riedi & Smith 2015).
Reef regeneration in Loch Creran
There has been no detailed study of natural reef regeneration in Loch Creran but studies of growth on experimentally deployed substrata (Cook, 2016; Harries & Moore, 2022) provide some relevant information, which can be supplemented by observations made by other studies. Moore et al., (2020) noted that the first scientific observations of serpulid reefs in Loch Creran (Connor, 1990) mentioned relatively small-sized structures (diameter <20 cm). At the same localities in 1994 (Moore, 1996) and 2005 (Moore et al., 2006) considerably larger reefs (diameter >28 cm) were common. Moore et al., (2020) considered this as evidence for a trend of increasing reef size over the period 1989-2005. Unfortunately, due to the absence of data before 1989 it is not clear whether the first observations of the biogenic reefs were arising de novo or regenerating from a degraded condition.
In 1994, reefs were observed to be absent from a ~1 km stretch of the Loch Creran south shore in an area affected by organic waste discharge from the Barcaldine alginate factory (Moore, 1996; Moore et al., 1998). In 2005, nine years after the closure of the factory, reefs were found along four transects passing through this area, including “moderate numbers of medium and large reefs close to the outfall pipe” (Moore et al., 2006). This demonstrates a degree of recovery following the end of waste discharge. The reefs were found growing on cobbles and boulders on a seabed which consisted mostly of organic-rich mud. Given that urchins and sea stars are likely to be rare or absent on soft mud, the recruitment studies discussed in this section further highlight the potential impact of predation pressure on larvae settlement and establishment.
By contrast, areas of reef damaged by dredging show poor recovery (Moore et al., 2020). Dredge tracks 3 m wide first observed in 1998 were still clearly evident in 2005 and 2007 sidescan sonar imagery, and still visible in 2015 against a background of degraded reefs. The poor recovery may be due to the fragmented tube material being a poor surface for colonisation with benthic predators likely to be present in degraded reef habitat.
Unpublished observations made by one of the authors of this report (D.J. Hughes) indicate that S. vermicularis reefs can arise abruptly on artificial substrata and then disappear. In September 1999 four Dexion™ metal grids each measuring 3 x 3 m square were deployed in the upper basin of Loch Creran. Serpulid reefs are known to be very sparse in this area of the Loch (see Fig. 17 in Moore et al., 2006) and were completely absent from the grid deployment site. The grids were deployed at approximately 17 m depth on fine mud for use in a project studying megafaunal bioturbation (Hughes et al., 2004). The square grids had a leg at each corner, elevating the horizontal metal frame approximately 10 – 15 cm above the mud surface. The grids were visited on scientific dives throughout 2001-2003 and then occasionally up to April 2015. Observations are summarised in Table 2, with descriptions in italics reproduced exactly as recorded in the logbook shortly after each dive.
| Date | Observations |
|---|---|
| September 1999 | Grids installed on seabed |
| 7 November 2000 | Grids visited. No Serpula vermicularis. |
| 18-20 July 2001 | Grids visited. No Serpula vermicularis. |
| 17 August 2001 | Grids visited. No Serpula vermicularis. |
| 16-17 October 2001 | Grids visited. No Serpula vermicularis. |
| 20-22 February 2002 | Grids visited. No Serpula vermicularis. |
| 28 May 2002 | Grids visited. No Serpula vermicularis. |
| 15 February 2003 | Surprisingly, there was a lot of serpulid growth on the grids – erect tubes growing up from the dexion. Looks like the start of a new reef here, quite a bit deeper than normally seen in Creran. |
| 19 August 2003 | Vigorous upright serpulid growth on dexion grids. |
| 19 March 2006 | 3 out of 4 grids heavily colonized by gregarious Serpula, with clusters >20 cm tall. For some reason the westernmost grid had very little growth. Very successful enhancement of serpulid reef growth in an area where they didn’t exist before. |
| 15 August 2010 |
|
| 19 April 2015 | The two grids east of the rope carried many Protanthea, Ascidiella and Ophiothrix. No serpulids at all. Grids to the west were partly buried and had much less epifauna. |
These observations record reef development (and later disappearance) on artificial hard substrata placed on a seabed where serpulid reefs did not previously occur. The abrupt onset of reef growth by February 2003 (more than three years after grid deployment), presumably indicates larval settlement in the summer of 2002. Vigorous serpulid growth persisted until at least 2006, but was in decline by August 2010 and had entirely disappeared by April 2015. The trajectory here is consistent with the pattern outlined in the section Status of reefs in Loch Creran, with healthy reef development in the early-mid 2000s, followed by visible decline and eventual disappearance post-2010. The grids are still in place but have not been visited (to our knowledge) since April 2015.
In 2022, NatureScot commissioned Herriot-Watt University to conduct follow-up monitoring of serpulid regeneration on the different substrata (shells/cobbles with or without mesh bags) originally deployed on the Loch bed by Cook in March 2012 (Cook, 2016). As reported above in the section Recruitment, growth and mortality in Loch Creran, by December 2015, incipient reef growth was recorded on scallop shells and cobbles enclosed in large mesh bags, but not on boulders or shells/cobbles deployed in open piles. Observations continued until 2017, followed by a gap until the next monitoring visit in August 2022. The initial field report from the 2022 survey suggests that reef growth persisted on the large-bagged shells and cobbles, but was absent from boulders, open shell piles or shells in small mesh bags. However, this was true only at the deployment site in Sea Life Centre Bay, with no reef growth recorded at four other sites elsewhere in the Loch. Even at Sea Life Centre Bay, although reef height increased from 2017 to 2022, there was a significant reduction in percentage cover. These results led Harries & Moore (2022) to conclude that the initially promising reef development on introduced substrata may be only temporary, and highly dependent on location within the Loch.
Hughes (2011) suggested that the S. vermicularis reefs of Loch Creran may not have existed continuously since the 19th century observations of Smith (1887) and proposed a search for dateable tube material in sediment cores as a possible method of detecting any cyclical pattern in their occurrence. This has now been attempted but results so far are uninformative. Small fragments of tube are present in deep cores but sediment mixing by bioturbation has erased any stratification that may once have existed, and none of the material recovered so far has been successfully dated (Harbour 2017, cited in Schuler 2021, D.B. Harries, personal communication). The long-term temporal persistence of reefs in Loch Creran is therefore still unknown.
Linne Mhuirich, Loch Sween
S. vermicularis reefs formerly occurred in Linne Mhuirich, a very shallow and highly sheltered inlet of Loch Sween, mid-Argyll. The earliest report appears to date from 1975 (R. Mitchell, cited by Moore et al., 2020), with later observations in the mid-1980s (Earll 1984, Lumb 1986). No further studies were conducted until September 1994 when Scottish Natural Heritage personnel (O. Paisley and D. J. Hughes, unpublished observations) found that the reefs had died out completely, leaving only relict tube debris. This was confirmed by a more detailed survey in November 1999 (Selkie Associates 1999). The cause of disappearance is unknown but direct physical disturbance (e.g. dredging) can be ruled out as the shallow, enclosed situation of Linne Mhuirich renders it inaccessible to large boats. Later work by SAMS in 2004-2005 (Hughes et al., 2008) and in July 2009 (Hughes 2011) at former sites of reef occurrence found no evidence of regeneration. No serpulid reefs were observed on intertidal surveys by Scottish Natural Heritage personnel in 2013 (Moore et al., 2013). Relict reef fragments were reported by a Seasearch dive survey in May 2017 (Seasearch Argyll 2017) but no living reefs were found. However, evidence has recently emerged that suggests aggregations of S. vermicularis may again be forming at Linne Mhuirich (J. Dodd, NatureScot, personal communication, Nov 2023). However, at the time of writing no formal survey has yet been carried out.
Loch Teacuis
Small aggregations of S. vermicularis were discovered in July 2006 in the upper basin of Loch Teacuis, Morvern (Dodd et al., 2009), growing on rocks amongst kelp, mainly at a depth of 0 - 5 m. Aggregations occurred around most of the Loch periphery except for the extreme head. None had been observed on a 1996 survey of the Loch, suggesting that reef formation had begun subsequent to this. There was no further study or detailed monitoring of these incipient reefs, but they were reported to have largely disappeared by 2015 (Kamphausen 2015, cited by Moore et al., 2020). A 2022 survey by NatureScot confirmed that the last few remaining large aggregations present in 2015 had disappeared, although individual serpulids were still common on cobbles and other substrata within the Loch (NatureScot, personal communication).
Other Scottish sea lochs
Serpula vermicularis aggregations were known to be present at the head of Loch Ailort, just outside the boundary of the Sound of Arisaig SAC (Kamphausen et al., 2018, Moore 2019). A diver and drop-down video survey in April 2017 found that aggregations extended along the southern shore of the Loch head, and along the islands and skerries in the entrance to the head of the Loch in the south-west. They occurred in very shallow water (~ 2 – 4 m BCD) on cobbles and boulders among kelp (a very similar environmental setting to the aggregations formerly present in Loch Teacuis). Aggregations were predominantly small, with heights in the range 2 – 26 cm (Kamphausen et al., 2018) and were morphologically simple, consisting of clusters of tubes (up to ~50 individuals) of the same age class (Moore, 2019). This size range and simplicity contrasts with the large, complex, multi-generational structures typical of Loch Creran, and may be evidence that these structures developed more recently in Loch Ailort compared with Loch Creran. However, further monitoring carried out in March 2023 failed to identify any evidence of live or dead serpulid aggregations in the location of the baseline diver transects established in 2017 (Sayer-Mitchell et al., 2023).
There is evidence that S. vermicularis reefs formerly existed in West Loch Tarbert, mid-Argyll. Relict reef fragments were recorded by divers in January 2017, with later observations of tubes containing living worms growing upwards from collapsed reefs (Seasearch Argyll 2017). Further observations of possibly regenerating reefs were made in July 2018 (Seasearch Argyll 2018). Relict tube debris was again recorded in September 2021 but the brief report makes no mention of any living reefs (Seasearch West of Scotland 2021).
Galway, West of Ireland
S. vermicularis reefs have been recorded at sites in Galway, western Ireland, since the 1970s (Bosence 1973, Minchin 1987). Unfortunately, these examples have not been studied in detail and there is very little recent published information available (Buckman & Harries, 2020, Schoenrock et al., 2021). A recent video lecture posted on YouTube (Seasearch Ireland 2022) lists four localities on the Galway coast. At two of these locations (Blacksod Bay, Killary Harbour) the reefs were said to have been destroyed or severely damaged by towed fishing gear. Reefs at Leitir Caladh were reported to be in good condition, although no further details were given. The fourth Irish locality, Ardbear Salt Lake (also known as Ardbear Lough) is the best-studied. Most of the available information is contained in the unpublished PhD thesis by Leahy (1991).
Serpulid reef decline and regeneration in Ardbear Salt Lake, Galway
Ardbear Salt Lake is a small (~4.5 km2) inlet with very restricted communication with the wider Ardbear and Clifden Bays, Galway. Water depth is mostly 4 – 6 m, with one deep depression (27 m), and a predominantly muddy substratum. In July 1986, discrete S. vermicularis aggregations were found along the southern shoreline, mostly in the 4 – 8 m depth range, with no significant growth below 12 m. Aggregations were small, reaching a maximum height of ~0.5 m, and width ~0.3 m (Leahy 1991). In early August 1987 there was a mass mortality of invertebrate fauna (including S. vermicularis) in the inlet, driven by an episode of severe hypoxia in which dissolved oxygen concentrations dropped from ~6 ml L-1 to ~1.5 ml L-1 (July 1987). Anoxic conditions were recorded in the deepest part of the inlet. Serpulids and their associated epifauna did not appear stressed on 24 July 1987, but by 7 August all visible life below 5 m depth was moribund and most S. vermicularis were dead and decomposing (Leahy 1991). By 14 August 1987, living reefs persisted only above 5 m depth. The hypoxic episode was followed in September/October 1987 by a bloom of the toxin-producing flagellate Heterosigma akashiwo.
Dissolved oxygen concentrations in Ardbear Salt Lake began to recover from September 1987, and by May 1988 only the deep basin remained anoxic. After the phytoplankton bloom in late summer 1987, serpulid reefs appeared to have been almost completely wiped out, with the exception of a few isolated clumps at the inlet mouth. However, 3.5 years later, aggregations measuring up to 60 cm in height and 50 cm in width were recorded at the study site. Leahy (1991) considered that reefs of this size implied an unfeasibly rapid growth rate if arising entirely from larval settlement, and that some serpulids must have survived the hypoxia and plankton bloom to provide a nucleus for regeneration.
When Ardbear Salt Lake was surveyed from July to September 1972, S. vermicularis reefs were reported to occur over approximately 25% of the inlet bed, covering areas hundreds of metres across, with reef heights up to 2 m (Bosence, 1979). Reef growth extended down to 19 m depth. Leahy (1991) noted that no formations of this size existed at the time of their subsequent work in 1986 – 1987. Depth range was also more restricted, with no significant reef growth below 12 m. This contrast indicates a striking change in the extent and condition of the local reefs between the two surveys. It is also worth noting that the reef dimensions described by Bosence (1979) far exceed anything ever recorded in Loch Creran. The small size, shallow depth and limited water exchange of Ardbear Salt Lake predispose this water body to periodic hypoxia and consequent ecosystem collapse (Henry et al., 2008), and it is possible that the serpulid reefs here undergo repeated cycles of mass mortality and regeneration. Unfortunately, the wide gaps between intervals of scientific study mean that we cannot plot these changes in detail.
Recruitment and growth in Ardbear Salt Lake
Leahy (1991) studied the reproductive cycle of reef-building S. vermicularis by monthly sampling of adult worms in Ardbear Salt Lake and Killary Harbour (Connemara). Larvae and juvenile worms were maintained in aquaria to observe settlement preferences and measure growth rates. In both field localities spawning occurred during the summer, with at least two discrete episodes of egg/sperm release. In the aquarium, larval settlement began 34 days after fertilization. Of the 26 larval settlement events observed, 25 individuals settled on the anterior end of occupied adult tubes, close to the aperture. The sole exception settled on a tube from which the adult worm had been removed. Settlement was not recorded on rocks or cleaned fragments of old reef material. Tube growth of ~2 cm was observed on a portion of reef maintained in the aquarium for 12 months (Leahy, 1991), a value lower than the 3.3. cm y-1 recorded in Loch Creran (Hughes et al., 2008) but possibly attributable to the differing nutritional environments in the two settings.
Leahy (1991) proposed settlement in close proximity to an adult tube aperture as an adaptation to reduce the likelihood of burial in sediment and as a means to achieve a good elevated position for suspension-feeding. In the aquarium, the presence of newly-settled juveniles appeared to encourage further settlement: an initial settlement of 12 individuals was followed by 26 more within eight days. An attraction of settling larvae to established adults was suggested by Bosence (1979). Leahy’s observations may provide a solution to the apparent contradiction between the need for gregarious settlement to initiate and maintain reef growth, and the poor recruitment observed on dead tube fragments (Leahy, 1991; Hughes, 2011; Moore et al., 2020). Relict, unoccupied tubes lying prone on the seafloor will be exposed to sedimentation and grazing by benthic predators, and juvenile worms will not gain the benefit of an elevated feeding position. However, Chapman et al., (2007) found no difference in recruitment onto live and dead reef fragments deployed in a vertical orientation in Loch Creran, so the question of the factors promoting larval settlement remains unresolved.
Summary: dynamics of Serpula vermicularis reefs
The evidence presented throughout this literature review permits the following general conclusions:
- Outside localised areas of physical damage caused by dredging and boat moorings, Loch Creran’s serpulid reefs appeared to be in a healthy condition until at least 2005. The reefs may have been in an expanding phase from 1989 onwards, but due to the lack of earlier data, some uncertainty remains over this suggestion.
- Visible deterioration in local reef condition was apparent from 2011 at latest. A detailed survey in 2019 showed that decline had extended throughout most of the Loch, leading to a ~20% loss of reef habitat. Deterioration is manifested by reef collapse, reduced occupancy by living worms and overgrowth by algae and epifauna.
- Relict tube material provides a poor surface for colonisation, but worms can recruit successfully onto stones, shells or artificial hard substrata if shielded from benthic grazers/predators in the early juvenile stage. However, there is still uncertainty about the long-term persistence of reef growth on these shielded substrata.
- Observations in Loch Teacuis, Loch Ailort, in upper Loch Creran and at the site of the former Barcaldine alginate factory show that reefs can form de novo where previously absent, or where previously eliminated by organic pollution.
- Observations in Loch Teacuis, Linne Mhuirich and upper Loch Creran show that reefs can disappear completely in locations where anthropogenic physical disturbance can be ruled out as a cause.
- In Ardbear Salt Lake, Ireland, an episode of severe hypoxia in summer 1987 caused mass mortality of S. vermicularis reefs. There is some evidence for previous cycles of reef decline and regeneration in this locality.
Status of Serpula vermicularis in Scottish waters
The decline of Loch Creran’s serpulid reefs is documented in Status of Reefs in Loch Creran. However, it may be worth putting this into a broader context by examining whether there is evidence for any change in status of S. vermicularis across Scotland as a whole. Available data was downloaded from the Archive for Marine Species and Habitats portal (open source) using the ‘bounding box’ function and the species search criteria ‘Serpula vermicularis’. This dataset contained 1256 datapoints from surveys conducted between 1980 and 2019, of which 827 datapoints contained a categorical assessment of the abundance of S. vermicularis using the SACFOR (Superabundant, Abundant, Common, Frequent, Occasional, Rare) abundance scale. Additional data was downloaded from the Geodatabase of Marine features adjacent to Scotland (GeMS) habitat and point datasets. This database contains data of key habitat distribution in the form of shapefiles, and point data on locations of marine features, but not abundance data (Figure 1). Ordinal logistic regression analysis was used to compare the effect of the independent variable (in this case the year that the surveys were conducted) to the categorical variable (ranked and ordered by levels; S – Super abundant, A – Abundant, C – Common, F – Frequent, O – Occasional, R - Rare).
The data was split into two categories; Loch Creran data alone, and the rest of Scotland (excluding Loch Creran). Loch Creran observations were available from 1994-2019, while the rest of Scotland had data from 1980-2019, the length and size of this data set reduces the potential for annual survey effort to influence outcome. This categorisation of the data was applied to determine whether S. vermicularis decline is local to Loch Creran, or if it is widespread across the whole of Scotland. The following caveats apply; 1) Ordinal logistic regression does not consider spatial autocorrelation of samples (for example, two samples which are next to each other might have similar values). The significance of this caveat is somewhat diminished when comparing the annual effects on the outcome of surveys in the same area, 2) there are no recorded absence data for the serpulid reefs, instead only the categories which define presence along levels of abundance, 3) Differences in survey effort may bias the outcome of this test, bias potential is reduced by the size and length (temporally) of the datasets. More survey points included annually will result in greater accuracy in model predictions. A reasonable spread of data were observed although greater survey intensity is observed in more recent years in the Loch Creran data (Appendix 1), where data were insufficient, analysis was not performed. This analysis therefore provides insights into the likelihood of recording different abundance scores of S. vermicularis over the years that data have been collected.
Using the model output, the predicted annual values along the SACFOR scale were calculated (Figure 2). The coefficients of the model refer to the likelihood of recording an observation in ranked order. For example, a coefficient of +2 means a higher chance of an observation on the S, A and C, end of the scale, than a score of -2 which means a higher chance of observing a F, O, or an R observation. Plotting the coefficients shows a decline in the likelihood of abundant observations over time in both Loch Creran and Scotland as a whole. This may suggest a nationwide decline in S. vermicularis populations. However, without clear absence data no definitive conclusions can be drawn regarding the loss of the reefs in key areas. It should be noted that observations of both occurrence of individuals and the occurrence of reefs / aggregations of S. vermicularis are included in this dataset.
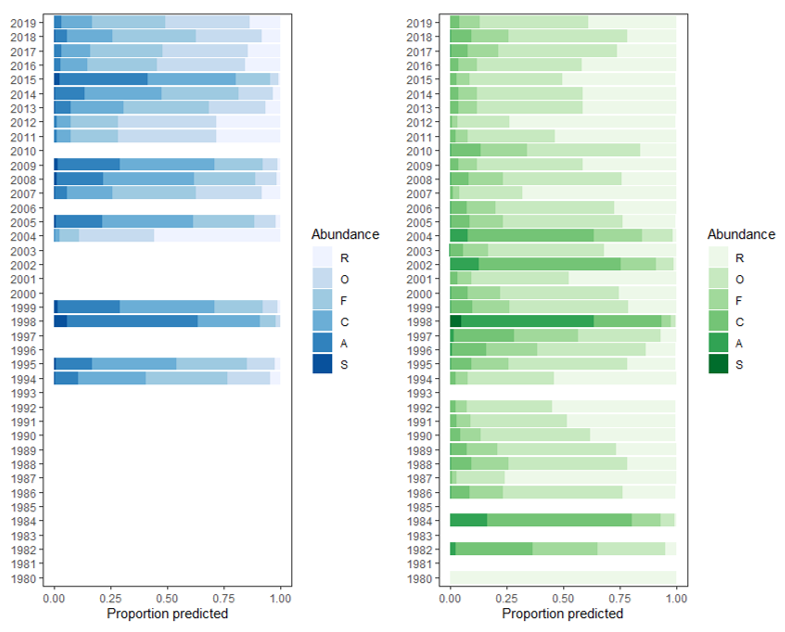
Figure 1. Likelihood of different observations on the SACFOR scale (x-axis) in different years (y-axis), in Loch Creran alone (left, blue bars) Scotland as a whole (right, green bars).
Click for a full description
Figure 1 shows bar graphs of the likelihood of different observations on the SACFOR scale (Superabundant, Abundant, Common, Frequent, Occasional, Rare) from 1980 to 2019. The values are representative of proportional odds logistics regression modelling. The left bar graph represents Loch Creran, with highest levels of superabundance in 1998. The right bar graph presents Scotland as a whole, with the highest levels of superabundance recorded in 1998.

Figure 2. Plotting the coefficients of the proportional odds logistic regression model shows a general trend towards observations on the Frequent, Occasional and Rare end of the SACFOR scale in both Loch Creran and in the rest of Scotland.
Click for a full description
Figure 2 is of two regression line graphs which plot the coefficients of the proportional odds logistic regression model showing a general trend towards observations on the Frequent, Occasional and Rare end of the SACFOR scale in both Loch Creran and in wider observations in the rest of Scotland. The blue straight lines represent the fit of a linear regression with standard error around the lines represented by grey shading. Here, the coefficient refers to a likelihood of observations in a ranked order; a score above zero means higher likelihood of more abundant observations being recorded (SAC), a score below zero means greater likelihood of recording less abundant (FOR) observations. The first graph plots serpulid data for Loch Creran, the second graph plots serpulid data for the rest of Scotland.
Other reef-building serpulids
Mass occurrences leading to the formation of reefs (or reef-like structures) have been recorded in approximately 10% of species in the family Serpulidae (ten Hove, 1979; ten Hove & van den Hurk, 1993). The potential importance of these structures in the marine carbon cycle and their effects on local habitat complexity and biodiversity has led to them being considered “ecosystem engineers” sensu Jones et al., (1994) (Montefalcone et al., 2022). Other than S. vermicularis, the best-studied reef-builders are Galeolaria hystrix from New Zealand (Smith et al., 2005) and the invasive Ficopomatus enigmaticus, a species possibly native to the Indian Ocean but now worldwide in brackish coastal environments (Schwindt et al., 2004b). The species is not currently found in Scottish waters but has been found elsewhere in the UK. However, there is very little published information on the growth or long-term dynamics of reefs of these (or any other) serpulid species. Mean individual tube extension rates of 4.0 cm y-1 and 1.7 cm y-1 have been estimated for G. hystrix and Spirobranchus cariniferus, a serpulid which forms aggregations in the New Zealand intertidal zone (Riedi & Smith 2015). Much faster extension rates (up to 2.0 cm month-1) were reported for two reef-building serpulids (F. enigmaticus and Hydroides dianthus) in the warm conditions of an Italian saline lagoon (Bianchi & Morri, 2001).
Schwindt et al., (2004a) provided some information on the decadal-scale dynamics of reefs of the invasive serpulid Ficopomatus enigmaticus in an Argentinean coastal lagoon. Over the 1975-1999 period, reef cover in the lagoon increased by 24%. Mean reef growth rate was estimated as 1.6 cm month-1, with smaller reefs growing faster than larger ones. Monthly growth rates were highest in summer and lowest in winter, correlating with the seasonal change in water temperature.
Santonnini et al., (2021) documented the abrupt appearance of dense serpulid aggregations in the Mar Menor, a shallow lagoon on the Southeastern coast of Spain. The lagoon was historically oligotrophic, with clear water and dense seagrass beds, but underwent a process of eutrophication as a result of nutrient inputs from agricultural fertilizers. In October 2019, heavy rainfall caused a massive inflow of fresh water into the eutrophic lagoon, leading to water column stratification and euxinia (anoxic conditions and generation of hydrogen sulphide). Just weeks after the euxinic episode, dense aggregations of serpulids (mostly Hydroides elegans and H. dianthus, but with some S. vermicularis and S. concharum) appeared on rocks, bivalveshells, boat hulls and other artificial substrata. Santonnini et al’s (2021) paper is mainly concerned with the identification and morphology of the serpulid species and provides little information on settlement or growth rates, but it provides an example of rapid reef development following a severe environmental perturbation. In this case, the serpulids proliferated opportunistically in a disturbed environment with little or no competition from other benthic organisms.
There appears to be no published information on serpulid reef decline and/or regeneration other than the S. vermicularis studies discussed throughout the Serpula vermicularis section above.
A timeline of environmental change and human activities in Loch Creran, 1980-2022
Introduction
It is widely accepted that large reef forming accumulations of S. vermicularis are declining in Loch Creran, and that the decline is widespread. The following section looks at available data highlighting the various environmental and anthropogenic pressures that have occurred in the Loch in recent times. The data presented is not interpreted in this section. The interpretation of potential impacts on S. vermicularis reefs follows in the Screening Assessment section. The interpretation of the data often requires a greater number of sampling points throughout the year to capture the true annual variations, limited sample dates often miss true maximum and minimum values. The source data presented is as provided. Significant trends and gaps in data are highlighted.
The data presented include those climatic parameters such as temperature, rainfall and storm events; physical and chemical parameters of seawater where available and as relevant to the Loch as possible; environmental parameters such as phytoplankton abundance and toxic algal blooms; and- anthropogenic impacts such as aquaculture, invasive species, boat traffic and forestry in the Loch Creran catchment.
The data was collected from a range of stakeholders to provide an overview of environmental changes that have occurred in the Loch, with an accompanying spreadsheet database of all known data sources. Where possible, open-source data were downloaded for this report. Most environmental and chemical data for the region were provided by the Scottish Environment Protection Agency (SEPA). Forestry data were downloaded (open source) from the National Forestry Inventory (NFI) and shipping data were obtained from the Marine Management Organisation (MMO). Multiple datasets were downloaded from the Scotland’s Aquaculture website and additional data were provided by Scottish Sea Farms (SSF). Further information was obtained through personal communications regarding the activities and changes to practices that have occurred in Loch Creran over the study period. While personal communications have been relied upon for some aspects of this section, care has been taken to ensure that no opinions or biases are reflected.
West Scotland climatic data
The highest resolution weather data for the UK are kept by the Met Office and the data are available for download for various weather stations across Scotland. The closest weather station to Loch Creran which regularly logs data is at Dunstaffnage Bay (Lat: 56.451, Long: 5.439), approximately 8 km south of Loch Creran. Monthly maximum and minimum air temperatures are recorded by the station (Figure 3). A general, significant upwards trend in both monthly air maximum and minimum temperatures is observed (Figure 3). This increase in temperature is only significant in the marine context if it directly relates to increased water temperature, for which data are scarce for Loch Creran (see below). Monthly rainfall is also measured at the Dunstaffnage Bay weather station (data available for the period 1972-2021) and appears to be relatively consistent with no clear increases observed.
However, some peak events are obvious. For example, during the winter of 2011-12 the west coast of Scotland had some severe storm events which are visible in the increased average rainfall that year. The most severe storm (highlighted in Figure 4) was caused by a deep Atlantic low-pressure system and Scotland had winds gusting between 60-70 knots. This storm is considered as the most significant since January 2005. The most significant impacts were felt across Scotland on 8th December 2011 but the event itself was just one of several storm events that occurred throughout November and December of that year. Maximum gusts of wind measured at the Dunstaffnage Bay station were 76 knots (Met Office, 2011).
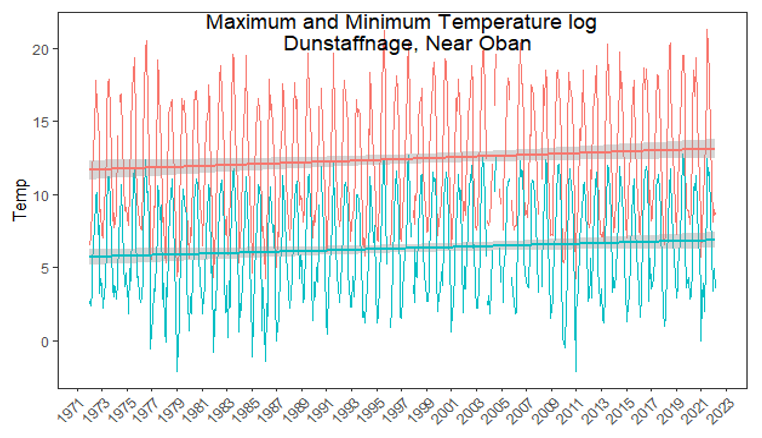
Figure 3. Monthly maximum (red) and minimum (blue) atmospheric temperatures from the closest Met Office weather station to Loch Creran (Dunstaffnage).
Click for a full description
Line graph showing the monthly maximum and minimum temperatures from the closest Met Office weather station to Loch Creran (Dunstaffnage). Red and blue lines indicate a straight line regression model with grey shaded areas representing standard error. Model analysis (straight fitted lines) finds that both maximum and minimum temperatures in the region are slowly increasing (maximum: y = 9 x 10-5x + 9.3, R2 = 0.0133, p = 0.005, minimum: y = 7 x 10-5x + 3.9 R2 = 0.01, p = 0.01) between the years of 1972 and 2022.
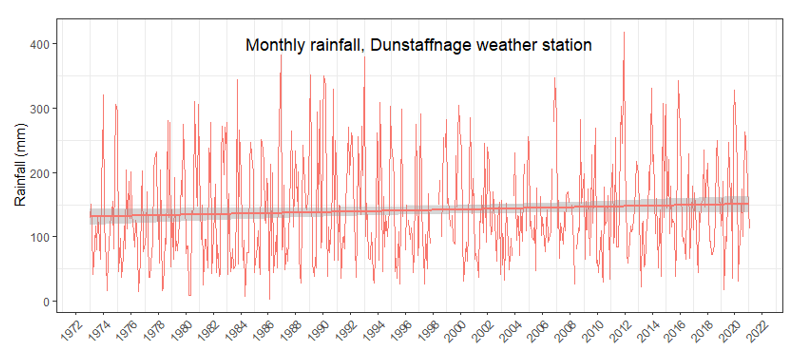
Figure 4. Monthly rainfall downloaded from Dunstaffnage Met Office weather station. Rainfall in 2012 was above average. The year 2012 indicates highest recorded rainfall in dataset.
Click for a full description
Figure 4 is a line graph showing monthly rainfall at Dunstaffnage weather station from 1972 to 2022. Rainfall is variable over the 50 year period. In 2012, rainfall was recorded as above average, over 400mm. The period from 2006 onwards covers the period of Loch Creran serpulid decline.
Specific water temperature data within Loch Creran are scarce, but the National Oceanic and Atmospheric Administration (NOAA) have open-source sea surface data. The closest available grid square to Loch Creran (Lat: 56.75, Long: -5.75, approximately 50 km from Loch Creran) was analysed (Figure 5). The sea surface temperature anomalies (as above or below the annual average temperatures) highlight more recent events which occurred in 2015, 2018 and 2021 where extreme cold weather caused temperatures to drop by more than 2°C below annual averages. In 2006, the data show a peak above average temperatures by 4°C (Figure 5).
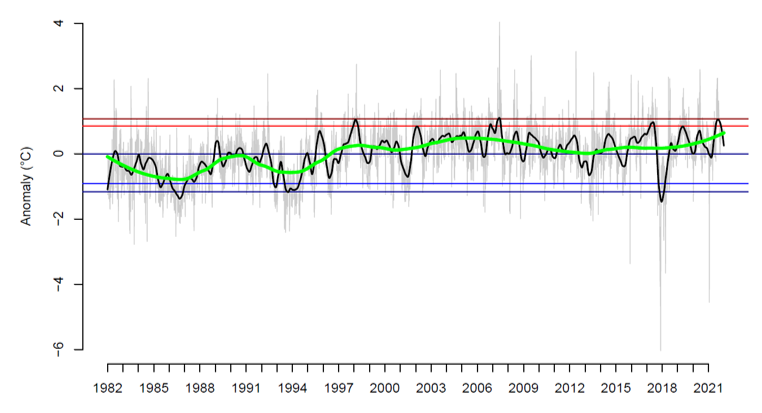
Figure 5. Sea surface temperature anomalies (above or below the average ranges).
Click for a full description
Figure 5 shows a line graph of sea surface temperature anomalies above or below the average temperatures for the whole series. Zero line represents the average temperature for that day of the year, the red line represents the 90th percentile of those temperatures and the dark line is the 95th percentile, the blue line (light) represents the 10th percentile, and the darker blue straight line represents the 5th percentile for the data. Any value above the light red line for periods of five or more days. The green line indicates decadal fluctuations (smoothed loess fitted line) and the black line indicates the annual temperature fluctuations (smoothed loess fitted line). The extreme cold spell in Feb – Mar 2018 corresponds to Anticyclone Hartmut (“The beast from the east”).
Data are interpolated from satellite records held at the National Oceanic and Atmospheric Administration website.
Temperature values which are more local to Loch Creran were made available by SEPA for the River Creran (1990 – 2018), Creagan Bridge (2000 – 2013) and North Shian (2000 – 2009) (Figure 6, see also the Riverine input and general water quality section). No obvious changes in water temperature are apparent at any of the three sites from the available data.
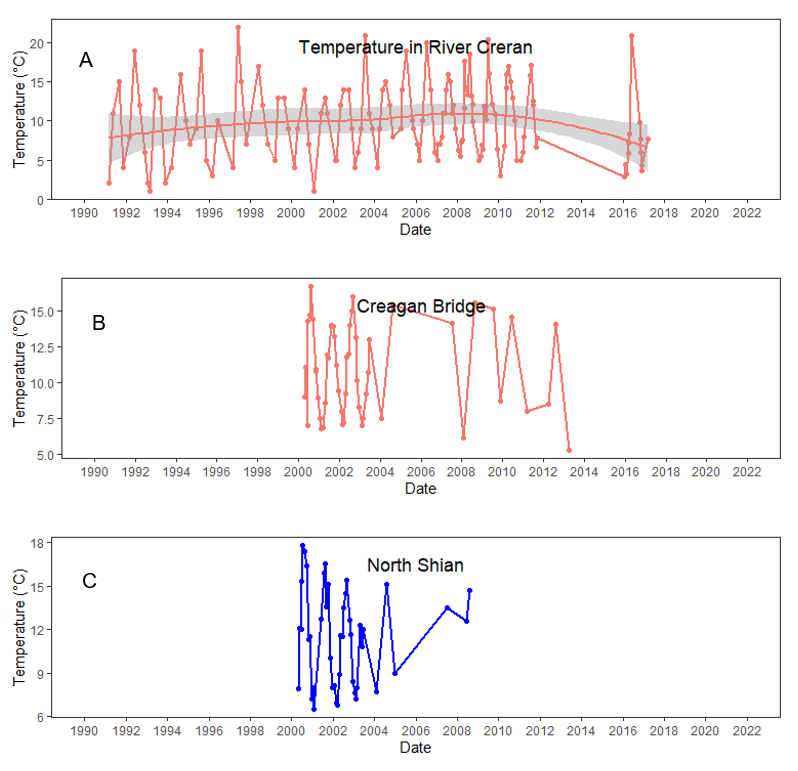
Figure 6. Local available water temperature values from Loch Creran as taken by SEPA. A from the River Creran, B from close to the Creagan Bridge and C from North Shian.
Click for a full description
Figure 6 shows three line graphs, of water temperature values from Loch Creran taken by SEPA. (A) shows temperature values from River Creran between 1991 and 2017; (B) shows temperature values from Creagan Bridge between 2000 and 2013; (C) shows temperature values from North Shian between 2000 and 2009. Red curved line in River Creran plot (A) represents Loess regression fitted curve with standard error in grey shading.
Physical characteristics of Loch Creran
Much of the baseline knowledge surrounding the physical characteristics of Loch Creran is derived from the Scottish Sea Lochs Catalogue (see Edwards & Sharples, 1986). Loch Creran is 12.8 km long with a maximum depth of 49 m. It has a recorded mean depth of 13.4 m, and it has a low water area of around 13.3 km2. The Loch can be divided into two basins: a larger lower basin and a smaller upper basin. The basins are separated by a sill with a mean depth of approximately 1 m. There is another 4 m sill at the narrow Loch entrance (Edwards and Sharples, 1986). In the main basin, the salinities are usually in the range of 30-34 PSU but salinities in the surface 4 metres can drop to approximately 23 PSU. The temperature in the main basin is similar to that of the adjacent sea with lows of about 6°C in February/March and highs of 13–15°C in August/September. Historically, in the main basin the dissolved oxygen concentration did not fall below 87% saturation (Gage, 1974). In the smaller upper basin, there is a halocline present, usually at 0.5 metres depth but occasionally this can be as deep as 3 metres. The surface salinity in this basin is usually around 28–29 PSU but can sometimes fall to 5 PSU. The bottom salinity of this basin is fairly constant and stays around 30-32 PSU (Gage, 1974). Loch Creran has a flushing time of around three days (Turrell et al., 2020).
Loch Creran phytoplankton
Phytoplankton cycles were first described in Loch Creran in 1978, in a six-year study which regularly measured phytoplankton species from stations C3 and C5 (Figure 7, Tett & Wallis, 1978). The C5 and C3 monitoring stations in Loch Creran are in central parts of the Loch where the depth is greater (Figure 7) (Moschonas et al., 2017; Tett & Wallis, 1978). The two sites were visited twice a month between 1972-1976 and, on less regular occasions, between 1970 and 1971. Daily samples were taken to capture March spring blooms in 1972 and 1973 at C3 and C5 respectively (Tett & Wallis, 1978). Intense sampling continued through the 70’s and these conditions are often used as a baseline which demonstrates change in the Loch, mainly because the data were gathered before the onset of aquaculture in the region (Tett et al., 2008). However, it is important to highlight that while aquaculture production was not taking place in the Loch at that time, there were significant anthropogenic pressures occurring, particularly the operation of the former alginate factory at Barcaldine. Later studies which modelled the nutrient composition of water in Loch Creran found that, between 1975 and 2003, local nutrient input (N and P) in Loch Creran increased at these two sites. However, no apparent increases in concentration were observed across the whole loch (Laurent, 2009). The comparisons were made between five main sampling sites in the 1970’s dataset and nine sampling stations during the 2002-2003 sampling campaign (Laurent, 2009).
Modelled nitrogen annual fluxes in 1975 were 33.4 mmol year-1 Dissolved Inorganic Nitrogen (DIN) in seawater and 1.2 mmol year-1 DIN in riverine input, compared with 1.6 mmol year-1 DIN (seawater) and 1.6 mmol year-1 DIN (riverine) in 2003, with additional input of 8.8 mmol year-1 DIN from fish farm aquaculture (Laurent et al., 2006). Observed chlorophyll concentrations decreased substantially in the region despite the increased nutrient input. The findings of Laurent et al., (2006) led to more research into the effects of nutrient input and chlorophyll levels on plankton communities. A study in 2008 proposed a structural indicator of phytoplankton community function, referred to as the Phytoplankton Community Index (PCI) (Tett et al., 2008). Data taken from a baseline or ‘type specific’ condition (i.e., 1971-1981) are enveloped as a reference and compared to more current (or differently enveloped) data. The resulting proportion of new data that falls within the original referenced data form the PCI (between 0-1). When analysed in this way, Loch Creran showed a change in the balance of phytoplankton, a reduced PCI leading to decreased chlorophyll a but no associated change in nutrient input (Tett et al., 2008).
The dominant phytoplankton species in Loch Creran in the spring bloom in 1972 were chain-forming diatoms, Skeletonema spp., which were later studied by Whyte (2012). Although exact numbers were not provided, the major trends were described. In general, diatoms in Loch Creran significantly decreased between 1970 and 2009, as did the dominant diatom (Skeletonema spp.) and both dinoflagellates and small flagellates (Whyte, 2012). These findings were localised to Loch Creran, with the nearby Lynn of Lorn observatory (situated to the south of the entrance to Loch Creran) showing no significant change (Whyte, 2012; Tett, 2013). Furthermore, between 2011 and 2013, the community structure showed a rise in the occurrence of autotrophic/mixotrophic dinoflagellates with a concomitant and substantial drop in the proportion of the community composed of diatoms, with the exception of the Pseudo-nitzschia genus which showed a significant increase (Whyte et al., 2017). A community shift from diatom-dominated systems to dinoflagellates is often associated with eutrophication events (Paerl, 1997). Later, change was observed at the Lynn of Lorn observatory where similar patterns were noted, namely a decrease in diatoms from 1970 to 2015 and an increase in dinoflagellates and ciliates which correlated with an increase in sea surface temperature (Tett, 2019).
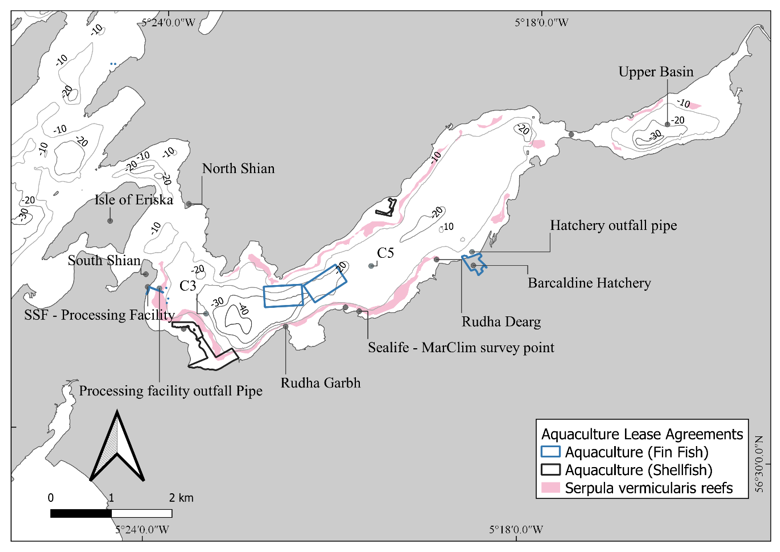
Figure 7. Overview of various factors in Loch Creran, the location of aquaculture agreements with Crown Estate Scotland (CES) and the location of sample stations and outfall pipes into the Loch (as discussed in phytoplankton and chemical sections).
Click for a full description
Note that the Barcaldine Hatchery pipeline is shared with the Local Caravan and Camping Site outfall. The MarClim survey points and outfall pipe locations are presented in Human Activities section. Data on S. vermicularis presence were downloaded from the GeMS habitat database, aquaculture agreements were downloaded from Crown Estate Scotland.
As part of the Official Control (OC) programme for ensuring shellfish safety, Food Standards Scotland (FSS) regularly monitor phytoplankton that may be harmful to human consumers of shellfish. The following analysis is based on freely available FSS data downloaded from Scotland’s Aquaculture online portal (Scotland's Aquaculture | Search Data). Data from Rubha Mor, Loch Creran, is available from 2011 – 2019 for the following phytoplankton species/genera; Pseudo-nitzschia spp., Alexandrium spp., Dinophysis spp (reported at genus level)., Prorocentrum lima, Prorocentrum minimum, and Lingulodinium polyedra. The resolution needed to conduct in-situ sampling of phytoplankton to an accurate level is generally considered to be weekly (Wang et al., 2022). The recommended sampling frequency set by FSS is weekly between March and September, fortnightly in October and from November to February, monthly sampling is advised (Murray, 2009). Therefore, the data are presented with the caveat that spring bloom events may have been missed and trends are discussed.
Some species within the Pseudo-nitzschia genus are associated with harmful blooms that produce neurotoxins (specifically domoic acid). If present in human foods at sufficient concentration quantities this toxin can cause amnesic shellfish poisoning (Lelong et al., 2012). The highest density of this genus in Loch Creran was found in 2011, where numbers associated with the spring bloom reached over 600,000 cells L-1 of seawater sampled. The Pseudo-nitzschia genus is plotted separately from other species given the orders of magnitude difference in abundance (Figure 8). In examining the different toxin producing species within the dataset, the various toxin alert levels from the National Reference Laboratory network (a network of laboratories required by countries in the World Health Organisation to implement quality assurance), Food Standards Scotland as referenced in the Algal Toxins monitoring Programme can be used (FFS, 2016; Murray, 2009). It should be noted that density of Pseudo-nitzschia species where not categorised to species level and different Pseudo-nitzschia species may or may not produce toxins (Fehling et al., 2006).
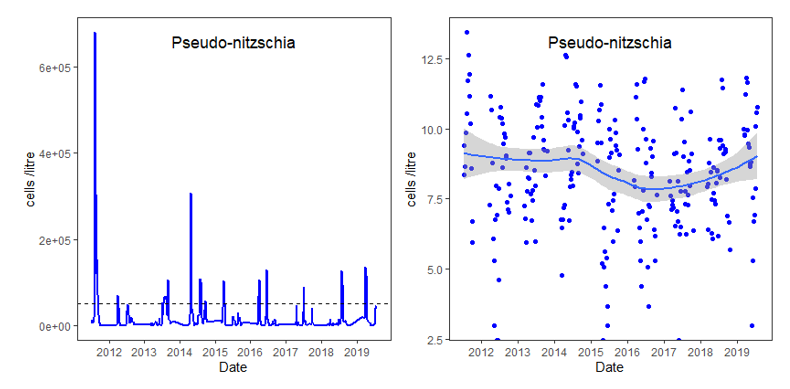
Figure 8. Measurements of the diatom Pseudo-nitzschia in Loch Creran as recorded by Food Standards Scotland (FSS).
Click for a full description
Line and scatter graph of measurements of the diatom Pseudo-nitzschia in Loch Creran. Peaks (left hand plot) represent the spring/summer blooms of this genus which is plotted separately from other potentially harmful species because of its much higher abundance. Dashed line indicates the alert limit for this genus (50,000 cells L-1). Right hand plot is logged cells-1 no significant changes are apparent in these data. The fitted blue line shows the Loess regression fitted curve for the logged data and the shaded grey area shows the standard error around the fitted line.
More recent years have seen an increase in the number of the phytoplankton genus Alexandrium in Loch Creran, with certain species within this particular genus related to paralytic shellfish poisoning (PSP) in humans (Collins et al., 2009; Brown et al., 2010). In the years since 2016, the spring-summer blooms have seen over 4,000 cells L-1 of seawater sampled in the Loch (Figure 9). However, not all Alexandrium species produce toxins and the most common species found in Loch Creran has been A. tamutum, a non-toxic species (Parks et al., 2019).
Several Dinophysis sp. are known to cause gastrointestinal illness and Diarrhetic Shellfish Poisoning (DSP) indirectly in humans through the production of diarrhoetic toxins and pectenotoxins, even at low concentrations (Reguera et al., 2014). This genus is considered the main threat to shellfish aquaculture production in Japan, Chile and European countries due to shellfish harvesting bans, which can occur as a result of their presence. Similarly, the genus Prorocentrum produces diarrhoetic toxins, although to a lesser extent than Dinophysis (Solter & Beasley, 2013).
Both Lingulodinium sp. and Protoceratium sp. produce toxins which are classified among diarrhoeic shellfish toxins, known as yessotoxins, but are not known to provoke DSP symptoms in isolation (Franchini et al., 2010). Unlike toxins produced by Dinophysis and Prorocentrum, yessotoxins induce dehydration and shock at high concentrations (Hinder et al., 2011).
The final genus, Protoperidinium sp., which was previously considered to be toxicologically benign (James et al., 2004), has since been discovered to consume Azadinium spinosum (too small and morphologically indistinct to monitor regularly), thought to produce azaspiracid toxins (Tillmann et al., 2009). Azaspiracid poisoning results from the ingestion of contaminated shellfish, but is especially common in mussels (Kliegman et al., 2020) with symptoms resembling those of DSP (Wu et al., 2019).
FSS monitoring samples were taken from relatively consistent locations in Loch Creran which were close to Rubha Mor, near the current South Shian Processing Facility and the region where oyster harvesting and farming occurs (Figure 10).
Long term change from historic sampling
To represent sampling periods before and after the establishment of the fish farm, data from 1979-1981 were enveloped (or gathered together as a single dataset) for comparison to data from 2011-2016 for Pseudo-nitzschia species recorded in Loch Creran (for methods and full dataset see Whyte, 2012; Whyte et al., 2017). When compared to the pre-fish farm dataset (1979-1981), an observed delay in spring biomass (as measured by cells L-1) is recorded with summer biomass lower and a delayed onset of autumnal blooms in the 2011-2016 dataset (Figure 11).
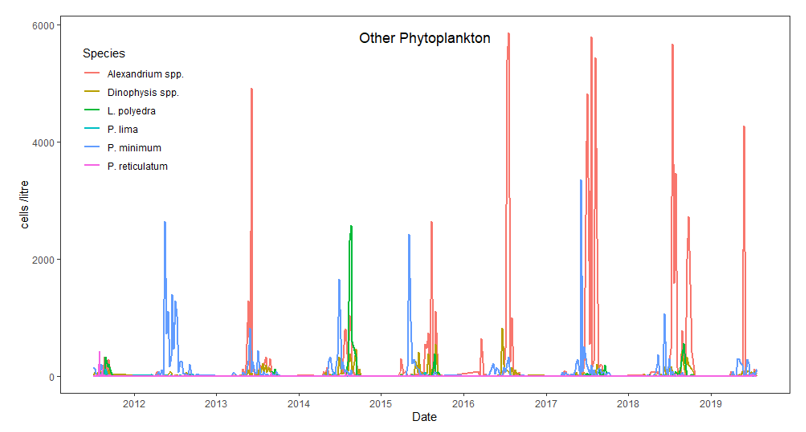
Figure 9. Cell density of dinoflagellate taxa in Loch Creran from FSS regulatory monitoring. For Alexandrium the FSS alert limit is 40 cells L-1 and for Dinophysis the alert level is triggered at 100 cells L-1.
Click for a full description
Figure 9 shows the cell density of dinoflagellate taxa; Alexandrium sp., Dinophysis sp., Prorocentrum sp., Lingulodinium sp, Protoceratium sp, and Protoperidinium sp in Loch Creran from FSS regulatory monitoring. For Alexandrium the FSS alert limit is 40 cells L-1 and for Dinophysis the alert level is triggered at 100 cells L-1. The graph covers the years 2011 to 2020. In general, Alexandrium sp. shows the highest cell L-1 across six of the years shown.

Figure 10. The location of phytoplankton sampling sites in Loch Creran.
Click for a full description
Data on S. vermicularis presence were downloaded from the GeMS habitat database, aquaculture agreements were downloaded from Crown Estate Scotland.
The diatom Skeletonema costatum dominated spring blooms and made up approximately 90% of the diatoms in Loch Creran between 1972-1981, the shift in PCI is noted and modern datasets indicate that S. costatum have decreased in numbers by an order of magnitude during March and April in both 2006-2007 data (Tett et al., 2008), 2008-2009 data (Whyte, 2012) and in the data presented in the present study from 2011-2016 (Figures 11-13). No change in nutrient levels were noted in 2006-2007, but chlorophyll, that can be used as an indicator of phytoplankton abundance, was noted as lower. However, in 2009, no change in silicate and nitrates were observed but phosphate levels were significantly lower than previous years (Tett et al., 2008; Whyte, 2012).
The dinoflagellate genus Alexandrium belongs to the group of phytoplankton thought to have increased in biomass in Loch Creran in recent years. In plotting recorded levels as above, the onset of biomass appears to have shifted earlier, with greater peaks in late summer/autumn (Figure 13).
The dinoflagellate genus Alexandrium is within the order Gonyaulacales, some species of which are known to produce toxins. The genus is amongst the phytoplankton thought to have increased in biomass in Loch Creran in recent years. In plotting recorded levels as above, the onset of biomass appears to have shifted earlier, with greater peaks in late summer/autumn (Figure 13). The genus was originally considered to be a different genus (Gonyaulacales) and so is grouped together with the 1970s data as such.
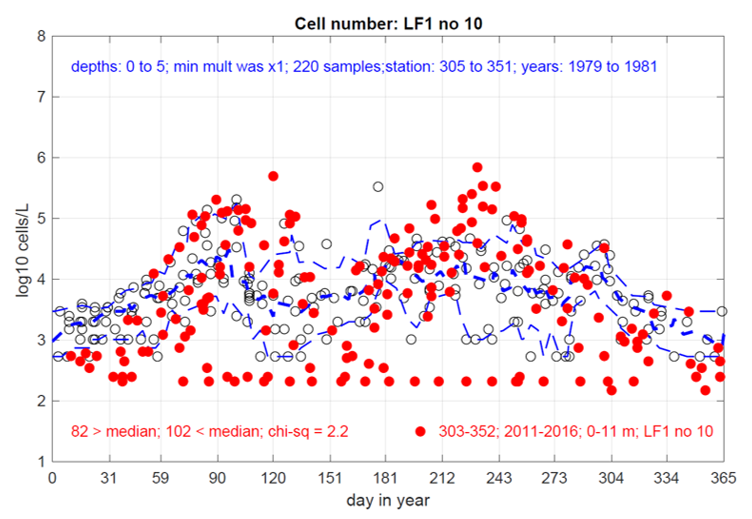
Figure 11. Comparison of Pseudo-nitzschia sp. in Loch Creran from 1979-1981 (Grey, clear circles) and 2011-2016 (red, full circles). Plot published with permission from P. Tett 2023.
Click for a full description
Scatter graph showing comparison of Pseudo-nitzschia species in Loch Creran from enveloped datasets gathered between 1979-1981 (grey, open circles) and 2011-2016 (red, full circles). Data are log transformed, the thick dashed blue line shows the median recorded counts in the 1979-1981 dataset and the thin dashed blue lines show the 10th and 90th percentiles of the 1979-1981 enveloped dataset. A chi squared test is applied to show a difference in the fit of the lines between the two enveloped datasets.
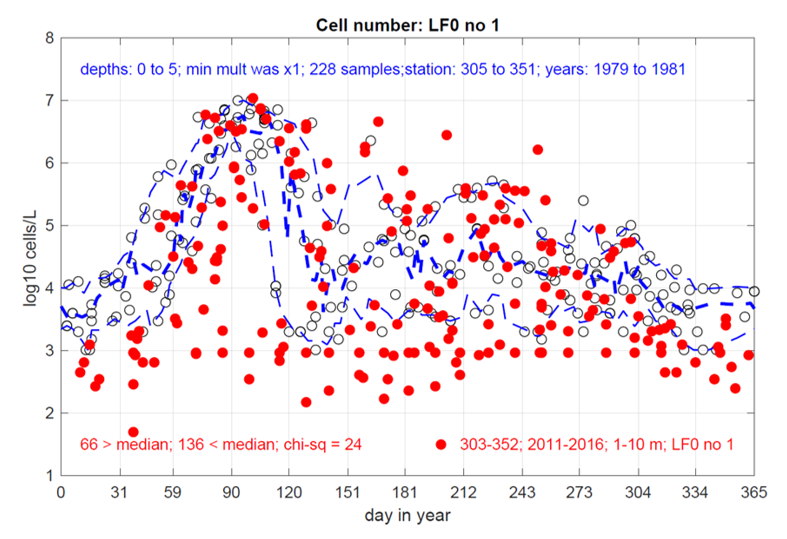
Figure 12. Comparison of Skeletonema sp. in Loch Creran from 1979-1981 (Grey, clear circles) and 2011-2016 (red, full circles). Plot published with permission from P. Tett, 2023.
Click for a full description
Scatter graph showing comparison of Skeletonema species in Loch Creran from 1979-1981 (grey, empty circles) and 2011-2016 (Full, red circles). Data are log transformed, the thick dashed blue line shows the median recorded counts in the 1979-1981 dataset and the thin dashed blue lines show the 10th and 90th percentiles of the 1979-1981 enveloped dataset.
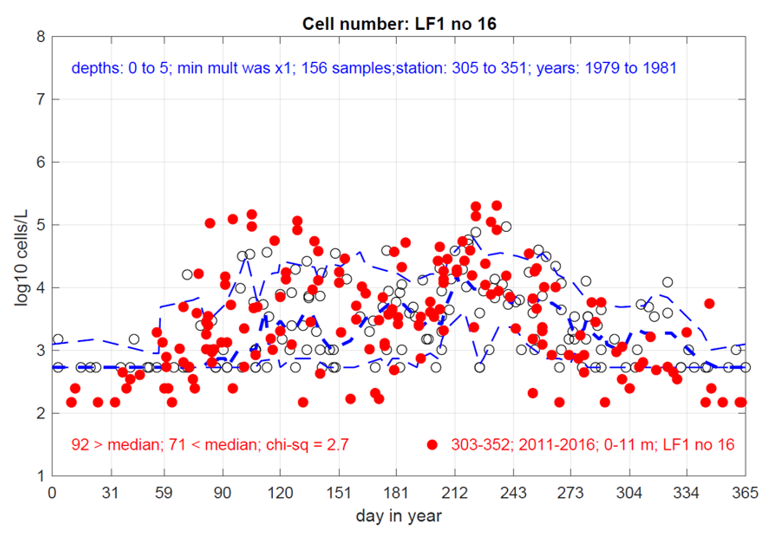
Figure 13. Comparison of Gonyaulacales in Loch Creran from 1979-1981 (Grey, clear circles) and 2011-2016 (red, full circles). Plot published with permission from P. Tett, 2023.
Click for a full description
Scatter graph showing comparison of Gonyaulacales in Loch Creran from 1979-1981. Data are log transformed, the thick dashed blue line shows the median recorded counts in the 1979-1981 dataset and the thin dashed blue lines show the 10th and 90th percentiles of the 1979-1981 enveloped dataset.
Human activities
This section lists the various activities which take place or have previously occurred within Loch Creran. While quantitative data was available for some activities, it was not possible to obtain information for all activities which occur in the Loch, which presents some challenges in terms of assessing trends over the timeframe of interest.
Aquaculture
Finfish aquaculture production has taken place in Loch Creran since Golden Sea Produce established the first farm in the 1970s (Black et al., 2000). Scottish Sea Farms Ltd. are currently the only finfish producer operating in Loch Creran.
South Shian
The Scottish Sea Farms processing plant at South Shian has held a consent to discharge prior to the publication of the Loch Creran Management Plan in 2007 (from grid reference NM 91030 41799, Figure 7). A benthic survey was undertaken in August 2001 which showed that there was no significant impact observed around the processing plant discharge point (Argyll and Bute Council, 2007). The discharge from this effluent pipe is monitored and recorded by SEPA and the information was shared for the purpose of this report (Figures 14-17). Units are reported but the rate of discharge (i.e., the amount of water per unit of time) is not reported.
Low levels of pH (~3.2) in the effluent water were recorded in 2003 in the outfall near the South Shian processing plant. However, pH did not decline to significantly low values again until 2018-2019 (Figure 14). The pH of seawater varies between 8.03-8.33 so in general, levels of pH from this outfall are lower than that of the surrounding seawater (Marion et al., 2011).
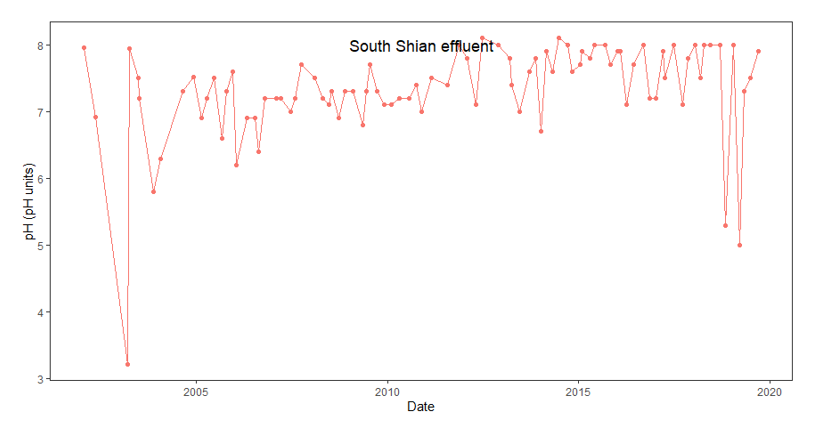
Figure 14. Regular measurements of pH in effluent from the outfall pipe at the South Shian processing plant from 2002 – 2020. Data and permission were provided by Scottish Sea Farms.
Click for a full description
Line graph showing pH values obtained from measurements taken at the outfall pipe associated with the South Shian processing plant between 2002 and 2020. The pH level begins just below 8 before decreasing significantly to just above 3 around 2003 before increasing to 8 again. The pH then fluctuates considerably but generally remains between 7 and 8 until around 2019 where two instances of pH below 5.5 occurs before returning to levels between 7 and 8.
The following chemical properties of the effluent are monitored by SEPA; ammoniacal nitrogen (amount of ammonia in mg/L), reactive phosphorous (mg/L), total oxidised nitrogen (sum of nitrate and nitrite, mg/L), chloride (mg/L) and pH. The following physical properties of the effluent were also measured by SEPA as part of the same monitoring programme; electrical conductivity (a measurement usually associated with suspended solids, pure water is 0.05 µS/cm at 25°C, drinking water between 200-800 µS/cm at 25°C), suspended solids (mg/L) and biochemical oxygen demand (mg/L). Records from 2005 suggest that the level of Chloride detected has increased, as seen by an increase in mg/L in the outfall pipe water (Figure 15), which corresponds with increased levels of electrical conductivity at 25 °C (Figure 15). The increases appear to begin between 2011-12 (Figure 15).
A peak in suspended solids is observed in 2007 and above normal biochemical oxygen demand in early 2010, while ammonia levels appear to have dropped in years 2010 – 2020 but have been below 100 mg/L since recordings began (Figure 16).
In 2008, 2014 and 2021 peaks of total oxidised nitrogen were recorded in the outfall water (nitrates and nitrites), 2008 recorded the highest levels at over 8 mg/L. An above normal amount of reactive phosphorous was recorded in 2005 from the pipe (just under 4 mg/L) but since then, nothing above normal was seen (Figure 17).
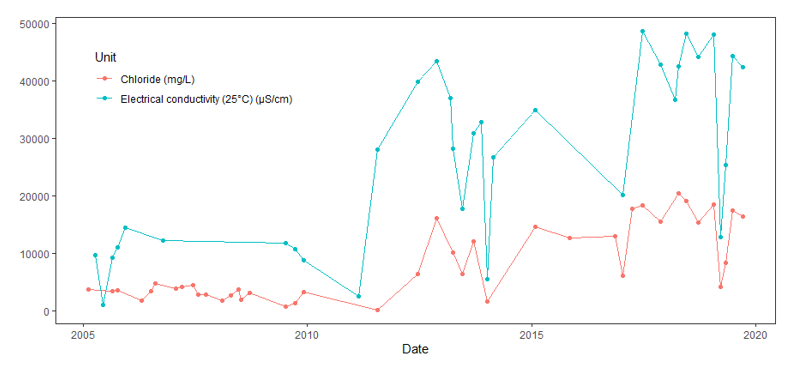
Figure 15. Measurements of the chloride and electrical conductivity of the water released from the outfall pipe at the South Shian processing plant. Units for y-axis are within legend box.
Click for a full description
Line graph showing measurements taken of chloride and electrical conductivity of the water discharged from the outfall pipe at the South Shian processing plant. Measurements were taken between 2005 and 2020. Chloride levels remained relatively constant between 2005 and 2010, before decreasing slightly until 2011 and then rapidly increasing between 2011 and 2012. Chloride levels beyond this year fluctuated considerably, although generally remained above levels prior to 2010, with one exception in 2019. Similarly, electrical conductivity remained relatively constant between 2005 and 2012, increasing around the same time period as chloride levels. Beyond 2012, electrical conductivity also fluctuated considerably but generally remained above levels observed prior to 2010.
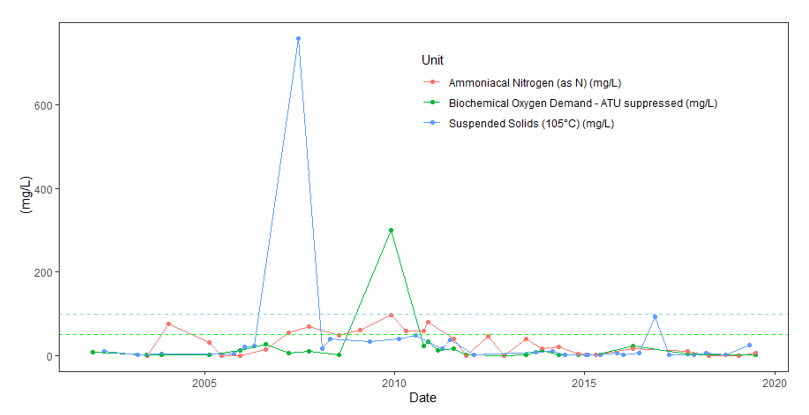
Figure 16. Measurements of ammonia, biochemical oxygen demand and suspended solids in water released from outfall pipe at the South Shian processing plant. Water treatment discharge limits for BOD and SS are represented by dashed lines.
Click for a full description
Line graph showing ammoniacal nitrogen, biochemical oxygen demand (BOD) and suspended solid levels in the effluent released from the outfall pipe at the South Shian processing plant between 2002 and 2020. Dashed lines of the same colour represent the discharge limits from the Environment Agency as set for water treatment works. Ammoniacal nitrogen levels fluctuate slightly between 2003 and 2014 but do not increase above 150 mg L-1. Biochemical oxygen demand and suspended solids remain relatively consistent throughout the same time period however, there are two notable peaks. An observed peak for suspended solids was observed around 2007, during which levels exceeded 600 mg L-1 while BOD experienced a peak of around 300 mg L-1 in 2010. An additional smaller peak was observed for suspended solids around 2017, although much smaller than the previous peak at around 100 mg L-1.
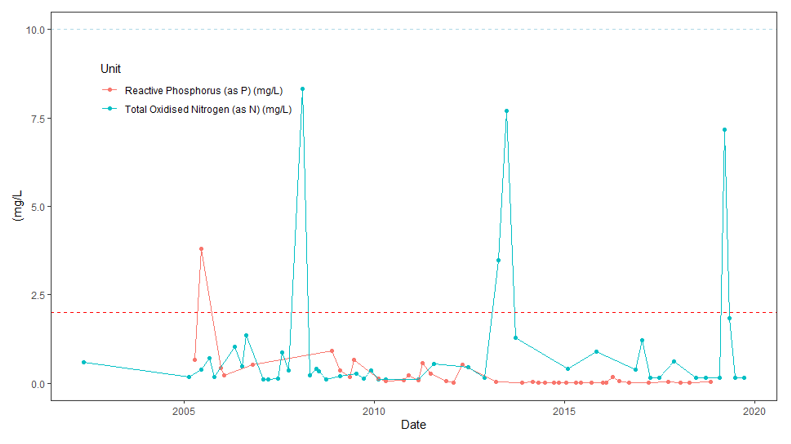
Figure 17. Measurements of reactive phosphorous (red) and total oxidised nitrogen (blue) in water released from outfall pipe at the South Shian processing plant. The Annual mean compliance limits for both chemicals are represented by dashed lines.
Click for a full description
Line graph showing measurements of reactive phosphorus (as P) and total oxidised nitrogen (as N) taken from the South Shian processing plant outfall pipe. The annual mean compliance limits are shown in dashed lines of the representative colour for each chemical group (these data are reported in the Environment Agency’s treatment and compliance limits). Reactive phosphorous has decreased since it was first measured in 2005, with a sharp peak shortly after measurements began in 2005. Total oxidised nitrogen measurements have been variable, with three peaks measured before 2020. The highest peak measured over 8 mg L-1.
Barcaldine Hatchery RAS (recirculating aquaculture system)
In 2018 a land-based hatchery was opened in Barcaldine. A discharge pipe, shared with the local Barcaldine Camping and Caravanning Club Site runs into the Loch (grid reference NM 96187 42400, Figure 7). Juvenile Atlantic salmon in the hatchery may be treated (‘bath treatments’) for water-borne micro-organisms (Saprolegnia sp) using formaldehyde and bronopol (bromo-2-nitropropane-1,3-diol). By condition of the CAR licence, these chemicals are only released into the Loch at high tide to increase the dispersal potential and ensure compliance with the licenced Environmental Quality Standards, minimising the risk to the marine environment.
An assessment to understand the impact of the Barcaldine Hatchery on chlorophyll distribution and nutrient concentrations was conducted in 2020-2021 (Anderson Marine Surveys, 2021). To investigate the potential effects of the hatchery, sampling and analysis of chlorophyll distribution and dissolved inorganic nitrogen (DIN) during winter and spring of 2020-2021 was performed. Overall, the surveys suggested no significant changes in chlorophyll or DIN in Loch Creran associated with the outfall pipe at the Barcaldine Hatchery. However, an increase in chlorophyll was observed in the summer of 2020. The summer mean concentrations were measured at between 3.0-3.3 mg(DIN)/m3, which is consistent with modelled predictions of cage and hatchery impacts.
Cage Sites
Two fish farm sites have consent to operate in Loch Creran (Site A and Site B) but they are operated in rotation. As such, there is currently only ever one site in operation in the Loch at any given time. The Creran A site has been in operation since 1983 (but was relocated from its original location in 2002) and the Creran B site has been farmed since 1989. Between September 1994 and May 1998, a maximum biomass of 1584 tonnes was reported (SSF, personal communication, 14 November 2022), which is largely consistent with the current maximum permitted biomass of 1500 tonnes. Biomass (and any associated nutrient input) in the Loch has therefore remained stable, with no significant increases since at least September 1994.
While active sites have been present since as early as 1983, the current pattern of production in the Loch was first introduced in 2002, when Site A was relocated from its original location in shallower water near Rubha Riabhach, to its current location in deeper water close to Site B (see Figure 10). A planning committee report from Jan 2002 indicates that the two sites were farmed in rotation prior to site A being moved in 2002. However, there appears to be no formal record of how long this rotational system of production had occurred for prior to this. Following the relocation of site A in 2002, the sites continued to be operated on a rotational basis, with a maximum biomass of approximately 1,500 tonnes being reached on a biennial basis, at only one of the sites at a time (Figure 18). This remains the current pattern of production in the Loch. The primary reason for moving Site A was to relocate it into deeper water to avoid overlap with serpulid reef habitat. Prior to 1998, when the cages were last present in the Rubha Riabhach area, it was considered likely that some physical damage to serpulid reefs had occurred as a result of the presence of the farm infrastructure. To prevent any further damage, and to allow recovery, the site was moved into deeper water closer to Site B.
Biomass in Loch Creran was notably lower in 2006-2008 (Creran A) and 2008-2010 (Creran B), which was not associated with farmed Atlantic salmon mortalities. However, in 2020 a salmon mortality event was recorded at Creran B with an associated loss of biomass (Figure 18).
Figure 18. Reported biomass in salmon aquaculture in Loch Creran (black columns), with reported mortalities (red bars).
Click for a full description
Column charts showing the reported biomass of salmon in tonnes for aquaculture sites Creran A and Creran B year is along x-axis. The charts indicate the rotation between the two sites with maximum biomass peaking at different time periods for each site. The maximum reported biomass for either site peaks at around 1500 tonnes which both sites showing slightly lower biomass between 2006 and 2010. Reported salmon mortalities are also included in the graph, with a mass mortality rate reported at the Creran B Site in 2020 which resulted in lower biomass for the site.
The benzoate salt of emamectin was introduced in the late 1990s with first registrations for use as an agricultural insecticide in the U.S. and Japan in 1997 (Jansson et al., 1997). Trials using emamectin benzoate to orally treat Atlantic Salmon (Salmo salar) infested with sea-lice (Lepeophtheirus salmonis) were conducted in Scotland in the late 1990s – early 2000s (Stone et al., 2000, 2002). Atlantic salmon have been produced in Loch Creran since 1983, but the location and methods used at the two cage sites changed in 2002 (Wilson et al., 2006). Regular monitoring of the pollutants released from Site A and Site B in Loch Creran began in 2002. Routine monitoring is carried out by Scottish Sea Farms under performance standards and is reported to the Scottish Environment Protection Agency (SEPA, see Scottish Pollution Release Inventory).
Fish farms are required to report any (above 0 g) release of emamectin benzoate, cypermethrin, deltamethrin and azamethiphos during treatment of salmon in fish farms. Use of these chemicals should be in accordance with limits detailed in the CAR license, which are established on the basis of site-specific modelling. Total quantities released are recorded by the Scottish Environment Protection Agency (Table 3). No data relating to chemical discharges is available prior to 2002. However, the available data shows that a range of chemical treatments have been employed in Loch Creran since at least 2002. The use of emamectin benzoate has remained relatively constant over the period, whereas the range and quantity of bath treatments has varied significantly. Between 2002 and 2021, release of emamectin benzoate into Loch Creran did not exceed 1 kg per year (Figure 19). Data shows that cypermethrin was in use between 2003 and 2009, with use appearing to increase significantly from 2007. Deltamethrin appears to have been introduced in 2009 and its use continued until 2017. However, peak use appeared to take place in 2009 and 2011, with reduced quantities applied in 2013, 2014 and 2017. Azamethiphos has been in use from 2012 to present, with the last recorded treatment occurring in 2022. However, its application appears to have peaked between 2013 and 2016. Use reduced significantly after this, with treatments taking place in only two subsequent months since (Oct 2021 and Jan 2022). In addition to information reported on Scotland’s Aquaculture website and the Scottish Pollutant Release Inventory (SPRI), two records of hydrogen peroxide release into the Loch are available. In 2013, hydrogen peroxide release was recorded as 5,544 litres and in 2015, 79,108 litres were recorded. However, it is understood that these quantities relate to the total product released, which is approximately 50% hydrogen peroxide, not the total quantity of hydrogen peroxide itself.
| Input | From-Until | Total |
|---|---|---|
| Azamethiphos (g) | 2012-2021 | 11870 |
| Cypermethrin (g) | 2007-2009 | 1264 |
| Deltamethrin (g) | 2007-2017 | 1081 |
| Emamectin benzoate (g) | 2007-2021 | 7579 |
| Nitrogen - total as N (t) | 2007-2017 | 762 |
| Phosphorus - total as P (t) | 2007-2020 | 122 |
| Total organic carbon or COD/3 (t) | 2007-2021 | 3077 |
| Zinc (kg) | 2007-2019 | 2333 |
| Copper (kg) | 2007-2017 | 12357 |
| Hydrogen Peroxide (litres) | 2013, 2015 | 5544, 79108 * |
* Only two records are present in SEPAs databases for hydrogen peroxide use.
The finfish aquaculture sector are also required to report on the release of certain metals such as copper (utilised as a feed additive and in some cases as a net antifoulant) and zinc (also included as a feed additive). The reported level of copper released from the farms appears to have increased from 2007, peaking in 2014 thereafter reducing significantly with very low levels reported from 2018 onwards (Figure 20). Any nutrients (N, P and C) released into the water column are also reported with no significant increases in nutrients obvious from the dataset (Figure 21). As biomass in the Loch has remained stable since at least 1994, it is unlikely that finish production has contributed significantly to any increased nutrient input over the period.
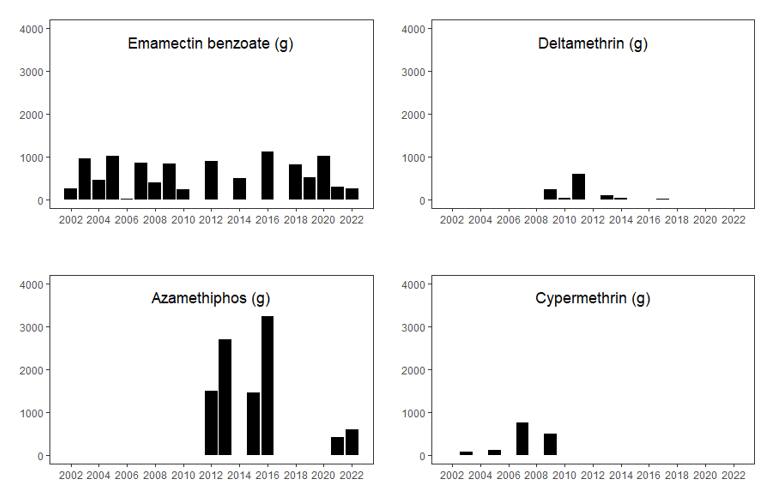
Figure 19. Reported annual inputs from Loch Creran aquaculture (fish farm) sites A and B of sea-lice treatment chemicals released into the Loch from 2007-2021. Information downloaded from the Scottish Environment Protection Agency (SEPA).
Click for a full description
Figure 19 shows the reported annual inputs from Loch Creran aquaculture (fish farm) sites A and B (alternating sites) of sea-lice treatment chemicals released into the Loch from 2007-2021 (x-axis). This includes Emamectin benzoate, Deltamethrin, Azamethiphos and Cypermethrin. No teflubenzuron is reported in use in Loch Creran. The data are open source and were downloaded from Scotland’s Aquaculture website. A monthly breakdown of these data are shown in Appendix 2.
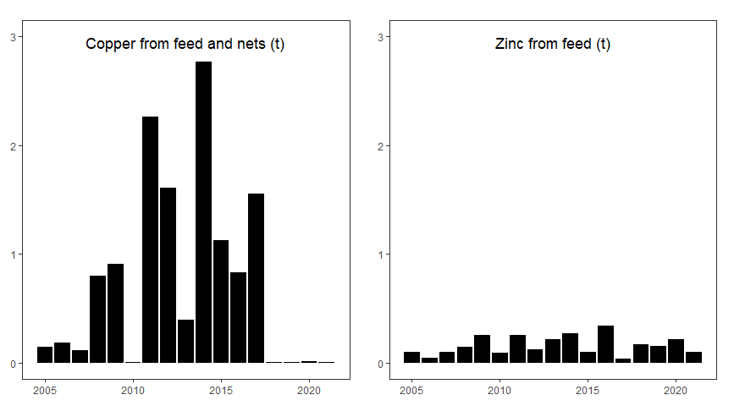
Figure 20. Reported annual inputs from Loch Creran aquaculture (fish farm) sites A and B of heavy metals released from fish farm infrastructure into the Loch from 2005-2021. Information downloaded from Aquaculture Scotland, also available from SEPA.
Click for a full description
Bar graphs of reported annual inputs of copper from feed and nets and zinc from feed from Loch Creran aquaculture sites A and B (alternating sites). Between 2005 and 2021 (x – axis), copper from feed and nets has been variable, with highest levels in 2014 and 2011. From 2018 to 2021, input has been low. Annual input of zinc from feed has been variable, with highest levels in 2016.
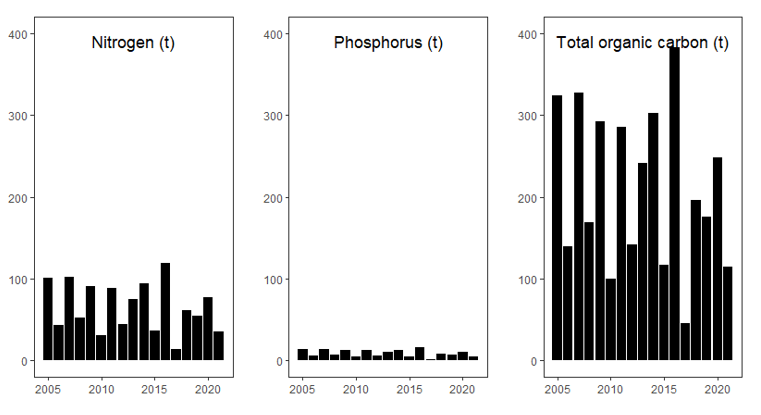
Figure 21. Reported annual inputs from Loch Creran aquaculture (fish farm) sites A and B of organic matter released into the Loch from 2005-2021. Information downloaded from Scottish Aquaculture.
Click for a full description
Bar graphs of reported annual inputs of nitrogen, phosphorous and total organic carbon (TOC) from Loch Creran aquaculture sites A and B (alternating sites). Between 2005 and 2021 (x – axis), nitrogen has been variable, with highest levels recorded in 2016. Lowest levels were recorded in 2017. Annual input of phosphorus has been variable, with highest input also recorded in 2016 and lowest levels recorded in 2017. Annual input of TOC again recorded highest input in 2016 and lowest input in 2017.
Shellfish harvesting and monitoring
FSS is responsible for ensuring that shellfish from designated harvesting areas meet the health standards laid down in Retained EU Regulation 853/2004. Classifications are assigned according to the FSS Protocol for Classification and Management of E. coli results. Routine Official Control (OC) sampling is carried out by FSS on a monthly basis with occurrences published on their website however, this data is only available from 2011 onwards. The data is, however, available from Scotland’s aquaculture website for E. coli levels surrounding the oyster farm on the south shore of Loch Creran from 1999 onwards (Figure 22). These data sets show three high recordings of E. coli in the region where the number of cells L-1 exceeded 15,000 in 2003 and 2020 and was recorded just under 10,000 in 2010 (Figure 23). Specifically, E. coli were measured at 18,100 per 100g on 18 June 2003, at 9,200 per 100g on 12 July 2010 and 1 July 2020, and at 18,000 per 100 g on 4 August 2020 (Figure 23).
SEPA monitoring of shellfish growing waters in Loch Creran show the compliance history for faecal coliforms in the water body from 2000-2010. Over this ten year period guideline standards for faecal coliforms were only achieved in three of the ten years (2001, 2002 and 2003), with the remaining years failing to meet standards. The Loch Creran, SEPA report (2011) identifies the most likely reason for these failures to be diffuse source pollution from livestock farming and/or sewage disposal.
Riverine input and general water quality
Sanitary surveys (‘Sani surveys’) have been conducted in the large outer basin of Loch Creran by Cefas, the Food Standards Agency (Scotland) and SAMS in 2007, 2013 and most recently in 2021. The main purpose of the reports is to identify any unsanitary water, of either human or animal origin, flows into the Loch which might impact near-shore fisheries. There were a significant number of septic tanks registered adjacent to Loch Creran between 2007-2013 (total 68 discharge points) and 16 discharges which flowed directly into the Loch. While a variety of sewage outlets were reported in the 2021 survey, they were not counted. High levels of E. coli in certain watercourses (two tributaries at Shian) were reported in 2013 but not in 2007 or 2021.
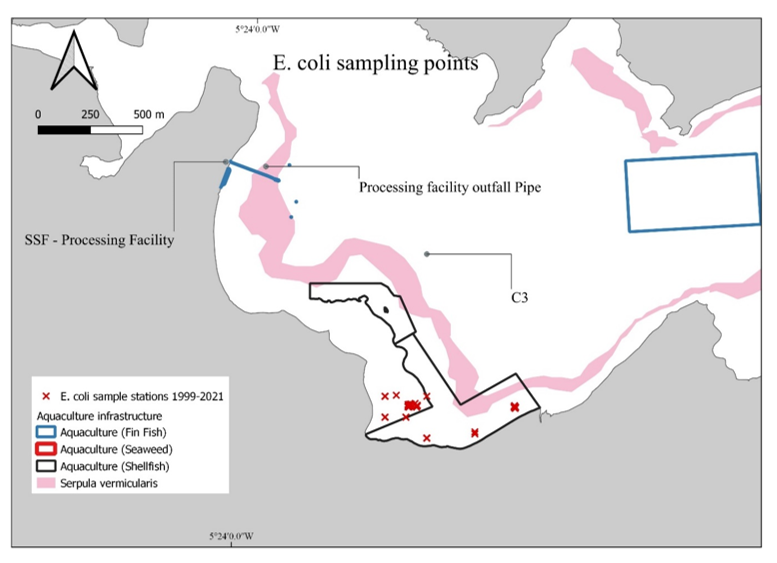
Figure 22. E. coli sampling points from 1999-2021 in Loch Creran as recorded by Scottish Aquaculture. Most samples were taken around the current oyster farming site.
Click for a full description
Data on S. vermicularis presence were downloaded from the GeMS habitat database, aquaculture agreements were downloaded from Crown Estate Scotland.
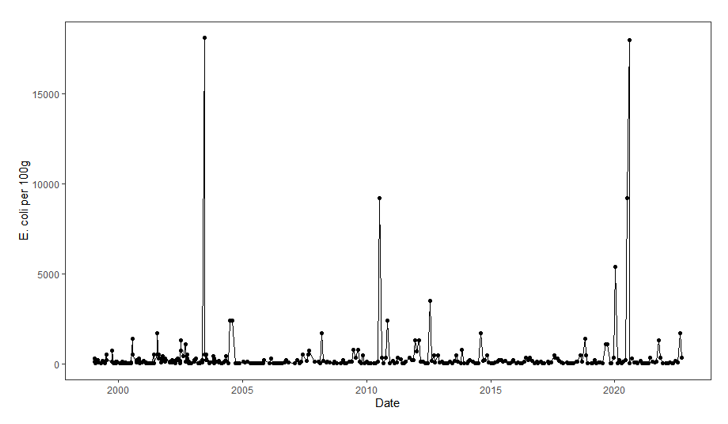
Figure 23. Recorded E. coli counts per 100 g of water measured at Rubha Mor in Loch Creran, near the oyster aquaculture sites
Click for a full description
Chart of E. coli counts per 100 g of water measured at Rubha Mor in Loch Creran. The graph shows several peaks between the years of 2000 and 2022. Two of these peaks are greater than 15000 E. coli per 100 g. There were 11 occurrences of values in this dataset that were indicated as less than (<, meaning insignificant) which were plotted as integers removing the < symbol from the dataset.
Loch Creran is not listed as one of the Scottish bathing waters (SEPA, 2016) but three regions in Loch Creran have been monitored for multiple chemicals by SEPA. Beginning in the 1980s with the River Creran, and then subsequent (less frequent) monitoring of two points in Loch Creran at North Shian and at the Creran Bridge which has data from 2000 – 2005 (Figure 24).

Figure 24. Regular chemical testing by SEPA of the River Creran began in the 1980s and two sites have been tested from 2000-2005 deeper into the two Loch basins at the Creagan Narrows (Creran Bridge) and at North Shian.
Click for a full description
Data on S. vermicularis presence were downloaded from the GeMS habitat database, aquaculture agreements were downloaded from Crown Estate Scotland.
There are no major changes or trends in alkalinity or pH (Figure 25) in the available data for the River Creran. The pH data for Creagan Bridge and North Shian do not appear to show obvious change but are insufficient to effectively show long-term change (Figure 26). Similarly with temperature and dissolved oxygen (DO) for the aforementioned sites (Figure 27).
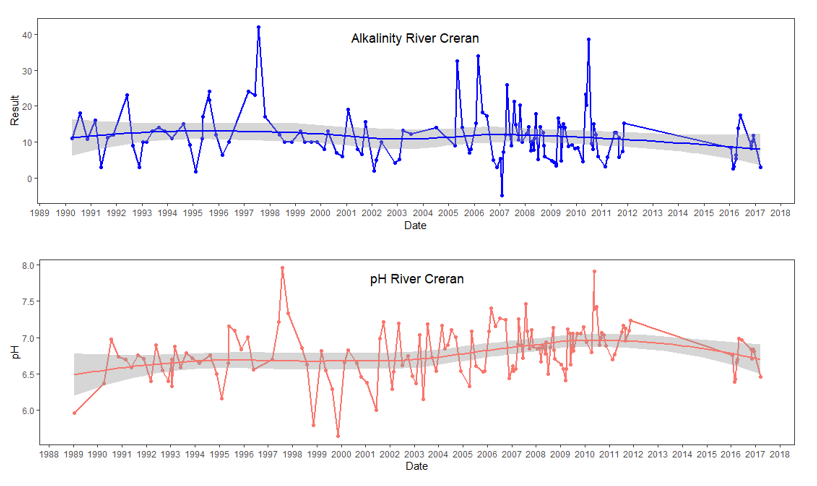
Figure 25. Measurements from River Creran which flows into the Loch from the east (landward) side. Alkalinity is measured as calcium carbonate (CaCO3) in mg L-1, pH is measured in pH units.
Click for a full description
Blue and red lines represent fitted loess regression curves with standard error in grey shading. Alkalinity in River Creran between 1990 and 2017 shows fluctuating levels with the highest peak over 40 mg/L in 1997. pH in River Creran between 1989 and 2017 shows fluctuating levels over time with the highest peaks in 1997 and 2010.
Figure 26. The pH of water at the Creagan Bridge and at North Shian as measured by SEPA between 2000 and 2005. There appears to have been a drop in pH between 2001 and 2002 measured below 7.0 at both sites.
Click for a full description
A line graph of pH at two locations in the Loch Creran between 2000 and 2005. At Creagan Bridge, pH level decreases below 7 in 2001, before increasing to above 8 in 2002, and remaining around 8 to 2005. At North Shian, pH level decreases from 8 in 2001 to below 7 in 2002. This then increases and remains around 8, although there is fluctuation.
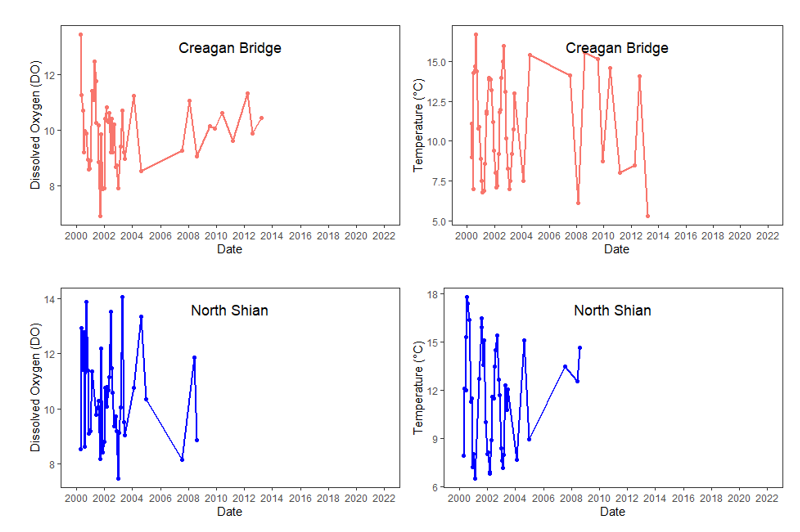
Figure 27. Dissolved oxygen (left) and temperature (right) as measured by SEPA in Loch Creran at two locations in the outer basin.
Click for a full description
Four line graphs depicting dissolved oxygen (DO) and temperature (°C) measurements taken between 2000 and 2014 for Creagan Bridge, and 2000 and 2010 for North Shian. Dissolved oxygen values fluctuate for both sites between 7 and 14 ml L-1, while temperature fluctuates between 5 and 18 °C.
Metals were also recorded by SEPA in the water of North Shian, Creagan Bridge and in the River Creran waterways. For the two sites in Loch Creran, water metal concentrations appear relatively consistent apart from in 2004 where zinc levels reached over 30 µg/L at Creagan Bridge and over 6 µg/L at North Shian (Figure 28).

Figure 28. Metals measured in two sites in Loch Creran, all metals plotted are those that can pass through a 45 µm membrane. Concentration is reported in µg/L.
Click for a full description
Two line graphs of metal concentrations at two locations in Loch Creran; North Shian and Creagan Bridge. Zinc shows the highest concentrations at both locations. Fluctuation can be seen in the concentrations. Zinc has a peak in 2004, with higher levels seen at Creagan Bridge than at North Shian.
Mussel production
One point to note from the Sanitary surveys is the observed cessation of blue mussel (Mytilus edulis) aquaculture. In 2009, approximately 500 tons of M. edulis were produced in Loch Creran (Ferreira et al., 2009). However, at some stage since then M. edulis production in the Loch ceased. The Loch Creran Sanitary Survey Review report, published in April 2013 noted that the two mussel farms appeared to be in a semi-derelict state, suggesting that the sites were no longer in active production at this stage. Alongside this, a decline of natural populations has also been recorded at one location. Mytilus edulis decline was observed during surveys conducted by researchers as part of a long-term nationwide monitoring project known as ‘MarClim' (see Mieszkowska et al., 2005). As part this survey, a sample station at the old Sealife Centre was monitored in 2003 and then subsequently in 2022 (for sample site location see Figure 7). Mytilus edulis was recorded as ‘Abundant’ (20-49% cover) in 2003 but ‘Absent’ at the same place in 2022. After 45 minutes of searching, no mussels were seen at the site (Figure 29, personal communications, M. Burrows, November 2022). This is consistent with trends observed across coastal waters in the North Atlantic (Baden et al., 2021; Barbosa et al., 2022).

Figure 29. Loss of the filter feeding mollusc, the blue mussel (Mytilus edulis) has also been observed in certain parts of Loch Creran. The images were taken in 2003 (left) and 2022 (right) at the same sample site. Image credit M.T. Burrows.
Oyster production
Overall, the production of Pacific oyster (Crassostrea gigas) in the Loch Creran area has increased between 2009-2021 with combined table and on-growing production increasing 167% (personal communications, Marine Scotland Science, November 2022).
Fishing
Loch Creran once provided grounds for the dredging of King (Pecten maximus) and Queen (Aequipecten opercularis) scallops. Historically, benthic trawling was carried out in Loch Creran (particularly the deeper, muddier sediments) for prawns (Nephrops norvegicus). Trawl gear could come into direct contact with the serpulid reefs when the gear was manoeuvred, due to the restricted size of the Loch. Trawling could also generate plumes of sediment with the potential to smother the reefs.
The use of bottom-contact mobile fishing gear associated with dredging and the use of benthic trawling gear have potential to damage biogenic reef features, such as serpulid reefs. As a result, on 29 March 2007, the Inshore Fishing (Prohibited Methods of Fishing-Loch Creran) Order 2007 came into force to prohibit scallop dredging and benthic trawling within the Loch. This legislation has been superseded by Inshore Fishing (Prohibition of Fishing and Fishing Methods) (Scotland) Order 2015 which prohibits all mobile gear fishing in Loch Creran.
Creeling for prawns (Nephrops norvegicus) and velvet swimming crab (Necora puber) also took place in Loch Creran in the past. Creels can weigh over 20kg and when dropped in fleets on the reefs, could potentially result in serious damage to their structure. Recovering the creels could also potentially damage the reef if they were dragged along the seabed. The Inshore Fishing (Prohibited Methods of Fishing) (Loch Creran) Order 2007 permitted creel fishing only outside designated exclusion zones (where reef occurred) and within the permitted creeling area in the upper Loch. This legislation has been superseded by Inshore Fishing (Prohibition of Fishing and Fishing Methods) (Scotland) Order 2015 which restricts creel fishing to the deepest parts of the Loch away from the reef.
Boating and moorings
Loch Creran experiences regular marine traffic of small- to medium-sized vessels, with larger vessels only occasionally passing through the area. Activity is mostly concentrated around the depot at Rubha Garbh, which serves the quarry at Glensanda, and a pier complex at the Barcaldine Hatchery and the adjacent Creran Marina, which serves both industrial and recreational vessels. While no industrial activity takes place at the Rubha Garbh depot, the pier complex at Barcaldine has a marina/boatyard, a fish hatchery and several other small-scale industrial activities. The marina at Barcaldine offers between 70 and 100 moorings, with yacht and pleasure craft activity increasing during the summer months. Larger workboats and pleasure craft are also frequently observed at the fish processing facility at South Shian (Scottish Sea Farms).
Mooring and anchor placement for both recreational and commercial use has the potential to cause significant damage to serpulid reefs. There are currently numerous leisure mooring agreements around Loch Creran, leased by Crown Estate Scotland, with recreational vessels frequently observed during summer months. It has previously been acknowledged that removing existing mooring placements may cause more damage than leaving them in place, as removal may cause damage to biogenic reefs which have grown over the area. Personal communications with local NatureScot staff indicate that marine licences have not been renewed where moorings were identified to be overlapping with reef habitat. Sailing directions were also edited to remove anchorages in locations where reefs were present, and a sign was erected at Eriska Narrows to direct vessels to anchorages where no reef is present (J Dodd, NatureScot, personal communication, 09 March 2023). Anchoring is more likely to cause damage than mooring placements, as they may be dropped indiscriminately, and will generally drag before settling. However, while existing moorings may not contribute to a further decline, it should be noted significant damage can occur in discreet areas around individual moorings as a result of the scour effect of the mooring chain moving across the seabed.
Vessel activity
The Marine Management Organisation (MMO) keeps records of vessel movement throughout the UK. Vessels are fitted with an Automatic Identification System (AIS) to provide information for the MMO mainly for the purpose of safety. Datasets are available for download and provide insight into the types of vessels that are active in Loch Creran (see AIS Derived Track Lines data, Figure 30). The outer, larger basin is the main region of activity in the Loch, the upper basin of the Loch is beyond the Creagan Bridge, preventing large vessels from entering. As a result, the upper basin remains relatively free of boat activity and since the ban on trawling and dredging in 2007 the region has been clear of most boating activity. The three busiest points of activity are the South Shian Processing facility, the Glensanda Quarry port and the Barcaldine Hatchery (Figure 30). Vessel tracks do not appear to pass through the reef-occupied regions in Loch Creran. A combined dataset collated by Marine Scotland shows the average weekly shipping density of all vessels, at a 2km grid resolution (see Marine Scotland Information for further details and links to maps on NMPi). Loch Creran is divided into 9 x 2km grid squares and the busiest point in the Loch is the region around Eriska and the South Shian processing plant, with records between 50 – 150 transects a week on average (see Figure 31). A high proportion of this activity relates to the Glensanda Quarry ferry, operating from Rubha Garbh, which functions as a passenger vessel for workers at the quarry site on the opposite side of Loch Linnhe.
There are several vessels in operation in Loch Creran, most serving the aquaculture sector, with three passenger ferries which carry staff and other passengers to the Glensanda site. The draught beneath these boats ranges from less than 1 m (passenger ferries) to 4.8 m on the larger thermo-licer vessels used by aquaculture in the Loch (Table 4).
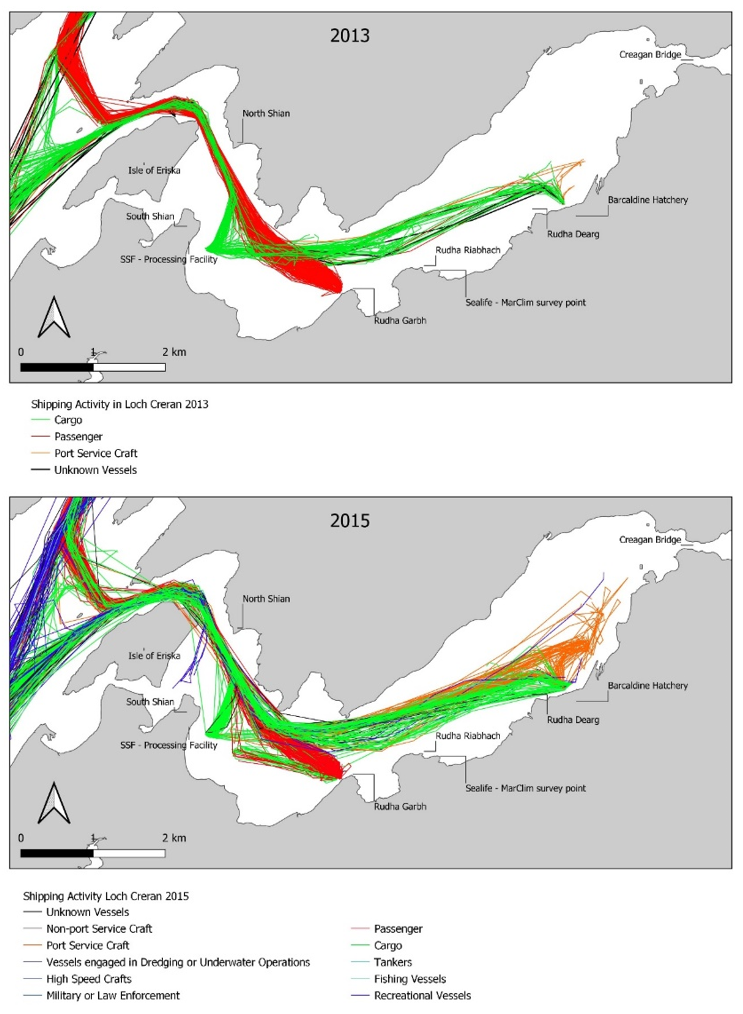
Figure 30. Map displaying major shipping activity within Loch Creran for cargo, passenger, port service craft and unknown vessels.
Click for a full description
The major activity in Loch Creran comes from cargo ships in and out of the three processing facilities on shore, the South Shian processing plant, Glensanda quarry port and the Barcaldine Hatchery. The types of vessels are predominantly cargo vessels (green), passenger vessels (red) and port service craft (orange) in a) 2015, and b) 2013. Limited recreational vessels are active in the Loch and since the creation of the SAC in 2005.
Figure 31. Average weekly shipping transects from the period 2012-2017, using AIS Shipping Traffic Data.
Click for a full description
Loch Creran is divided into 9 x 2km square grids, the busiest of which sees transects from most vessel types on average between 50-150 per week. NOT FOR NAVIGATION. © Crown copyright and database rights (2022). OS 100024655. OceanWise EK001-20140401. Use of this data is covered by copyright and privacy & content disclaimers. © OpenStreetMap contributors, tiles © CARTO.
| Vessel Name | Type (listed) | Use | Sector | Length (m) | Draught (m) |
|---|---|---|---|---|---|
| Simon Princess | Standby Safety Vessel | Thermolicer | Aquaculture | 42.25 | 4.8 |
| Fox Challenger | Tug | Thermolicer | Aquaculture | 29.8 | 2.8 |
| Sally Ann | Tug | Hydrogen peroxide tanks | Aquaculture | 23 | 2.4 |
| Helen Rice | Other | Not Known | Aquaculture | 21 | 2 |
| Viking Saga | Fish Carrier | Fish Carrier | Aquaculture | 42 | 4 |
| Aqua Lass | Tug | Cage Cleaning | Aquaculture | 26 | 2.8 |
| Ocean Aquila | Fish Carrier | Fish Transport | Aquaculture | 40.06 | 4.6 |
| Ella May | Unknown | Crane/barge | Aquaculture | 14 | 2 |
| Paul B | Tug | Cage tower | Aquaculture | 19 | 2.1 |
| Rose of Lorne | Pontoon | Passenger/car ferry | Glensanda | 36.4 | NA |
| Lady Iona | Passenger | Passenger/ferry | Glensanda | 17 | NA |
| Lady Heather | Passenger | Passenger/ferry | Glensanda | 15 | NA |
Tourism and recreation
Marine recreation and tourism is a significant sector within the wider Argyll and Bute region, with Loch Creran offering multiple activities including sailing, sea kayaking, wildlife watching and SCUBA diving. As a result of the wide variety of activities on offer, marine traffic is significantly increased within the Loch. Marine tourism has important economic implications for the area, accounting for a significant number of jobs and supporting other infrastructure within the sector such as hotels, guesthouses, B&Bs, restaurants and marinas. There were no quantitative data available at the time of review for activities that occur in the Loch therefore trends over the period of interest could not be assessed.
Agriculture
In Argyll and Bute agriculture is an important industry for economic output. Over a number of centuries, agriculture has shaped the natural environment and landscape of the region. On the northern and far western shores of Loch Creran there is extensive livestock agriculture. Many agricultural activities are regulated under The Control of Pollution (Silage, Slurry and Agricultural Fuel Oil) (Scotland) Regulations 2001. Farmers are also encouraged to demonstrate good practice which is stated in documents such as the Four Point Plan, the Farm Soils Plan, and the Prevention of Environmental Pollution from Agricultural Activity (PEPFAA) Code (Argyll and Bute Council, 2007).
Forestry
Deforestation of regions close to shore may increase freshwater run-off which will lead to increased input of nutrients, turbidity and sediments as well as potentially impacting pH (Howe et al 2010). Forestry and Land Scotland (FLS) manage the majority of the commercial forestry surrounding Loch Creran. The region of Glen Creran is carefully managed with a clear-fell and re-stock approach. Additionally, felling of trees on private land requires permission and all the data is open source. The National Forest Inventory has information on tree felling, ground clearing and inventories on forest and grassland regions. Between 2011 and 2018, felling occurred in the region of the old Sea Life Centre and in parts of the upper basin close to shore (Figure 32). Specific dates of the felling operations were not available. Private applications to fell trees are observed at the old Sea Life Centre, near Rubha Mor and in regions north of the Loch, but it is not clear how much of the permitted felling has taken place to date (Figure 33).
Ground preparation for new planting may involve the use of excavators or similar machinery to clear and prepare new planting positions ready for replanting of a felled site. One form this can take is mounding, where raised positions are created. Another is scarification, where ground vegetation is removed, dependent on the site. Access tracks and drainage are also installed at this time, in order to facilitate maintenance and deer control and to reduce waterlogging. Some sites are not prepared mechanically and are planted without it, such as steep sites or where there are very shallow soils. On average across the West Region, FLS sites lay fallow for 1-2 years after felling. The length of fallow period depends on various factors; soil types and vegetation issues, risk from weevil and future prescription. Some sites are left to naturally regenerate rather than being planted. This can also be affected by tree supply and weather during the planting season (FLS, personal communication, 22 November 2022).
In recent years, there has been no fertiliser used by FLS in Glen Creran. The only pesticide/herbicide use has been glyphosate for rhododendron control. This is applied by stump treatment (following cutting) and then minor follow-up foliar spray. Work of this type has not been carried out since 2020 (FLS, personal communication, 22 November 2022).
There has also been a water catchment agreement in place with Marine Resource Centre Ltd. (MRC), now Scottish Sea Farms, since 2000. This agrees to no pesticide or fertiliser use within the Loch Dubh Catchment in order to ensure water quality feeding into the lochan/reservoir. Although this would only affect the lower basin, this may provide additional benefits for the upper basin (FLS, personal communication, 22 December 2022).
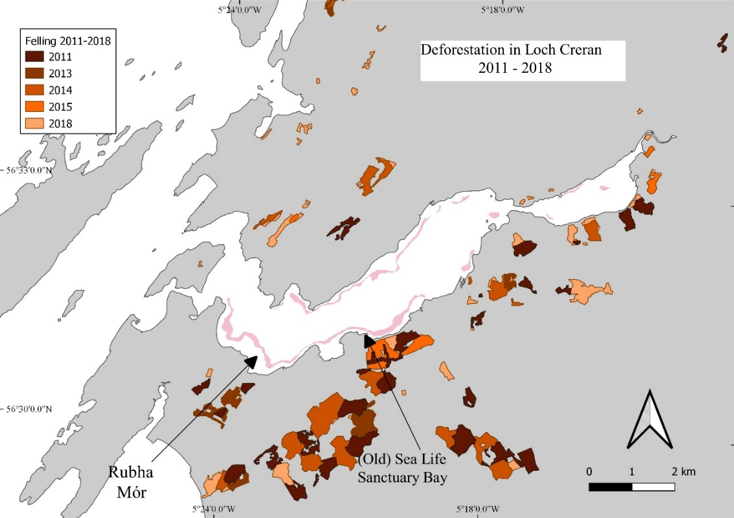
Figure 32. The state of forestry in the region surrounding Loch Creran from the National Forest Inventory (NFI).
Click for a full description
Data downloaded from
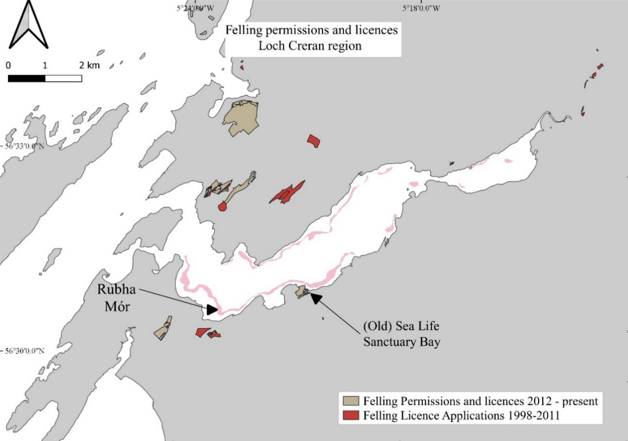
Figure 33. Felling permissions and licences in the region surrounding Loch Creran.
Click for a full description
Felling Permissions (before 2012 known as Felling Licence applications) received by Scottish Forestry and administered using Scottish Forestry’s case management system.
Figure 34. Map showing areas that have been restocked between 1992 and 2013; assuming a two-year fallow period between felling and restocking, this would approximate to areas felled between 1990 and 2011.
Observations of the invasive ascidian Didemnum vexillum
Systematic monitoring of sites in Loch Creran for the invasive ascidian Didemnum vexillum began in 2017 (Moore & Harries, 2020). The subtidal serpulid reef monitoring programme was initiated as the invasive, mat-forming species was first recorded in Loch Creran in 2016 (Moore & Harries, 2020). Between 2017 and 2021, 12 subtidal transects and 7 artificial structures were surveyed annually. No observations of D. vexillum were recorded on any of the occasions, leading surveyors to conclude that the ascidian is not exerting any pressure on S. vermicularis in the Loch (Moore & Harries, 2020).
Alginate factory
During World War 2 (1941) the Ministry of Supply built an alginate factory on the shores of Loch Creran. After the war it was renamed to Alginate Industries Ltd (Donovan, 2006). Production peaked in the 1970s and at that time it was discharging alkaline liquid whey, an acidic leachate, formaldehyde, and organic particulates directly into Loch Creran (Black et al., 2000). Operations ceased at the factory in 1996 but much of the waste that had already been discharged had coated the seabed near the effluent pipe which caused a large surface mat of Beggiatoa sp. (a Gammaproteobacteria of the order Thiotrichales) (Moore et al., 1998). The cessation of alginate waste entering Loch Creran as a result of the Alginate Factory is discussed as an external cause of nutrient reductions over the course of three decades by Laurent (2009). The precise composition of effluents is unknown but solid discharge of seaweed remains, after alginate is extracted is thought to contain significant quantities of carbon, phosphorus and nitrogen in particulate and dissolved (inorganic and organic) forms (Jones, 1979; Laurent, 2009). In 2007 the former alginate factory was purchased by Marine Resource Centre Ltd. (MRC) with a consent in place to discharge trade effluent from a smolt production unit into the Loch. At the time, it was not being used and was not expected to be used in the foreseeable future (Argyll and Bute Council, 2007).
In 2016, the MRC was purchased by Scottish Sea Farms (SSF), with plans to build a new land-based freshwater salmon hatchery at the site (Findlay, 2016). This is now the Barcaldine Hatchery, mentioned in the section Aquaculture.
Summary
Compilation of a detailed timeline of environmental and anthropogenic change in Loch Creran over the period 1980-2022 is hindered by the incompleteness of the available data. For example, data on some potentially important parameters (such as phytoplankton biomass and community structure) are only semi-quantitative and are based on a small number of measurements taken at widely-separated intervals. In the case of human activities, such as the quantities of chemical treatments used in salmon farming, or the extent of deforestation in the Loch Creran watershed, quantitative data are available, but there is no information on the degree (if any) to which the serpulid reefs experienced an environmental change as a result of these actions. The following list summarises the main findings of the Environmental Data Analysis, highlighting any changes or events coincident with the observed period of serpulid reef deterioration, taken to have begun in 2011 (at latest) and extending to the present day.
- West of Scotland climatic data show a general trend of increasing air temperature, sea surface temperature and rainfall from 1971 to the present, but there is no evidence for any singular event or alteration in the rate of change coincident with the period of serpulid reef decline.
- Phytoplankton biomass in Loch Creran appears to have declined substantially since the 1970s, accompanied by a shift in community dominance from diatoms to dinoflagellates. However, the patchy and semi-quantitative data do not allow these long-term trends to be plotted in detail and it is not known whether any significant changes took place in or around 2011.
- The potentially-toxin producing diatom genus Pseudo-nitzschia showed higher abundance in 2011 than at any time since, but there are no earlier data to put this peak into longer-term context.
- Atlantic salmon production has taken place in Loch Creran throughout the known period of serpulid reef occurrence (1980s – present) but at a fairly constant level and with no trend towards higher biomass or increased organic carbon discharge from cage sites.
- A variety of anti-sea lice chemical treatments have been employed since at least 2002 and data presented include those discharged by well boats. Azamethiphos application was highest from 2012-2016, but with little subsequent use. This reflects a broad trend across the wider industry, with use of Azamethiphos peaking around 2016/17 but reducing in the years since. Reported quantities of Emamectin benzoate have remained at relatively constant levels from 2002 to the present.
- Escherichia coli counts in water discharged into Loch Creran show intermittent high peaks compared to the normal background levels but with no increasing trend. Other measured parameters in seawater, outfall or riverine input show no evidence for a decline in water quality in the Loch.
- Blue mussels (Mytilus edulis) have suffered a decline in Loch Creran. This is expressed by an observed disappearance of wild populations at one monitoring station between 2003 and 2022. This decline broadly coincides with the observed deterioration of serpulid reefs. However, this does not appear to be unique to Loch Creran with similar declines observed across coastal waters throughout much of the North Atlantic (Baden et al., 2021; Barbosa et al., 2022). The blue mussel cultivation sector ceased at some stage after 2009, with evidence to suggest that former mussel farms sites had become derelict by 2013.
- Mobile fishing gear has been shown to cause localised damage to the serpulid reefs. However, its use in the Loch has been restricted to avoid overlap with areas of serpulid reef since 2007 and prohibited entirely since 2017.
- There has been extensive tree-felling in the Loch Creran watershed since 2011, especially in areas to the south of the Loch. However, there are no data to indicate whether this activity has led to any increase in freshwater runoff into the Loch, or, if so, whether the serpulid reefs would suffer any adverse consequences from this.
- A range of other commercial and recreational activities take place in and around Loch Creran but there are few or no data on their frequency, intensity or changes over the period of interest.
Figure 35 provides a visual summary of the health of the serpulid reefs using information extracted from the literature review. The available data on environmental and anthropogenic changes in Loch Creran which have been discussed throughout this report section has been used to create timelines which can be seen in Figures 36 and 37.
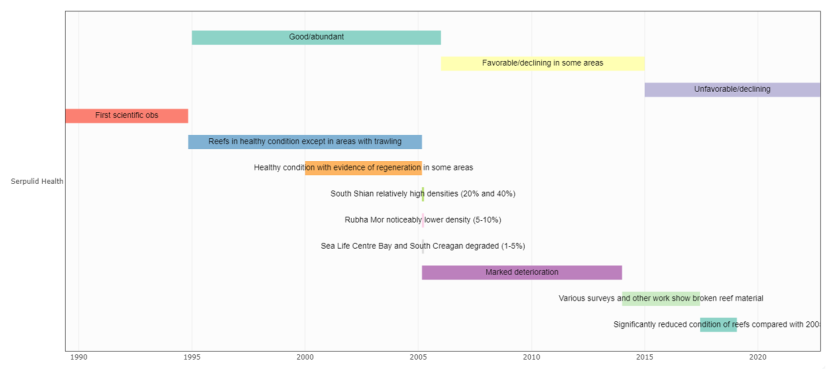
Figure 35. Available information from published literature showing the decline of S. vermicularis in Loch Creran, highlighting the key time points when S. vermicularis were considered in decline throughout the whole of Loch Creran.
Click for a full description
A timeline from 1990 to present day of Serpula vermicularis condition within Loch Creran based on observations in literature. The reefs were considered in good condition between 1995 and 2006/07, when reef decline began to be observed in some areas of the Loch. From 2015 onwards, the reefs as a whole have been considered unfavourable/declining.
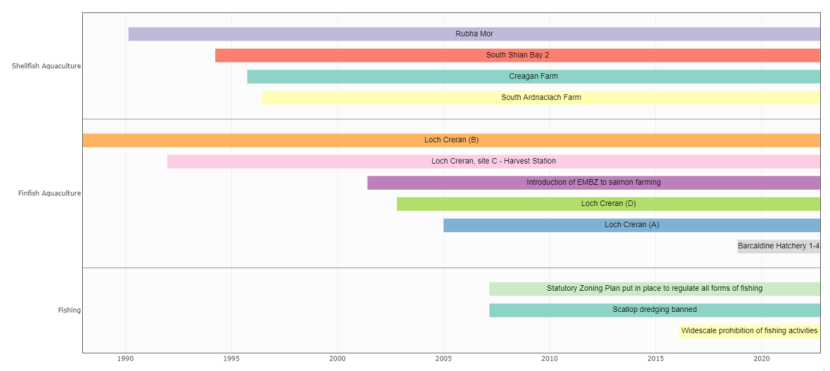
Figure 36. A timeline of fishing and aquaculture activity in Loch Creran.
Click for a full description
A timeline of the fishing and aquaculture industries in Loch Creran from 1990 to present day. Aquaculture has a longstanding history within the Loch that predates 1990, and has continued to grow with the introduction of multiple other finfish and shellfish sites. Fishing was previously a well established activity in the Loch however, has since become prohibited from some areas in 2007 with more widespread prohibition in 2015.
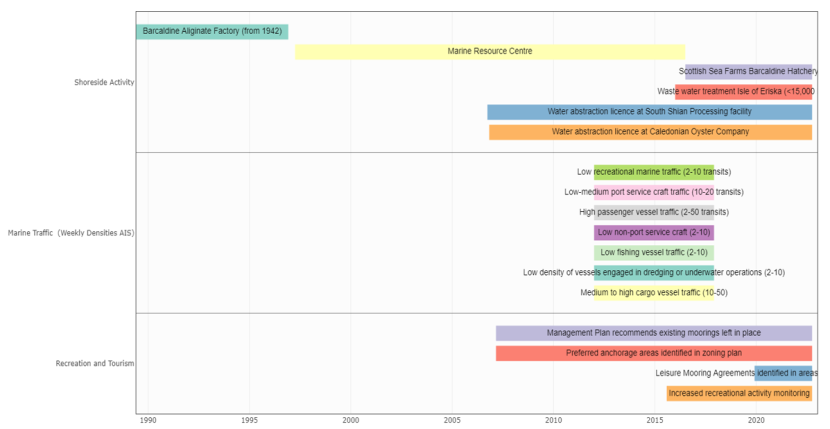
Figure 37. Available data on marine traffic, tourism and shoreside activities in Loch Creran.
Click for a full description
A timeline of shoreside activity, marine traffic and recreation and tourism activities in the area surrounding Loch Creran. These are divided into shoreside activity, marine traffic and recreation and tourism.
Screening assessment
Introduction
The screening assessment was structured around the following questions:
- Can we identify temporal change in any of the environmental or anthropogenic parameters discussed in the Environmental Data Analysis? Review will focus on the period from 2005-2011, when observations suggest that reef condition began to deteriorate.
- If any temporal change(s) can be identified, is there a plausible causal link between the parameter(s) in question and the decline of the serpulid reefs? This stage is driven by the maxim that “correlation is not causation” and will involve a review of the published literature to assess whether the observed changes may be connected.
If robust evidence exists to suggest an impact on S. vermicularis in Loch Creran, conclusions can be drawn on the potential future efforts for restoration and protection. However, the following section aims to make a clear distinction between insufficient evidence of impact and robust evidence of no impact or robust evidence of impact on the reefs.
Table 5 below presents the factors discussed in previous sections (Literature Review and Environmental Data Assessment) and whether these factors have been included or excluded from subsequent analysis, along with rationale for their inclusion or exclusion.
| Factor | Included in SA | Rationale for inclusion/exclusion |
|---|---|---|
| West Scotland climate | No | Some consistent gradual trends over the period of interest, but no sharp change in gradient or significant singular events coincident with reef decline. |
| Phytoplankton biomass / composition | Yes | Fundamental importance of food supply to serpulid reefs and the wider Loch Creran ecosystem. Some evidence of significant change over time, although data are insufficient to plot this in detail. Some phytoplankton species are implicated in HABs. This could also link to M. edulis decline |
| South Shian outfall water | No | No evidence for any consistent trends or significant changes over the period of interest. |
| Farmed salmon biomass/TOC output | No | Salmon tonnage is consistent over entire period of interest (if anything, lower now than in 1990s, when reefs were healthy). |
| Sea lice chemical treatments (Azamethiphos, EMB etc.) | Yes | These chemicals are toxins specifically applied to kill pests, so could potentially also have impacts on non-target species. |
| Heavy metals (Cu, Zn) | No | Antifoulants could potentially have impacts on non-target species. |
| E. coli | No | No evidence of any consistent trends or significant changes over the period of interest. |
| Riverine & general water quality | No | No evidence of any consistent trends or significant changes over the period of interest. |
| Mussels & oysters | (Yes) | Not a factor in themselves but may come into the discussion on phytoplankton. |
| Fishing | No | Trawl damage well-documented by Moore et al., (2003, 2009). Current reef decline is clearly something different. Mobile fishing has been banned since 2007, so it is no longer relevant. |
| Boating & moorings | No | No data on small boat use or illegal anchorage, other than general location of harbours etc. Mooring damage well-documented by Moore et al., (2003, 2009) but very localised. Current reef decline far too widespread for this to be a factor. |
| Vessel activity | No | Anecdotal evidence of vessel activity within the Loch. High resolution tracks of large vessels are recorded annually but cannot currently be related to reef damage unless precise locations of damaged areas are known. |
| Tourism & recreation | No | Insufficient data available to assess. |
| Agriculture | No | No evidence of change in local practices, and no intensive agriculture (e.g. herbicide use) anywhere near the Loch. |
| Forestry | No | Difficulties arise in defining the impacts of deforestation on the Loch (e.g. % change in runoff) from the maps of tree-felling activity. Personal communication suggests herbicides or fertilisers have not been used since 2000, so no chemical input of concern. |
| Didemnum vexillum | No | Not recorded in association with serpulid reefs, or currently considered widespread in Loch Creran. |
| Alginate factory | No | Closed in 1996 before the reef declined. |
West Scotland climatic data
The climatic data available for the Loch Creran area show no obvious extreme events suggestive of a possible association with the serpulid reef decline. During the winter of 2011/12, the west coast of Scotland experienced some storm events but without accurate wind data we cannot determine whether these events were worse than in previous or subsequent years. Both minimum and maximum air temperatures have shown a general upwards trend since the early 1970s however, no data were available to suggest that similar trends have been observed for water temperature in Loch Creran. The broad geographical range of S. vermicularis, which has been recorded in the Mediterranean, suggests that if water temperatures had increased within Loch Creran, this would likely not have caused a significant decline in serpulid populations. Overall, rainfall figures appear to be relatively consistent over the time period during which serpulid reefs have been considered in decline so it is unlikely that increased freshwater input due to more intense rainfall is a factor.
Changes in freshwater input (as related to precipitation) are implicated in studies exploring the changes in phytoplankton in Loch Creran (Whyte, 2012, Whyte et al., 2017). These studies largely consider changes in weather patterns from ‘baseline’ datasets gathered in the 1970s. Comparisons of this nature fail to explain why abundance of S. vermicularis was so high in the late 1990s and early 2000s. Overall the UK is observing warmer, wetter and sunnier conditions than those recorded in the 20th century. From 2012-2021 the UK was 2% wetter than 1991-2020 and 10% wetter than 1961-1990 (Kendon et al., 2021). Western Scotland is most influenced by the Atlantic frontal systems. In terms of average rainfall local to Loch Creran however, the Dunstaffnage weather station records show the highest rainfall peak (between the years 1972-2022) occurred in 2012 (See Figure 4, Environmental data report). Although in general, there do not appear to be multiple above average rainfall events for the decades in which S. vermicularis decline is recorded. Modelled hydrography of Loch Creran shows it is a well-mixed Loch with strong vertical diffusion and that the flushing times are not easily influenced by rainfall (Gillibrand et al., 2013). Implied correlation between increased rainfall and S. vermicularis decline would therefore be highly uncertain. Related changes to phytoplankton biomass are discussed below.
Phytoplankton
A summary of studies conducted in Loch Creran since the early exploratory studies in the 1970s (see Tett and Wallis, 1978; Tett et al., 1985) is below;
- Between 1975 and 2003 local nutrient input increased. However, there were few increases observed throughout the whole Loch and chlorophyll concentrations decreased (Laurent et al., 2006).
- In 2006-2007 datasets which were analysed against the same baseline envelope (1979-1981), proportionally diatoms decreased, and dinoflagellates increased. PCI indicates a significant change in the balance of organisms in the Loch, decreased chlorophyll but again, no nutrient change (Tett et al., 2008), numbers overall are thought to have decreased.
- In 2009, a study conducted to understand the assimilatory capacity of Loch Creran (using 1979 and 2003 data) found that temperature had increased but particulate organic nitrogen and particulate organic carbon had decreased in the Loch, with changes of some phytoplankton species as well. Nutrient reductions were largely attributed to the closure of the alginate factory in the late 1990s (Laurent, 2009).
- No significant changes in silicate or nitrate were noted in samples collected in 2009, but phosphate levels were found to be significantly lower, the decline in PCI and phytoplankton numbers overall were attributed to weather pattern changes and flushing in the Loch (Whyte, 2012).
- Further data gathered in 2011-2013 show community structure has changed considerably and supports the hypothesis by Tett et al., (2008). A rise in mixotrophic/autotrophic dinoflagellates and fewer diatoms, except for Pseudo-nitzschia species which appeared to be increasing, directly related to the intensity and timing of local precipitation patterns (Whyte et al., 2017).
- Monitoring of phytoplankton across broader regions (i.e., the northeast Atlantic) suggests the opposite patterns are occurring in offshore waters compared to inshore waters. But phytoplankton data from Loch Ewe in Scotland, show similar trends to Loch Creran, a decreased diatom occurrence and increased dinoflagellate presence (between a 1958-1967 envelope and a 1998 – 2017 time set) (Bedford et al., 2020).
Overall, nutrient change is hard to show effectively, but chlorophyll a appears to have reduced significantly since early datasets were gathered (1971-1981). The shift in PCI from one dominated by Skeletonema spp., to a more Pseudo-nitzschia spp. dominated system, is also shown in the 2011-2016 envelope presented in the present study. However, since the state of S. vermicularis between 2003 and 2009 was favourable even in the conditions highlighted above, it does seem that the onset of decline occurred after the change in PCI.
As stated in the Environmental Data Analysis, between 1975 and 2003, nutrient input to Loch Creran increased (Laurent et al., 2006). Nitrogen sources from the sea increased from 33.4 mmol DIN year−1 in 1975 to 35.0 mmol DIN year−1 in 2003. Nitrogen sources from River Creran increased from 1.2 mmol DIN year−1 in 1975 to 1.6 mmol DIN year−1 in 2003, while fish farm nitrogen sources increased from 0.0 mmol DIN year−1 in 1975 to 8.8 mmol DIN year−1 in 2003. However, despite the increased nutrient input, observed chlorophyll concentrations decreased substantially. In summer 1975 chlorophyll concentrations were observed at 2 to 3 mg m−3, compared to 0.3 to 2 mg m−3 observed in 2003. Although exact numbers were not provided, changes in the phytoplankton community of Loch Creran were described by Whyte (2012).
In Loch Creran the dominant phytoplankton species in the spring bloom of 1972 were chain-forming diatoms (Skeletonema spp.) (Whyte, (2012). Diatoms in Loch Creran significantly decreased between 1970 and 2009 as did other small flagellates with respect to the abundance of dinoflagellates (Whyte, 2012). Whyte et al., (2017) also found that between 2011 and 2013 community structure had changed substantially, with a substantial drop in diatoms, a rise in autotrophic/mixotrophic dinoflagellates and an increase in the Pseudo-nitzschia genus. In 2011 diatoms vs. autotrophic dinoflagellates were 0.25 cells L-1, in 2012 this was measured at 0.58 cells L-1 and in 2013, 0.00 cells L-1. A move from a diatom- dominated system to one dominated by dinoflagellates is often associated with eutrophication events (Paerl, 1997). We do not currently have any more recent information on phytoplankton abundance and community structure other than the change up to 2013. Similarly, there are limited data available for nutrient input prior to 2017 for Loch Creran. However, chlorophyll concentrations were shown to decrease substantially despite increased nutrient input (Laurent et al., 2006).
While precise preference for feeding is likely unknown and un-recorded for S. vermicularis in Loch Creran, studies on similar species are useful. Most of these focus on non-toxic phytoplankton species within the three key groups; diatoms, dinoflagellates and ciliates (Leung & Cheung, 2017; Pan & Marcoval, 2014). The diet of the serpulid Ditrupa arietina includes the flagellate microphyte Dunaliella tertiolecta (Jordana et al., 2000). The serpulid Hydroides elegans was found to be able to consume and utilise several phytoplankton species including the diatom Chaetoceros gracilis and the dinoflagellates Dunaliella tertiolecta and Isochrysis galbana for development and growth. It was unable to use the diatom Thalassiosira pseudonana as a food source (Leung and Cheung, 2017). The serpulid Hydroides elegans is used as a model organism in feeding studies in Hong Kong (Leung & Cheung, 2017). Both the adult and larval stages were investigated. Adult tube growth was highest when fed on mixed suspension of key phytoplankton species, showing that H. elegans can consume a range of phytoplankton. The effects of toxin- producing phytoplankton were not studied but the authors concluded that H. elegans could be suitable for nutrient removal in eutrophic events because it is capable of filtering algal cells from water efficiently (Leung & Cheung, 2017). Other species of serpulids have been shown to be highly efficient at regulating phytoplankton biomass. For example, Ficopomatus enigmaticus is an invasive species recorded in the Southwest Atlantic and has been shown to clear pennate diatoms twice as fast (on average) as ciliate diatoms (Pan & Marcoval, 2014). Ditrupa arietina have been shown to be capable of ingesting large amounts of phytoplankton cells with thresholds reported at 1 x104 and 4 x 104 cells ml‑1, well above any naturally reported or recorded cell biomass within Loch Creran (Jordana et al., 2000).
Studies have found that S. vermicularis appears to be able to track short term availability of food, and when food is available S. vermicularis is able to detect it. S. vermicularis use their operculum as a lid to cap their opening, hide from predation and stop feeding (Capa et al., 2021). Hiding time is adjusted in response to food availability (Dill & Fraser, 1997; Ten Hove & Kupriyanova, 2009). Serpulids re-emerge sooner when hiding if food is abundant to take advantage of intermittent food availability (Dill & Fraser, 1997). These studies all suggest that serpulids are not limited by an over-abundance of food but might be limited by a lack of food sources. However, lack of specific diet information, feeding thresholds and tolerance to toxin-producing species would again render any interpretation of these data highly uncertain.
Human activities
Aquaculture
The aquaculture industry is likely to be one of the main sources of nutrient and chemical input to Loch Creran, with a number of finfish and shellfish sites currently in operation in the Loch. It should be noted that two active finfish farms exist in Loch Creran however, only one is operational at any given time as the sites are operated in rotation. Two fish farm sites have been in operation in Loch Creran since the early 1980’s and have continued to produce salmon throughout the timeline examined, with no significant changes in biomass having occurred in the Loch since at least 1994. The Loch has also been utilised for shellfish aquaculture, specifically Pacific oyster and blue mussel production.
South Shian outfall water
Data from the processing plant discharge point at South Shian has shown multiple spikes across different measurements, including pH, chloride use, suspended solids and reactive phosphorous. However, none of these anomalies coincide with the decline of the serpulid reefs within Loch Creran and no consistent trends have been observed which might indicate an effect of effluent from this processing plant. These impacts would demonstrate as point source, and therefore decline in the reefs would occur in proximity to the release first. Decline is thought to be widespread throughout Loch Creran and available data do not highlight any spatial differences.
Barcaldine Hatchery
Data provided by SEPA for the years, 2021 and 2022 show the Barcaldine Hatchery is operating well within their licensed levels of discharge, with bath chemicals (formaldehyde and bronopol) only released into the Loch at high tide to increase dispersal potential.. The Barcaldine Hatchery depends upon a freshwater source for the rearing of juvenile fish, so discharges are contained within this freshwater. Freshwater effluent is buoyant, which may assist with dispersal of the discharge in the marine environment. Any discharges must be in line with CAR license conditions to ensure that Environmental Quality Standards (EQS) are maintained Although it forms a significant development in the area, the hatchery was not in operation until after initial declines of serpulid reefs were observed, so the Barcaldine Hatchery is unlikely to be implicated in historic S. vermicularis decline.
Salmon farming
Biomass and TOC output
Up until September 2022, two active finfish farms existed in Loch Creran (Site A and Site B) but only one site is ever in operation at any given time. Despite a significant increase in overall tonnage of finfish aquaculture production in Scotland, maximum biomass within Loch Creran has remained at ~1500 tonnes since at least 1994, before any reported changes in S. vermicularis began.
Changes related to organic matter are discussed with their impacts on phytoplankton above. Monitored releases from aquaculture sites show that organic carbon contributes significantly more than nitrogen or phosphorous, although there is considerable variation in the amount of total organic carbon (TOC) released between years whereas nitrogen and phosphorous remained relatively constant across sample years. This correlates with the reported two-year production cycle for the salmon farm as a reduced biomass at certain points in the cycle would result in reduced TOC released.
Due to the maximum biomass of salmon farming in Loch Creran remaining consistent over the entire period of interest, and no obvious trends on TOC released from sites, it seems unlikely that biomass alone, or nutrient release from salmon farming has caused a decline in serpulid reefs in the area.
Chemical treatments
A review of available environmental data indicates that in terms of total grams applied, azamethiphos contributes the greatest amount of sea lice medical treatments, followed by emamectin benzoate, cypermethrin and deltamethrin. However, it is important to highlight that the key parameter relates to the relevant environmental quality standard, rather than to outright quantity of chemical applied.
Cypermethrin is no longer available for use by the aquaculture industry in Scotland and has not been used in Loch Creran since 2009. Deltamethrin is still licensed for use and may be applied in the future, however its use in the Loch has been limited in recent years, with no applications having taken place since 2017.
Azamethiphos, deltamethrin and cypermethrin are applied externally as a bath treatment so are considered potentially more hazardous to plankton than emamectin benzoate (EMBZ), which is given via salmon feed and reaches lice through their bloodstream. However, EMBZ may reach bottom sediments via fish farm waste in the form of fish faeces and uneaten food (Tett, 2008).
Azamethiphos is highly water soluble, so it is not expected to be found in sediments in detectable concentrations during normal commercial applications. The amphipod crustacean, Corophium volutator, has been shown to experience a 50% mortality rate at high concentrations (Mayor et al., 2008). Mayor et al., (2008) determined the LC50 (lethal concentration) for the burrowing polychaete Hediste diversicolor was 1368 µg EB kg-1 (wet weight) of sediment. The use of azamethiphos in salmon farming in Chile has been shown to reduce the capacity of both chemo- and photoautotrophic production (i.e., bacterial and phytoplankton, respectively) in certain situations, but the impacts were quite varied across the sites tested (Rain-Franco et al., 2018). If azamethiphos potentially acts to depress carbon fixation in phytoplankton, it cannot be ruled out as an indirect factor limiting food sources in Loch Creran, but the same study found that azamethiphos could stimulate primary production if the conditions were favourable (Rain-Franco et al., 2018). These findings contrast with earlier studies that found no impact on primary production by azamethiphos (Burridge et al., 2010). Given the intermittent nature of treatments and the area of Loch that may potentially be affected, it should be highlighted that any potential effect would be limited spatially and temporarily. In contrast to this, sublethal effects of deltamethrin have been observed in polychaetes (Alitta virens, formerly known as Nereis virens), at concentrations of 11µg/g (Van Geest et al., 2014). The effects observed related to burrowing behaviour and worm condition. The chemical does, however, have rapid degradation in water (60% in 5 mins), high adsorption on sediment (particles stick to the surface of the sediment) and low bioconcentration capacities which indicate that it will not accumulate through food chains (Haya et al., 2005). Tett et al. (2018) noted that application of this chemical treatment in Scotland was decreasing and no applications have occurred in Loch Creran since 2017.
As highlighted above, cypermethrin is no longer in active use as a sea lice treatment in Scotland and has not been applied in Loch Creran since 2009. However, it is still used widely in the terrestrial environment, including in agriculture (as a treatment for crops and livestock, including cattle and sheep), forestry (used to treat young trees, both pre- and post- planting to control large pine weevil) and in domestic application as a household insecticide. The wide range of uses of cypermethrin means there are a number of routes by which it can enter the water environment (Environment Agency, 2019). Monitoring carried out by the Environmental Agency in England between 2016 and 2018 found the average concentration was above the EQS at 20 of the 40 saline sites monitored (Environment Agency, 2019). There does not appear to be any available data relating to any similar monitoring programmes in Scotland.
Systematic studies of the responses of marine polychaetes to cypermethrin appear to be lacking. However, laboratory experiments considered the growth of three marine phytoplankton species Skeletonema costatum, Scrippsiella trochoidea and Chattonella marina and their response to exposure to cypermethrin for 96 h and 32 days in a co-culture system. Based on the results of this study, the authors suggested that in some circumstances, cypermethrin might have potential to lead to a shift in phytoplankton community structure, from diatom to harmful dinoflagellate species, and thus potentially be stimulatory for harmful algal blooms (Wang et al. 2011). Further laboratory experiments have found that cypermethrin reduced zooplankton density and biodiversity not only directly, by killing copepods, but also indirectly, by increasing the numbers of rotifers, which feed on phytoplankton (Medina et al. 2004). The same study found that zooplankton density recovered after treatment, but zooplankton biodiversity remained altered. The authors theorised that in an open environment, the rapid dissipation of the chemical and population processes of compensation, migration, and immigration, may lead to recovery of the affected zooplankton communities. The findings of these studies are of note, given the observed reduction in biomass and shift in phytoplankton community in Loch Creran.
There is limited evidence relating to the sensitivity of serpulid species to hydrogen peroxide. However, a laboratory based study considering the tolerance of two marine polychaete species suggests that hydrogen peroxide has the potential to result in negative effects. The study considered the tolerance of Capitella sp. and Ophryotrocha spp. to a 1 h exposure to H2O2 at varying concentrations, ranging from 0-1800 mg L‑1 (Fang et al. 2018). High cumulative mortality during a 72 h post-exposure period were observed for both polychaete species. The study found a positive correlation between observed mortality rates in both species with increasing concentrations of hydrogen peroxide.
While evidence exists that emamectin benzoate may persist in sediments for up to a year after treatments, it was previously found to not have toxic effects on either water column or benthic organisms in a study by Telfer et al., (2006). Furthermore, no adverse effect on zooplankton communities and copepod abundance in the vicinity of sea-farms was observed in studies by Willis et al. (2005) and Willis & Ling (2003). Evidence has however emerged more recently to suggest that EMBZ has a significantly greater effect on total crustacean abundance and species richness than previously thought (SEPA, 2018). Ingestion of EMBZ has also been shown to induce premature moulting in American lobsters (Waddy et al., 2002) and with accumulation in sediment, may pose a risk to other sensitive benthic organisms such as polychaete worms. Evidence of detrimental impacts of emamectin benzoate on organisms such as lobsters (Daoud et al., 2018), treated salmon smolts (Lennox et al., 2020), polychaetes (McBriarty et al., 2018) and Daphnia magna is present in the literature.
Although there have been no studies into the effects of EMBZ on S. vermicularis, it has been found to have an effect on the polychaete Alitta virens (McBriarty et al., 2018). When exposed to concentrations of 400 µg/kg dry weight (nominal) for 30 days in sand, McBriarty et al., found that no treatment-related mortality occurred. However, a significant decrease in worm mass and marked behavioural changes were observed compared with control populations. This is considerably higher than concentrations observed in sediments around fish farm sites. The effects are similar to those observed when A. virens was exposed to deltamethrin (Van Geest et al., 2014).
The studies outlined above are useful in understanding acute toxicity. However, studies to consider potential chronic effects (i.e. effects on growth rate, reproduction and mortality) appear to be lacking. Experiments to investigate chronic toxicity are possible but challenging, as they would likely require maintenance of serpulids in aquaria for periods of at least several years. Such long-term experiments would be logistically demanding, unlikely to accurately reproduce conditions experienced by populations in the field, and as such, any results would therefore be of limited value relative to the investment of time and resources.
Given the timeline of observed reef decline in Loch Creran, there is temporal overlap with chemical treatments applied historically in the Loch. Application of bath treatments in the Loch appears to have peaked from 2009 – 2016, which broadly coincides with the observed reef declines in the Loch. However, applications have decreased significantly since 2016, with no bath treatments taking place between August 2016 and October 2021 and observed deterioration of the habitat appears to have continued during this period.
Impacts of chemical treatments on larval stage S. vermicularis
While present in the water column, there may be potential for S. vermicularis larvae to be affected by the release of chemicals into the water column as a result of the use of bath treatments associated with finfish aquaculture. The duration of the planktonic stage of S. vermicularis is unknown but comparison with other serpulid species suggests it may be between six days and two months (Holt et al., 1998).
Available evidence suggests spawning occurs in the summer. In the Clyde area, Elmhirst (1922) observed spawning to occur in June to August and in Plymouth ripe individuals were seen in August and September (Allen, 1915). Studies in Loch Creran (Cook et al. 2021) suggest that July may be the peak month for spawning. Chapman et al., (2007) recorded larval settlement in Loch Creran from mid-June to mid-October, peaking in late August/early September. Based on these studies it seems reasonable to conclude that planktonic S. vermicularis are unlikely to be present in the water column beyond the month of October. Therefore, any activities that could affect the viability of larvae while present in the water column would have potential to have an effect during the period June – October. However, it seems probable that the greatest effect would occur where the activity overlaps with the period of peak spawning, which is suggested to occur in Loch Creran between July and early September (Cook et al., 2021; Chapman et al., 2007).
On this basis, data on the release of bath treatments were analysed to assess potential temporal overlap between bath treatments released from the fish farms and planktonic stage S. vermicularus potentially present in the water column during a five month period between June and October each year. The results of this basic analysis indicate that temporal overlap occurred in 2005 (cypermethrin discharged in October), 2007 (cypermethrin discharged in June, September and October); 2009 (deltamethrin discharged in June, July, August, September and October); 2011 (deltamethrin discharged in June, July, August, September and October); 2013 (azamethiphos discharged in July); 2014 (deltamethrin discharged in September), 2016 (azamethiphos discharged in August and September) and 2021 (azamethiphos discharged in October).
Figure 38, 39 and 40 provide a visual representation of the overlap between bath treatments and the five month period between June and October.
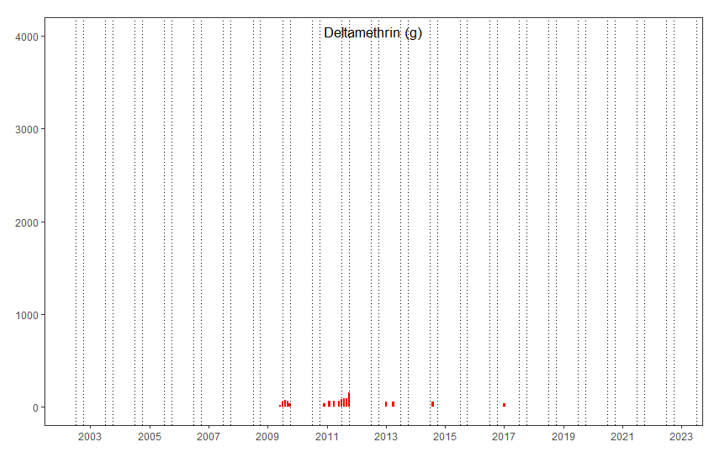
Figure 38. Deltamethrin use between May – October (2002–2023).
Click for a full description
The data presented indicates that deltamethrin was applied during this period in 2009 (discharged in June, July, August, September and October); 2011 (discharged in June, July, August, September and October); and 2014 (discharged in September). This is based on data from Creran A and B fish farms between 2002 and 2023.
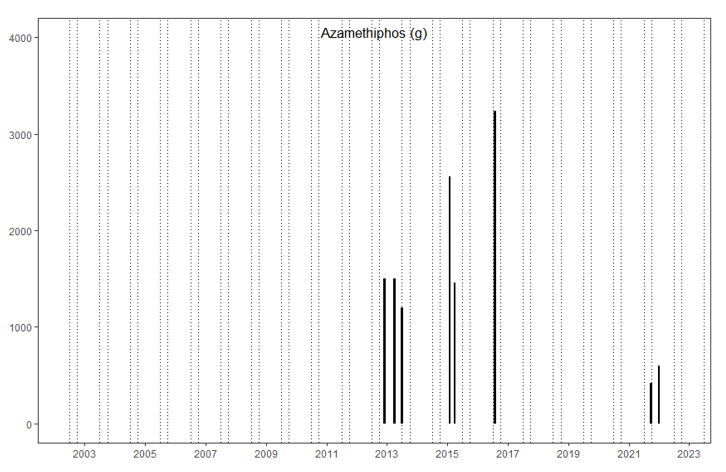
A visual summary to represent the use of azamethiphos where it has taken place during the period June to October, when S. vermicularus larvae may be present in the water column.
Click for a full description
The data presented indicates that azamethiphos was applied during this period in 2013 (discharged in July), 2016 (discharged in August and September) and 2021 (discharged in October). This is based on data from Creran A and B fish farms between 2002 and 2023.
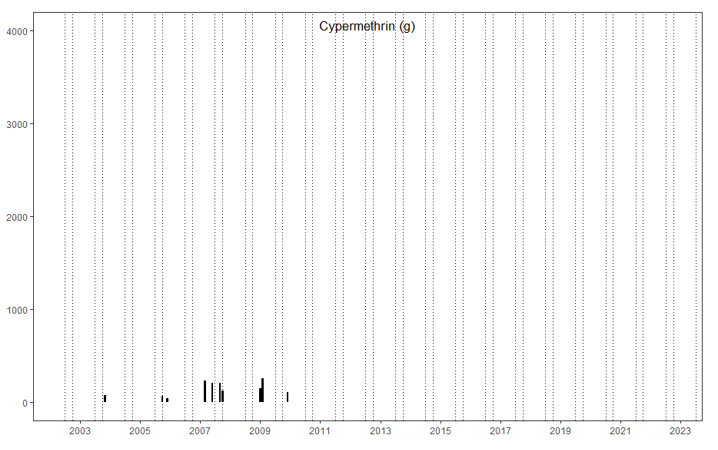
Figure 40. Cypermethrin use between May–October (2002–2023).
Click for a full description
The data presented indicates that cypermethrin was applied during this period in 2005 (discharged in October) and 2007 (discharged in June, September and October). This is based on data from Creran A and B fish farms between 2002 and 2023.
Loch Creran is considered to be a well-mixed water body with no isolation of high and low salinity areas. The flushing time for the Loch is 3 days, during which time around 60% of the Loch’s volume is exchanged with the neighbouring coastal waters, it has been suggested that this reduces the risk of problematic levels of soluble contaminants (Edwards and Sharples, 1986). Based on the known flushing rate, it seems unlikely that any significant concentration of chemicals released following bath treatments would remain in the water column for prolonged periods. Any chemicals licensed for use will be assessed by SEPA to ensure that the relevant environmental quality standard will be met. The relevant EQS for each chemical will differ. The chemicals licenced for use in Loch Creran were licensed before SEPA introduced their revised regulatory framework for aquaculture in 2019, so were assessed under the previous framework. Detailed modelling as required under the revised framework may provide a greater understanding of the dispersion and fate of any chemicals released into the water column.
Previous studies in the Loch (Moore et al., 2020; Cook, 2016; Hughes, 2011) provide evidence to demonstrate that resettlement was occurring during periods where temporal overlap with bath treatments released from the farms exists. While this is not evidence of no effect, it does indicate that larvae present in the water column during these periods were able to survive and settle despite these treatments taking place. No evidence appears to exist to demonstrate that the chemicals associated with historic bath treatments in the Loch result in any direct effect on S. vermicularus during the planktonic stage of its lifecycle. However, without evidence to conclude otherwise, and where temporal overlap exists between the release of chemicals associated with bath treatments and the key spawning / settlement period for S. vermicularus, it is not possible to rule out potential effects entirely, although widespread effects across the whole Loch, as have been observed, appear to be unlikely.
Due to the knowledge gaps that exist, it is not possible to rule out potential effects. To consider effects fully, an improved understanding of the sensitivity of S. vermicularus larvae to the chemicals applied as bath treatments in the Loch would be required. In addition, detailed modelling, which could be carried out relatively easily, would provide further information regarding the predicted dispersal and fate of bath treatments in the Loch. This information may provide further evidence to help understand the likelihood that any negative effects on planktonic stage S. vermicularus larvae could have occurred as a result the release of bath treatments in the Loch.
Heavy metals
Reported annual inputs of heavy metals from Sites A and B indicate that copper input is at far greater levels compared with zinc input however, copper input fluctuated considerably whereas zinc remained relatively constant. There is some evidence to suggest that the release of aquaculture associated heavy metal and organic pollutants can induce stress in bioaccumulating organisms in Loch Creran, such as the blue mussel Mytilus edulis (Nippard & Ciocan, 2019).
Copper is an essential component of fish feed as it is required for normal growth and metabolism. This can result in increased concentrations of copper around fish farms due to either over feeding (excess food reaching the seabed) or by excretion in fish faeces (Tett, 2008). The copper reaching the seabed dissolves to increase the concentration of copper ions in the sediment pore waters (the water around the sediment on the seabed) and may diffuse back into the water column. Copper has been shown to affect fertilisation success in marine invertebrates although there are relatively few studies on polychaetes. Using Galeolaria caespitosa, copper was shown to have much stronger effects on fertilisation at low sperm concentrations than at high concentrations (Kupriyanova & Havenhand, 2005). This suggests that effects in natural field populations would be much greater than those in laboratory conditions, as sperm dispersal is likely to be greater in the field. Exposure of gametogenic A. virens to higher concentrations of copper induced more males to spawn and they emerge earlier in the year. Another study found that copper pulses from impregnated blocks had a profound negative effect on settlement of certain marine invertebrates (for example large space-occupying tunicates) in the field but found that serpulid polychaetes responded positively. Serpulid settlement densities were an order of magnitude higher on copper exposed plates (Johnston & Keough, 2002). In contrast, Watson et al., 2008 found copper affected settlement behaviour in the nechtochaete larval stage of A. virens. Larvae avoided settling on sediment with concentrations of copper greater than 50 mg/kg per dry weight of sediment.
The input of heavy metals into Loch Creran from finfish aquaculture production, either via feed or antifouling paint, indicates this factor requires due consideration, as numerous detrimental effects have been highlighted in this section which suggest serpulid reefs, and other species, could be negatively impacted. It should however, be noted that aquaculture is not the only possible source of heavy metals in the marine environment and the relative contribution (and distribution) of copper from feed to the surrounding environment in Loch Creran is not known.
Shellfish production
During the period in which serpulid reef decline has been observed, blue mussel aquaculture production has ceased. However, the production of Pacific oyster (Crassostrea gigas) in the Loch Creran area has increased significantly between 2009-2021 with combined table and on-growing production increasing 167% (personal communications, Marine Scotland Science, November 2022).
It seems unlikely that the observed changes in shellfish production would have any significant effect on serpulid reefs in the Loch. However, the observed period of decline for blue mussel production in the Loch does coincide with declining conditions of serpulid reefs. However, the cause of the cessation of blue mussel production has not been explored in this study.
E. coli
The Food Standards Agency (FSA) / FSS (formed in 2015 and taking on the former remit of FSA) have recorded occurrences of Escherichia coli since 1999. Sampling has focused on areas along the southern shore of Loch Creran as part of statutory monitoring required because of the shellfish harvesting and production taking place in the area. The data discussed in the previous section highlights three occurrences of high E. coli loadings in water discharged into the Loch. However, there was no overall significant increasing or decreasing trend, nor is there currently any evidence to suggest that E. coli would likely impact serpulid reefs.
Riverine and general water quality
Data provided from the three monitoring stations within Loch Creran suggest there has been no significant changes in the overall quality of water being discharged into the Loch. Patterns of alkalinity, pH, dissolved oxygen and temperature seem to fluctuate over the course of the monitoring period with no clear trends. Similarly, metals have also been monitored and appear to be relatively consistent with the exception of a spike in zinc levels in 2004 at Creagan Bridge. However, this anomaly does not appear to coincide with the decline of serpulid reefs.
Fishing
Dredging, trawling, creeling and whelk fishing have all occurred in Loch Creran. On 29 March 2007, the Inshore Fishing (Prohibited Methods of Fishing) (Loch Creran) Order 2007 came into force to prohibit scallop dredging and benthic trawling in parts of the Loch. In 2015 a new Order came into force banning all mobile gear in Loch Creran. Creeling is only allowed in the centre of the Loch away from all the protected features including serpulid reefs. This is further supported by the Senior Fishery Officer for the Oban District, who confirmed that creel fishing is the only commercial fishing which still occurs in Loch Creran.
Previous reef damage by trawling activity in the Loch has been well documented by Moore and colleagues (2003 and 2009) however, the damage at the time was considered to be more localised to areas where commercial fishing took place. The banning of mobile fishing gear since 2007, and the current widespread decline of the serpulid reefs indicates that commercial fishing cannot be a causal factor.
Boating and moorings
Loch Creran experiences mainly marine traffic of small to medium sized vessels, with larger vessels only occasionally passing through the area. Activity is mostly concentrated around the depot at Rubha Garbh, which serves the quarry at Glensanda, the pier complex at the Barcaldine Hatchery and Creran Marine, which serves both industrial and recreational vessels and the pump ashore facility at South Shian. While no industrial activity takes place at the Rubha Garbh depot, the pier complex at Barcaldine has a marina/boatyard, a fish hatchery, and several other small scale industrial activities. The marina at Barcaldine offers between 70 and 100 moorings, with yacht and pleasure craft activity increasing during the summer months. Larger workboats are also frequently observed at South Shian, the main activity here is well boats coming alongside at the pontoon pump ashore facility (Scottish Sea Farms).
Mooring and anchor placement for both recreational and commercial use has the potential to cause significant damage to serpulid reefs. Anchoring is more likely to cause damage than mooring placements, as they may be dropped indiscriminately, and will generally drag before settling. If the wind direction and currents change this can cause the anchor chain to drag and could damage larger areas of the reef. There are currently numerous leisure mooring agreements around Loch Creran, leased by Crown Estate Scotland, with recreational vessels frequently observed during summer months. In the 2007 management plan, Argyll and Bute Council recognised that mariners have a right to anchor in safe areas. In this plan, a strategy was decided that only emergency anchoring would be allowed in serpulid reef areas. There were also preferred anchorage sites identified which mariners will be informed of and be encouraged to use (Argyll and Bute Council, 2007). Since the management plans were in place, there are now limited anchorages for recreational yachts remaining in Loch Creran, any that were in proximity to serpulid reefs have been removed. There is currently a sign at the entrance to Loch Creran directing yachts to appropriate anchorage zones.
Mooring damage is well documented by Moore et al., (2003, 2009) but any damage is localised. The current pattern of reef decline is too widespread for this to be a significant factor contributing to the observed decline.
Tourism and recreation
Marine recreation and tourism are significant sectors within the Argyll and Bute region, with Loch Creran offering multiple activities including sailing, sea kayaking, wildlife watching and scuba diving. Loch Creran is a common diving location as it offers shallow sheltered sites when the weather is inclement. Diving is common at a number of sites that have easy access from the road. As serpulid reefs are fragile this makes them vulnerable to physical damage which could be caused by novice divers who have poor spatial awareness or buoyancy control. A dive code was created prior to the publication of the Loch Creran Management Plan in 2007, for recreational diving in the Firth of Lorn and Loch Creran marine SAC. It has been distributed to all SSAC, BSAC, Sub-Aqua Association (SAA), Professional Association of Diving Instructors (PADI) and National Association of Underwater Instructors (NAUI) clubs. Signage was also placed at strategic points around the Loch (Argyll and Bute Council, 2007).
The lack of data regarding recreational and tourist activities in the Loch precludes any further consideration of this factor. However, it is highly unlikely that this activity has contributed significantly to the widespread declines that have been observed across much of the Loch.
Agriculture
On the northern and far western shores of Loch Creran there are small numbers of cattle livestock. Many agricultural activities are regulated under The Control of Pollution (Silage, Slurry and Agricultural Fuel Oil) (Scotland) Regulations 2001. Farmers are also encouraged to have good practice, which is stated in documents such as Four Point Plan, the Farm Soils Plan, and the Prevention of Environmental Pollution from Agricultural Activity (PEPFAA) Code. Agriculture does have the potential to reduce a loch’s water quality due to run-off entering the marine environment. In the 2007 management plan it was stated that the level in 2007 was having no negative impacts on the conservation interests within Loch Creran (Argyll and Bute Council, 2007). It should be noted that any agricultural data employed for the purposes of this report predate the period during which serpulid reefs were considered to decline so may no longer be relevant.
Based on the lack of recent information on agricultural practices in the area, it cannot be determined with certainty whether or not local agriculture has had an impact on serpulid reef decline. Monitoring data from 2007 indicates that no impact occurred previous to this however, more recent data would be required to determine current impacts.
Forestry
Commercial forestry in the surrounding area of Loch Creran is managed by FLS. There is a significant amount of forestry in the catchment area around Loch Creran. Glen Creran is carefully managed with a clear-fell and re-stock approach. Deforestation of regions close to shore can increase freshwater run-off which in turn will increase nutrients, turbidity and sediments as well as potentially impacting the pH (Howe et al., 2010). Land preparation and development for the forestry could have an impact on water quality through siltation, eutrophication or the translocation of herbicides and insecticides. The possibility of impacting water quality can increase in freshwater catchments or marine areas experiencing restricted water exchange and therefore large-scale plantation or felling proposals could have an impact on the conservation interests of Loch Creran. The forestry industry currently adheres to the Forests and Water Guidelines, which were created following extensive consultation throughout the forest and water industries, government departments, including Forestry Commission Scotland and SEPA, research institutes and other relevant organisations. The Guidelines provide guidance to forest managers and practitioners on how forests should be designed, planned and operations carried out to protect and enhance the water environment. Any new forestry developments around the boundary of the SAC should be subject to the Habitats Regulations (Argyll and Bute council, 2007).
Although it is clear there has been extensive tree-felling in the area surrounding Loch Creran since 2011, particularly in areas to the south of the Loch, the absence of data to suggest there has been an increase in freshwater runoff as a result provides little clarity on whether serpulid reef decline could be linked to this activity.
Didemnum vexillum
Monitoring of the invasive, non-native ascidian Didemnum vexillum began in 2017 after first being observed in Loch Creran in 2016. Data collected indicates that the species has not been observed on any occasion between 2017 and 2021 on or near serpulid reefs. Didemnum vexillum is therefore, unlikely to have caused the current widespread decline observed throughout the Loch.
Alginate factory
The alginate factory in Barcaldine commenced operations in 1939 and reached peak production in the 1970s. The factory discharged alkaline liquid whey, an acidic leachate, formaldehyde, and organic particulates (Black et al., 2000). The debate surrounding the closure of the alginate factory stems around the diet and nutrient requirements of S. vermicularis. Serpulid aggregations, in some places, develop in brackish, nutrient rich waters. The removal of this source of nutrients has previously been suggested as a factor that had an impact on phytoplankton communities, potentially explaining some of reduction in nutrients observed between the 1970’s and 2003-2006 (Laurent, 2009). Furthermore, algal mats have been shown to provide habitats for certain species of serpulid in other parts of the world (Rossbach et al., 2021). On the other hand, anoxia caused by excessive nutrient input, also reported as a local impact of the alginate factory (see Jones, 1979), is likely to have a detrimental impact on recruitment of juveniles and reduced respiration in serpulid reefs immediately offshore from the factory, as has been shown in other species (namely Hydroides elegans) (Leung et al., 2013). The closure of the factory in 1996 (a decade before the height of S. vermicularis abundance was recorded) has previously been linked to the de novo success with new formations recorded (Moore, 1996; Moore et al., 1998). The closure of the alginate factory has previously been linked to a reduction in nutrients in the water column (Laurent et al., 2006) and the regrowth of immediately adjacent S. vermicularis aggregations along the south shore of Loch Creran. It is unlikely that the reefs of S. vermicularis in the rest of the Loch would be significantly impacted by this closure, given the efficient mixing time of the Loch and their distance from the source (Gillibrand et al., 2013). It is therefore unlikely that this closure had an impact on the broad deterioration of S. vermicularis reefs in Loch Creran.
Assessment of activities, pressures and knowledge gaps
Introduction - Explaining ecological change in the sea
Marine ecosystems provide many documented cases of species undergoing population decline or disappearance. Examples range from the mass extinctions preserved in the fossil record (Harnik et al. 2012) to the extinctions of individual species in historical time (Carlton et al. 1999). These events have attracted, and continue to attract, much scientific interest, but with the exception of marine vertebrate megafauna depleted or exterminated by direct human predation (e.g. Turvey & Risley 2006, Ward-Paige et al. 2010) it has proved difficult to identify the causal factors behind some well-documented instances of population collapse or change. Recent examples illustrate the challenges involved:
- Diadema antillarum mass mortality, western Atlantic, 1983-1984
- Sea star wasting, North American Pacific coast, 2013 – present
- Crustacean mass mortality, north-eastern England, 2021-2022
- Recent evidence of widespread decline in blue mussel populations observed across and beyond the North Atlantic
Diadema antillarum mass mortality, western Atlantic, 1983-1984
In 1983-1984 the long-spined black sea urchin Diadema antillarum suffered mass mortality throughout the Caribbean Sea and western Atlantic as far as Bermuda (Lessios, 1988). Local mortality rates of 93-100% were observed within two weeks. The loss of this keystone herbivore led to profound changes in the ecology of Caribbean reef ecosystems, most visibly a phase shift from coral-dominated to algal-dominated communities (Hughes 1994). Population recovery was extremely slow, and in some areas the residual populations were hit by a second die-off in 2022 (Hylkema et al., 2023). The progression of mortality across the region in 1983-1984 was suggestive of a water-borne pathogen as the causal agent (Lessios 1988), and its initiation on the Caribbean coast of Panama (Lessios et al., 1984) raised suspicions that the agent responsible might have been introduced in ballast water released by a ship transiting the Panama Canal. Both suggestions are plausible but unconfirmed, (Lessios 2016). The causative agent of the 2022 mass mortality has recently been identified as a scuticociliate protist (possibly Philaster sp.) (Hewson et al. 2023), but it is not known whether the same pathogen was also responsible for the 1983-1984 event.
Sea star wasting, North American Pacific coast, 2013 – present
Since 2013, more than 20 species of sea star (Echinodermata: Asteroidea) found along the Pacific coast of North America have been afflicted by a condition usually termed sea star wasting disease (SSWD) or sea star wasting syndrome (SSWS) (Oulhen et al., 2022). Wasting typically begins with breakdown of body wall connective tissue, followed by disintegration of the epidermis and death. Earlier mass mortality events affecting sea stars have been noted in North America (e.g. Dungan et al., 1982), but the episode unfolding since 2013 has been particularly severe, both in terms of geographic extent and the number of species affected. A similar syndrome affecting the sea star Crossaster papposus has recently been recorded from the Irish Sea (Smith et al., 2022). Viruses have been proposed as transmissible agents responsible for SSWD (Hewson et al., 2014), with their effects possibly synergistic with environmental factors such as higher sea temperature. No infectious agent has yet been identified and the cause(s) of SSWD remain uncertain (Oulhen et al., 2022).
Crustacean mass mortality, northeastern England, 2021-2022
Recent stranding of thousands of dead or dying crabs and lobsters was observed along at least 70 km of the north-eastern English coast, first noted in early October 2021 and continuing intermittently through 2022 (BBC 2021, 2023). The cause of this unusual crustacean mortality event is unclear, with potential suggested causes including disease, harmful algal blooms and toxic chemicals released from bottom sediments by local dredging activity. A Crustacean Mortality Expert Panel (CMEP) was convened in December 2022 to assess the evidence and, if possible, identify the likely cause. Their report, published in January 2023 (CMEP, 2023) could not identify a clear and convincing single cause for the event. Harmful algal blooms, chemical toxicity and dredging were judged “unlikely” or “very unlikely” to be the causal factors. A novel pathogen was considered the most likely cause of mortality, although no direct evidence for one was found.
Recent evidence of widespread decline in blue mussel populations observed across and beyond the North Atlantic
Within coastal waters of the North Atlantic, and further afield, there has been sustained evidence of decline in Mytilus spp. distributions (Baden et al., 2021; Christie et al., 2020). The reasons for the declines are uncertain. No single factor has been identified or demonstrated to account for the observed losses of mussel populations. In review, Baden et al. (2021), found some factors were more likely to play a part in the mussel decline than others and several factors have been investigated, many of which may be acting together as multiple stressors. Such potential causes of decline include but are not limited to: changes in predation by eider ducks (Somateria molissima) and the green crab (Carcina maenas) (which have increased populations by 10-fold and 3-fold respectively), climate change, overfishing, competition from invasive species, pollution, pathogens/ disease and reproduction. Similarly, Christie et al., (2020) highlight that meso-predation by increased abundance of C. maenas and the goldsinny wrasse (Ctenolabrus rupestris) exert grazing pressure on small blue mussels. The overlying cause is however, as highlighted by Baden et al., (2021), likely to be a result of multiple stressors acting in unison rather than a single underlying factor.
Serpulid reef decline in Loch Creran: barriers to explanation
The case studies cited above illustrate the difficulty of establishing the causal factor(s) underlying significant population changes or mass mortality events in the marine environment. Some of these examples, represent events which have well-documented starting points and an abundance of specimens available for laboratory examination, chemical and microbiological analysis. Considerable scientific effort has been put into all three, comprising almost 40 years of research in the case of Diadema antillarum, yet in each case, the causes of the events remain unresolved.
Several aspects of the serpulid reef decline in Loch Creran pose problems to any attempt to identify the causal factor(s):
- We are not dealing with an abrupt mass mortality event (indicative of a discrete triggering event or factor) but a protracted decline extending over more than a decade, and therefore potentially overlapping with a number of natural or anthropogenic environmental changes.
- A protracted decline might be driven by changes in any or all of the key demographic parameters of the serpulid population, i.e. recruitment, growth and mortality. Data on all three are very limited (e.g. Chapman et al. 2007, Hughes et al., 2008, Hughes 2011) and none were monitored regularly across the period of interest.
- The starting point of the decline cannot be firmly identified. The Literature Review in this report suggests that reefs were in a healthy condition in 2005 with visible deterioration apparent from 2011 onwards, but their status in the intervening period is not well-known.
- The reef decline occurred in the subtidal zone, with no visible manifestation above water. This contrasts with the mass mortalities discussed above, in all of which large numbers of stranded specimens were available for scientific study.
- In situ research on the Loch Creran serpulid reefs was infrequent and intermittent across the period of interest, with long gaps between surveys or field experiments.
- The data on some potentially important natural variables and human activities are patchy or absent, and we do not have a comprehensive picture of environmental change in Loch Creran over the period of interest.
- Information on serpulid reefs in other Scottish sea lochs is extremely limited. The few available datasets suggest that reefs can appear and disappear, but the lack of a detailed study in any locality means that the determining factors are unknown and these observations provide no insight into the possible causes of change in Loch Creran.
- The only well-documented case study of S. vermicularis mortality triggered by environmental change (Ardbear Salt Lake, Ireland, Leahy 1991) is of limited relevance owing to the differences between the Irish locality and Loch Creran. A hypoxic episode of the severity described in Ardbear Salt Lake has never been recorded in Loch Creran, and it is unlikely that any such event could occur there. There are indications of earlier cycles of reef growth and decline in Ardbear Salt Lake (Bosence 1979, Leahy 1991) but the timing and details of these are not known.
Testing hypotheses on the causes of reef decline
Taking account of the limited evidence base described above, it may be useful to consider how the issue of reef decline in Loch Creran could be examined from first principles using a hypothesis-testing approach. This approach, summarised in Table 6, could be applied to any natural environmental variable or human activity (here termed “Factor X”) proposed as a cause of reef decline. The hypothesis that Factor X is responsible for reef decline is addressed through a series of simple questions relating to the susceptibility of S. vermicularis to X, and the expected spatial and temporal pattern of decline, were X to be the causal factor. This may help to illustrate the type of information needed to evaluate the specific factors discussed in the Data Analysis and Screening assessment sections of this report.
Hypothesis: Factor X is the cause of serpulid reef decline in Loch Creran
| Evidence supporting the hypothesis | Evidence against the hypothesis |
|---|---|
| Susceptibility to X | Susceptibility to X |
Observational or experimental evidence that X has an adverse effect, e.g.
(See Note 1 below) | Observational or experimental evidence that X has no adverse effects. (See Note 2 below) |
| Timing of reef decline | Timing of reef decline |
|
|
|
|
| Spatial pattern of reef decline | Spatial pattern of reef decline |
|
|
|
|
Notes:
- Regarding susceptibility to Factor X, published evidence can be ranked in quality and relevance, from highest to lowest, on the following scale:
- Adverse effects on S. vermicularis >
- Adverse effects on serpulids other than S. vermicularis >
- Adverse effects on polychaetes other than Serpulidae >
- Adverse effects on animals other than polychaetes
- An assessment of non-susceptibility to Factor X is not determined by the absence of evidence (i.e. no relevant data at all), but by observational or experimental evidence that X does not have adverse effects. Quality of evidence is ranked on the scale outlined in Note 1.
The framework outlined could, in theory, be used to evaluate any single factor proposed as a cause of reef decline. A hypothesis entailing the interaction of two factors (i.e. “Factors X and Y in combination are the cause of reef decline in Loch Creran”) would be much more difficult to express as a set of testable predictions, and multi-factorial hypotheses (e.g. “Factors X, Y and Z…”) are likely to be untestable and therefore purely speculative.
Application of the hypothesis-testing approach
The approach outlined is applied to three of the factors discussed in the Data Analysis and Screening Assessment sections of this report, to examine whether the available data is sufficient to test hypotheses on the causes of reef decline. Although additional factors could have been tested, these were screened out in the Screening Assesment section. For a summary of the rationale for exclusion of factors see Table 5. The first hypothesis tested against the approach relates to phytoplankton community change, which was selected due to the fundamental importance of food supply to serpulid reefs and would be expected to affect the entire Loch simultaneously. The other two factors tested, (emamectin benzoate and azamethiphos) are point-source anthropogenic factors associated with fish farms. These were selected due to their potential to have an impact on non-target species.
Hypothesis 1: Change in the phytoplankton community (biomass and/or composition) is the cause of serpulid reef decline in Loch Creran
| Evidence supporting the hypothesis | Evidence against the hypothesis |
|---|---|
| Susceptibility to change in phytoplankton community | Susceptibility to change in phytoplankton community |
Unknown: insufficient data on diet of S. vermicularis in Loch Creran
Phytoplankton selectivity not known, i.e. would a shift from diatoms to dinoflagellates have any adverse effect? | - |
| Timing of reef decline | Timing of reef decline |
| - | Phytoplankton community change seems to have been gradual when measured against 1970s baseline. Change was underway before modern reefs confirmed to exist. Numbers of diatoms, dinoflagellates and ciliates ± steady over period 2004-2022. No abrupt changes at or just before 2011. Lack of phytoplankton community evidence in the 1990s, lack of evidence of S. vermicularis abundance in the 1970s for comparison. |
| Spatial pattern of reef decline | Spatial pattern of reef decline |
| Reef decline throughout the Loch is consistent with a non- point-source factor, but early stages are poorly-documented. Not known whether start of deterioration was localised or general. | - |
The overall conclusion from this example is that the available data provide no strong support for the hypothesis that reef decline was triggered by changes in Loch Creran’s phytoplankton community. Important information on the diet of Serpula vermicularis in the Loch is lacking. There are no indications of abrupt change in phytoplankton biomass or composition around the time when reef deterioration was first detected. The decline of reefs throughout Loch Creran is consistent with a non-localised factor such as phytoplankton, but crucially we do not know whether the process of decline was always widespread or initially confined to specific areas of the Loch.
Hypothesis 2: Emamectin benzoate (EMBZ) from salmon cages is the cause of serpulid reef decline in Loch Creran
| Evidence supporting the hypothesis | Evidence against the hypothesis |
|---|---|
| Susceptibility to Emamectin Benzoate | Susceptibility to Emamectin Benzoate |
| Exposure to EMBZ associated with lower growth rate and behavioural change in the polychaete Alitta (formerly Nereis) virens. |
|
| Timing of reef decline | Timing of reef decline |
| Use of EMBZ in Loch Creran precedes the observed start of reef decline, and continues to the present. | - |
| Spatial pattern of reef decline | Spatial pattern of reef decline |
| - | Reef decline general throughout the Loch. No indication of greater severity closer to salmon farm. |
The data summarised in Table 8 provide no strong support for emamectin benzoate (EMBZ) as the cause of reef decline. Chronic effects of EMBZ exposure have been shown for one species of burrowing polychaete (Alitta virens, McBriarty et al., 2018), but none were found for Arenicola marina exposed to high concentrations of EMBZ (Cheng et al. 2020). Emamectin benzoate binds strongly to sediments (Strachan & Kennedy 2021). As would be expected, concentrations in sediment follow a spatial gradient, with highest values in the immediate vicinity of fish cages (Bloodworth et al., 2019). The initial stages of reef decline in Loch Creran were not recorded, and we therefore do not know where the process began. To help understand the role of EMBZ, it is important to note recent surveys, which show no evidence for a spatial gradient of impact with distance from the farm (Moore et al., 2020). Moreover, it is not clear that serpulids inhabiting tubes standing erect above the seabed would have any exposure to sediment-bound EMBZ, whatever its local concentration or its effects on burrowing polychaetes.
Hypothesis 3: Azamethiphos (AZ) from salmon farm bath treatments is the cause of serpulid reef decline in Loch Creran
| Evidence supporting the hypothesis | Evidence against the hypothesis |
|---|---|
| Susceptibility to AZ | Susceptibility to AZ |
| Unknown: there have been no systematic studies of the responses of polychaetes to AZ exposure. | - |
| Timing of reef decline | Timing of reef decline |
| Use of AZ in Loch Creran began in 2012, this coincides approximately with the observed start of reef decline. | Frequent use of AZ ended in 2016 (with smaller-scale use resumed in 2021 and 22). Reef decline appears to have continued, with no evidence of recovery, throughout the period of non-use. |
| Spatial pattern of reef decline | Spatial pattern of reef decline |
| No clear predictions are possible. Although the early stages of the decline are poorly documented, the general reef decline in the Loch is consistent with a non-point source factor. Despite AZ being a point source factor, it is also highly water-soluble. This facilitates both its dispersal into the Loch but this would also be accompanied by a rapid dilution, reducing the acute toxicity over time and distance. If AZ is a factor of serpulid reef decline, a discrete pattern of decline in respect of the salmon farms would be expected. The limited information on the spatial pattern of reef decline neither supports nor contradicts the hypothesis. | - |
The information summarised in Table 9 is highly inconclusive. There is no published evidence that azamethiphos has adverse effects on S. vermicularis, with limited evidence of the effects on polychaetes in general. The initiation of azamethiphos use in 2012 is close to the observed start of reef decline, but as noted in the Literature Review, the decline may have preceded the first application of this chemical treatment. The combination of point-source application and azamethiphos being highly soluble in water does not allow any clear prediction of the spatial pattern of reef decline.
The hypothesis-based approach cannot easily be applied to heavy metals (copper, zinc) because the multiple potential sources of these metals (riverine input, aquaculture, shipping, domestic water discharge) do not allow clear predictions.
These tables focus on three environmental factors that have been suggested as potential drivers of reef decline, and as examples of non-localised and point-source factors operating in Loch Creran. In all three of these cases the available data is inadequate to disprove the hypotheses but also provide no strong support for them. The data gaps would also lead to inconclusive results if the approach was applied to the other environmental factors discussed in earlier sections of this report.
Factors influencing the presence of serpulid reefs in Loch Creran
The evaluation of the proposed factors which may contribute to the reef decline in Loch Creran would be aided by an understanding of why the reefs occur there in the first place. Unfortunately, our knowledge on this point is very limited. Bosence (1979) proposed a scenario for reef development beginning with larval settlement on discrete, isolated hard substrata (e.g. small stones or shells on a muddy seabed). With a restricted choice of available hard substrata, subsequent larval settlement occurs on the tubes of established adults, with upwards growth forming bush-like aggregations. While this may well be an accurate description of how S. vermicularis reefs became established in Loch Creran, it fails to explain why they exist there and not in many other similar localities. Shallow muddy or muddy sand seabeds with scattered stones and shells are found around the margins of many (perhaps all) sea lochs, most of which have no record of serpulid reefs. Among the sea lochs in the Firth of Lorn / Loch Linnhe system, Loch Etive has an unusual hydrography that makes it very different from the neighbouring Loch Creran (Gage 1972). However, there is no obvious reason why reefs should be absent from, for example, Lochs Craignish, Melfort, Feochan, Leven or Eil, which are more similar to Creran in size, depth and hydrography.
Loch Creran contains the largest extent of S. vermicularis reefs in the world and therefore is the focal point for discussion of their ecology. However, in some respects Loch Creran is atypical of the sites known to support (or to have supported) this feature. Linne Mhuirich and Teacuis (formerly known to support reefs and aggregations respectively) are both small inlets with narrow, restricted connections to larger sea lochs which lack reefs whereas Loch Creran opens directly to the Lynne of Lorne. The best-known Irish locality, Ardbear Salt Lake, also fits this description. The incipient reefs or aggregations in Loch Ailort occupied a very small area at the furthest extent from the Loch entrance. By contrast, Loch Creran’s reefs occur around most of the margin of the main Loch basin with high abundance (at least formerly) in South Shian Bay and neighbouring areas close to the Loch entrance (Moore et al., 1998) where they will be exposed to a high flushing rate from the incoming tide. The upper basin of Loch Creran, which more closely resembles the other Scottish and Irish localities in size and topography, has very little reef growth (Moore et al., 1998). These contrasts illustrate our poor understanding of the factors promoting reef development in sea loch environments.
It is also worth remembering that in situ recording of subtidal benthic ecosystems in most Scottish sea lochs dates back only as far as the MNCR sea loch surveys of the late 1980s/early 1990s (Howson et al. 1994), a time perspective of less than 40 years. Assessments of marine environmental change are vulnerable to the “Shifting Baseline Syndrome” (Pauly, 1995; Papworth et al., 2009), one aspect of which is the assumption that the state of a marine ecosystem when first observed by scientists represents its “natural” condition, and that any subsequent change is “unnatural” (Jackson, 2001). In 1882, Smith (1887) observed serpulids growing amongst eelgrass in the Loch Creran intertidal zone, a phenomenon which exists nowhere in the Loch today. In the late 1960s, the benthos of Loch Creran was surveyed by trawling (Gage, 1972) and in situ sampling using scuba (Gage, 1974). Neither of Gage’s papers mentions serpulid reefs, raising the possibility that they may not have existed at the time of writing. Given the short duration of scientific study we should not assume that the presence of reefs in Loch Creran is “natural” and their absence “unnatural”. Both states may be equally “natural”, depending on poorly-understood aspects of serpulid population dynamics. The same may be true for other sea lochs where S. vermicularis reefs are currently absent.
A hypothetical timeline for serpulid reefs in Loch Creran
Smith’s (1887) observations demonstrate that serpulid aggregations of some kind were present in Loch Creran in the 1880s, but this record is too isolated in time, and too different from anything that exists today, to be of much use in interpreting the current process of decline. Evidence discussed in the Literature Review section of this report, together with additional observations cited above, allow the following hypothetical timeline of reef development and decline over the past four decades:
- S. vermicularis reefs did not exist in the late 1960s but began to form during the 1980s by the process described by Bosence (1979). The cause of large-scale larval settlement at this time is unknown, but accidental observations in Loch Creran’s upper basin (see the section Reef regeneration in Loch Creran) show that previously-vacant hard substrata can be rapidly colonised in a single settlement episode. Growth of established tubes through the late 1980s gave rise to the relatively small aggregations observed in June 1989 (Connor, 1990).
- Aggregations increased in size and complexity through the 1990s by a combination of tube growth and settlement of additional larvae near the distal ends of established adult tubes (Leahy, 1991). This process gave rise to the flourishing reefs surveyed in November-December 1994 (Moore et al., 1998). Apart from localised areas of physical damage caused by trawling and boat moorings, the reefs remained in a healthy condition until at least 2005 (Moore et al., 2006; Hughes et al., 2008).
- A deterioration in reef condition was underway by at least 2011 and has continued to the present (Schuler, 2021). A failure of recruitment is a likely proximate factor contributing to decline (Harries & Moore, 2022), even if the ultimate cause(s) of this failure remain unknown. Death of older worms and bioerosion of basal tube sections will lead to the collapse and fragmentation of established aggregations. If collapsed, relict tube debris is an unattractive surface for larval settlement and/or is unsuitable for juvenile survival, as experiments suggest (Hughes, 2011; Moore et al., 2020), then reef decline driven by shortage of recruits may become a self-sustaining process.
The first section of the hypothetical timeline could be reviewed by taking account of evidence (specimens, diver observations) that modern serpulid reefs did exist in Loch Creran before the MNCR record in 1989. The logbooks of recreational divers active in the 1970s and 1980s (or scallop divers, if any were fishing Loch Creran at that time) might contain relevant observations, but to date we are not aware of any further evidence from such sources.
Knowledge gaps and future focus
The hypothesis-testing approach outlined above can produce only inconclusive results owing to the lack of key information, including:
- Demographic parameters of the serpulid population during the periods of healthy and declining reef condition, i.e. rates of recruitment, growth and mortality.
- The spatial and temporal pattern of reef deterioration.
Because the data relates to past processes, the data cannot now be recovered. Consequently, it is unlikely that the process of reef decline since (approximately) 2011 can be resolved or understood in any greater detail than it is now.
Assuming that regular SCM will be continued using established techniques, recommended additional future work in Loch Creran should concentrate on two aspects of serpulid biology that relate to the capacity of the reefs to recover and adapt to future environmental change: 1. Recruitment and 2. Diet of S.vermicularis in Loch Creran:
1. Recruitment of Serpula vermicularis in Loch Creran
If recruitment failure is a factor in reef decline, as suggested by Harries & Moore (2022), it would be informative to measure annual rates of settlement and juvenile survival at a number of sites of present (or former) reef occurrence, and non-occurrence. The selection should include sites in close proximity to the salmon farms and any other point-source inputs to the Loch. Recruitment should be measured using a standardised method based on earlier published studies (Chapman et al., 2007, Cook et al., 2021). Experiments (i.e. deployment of panels or other substrata) should be timed to cover the known late summer recruitment period in the Loch (Chapman et al., 2007). Ideally it would be useful to monitor recruitment annually on an open-ended basis, but the feasibility of this would inevitably depend on available funding and other logistical factors.
2. Diet of Serpula vermicularis in Loch Creran
There appears to be no specific information on the diet of S. vermicularis in Loch Creran. As noted previously, this lack of information is an obstacle to testing the hypothesis that change in the phytoplankton community is the cause of reef decline. In laboratory experiments, Dales (1957) found that free-swimming microalgae passed through the tentacle crowns of sabellid and serpulid tubeworms, leading him to conclude that the worms feed on suspended detritus alone. By contrast, later work has found that serpulids (including some reef-building species) do consume phytoplankton (Bruschetti et al., 2008; Leung & Cheung, 2017), a mixture of phytoplankton and heterotrophic ciliates (Pan & Marcoval, 2014) or a wide range of both eukaryotic and prokaryotic microorganisms (Jordana et al., 2001). It would be useful to clarify the diet of S. vermicularis in Loch Creran, in particular the relative importance of phytoplankton versus non-living detritus and, assuming phytoplankton are consumed, the level of selectivity (e.g. diatoms versus dinoflagellates). This information could be obtained using some or all of the following techniques:
- Laboratory measurements of retention efficiency for different phytoplankton species.
- Microscopic examination of gut contents of worms collected in the field.
- Stable isotope measurements of worm tissue to determine the trophic level occupied by S. vermicularis (i.e. relative importance of detritus and phytoplankton in the diet).
- Measurement of chemical biomarkers for different phytoplankton groups in worm tissue.
Gut content analysis should be carried out at the time of the spring phytoplankton bloom, when this potential food source is most abundant, and at other points in the year when phytoplankton are sparse. Stable isotope and biomarker analyses will provide a time-averaged index of diet over the lifetime of an individual worm. If historical specimens from Loch Creran can be located in museum or university collections, the same analyses could be carried out to investigate whether the diet of S. vermicularis in the Loch has changed over the time period covered by this report.
Serpulid reef development in Loch Ailort
Recent surveys suggested that incipient S. vermicularis aggregations / reefs were forming near the head of Loch Ailort. Unfortunately, a follow-up survey carried out in March 2023 (Sayer-Mitchell et al., 2023) failed to identify any evidence of S. vermicularis aggregations in the location. These findings suggest that a similar decline may have also occurred in Loch Ailort, as has been observed in Loch Teacuis and Loch Creran. Had the aggregations in this location persisted, it would have provided an opportunity to monitor these features and the surrounding ecology of the pelagic environment to improve our understanding of the early stages of reef development. Had such a study been in place prior to the incipient reefs disappearing, we may have been able to gain insight into the factors responsible, information we lack for the extinct reefs from other locations in Scotland. If a suitable site, supporting newly formed incipient reefs were identified again in the future, this would provide an opportunity to obtain data, which may help us to understand the factors influencing the process of reef development and decline.
Current status and extent of serpulid reefs in Loch Creran
Loss of habitat is currently estimated at around 20% and the condition of the reefs is currently ‘Unfavourable Declining’. This status was set by Moore and colleagues (2020) after diver transects between 2017-2019 and MNCR video transects in 2014. To our knowledge, the extent (in shapefile or polygon form) remains from 2005 and likely needs revision. Knowledge of the current spatial extent of the reefs could potentially form a better understanding of regional or localised impacts.
Recommendations
In the absence of any robust causal link between the widespread decline of serpulid reefs and the activities which occur in Loch Creran, it is difficult to define recommendations for future work.
If the hypothesis that decline is part of a phase of reef degradation that forms the downward section of a fluctuation in a long-term natural cycle is to be accepted, then a ‘business as usual’ scenario can be recommended, whereby continued monitoring of Loch Creran occurs and data continue to be gathered as they have since 2005. In this case, the regeneration of reefs may occur naturally but only through time and monitoring will this be shown. The main risk with this approach is that some un-recorded or unknown factor that has caused decline may have been missed and subsequently the reefs will fail to recover unless it is addressed. The status of an SAC being set to ‘unfavourable declining’ means that decline is occurring and will continue, and that more drastic action is needed than just a ‘business as usual’ response.
- Produce new shapefiles to update the 2005 data in order to estimate the current extent of S. vermicularis reefs and allow further spatial analysis mapping their decline. For example, comparison to wind speed and directions, or proximity to outfall or structures.
- If incipient aggregations are identified elsewhere in the future, tailored monitoring could be implemented to help understand optimum conditions for growth and should decline subsequently occur, the key factors influencing this.
- There are several hypothesis testing studies which could be carried out to determine or better understand key factors that may be important for the survival of S. vermicularis. For example, ex situ experiments which show optimum feeding rates or preference, or the analysis of stomach contents of some specimens around phytoplankton bloom times. As well as targeted, site-specific sampling to test for chemicals or heavy metals that might be having an impact on reef formation. Recommendations for hypothesis testing studies are outlined in sections 1 and 2 above in Knowledge gaps and future focus.
- As discussed in Knowledge gaps and future focus, deployment of settlement panels to measure annual rates of settlement and juvenile survival may help to improve our understanding of the role that recruitment plays in terms of both the observed decline and potential recovery of reef in the Loch.
- It would be useful to clarify the diet of S. vermicularis in Loch Creran, in particular the relative importance of phytoplankton versus non-living detritus and, assuming phytoplankton are consumed, the level of selectivity (e.g. diatoms versus dinoflagellates). A number of potential experimental approaches are identified in Knowledge gaps and future focus.
- Lethal Concentration studies (LC50) and Lethal Dose (LD50) experiments would demonstrate the concentrations of certain chemicals that S. vermicularis can tolerate in the water column. (Lethal Dose and Lethal Concentration studies are designed to assess acute toxicity (Lethal Dose, the amount of a substance that causes the death of 50% of the test group, so called LD50) and exposure (Lethal concentration, or the concentration of a chemical in water (or air) that causes 50% mortality in a test group, so called LC50) are usually performed over set periods of time).
- An external body or monitoring committee could be introduced to oversee and regulate activities in Loch Creran. The group could be tasked with looking after the interests of the environment and priority features in Loch Creran and increase the monitoring effort currently in place. The feasibility of this as a long-term option would be dependent on availability of public funding, which may be a limiting factor.
Conclusions
- Identification of the causal factor(s) behind reef decline in Loch Creran is not possible owing to the incompleteness of the environmental data and limited information on the spatial and temporal course of the decline.
- The hypothesis that the observed reef decline is a natural process, reflecting a decadal-scale cycle of development and senescence, is not disproven by any of the available evidence.
- There is no convincing support for any hypothesis attributing reef decline to human activities in the Loch, but in the absence of robust evidence the possibility that such a link exists cannot be ruled out.
Appendix 1
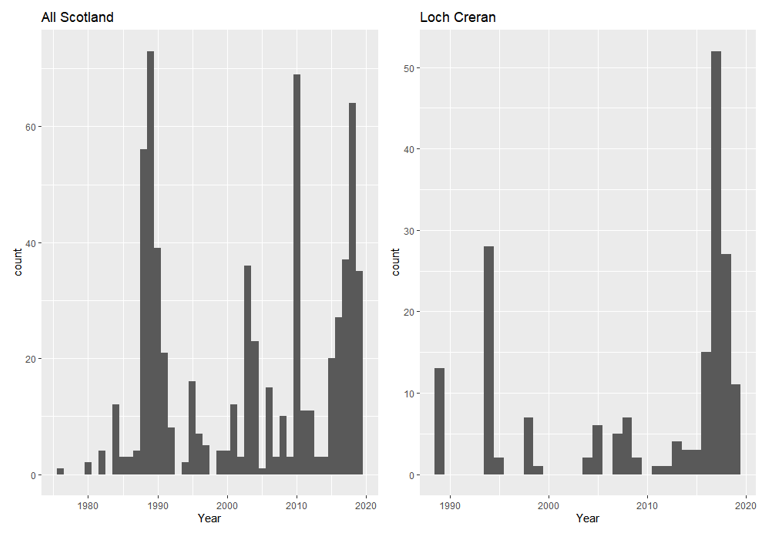
Figure A1.1. Histogram of data points included in the ordinal logistics regression models (Figure 1).
Click for a full description
Two histograms of data points, one of all of Scotland (1975 – 2020) and the second of Loch Creran (1980s – 2020). These data points were included in the ordinal logistics regression models.
Appendix 2
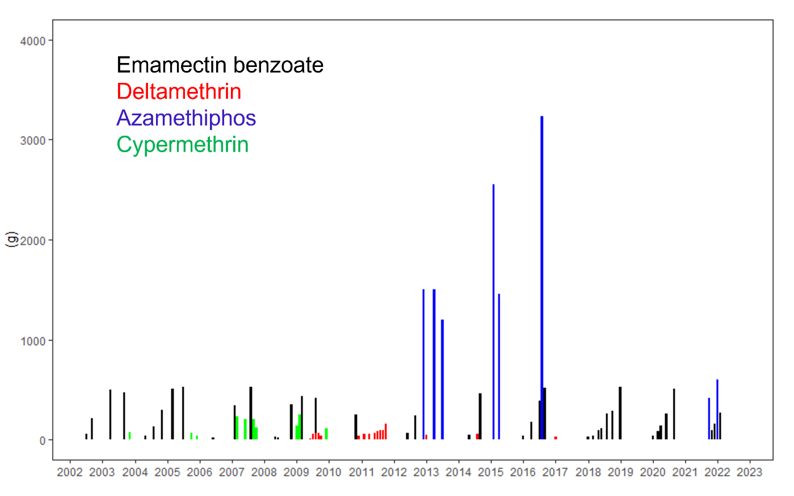
Appendix 2. Reported monthly (the same data are represented in Figure 19 but in annual form) releases (including well-boat discharges) of chemicals used in the treatment of sea-lice into Loch Creran from sites A and B.
Click for a full description
Reported monthly (the same data are represented in Figure 19 but in annual form) releases (including well-boat discharges) of chemicals used in the treatment of sea-lice into Loch Creran from sites A and B. Y axis is labelled yearly but tick marks represent the mid-point of the year (June 15th).
References
Allen, E.R. 1915. Polychaeta of Plymouth and the south Devon coast, including a list of the Archiannelida. Journal of the Marine Biological Association of the United Kingdom, 10, 592-646.
Anderson Marine Surveys, 2021. Barcaldine Hatchery Assessment of Chlorophyll Distribution and Nutrient Concentrations (Comissionned Report 21/08.1 rev0). AMSL.
Argyll and Bute Council, 2007. Loch Creran Marine Special Area of Conservation Management Plan.
Baden, S., Hernroth, B. & Lindahl, O. 2021. Declining populations of Mytilus spp. in North Atlantic coastal waters – a Swedish perspective Journal of Shellfish Research, 40, 2.
Barbosa, R.V., Jaud, M., Bacher, C., Kerjean, Y., Jean, F., Ammann, J. & Thomas, Y. 2022. High-resolution drone images show that the distribution of mussels depends on microhabitat features of intertidal rocky shores. Remote Sensing, 14, 21.
BBC, 2021. Dead crabs and fish washed up on Teeside beaches BBC England News 25 October 2021.
BBC, 2023. Shellfish deaths: minister rejects call for further probe BBC England News 10 February 2023.
Bedford, J., Ostle, C., Johns, D. G., Atkinson, A., Best, M., Bresnan, E., Machairopoulou, M., Graves, C. A., Devlin, M., Milligan, A., Pitois, S., Mellor, A., Tett, P., & McQuatters-Gollop, A. 2020. Lifeform indicators reveal large-scale shifts in plankton across the North-West European shelf Global Change Biology, 26(6), 3482–3497.
Bianchi, C.N. & Morri, C. 2001. The battle is not to the strong: serpulid reefs in the Lagoon of Orbatello (Tuscany, Italy). Estuarine, Coastal and Shelf Science, 53, 215–220.
Black, K.D., Hughes, D.J., Provost, P.G., & Pereira, P.M.F. 2000. Broad scale survey and mapping of seabed biota in Loch Creran, Argyll. Scottish Natural Heritage Commissioned Report F98AA408.
Bloodworth, J. W., Baptie, M. C., Preedy, K. F., & Best, J. 2019. Negative effects of the sea lice therapeutant emamectin benzoate at low concentrations on benthic communities around Scottish fish farms Science of The Total Environment, 669, 91–102.
Bosence, D.W.J. 1979. The factors leading to aggregation and reef formation in Serpula vermicularis L. In Larwood G. and Rosen B.R. (eds) Biology and Systematics of Colonial Organisms. Systematics Association Special Volume No. 11. London and New York: Academic Press, pp. 299–318.
Bosence, D.W.J. 1973. Recent serpulid reefs, Connemara, Eire. Nature, 242, 40-41.
Brown, L., Bresnan, E., Graham, J., Lacaze, J.-P., Turrell, E., & Collins, C. 2010. Distribution, diversity and toxin composition of the genus Alexandrium (Dinophyceae) in Scottish waters European Journal of Phycology, 45(4), 375–393.
Bruschetti., M., Luppi, T., Fanjul, E., Rosenthal, A. & Iribarne, O. 2008. Grazing effect of the invasive reef-forming polychaete Ficopomatus enigmaticus (Fauvel) on phytoplankton biomass in a SW Atlantic coastal lagoon Journal of Experimental Marine Biology and Ecology, 354, 212–219.
Buckman, J.O. & Harries, D.B. 2020. Reef-forming Serpula vermicularis from Scotland and Ireland: tube structure, composition and implications. Zoologischer Anzeiger, 288, 53–65.
Burridge, L., Weis, J.S., Cabello, F., Pizarro, J. & Bostick, K. 2010. Chemical use in salmon aquaculture: a review of current practices and possible environmental effects. Aquaculture, 306(1–4), 7–23.
Capa, M., Kupriyanova, E., Nogueira, J. M. de M., Bick, A., & Tovar-Hernández, M.A. 2021. Fanworms: Yesterday, Today and Tomorrow Diversity, 13(3), 130.
Carlton, J.T., Geller, J.B., Reaka-Kudler, M.L. & Norse, E.A. 1999. Historical extinctions in the sea. Annual Review of Ecology and Systematics, 30, 515–538.
Chapman, N.D., Moore, C.G., Harries, D.B. & Lyndon, A.R. 2007. Recruitment patterns of Serpula vermicularis L. (Polychaeta, Serpulidae) in Loch Creran, Scotland. Estuarine, Coastal and Shelf Science, 73, 598–606.
Cheng, B., Van Smeden, J., Deneer, J., Belgers, D., Foekema, E., Roessink, I., Matser, A., & Van den Brink, P. J. 2020. The chronic toxicity of emamectin benzoate to three marine benthic species using microcosms Ecotoxicology and Environmental Safety, 194, 110452.
Christie, H., Kraufvelin, P., Kraufvelin, L., Niemi, N., & Rinde, E. 2020. Disappearing Blue Mussels – Can Mesopredators Be Blamed? Frontiers in Marine Science, 7.
Collins, C., Graham, J., Brown, L., Bresnan, E., Lacaze, J.-P., & Turrell, E. A. 2009. Identification and Toxicity of Alexandrium Tamarense (dinophyceae) in Scottish Waters Journal of Phycology, 45(3), 692–703.
Connor, D.W. 1990. Survey of Loch Linne, Eil, Creran and Aline, Nature Conservancy Council CSD Report 1073 (MNCR/SR/12), Nature Conservancy Council, Peterborough.
Cook, R. L., Sanderson, W. G., Moore, C. G., & Harries, D. B. 2021. The right place at the right time: improving the odds of biogenic reef restoration Marine Pollution Bulletin, 164, 112022.
Cook, R.L. 2016. Development of Techniques for the Restoration of Temperate Biogenic Reefs. PhD dissertation. Heriot-Watt University, Edinburgh.
Crustacean Mortality Expert Panel, 2023. Independent Expert Assessment of Unusual Crustacean Mortality in the North-east of England in 2021 and 2022
Dales, R.P. 1957. Some quantitative aspects of feeding in sabellid and serpulid fan worms. Journal of the Marine Biological Association of the United Kingdom, 36, 309-316.
Daoud, D., McCarthy, A., Dubetz, C., & Barker, D.E. 2018. The effects of emamectin benzoate or ivermectin spiked sediment on juvenile American lobsters (Homarus americanus) Ecotoxicology and Environmental Safety, 163, 636–645.
Dill, L.M., & Fraser, A.H.G. 1997. The worm re-turns: Hiding behavior of a tube-dwelling marine polychaete, Serpula vermicularis Behavioral Ecology, 8(2), 186–193.
Dodd, J., Baxter, L. & Hughes, D.J. 2009. Mapping Serpula vermicularis (Polychaeta: Serpulidae) aggregations in Loch Teacuis, western Scotland, a new record. Marine Biology Research, 5, 200-205.
Donovan, T. 2006. A comprehensive guide to Loch Creran Marine Special Area of Conservation. Creran Publishing. ISBN 978-0-9556599-0-4.
Dungan, M.L., Miller, T.E., & Thomson, D.A. 1982. Catastrophic Decline of a Top Carnivore in the Gulf of California Rocky Intertidal Zone Science, 216(4549), 989–991.
Earll, R. 1984. Species and communities of Loch Sween. Marine Conservation Society (Nature Conservancy Council) Ross-on-Wye, Herefordshire.
Edwards, A. & Sharples, F. 1986. Scottish Sea Lochs: A Catalogue, Scottish Marine Biological Association, Oban.
Elmhirst, R., 1922. Notes on the breeding and growth of marine animals in the Clyde Sea area. Report of the Scottish Marine Biological Association, 19-43.
Environment Agency, 2019. Cypermethrin: Sources, pathways and environmental data
Fang, J., Samuelsen, O.B., Strand, Ø., & Jansen, H. 2018. Acute toxic effects of hydrogen peroxide, used for salmon lice treatment, on the survival of polychaetes Capitella sp. and Ophryotrocha spp Aquaculture Environment Interactions, 10, 363–368.
Fehling, J., Davidson, K., Bolch, C., & Tett, P. 2006. Seasonality of Pseudo-nitzschia spp. (Bacillariophyceae) in western Scottish waters Marine Ecology Progress Series, 323, 91–105.
Ferreira, J. G., Sequeira, A., Hawkins, A. J. S., Newton, A., Nickell, T. D., Pastres, R., Forte, J., Bodoy, A., & Bricker, S. B. 2009. Analysis of coastal and offshore aquaculture: Application of the FARM model to multiple systems and shellfish species Aquaculture, 289(1), 32–41.
Findlay, K. 2016. Fish farm firm buys up Marine Resource Centre for £35million The Press and Journal, 7 July.
Food Standards Agency, 2013. Sanitary Survey Review Loch Creran 2013
Food Standards Scotland, 2016. Shellfish Safety and Sanitation
Franchini, A., Malagoli, D., & Ottaviani, E. 2010. Targets and Effects of Yessotoxin, Okadaic Acid and Palytoxin: A Differential Review. Marine Drugs, 8(3), Article 3.
Gage, J.D. 1972. A preliminary survey of the benthic macrofauna and sediments in Lochs Etive and Creran, sea-lochs along the west coast of Scotland. Journal of the Marine Biological Association of the United Kingdom, 52, 237-276.
Gage, J. 1974. Shallow water zonation of sea loch benthos and its relation to hydrographic and other physical features. Journal of the Marine Biological Association of the United Kingdom, 54, 223–249.
Gillibrand, P.A., Inall, M.E., Portilla, E., & Tett, P. 2013. A box model of the seasonal exchange and mixing in Regions of Restricted Exchange: Application to two contrasting Scottish inlets. Environmental Modelling & Software, 43, 144–159.
Harnik, P.G., Lotze, H.K., Anderson, S.C., Finkel, Z.V., Finnegan, S., Lindberg, D.R., Liow, L.S., Lockwood, R., McClain, C.R., McGuire, J.L., O’Dea, A., Pandolfi, J.M., Simpson, C. & Tittensor, D.P. 2012. Extinctions in ancient and modern seas. Trends in Ecology and Evolution, 27, 608-617.
Harries, D.B. & Moore, C.G. 2022. Loch Creran serpulid regeneration monitoring – field report. Report to NatureScot from Heriot-Watt University, September 2022.
Haya, K., Burridge, L.E., Davies, I.M. and Ervik, A. 2005. A review and assessment of environmental risk of chemicals used for the treatment of sea lice infestations of cultured salmon. Environmental effects of marine finfish aquaculture, 305-340.
Henry, L.M., Kennedy, R. & Keegan, B.F. 2008. An investigation of periodic hypoxia in Ardbear Salt Lake. Journal of the Marine Biological Association of the United Kingdom, 88, 1297-1307.
Hewson, I., Button, J. B., Gudenkauf, B. M., Miner, B., Newton, A. L., Gaydos, J. K., Wynne, J., Groves, C. L., Hendler, G., Murray, M. 2014. Densovirus associated with sea-star wasting disease and mass mortality. Proceedings of the National Academy of Sciences U.S.A., 111, 17278–17283.
Hewson, I., Ritchie, I.T., Evans, J.S. et al. 2023. A scuticociliate causes mass mortality of Diadema antillarum in the Caribbean Sea. Science Advances, 9, eadg3200.
Hinder, S.L., Hays, G.C., Brooks, C.J., Davies, A.P., Edwards, M., Walne, A.W., & Gravenor, M.B. 2011. Toxic marine microalgae and shellfish poisoning in the British isles: history, review of epidemiology, and future implications Environmental Health, 6, 10-54.
Holt, T.J., Rees, E.I., Hawkins, S.J. & Seed, R., 1998. Biogenic reefs (Volume IX). An overview of dynamic and sensitivity characteristics for conservation management of marine SACs Scottish Association for Marine Science (UK Marine SACs Project), 174 pp.
Howe, J.A., Austin, W.E.N., Forwick, M., Paetzel, M., Harland, R., & Cage, A.G. 2010. Fjord systems and archives: A review. Geological Society, London, Special Publications, 344(1), 5–15.
Howson, C.M., Connor, D.W. & Holt, R.H.F. 1994. The Scottish sea lochs – an account of surveys undertaken for the MNCR. Joint Nature Conservation Committee Report, no. 164. [Marine Nature Conservation Review Report no. MNCR/SR/27.]
Hughes, D.J. 2011. Where’s the ‘reef’? A five year study of serpulid tube bioerosion in a Scottish sea loch Marine Ecology Progress Series, 430, 273–280.
Hughes, D.J., Hatton, A.D. & Nickell, L.A. 2004. Do biogenic sulphur compounds in phytodetritus act as cues for deposit-feeder activity? Field experiments and observations on the echiuran worm Maxmuelleria lankesteri. Journal of Experimental Marine Biology and Ecology, 313, 175-189.
Hughes, D.J., Poloczanska, E.S. & Dodd, J. 2008. Survivorship and tube growth of reef-building Serpula vermicularis (Polychaeta: Serpulidae) in two Scottish sea lochs. Aquatic Conservation: Marine and Freshwater Ecosystems, 18, 117-129.
Hughes, T.P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science, 265, 1547-1551.
Hylkema, A., Kitson-Walters, K., Kramer, P.R., Patterson, J.T., Roth, L., Sevier, M.L.B., Vega-Rodriguez, M., Warham, M.W., Williams, S.M. & Lang, J.C. 2023. The 2022 Diadema antillarum die-off event: comparisons with the 1983-1984 mass mortality. Frontiers in Marine Science, 9, 1067449.
Jackson, J.B.C. 2001. What was natural in the coastal oceans? Proceedings of the National Academy of Sciences USA, 98, 5411-5418.
James, K. J., Fidalgo Sáez, M. J., Furey, A., & Lehane, M. 2004. Azaspiracid poisoning, the food-borne illness associated with shellfish consumption Food Additives & Contaminants, 21(9), 879-892.
Jansson, R. K., Brown, R., Cartwright, B., Cox, D., Dunbar, D. M., Dybas, R. A., Eckel, C., Lasota, J. A., Mookerjee, P. K., Norton, J. A., Peterson, R. F., Starner, V. R., & White, S. 1997. Emamectin benzoate: a novel avermectin derivative for control of lepidopterous pests. Proceedings of the 3rd International Workshop on Management of Diamondback Moth and Other Crucifer Pests., 8.
JNCC, 2017. SACFOR abundance scale
Johnston, E.L. & Keough, M.J. 2002. Direct and indirect effects of repeated pollution events on marine hard‐substrate assemblages. Ecological Applications, 12(4), 1212-1228.
Jones, K.J. 1979. Studies on nutrient levels and phytoplankton growth in a Scottish sea-loch. PhD thesis. University of Strathclyde, Glasgow.
Jones C.G., Lawton, J.H. & Shachak, M. 1994. Organisms as ecosystem engineers. Oikos, 69, 373–386
Jordana, E., Charles, F., Grémare, A., Amouroux, J.-M. & Chrétiennot-Dinet, M.-J. 2001. Food sources, ingestion and absorption in the suspension-feeding polychaete Ditrupa arietina (O.F. Müller). Journal of Experimental Marine Biology and Ecology, 266, 219-236.
Jordana, E., Duchêne, J.-C., Charles, F., Grémare, A., & Amouroux, J.-M. 2000. Experimental study of suspension-feeding activity in the serpulid polychaete Ditrupa arietina (O.F. Müller) Journal of Experimental Marine Biology and Ecology, 252(1), 57–74.
Kamphausen, L. 2015. Loch Teacuis serpulid survey March 2015. Report to Scottish Natural Heritage.
Kamphausen, L., Henderson, S., Kent, F., MacMinn, R., Beaton, J., Dodd, J., McGrath, O. & James, B. 2018. Loch Ailort serpulid diving survey and drop-down video survey April 24-28, 2017. Scottish Natural Heritage Post-Survey Summary.
Kendon, M., McCarthy, M., Jevrejeva, S., Matthews, A., Sparks, T., & Garforth, J. 2021. State of the UK Climate 2020 International Journal of Climatology, 41(S2), 1–76.
Kliegman, R.M., Geme III, J.W.S., Blum, N., Shah, S.S. & Tasker, R.C. eds. 2020. Nelson Textbook of Pediatrics. Elsevier Health Sciences.
Kupriyanova, E.K., Nishi, E., ten Hove, H.A. & Rzhavsky, A.V. 2001. Life-history patterns in serpulimorph polychaetes: ecological and evolutionary perspectives. Oceanography and Marine Biology: An Annual Review, 39, 1–101.
Kupriyanova, E.K. and Havenhand, J.N. 2005. Effects of temperature on sperm swimming behaviour, respiration and fertilization success in the serpulid polychaete, Galeolaria caespitosa (Annelida: Serpulidae). Invertebrate Reproduction & Development, 48(1-3), 7-17.
Laurent, C.D.C. 2009. Studying the Assimilative Capacity of Loch Creran. Edinburgh Napier University.
Laurent, C., Tett, P., Fernandes, T., Gilpin, L., & Jones, K. 2006. A dynamic CSTT model for the effects of added nutrients in Loch Creran, a shallow fjord Journal of Marine Systems, 61(3), 149–164.
Leahy, Y. 1991. Polychaete studies on the west coast of Ireland. I. Benthic characterisation of the Dunkellin estuary with particular reference to the Polychaeta. II. Aspects of the biology and ecology of the serpulid Serpula vermicularis Linnaeus from Ardbear Salt Lake. PhD thesis. University College Galway, Ireland.
Lelong, A., Hégaret, H., Soudant, P., & Bates, S.S. 2012. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms Phycologia, 51(2), 168–216.
Lennox, R. J., Salvanes, A. G. V., Barlaup, B. T., Stöger, E., Madhun, A., Helle, T. M., & Vollset, K. W. 2020. Negative impacts of the sea lice prophylactic emamectin benzoate on the survival of hatchery released salmon smolts in rivers Aquatic Toxicology, 224, 105519.
Lessios, H.A. 1988. Mass mortality of Diadema antillarum in the Caribbean: what have we learned? Annual Review of Ecology and Systematics, 19, 371-393.
Lessios, H.A. 2016. The great Diadema antillarum die-off: 30 years later. Annual Review of Marine Science, 8, 267-283.
Lessios, H.A., Cubit, J.D., Robertson, D.R., Shulman, M.J., Parker, M.R., Garrity, S.D. & Levings, S.C. 1984. Mass mortality of Diadema antillarum on the Caribbean coast of Panama. Coral Reefs, 3, 173-182.
Leung, J.Y.S., & Cheung, N.K.M. 2017. Feeding behaviour of a serpulid polychaete: Turning a nuisance species into a natural resource to counter algal blooms? Marine Pollution Bulletin, 115(1–2), 376–382.
Leung, Y.S., Shin, P.K.S., Qiu, J.W., Ang, P.O., Chiu, J.M.Y., Thiyagarajan, V., & Cheung, S.G. 2013. Physiological and behavioural responses of different life stages of a serpulid polychaete to hypoxia Marine Ecology Progress Series, 477, 135–145.
Lumb, C.M. 1986. Loch Sween sublittoral survey, August 27 to September 8, 1984. Nature Conservancy Council CSD Rep No 722. Nature Conservancy Council, Peterborough.
Marion, G.M., Millero, F.J., Camões, M.F., Spitzer, P., Feistel, R. and Chen, C.T. 2011. pH of seawater. Marine Chemistry, 126(1-4), 89-96.
Mayor, D.J., Solan, M., Martinez, I., Murray, L., McMillan, H., Paton, G.I. and Killham, K. 2008. Acute toxicity of some treatments commonly used by the salmonid aquaculture industry to Corophium volutator and Hediste diversicolor: Whole sediment bioassay tests. Aquaculture, 285(1-4), 102-108.
McBriarty, G. J., Kidd, K. A., & Burridge, L. E. 2018. Short-term effects of the anti-sea lice therapeutant emamectin benzoate on clam worms (Nereis virens)Archives of Environmental Contamination and Toxicology, 74(4), 539–545.
Medina, M., Barata, C., Telfer, T., & Baird, D.J. 2004. Assessing the Risks to Zooplankton Grazers of Continuous Versus Pulsed Cypermethrin Exposures from Marine Cage Aquaculture. Archives of Environmental Contamination and Toxicology, 47(1), 67–73.
Met Office, 2011. Early winter storms, late 2011
Mieszkowska, N., Leaper, R., Moore, P., Kendall, M., Burrows, M., Lear, D., Poloczanska, E., Hiscock, K., & Thompson, R. 2005. Marine Biodiversity and Climate Change (MarClim) Assessing and predicting the influence of climatic change using intertidal rocky shore biota. Final Report for United Kingdom Funders. Marine Biological Association Occasional Publications, 20, 53p.
Minchin, D. 1987. Serpula vermicularis L. (Polychaeta: Serpulidae) reef communities from the west coast of Ireland. Irish Naturalists’ Journal, 22, 314–316.
Montefalcone, M., Oprandi, A., Azzola, A., Morri, C. & Bianchi, C.N. 2022. Serpulid reefs and their role in aquatic ecosystems: a global review. Advances in Marine Biology, 92, 1–54.
Moore, C.G., & Harries, D.B. 2020. Monitoring for the possible spread of Didemnum vexillum into the subtidal environment of Loch Creran. Scottish Natural Heritage Research Report 1155.
Moore C.G., Bates C.R., Mair J.M., Saunders G.R., Harries D.B. & Lyndon A.R. 2009. Mapping serpulid worm reefs (Polychaeta: Serpulidae) for conservation management. Aquatic Conservation: Marine and Freshwater Ecosystems, 19, 226–236.
Moore C.G., Saunders G.R., Harries D.B., Mair J.M., Bates C.R. & Lyndon A.R. 2006. The establishment of site condition monitoring of the subtidal reefs of Loch Creran Special Area of Conservation. Scottish Natural Heritage Commissioned Report No. 151.
Moore, C.G. 1996. The distribution of serpulid reefs in Loch Creran, Argyll. Scottish Natural Heritage Research, Survey and Monitoring Report No. 53.
Moore, C.G., Saunders, G.R., & Harries, D.B. 1998. The status and ecology of reefs of Serpula vermicularis L. (Polychaeta: Serpulidae) in Scotland Aquatic Conservation: Marine and Freshwater Ecosystems, 8(5), 645–656.
Moore, C.G. 2019. Biological analyses of underwater video from monitoring and research cruises in Lochs Ailort and Fyne, the Sounds of Barra and Mull, inner Moray Firth, off Wester Ross, Noss Head and Rattray Head, and around the Southern Trench in outer Moray Firth. Scottish Natural Heritage Research Report No. 1085.
Moore, C.G., Harries, D.B., Atkinson, R.J.A., Clark, L., Cook, R.L., Hirst, N.E., Saunders, G.R., Lyndon, A.R., Sanderson, W.G., & Porter, J.S. 2013. The distribution and condition of proposed protected features within the Loch Sween possible Nature Conservation MPA (No. 621).
Moore, C.G., Harries, D.B., Lyndon, A.R., Saunders, G.R. & Conway, T.R. 2003. Quantification of serpulid biogenic reef coverage of the sea bed (Polychaeta: Serpulidae) using a video transect technique. Aquatic Conservation: Marine and Freshwater Ecosystems, 13, 137-146.
Moore, C.G., Harries, D.B., Tulbure, K.W., Cook, R.L., Saunders, G.R., Lyndon, A.R., Kamphausen, L. & James, B. 2020. The current status of serpulid reefs, horse mussel beds and flame shell beds in Loch Creran SAC and MPA. Scottish Natural Heritage Research Report No. 1156.
Moschonas, G., Gowen, R.J., Paterson, R.F., Mitchell, E., Stewart, B.M., McNeill, S., Glibert, P.M., & Davidson, K. 2017. Nitrogen dynamics and phytoplankton community structure: The role of organic nutrients Biogeochemistry, 134(1), 125–145.
Murray, L. 2009. Algal Toxins in Shellfish Monitoring Programme Scotland (Implementation of EC 854/2004).
Nippard, L., & Ciocan, C. 2019. Potential impact of aquaculture effluents in Loch Creran, Scotland. Vie et Milieu, 1(69), 47–52.
Oulhen, N., Byrne, M., Duffin, P., Gomez-Chiarri, M., Hewson, I., Hodin, J., Konar, B., Lipp, E.K., Miner, B.G., Newton, A.L., Schiebelhut, A.M., Smolowitz, R., Wahltinez, S.J., Wessel, G.M., Work, T.M., Zaki, H.A. & Wares, J.P. 2022. A review of asteroid biology in the context of sea star wasting: possible causes and consequences The Biological Bulletin, 243(1), 50–75.
Paerl, H.W. 1997. Coastal eutrophication and harmful algal blooms: Importance of atmospheric deposition and groundwater as “new” nitrogen and other nutrient sources. Limnology and oceanography, 42(5part2), 1154–1165.
Pan, J. & Marcoval, M.A. 2014. Top-down effects of an exotic serpulid polychaete on natural plankton assemblage of estuarine and brackish systems in the SW Atlantic Journal of Coastal Research, 30(6), 1226-1235.
Papworth, S.K., Rist, J., Coad, L. & Millner-Gulland, E.J. 2009. Evidence for shifting baseline syndrome in conservation. Conservation Letters, 2, 93–100.
Parks, R., Swan, S., Davidson, K., Turner, A., Powell, A., Ford, C., & Petch, R. G. 2019. Annual report on the results of the E. coli, biotoxin, phytoplankton and chemical contaminants Official Control Monitoring Programmes for Scotland – 2018 (C7711 to C7715). Cefas.
Pauly, D. 1995. Anecdotes and the shifting baseline syndrome of fisheries. Trends in Ecology and Evolution, 10, 430.
Poloczanska, E.S., Hughes, D.J. & Burrows, M.T. 2004. Underwater television observations of Serpula vermicularis (L.) reefs and associated mobile fauna in Loch Creran, Scotland. Estuarine, Coastal and Shelf Science, 61, 425–435.
Rain-Franco, A., Rojas, C. and Fernandez, C. 2018. Potential effect of pesticides currently used in salmon farming on photo and chemoautotrophic carbon uptake in central–southern Chile. Aquaculture, 486, 271–284.
Reguera, B., Riobó, P., Rodríguez, F., Díaz, P.A., Pizarro, G., Paz, B., Franco, J.M., & Blanco, J. 2014. Dinophysis toxins: causative organisms, distribution and fate in shellfish Marine Drugs, 12(1), 394-461.
Riedi, M.A. & Smith, A.M. 2015. Tube growth and calcification of two reef-building ecosystem engineers in southern New Zealand: Galeolaria hystrix and Spirobranchus cariniferus (Polychaeta: Serpulidae). Marine Geology, 367, 212-219.
Rossbach, F. I., Casoli, E., Beck, M., & Wild, C. 2021. Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes Diversity, 13(6), 6.
Santonnini, J., Ruso, Ruso, Y.D.P., Melendreras, E.C. & Casalduero, F.G. 2021. Massive aggregations of Serpulidae associated with eutrophication of the Mar Menor, southeast Iberian Peninsula. Frontiers in Marine Science, 7, 531726.
Sayer-Mitchell, J.D., Roberts, K.C., Schulz, P.H. and Brown, H. 2023. Loch Ailort serpulids: diving, ROV, and multibeam survey, 27 February – 3 March 2023. NatureScot Survey Report.
Schoenrock, K.M., O’Callaghan, R., O’Callaghan, T., O’Connor, A. & Stengel, D.B. 2021. An ecological baseline for Laminaria hyperborea forests in western Ireland. Limnology and Oceanography, 6, 3439-3454.
Schuler, M. 2021. Condition assessment of Priority Marine Feature Serpula vermicularis reefs in Loch Creran MPA, Scotland, using video analysis. BSc. (Hons.) project, Institute of Earth and Life Sciences, Heriot-Watt University.
Schwindt, E., De Francesco, C.G. & Iribarne, O.O. 2004a. Individual and reef growth of the introduced reef-building polychaete Ficopomatus enigmaticus in a south-western Atlantic coastal lagoon. Journal of the Marine Biological Association of the United Kingdom, 84, 987–993.
Schwindt, E., Iribarne O.O. & Isla, F.I. 2004b. Physical effects of an invading reef-building polychaete on an Argentinean estuarine environment. Estuarine Coastal and Shelf Science, 59, 109-120
Seasearch, 2017. Annual report – Argyll 2017
Seasearch, 2018. Annual report – Argyll 2018
Seasearch, 2021. Annual report – West of Scotland 2021
Seasearch, 2022. Serpula project update – 27 February 2022
Selkie Associates, 1999. Serpulid reef survey in the Linne Mhuirich, Loch Sween, November 6th, 7th and 20th 1999. Report to Scottish Natural Heritage, Edinburgh.
SEPA, 2011. Loch Creran, Argyll – UKS7992332
SEPA, 2018. SEPA Position Statement to support the implementation of the Water Environment (Controlled Activities) (Scotland) Regulations 2011 Interim position statement for protecting the marine environment until such time as a Ministerial direction is issued on environmental standards for emamectin benzoate.
Smith, A.M., McGourty, C.R., Kregting, L. & Elliot, A. 2005. Subtidal Galeolaria hystrix (Polychaeta: Serpulidae) reefs in Paterson Inlet, Stewart Island, New Zealand. New Zealand Journal of Marine and Freshwater Research, 39, 1297–1304.
Smith, A.W. 1887. Loch Creran: Notes from the West Highlands. Paisley: Alexander Gardner.
Smith, S., Hewson, I. & Collins, P. 2022. The first records of sea star wasting disease in Crossaster papposus in Europe. Biology Letters, 18, 20220197.
SNH, 2010. Condition of designated sites.
Solter, P.F., & Beasley, V.R. 2013. Chapter 38 – Phycotoxins. In: Haschek, W.M., Rousseaux, C.G. & Wallig, M.A. eds. Haschek and Rousseaux’s Handbook of Toxicologic Pathology, third edition. Academic Press, 1155–1186.
Stone, J., Sutherland, I.H., Sommerville, C.S., Richards, R.H., & Varma, K.J. 2002. The efficacy of emamectin benzoate as an oral treatment of sea lice, Lepeophtheirus salmonis (Kròyer), infestations in Atlantic salmon, Salmo salar L. Journal of Fish Diseases, 22(4), 261–270.
Stone, J., Sutherland, I.H., Sommerville, C., Richards, R.H., & Varma, K.J. 2000. Commercial trials using emamectin benzoate to control sea lice Lepeophtheirus salmonis infestations in Atlantic salmon Salmo salar Diseases of Aquatic Organisms, 41(2), 141–149.
Strachan, F. & Kennedy, C.J. 2021. The environmental fate and effects of anti-sea lice chemotherapeutants used in salmon aquaculture. Aquaculture, 544, 737079.
Telfer, T.C., Baird, D.J., McHenery, J.G., Stone, J., Sutherland, I., & Wislocki, P. 2006. Environmental effects of the anti-sea lice (Copepoda: Caligidae) therapeutant emamectin benzoate under commercial use conditions in the marine environment Aquaculture, 260(1), 163–180.
ten Hove H.A & van den Hurk P. 1993. A review of recent and fossil serpulid ‘reefs’, actuopaleontology and the ‘Upper Malm’ serpulid limestones in NW Germany. Geologie en Mijnbouw, 72, 23–67.
ten Hove, H.A.T., & Kupriyanova, E.K. 2009. Taxonomy of Serpulidae (Annelida, Polychaeta): The state of affairs Zootaxa, 2036(1), 1–126.
ten Hove, H.A. 1979. Different causes of mass occurrence in serpulids. In: Larwood, G. & Rosen, B.R. eds. Biology and Systematics of Colonial Organisms. Systematics Association Special vol. 11. Academic Press, London, 281–298.
Tett, P. 2019. Long-term changes in Phytoplankton Lifeforms and SST at the Lorn Pelagic Observatory, v.1 (296; SAMS Internal Report, p. 36).
Tett, P., & Wallis, A. 1978. The general annual cycle of chlorophyll standing crop in Loch Creran Journal of Ecology, 66(1), 227–239.
Tett, P., Droop, M.R. & Heaney, S.I. 1985. The Redfield ratio and phytoplankton growth rate. Journal of the Marine Biological Association of the United Kingdom, 65(2), 487–504.
Tett, P., Carreira, C., Mills, D.K., Van Leeuwen, S., Foden, J., Bresnan, E. and Gowen, R.J. 2008. Use of a Phytoplankton Community Index to assess the health of coastal waters. ICES Journal of Marine Science, 65(8), 1475–1482.
Tett, P. 2013. Creran Microplankton in 2012 SAMS internal report 285, Scottish Association for Marine Science, Oban.
Tett, P., Benjamins, S., Black, K.D., Coulson, M., Davidson, K., Fernandes, T., Fox, C., Hart, M., Hicks, N., Hughes, A., Hunter, D-C., Nickell, T., Risch, D., Tocher, D., Vare, L., Vespoor, E., Wilding, T., Wilson, B., & Wittich, A. 2018. Review of the environmental impacts of salmon farming in Scotland. Report for the Environment, Climate Change and Land Reform (ECCLR) Committee. The Scottish Parliament, 196 pp.
Tillmann, U., Elbrächter, M., Krock, B., John, U., & Cembella, A. 2009. Azadinium spinosum gen. Et sp. Nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins European Journal of Phycology, 44(1), 63–79.
Tulbure, K. 2015. Investigating the condition of the Priority Marine Feature, ‘Serpulid aggregations’ (Serpula vermicularis) in Loch Creran. Honours Project (BSc) Heriot Watt University.
Turrell, W., R., Dalgarno, E., Hermann, G., Graham, J., Lacaze, J.-P., & Rabe, B. 2020. The Thermal Environment of an Inter-tidal Pacific Oyster Farm. Scottish Marine and Freshwater Science Report, 11(15). Marine Scotland.
Turvey, S.T. & Risley, C.L. 2006. Modelling the extinction of Steller’s sea cow. Biology Letters, 2, 94–97.
Van Geest, J.L., Burridge, L.E. and Kidd, K.A. 2014. The toxicity of the anti-sea lice pesticide AlphaMax® to the polychaete worm Nereis virens. Aquaculture, 430, 98–106.
Waddy, S.L., Burridge, L.E., Hamilton, M.N., Mercer, S.M., Aiken, D.E. and Haya, K. 2002. Rapid communication/communication rapide emamectin benzoate induces molting in American lobster, Homarus americanus. Canadian Journal of Fisheries and Aquatic Sciences, 59(7), 1096-1099.
Wang, Z., Nie, X. & Yue, W. 2011. Toxicological effects of cypermethrin to marine phytoplankton in a co-culture system under laboratory conditions, Ecotoxicology, 20, 1258–1267.
Wang, W., Shi, K., Zhang, Y., Li, N., Sun, X., Zhang, D., Zhang, Y., Qin, B., & Zhu, G. 2022. A ground-based remote sensing system for high-frequency and real-time monitoring of phytoplankton blooms. Journal of Hazardous Materials, 439, 129623.
Ward-Paige, C.A., Mora, C., Lotze, H.K., Pattengill-Semmens, C., McClenachan, L., Arias-Castro, E. & Myers, R.A. 2010. Large-scale absence of sharks on reefs in the Greater Caribbean: a footprint of human pressures. PloS One, 5(8), 11968.
Watson, G. J., Leach, A., & Fones, G. 2008. Effects of copper and other metals on fertilization, embryo development, larval survival and settlement of the polychaete Nereis (Neanthes) virens Invertebrate Reproduction & Development, 52(1–2), 101–112.
West, H., Quinn, N., & Horswell, M. 2019. Regional rainfall response to the North Atlantic Oscillation (NAO) across Great BritainHydrology Research, 50(6), 1549–1563.
Whyte, C. T. 2012. An Investigation into changes in the phytoplankton community in Loch Creran, a Scottish sea loch. Edinburgh Napier University.
Whyte, C., Davidson, K., Gilpin, L., Mitchell, E., Moschonas, G., McNeill, S., & Tett, P. 2017. Tracking changes to a microplankton community in a North Atlantic sea loch using the microplankton index PI(mp). ICES Journal of Marine Science: Journal du Conseil.
Willis, K.J., & Ling, N. 2003. The toxicity of emamectin benzoate, an aquaculture pesticide, to planktonic marine copepods Aquaculture, 221(1), 289–297.
Willis, K.J., Gillibrand, P.A., Cromey, C.J., & Black, K.D. 2005. Sea lice treatments on salmon farms have no adverse effects on zooplankton communities: A case studyMarine Pollution Bulletin, 50(8), 806–816.
Wilson, A., Nickell, T., Cromey, C., & Black, K. 2006. ECASA Study Site Report Loch Creran Scotland [ECASA]. Scottish Association for Marine Science.
Wu, X., Hou, L., Lin, X., & Xie, Z. 2019. Application of novel nanomaterials for chemo-and biosensing of algal toxins in shellfish and water. Novel Nanomaterials for Biomedical, Environmental and Energy Applications, 353-414. Elsevier.
Last updated:







