NatureScot Research Report 1213 - Exploring biological trait analysis and inter-habitat linkages in three seabed habitat PMFs in Scotland
Year of publication: 2023
Authors: James, K., Kamphausen, L., Cunningham, S. & Kent, F.
Cite as: James, K., Kamphausen, L., Cunningham, S. & Kent, F. 2023. Exploring biological trait analysis and inter-habitat linkages in three seabed habitat PMFs in Scotland. NatureScot Research Report No. 1213.
Keywords
Priority Marine Features; habitat linkages; biodiversity; functional traits analysis; Biological Trait Analysis
Background
To date, marine conservation work has focused predominantly on assessing the status of single species or habitats. This approach has obvious limitations, ignoring the interactions between species and habitats which create ecosystems as a whole. While ecosystem functioning has been identified as a more important and more useful indicator of the status of the marine environment, it remains very difficult to quantify and assess.
By exploring the effects of linkages between a small number of Scottish Priority Marine Feature (PMF) habitats this study aimed to explore methodologies which might assist in more systems-based nature conservation work.
Interactions exist between organisms over many trophic levels. Interactions can occur directly, for example, through predation or competition, or indirectly, for example, where the modification of the environment by one species favours the presence of another (Yakovis & Artemieva, 2017; Passarelli et al., 2014; Trussel et al., 2006) or when species move between habitats (for example, the movement of fish between habitats such as kelp forests and adjoining sedimentary habitats). Linkages between habitats can promote biodiversity (Angelini et al., 2015; Tews et al., 2014), enhance ecosystem services (e.g. increase sediment stability (Alteri et al., 2007)) and support fisheries (Jones, 2010; Saintilan et al., 2007).
Understanding if and how habitats are linked is a crucial step in marine management, especially if trying to apply an ecosystem approach, as practices for one habitat may have cascading effects for the whole ecosystem (Bagur et al., 2016; Donadi et al., 2013). To date, research regarding marine inter-habitat linkages has primarily focussed on the tropics (Jones, 2010; Gribben et al., 2009; Saintilan et al., 2007) and there is little information regarding linkages in temperate and Scottish waters.
PMF habitats such as flame shell beds, maerl beds and horse mussel beds are considered to be some of the most biodiverse habitats in the north-east Atlantic (Trigg et al., 2011; Barbera et al., 2003; Farinas-Franco et al., 2014). These habitats can occur together (i.e. ‘linked’), either adjacent to one another (horizontal linkages) or on top of each other (vertical linkages). The implications of such linkages on overall biodiversity are, however, unknown.
The current study progresses research in this area by investigating inter-habitat linkages between maerl beds (formed by Phymatolithon calcareum or Lithothamnion glaciale), flame shell beds (Limaria hians) and horse mussel beds (Modiolus modiolus). Infaunal seabed samples from across Scotland were used to explore the effects of linkages between maerl beds and horse mussel beds, and flame shell beds and horse mussel beds.
Main findings
- This study aimed to investigate the effects of linkages between selected Priority Marine Feature (PMF) habitats in Scotland on their associated biological communities, and trialled a number of different approaches to assess these.
- The study established a useful process to assess linkages between habitats using biodiversity indices and biological trait analysis (BTA).
- In agreement with previous studies (Donadi et al., 2015; Bremner et al., 2006), BTA was found to be an effective way to investigate differences in functional diversity between locations from an ecosystem rather than a species perspective. In contrast, NMDS plots, SIMPER analyses and ANOSIM analyses did not examine differences between locations at an ecosystem level, with reported differences between sites caused by a few dominant taxa.
- Therefore, it is recommended that future studies use BTA to investigate the effects of linkages between habitats to provide a broader ecosystem perspective. The approach is also recommended for general monitoring and assessment. In addition to better reflecting changes in ecosystem functioning, it also greatly facilitates the interpretation of complex datasets and presents results in a format more accessible to external stakeholders or indeed anybody who does not have a good working knowledge of marine infaunal species communities.
- The results of the BTA identified that examples of some habitats used in this study were likely subject to unquantified levels of disturbance (of natural and / or anthropogenic origin) which may have had a role in shaping the benthic communities in these locations and their functional diversity. BTA may thus also be a useful method to assess the effects of pressures on habitats.
- With limited resources, marine management often requires prioritisation of features and sites for monitoring and protection. While results concerning particular effects of linkages between specific PMFs are preliminary and require further research (limited largely by the data available for the study), the results presented here from local scales suggest that habitat linkages are important in terms of increased species and functional diversity. Diverse habitats are recognised for the variety of ecosystem services they provide and for being more resilient to pressures such as climate change.
- Functional diversity appeared to be influenced strongly by environmental factors, and when attempting to compare habitats on a national scale the relatively subtle differences caused by the presence or absence of habitat linkages could not be detected using the samples assessed in this study. Future monitoring would benefit from inclusion of physical factors (e.g. exposure, seabed water temperature, light levels, etc.) and a better understanding of human activities alongside biological sampling, to enable differences in biodiversity in response to linked habitats to be disentangled from man-made pressures and disturbance or environmental factors. Using only samples collected by similar methodologies would further strengthen the power of the analyses.
- Further work is needed to determine if other linked PMFs exhibit similar patterns in terms of elevated diversity and differing community structure. Follow-up studies are needed to explore whether the effects of inter-habitat linkages at the local scale found in this study hold across other locations and habitats. If this is the case it would have important implications for prioritising inter-linked habitats in marine management in the future.
Introduction
1.1 Inter-habitat linkages
Inter-habitat linkages can exist both horizontally and vertically in the marine environment. Horizontal linkages can occur between habitats that exist next to each other. The presence of a habitat may change the surrounding environment enabling another habitat to form. Mobile species moving between adjacent habitats can also provide horizontal linkages. Vertical linkages occur when habitats co-exist in exactly the same place for example as layers or a mixed habitat. This could be where one habitat provides settlement area for another, or where epifaunal communities influence the associated infauna.
1.1.1 Horizontal linkages
A large proportion of existing research has focussed on linkages between tropical habitats such as mangroves, salt marshes, seagrass meadows, and coral reefs, and the movement of fish species amongst these (Jones et al., 2010; Saintilan et al., 2007). Less work has been conducted on temperate benthic habitats, which can also influence each other. The presence of one habitat may modify the surrounding physical environment and facilitate the presence of another habitat close by. For example, the recruitment and development of a cockle bed can be increased by a nearby mussel bed (Donadi et al., 2013). Mussel beds are ecosystem engineers and significantly alter the physical environment; as the mussel bed forms its surface becomes uneven, which causes a reduction in wave energy. By altering hydrodynamics, the mussel bed facilitates the settlement of cockles nearby. This linkage has a significant effect on the surrounding benthos and community structure (Donadi et al., 2013). As ecosystem engineers alter many physical properties in the surrounding environment, such as sediment stability and nutrient availability (Passarelli et al., 2014), horizontal linkages may exert effects over significant distances.
1.1.2 Vertical linkages
Vertical linkages between habitats can occur when two or more habitats co-exist in the same space. This can increase niche diversity and result in enhanced biodiversity (Petrosillo & Zurlini, 2016; Donadi et al., 2015). Vertical linkages can occur from a habitat (or facilitation) cascade, where the presence of one modifies and facilitates the presence of another (Yakovis & Artemieva, 2017; Angelini et al., 2015; Passarelli et al., 2014). For example, cordgrass (Spartina sp.) facilitates the establishment of ribbed mussels (Geukensia demissa) as the roots and rhizomes of the cordgrass stabilise the sediment (Altieri et al., 2007). The mussels then further modify the environment by filtering water and providing a hard substrate for other organisms to settle (Altieri et al., 2007).
Vertical linkages can also occur across the sediment-water interface (SWI; Austen et al., 2002), between pelagic and benthic systems (Lancaster et al., 2014), and between infaunal and epifaunal communities (Reiss et al., 2009). For example, the physical transport of particles and nutrients across the SWI is strongly influenced by the structure of the benthic boundary layer. Therefore the presence of a structurally complex benthic habitat (such as mussel beds) will alter nutrient cycling across the SWI, influencing the infaunal species found within this habitat (Austen et al., 2002). The presence of infaunal ecosystem engineers can also affect properties of the underlying sediments. For example, the presence of lugworms or cockles within a mussel bed can reduce the amount of organic matter in the underlying sediment (Donadi et al., 2015).
1.1.3 The effects of inter-habitat linkages
Inter-habitat linkages can have substantial effects on biodiversity, community structure and functioning of an ecosystem (Bagur et al., 2016; Donadi et al., 2013), which makes them an important element to consider in marine management. For example, Bagur et al. (2016) found that the presence of two structurally different ecosystem engineers - intertidal mussels, Perumytilus purpuratus, and rock boring bivalves, Lithophaga patagonica - increased overall species richness of the intertidal landscape.
On a local scale, inter-habitat linkages can aid the recovery of damaged habitats. For example, in Scotland, the presence of horse mussel beds can contribute to the stability and recovery of a flame shell bed (Trigg & Moore, 2009). As both habitat-forming species produce byssal threads, the presence of both bivalves increases stability of the bed, promoting recovery after disturbance.
On a larger scale, habitat linkages of the type where one habitat facilitates the presence of other habitats generally increase functional heterogeneity of the ecosystem through the presence of multiple different habitats (Petrosillo & Zurlini, 2016; Donadi et al., 2015; Donadi et al., 2013; Tews et al., 2014). For example, within the Wadden Sea the co-existence of linked cockle, lugworm and blue mussel habitats increases habitat heterogeneity and niche availability as each species modifies the physical environment in a unique way (Donadi et al., 2015).
The presence of an ecosystem engineer may also influence adjacent habitats through behavioural interactions which can shape community structure and biodiversity (Gribben et al., 2009). For example, by moving between mangroves and seagrass meadows depending on the lunar cycle, fish can take advantage of high abundances of zooplankton in mangroves when the tide is high (Saintilan et al., 2007). Migrating fish may also, through excretion and defecation for example, transfer essential nutrients and minerals from adjacent ecosystems (Lundberg & Moberg 2003). Such functional linkages are considered important components of ecosystem resilience as increased biodiversity often increases the resistance (Isbell et al., 2015) and resilience (Oliver et al., 2015) of a system. Thus habitat linkages also play a role in maintaining healthy ecosystems into the future (Austen et al., 2002) and may act as a buffer in response to climate stress for example (Bulleri et al., 2018).
1.2 The seabed habitat PMFs considered in this study
Linkages are thought to exist between a number of PMF habitats (see Table 1 for examples and mechanisms). This study focuses on exploring the linkages between maerl beds and horse mussel beds, and between horse mussel beds and flame shell beds, as a pilot to establish a framework for further research. This section describes the PMFs selected for the study and the nature of some of the connections in habitat examples that have been surveyed is set out in Table 1.
| Linkage | Description | Functional significance | References |
|---|---|---|---|
| Flame shell beds - Horse mussel beds | Horse mussel beds are sometimes found adjacent to or mixed with flame shell beds. Examples include Loch Alsh, Loch Carron and Loch Creran, where mixed flame shell and horse mussel bed exist, or Loch Linnhe and Loch Leven, where flame shell beds are found close to horse mussel beds. | When the byssal threads of flame shell beds and horse mussel beds intertwine, both PMFs are stabilised. Horse mussels provide a hard substrate for flame shell larvae to settle and increased structural complexity alters hydrodynamics and nutrient cycling across the surface-water interface, influencing the infaunal species found in the sediment. | Connor et al., 2004 Moore et al., 2012 Lancaster et al., 2014 Austen et al., 2002 Trigg & Moore 2009 Moore et al., 2018 Moore et al., 2020 |
| Flame shell beds - Maerl beds | Nests of the flame shell are sometimes found in conjunction with maerl beds. Examples of this linkage can be seen e.g. in the Strome Narrows in the Loch Carron MPA. | Flame shells bind maerl together with their byssal threads, helping to stabilise the maerl bed. | Birkett et al., 1998 Hall-Spencer et al., 2003 Lancaster et al., 2014 |
| Flame shell beds - Tide-swept algal communities | Flame shell beds often support tide-swept algal communities. | Flame shell beds provide a stable substratum for the attachment of seaweeds in an otherwise unsuitable location. | Minchin, 1995 Lancaster et al., 2014 |
| Flame shell beds – Kelp forests | Flame shell beds and kelp forests can be found in close proximity of each other, such as in e.g. the Loch Carron MPA. | The community structure of flame shell beds can be influenced by the presence of a nearby kelp forest. | Trigg et al., 2011 Moore et al., 2018 |
| Horse mussel beds - Maerl beds | Horse mussel and maerl beds can co-exist in a mosaic. Examples of this are within the Fetlar to Haroldswick MPA in Shetland. | Horse mussels bind maerl together with their byssal threads, helping to stabilise the maerl bed. | Birkett et al., 1998 Hirst et al., 2013 Lancaster et al., 2014 |
| Maerl beds - Maerl or coarse shell gravel with burrowing sea cucumbers | Areas of predominantly dead maerl and other coarse gravels with burrowing sea cucumbers often occur on the edge of maerl beds. | This linkage occurs when burrowing cucumbers utilise habitat within and at the edge of a maerl bed. | Lancaster et al., 2014 |
1.2.1 Maerl beds
Maerl beds are formed when individual pieces of free-living coralline algae come together to form a structurally and functionally complex 3D habitat (Riosmena-Rodríguez, 2017; Passarelli et al., 2014; Barbera et al., 2003; Jackson et al., 2004). As maerl beds offer a multitude of niches (for example, on the thalli of the coralline algae or within crevices in the matrix) they support a large number of infaunal and epifaunal invertebrates (Jackson et al., 2004). Maerl beds are also important for fisheries, providing nurseries for commercially important fish and shellfish (Kamenos et al., 2004; Hall-Spencer et al., 2003).
Maerl beds can be significantly affected by physical disturbance and activities such as demersal fishing can break up the brittle maerl thalli or bury them (removing access to light), which reduces the structural heterogeneity and biodiversity of the bed (Kamenos, 2003). As coralline algae are slow-growing, with thalli growing less than 1 mm per year (Riosmena-Rodríguez, 2017), the effects of demersal fishing can be detrimental and irreversible (Riosmena-Rodríguez, 2017; Kamenos, 2003). However, dead maerl beds with intact thalli still provide a habitat to multiple species and are considered an important habitat (Passarelli et al., 2014; Jackson, 2004). Maerl beds are predicted to decline significantly in future due to the effects of climate change under all IPCC Representative Concentration Pathways (ranging from 38% decline under RCP 2.6 up to 84% decline under RCP 8.5), with the most rapid rate of decline up to 2050 (Simon-Nutbrown et al., 2020).
1.2.2 Flame shell beds
Flame shells make byssus tubes or nests to live within and these can be connected to create a woven turf - a ‘bed’ of flame shell nests which is structurally and functionally complex, supporting an array of infaunal and epifaunal species (Moore et al., 2018; Tyler-Walters et al., 2016; Hall-Spencer & Moore, 2000a). Flame shells are considered to be ecosystem engineers, as their nests stabilise the sediment by incorporating gravel, shells and maerl with their byssal threads, whilst also providing a substrate for sessile organisms to attach to (Tyler-Walters et al., 2016; Hall-Spencer et al., 2000). Community structure can vary both spatially between beds and temporally within a bed, partially due to an influence of nearby habitats such as rocky reefs (Trigg et al., 2011).
Flame shell beds are sensitive to physical disturbance, including that caused by towed, bottom-contacting fishing activity, with beds in the Clyde and Loch Carron having been destroyed or damaged by scallop dredging (Moore et al., 2018; Hall-Spencer & Moore, 2000b). Once damaged, flame shell beds can take decades to recover (Trigg & Moore, 2009). The presence of other ecosystem engineers that can also produce byssal threads (such as Modiolus modiolus), can contribute to the stability and recovery of a flame shell bed (Trigg & Moore, 2009).
1.2.3 Horse mussel beds
When individual horse mussels aggregate, they can form structurally complex biogenic reefs that harbour a high number of species (Farinas-Franco et al., 2018). Horse mussel beds are formed from live mussels, shell fragments and pseudo-faeces that are bound together by strong byssal threads (Farinas-Franco et al., 2018). As beds are structurally complex, increasing niche availability, they harbour a high biodiversity of both infaunal and epifaunal species (Farinas-Franco et al., 2018; Farinas-Franco and Roberts, 2014; Sanderson et al., 2008) as well as supporting commercially important fish and shellfish (Kent et al., 2017).
As with maerl beds and flame shell beds, horse mussel beds are also sensitive to physical impact (Cook et al., 2013). Furthermore, a loss of horse mussel bed habitat extent is expected as a result of climate change (Gormley et al., 2013)
1.3 Background to techniques used to investigate linkages
This study aimed to investigate inter-habitat linkage effects on community structure, biodiversity, and the functional diversity of the specified PMFs.
Methods used to explore community structure are well developed, with non-metric multidimensional scaling and ANOSIM analysis used extensively in the literature (Pacheco & Vasconcelos, 2012; Trigg, 2011; Pister, 2009; Ellingsen & Gray, 2002). Similarly, biodiversity measurements such as the Shannon-Wiener diversity index, Pielou’s evenness and species richness are widely used in ecology to investigate how rich and even a habitat’s species composition is.
On the other hand, techniques used to investigate functional diversity are still relativity novel with multiple studies suggesting a number of different approaches (Paganelli et al., 2011; Schleuter et al., 2010; Bremner et al., 2006; Usseglio-Polatera et al., 2000). In 2010, Schleuter et al. (2010) published a review that summarised different functional diversity indices that could be used in ecology to measure functional richness and evenness. However, for their calculations, functional traits had to be numerical variables (such as the amount of organic carbon in a leaf, or the size of a plant), while categorical variables (such as feeding type) could not be used in the calculations (Schleuter et al., 2010). As a lot of functional traits in ecology are categorical (feeding type, bioturbation, substrate type), the usefulness of these calculations to measure functional diversity is limited.
Biological trait analysis (BTA) is a different technique that can be used to measure functional diversity in marine benthic assemblages (Bremner et al., 2006). BTA considers a number of behavioural, morphological, and life history traits to describe the ecological functioning of taxa (Bremner et al., 2006). An advantage of BTA is that when assigning trait values, a ‘fuzzy coding’ procedure is used which allows inter-specific variation in traits to be accounted for (Bremner et al., 2006). For example, if a species is both a deposit feeder and a suspension feeder, fuzzy coding allows for the classification of both deposit feeder and suspension feeder for that species. Furthermore, BTA considers both categorical and discrete functional traits, with discrete variables transformed to categorical data. For example, body size may be split into small, medium and large categories (Donadi et al., 2015). As well as investigating inter-habitat linkages (Donadi et al., 2015), BTA analysis can be used to assign biotopes to habitats (Demars et al., 2011), and for bio-monitoring to assess the effects of difference pressures on communities (Paganelli et al., 2011; Bremner et al., 2006; Usseglio-Polatera et al., 2000).
Given the greater flexibility and biological relevance of BTA mentioned above, it was considered that BTA should be used to investigate effects on functional diversity of inter-habitat linkages in Scotland. Habitats were considered to be linked when they were adjacent to one another, or co-existed in the same space.
1.4 Aims and hypothesis
This study aimed to investigate the effects of habitat linkages at a localised scale (using samples <5 miles apart) and compare linkage effects through reference to linked and non-linked examples of the different habitats at a ‘national’ scale (i.e. study areas >100 miles apart). Specifically the work set out to:
- Investigate if linked habitats have different biological communities to non-linked (independent) habitats.
- Investigate if Shannon-Wiener diversity, Pielou’s evenness, species richness, and abundance are higher for linked or non-linked habitats.
- To investigate if inter-habitat linkages affect functional diversity.
2. Methods
2.1 Sample collection
Quantitative infaunal samples from a small subset of surveys were used in this proof-of-concept study. Samples were collected between 2011 and 2015 from the west coast of Scotland (Loch Alsh), Outer Hebrides (Sound of Barra), Orkney (Wyre Sound, Eday and Veantrow Bay) and Shetland (Fetlar to Haroldswick) (see Appendix 4: Sample details, and Mercer et al., 2018; Hirst et al., 2013; Moore et al., 2013; Hirst et al., 2012b).
For all surveys, infaunal samples were passed through a 1 mm mesh sieve and identified to the finest possible taxonomic level. At the Shetland and the Orkney stations, infaunal data came from 4 replicate cores per sampling station. The cores were 20 cm (length) by 5 cm (diameter) giving a volume of approximately 1.6 l. Within Loch Alsh, infaunal data from the linked flame shell and horse mussel beds and the eastern flame shell bed came from cores of the same dimensions. For the Sound of Barra maerl beds, as well as the western flame shell bed in Loch Alsh, infaunal data came from single grab samples with a volume of 15 l each. Grab samples were thus roughly 9 times larger in volume than core samples, which makes direct comparison of some community statistics impossible. This study examined whether there are nevertheless metrics which can be used to compare grab and core data. To do this, infaunal species records from each grab sample were divided by 9 before they were used in data analysis when compared to core samples.
Each grab and core was treated as a separate sample in these analyses.
2.2 Study sites and feature comparisons
2.2.1 Flame shell and horse mussel bed communities: local comparisons in Loch Alsh
Vertical linkages between intermixed flame shell beds and horse mussel beds were investigated within the Lochs Duich, Long and Alsh MPA (Moore et al., 2013). Independent flame shell beds are present on either side of the intermixed bed at the mouth of Loch Alsh (termed eastern and western flame shell bed respectively - see Figure 1).

Figure 1: Map showing flame shell (red points) and mixed flame shell and horse mussel bed (green points) infaunal sampling stations at the mouth of Loch Alsh (part of the Lochs Duich, Long and Alsh MPA).
Click for a full description
Map Loch Alsh (part of the Lochs Duich, Long and Alsh MPA). Showing infaunal sampling stations delineated as points:
W FS = western flame shell bed (red points)
E FS = eastern flame shell bed (red points)
FS & HM = mixed flame shell and horse mussel bed (green points)
Ordnance Survey © Crown copyright [and database rights] 2021 OS 100017908. GeMS Habitat Data ©Scottish Natural Heritage, licensed under the Open Government Licence. Contains Marine Recorder data, database provided under Open Government License v3. Individual data contained within are subject to data limitations described within the database. Contains information from Scottish Government (Marine Scotland) licensed under the Open Government Licence v3.0. Map © Scottish Natural Heritage.
2.2.2 Maerl bed and horse mussel bed communities: local comparisons in Fetlar to Haroldswick
Horizontal linkages between maerl beds and horse mussel beds were investigated within the Fetlar to Haroldswick MPA in Shetland.
Maerl beds and horse mussel beds are considered to be linked horizontally in this location as they co-exist next to one another. The habitat is considered a mosaic with horse mussel beds changing into maerl beds over a small area (e.g. 200 m2) (Hirst et al., 2013). Maerl and horse mussel bed cores from mosaic habitat in Uyea Sound, Hascosay, North Hascosay and Basta Voe were used in this study (see Figure 2).
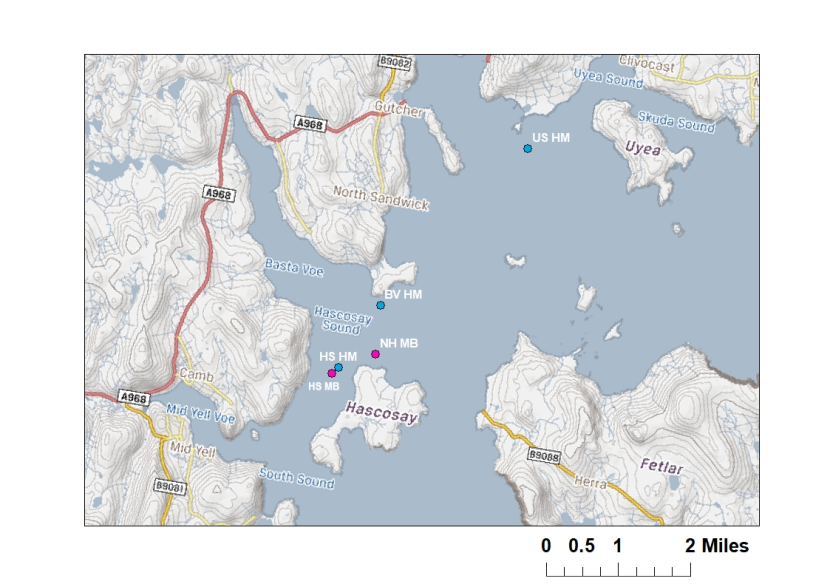
Figure 2: Map showing maerl bed (pink) and horse mussel bed (blue) infaunal sampling stations within the maerl and horse mussel bed mosaic in the Fetlar to Haroldswick MPA.
Click for a full description
Map of the Fetlar to Haroldswick MPA. Showing infaunal sampling stations delineated as points:
HS MB = Hascosay maerl bed (pink points)
NH MB = North Hascosay maerl bed (pink points)
HS HM = Hascosay horse mussel bed (blue points)
BV HM = Basta Voe horse mussel bed (blue points)
US HM = Uyea Sound horse mussel bed (blue points)
Ordnance Survey © Crown copyright [and database rights] 2021 OS 100017908. GeMS Habitat Data ©Scottish Natural Heritage, licensed under the Open Government Licence. Contains Marine Recorder data, database provided under Open Government License v3. Individual data contained within are subject to data limitations described within the database. Contains information from Scottish Government (Marine Scotland) licensed under the Open Government Licence v3.0. Map © Scottish Natural Heritage.
2.3 Maerl bed communities: national comparison
On a national scale (Figure 3), the maerl beds linked with horse mussel beds in Fetlar to Haroldswick MPA were compared to non-linked maerl beds in Orkney (Figure 4) and the Sound of Barra SCI (Figure 5) with regards to their community structure. Maerl beds in Orkney and the Sound of Barra were selected as both beds are ‘wave formed’ rather than ‘tidally-formed’ so have a similar structure to those in Fetlar to Haroldswick.
Functional diversity was also compared on a local and national level; however, to simplify the analysis, infaunal data from the maerl beds in Fetlar to Haroldswick were only compared with infaunal data from grabs in Barra, and infaunal data from cores in Wyre Sound, Orkney. Wyre Sound was selected as a site because it had the most similar degree of exposure to Barra and Fetlar to Haroldswick.
As sample type (core vs grab) was found to have a large effect on species diversity measurements, the effects of linkages on biodiversity were not explored for the national scale maerl bed analysis.
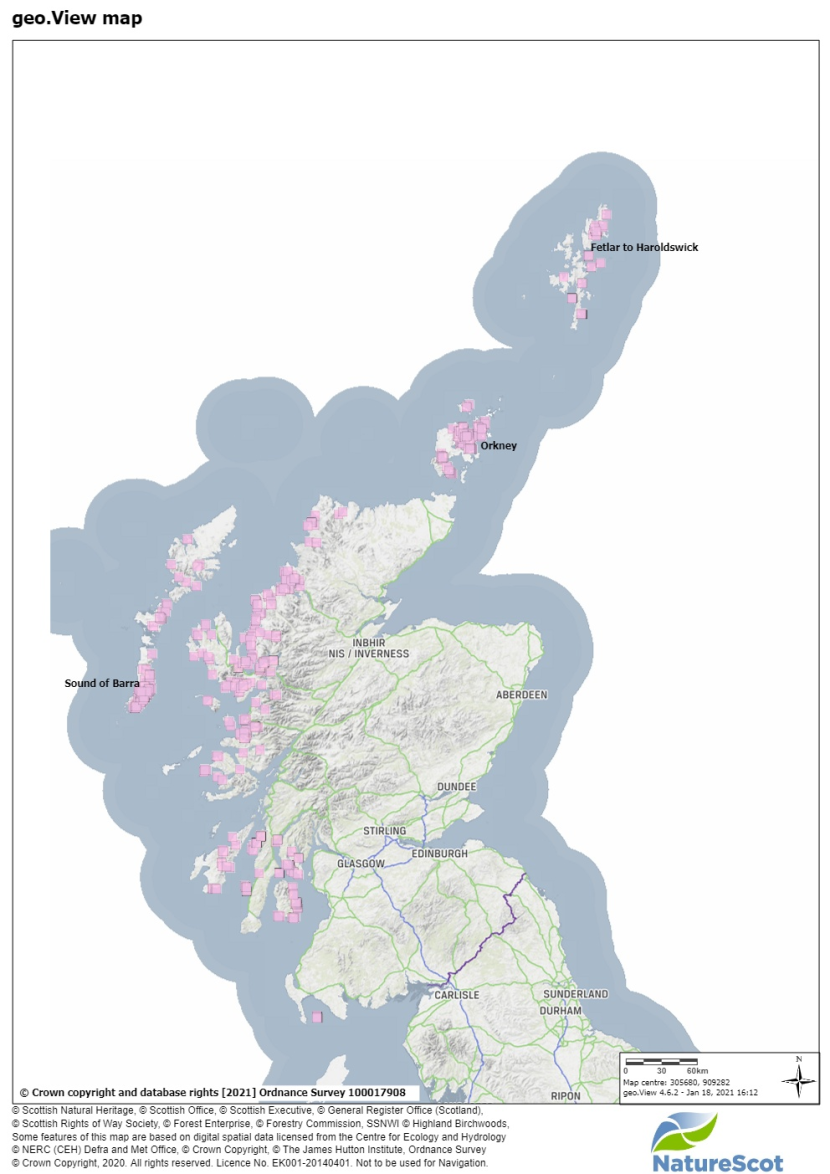
Figure 3: Map of Scotland. Showing known records of maerl beds (pink squares) and general locations of study areas.
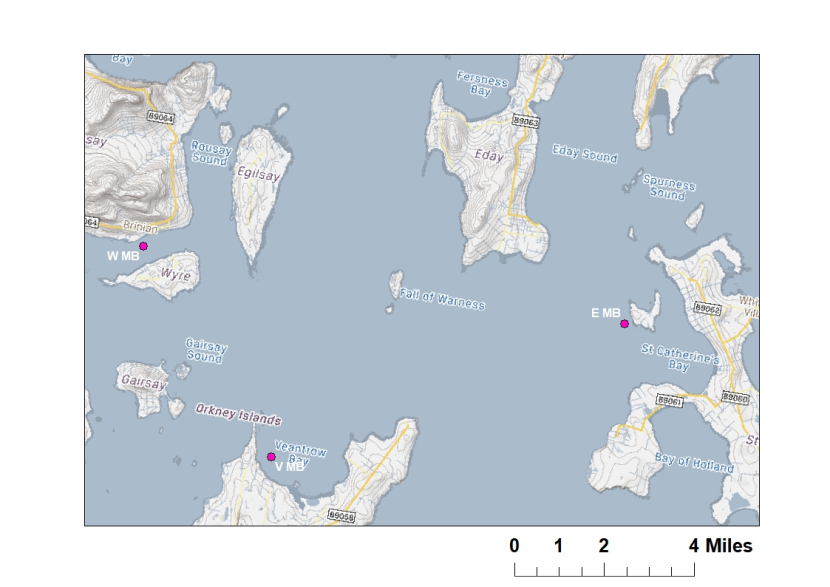
Figure 4: Map showing independent maerl bed sampling locations (pink points) in Orkney waters.
Click for a full description
Stations are Wyre maerl bed (W MB), Eday maerl bed (E MB), and Veantrow Bay maerl bed (V MB). Ordnance Survey © Crown copyright [and database rights] 2021 OS 100017908. GeMS Habitat Data ©Scottish Natural Heritage, licensed under the Open Government Licence. Contains Marine Recorder data, database provided under Open Government License v3. Individual data contained within are subject to data limitations described within the database. Contains information from Scottish Government (Marine Scotland) licensed under the Open Government Licence v3.0. Map © Scottish Natural Heritage.
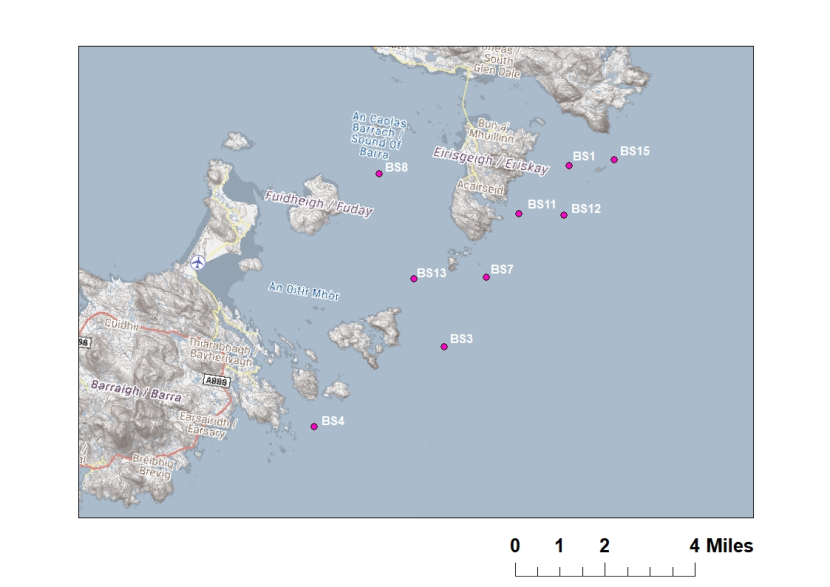
Figure 5: Map showing independent maerl bed sampling locations (pink points) in the Sound off Barra (BS).
Click for a full description
Maerl beds sampling points in the Sound of Barra (BS) are represented by pink circles. Ordnance Survey © Crown copyright [and database rights] 2021 OS 100017908. GeMS Habitat Data ©Scottish Natural Heritage, licensed under the Open Government Licence. Contains Marine Recorder data, database provided under Open Government License v3. Individual data contained within are subject to data limitations described within the database. Contains information from Scottish Government (Marine Scotland) licensed under the Open Government Licence v3.0. Map © Scottish Natural Heritage.
2.4 Data analysis
2.4.1 Community structure (aim 1)
All analysis was carried out in R Studio (R Core Team, 2019). As a first step, non-metric multidimensional scaling (NMDS) was used to investigate if samples showed differences in community structure, which might be caused by habitat linkages. NMDS uses Bray-Curtis dissimilarity to measure how similar sites are. Sites are then plotted in an NMDS plot which places sites that have a similar community structure closer together. NMDS plots were created using the ‘vegan’ package in R (Oksanen et al., 2019).
Initial data exploration also attempted using cluster analysis (‘fpc’ package; Hennig, 2020) to illustrate which sites were similar to each other. However, ANOSIM analyses showed that differences within clusters were often as large as differences between clusters, potentially because of high numbers of 1s and 0s within the dataset, or because there were more variables (species) than samples. Cluster analysis was therefore not an appropriate technique to be used to analyse similarities between samples.
To investigate differences in community structure via NMDS plots, a minimum species matrix was used. This matrix excluded taxa that were not identified to species level (e.g. nematodes) and combined records of the same species (e.g. juvenile and adult records). A minimum species matrix was used so that differences in community structure picked up by the analyses would be due to the species present, rather than due to larger variations in phylum or age structure. As Bray-Curtis dissimilarity uses the abundance of species to measure dissimilarity, species data from Sound of Barra and species data from the western flame shell bed at Loch Alsh (grab samples) were divided by 9. This was because the sample volume of the grabs collected at these sites was 9 times larger than that of the cores from Orkney, Fetlar to Haroldswick and the other sites in Loch Alsh. This volume-controlled minimum species matrix was used for the rest of the community structure and species diversity analysis.
Following the creation of NMDS plots, a non-parametric two-way analysis of variance (ANOSIM; Oksanen et al., 2019) was used to quantify the differences between bed types, locations, latitudes and depth. A similarity percentages analysis (SIMPER; Oksanen et al., 2019) was then run for each comparison to determine which 10 species accounted for the largest difference between bed type, location, latitude and depths. As sites in Loch Alsh were at the same latitude, and in a similar depth range, ANOSIM and SIMPER analyses were not used to investigate differences between latitude and depth. NMDS plots, ANOSIM and SIMPER analyses were performed both on log transformed and non-transformed data. In all cases, transforming the data did not change the outputs, indicating that the results were not skewed by dominant species. The report therefore only presents the analyses performed on non-transform the data, which show the true differences between samples.
The effects of inter-habitat linkages on community structure were investigated on a local scale in Loch Alsh and Fetlar to Haroldswick MPA, and on a national scale between maerl beds from Fetlar to Haroldswick, Wyre Sound (Orkney) and the Sound of Barra.
2.4.2 Species diversity (aim 2)
To investigate if inter-habitat linkages influence species diversity, the Shannon diversity index, Pielou’s evenness, species richness and abundance were calculated for each sample (vegan package; Oksanen et al., 2019). For this analysis, samples were only compared with other samples on a local scale.
Linear models were then used to investigate if the diversity indices varied between locations and bed type. Individual indices were calculated from non-transformed raw data. In general, the index-dataset was normally distributed and did not need to be transformed before using a linear model (Shapiro-Wilk test, p>0.05). For the Fetlar to Haroldswick local study, the abundance data was not normally distributed (Shapiro-Wilk test, p<0.05), and was log transformed before using a linear model. Likelihood ratio tests (using the ‘lmtest’ package; Zeileis and Hothorn, 2002), were used to test the statistical significance of the impact of location and bed type on diversity indices.
As grab samples are 9 times larger in volume than core samples, the grabs consistently had a higher Shannon Diversity Index and Species Richness Index. The differences in diversity could not be controlled for by dividing the infaunal abundance data from the grab samples by 9. Therefore, the effects of inter-habitat linkages on species diversity on a national scale (which would have required comparing grab samples to core samples) was not considered further in this study.
2.4.3 Functional diversity - Biological Trait Analysis (aim 3)
Creating a dataset for BTA analysis
Biological trait analysis (BTA) was used to investigate differences in functional diversity between beds that were independent and beds that were linked. Following recommendations by Bremner et al. (2006), species were only included in BTA if they were present in 50 % of the sites or the top 90 % of abundance. For the Fetlar to Haroldswick analysis, species were only included if they were in the top 75 % of abundance, as SIMPER analysis showed that the sites were more dominated by few taxa such as Jasmineira elegans, compared to sites at Loch Alsh. Taxa that were identified only to family or phylum level were included if species within the taxonomic category were known to have similar functional traits.
Biological traits where chosen to reflect lifestyle and behaviour characteristics of the infaunal taxa (Paganelli et al., 2012). Following Donadi et al. (2015), four trait categories - bioturbation, size, longevity, and feeding mode - were selected (Table 2). Bioturbation was chosen as a behavioural trait and refers to the movement of sediment by organisms which plays an important role in many geochemical processes including carbon burial (Queiros et al., 2013). Feeding mode was also selected as a behavioural trait, with feeding type indicating, for example, how stressed an environment is and the sediment type (Paganelli et al., 2012). Size and longevity were chosen as lifestyle characteristics, with each trait able to indicate if a site is disturbed. A fifth category, substratum affinity, was included in this analysis as a proxy to investigate changes in the substrate between the beds (Table 2; Paganelli et al., 2012). Each trait category was separated into several modalities, for example, bioturbation was split into biodiffuser, upward-conveyor, downward-conveyor, bio-irrigator and surficial modifier (Queiros et al., 2013).
| Trait | Category | Abbreviation |
|---|---|---|
| Bioturbation | None | none |
| Bioturbation | Biodiffuser | biodiff |
| Bioturbation | Upward-conveyor | upw |
| Bioturbation | Downward-conveyor | down |
| Bioturbation | Bio-irrigator | bioirr |
| Bioturbation | Surface modifier | surf_mod |
| Size | <5mm | s_5 |
| Size | 5-10mm | s5_10 |
| Size | 10-20mm | s10_20 |
| Size | 20-40mm | s20_40 |
| Size | 40-80mm | s40_80 |
| Size | >80mm | s80 |
| Feeding mode | Deposit feeder | dep |
| Feeding mode | Suspension feeder | susp |
| Feeding mode | Detritus feeder | detritus |
| Feeding mode | Opportunist/Scavenger | opp |
| Feeding mode | Grazer | graz |
| Feeding mode | Predator | pred |
| Longevity | <1 year | <1yr |
| Longevity | 1-2 years | 1_2y |
| Longevity | 3-5 years | 3_5y |
| Longevity | 6-10 years | 6_10y |
| Longevity | >10 years | >10yr |
| Substratum affinity | Rock | rock |
| Substratum affinity | Gravel | gravel |
| Substratum affinity | Coarse Sand | coarse sand |
| Substratum affinity | Sand | sand |
| Substratum affinity | Muddy Sand | muddy sand |
| Substratum affinity | Mud | mud |
| Substratum affinity | Other species | other_sp |
When carrying out BTA, first a trait-by-taxon matrix is created, where each species is given a code for each trait category. A fuzzy coding procedure was used to assign functional trait codes to each species (Chevenet et al., 1994). The fuzzy coding procedure allows for intra-specific variability in traits, as some taxa may exhibit multiple traits within the same category. Species were scored from 0 to 3 for each trait category, with 0 indicating that the taxa have no affinity for the trait and 3 indicating that the taxa have total affinity for the trait. For example, with regards to bioturbation, the polychaete Aonides paucibranchiata is both an upward-conveyor and a downward-conveyor. Therefore, this species has a code of 1.5 for both the upward-conveyor and downward-conveyor trait, and a 0 for all other bioturbation traits. If no information was available for a species for a specific trait category, then a code of 0 was given for all traits within that category. In some cases, if trait information was unavailable, species were given the same code as other species within the same genus, or family. This only occurred if the species shared the same codes for other trait categories indicating it had a similar function. Trait information was obtained from biological trait databases (WoRMS, Polytraits, BIOTIC), and primary and secondary literature (Appendix 1; Fish and Fish 2002).
Following the creation of the trait-by-taxon matrix, the trait category scores for each taxon were multiplied by the abundance of the taxon in each sample. As before, the abundances for samples from Sound of Barra, and for the western flame shell bed at Loch Alsh were divided by 9 to account for differences in the sample volume. Trait categories were then summed for each sample and added to a sample-by-trait matrix containing total scores for each trait category for each sample. For the Loch Alsh analysis, the matrix included 16 samples and 28 traits. For the Fetlar to Haroldswick analysis, the matrix included 36 samples and 28 traits.
To investigate which traits were dominant in each sample, an additional BTA data-frame was created containing proportion data. In this matrix, each trait value was divided by the total sum of trait values within that category. This resulted in a matrix that had a relative proportion of each trait for each sample. For example, for the western flame shell bed 1, the trait proportion values were: 0.12 (none), 0.23 (bio-diffuser), 0.07 (upward-conveyor), 0.01 (downward-conveyor), 0.02 (upward-conveyor) and 0.55 (surface modifier), giving a total sum of 1.
Maerl bed samples from Wyre Sound were strongly dominated by the caprellid Caprella acanthifera. As this species had a dominant effect on the BTA analysis, even when the dataset was transformed, it was removed from the analysis for the national-scale maerl bed comparisons.
The above procedure resulted in the creation of two datasets for each analysis. The first, a sample-by-trait matrix from abundance data, was used to show how functionally rich sites were. The second, a sample-by-trait matrix from proportion data, was used to show which functional categories were associated with which locations.
Principle component analysis (PCA) and regression
Principle component analysis (PCA) and regression were used to explore differences in functional diversity between sites. PCA analysis was first used to visualise differences in functional diversity between sites with PC1 and PC2 scores corresponding to the x and y-axis (using the ‘factoextra’ package; Kassambara and Mundt, 2019). Following plot visualisation, PC rotation scores were obtained, showing the correlation between PC1 and PC2 values and specific traits. PC rotation scores were plotted using the package ‘corrplot’ (Weio and Simko, 2017). This information was then used to determine which traits were more abundant in certain samples. For example, if PC1 was positively correlated with suspension feeders and short-lived species, and samples from Basta Voe had a positive PC1 value, then this indicated that Basta Voe was characterised by suspension feeders and short-lived species. If the direction of all, or most functional trait vectors pointed away from a sample, then it indicated that the sample was not functionally diverse.
Following PCA visualisation, PCA regression was used to quantify the differences in PC1 and PC2 axis values on location and bed type. Linear models were then run, with PC1 and PC2 axis values as dependent variables and location and bed type as independent variables. Likelihood ratio tests were used to determine if the relationship was significant, and post hoc analysis was used to investigate the specific sample to sample differences.
PCA analysis and regression were run on abundance and proportion sample-by-trait matrices for both the local and national scale analyses.
3. Results
3.1 Local scale comparisons: Loch Alsh study - vertical linkages – flame shell beds and horse mussel beds
3.1.1 Community structure
When analysing differences in community structure between flame shell (FS) beds that were linked with horse mussel (HM) beds, and flame shells beds that were independent, there were distinct locational patterns in community composition (Figure 6). Locational differences in community composition were confirmed by ANOSIM analysis with cores from the same site having a more similar community structure compared to cores from different sites (p<0.001, R2=0.89). There was also a significant difference in bed type with linked beds (FS & HM 1 and 2) having a more similar community structure than the independent beds (W FS and E FS) (p<0.001), although the R2 was much lower for this comparison (R2=0.66). The results are not shown for the logged dataset as transforming the data had no impact on the NMDS plots. The western flame shell bed samples were grabs while all other samples from Loch Alsh were cores. The mean abundance in the western flame shell bed was divided by 9 to account for differences in sample volume (other metrics were calculated off full original sample volume), but nevertheless sample volume and collection techniques may be driving some of the differences seen here.
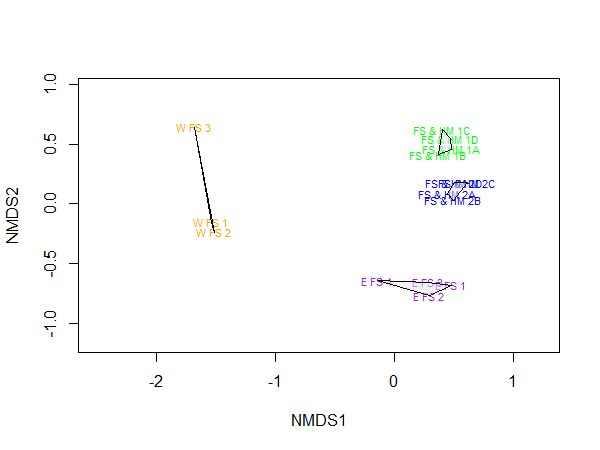
Figure 6: Two-dimensional non-metric multidimensional scaling (NMDS) plot, showing variation in community composition between sites in Loch Alsh.
Click for a full description
Stress level of plot was 0.04 after 20 runs. Polygons represent clustering by location. Sites are eastern FS bed (purple; E FS), western FS bed (orange; W FS), FS and HS bed 1 (green; FS & HM 1), and FS and HS bed 2 (blue; FS & HM 2).
A similarity percentages (SIMPER) analysis on the data identified which species contributed to differences between the flame shell and horse mussel beds and the independent flame shell beds (Tables 3, 4 and 5). Differences between beds were driven by only a few taxa. When comparing the eastern flame shell bed to the linked flame shell / horse mussel beds, the top 10 species contributed to ~30.3 % of the average dissimilarity between the bed types, with the polychaete Jasmineira elegans contributing the greatest amount (7.9 %; Table 3). When comparing the western flame shell bed to the linked flame shell / horse mussel beds, the top 10 species contributed to ~48.3 % of the average dissimilarity between the bed types, with Jasmineira elegans again contributing the greatest amount (15.4 %; Table 4). The two independent flame shell beds (E FS and W FS) were also distinguished most strongly by Jasmineira elegans though less strongly so (3.4 %, Table 5). Horse mussel Modiolus modiolus abundances made the second largest contribution to dissimilarity here (2.8 %, Table 5).
In all three comparisons, dissimilarities were dominated by Polychaeta (Jasmineira elegans, Nereimyra punctata, Pholoe inornata, Aonides oxycephala, Spirobranchus triqueter, and Psamathe fusca) which were more abundant in the linked flame shell / horse mussel bed than in the independent flame shell beds.
| Species | Class | Abundance in E FS | Abundance in FS & HM | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Jasmineira elegans | Polychaeta | 61 | 112 | 7.9 | 7.9 |
| Nereimyra punctata | Polychaeta | 9 | 73 | 7.7 | 15.6 |
| Aonides oxycephala | Polychaeta | 4 | 26 | 2.6 | 18.2 |
| Spirobranchus triqueter | Polychaeta | 0 | 22 | 2.5 | 20.7 |
| Ophiopholis aculeata | Ophiuroidea | 0 | 17 | 2.2 | 22.9 |
| Pholoe inornata | Polychaeta | 28 | 40 | 2.0 | 24.9 |
| Psamathe fusca | Polychaeta | 9 | 21 | 1.5 | 26.4 |
| Janira maculosa | Malacostraca | 1 | 10 | 1.4 | 27.8 |
| Ophiothrix fragilis | Ophiuroidea | 0 | 10 | 1.3 | 29.1 |
| Flabelligera affinis | Polychaeta | 7 | 17 | 1.2 | 30.3 |
| Species | Class | Abundance in W FS | Abundance in FS & HM | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Jasmineira elegans | Polychaeta | 3 | 112 | 15.4 | 15.4 |
| Nereimyra punctata | Polychaeta | 2 | 73 | 10.7 | 26.1 |
| Pholoe inornata | Polychaeta | 10 | 40 | 4.3 | 30.4 |
| Aonides oxycephala | Polychaeta | 1 | 26 | 3.8 | 34.2 |
| Spirobranchus triqueter | Polychaeta | 0 | 22 | 3.1 | 37.3 |
| Psamathe fusca | Polychaeta | 2 | 21 | 2.9 | 40.2 |
| Ophiopholis aculeata | Ophiuroidea | 0 | 17 | 2.8 | 42.0 |
| Eumida sanguinea | Polychaeta | 0 | 16 | 2.2 | 44.2 |
| Flabelligera affinis | Polychaeta | 3 | 17 | 2.1 | 46.3 |
| Aurospio banyulensis | Polychaeta | 0 | 13 | 2.0 | 48.3 |
| Species | Class | Abundance in W FS | Abundance in E FS | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Jasmineira elegans | Polychaeta | 1 | 4 | 3.4 | 3.4 |
Modiolus modiolus/ Modiolula phaseolina | Bivalvia | 0 | 2 | 2.8 | 6.2 |
| Aurospio banyulensis | Polychaeta | 0 | 2 | 2.5 | 8.7 |
| Eumida sanguinea | Polychaeta | 0 | 2 | 1.2 | 9.9 |
| Golfingia (golfingia) elongata | Sipunculidea | 0 | 2 | 1.8 | 11.7 |
| Psamathe fusca | Polychaeta | 1 | 2 | 1.7 | 13.4 |
| Thracia distorta | Bivalvia | 0 | 1 | 1.7 | 15.1 |
| Nereimyra punctata | Polychaeta | 1 | 2 | 1.6 | 16.7 |
| Metaphoxus fultoni | Malacostraca | 0 | 2 | 1.5 | 18.2 |
| Polyophthalmus pictus | Polychaeta | 0 | 1 | 1.5 | 19.7 |
3.1.2 Species diversity
In general, inter-habitat linkages between flame shell beds and horse mussel beds had a positive effect on diversity indices (Table 6 and Table 7). Shannon’s diversity index, species richness and abundance were all significantly higher in the linked flame shell and horse mussel beds compared to the independent flame shell beds (Table 7). Pielou’s evenness did not vary significantly between linked and independent beds, with both bed types being relatively even (p>0.05 for both comparisons).
| Location | Shannon Diversity Index | Pielou's Evenness | Species Richness | Abundance |
|---|---|---|---|---|
| W FS | 2.99 | 0.777 | 54 | 46.3* |
| E FS | 2.92 | 0.774 | 44 | 212 |
| FS & HM 1 | 3.46 | 0.732 | 113 | 728 |
| FS & HM 2 | 3.44 | 0.794 | 77 | 522 |
| - | Bed Type | Bed Type |
|---|---|---|
| Diversity Index | LRT | p-value |
| Shannon Diversity Index | 13.11 | <0.001 |
| Species Richness | 18.64 | <0.001 |
| Species Abundance | 23.65 | <0.001 |
3.1.3 Functional richness - abundance data
Principle Component Analysis suggested that linked flame shell and horse mussel beds were more functionally rich than the independent flame shell beds (Figure 7 and Figure 8). The first two PCA axes explained 97.9% of the total variance in function traits, with the first principal component axis (PC1) accounting for 91.7% of the variance, and the second principle component axis (PC2) accounting for 6.2%. The linked beds and individual flame shell beds were separated along the PC1 axis, which suggests that differences in functional diversity were driven by inter-habitat linkages. Differences in functional diversity due to bed type were confirmed by statistical modelling, with linked beds having a significantly higher PC1 score than independent flame shell beds (LRT: χ(1) =20.2, p<0.001). Sites also had different PC2 values, with FS & HM 2 having a significantly higher PC2 value than all other sites (LRT: χ(3) =20.56, p<0.001; post hoc analysis, p <0.02 for all FS & HM 2 - other site comparisons). Interestingly, samples from individual flame shell beds (E FS and W FS) were more closely clustered than samples from linked flame shell beds (FS & HM 1 and 2), suggesting that functional diversity within the linked flame shell bed is more heterogeneous than within the independent flame shell beds.
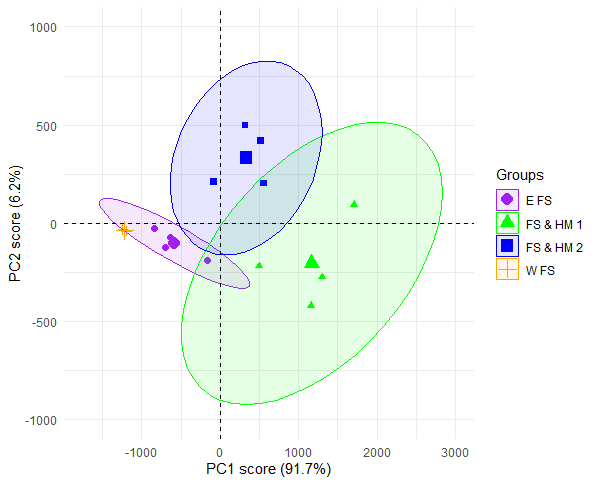
Figure 7: PCA plot comparing functional richness between independent flame shell bed communities and inter-linked flame shell bed and horse mussel beds communities within Loch Alsh. Each point represents an infaunal sample with text coloured by site.
Click for a full description
Position of samples in relation to PC1 and PC2 scores with 95% confidence ellipses. Locations are: Flame shell bed and horse mussel bed 1 (FS & HM 1; green), flame shell bed and horse mussel bed 2 (FS & HM 2; blue), eastern flame shell bed (E FS; purple) and western flame shell bed (W FS; orange). Larger symbols represent the centre of the ellipse for each group.
The direction of functional traits plot PC1 and PC2 rotation scores (Figure 8) show that linked flame shell and horse mussel beds are more functionally rich than the individual flame shell beds. With regards to the PC1 rotation scores, scores were mainly positive. In general, samples from the linked flame shell beds were also positive indicating that traits were found in a higher abundance in these beds. In particular, organisms that were suspension feeders with a short life span (1-2 years) were found in abundance in FS & HM bed 1, and organisms that were deposit feeders and biodiffusers were found in abundance in FS & HM bed 2. Furthermore, whilst the FS & HM 1 bed tended to support organisms that favoured rocky substrate, the FS & HM 2 bed supported organisms that favoured both gravel and muddy sand substrates.
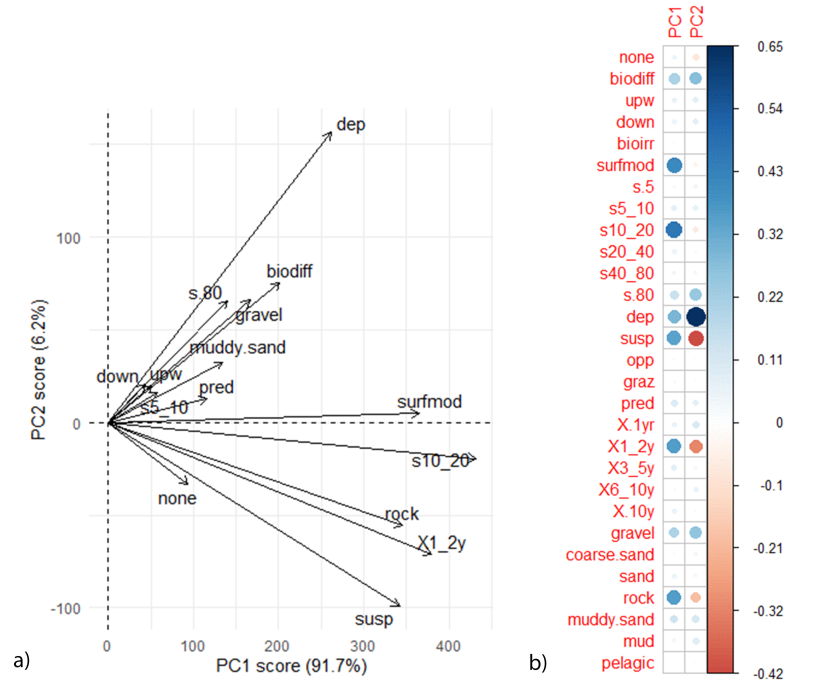
Figure 8: PCA plot showing a) the direction of functional traits in relation to PC1 and PC2 and b) the correlation matrix. Comparisons with Figure 6 show functional traits that result in differences between communities.
Click for a full description
PCA analysis contained both linked flame shell and horse mussel beds and individual flame shell beds. For the correlation matrix, red dots represent a negative correlation and blue dots a positive correlation. Size of dots correspond to the strength of the correlation. For functional trait acronyms refer to the Table 2.
3.1.4 Functional evenness - proportion data
PCA analysis showed that flame beds within Loch Alsh (E FS, FS & HM 1 and FS & HM 2) had distinct functional differences to the bed at the mouth of Loch Alsh (W FS) (Figure 9 and Figure 10). The first two PCA axes explained 80.5% of the total variance in function traits, with the first principle component axis (PC1) accounting for 54.9% of the variance, and the second principal component axis (PC2) accounting for 25.6%. As beds within Loch Alsh and beds at the mouth of Loch Alsh (W FS) were separated along the PC1 axis, differences in functional diversity were driven by locational patterns. Differences in functional diversity due to the location were confirmed by statistical modelling, with beds within Loch Alsh having a significantly higher PC1 score than the bed at the mouth of Loch Alsh (LRT: χ(3) =32.01, p<0.001, post hoc analysis, p <0.001 for all FS & HM 2 - site comparisons). PC2 values were dependent on site, with FS & HM bed 2 having a significantly lower PC2 value than the other beds (LRT: χ(3) =18.4, p<0.001; post hoc analysis, p <0.04 for all FS & HM 2 - site comparisons).

Figure 9: PCA plot comparing functional eveness between independent flame shell bed communities and inter-linked flame shell bed and horse mussel beds communities within Loch Alsh. Each point represents an infaunal sample with text coloured by site.
Click for a full description
Position of samples in relation to PC1 and PC2 scores with 95% confidence ellipses. Locations are: Flame shell bed and horse mussel bed 1 (FS & HM 1; green), flame shell bed and horse mussel bed 2 (FS & HM 2; blue), eastern flame shell bed (E FS; purple) and western flame shell bed (W FS; orange). Larger symbols represent the centre of the ellipse for each group.
The direction of functional traits plot and PC1 and PC2 rotation scores (Figure 10) show that beds within Loch Alsh have a different functional community than the flame shell bed at the mouth of Loch Alsh (W FS). PCA analysis indicated that the flame shell bed at the mouth of Loch Alsh (W FS) is dominated by predators, medium long lived species (3-5 year life span) and muddy sand. The eastern flame shell bed is dominated by surface modifiers and suspension feeders with a small body size (10-20 mm). Linked flame shell and horse mussel beds did not cluster in the PCA plots, with multiple functions correlating with different samples. For example, whilst FS & HM 1 had a higher proportion of short lived species (1-2 year life span) that preferred to inhabit a rocky substrate, FS & HM 2 had a higher proportion of deposit feeders and biodiffusers that were large (body size <80 mm) and short lived (<1 year life span) and preferred to inhabit a gravel substrate. This shows that these sites are functionally heterozygous, with multiple traits found within each site.
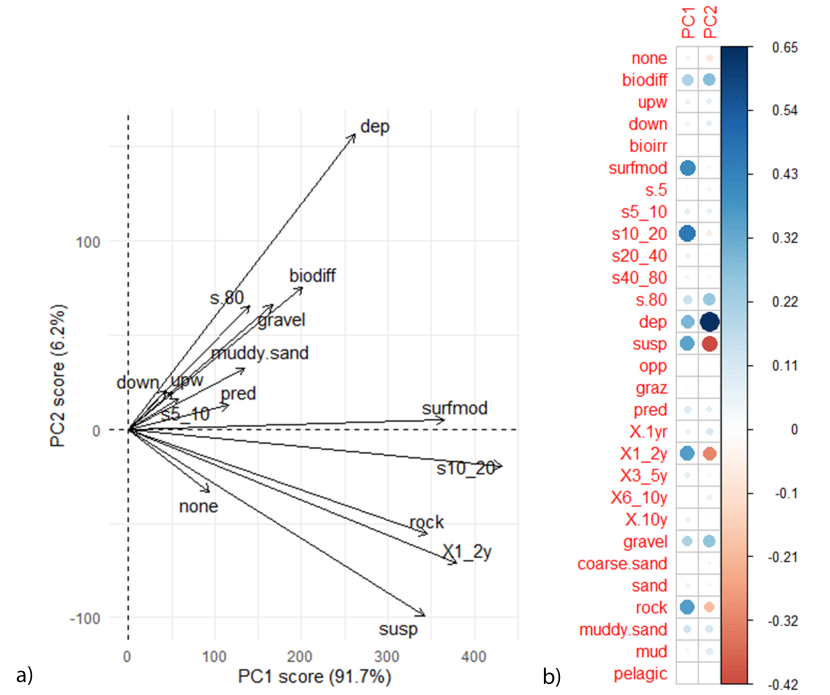
Figure 10: PCA plot showing a) the direction of functional traits in relation to PC1 and PC2 and b) the correlation matrix. Comparisons with Figure 9 show functional traits that result in differences between communities.
Click for a full description
PCA analysis contained both linked flame shell and horse mussel beds and individual flame shell beds. For the correlation matrix, red dots represent a negative correlation and blue dots a positive correlation. Size of dots correspond to the strength of the correlation. For functional trait acronyms refer to the Table 2.
3.2 Local scale comparison: Fetlar to Haroldswick study - horizontal linkages – maerl beds and horse mussel beds
3.2.1 Community structure
As illustrated in the NMDS plots (Figure 11), on a local scale, clustering according to community composition was largely driven by location. Samples grouped into 3 distinct clusters which did not correspond to geographical proximity. Community structure in the linked horse mussel beds in Uyea Sound and Hascosay was similar despite Uyea Sound being at the greatest physically distance from the other samples (cluster 1). The linked horse mussel bed at Basta Voe had a distinctly separate community structure (cluster 2). The linked maerl beds in Hascosay and North Hascosay shared a similar community composition (cluster 3). The community structure of the two Hascosay sites (a horse mussel bed and a maerl bed, HS HM and HS MB) did not overlap. This result suggests that even within the horse mussel maerl bed habitat mosaic community composition is driven by the biogenic bed type, with samples taken from distinct maerl and horse mussel bed areas within the ‘mosaic’ having different communities. Differences in community composition due to location were confirmed by ANOSIM analysis (R2=0.81, p<0.001). ANOSIM analysis also confirmed that difference in community between clusters was statistically significant (R2=0.81, p<0.001).
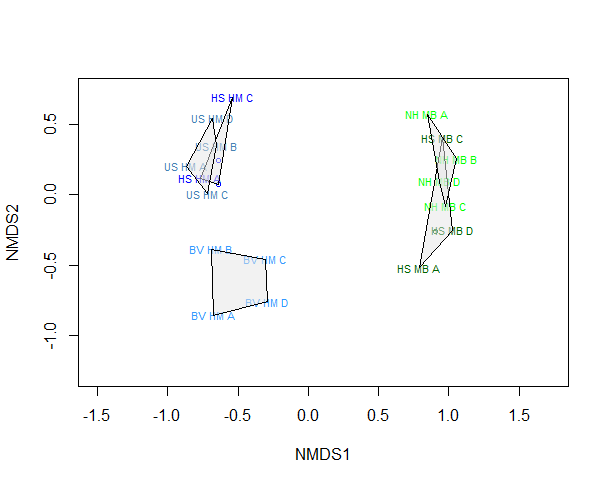
Figure 11: Two-dimensional non-metric multidimensional scaling (NMDS) plot, showing variation in community composition between sites within the Fetlar to Haroldswick MPA in Shetland.
Click for a full description
Stress level of plot was 0.04 after 20 runs. Polygons represent clustering by location. Sites (all from within the habitat mosaic, i.e. linked habitats) are Hascosay horse mussel bed (HS HM), Hascosay maerl bed (HS MB), North Hascosay maerl bed (NH MB), Uyea Sound horse mussel bed (US HM), Basta Voe horse mussel bed (BV HM). Samples are coloured by site, with horse mussel beds blue and maerl beds green.
As the NMDS analysis indicated that there were 3 distinct communities present in Fetlar to Haroldswick (Figure 11), a similarity percentages (SIMPER) analysis was run to identify which species contributed to differences between these communities (Table 8 to Table 10). When comparing differences between the first two clusters, the top 10 species contributed to 60.3% showing that differences between the clusters was driven by a few key species (Table 8 and Figure 8). Species abundant in cluster 1 contributed the greatest amount of dissimilarity between the communities, with Capitella capitata, a polychaete (18.9%), followed by the oligochaete Tubificoides pseudogaster (14.4%). Although species between cluster 1 and cluster 2 differed, similar functional groups appeared to be present in both bed types. For example, polychaetes, which play a role in the bioturbation of sediment, were present in both bed types. The brittle star Ophiopholis aculeata, a detritus feeder, was more abundant in cluster 1.
When comparing differences between cluster 1 and cluster 3, the top 10 species contributed to 65.7% showing that differences between the clusters was driven by a few key species (Table 9). As before, Capitella capitata, a polychaete, contributed the greatest amount of dissimilarity between the communities (17.3%), followed by oligochaete Tubificoides pseudogaster (15.5%). As before, polychaetes and brittle starts were found in abundance in both beds, however, Malacostraca were more abundant in cluster 3, and oligochaetes were more abundant in cluster 1.
Although differences between the community structure in cluster 2 and cluster 3 were driven by a few key taxa, the top 10 taxa only contributed to 46.8 % of dissimilarity (Table 10). Therefore, compared to comparisons between the other clusters, differences in community structure between the cluster 2 and cluster 3 were driven by smaller changes in taxa. Although both communities had Malacostraca species present, polychaetes and oligochaetes were more abundant in cluster 2, and Ophiuroidea species were more abundant in cluster 3.
| Species | Class | Abundance in Cluster 1 | Abundance in Cluster 2 | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Capitella capitata | Polychaeta | 86 | 1 | 18.9 | 18.9 |
| Tubificoides pseudogaster | Oligochaeta | 81 | 12 | 14.4 | 33.3 |
| Ophiopholis aculeata | Ophiuroidea | 29 | 2 | 6.2 | 39.5 |
| Chaetozone setosa | Polychaeta | 1 | 17 | 4.9 | 44.4 |
| Tubificoides benedii | Oligochaeta | 23 | 0 | 4.7 | 49.1 |
| Mediomastus fragilis | Polychaeta | 24 | 9 | 3.5 | 52.6 |
| Pomatoceros triqueter | Polychaeta | 4 | 11 | 2.5 | 55.1 |
| Polydora caeca | Polychaeta | 0 | 8 | 2.0 | 57.1 |
| Crassicorophium bonellii | Malacostraca | 0 | 6 | 1.6 | 58.7 |
| Hiatella arctica | Bivalvia | 6 | 8 | 1.6 | 60.3 |
| Species | Class | Abundance in Cluster 1 | Abundance in Cluster 3 | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Capitella capitata | Polychaeta | 86 | 0 | 17.3 | 17.3 |
| Tubificoides pseudogaster | Oligochaeta | 81 | 1 | 15.5 | 32.8 |
| Ophiopholis aculeata | Ophiuroidea | 29 | 0 | 6.0 | 38.8 |
| Socarnes filicornis | Malacostraca | 0 | 25 | 5.9 | 44.7 |
| Mediomastus fragilis | Polychaeta | 24 | 1 | 5.2 | 49.9 |
| Amphipholis squamata | Ophiuroidea | 2 | 23 | 4.8 | 54.7 |
| Tubificoides benedii | Oligochaeta | 23 | 0 | 4.3 | 59.0 |
| Vaunthompsonia cristata | Malacostraca | 0 | 14 | 3.1 | 62.1 |
| Janira maculosa | Malacostraca | 0 | 8 | 1.9 | 64.0 |
| Paradialychone filicaudata | Polychaeta | 0 | 7 | 1.7 | 65.7 |
| Species | Class | Abundance in Cluster 2 | Abundance in Cluster 3 | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Socarnes filicornis | Malacostraca | 0 | 25 | 8.7 | 8.7 |
| Amphipholis squamata | Ophiuroidea | 0 | 23 | 7.4 | 16.1 |
| Chaetozone setosa | Polychaeta | 17 | 0 | 6.7 | 22.8 |
| Vaunthompsonia cristata | Malacostraca | 0 | 14 | 4.6 | 27.4 |
| Tubificoides pseudogaster | Oligochaeta | 12 | 1 | 4.5 | 31.9 |
| Pomatoceros triqueter | Polychaeta | 11 | 0 | 3.9 | 35.8 |
| Mediomastus fragilis | Polychaeta | 9 | 1 | 3.3 | 39.1 |
| Polydora caeca | Polychaeta | 8 | 1 | 2.7 | 41.8 |
| Janira maculosa | Malacostraca | 1 | 8 | 2.6 | 44.4 |
| Crassicorophium bonellii | Malacostraca | 6 | 5 | 2.4 | 46.8 |
3.2.2 Species diversity
In general, maerl bed habitat within the maerl and horse mussel mosaic had significantly higher species diversity measures than horse mussel bed habitat (Table 11 and Table 12). The maerl beds had a higher Shannon Diversity Index, a higher Pielou’s index and higher species richness values (Table 11 and Table 12). Abundance was not significantly different between habitat types.
| Location | Shannon Diversity Index | Pielou's evenness | Species Richness | Abundance |
|---|---|---|---|---|
| HS MB | 2.85 | 0.81 | 35 | 168 |
| NH MB | 2.92 | 0.82 | 36 | 160 |
| HS HM | 2.25 | 0.68 | 28 | 323 |
| US HM | 2.03 | 0.66 | 22 | 282 |
| BS HM | 2.77 | 0.85 | 31 | 113 |
| - | Bed Type | Bed Type |
|---|---|---|
| Diversity Index | LRT | p-value |
| Shannon Diversity Index | 8.49 | <0.01 |
| Pielou's Evenness | 5.57 | <0.02 |
| Species Richness | 5.76 | <0.02 |
3.2.3 Functional richness - abundance data
At Fetlar to Haroldswick, PCA analysis indicated that in general, horse mussel bed areas sampled within the mapped horse mussel and maerl bed mosaic were more functionally rich than the maerl bed areas (Figure 12 and Figure 13). The first two PCA axes explained 91.8 % of the total variance in functional traits, with the first principal component axis (PC1) accounting for 81.4 % of the variance, and the second principle component axis (PC2) accounting for 10.4 %. As samples from the Uyea Sound horse mussel bed (US HM), and the horse mussel bed at Hascosay (HS HM) were separated from the other sites along the PC1 axis, differences in functional diversity appeared to be driven by biogenic bed type. Differences in functional diversity were confirmed by statistical modelling, with the horse mussel beds at Uyea Sound and Hascosay having a significantly lower PC1 score than the other locations (LRT: χ(2) = 13.173, p<0.002; post hoc analysis, p<0.02 for all comparisons).
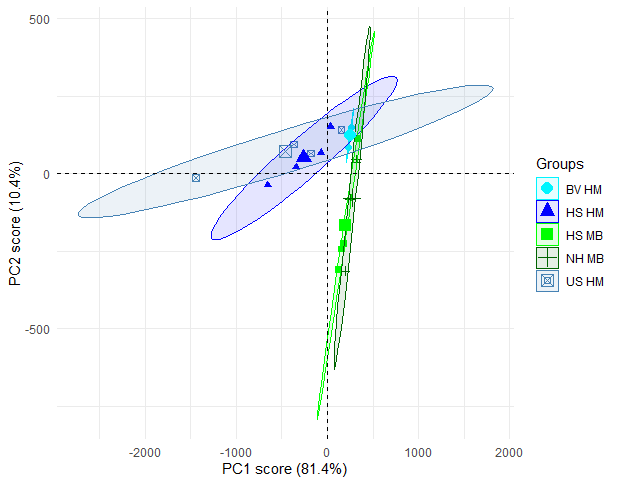
Figure 12: PCA plot comparing functional richness between independent maerl bed and horse mussel communities within the Fetlar to Haroldswick MPA. Each point represents an infaunal sample with text coloured by site.
Click for a full description
Position of samples in relation to PC1 and PC2 scores with 95% confidence ellipses. Locations are: Hascosay maerl bed (HS MB), Hascosay horse mussel bed (HS HM), North Hascosay maerl bed (NH MB), Basta Voe horse mussel bed (BV HM) and Uyea Sound horse mussel bed (US HM). Samples are coloured by biogenic bed type with horse mussel beds coloured blue and maerl beds coloured green. Larger symbols represent the centre of the ellipse for each group.
As the majority of functional traits are negatively correlated with PC1 values (Figure 13), the horse mussel bed areas within Uyea Sound and Hascosay are more functionally diverse than the other locations (reflected in the correspondence of the ellipses in Figure 12 with these negative PC1 values). Interestingly, different functional groups were found at the two locations. At Uyea Sound, taxa that were upward-conveyors, short lived (life span of less than 3 years), small (body size of 5-10 cm) with an affinity to muddy sand substrate were abundant. At Hascosay, a diverse range of taxa were abundant in the horse mussel bed including surface modifiers and biodiffusers, deposit feeders and suspension feeders, and species that ranged from medium to large (body size of over 80 mm and from 10-20 mm). Furthermore, taxa found at the Hascosay horse mussel bed had an affinity for a large range of substrate types including rock, coarse sand and sand.
The maerl beds at North Hascosay and Hascosay were not as functionally rich as the horse mussel beds. The positions of both maerl bed locations in Figure 12 overlap, indicating that they are functionally similar.
The horse mussel bed at Basta Voe was also not functionally rich with samples clustering tightly in Figure 12. This suggests that the habitat at Basta Voe may be relatively homozygous compared to the other sites, supporting a smaller range of functional groups.
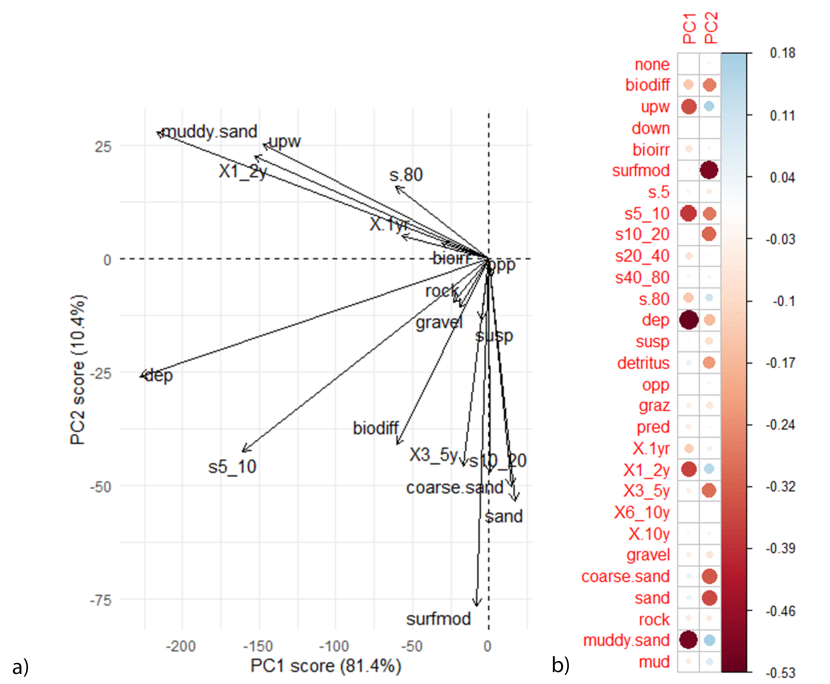
Figure 13: PCA plot showing a) the direction of functional traits in relation to PC1 and PC2 and b) the correlation matrix. Comparisons with Figure 12 show functional traits that result in differences between communities.
Click for a full description
PCA analysis contained independent maerl bed and horse mussel communities within the Fetlar to Haroldswick MPA. For the correlation matrix, red dots represent a negative correlation and blue dots a positive correlation. Size of dots correspond to the strength of the correlation. For functional trait acronyms refer to the Table 2.
3.2.4 Functional evenness - proportion data
PCA analysis showed that there were distinct functional differences between each location (Figure 14 and Figure 15). The first two PCA axes explained 68.8% of the total variance in functional traits, with the first principle component axis (PC1) accounting for 50.5% of variance, and the second principle component axis (PC2) accounting for 18.3%. Samples were separated by biogenic bed type (maerl bed vs. horse mussel bed) along the PC1 axis, indicating that differences in functional diversity were driven by differences in biogenic bed type within the mosaic. Differences in functional diversity due to bed type were confirmed by statistical modelling, with samples from maerl bed patches having a significantly lower PC1 score than samples from the horse mussel bed patches (LRT: χ(1) = 46.22, p<0.001). Samples from the Basta Voe horse mussel bed were also found to have a significantly lower PC2 value than the horse mussel bed samples from Hascosay and Uyea Sound (LRT: χ(4) = 41.50, p<0.001; post hoc analysis, p<0.001 for all comparisons).
The direction of functional traits plot and PC1 and PC2 rotation scores (Figure 15) show that sites varied in functional diversity. Maerl beds were dominated by organisms that were surface modifiers and biodiffusers, as well as opportunist feeders and organisms with a medium life span (3-5 years life span), and medium body size (body size of 10-20mm). Furthermore, organisms from maerl bed patches had an affinity for coarser substrate types including gravel, coarse sand and sand.
Horse mussel bed areas at Uyea Sound and Hascosay supported organisms that were upward-conveyors and deposit feeders and had an affinity for muddy sand substrates. The Basta Voe horse mussel bed was characterised by organisms that did not bioturbate the sediment, were suspension and detritus feeders and had a large range in life span (1-2 years and over 10 years) and in body size (<5mm and >80mm). The results suggest that differences in functional trait diversity were driven in small changes in the local habitats.
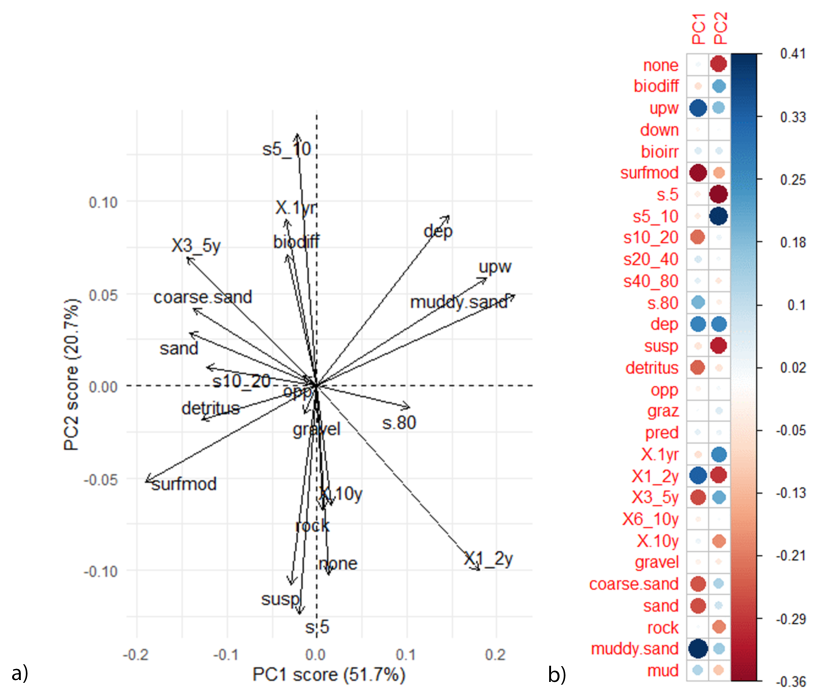
Figure 15: PCA plot showing a) the direction of functional traits in relation to PC1 and PC2 and b) the correlation matrix. Comparisons with Figure 14 show functional traits that result in differences between communities.
Click for a full description
PCA analysis contained independent maerl bed and horse mussel communities within the Fetlar to Haroldswick MPA. For the correlation matrix, red dots represent a negative correlation and blue dots a positive correlation. Size of dots correspond to the strength of the correlation. For functional trait acronyms refer to the Table 2.
3.3 National scale analysis: maerl beds
The maerl beds horizontally-linked with horse mussel beds in Fetlar to Haroldswick considered in Section 3.2 are compared here with non-linked / independent examples of maerl bed habitat from two areas >100 miles apart (no data was available for this study from independent maerl beds nearby Fetlar to Haroldswick).
3.3.1 Community structure
There were distinct locational patterns in community composition (Figure 16). Locational differences in community composition were confirmed by ANOSIM analysis (p<0.01, R2=0.84). The Sound of Barra and Orkney samples showed larger within site dissimilarity than the Fetlar to Haroldswick samples. Results for the log-transformed dataset are not shown, as transforming the data had no impact on the NMDS plots.
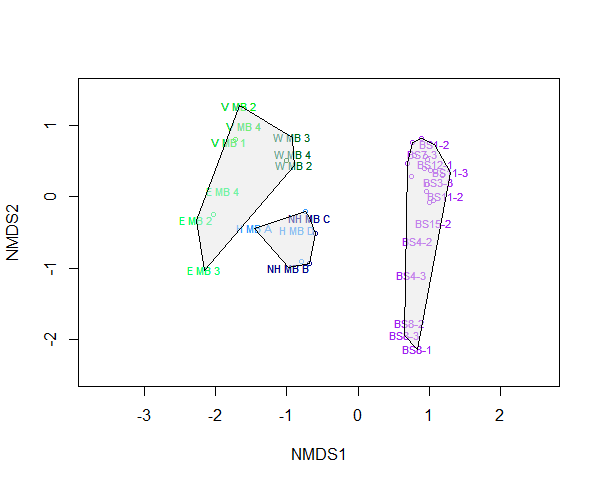
Figure 16: Two-dimensional non-metric multidimensional scaling (NMDS) plot, showing variation in community composition between maerl beds on a national scale. Sites include Orkney, Sound of Barra and Fetlar to Haroldswick.
Click for a full description
Stress level of plot was 0.04 after 20 runs. Polygons represent clustering by location. Sites are Wyre maerl bed (W MB), Eday maerl bed (E MB), Veantrow maerl bed (V MB), Hascosay maerl bed (H MB), North Hascosay maerl bed (NH MB), and Sound of Barra maerl bed (BS). Sites are coloured by location: Orkney is green, Sound of Barra is purple, and Fetlar to Haroldswick is blue.
A similarity percentages (SIMPER) analysis on the data identified which species contributed to differences between Fetlar to Haroldswick and the other maerl bed locations (Table 13 and Table 14). Differences between maerl beds were dominated by a few particular taxa, especially between Orkney and Fetlar to Haroldswick. When comparing Orkney and Fetlar to Haroldswick, the top 10 species contributed ~47.5 % of the average dissimilarity between the bed types, with Caprella acanthifera contributing the greatest amount (13.6 %; Table 13). When comparing Barra and Fetlar to Haroldswick, the top 10 species contributed ~49.9 % of the average dissimilarity between the bed types, with Socarnes filicornis contributing the greatest amount (12.4%; Table 14). In both comparisons, dissimilarities were dominated by Socarnes filicornis, Amphipholis squamata, Vaunthompsonia cristata, Janira maculosa, Maera othonis, and Paradialychone filicaudata, which were more abundant in the Fetlar to Haroldswick site.
| Species | Class | Abundance in Orkney | Abundance in FtoH | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Caprella acanthifera | Malacostraca | 108 | 0 | 13.6 | 13.6 |
| Socarnes filicornis | Malacostraca | 0 | 25 | 6.6 | 20.2 |
| Phtisica marina | Malacostraca | 37 | 0 | 6.4 | 26.6 |
| Amphipholis squamata | Ophiuroidea | 4 | 23 | 5.2 | 31.8 |
| Musculus discors | Bivalvia | 22 | 3 | 5.1 | 36.9 |
| Vaunthompsonia cristata | Malacostraca | 3 | 14 | 3.1 | 40 |
| Janira maculosa | Malacostraca | 5 | 8 | 2.2 | 42.2 |
| Lepidochitona cinerea | Polyplacophora | 4 | 0 | 1.9 | 44.1 |
| Paradialychone filicaudata | Polychaeta | 0 | 7 | 1.9 | 46 |
| Maera othonis | Malacostraca | 3 | 4 | 1.5 | 47.5 |
| Species | Class | Abundance in Barra | Abundance in FtoH | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| Socarnes filicornis | Malacostraca | 0 | 22 | 12.4 | 12.4 |
| Amphipholis squamata | Ophiuroidea | 1 | 23 | 9.9 | 22.3 |
| Vaunthompsonia cristata | Malacostraca | 0 | 14 | 6.3 | 28.6 |
| Janira maculosa | Malacostraca | 0 | 8 | 3.8 | 32.4 |
| Paradialychone filicaudata | Polychaeta | 0 | 7 | 3.5 | 35.9 |
| Goodallia triangularis | Bivalvia | 6 | 0 | 2.9 | 38.8 |
| Apherusa bispinosa | Malacostraca | 0 | 5 | 2.7 | 41.5 |
| Maera othonis | Malacostraca | 0 | 4 | 2.6 | 44.1 |
| Cymodoce truncata | Malacostraca | 0 | 6 | 2.4 | 46.5 |
| Crassicorophium bonellii | Malacostraca | 0 | 5 | 2.3 | 49.8 |
3.3.2 Functional richness - abundance data
Functional diversity varied between sites (Figure 17 and Figure 18). The first two PCA axes explained 66.9% of the total variance in functional traits, with the first principle component axis (PC1) accounting for 48.1% of the variance, and the second principal component axis (PC2) accounting for 18.8%. Differences in PC1 values and PC2 values were driven by site (LRT: χ(3) = 30.56, p<0.001 and LRT: χ(3) = 38.07, p<0.001 respectively), with Wyre Sound having a significantly lower PC1 value, and Fetlar to Haroldswick having a significantly higher PC2 value (post hoc analysis, p<0.001 for both comparisons). As PC1 values accounted for a larger amount of variance than PC2 values, maerl beds in Wyre are more dominated by different functional groups compared to maerl beds in Barra and Fetlar to Haroldswick.

Figure 17: PCA plot comparing functional richness between inter-linked maerl and horse mussel bed communities in Fetlar to Haroldswick and independent maerl bed communities within Orkney and the Sound of Barra.
Click for a full description
Position of samples in relation to PC1 and PC2 scores with 95% confidence ellipses. Sites are coloured by location with samples from Fetlar to Haroldswick coloured blue, samples from Wyre in Orkney coloured green and samples from the Sound of Barra coloured purple. Larger symbols represent the centre of the ellipse for each group.
The direction of functional traits plot and PC1 and PC2 rotation scores (Figure 18) show that Wyre maerl beds had a high abundance of organisms that were surface modifiers, detritus feeders, and short-lived (less than 1-year life span). In comparison, beds at Fetlar to Haroldswick had a higher abundance of traits such as upward-conveyors and biodiffusers, deposit feeders, and organisms with an affinity with coarse sand (Figure 17 and Figure 18). Maerl bed samples at Barra were found to have a low functional richness, with the direction of functional traits often pointing in the opposite direction to the samples (Figure 17 and Figure 18), however, some functional traits (such as long lived species and predators) were abundant in Barra.
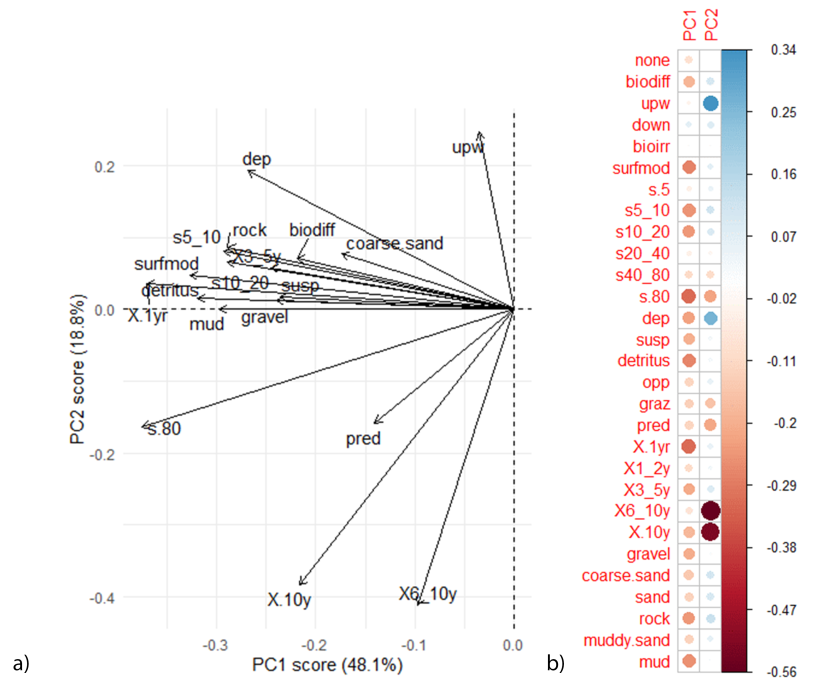
Figure 18: PCA plot showing a) the direction of functional traits in relation to PC1 and PC2 and b) the correlation matrix. Comparisons with Figure 17 show functional traits that result in differences between communities.
Click for a full description
PCA analysis contained inter-linked maerl and horse mussel bed communities in Fetlar to Haroldswick and independent maerl bed communities within Orkney and the Sound of Barra. For the correlation matrix, red dots represent a negative correlation and blue dots a positive correlation. Size of dots correspond to the strength of the correlation. For functional trait acronyms refer to the Table 2.
3.3.3 Functional evenness - proportion data
Functional diversity also varied between sites when using the proportion dataset (Figure 19 and Figure 20). The first two PCA axes explained 66.7 % of the total variance in function traits, with the first principle component axis (PC1) accounting for 42.8 % of variance, and the second principle component axis (PC2) accounting for 24.0 %. Differences in PC1 values and PC2 values were driven by site (LRT: χ(3) = 35.73, p<0.001 and LRT: χ(3) = 56.83, p<0.001 respectively), with Wyre having a significantly higher PC1 value, and Fetlar to Haroldswick having a significantly lower PC2 value (Figure 19; p<0.0012 for all comparisons). As PC1 values accounted for a larger amount of variance than PC2 values, maerl beds in Wyre are more functionally separate to maerl beds in Barra and Fetlar to Haroldswick.
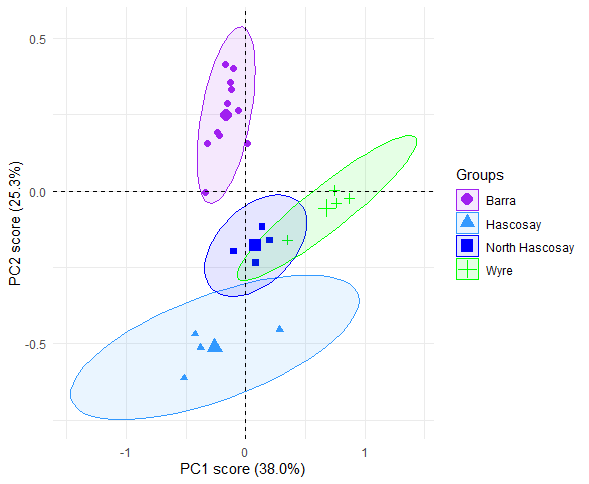
Figure 19: PCA plot comparing functional evenness between inter-linked maerl and horse mussel bed communities in Fetlar to Haroldswick and independent maerl bed communities within Orkney and the Sound of Barra.
Click for a full description
Position of samples in relation to PC1 and PC2 scores with 95% confidence ellipses. Sites are coloured by location with samples from Fetlar to Haroldswick coloured blue, samples from Wyre in Orkney coloured green and samples from the Sound of Barra coloured purple. Larger symbols represent the centre of the ellipse for each group.
The direction of functional traits plot and the PC1 and PC2 rotation scores (Figure 20) show similar results as above, with Wyre maerl beds dominated by species that were surface modifiers, detritus feeders and had a short life span of less than 1 year (Figure 19 and Figure 20). The Sound of Barra maerl bed supported small species (body size of less than 5mm) with a long life span (6-10 years and >10 years), as well predators. In Fetlar to Haroldswick, the maerl beds supported more species that were biodiffusers and had a small-medium body size (5-10 mm and 10-20mm) and a medium lifespan (3-5 years).
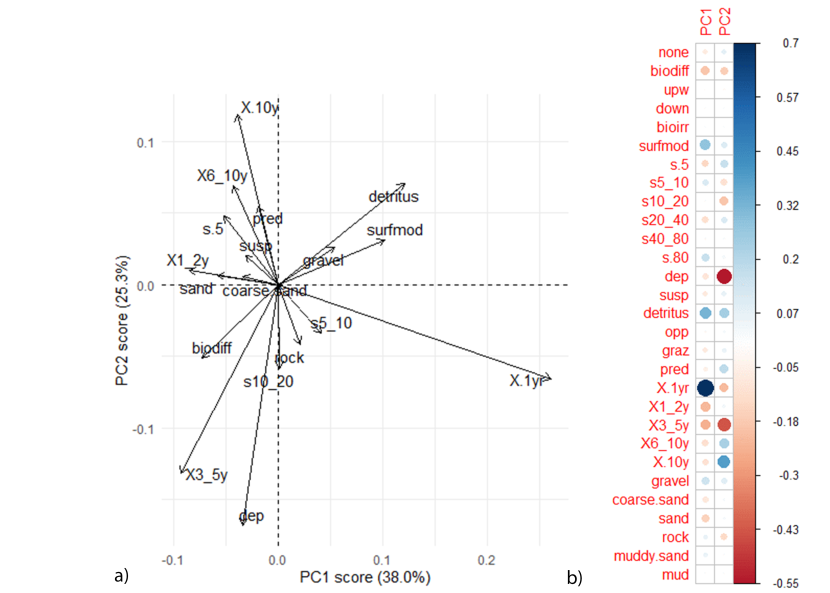
Figure 20: PCA plot showing a) the direction of functional traits in relation to PC1 and PC2 and b) the correlation matrix. Comparisons with Figure 19 show functional traits that result in differences between communities.
Click for a full description
PCA analysis contained inter-linked maerl and horse mussel bed communities in Fetlar to Haroldswick and independent maerl bed communities within Orkney and the Sound of Barra. For the correlation matrix, red dots represent a negative correlation and blue dots a positive correlation. Size of dots correspond to the strength of the correlation. For functional trait acronyms refer to the Table 2.
4. Discussion
This study aimed to investigate the effects of linkages between selected Priority Marine Feature (PMF) habitats in Scotland on their associated biological communities, and trialled a number of different approaches to assess these.
While results concerning particular effects of linkages between specific PMFs are preliminary and require further research (limited largely by the data available for the study), this study has identified a successful framework for investigating the effects of inter-habitat linkages between PMFs.
4.1 Methods review and recommendations
In agreement with previous studies (Donadi et al., 2015; Bremner et al., 2006), BTA was found to be an effective way to investigate differences in functional diversity between locations from an ecosystem rather than a species perspective. In contrast, NMDS plots, SIMPER analyses and ANOSIM analyses were not able to examine differences between locations from an ecosystem perspective, and instead reported differences between sites caused by a few dominant taxa. Although locations may appear to differ greatly in the NMDS plots, SIMPER analysis showed that often species with similar traits were abundant at both locations. For example, in Fetlar to Haroldswick, both horse mussel beds and maerl beds often had a high abundance of brittle stars, with differences in specific species of brittle stars causing the bed types to appear distinct in the analyses. However, whilst NMDS plots may indicate that community composition between locations differs, the locations may be functionally similar, as different species of brittlestar do not fulfil vastly different functional roles.
Therefore, it is recommended that future studies use BTA to investigate the effects of linkages between habitats to provide a broader ecosystem perspective. The approach is also recommended for general monitoring and assessment. In addition to better reflecting changes in ecosystem functioning, it also greatly facilitates the interpretation of complex datasets and presents results in a format more accessible to external stakeholders or indeed anybody less familiar with the species of marine infaunal communities.
For the BTA, trait information was obtained for 94% of taxa in the Fetlar to Haroldswick study and 96.4% of taxa in the Loch Alsh study. Similar to previous studies, information on life span was the main category of missing data (Paganelli et al., 2012). Also similar to previous research, this study found that searching for literature regarding biological traits was time-consuming, and the available information on life-cycle traits was limited. As the number and type of biological traits selected can influence the results (Paganelli et al., 2011), future studies should aim to include as many traits as possible. However, because information regarding the functional traits of many species is limited there must be a trade-off between time spent researching trait data and the number of traits included in the analysis. Future studies could ascertain that functional traits are not having a dominant effect on the result by removing traits and re-running the analysis. Dominant functional trait identification can be an informative process in its own right as well.
A key limitation in the study was the variation in infaunal sample data collection methods which influenced the results. This highlights the importance of using data collected by the same methods for comparisons. While one uniform sampling method and strategy for all habitats would be ideal statistically, practical considerations dictate that different methods will continue to be used, not least because no one method is suitable for every habitat. Future sampling programmes should however aim to use the same methods as used for existing data.
A further limitation of the study was the effects of environmental variables and pressures on the results. When comparing sites on a national scale, both BTA and NMDS plots showed distinct differences between locations, which was likely a result of environmental parameters and / or anthropogenic pressures. While efforts were made to only make comparisons between sites with similar physical attributes (and any that were extremely different were excluded from the analysis), future inter-habitat linkage studies should aim to select samples from beds within a narrow environmental and pressure envelope. A potential study site for this would be Loch Carron, where infaunal core samples were taken from a mixed flame shell and maerl bed as well as independent flame shell and maerl beds. Future research should consider available environmental data (such as e.g. wave energy, current speeds, depth, salinity) to investigate which environmental factors are driving differences between locations.
4.2 Effects of linkages
At a local scale, vertically linked habitats (i.e. flame shell beds overlaying or mixed with horse mussel beds) had a distinct biological community which was associated with higher biodiversity compared to non-linked examples of the habitats nearby.
Horizontally linked habitats (i.e. maerl bed and horse mussel bed patches adjacent to each other in a mosaic) did not share similar biological communities, suggesting that infaunal community structure was particular to biogenic bed type, and not driven by proximity to other PMFs. It was not possible to look at differences in diversity between horizontally linked habitats and independent habitats on a local scale because there is no data from locations which contain both independent as well as linked beds. When trying to compare linked and unlinked habitats on a national scale, community structure of maerl beds varied so strongly between locations that it was impossible to assess the potential impact of linkages. Temperature, depth and salinity executed the strongest influence on functional diversity of the habitats considered.
There was evidence that habitats with vertical linkages (i.e. flame shell beds overlaying or mixed with horse mussel beds) increased functional diversity on a local scale. Patterns in functional diversity were driven by factors such as substrate type and the type of biogenic structure (i.e. the presence of either maerl or horse mussels), and an increase in habitat and niche heterogeneity.
As an incidental finding, the results indicated that disturbance (both natural and / or anthropogenic) likely plays a role in shaping communities and their functional diversity, a finding made when comparing the functional diversity of communities from different sites with different degrees of disturbance. Further research is needed across multiple environmental and pressure gradients to understand this relationship.
The impact of linked habitats and habitat interactions on overall biodiversity and ecosystem functioning at a site is likely to have an important role in enhancing ecosystem resilience in the face of pressures, for example climate change. Previous studies have found that habitats with linkages can act as a buffer to climate stress (Bulleri et al., 2018). For example, in addition to increasing the species diversity (Angelini et al., 2015), the linkage between ribbon mussels and cordgrass (Alteri et al., 2007) also enhances the resistance and resilience of the system to drought (Angelini et al., 2016) and thermal stress (Crotty and Bertness, 2015). As another example, coastal plants can increase the pH in the surrounding water column via photosynthesis, protecting nearby mussel beds and coral reefs from the stress of ocean acidification (Wahl et al., 2017; Unsworth et al., 2012). As interactions between habitats can reduce the environmental stress experienced by individual organisms, understanding existing linkages in the marine environment is crucial for effective conservation in a changing world (Gaines and Roughgarden 1987, Rilov and Schiel 2011).
4.2.1 Community structure
Local-scale comparisons
When comparing community composition in Loch Alsh, habitat linkages between flame shell beds and horse mussel beds appeared to influence community composition. Linked beds had higher abundances of most species, including polychaetes, such as Jasmineira elegans and Nereimyra punctata, and oligochaetes such as Aonides oxycephala. As horse mussels and flame shells co-existed spatially, the higher abundances in the mixed bed may have been a consequence of increased niche availability by effectively providing two habitats in one physical location (Petrosillo and Zurlini, 2016; Donadi et al., 2015).
The community structures of the eastern and western independent flame shell beds in Loch Alsh differed from each other; the eastern flame shell bed had a higher abundance of polychaetes, bivalves and Malacostraca species than the western bed. The western bed was a little shallower than the eastern bed 6 – 10 m and 16.5 m respectively (see Appendix 4 for details). The observed differences between the communities of the independent flame shell beds may be caused by both locational and temporal parameters, as well as differences in sampling methodology. The western flame shell bed infaunal samples were collected using a Day grab and those from the eastern flame shell bed were from diver-collected cores. Grab samples may preferentially sample infaunal taxa in the surface layers of the bed, while core samples may more consistently target infaunal taxa at greater depths in the sediments below. Locational patterns may have driven the differences since the western bed is situated outside the Kyle Akin Narrows, where conditions (e.g. greater exposure and reduced current speeds) may be less favourable for certain species. The effects of locational patterns on community structure has been observed in the literature previously, with environmental parameters, such as organic enrichment, sediment instability and oxygen levels leading to locational differences in community structure (Paganelli et al., 2012; Trigg et al., 2011). Differences between sites may also be due to seasonal patterns, with the western flame shell bed sampled in March 2014 and the eastern flame shell bed in August 2014. Organisms that exhibited seasonal changes were not excluded from this analysis and could have driven changes in community composition between the two sites (Trigg et al., 2011).
In Fetlar to Haroldswick, community structure within the maerl and horse mussel mosaic varied between different patches of biogenic habitat. Horse mussel bed samples from Hascosay and Uyea Sound shared a similar community that was distinct to that from the horse mussel bed at Basta Voe. Differences between horse mussel bed communities were often driven by species associated with organic enrichment, such as the polychaete Capitella capitata and the oligochaete Tubificoides pseudogaster, which suggests some horse mussel beds may have been organically enriched. The pattern was not mirrored in maerl bed communities at the same locations though, so it is unlikely that organic enrichment had been imposed on the wider environment. The findings of the BTA indicate that community composition within the maerl and horse mussel bed mosaic in Fetlar to Haroldswick may also be influenced by environmental parameters that were not considered in this study.
Whilst mobile species were not included in this analysis, it is likely that habitat mosaics are beneficial for cross-habitat foragers (Lundberg and Moberg 2003). In the absence of data from unlinked examples of either habitat in this location it was, however, not possible to quantify the biodiversity benefits conferred by the horizontal linkages.
National-scale comparisons
During the investigation of differences between linked and independent maerl beds on a national scale, results indicated that species composition seemed to be driven by location-specific environmental differences more strongly than by the presence or absence of habitat linkages.
The differences in community structure between locations were defined to a large degree by a few key species. At Fetlar to Haroldswick, samples had a higher abundance of the oligochaete Tubificoides Pseudogaster, the polychaete Capitella capitata, the crustacean Socarnes filicornis, and the brittle stars Ophiopholis aculeata and Amphipholis squamata, than samples from the other locations. Species that had a higher abundance in the other locations were mainly bivalves (such as Goodallia triangularis, which was more abundant in the Sound of Barra), although samples from Orkney were dominated by the skeleton shrimp species Caprella acanthifera and Phtisica marina.
As an example of location-specific factors which drive differences in community structure, in Wyre Sound the very shallow maerl bed (0-5 m) supported a dense macro-algal coverage in the summer which could explain the high abundance of Caprella acanthifera and Phtisica marina species, which live on fronds of algae (Isaac and Moyse, 1990). In contrast, the sites at Barra were deeper at mainly >20 m and in a more open-coast setting, which could explain the presence of the small robust bivalve Goodallia triangularis, which has a depth range of up to 150m (Dall et al., 1803). Fetlar to Haroldswick is characterised by species that bio-turbate the sediment (such as Tubificoides Pseudogaster and Capitella capitata), suggesting that the sediment at this site may be siltier. This could be a consequence of horizontal linkages between maerl and horse mussel beds, as mussels are known to dissipate wave energy and create accumulations of pseudofaeces, which could lead to a siltier environment (Donadi et al., 2013).
4.2.2 Biodiversity
Findings on the impacts of linkages on biodiversity followed the same pattern as community structure.
Local-scale comparisons
On a local scale, vertical habitat linkages in Loch Alsh appeared to influence biodiversity. The Shannon Wiener Index, species richness, and abundance were all significantly higher in the linked flame shell and horse mussel bed samples than in the independent flame shell beds nearby. As the habitats are linked vertically, it is assumed that the increased biodiversity at the linked sites was a result of increased niche availability (Donadi et al., 2015). For example, organisms may attach to the shell of the horse mussel (Alteri et al., 2007), or bury themselves in the stable mat created by the byssal threads of both the flame shells and the horse mussels.
In Fetlar to Haroldswick, where horse mussel beds and maerl beds occur in a habitat mosaic, the maerl bed areas were more diverse than horse mussel bed areas, with horse mussel beds containing more dominant species. However, it was not possible to compare the diversity of the linked habitats in Fetlar to Haroldswick to independent habitats on a local scale because there were no comparable datasets from independent beds in the site.
National-scale comparisons
It was also not possible to compare the diversity of the linked habitats to independent habitats at a national scale because of the limited dataset used in this study. Therefore it cannot be concluded that the high biodiversity of the Fetlar to Haroldswick maerl beds is related to the presence of adjacent horse mussel beds.
4.2.3 Functional diversity
Functional trait analysis was found to be an effective method to assess differences and similarities between communities.
Local-scale comparisons
In Loch Alsh it was possible to demonstrate that functional diversity differed between linked and independent habitats, with vertically linked habitats (flame shell beds and horse mussel beds) having a higher functional diversity compared to the independent flame shell beds.
Biological trait analysis (BTA) indicated that on a local scale in Loch Alsh, habitat linkages increased functional richness. When comparing linked flame shell (FS) and horse mussel beds (HS) to the independent flame shell beds, there was a greater range of functional trait categories in the linked habitats. Unsurprisingly, functional groups were connected to substrate type, with deposit feeders found in muddier sediments, and suspension feeders found in coarser sediments. Fine sediment can clog the feeding appendages of suspension feeders (Paganelli et al., 2012). The results suggest that substrate type varied within the linked bed in Loch Alsh (FS & HM 2 having finer sediment and FS & HM 1 having coarser sediment). Small changes in substrate type may be one of the key drivers of increased functional diversity. Further research is needed, using substrate type as an additional environmental parameter, to explore the link between substrate type and functional heterogeneity and between the key species and the substrate present.
Interestingly, dominant functional traits present within the linked habitat FS & HM 1 were similar to the functional traits found within the independent eastern flame shell bed station, namely suspension feeders and bio-irrigators. In comparison, samples from the western flame shell bed were dominated by different functional traits such as predators and organisms with a medium life span (3-5 years). There was a slight overlap between samples from the western and eastern flame shell beds, although sites shared fewer functional groups compared to the linked beds and the eastern flame shell bed. As the eastern flame shell bed and the two linked beds are located within Loch Alsh, and the western flame shell bed is found just outside the Kyle Akin Narrows, these results indicate that functional traits may be influenced quite strongly by local environmental parameters, such as exposure and current speed. Further research is needed to understand the effects of environmental parameters, such as wave energy, bottom temperature and salinity on driving functional diversity.
Within Fetlar to Haroldswick, functional traits varied considerably between sampling areas within the linked horse mussel and maerl beds. Horse mussel bed samples at Hascosay and Uyea Sound were more functionally rich compared to the other sites, supporting a large range of functional traits. Functional traits varied between biogenic bed type, with maerl beds and horse mussel beds having different dominant traits. The small-scale habitat heterogeneity across the biogenic habitat mosaic in Fetlar to Haroldswick enhances the functional diversity of the overall environment, but no conclusions could be drawn on whether the communities of the individual horse mussel and maerl bed patches were influenced by the proximity of patches of the other habitat in the mosaic.
National-scale comparisons
Biological trait analysis (BTA) indicated that on a national scale, the types of functional traits present varied greatly between locations. The results suggest that on a larger scale, functional diversity may be influenced more strongly by environmental factors and pressure from anthropogenic activities than by the presence or absence of habitat linkages. Assessments of datasets that share similar environmental parameters and pressures would be needed to identify the role of inter-habitat linkages on functional diversity on a national scale.
In Wyre Sound, the maerl bed was dominated by short lived species with a life span of less than 1 year, as well as by detritus feeders. Small, short lived, opportunist species (such as detritus feeders) often characterise stressed benthic habitats, which suggest that the Wyre Sound maerl bed might be exposed to stressors (Paganelli et al., 2012). This high occurrence of opportunist species in Wyre Sound may be a result of the high wave energy experienced by the benthic communities in the very shallow waters.
An interesting difference between the Wyre Sound and Fetlar to Haroldswick beds is that unlike the maerl beds in Fetlar to Haroldswick, the maerl bed in Wyre Sound was dominated by surface modifiers and had a lower abundance of species that work the sediment. Bioturbators play an important role in the breakdown of organic material within the sediment. This could mean that, compared to Fetlar to Haroldswick, carbon in Wyre Sound is broken down less in the top layers of sediment in the bed. This may be driven by the higher levels of wave energy causing mixing of the bed to a deeper level.
The communities in the Sound of Barra maerl beds were less functionally rich than those in Wyre Sound and Fetlar to Haroldswick, and were dominated by species that were small (less than 5 mm body size) but relatively long lived (known maximum life span of over 6 years – though obviously it is unknown whether individuals at the location achieve this age), as well as by predators (such as Psamathe fusca). Like in Wyre Sound, the high abundance of small organisms suggests that the Barra maerl beds may be subject to regular disturbance. Additional work is needed to clarify whether environmental parameters or anthropogenic pressures which were not considered in this study are driving this difference in community composition. The maerl beds in the Sound of Barra are deeper than in Wyre Sound and Fetlar to Haroldswick, potentially more exposed in a more open-coast location, and are made up of mega-ripples, where maerl accumulates in troughs between waves of coarse sand and shell gravel, all of which influence community composition. Dredging pressure, which is present in the Sound of Barra, could also have increased the proportion of smaller species which are not trapped and retained in dredges (Mercer et al., 2018).
5. Conclusions and future work
This study established a useful process to assess linkages between PMFs using biodiversity measurements and biological trait analysis. With limited resources, marine management often requires prioritising of features and sites for monitoring and protection. The results presented here from local scales suggest that linked habitats are important in terms of their increased species and functional diversity. Diverse habitats are recognised for the variety of ecosystem services they provide and being more resilient to pressures such as climate change. Further work is needed to determine if other linked PMFs exhibit similar patterns in terms of elevated diversity and differing community structure. Follow-up studies are needed to explore whether the effect of inter-habitat linkages at the local scale found in this study holds across other locations and habitats. If this is the case it would have important implications for prioritising inter-linked habitats in marine management in the future.
The methodology presented here offers the potential for much better quantification and interpretation of the services provided by PMF habitats, and by systems of multiple habitats. Adoption of such methodologies in wider monitoring and assessment work could make a significant contribution to moving towards a focus on ecosystem functioning in marine nature conservation.
The results from this study also indicated that BTA could be used to explore the effects of anthropogenic and natural disturbances on ecosystems. Functional traits that are prevalent in a site may vary depending on how stressed the system is. By researching the effects of disturbance on functional traits, it may be possible to identify how disturbance affects the ecosystem as a whole, rather than its effects on individual species which may be replaced by species with similar functional traits (Bremner et al., 2006). This research could provide valuable evidence in a marine management context, allowing habitats whose functions and services are being altered through anthropogenic impacts such as demersal fishing to be identified.
There is also potential to investigate the effects of functional diversity on blue carbon sequestration. At certain sites in this study, bioturbators were prevalent, which mix the upper layers of sediment, in turn affecting carbon breakdown and sequestration. The effects of bioturbators on blue carbon sequestration is currently poorly understood, however and remains a vital research gap (Thomson, 2019). By analysing the organic content and infaunal data within sediment cores, future research could use BTA analysis, with organic content as an additional environmental parameter, to explore the effects of bioturbators on carbon sequestration.
In general, this study highlighted the effects of location on community structure, biodiversity and functional diversity. Future work should aim to include additional environmental parameters in the analysis (such as temperature, substrate type and exposure), as variable environmental conditions appeared to override effects of linkages in this study.
REFERENCES
Altieri, A.H., Silliman, B.R., and Bertness, M.D., 2007. Hierarchical Organisation via a Facilitation Cascade in Intertidal Cordgrass Bed Communities. The American Naturalist, 169 (2): 195-206.
Angelini, C., Griffin, J.N., van de Koppel. J., Lamers, L.P.M., Smolders, A.J.P., Derksen-Hooijberg, M., van der Heide, T., and Silliman, B.R., 2016. A keystone mutualism underpins resilience of a coastal ecosystem to drought. Nature Communications, 7: 12473.
Angelini, C., van der Heide, T., Griffin, J.N., Morton, J.P., Derksen-Hooijberg, M., Lamers, L.P.M., Smolders, A.J.P., and Silliman, B.R., 2015. Foundation species’ overlap enhances biodiversity and multifunctionality from the patch to landscape scale in southeastern United States salt marshes. Proceedings of the Royal Society B: Biological Sciences, 82 (1811): 20150421.
Austen, M.C., Lambshead, P.J.D., Hutchings, P.A., Boucher, G., Snelgrove, P.V.R., Heip, C., King, G., Koike, I., and Smith, C., 2002. Biodiversity links above and below the marine sediment-water interface that may influence community stability. Biodiversity and Conservation, 11: 113-136.
Bagur, M., Gutiérrez, J.L., Arribas, L.P., and Palomo, G.M., 2016. Complementary influences on co-occurring physical ecosystem engineers on species richness: insights from a Patagonian rocky shore. Biodiversity and Conservation, 25: 2787-2802.
Barbera. C., Bordehore, C., Borg, J.A., Glémarec, M., Grall, J., Hall‐Spencer, J.M., de la Huz, Ch., Lanfranco, E., Lastra, M., Moore, P.G., Mora, J., Pita, M.E., Ramos‐Esplá, A.A., Rizzo, M., Sánchez‐Mata, A., Seva, A., Schembri, P.J., and Valle, C., 2003. Conservation and management of northeast Atlantic and Mediterranean maerl beds. Aquatic Conservation: Marine and Freshwater Ecosystems, 13: 65-76.
Barton, K., 2019. MuMIn: Multi-Model Inference. R package version 1.43.15. https://CRAN.R-project.org/package=MuMIn
Bertolini, C., Montgomery, W.I., O’Connor, N.E., 2018. Habitat with small inter-structural spaces promotes mussel survival and reef generation. Marine Biology, 165 (163):1-11.
Bremner, J., Rogers, S.I., Frid, C.L.J. 2006. Methods for describing ecological functioning of marine benthic assemblages using biological traits analysis (BTA). Ecological Indicators, 6: 609-622.
Bulleri, F., Eriksson, B.K., Queirós, A., Airoldi, L., Arenas, F., Arvanitidis, C., Bouma, T.J., Crowe, T.P., Davoult, D., Guizien, K., Iveša, L., Jenkins, S.R., Michalet, R., Olabarria, C., Procaccini, G., Serrão, E.A., Wahl, M., Benedetti-Cecchi, L., 2018. Harnessing positive species interactions as a tool against climate-driven loss of coastal biodiversity, PLOS Biology, 16(9): e2006852.
Chevenet, F., Dolédec, S., Chessel, D., 1994. A fuzzy coding approach for the analysis of long-term ecological data. Freshwater Biology, 31: 295-309.
Crotty, S., and Bertness, M.D., 2015. Positive interactions expand habitat use and the realized niches of sympatric species. Ecology, 96 (10): 2575-2582.
Cook, R., Fariñas-Franco, J.M., Gell, F.R., Holt, R.H.F., Holt, T., Lindenbaum, C., Porter, J.S., Seed, R., Skates, L.R., Stringell, T.B., Sanderson, W.G., 2013. The substantial first impact of bottom fishing on rare biodiversity hotspots: A dilemma for evidence-based conservation. PLOS One, 8 (8): e69904.
Dall, W., Dorville, E., Montagu, G., 1803. Testacea Britannica, or natural history of British shells, marine, land and the fresh-water, including the most minute: systematically arranged and embellished with figures. Sold by J. White, London, 1-924.
Demars, B. O. L., Kemp, J. L., Friberg, N., Usseglio-Polater, P., and Harper, D. M. 2011. Linking biotopes to invertebrates in rivers: Biological traits, taxonomic composition and diversity. Biological Indicators, 23: 301-311.
Donadi, S., van der Heide, T., Piersma, T., van der Zee, E.M., Weerman, E.J., van de Koppel, J., Olff, H., Devine, C., Hernawan, U.E., Boers, M., Planthof, L., Eriksson, B.K., 2015. Multi-scale habitat modification by coexisting ecosystem engineers drives spatial separation of microbenthic functional groups. Nordic Society Oikos, 124 (11): 1502-1510.
Donadi, S., van der Heide, T., van der Zee, E.M., Eklöf, J.S., van de Koppel, J., Weerman, E.J., Piersma, T., Olff, H., Eriksson, B.K., 2013. Cross-habitat interactions among bivalve species control community structure on intertidal flats. Ecology, 94 (2), 489-498.
Dutertre, M., Grall, J., Ehrhold, A., Hamon, D., 2015. Environmental factors affecting maerl bed structure in Brittany (France). European Journal of Phycology, 50 (4): 371-383.
Ellingsen, K.E., Gray, J.S. (2002). Spatial patterns of benthic diversity: is there a latitudinal gradient along the Norwegian continental shelf? Journal of Animal Ecology, 71: 373-389.
Farinas-Franco, J.M., Allcock, L.A., Roberts, D., 2018. Protection alone may not promote natural recovery of biogenic habitats of high biodiversity damaged by mobile fishing gears. Marine Environmental Research, 135: 18-28.
Fariñas-Franco, J.M., Pearce, B., Porter, J., Harries, D., Mair, J.M., Woolmer, A.S. & Sanderson, W.G., 2014. Marine Strategy Framework Directive Indicators for Biogenic Reefs formed by Modiolus modiolus, Mytilus edulis and Sabellaria spinulosa. Part 1: Defining and validating the indicators, JNCC Report No. 523, ISSN 0963-8091.
Farinas-Franco, J.M., and, Roberts, D., 2014. Early faunal successional patterns in artificial reefs used for restoration of impacted biogenic habitats. Hydrobiologia, 727 (1): 75-94.
Gaines, S.D., and Roughgarden, J., 1987. Fish in offshore kelp forests affect recruitment to intertidal barnacle populations. Science, 235:479-481.
Gormley, K.S., Porter, J.S., Bell, M.C., Hull, A.D., & Sanderson, W.G., 2013. Predictive habitat modelling as a tool to assess the change in distribution and extent of an OSPAR priority habitat under an increased ocean temperature scenario: consequences for marine protected area networks and management. PlOS ONE, 8(7): e68263.
Gribben, P.E., Byres, J.E., Clements, M., McKenzie, L.A., Steinberg, P.D., Wrights, J.T., 2009. Behavioural interactions between ecosystems engineers control community species richness. Ecology Letters, 12: 1127-1136.
Hall-Spencer, J.M., Grall, J. Moore, P.G., and Atkinson, R.J.A., 2003. Bivalve fishing and maerl-bed conservation in France and the UK - retrospect and prospect. Aquatic Conservation: Marine and Freshwater ecosystems, 13: 33-41.
Hall-Spencer, J.M., and Moore, P.G., 2000a. Limaria hians (Mollusca: Limacea): a neglected reef-forming keystone species. Aquatic Conservation: Marine and Freshwater Ecosystems, 10 (4): 267-277.
Hall-Spencer J.M. and Moore P.G., 2000b. Scallop dredging has profound long-term impacts on maerl habitats. ICES Journal of Marine Science, 57: 1407-1415.
Hennig, C., 2020. fpc: Flexible Procedures for Clustering. R package version 2.2-4. https://CRAN.R-project.org/package=fpc
Hirst, N.E., Kamphausen, L.M., Cook, R.L., Porter, J.S., and Sanderson, W.G., 2013. Distribution and status of proposed protected features in the Fetlar to Haroldswick MPA proposal. Scottish Natural Heritage Commissioned Report No. 599.
Hirst, N. E., Clark, L., Sanderson, W.G., 2012a. The distribution of selected MPA search features and Priority Marine Features off the NE coast of Scotland. Scottish Natural Heritage Commissioned Report 500.
Hirst, N.E., Cook, R., James, B., Kent, F., Loxton, J., Porter, J. and Sanderson, W.G., 2012b. The distribution of Priority Marine Features and MPA search features in Orkney waters: Rousay to Copinsay including Eday Sound and points in Scapa Flow. Scottish Natural Heritage Commissioned Report No. 509.
Isbell, F., Craven, D., Connolly, J., Loreau, M., Schmid, B., Beierkuhnlein, C., Bezemer, T.M., Bonin, C., Bruelheide, H., de Luca, E., Ebeling, A., Griffin, J.N., Guo, Q., Hautier, Y., Hector, A., Jentsch, A., Kreyling, A., Lanta, V., Manning, P., Meyer, S.T., Mori, A.S., Naeem, S., Niklaus, P.A., Polley, H.W., Reich, P.B., Roscher, C., Seabloom, E.W., Smith, M.D., Thakur, M.P., Tilman, D., Tracy, B.F., van der Putten, W.H., van Ruijven, J., Weigelt, A., Weisser, W.W., Wilsey, B., and Eisenhauer, N., 2015. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature, 526: 574-589.
Jackson, C. M., Kamenos, N.A., Moore, P.G., and Young, M., 2004. Meiofaunal bivalves in maerl and other substrata; their distribution and community structure. Ophelia, 58 (1): 49-60.
Jones, D. L., Walter, J.F., Brooks, E.N., and Serafy, J.E., 2010. Connectivity through ontogeny: fish population linkages among mangrove and coral reef habitats. Marine Ecology Progress Series, 401: 245-258.
Kamenos, N.A., Moore, P.G., and Hall-Spencer, J.M., 2004. Small-scale distribution of juvenile gadoids in shallow inshore waters; what role does maerl play? ICES Journal of Marine Science, 61 (3): 422-429.
Kamenos, N.A., Moore, P.G., and Hall-Spencer, J.M. 2003. Substratum heterogeneity of dredged vs un-dredged maerl grounds. Journal of the Marine Biological Association of the United Kingdom, 83 (2): 411-413.
Kassambara, A., and Mundt, F., 2019. factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.6.
Kent, F. E., Mair, J. M., Newton, J., Lindenbaum, C., Porter, J. S., & Sanderson, W. G., 2017. Commercially important species associated with horse mussel (Modiolus modiolus) biogenic reefs: a priority habitat for nature conservation and fisheries benefits. Marine Pollution Bulletin, 118 (1-2): 71-78.
Lancaster, J. (Ed.), McCallum, S., Lowe A.C., Taylor, E., Chapman A. & Pomfret, J. 2014. Development of detailed ecological guidance to support the application of the Scottish MPA selection guidelines in Scotland’s seas. Scottish Natural Heritage Commissioned Report No. 491.
Lundberg, J., and Moberg, F., 2003. Mobile link organisms and ecosystem functioning: implications for ecosystem resilience and management. Ecosystems, 6(1): 87-98.
Mercer, T.M., Bunker, F.StP.D, Howson, C.M. & Kamphausen, L. 2018. Sound of Barra macrobenthic infaunal survey and Eriskay Causeway sediment survey 2015. Scottish Natural Heritage Research Report No. 951.
Moore, C.G., Harries, D.B., Tulbure, K.W., Cook, R.L., Saunders, G.R., Lyndon, A.R., Kamphausen, L., & James, B. 2020. The current status of serpulid reefs, horse mussel beds and flame shell beds in Loch Creran SAC and MPA. Scottish Natural Heritage Research Report No. 1156.
Moore, C.G., Harries, D.B., James, B., Cook, R.L., Saunders, G.R, Tulbure, K.W., Harbour, R.P. & Kamphausen, L. 2018. The distribution and condition of flame shell beds and other Priority Marine Features in Loch Carron Marine Protected Area and adjacent waters. Scottish Natural Heritage Research Report No. 1038.
Moore, C.G., Harries, D.B., Cook, R.L., Hirst, N.E., Saunders, G.R., Kent, F.E.A., Trigg, C. and Lyndon, A.R., 2013. The distribution and condition of selected MPA search features within Lochs Alsh, Duich, Creran and Fyne. Scottish Natural Heritage Commissioned Report No. 566.
Moore, C. G., Harries, D. B. & Trigg, C. 2012. The distribution of selected MPA search features within Lochs Linnhe, Etive, Leven and Eil: a broadscale validation survey (Part B). Scottish Natural Heritage Commissioned Report No. 502.
Oksanen, J F. et al., 2019. vegan: Community Ecology Package. R package version 2.5-6. https://CRAN.R-project.org/package=vegan
Oliver, T. H., Heard, M.S., Isaac, N.J.B., Roy, D.B., Procter, D., Eigenbrod, F., Freckleton, R., Hector, A., Orme, C.D.L., Petchey, O.L., Proenca, V., Raffaelli, D., Suttle, K.B., Mace, G.M., Martin-Lopez, B., Woodcock, B.A., and Bullock, J.M., 2015. Biodiversity and Resilience of Ecosystem Functions. Trends in Ecology and Evolution, 30 (11): 673-684.
Paganelli, D., Marchini, A., Forni, G., Occhipinti-Ambrogi, A. 2011. Functional diversity of the soft-bottom macrobenthos community 3km off the Emilia-Romagna coastline. Studi trentini di scienze naturali. 89: 99-105.
Passarelli, C., Olivier, F., Paterson, D.M., Meziane, T., Hubas, C., 2014. Organisms as cooperative ecosystem engineers in intertidal flats. Journal of Sea Research, 92: 92-101.
Petrosillo, I., and Zurlini, G. 2016. The important role of ecological engineers in providing ecosystem services at landscape level. Animal Conservation, 19 (6): 500-501.
Pister, B. 2009. Urban marine ecology in southern California: the ability of riprap structures to serve as rocky intertidal habitat. Marine Biology, 156: 861-873.
Queiros, A. M., Birchenough, S.N.R., Bremner, J., Godbold, J.A., Parker, R.E., Romero-Ramirez, A., Reiss, H., Solan, M., Somerfield, P.J., Colen, C.V., Hoey, G.V., Widicombe, S. 2013. A bioturbation classification of European marine infaunal invertebrates. Ecology and Evolution, 3(11): 3958-3985.
Reiss, H., Degraer, S., Duineveld, G.C.A., Kroncke, I., Aldridge, J., Craeymeersch, J.A., Eggleton, J.D., Hillewaert, H., Lavaleye, M.S.S., Moll, A., Pohlmann, T., Rachor, E., Robertson, M., Berghe, E.V., van Hoey, G., and Ress, H.L. 2009. Spatial patterns of infauna, epifauna, and demersal fish communities in the North Sea. ICES Journal of Marine Science, 67 (2): 278-293.
Rilov, G., and Schiel, D.R., 2011. Community regulation: the relative importance of recruitment and predation intensity of an intertidal community dominant in a seascape context. PLOS ONE, 6 (8): e23958.
Riosmena-Rodríguez, R. 2017. “Natural History of Rhodolith/Maërl Beds: Their Role in Near-Shore Biodiversity and Management”, In: Riosmena-Rodríguez, R., Nelson, W., and Aguirre, J. (ed.) Rhodolith/Maërl Beds: A Global Perspective. Springer International Publishing, pp. 3-26.
Saintilan, N., Hossain, K., and Mazumder, D., 2007. Linkages between seagrass, mangrove and saltmarsh as fish habitat in the Botany Bay estuary, New South Wales. Wetlands Ecology and Management, 15:277-286.
Sanderson, W., Holt, R., Kay, L., Ramsay, K., Perrins, J., McMath, A., and Rees, E., 2008. Small-scale variation within a Modiolus modiolus (Mollusca: Bivalvia) reef in the Irish Sea. II. Epifauna recorded by divers and cameras. Journal of the Marine Biological Association of the United Kingdom, 88 (1): 143-149.
Schleuter, D., Daufresne, M., Massol, F., and Argillier, C., 2010. A user’s guide to functional diversity indices. Ecological monographs, Ecological Society of America, 80 (3), 469- 484.
Simon-Nutbrown, C., Hollingsworth, P.M., Fernandes, T.F., Kamphausen, L., Baxter, J.M. and Burdett, H.L. 2020. Species Distribution Modeling Predicts Significant Declines in Coralline Algae Populations Under Projected Climate Change With Implications for Conservation Policy. Fronties in Marine Science, 7: 575825. doi: 10.3389/fmars.2020.575825
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M.C., Schwager, M., and Jeltsch, F., 2014. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography, 31: 79-92.
Thomson, A.C.G., Trevathan‐Tackett, S.M., Maher, D.T., Ralph, P.J., and Macreadie, P.I., 2019. Bioturbator-stimulated loss of seagrass sediment carbon stocks. Limnology and Oceanography 64(1), pp. 342-356.
Trigg, C., Harries, D., Lyndon, A., Moore, C.G., 2011. Community composition and diversity of two Limaria hians (Mollusca: Limacea) beds on the west coast of Scotland. Journal of the Marine Biological Association of the United Kingdom, 91 (7): 1403-1412.
Trigg C. and Moore C.G., 2009. Recovery of the biogenic nest habitat of Limaria hians (Mollusca: Limacea) following anthropogenic disturbance. Estuarine, Coastal and Shelf Science, 82: 351-356.
Trussel, G. C., Euanchuck, P.J., and Matassa, C.M., 2006. Habitat effects on the relative importance of trait and density-mediated indirect interactions. Ecology Letters, 9(11): 1245-1252.
Tyler-Walters, H., James, B., Carruthers, M. (eds.), Wilding, C., Durkin, O., Lacey, C., Philpott, E., Adams, L., Chaniotis, P.D., Wilkes, P.T.V., Seeley, R., Neilly, M., Dargie, J. & Crawford-Avis, O.T. 2016. Descriptions of Scottish Priority Marine Features (PMFs). Scottish Natural Heritage Commissioned Report No. 406.
Unsworth, R.K.F., Collier, C.J., Henderson, G.M., and McKenzie, L.J., 2012. Tropical seagrass meadows modify seawater carbon chemistry: implications for coral reefs impacted by ocean acidification. Environmental Research Letters, 7(2): 024026.
Usseglio-Polatera, P., Bournaud, M., Richoux, P., and Tachet, H., 2000. Biomonitoring through biological traits of benthic macroinvertebrates: how to use species trait databases? Hydrobiologia, 422/423: 153-162.
Wahl, M., Schneider Covachã, S., Saderne, V., Hiebenthal, C., Müller, J.D., Pansch, C., and Sawall, Y., 2018. Macroalgae may mitigate ocean acidification effects on mussel calcification by increasing pH and its fluctuations. Limnology and Oceanography, 63: 3-21.
Wei, T and Simko, V., 2017. R package "corrplot": Visualization of a Correlation Matrix (Version 0.84). Available from https://github.com/taiyun/corrplot
Wilcox, M., and Jeffs, A., 2017. Is attachment substrate a prerequisite for mussels to establish on soft-sediment substrate? Journal of Experimental Marine Biology and Ecology, 495: 83-88.
Yakovis, E., and Artemieva, A. 2017. Cockles, barnacles and ascidians compose a subtidal facilitation cascade with multiple hierarchical levels of foundation species. Nature Scientific Reports, 7 (237): 1-11.
Zeileis, A., and Hothorn, T. 2002. Diagnostic Checking in Regression Relationships. R News 2(3), 7-10. https://CRAN.R-project.org/doc/Rnews
APPENDIX 1: REFERENCES OF EACH BIOLOGICAL TRAIT FOR EACH SPECIES
Antoniadou, C., and Chintiroglou, C. 2006. Trophic relationships of polychaetes associated with different algal growth forms. Helgoland Marine Research, 60: 39-49.
Alexandridis, N., 2017. Models of general community assembly mechanisms simulating the spatial and temporal dynamics of benthic biodiversity. Thesis. Earth Sciences. Université de Bretagne occidentale: 1-150.
Bitlis, B., and Ozturk, B., 2016. Rissooidea species distributed along the Turkish levantine coast. Turkish Journal of Fisheries and Aquatic Sciences, 16: 443-454.
Cadien, D.B. 2004. Amphipoda of the Northeast Pacific (Equator to Aleutians, intertidal to abyss): VI. Corophioidea. Sanitation District of Los Angeles County: 1-42.
Fauchald, K., and Jumars, J.A., 1979. The diet of worms: a study of polychaete feeding guilds. Oceanography and Marine Biology: An Annual Review 17:193-284.
Faulwetter, S., Markantonatou, V., Pavloudi, C., Papageorgiou, N., Keklikoglou, K., Chatzinikolaou, E., Pafilis, E., Chatzigeorgiou, G., Vasileiadou, K., Dailianis, T., Fanini, L., Koulouri, P., and Arvanitidis, C., 2014. Polytraits: A database on biological traits of marine polychaetes. Biodiversity Data Journal, 2: e1024.
Fish, J. and Fish, S., 2011. A Student's Guide To The Seashore. 2nd ed. Cambridge, UK: Cambridge University Press.
Fontaine, A.R. 1965. The feeding mechanisms of the ophiuroid Ophiocomina nigra. Journal of the Marine Biological Association of the United Kingdom, 45 (2): 373-385.
Foveau, A. 2009. Benthic Habitats and Communities of the Eastern Basin from the Handle: Condition at the Beginning of the 21st Century. Thesis, University of Lille Nord de France.
Gallucci, A. M., and Gambi, M. C. 2014. SYLLIS PROLIFERA KROHN, 1852 (ANNELIDA, SYLLIDAE): A STANDARDISED PROTOCOL FOR LABORATORY CULTURING OF A MODEL SPECIES. Technical Report.
Gibson, R.N., and Atkinson, R.J.A., 2001. Oceanography and Marine Biology: An annual review. Vol 39. Taylor and Francis.
Mackie, A.S.Y., 1991. Scalibregma celticum new species (Polychaeta: Scalibregmatidae) from Europe, with a redescription of Scalibregma inflatum Rathke, 1843 and comments on the genus Sclerobregma Hartman, 1965. Bulletin of Marine Science, 48 (2): 268-276.
MarLIN, 2006. BIOTIC - Biological Traits Information Catalogue. Marine Life Information Network. Plymouth: Marine Biological Association of the United Kingdom. [Cited Dec 2019]
Marlin.ac.uk. 2020. Marlin - The Marine Life Information Network.
Martins, R., Magalhães, L., Moreira, A., Peter, A., Martin, G.S., Rodrigues, A.M., and Quintino, V., 2013. Diversity, distribution and ecology of the family Syllidae (Annelida) in the Portuguese coast (Western Iberian Peninsula). Helgoland Marine Research, 67:775-788.
Marque, J.C., and Bellan-Santi, D., 1991. Gammaridea and Caprellidea (Crustacea - Amphipoda) of the Portuguese south-western continental shelf: taxonomy and distributional ecology. Bijdragen tot de Dierkunde, 61 (2): 65-87.
Mhc.jncc.gov.uk. 2020. Infralittoral Muddy Sand - JNCC Marine Habitat Classification
Morten, B., Cunha, R., and Martins, A., 2014. Species richness, relative abundance and dwarfism in Azorean bivalves: consequences of latitude, isolation or productivity? Or all three? Journal of the Marine Biological Association of the United Kingdom, 94(3): 567-578.
Myres, A. A., and Lowely, J. K. 2003. A phylogeny and a new classification of the corophiidea leach, 1814 (amphipoda). Journal of Crustacean Biology, 23(2): 443-485.
Navarro-Barranco, C., Tierno de Figueroa, J.M., Guerra-García, J.M., Sánchez-Tocino, L., and García-Gómez, J.C., 2013. Feeding habits of amphipods (Crustacea: Malacostraca) from shallow soft bottom communities: Comparison between marine caves and open habitats. Journal of Sea Research, 78: 1-7.
Nishi, E., Tanaka, K., Tovar-Hernández, M.A., and Giangrande, A., 2009. Dialychone, Jasmineira and Paradialychone (Annelida: Polychaeta: Sabellidae) from Japan and adjacent waters, including four new species descriptions. Zootaxa, 2167: 1-24.
Oliver, P.G., Holmes, A.M., Killeen, I.J. and Turner, J.A., 2016. Marine Bivalve Shells of the British Isles. Amgueddfa Cymru - National Museum Wales. Available from: http://naturalhistory.museumwales.ac.uk/britishbivalves [Accessed: 2 April 2020].
Palomares, M.L.D. and Pauly, D., 2019. SeaLifeBase. World Wide Web electronic publication. www.sealifebase.org, version (12/2019).
Parr, C.S., Wilson, N., Leary, P., Schulz, K.S., Lans, K., Walley, L., Hammock, J.A., Goddard, A., Rice, J., Studer, M., Holmes, J.T.G, and Corrigan, R.J., 2014. The Encyclopedia of Life v2: Providing Global Access to Knowledge About Life on Earth. Biodiversity Data Journal, 2: e1079.
Guerra-García, J.M., and de Figueroa, J.M.T., 2009. What do caprellids (Crustacea: Amphipoda) feed on? Marine Biology, 156 (9):1881-1890.
Guerra-García, J. M., Tierno de Figueroa, J.M., Navarro-Barranco, C., Ros, M., Sanchez-Moyano, J.E., and Moreira, J., 2014. Dietary analysis of the marine Amphipoda (Crustacea: Peracarida) from the Iberian Peninsula. Journal of Sea Research, 85: 508-517.
Pleijel, F., and Rouse, G.W., 2012. A revision of Nereimyra (Psamathini, Hesionidae, Aciculata, Annelida). Zoological Journal of the Linnean Society, 164(1): 36-51.
Queiros, A., Birchenough, S.N.R., Bremner, J., Godbold, J.A., Parker, R.E., Romero-Ramirez, A., Reiss, H., Solan, M., Somerfield, P.J., Colen, C.V., Hoey, G.V., and Widdicombe, S., 2013. A bioturbation classification of European marine infaunal invertebrates. Ecology and Evolution, 3(11): 3958-3985.
Ramey-Balci, P.A., and Bodnar, E., 2009. Selection by a deposit-feeding polychaete, Polygordius jouinae, for sands with relatively high organic content. Limnology and Oceanography, 54:1512-1520.
San Martin, G., and Lopez, E., 2000. Three new species of Syllis (Syllidae: Polychaete) from Iberian coasts. Cahiers de Biologie Marine, 41: 425-433.
Schnedl, S.M., Haselmair, A., Gallmetzer, I., Mautner, AK., Tomasovych, A., and Zuschin, M., 2018. Molluscan benthic communities at Brijuni Islands (northern Adriatic Sea) shaped by Holocene sea-level rise and recent human eutrophication and pollution. The Holocene, 28 (11): 1801-181.
Sejr, M. K., Sand, M.K., Jensen, K.T., Petersen, J.K., Christensen, P.B., and Rysgaard, S., 2002. Growth and production of Hiatella arctica (Bivalvia) in a high-Arctic fjord (Young Sound, Northeast Greenland). Marine Ecology Progress Series, 244: 163-169.
Species-identification.org. 2020. Marine Species Identification Portal.
Thiel, M. and Watling, L., 2015. Lifestyles and Feeding Biology. Oxford: Oxford University Press.
Tovar-Hernández, M.A., Licciano, M., and Giangrande, A., 2007. Revision of Chone Krøyer, 1856 (Polychaeta: Sabellidae) from the Eastern Central Atlantic and Mediterranean Sea with descriptions of two new species. Scientia Marina, 71 (2): 315-338.
APPENDIX 2: FUZZY CODES FOR LOCH ALSH SPECIES
|
- |
Bioturbation |
Bioturbation |
Bioturbation |
Bioturbation |
Bioturbation |
Bioturbation |
Body Size |
Body Size |
Body Size |
Body Size |
Body Size |
Body Size |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
- |
none |
biodiff |
upw |
down |
bioirr |
surfmod |
s<5 |
s5_10 |
s10_20 |
s20_40 |
s40_80 |
s>80 |
|
Ampelisca diadema |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Amphipholis squamata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
2.5 |
0.5 |
0 |
0 |
|
Aonides oxycephala |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Arcopagia crassa |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
|
Asclerocheilus intermedius |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
0.5 |
0.5 |
0 |
|
Aurospio banyulensis |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Balanus balanus |
3 |
0 |
0 |
0 |
0 |
0 |
0.5 |
2 |
0.5 |
0 |
0 |
0 |
|
Clytia hemisphaerica |
3 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Dipolydora caulleryi |
2 |
0 |
0 |
0 |
1 |
0 |
0 |
0.5 |
2 |
0.5 |
0 |
0 |
|
Eumida bahusiensis |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Eumida sanguinea |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Flabelligera affinis |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
0 |
|
Galathea nexa |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Gammaropsis maculata |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Glycera lapidum |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Golfingia (Golfingia) elongata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Golfingia (Golfingia) vulgaris vulgaris |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Harpinia crenulata |
0 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Hiatella arctica |
3 |
0 |
0 |
0 |
0 |
0 |
0.5 |
0.5 |
0.5 |
0.5 |
0.5 |
0.5 |
|
Hydroides elegans |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
2 |
1 |
0 |
|
Janira maculosa |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Janua pagenstecheri |
0 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Jasmineira elegans |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Kurtiella bidentata |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Lepidonotus squamatus |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Leptochiton asellus |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Leptochiton cancellatus |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Leucosolenia variabilis |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Limaria hians |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
|
Mediomastus fragilis |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Metaphoxus fultoni |
0 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Minuspio cirrifera |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Modiolus modiolus |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Modiolus modiolus/Modiolula phaseolina juv. |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Monia patelliformis |
3 |
0 |
0 |
0 |
0 |
0 |
0.6 |
0.6 |
0.6 |
0.6 |
0.6 |
0 |
|
Musculus subpictus |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Nereimyra punctata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Nucula nucleus |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Onoba semicostata |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Ophiocomina nigra |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Ophiopholis aculeata |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Ophiothrix fragilis |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Othomaera othonis |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Parvicardium pinnulatum |
0 |
0 |
0 |
0 |
0 |
3 |
1.25 |
1.25 |
0.5 |
0 |
0 |
0 |
|
Pholoe baltica |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Pholoe inornata |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Pione vastifica |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Polyophthalmus pictus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
|
Psamathe fusca |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Scalibregma celticum |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Scolelepis fuliginosa |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
|
Sphaerosyllis hystrix |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Sphaerosyllis taylori |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0.5 |
2 |
0.5 |
0 |
|
Spirobranchus triqueter |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
0 |
0 |
|
Syllis armillaris |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
2 |
1 |
0 |
|
Syllis columbretensis |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
2 |
1 |
0 |
|
Terebellides stroemii |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Testudinalia testudinalis |
3 |
0 |
0 |
0 |
0 |
0 |
0.75 |
0.75 |
0.75 |
0.75 |
0 |
0 |
|
Thracia distorta |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Timoclea ovata |
0 |
0 |
0 |
0 |
0 |
3 |
1 |
1 |
1 |
0 |
0 |
0 |
|
Trichobranchus glacialis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Trypanosyllis (Trypanosyllis) coeliaca |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Tubificoides amplivasatus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
2 |
0 |
0 |
|
Urothoe elegans |
0 |
0 |
0 |
0 |
0 |
3 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
- |
Feeding mode |
Feeding mode |
Feeding mode |
Feeding mode |
Feeding mode |
Feeding mode |
Life Span |
Life Span |
Life Span |
Life Span |
|---|---|---|---|---|---|---|---|---|---|---|
|
- |
dep |
susp |
opp |
graz |
pred |
<1yr |
1_2y |
3_5y |
6_10y |
>10y |
|
Ampelisca diadema |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Amphipholis squamata |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Aonides oxycephala |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Arcopagia crassa |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Asclerocheilus intermedius |
1.5 |
1.5 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Aurospio banyulensis |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Balanus balanus |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Clytia hemisphaerica |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Dipolydora caulleryi |
0.75 |
2 |
0 |
0.25 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Eumida bahusiensis |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
|
Eumida sanguinea |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
|
Flabelligera affinis |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Galathea nexa |
1.5 |
0 |
1.5 |
0 |
0 |
2 |
1 |
0 |
0 |
0 |
|
Gammaropsis maculata |
0 |
3 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Glycera lapidum |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Golfingia (Golfingia) elongata |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Golfingia (Golfingia) vulgaris vulgaris |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Harpinia crenulata |
1.5 |
0 |
1.5 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Hiatella arctica |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Hydroides elegans |
0 |
3 |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
0 |
|
Janira maculosa |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Janua pagenstecheri |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Jasmineira elegans |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Kurtiella bidentata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Lepidonotus squamatus |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
3 |
0 |
0 |
0 |
|
Leptochiton asellus |
0 |
0 |
0 |
3 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
|
Leptochiton cancellatus |
0 |
0 |
0 |
3 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
|
Leucosolenia variabilis |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Limaria hians |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Mediomastus fragilis |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Metaphoxus fultoni |
1.5 |
0 |
1.5 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Minuspio cirrifera |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Modiolus modiolus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Modiolus modiolus/Modiolula phaseolina juv. |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Monia patelliformis |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Musculus subpictus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Nereimyra punctata |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Nucula nucleus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Onoba semicostata |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Ophiocomina nigra |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ophiopholis aculeata |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ophiothrix fragilis |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Othomaera othonis |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Parvicardium pinnulatum |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0.5 |
2.5 |
0 |
|
Pholoe baltica |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
|
Pholoe inornata |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
|
Pione vastifica |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Polyophthalmus pictus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Psamathe fusca |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
|
Scalibregma celticum |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Scolelepis fuliginosa |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Sphaerosyllis hystrix |
1.5 |
0 |
0 |
0 |
1.5 |
0 |
3 |
0 |
0 |
0 |
|
Sphaerosyllis taylori |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Spirobranchus triqueter |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Syllis armillaris |
0 |
0 |
0 |
1.5 |
1.5 |
3 |
0 |
0 |
0 |
0 |
|
Syllis columbretensis |
0 |
0 |
0 |
1.5 |
1.5 |
3 |
0 |
0 |
0 |
0 |
|
Terebellides stroemii |
3 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
0 |
|
Testudinalia testudinalis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Thracia distorta |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Timoclea ovata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
|
Trichobranchus glacialis |
3 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
0 |
|
Trypanosyllis (Trypanosyllis) coeliaca |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
|
Tubificoides amplivasatus |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
1 |
2 |
0 |
0 |
|
Urothoe elegans |
3 |
0 |
0 |
0 |
0 |
2.5 |
0.5 |
0 |
0 |
0 |
|
- |
Substrate - gravel |
Substrate - coarse sand |
Substrate - sand |
Substrate - rock |
Substrate - muddy sand |
Substrate - mud |
Substrate - pelagic |
|---|---|---|---|---|---|---|---|
|
Ampelisca diadema |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Amphipholis squamata |
1 |
1 |
1 |
0 |
0 |
0 |
0 |
|
Aonides oxycephala |
0.6 |
0.6 |
0.6 |
0 |
0.6 |
0.6 |
0 |
|
Arcopagia crassa |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Asclerocheilus intermedius |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
|
Aurospio banyulensis |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Balanus balanus |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Clytia hemisphaerica |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Dipolydora caulleryi |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Eumida bahusiensis |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Eumida sanguinea |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Flabelligera affinis |
0 |
0 |
0 |
1 |
1 |
1 |
0 |
|
Galathea nexa |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Gammaropsis maculata |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Glycera lapidum |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Golfingia (Golfingia) elongata |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Golfingia (Golfingia) vulgaris vulgaris |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Harpinia crenulata |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
|
Hiatella arctica |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Hydroides elegans |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Janira maculosa |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Janua pagenstecheri |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Jasmineira elegans |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Kurtiella bidentata |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
|
Lepidonotus squamatus |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Leptochiton asellus |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Leptochiton cancellatus |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Leucosolenia variabilis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Limaria hians |
1.5 |
0 |
0 |
0 |
1.5 |
0 |
0 |
|
Mediomastus fragilis |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Metaphoxus fultoni |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
|
Minuspio cirrifera |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Modiolus modiolus |
0.6 |
0 |
0.6 |
0.6 |
0.6 |
0.6 |
0 |
|
Modiolus modiolus/Modiolula phaseolina juv. |
0.6 |
0 |
0.6 |
0.6 |
0.6 |
0.6 |
0 |
|
Monia patelliformis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Musculus subpictus |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Nereimyra punctata |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Nucula nucleus |
1.5 |
0 |
1.5 |
0 |
0 |
0 |
0 |
|
Onoba semicostata |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Ophiocomina nigra |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Ophiopholis aculeata |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Ophiothrix fragilis |
0.75 |
0.8 |
0.75 |
0.75 |
0 |
0 |
0 |
|
Othomaera othonis |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Parvicardium pinnulatum |
1 |
0 |
1 |
0 |
1 |
0 |
0 |
|
Pholoe baltica |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Pholoe inornata |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Pione vastifica |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Polyophthalmus pictus |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Psamathe fusca |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Scalibregma celticum |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Scolelepis fuliginosa |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Sphaerosyllis hystrix |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Sphaerosyllis taylori |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Spirobranchus triqueter |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Syllis armillaris |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Syllis columbretensis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Terebellides stroemii |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Testudinalia testudinalis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Thracia distorta |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Timoclea ovata |
1.5 |
0 |
1.5 |
0 |
0 |
0 |
0 |
|
Trichobranchus glacialis |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Trypanosyllis (Trypanosyllis) coeliaca |
0 |
0 |
0.75 |
0.75 |
0.75 |
0.75 |
0 |
|
Tubificoides amplivasatus |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Urothoe elegans |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
APPENDIX 3: FUZZY CODES FOR FETLAR TO HAROLDSWICK SPECIES
|
- |
Bioturbation none |
Bioturbation biodiff |
Bioturbation gall |
Bioturbation upw |
Bioturbation down |
Bioturbation bioirr |
Bioturbation surfmod |
Body Size s<5 |
Body Size s5_10 |
Body Size s10_20 |
Body Size s20_40 |
Body Size s40_80 |
Body Size s>80 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Alvania beanii |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Amphipholis squamata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
2.5 |
0.5 |
0 |
0 |
|
Aonides paucibranchiata |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Capitella capitata |
0 |
0.5 |
0 |
2 |
0 |
0.5 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Caprella acanthifera |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Ceradocus semiserratus |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Chaetozone setosa |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Chone filicaudata |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Clausinella fasciata |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0.75 |
0.75 |
0.75 |
0.75 |
0 |
0 |
|
Crassicorophium bonnellii |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Dialychone dunerificta |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Echinocyamus pusillus |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
1 |
1 |
1 |
0 |
0 |
0 |
|
Galathea intermedia |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Gammaropsis lobata |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Gari tellinella |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Gibbula tumida |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Glycera lapidum |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Goodallia triangularis |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Gouldia minima |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Hiatella arctica |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0.5 |
0.5 |
0.5 |
0.5 |
0.5 |
0.5 |
|
Janira maculosa |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Leptochiton asellus |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Maera othonis |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Mediomastus fragilis |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Minuspio cirrifera |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Modiolus modiolus |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Nematoda spp. |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Nemertea spp. |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
|
Nephasoma (Nephasoma) minutum |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Onoba semicostata |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Ophiocomina nigra |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Ophiopholis aculeata |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Phtisica marina |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
1 |
1 |
1 |
0 |
0 |
0 |
|
Pisione remota |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0.75 |
0.75 |
0.75 |
0.75 |
0 |
0 |
|
Polygordius |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Polynoidae sp. |
0.5 |
2.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Pomatoceros triqueter |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Psamathe fusca |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Socarnes erythrophthalmus |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Socarnes filicornis |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Sphaerosyllis bulbosa |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Spirobranchus triqueter |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1 |
1 |
1 |
0 |
0 |
|
Syllis pontxioi |
0 |
0 |
0 |
0 |
0 |
|
3 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Thracia villosiuscula |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Timoclea ovata |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
1 |
1 |
1 |
0 |
0 |
0 |
|
Trypanosyllis coeliaca |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Tubificoides sp. |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
2 |
0 |
0 |
|
Vaunthompsonia cristata |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
- |
Feeding Mode |
Feeding Mode |
Feeding Mode |
Feeding Mode |
Feeding Mode |
Feeding Mode |
Life Span |
Life Span |
Life Span |
Life Span |
Life Span |
|---|---|---|---|---|---|---|---|---|---|---|---|
|
- |
dep |
susp |
detritus |
opp |
graz |
pred |
<1yr |
1_2y |
3_5y |
6_10y |
>10y |
|
Alvania beanii |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Amphipholis squamata |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Aonides paucibranchiata |
3 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Capitella capitata |
3 |
0 |
0 |
0 |
0 |
0 |
1 |
2 |
0 |
0 |
0 |
|
Caprella acanthifera |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Ceradocus semiserratus |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Chaetozone setosa |
1.5 |
0 |
1.5 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Chone filicaudata |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Clausinella fasciata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
|
Crassicorophium bonnellii |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Dialychone dunerificta |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Echinocyamus pusillus |
1 |
0 |
1 |
0 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Galathea intermedia |
0 |
3 |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
0 |
0 |
|
Gammaropsis lobata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Gari tellinella |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Gibbula tumida |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Glycera lapidum |
0 |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Goodallia triangularis |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Gouldia minima |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Hiatella arctica |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Janira maculosa |
1.5 |
0 |
0 |
0 |
1.5 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Leptochiton asellus |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
|
Maera othonis |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Mediomastus fragilis |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Minuspio cirrifera |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Modiolus modiolus |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Nematoda spp. |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Nemertea spp. |
0 |
0 |
0 |
0.5 |
0 |
2.5 |
0 |
0 |
0 |
0 |
0 |
|
Nephasoma (Nephasoma) minutum |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Onoba semicostata |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Ophiocomina nigra |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ophiopholis aculeata |
1.5 |
0 |
0 |
0 |
0 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Phtisica marina |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Pisione remota |
0 |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
3 |
0 |
0 |
|
Polygordius sp. |
3 |
0 |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
|
Polynoidae sp. |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
3 |
0 |
0 |
|
Pomatoceros triqueter |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Psamathe fusca |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
|
Socarnes erythrophthalmus |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Socarnes filicornis |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Sphaerosyllis bulbosa |
1 |
0 |
1 |
0 |
0 |
1 |
0 |
3 |
0 |
0 |
0 |
|
Spirobranchus triqueter |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Syllis pontxioi |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
3 |
0 |
0 |
0 |
|
Thracia villosiuscula |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Timoclea ovata |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
0 |
|
Trypanosyllis coeliaca |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0 |
0 |
0 |
|
Tubificoides sp. |
1.5 |
0 |
0 |
0 |
1.5 |
0 |
0 |
1 |
2 |
0 |
0 |
|
Vaunthompsonia cristata |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
- |
Substrate - gravel |
Substrate - coarse sand |
Substrate - sand |
Substrate - rock |
Substrate -muddy sand |
Substrate - mud |
Substrate - other |
|---|---|---|---|---|---|---|---|
|
Alvania beanii |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
|
Amphipholis squamata |
1 |
1 |
1 |
0 |
0 |
0 |
0 |
|
Aonides paucibranchiata |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Capitella capitata |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Caprella acanthifera |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Ceradocus semiserratus |
1 |
1 |
0 |
1 |
0 |
0 |
0 |
|
Chaetozone setosa |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Chone filicaudata |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
|
Clausinella fasciata |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Crassicorophium bonnellii |
0 |
0 |
0.75 |
0.75 |
0.75 |
0.75 |
0 |
|
Dialychone dunerificta |
0 |
0 |
1.5 |
0 |
1.5 |
0 |
0 |
|
Echinocyamus pusillus |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Galathea intermedia |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Gammaropsis lobata |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Gari tellinella |
0.75 |
0.75 |
0.75 |
0 |
0.75 |
0 |
0 |
|
Gibbula tumida |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Glycera lapidum |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Goodallia triangularis |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Gouldia minima |
0 |
1 |
1 |
0 |
1 |
0 |
0 |
|
Hiatella arctica |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Janira maculosa |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Leptochiton asellus |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Maera othonis |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Mediomastus fragilis |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
|
Minuspio cirrifera |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
|
Modiolus modiolus |
0.6 |
0 |
0.6 |
0.6 |
0.6 |
0.6 |
0 |
|
Nematoda spp. |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Nemertea spp. |
0 |
0 |
1 |
1 |
1 |
0 |
0 |
|
Nephasoma (Nephasoma) minutum |
0 |
0 |
0 |
0 |
1.5 |
1.5 |
0 |
|
Onoba semicostata |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Ophiocomina nigra |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Ophiopholis aculeata |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Phtisica marina |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Pisione remota |
0 |
3 |
0 |
0 |
0 |
0 |
0 |
|
Polygordius sp. |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Polynoidae sp. |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Pomatoceros triqueter |
1.5 |
0 |
0 |
1.5 |
0 |
0 |
0 |
|
Psamathe fusca |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Socarnes erythrophthalmus |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Socarnes filicornis |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
|
Sphaerosyllis bulbosa |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Spirobranchus triqueter |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
|
Syllis pontxioi |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
|
Thracia villosiuscula |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
0 |
|
Timoclea ovata |
1.5 |
0 |
1.5 |
0 |
0 |
0 |
0 |
|
Trypanosyllis coeliaca |
0 |
0 |
0.75 |
0.75 |
0.75 |
0.75 |
0 |
|
Tubificoides sp. |
0 |
0 |
0 |
0 |
3 |
0 |
0 |
|
Vaunthompsonia cristata |
0 |
1.5 |
1.5 |
0 |
0 |
0 |
0 |
APPENDIX 4: SAMPLE DETAILS
|
Station name |
Location |
Sample Type |
No of replicates |
Date |
Depth (m bcd) |
Original station ID |
Survey name |
Latitude |
Longitude |
|---|---|---|---|---|---|---|---|---|---|
|
West Flame shell bed (W FS) |
Loch Alsh |
Grabs |
3 |
25/03/2014 25/03/2014 26/03/2014 |
6 10 10 |
G01 G03 GEX7 |
2014 Marine Scotland Loch Alsh benthic survey - infaunal data |
57.277528 57.275962 57.281666 |
-5.753962 -5.749504 -5.762287 |
|
East Flame Shell bed (E FS) |
Loch Alsh |
Cores |
4 |
10/08/2012 |
16.5 |
FS01 |
2012 Marine Scotland benthic survey of Lochs Alsh, Duich, Creran and Fyne |
57.278602 |
-5.704505 |
|
Flame Shell bed and Horse Mussel bed 1 (FS & HM 1) |
Loch Alsh |
Cores |
3 |
08/08/2012 |
19 |
HM01 |
2012 Marine Scotland benthic survey of Lochs Alsh, Duich, Creran and Fyne |
57.272978 |
-5.714097 |
|
Flame Shell bed and Horse Mussel bed 2 (FS & HM 2) |
Loch Alsh |
Cores |
4 |
11/08/2012 |
15 |
HM02 |
2012 Marine Scotland benthic survey of Lochs Alsh, Duich, Creran and Fyne |
57.276917 |
-5.732836 |
|
Basta Voe Horse Mussel Bed (BV HM) |
Fetlar to Haroldswick |
Cores |
4 |
02/09/2012 |
10 |
BVMNCR1 |
2012 Marine Scotland Fetlar to Haroldswick benthic MPA search feature survey |
60.631600 |
-0.994417 |
|
Hascosay Horse Mussel Bed (HS HM) |
Fetlar to Haroldswick |
Cores |
4 |
04/09/2012 |
8 |
HSMNCR3 |
2012 Marine Scotland Fetlar to Haroldswick benthic MPA search feature survey |
60.618100 |
-1.007570 |
|
Hascosay Maerl Bed (HS MB) |
Fetlar to Haroldswick |
Cores |
4 |
04/09/2012 |
8 |
H |
2012 Marine Scotland Fetlar to Haroldswick benthic MPA search feature survey |
60.616900 |
-1.009870 |
|
North Hascosay Maerl Bed (NH MB) |
Fetlar to Haroldswick |
Cores |
4 |
03/09/2012 |
14 |
NH |
2012 Marine Scotland Fetlar to Haroldswick benthic MPA search feature survey |
60.621900 |
-0.993283 |
|
Uyea Sound Horse Mussel Bed (US HM) |
Fetlar to Haroldswick |
Cores |
4 |
03/09/2012 |
27 |
USM |
2012 Marine Scotland Fetlar to Haroldswick benthic MPA search feature survey |
60.667000 |
-0.944023 |
|
Wyre Sound Maerl Bed (W MB) |
Orkney |
Cores |
4 |
03/09/2011 |
0-5* |
W4 |
2011 Marine Scotland benthic survey |
59.126 |
-2.9899 |
|
Eday Sound Maerl Bed (E MB) |
Orkney |
Cores |
4 |
05/09/2011 |
5-10* |
E15 |
2011 Marine Scotland benthic survey |
59.127 |
-2.6829 |
|
Veantrow Bay Maerl Bed (V MB) |
Orkney |
Cores |
4 |
02/09/2011 |
0-5* |
K21 |
2011 Marine Scotland benthic survey |
59.067 |
-2.8876 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
21/07/2015 |
22 |
GM4 |
2015 Sound of Barra grab sampling survey |
56.98456 |
-7.35684 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
21/07/2015 |
20 |
GM3 |
2015 Sound of Barra grab sampling survey |
57.01891 |
-7.29241 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
21/07/2015 |
21 |
GM7 |
2015 Sound of Barra grab sampling survey |
57.0433 |
-7.27759 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
21/07/2015 |
26 |
GM11 |
2015 Sound of Barra grab sampling survey |
57.06539 |
-7.26738 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
21/07/2015 |
21 |
GM1 |
2015 Sound of Barra grab sampling survey |
57.08396 |
-7.24494 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
21/07/2015 |
26 |
GM15 |
2015 Sound of Barra grab sampling survey |
57.0892 |
-7.21962 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
22/07/2015 |
7.8 |
GM8 |
2015 Sound of Barra grab sampling survey |
57.06719 |
-7.35454 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
22/07/2015 |
27 |
GM12 |
2015 Sound of Barra grab sampling survey |
57.06832 |
-7.24133 |
|
Sound of Barra Grab (BS) |
Sound of Barra |
Grabs |
3 |
22/07/2015 |
20 |
GM13 |
2015 Sound of Barra grab sampling survey |
57.03746 |
-7.31943 |
*cores collected at one depth per site, but depth information is approximate only because it was derived from charts in retrospect.






