Interim Guidance on apportioning impacts from marine renewable developments to breeding seabird populations in SPAs
A key element of impact assessments for proposed marine renewable (offshore wind, wave and tidal) energy sites is the possible impact that such developments will have on breeding seabird populations.
Contents
- Introduction
-
Scope of this paper
- Future Approaches to Apportioning
- A theoretical approach
- Foraging Range
- Weighting by colony size
- Weighting by distance from the colony
- Weighting by sea area
- Combining the three weighting factors
- Example Table 1
- Disadvantages of the theoretical approach
- Advantages of the theoretical approach
- Requirements for theoretical apportioning impacts to SPAs
- Data led empirical approaches
- Tagging studies (GPS equipped loggers or satellite tags)
- Field Surveys to Support Theoretical and Empirical Methods
- Integrating Theoretical and Empirical approaches
- Concluding remarks
- References
Introduction
A key element of impact assessments for proposed marine renewable (offshore wind, wave and tidal) energy sites is the possible impact that such developments will have on breeding seabird populations. Many of the seabirds using marine renewables sites will breed in protected areas, including Special Protection Areas (SPAs). The questions which then follow are: from which SPAs do these birds originate and in what proportion?
This guidance is based on original work by Mark Trinder (MacArthur Green) and Andy Douse (SNH). It focuses solely on effects on seabirds during the breeding season. The Statutory Nature Conservation Bodies (SNCBs) are also considering issues surrounding the assessment of impacts on seabirds outwith the breeding season, which will lead to further guidance for developers and regulators.
Scope of this paper
We present a range of methods to determine connectivity with breeding seabird SPAs. These include both data-led empirical approaches and a theoretical approach for estimating how birds using marine renewables development sites can be ‘apportioned’ to multiple source colonies.
Future Approaches to Apportioning
Marine Scotland have been developing a tool to use for attributing birds at sea to origin colonies. This approach uses seabird density predictions based on the work of Ewan Wakefield an others (Wakefield et al 2017). These predictive maps are available for four seabird species Shag, Common Guillemot, Kittiwake and Razorbill. When the outputs of this work are available the approach outlined in this guidance may no longer be relevant, at least for those species.
A theoretical approach
Connectivity to SPAs is largely based on determining seabird foraging ranges. This theoretical approach to apportioning uses published seabird foraging range information and generalised models. Empirically derived approaches (see below) use site-specific field-derived data on foraging ranges and locations. In the absence of sufficient survey or tagging data, theoretical approaches are the only option.
The simplest approach, that has been adopted at some inland development sites, is to assess impacts against each SPA within foraging range separately and in turn, assuming each time that all the birds within the development site originate from the SPA in question. For all its simplicity, this approach is fundamentally flawed in that impacts will inevitably be overestimated at every SPA and particularly at SPA sites with relatively small species populations. We do not recommend this approach in the marine environment where large foraging ranges mean that multiple SPAs will often be included in such an analysis.
In this theoretical approach, as available data are currently limited, we make some necessary and very general assumptions. We acknowledge that the resulting picture of seabird distribution and habitat use at sea is not accurate, but the approximations are aimed at creating a workable tool for the assessment of impacts on SPAs. Our aim is to present a practical solution that has an agreed basis and can be widely adopted. In future we hope that further data will be available to help refine this approach.
The method is based on foraging range and three colony-specific weighting factors:
(i) Colony size (in individuals);
(ii) Distance of colony from the development site1; and
(iii) Sea area (the areal extent of the open sea within the foraging range of the relevant species).
Foraging Range
Breeding seabirds are central place foragers i.e. they have to leave the colony (the central place) and disperse to find food before returning to the breeding colony. There is an energetic and time advantage in feeding as close to the colony as possible (all else being equal) and a limit to how far birds will travel, as beyond this distance a bird becomes unable to find enough food fast enough to bring back to the nest in order to breed successfully. This suggests that there is a limit to the distance travelled which will be approximated by the maximum recorded foraging range for that species (in that location). As seabirds generally avoid crossing any substantial land-mass the effective distance from colony should be measured as the ‘by-sea’ route.
To determine the SPAs for which there may be connectivity (i.e. which are within foraging range), we recommend using the single mean maximum value from Table 1 of Thaxter et al 2012 (which itself is a review of information across many studies available at the time). In some cases, where no estimate of foraging range for a species has yet been published, it may be justifiable to use values from a related species with strong ecological similarities when those are available.
The HRA of the Sectoral Plans included a systematic review of foraging ranges for seabirds in Scottish SPAs. The summary report is online. Table E1 of Appendix E in the report gives value of maximum ranges for seabird species which may be useful in some cases.
The amount of information on seabird foraging range is expanding rapidly due to the increasing numbers of tracking studies (e.g. the FAME2 project) using technology such as GPS tags. These studies provide temporally and spatially explicit data on the ranging and behaviour of key species. Although at present we advise use of the ranges published in Thaxter et al 2012, we recommend that developers conduct a thorough review of all the available data and contact SNH seabird specialists to agree appropriate foraging ranges if newer data are available.
Weighting by colony size
Large colonies will contribute more individuals to the number of birds found within a development site, all other factors being equal. Population size is best measured in numbers of individuals (which can be converted from breeding pairs or apparently occupied site). As long as count units are consistent between colonies, the actual unit is not particularly important, as weighting values are relative.
All colonies being considered should have been counted concurrently or at the very least at a similar time. A suitable reference point for most species will be Seabird 2000 data, and though we acknowledge that these data are now relatively old, at present this is the recommended dataset. A new seabird census is planned and will provide newer data in a few years’ time. Where count data are old , species-specific change metrics can be used to correct population counts to approximate the current likely population sizes but there must be clear justification for doing so. Such change metrics will usually be based on regional population trends, so the resultant weighting factor is insensitive to absolute values of population size.
For colonies that occupy long stretches of coastline (and may consist of separate ‘sub-sections’) a single weighting factor may not be appropriate. In such cases different sub-sections may be treated as separate colonies. Most seabird colonies will have separate count sections (see Seabird Colony Register)3. If a single SPA is made up of several sub sections or colonies the combined SPA impact can be reconstructed after the weighting for each sub section is completed.
The birds using proposed development areas will often include a mix of birds from SPA and non-SPA colonies. Non-SPA birds must be included in the analyses; otherwise impacts on SPA breeding birds will be exaggerated. This will require knowledge of the sizes of non-SPA as well as SPA colonies in the area of search.
If the apportioning is to relate to breeding adults only, the proportion of adult birds that do not breed in any one year (so called breeding sabbaticals) may be taken into account if known. More significantly, there will be a proportion of non-breeding age birds present in the proposed development area, and this will require determination of the age of seabirds during field surveys of the site. Where birds cannot be aged in the field, published life-tables may be used to estimate of the expected proportion of adults different age classes. Assessors should be aware that for some species, sub-adult and immature age classes may use different geographical areas to those frequented by adult birds. A good knowledge of seabird ecology is fundamental to such analyses.
Weighting by distance from the colony
The distance of the colony should be measured as the distance between the geometric centre of the development to the geometric centre of the colony. We acknowledge that colonies with complex boundaries might be more sensibly measured from a different location, which should be specified. As an example if it is known that a species has a strongly skewed distribution within a large SPA it might be more suitable to measure from the centre of that distribution. A cautious approach would be to use the ‘nearest boundary to nearest boundary’ for an individual SPA, although this would then have impacts on the apportioning to other sites being considered.
Development sites that are distant from a seabird colony/s might be expected to have fewer birds on them from that colony than development sites close to a colony/SPA. Thus the contribution of any particular colony to the number of birds seen at a given development site will be inversely related to distance between the development site and the colony.
As birds radiate out from a colony density will decrease by a factor proportional to 1/distance2 as area increases proportionally by π.r2. For simplicity a weighting factor based on 1/distancei2 is used here. The foraging range should be applied to the by-sea or sea route that birds would take to reach foraging locations. This prevents the implausible situation where species (particularly with very large foraging ranges e.g Gannet, Fulmar) from a colony on the west coast would be expected to be found at an east coast location by traversing the land between.
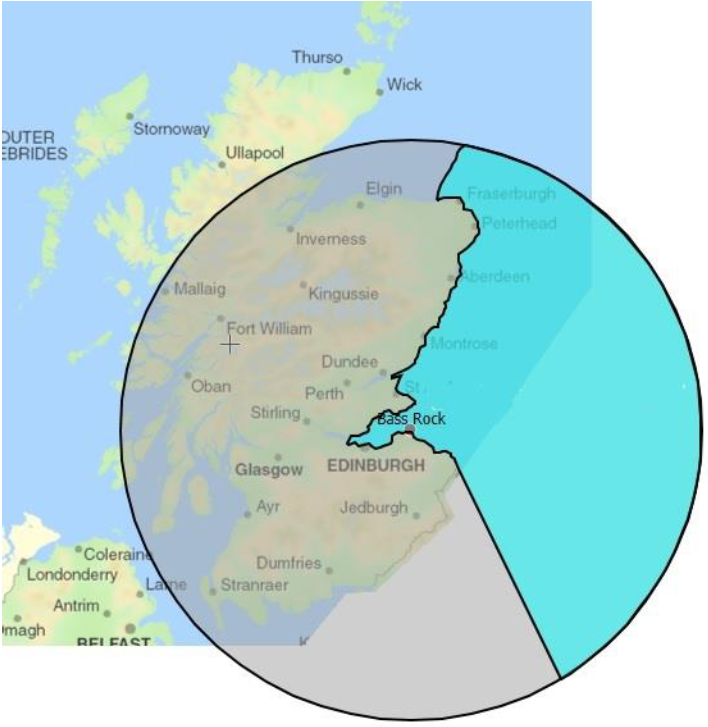
Figure 1. Illustration of effect of sea-route foraging range versus ‘straight-line’ foraging range. The blue area is the true area of sea accessible to a species with a foraging range of 230km from Bas Rock, whereas the grey circle shows the result of applying the straight-line distance, which includes sea areas on the west coast which would not be accessible.
Weighting by sea area
We suggest that a correction is also made for the available sea area. The logic behind this is that for a colony on an island, seabird foragers may have all 360o of marine habitat to forage over, whereas for a mainland colony, birds may only have about 180o of marine habitat. The available sea area can be measured by plotting a circle defined by species-specific foraging range round the colony in GIS and calculating the area of sea available to each seabird species. The fraction of the disc centred on the colony that is occupied by sea surface is then expressed as a decimal. As the density of birds will increase as the area of available forage area decreases this should be used in the formula as 1/estimated area.
We acknowledge that seabirds are very unlikely to be distributed evenly across all the available sea area; the distribution will be patchy and likely related to prey availability. However without specific evidence and in the interests of not overcomplicating any estimation this theoretical approach cannot provide finer scale differentiation.
Combining the three weighting factors
The weighting factors should be combined to produce an overall weighting. Each factor is given equal weight in the combined weighting, as there appears to be no compelling reason or specific evidence for any other treatment.
Example Table 1
This illustrates the apportioning approach for three colonies A, B and C for an imaginary bird species. Data on populations, distances from a theoretical development site and sea areas are given in the table, along with the resulting weightings and the combined weighting. Here the population considered is number of breeding adults - immature birds will not be included. As noted above if necessary the development site counts should be adjusted to account for this. In this case the maximum number of birds on the development site is 725 birds. 225 are aged as sub-adult or immature, with 500 birds in adult plumage. The calculations estimate the contribution of each SPA to this total.
A table using and embedded excel spread sheet showing these calculations is presented below. Double click on the table to edit values of the SPA population count, distance to the development and the proportion of the foraging area that is sea to change the value of weighting and the resultant calculation of the number of birds allocated to each SPA on the development site. (This function not available in pdf format version of this document use the following table).
| SPA name | Count of adult birds on SPA | Distance from SPA to development | 1/Proportion of Forage Range as Sea | Resulting Weight for SPA | Proportional Weight of SPA | Adult birds from each SPA on development site |
|---|---|---|---|---|---|---|
| A | 5000 | 10 | 2.000 | 2.905 | 0.769 | 385 |
| B | 6000 | 20 | 1.667 | 0.726 | 0.192 | 96 |
| C | 10000 | 50 | 1.250 | 0.145 | 0.038 | 19 |
| SUM | 21000 | 80 | 4.917 | 3.777 | 1.000 | 500 |
The calculation is made as follows:
Weight = (Colony Population / Sum of Populations) * (Sum of Distance2 / Colony Distance2) * (1/Colony Sea Proportion / Sum of 1/Sea Proportions).
(Note that the second part of the calculation (distance) is inverted as there is an inverse relationship with distance, following the inverse-square law.)
In the first line of the table (for colony A) the figures are:
(5000/21000)*(3000/100)*((2.0)/4.197) = 2.905
The weighting is then used to calculate the proportion of birds attributed to that SPA (SPA Prop) by calculating (SPA weight / sum of all weights) – in this case 2.905/3.777 = 0.769, this proportion is then used to calculate the number of birds on the site (multiply the total number on site by the proportion allocated to this SPA) 0.769 * 500 = 385 birds.
Disadvantages of the theoretical approach
This theoretical approach is a great simplification of a complex set of factors. It involves assumptions that are unlikely to be a good reflection of reality, but which have the merit of being simple and applicable through an easily understood working model. This approach offers a logical, potentially standard, basis for estimating the relevant contribution of birds from differing source colonies to the numbers seen in a given area where development(s) may be proposed. We suggest this approach is used until a more complex, evidence-based model is developed and agreed.
Some of the simplifications that underlie this calculation are:
- It assumes that birds are evenly distributed at sea. This is extremely unlikely, when key prey species (e.g. lesser sandeel) are known to be patchily distributed. Indeed, tracking studies have shown that some seabirds will travel very long distances, to particular foraging locations. In most studies the distributions of birds and prey in the area under investigation will be unknown.
- It assumes that seabird colonies are ‘independent’ of one another. There is some evidence that seabird colonies may ‘interfere’ with one another, thus affecting foraging range (Furness & Birkhead 1984, Cairns 1989, Wakefield et al 2012) but taking this into account for the purpose of apportioning impacts arising from marine renewables developments is likely to be impractical. Studies which show this effect are few or have small samples of tracked birds, although the theory is quite feasible. There is some contrary evidence that birds from different adjacent colonies can share foraging areas (Dean et al 2013) or a hybrid of separate areas and overlapping areas (Ainsley et al 2004).
- Seabirds breeding at large colonies may have larger foraging ranges, due in part to competition and prey depletion near to the colony. This effect, known as the Storer-Ashmole’s Halo (Elliot et al. 2009), has rarely been demonstrated in practice, though it has a sound theoretical basis (Gaston et al 2007). It is not known whether there is evidence for this from current bird-tagging studies, although something similar does appear to be found in bumble bees (Goulson and Osborne 2009). At present we regard it as impractical to factor Storer-Ashmole’s Halo into this analysis.
Advantages of the theoretical approach
- Seabird foraging ranges can vary considerably within and between years. The theoretical approach represents an ‘averaging’ over a long time period and across sites, in a way that tagging studies and other empirical approaches do not.
- The relative simplicity of the data needed and their application provides cost savings over empirical approaches
- It provides a mechanism for considering all species, from all relevant colonies even when the number of colonies to be considered is large, in which case empirical approaches are unlikely to be viable.
Requirements for theoretical apportioning impacts to SPAs
In summary to undertake this analysis the following is required.
- Use of proposed development site by seabirds, obtained from field survey (should such data not already exist)
- Agreed foraging range value for all species of interest (from which the proportion of sea area within foraging range will be calculated)
- Concurrent population estimates (in numbers of individuals) for all seabird colonies within foraging ranges
- Accurate determination of distances from colonies to proposed development site
Data led empirical approaches
In principle it is advantageous to apply field data for a specific development to help to determine the origin of birds within the development boundary. There are several methods of collecting data and supporting evidence. The results could be used jointly with the theoretical approach in some cases.
Tagging studies (GPS equipped loggers or satellite tags)
Data from birds tagged4 at different colonies can be used to estimate the proportion of birds from each colony using proposed marine renewable development sites. There is a need to correct for sample sizes of birds tagged from each colony, and to weight for the population size of each colony. Observations can also be weighted by time spent in the development site. We have already noted that seabird foraging ranges can vary considerably within and between years. The theoretical approach overcomes this by using ‘typical’ ranges derived from many studies over a long time period and across sites. To overcome this variation with site specific data would require a large number of tagged birds, from concurrent studies at all (or most) colonies under consideration, at each stage of the breeding season and over a number of years with considerable analysis costs. Almost all studies use only a relatively small number of tags, often deployed over a limited timescale. Most are behavioural or ecological studies, not strictly targeting the questions of apportioning origins of birds.
Tagging studies have a number of significant advantages over the theoretical method:
- Assumptions about how far birds travel, or indeed where they travel to, do not have to be made.
- Weighting by time spent in the development area is possible for GPS tagged birds.
- Time spent in the development area (and potentially elsewhere) can lead to estimates of turnover.
- Data can be analysed at smaller timescales than across a whole breeding season, depending on the quantity available. This means that the ‘contribution’ each SPA makes to birds present within a proposed marine renewables development site can be calculated at different stages of the breeding cycle if sufficient data exist.
Disadvantages include:
- Capture and handling for tagging studies may cause considerable disturbance and stress to breeding birds. As a result, and due to the added burden of carrying a tag, tagged birds may not behave normally. Some species are less amenable to tagging than others (Baron et al 2010, Vandebeele et al 2011).
- Tagging studies may be difficult or even impossible to conduct safely at some colonies due to topography.
- The demanding resource requirements. For any study a sufficient number of birds must be tracked (of potentially multiple species) at each stage of the breeding season and across more than one year. Furthermore, when there are several colonies within foraging range, studies are needed from all SPA (and non-SPA) sites under consideration. In addition, tagging studies must be carried out by trained and experienced personnel.
- Tagging studies are likely to be viable only in situations where there are just a few colonies, each of which can be well-studied. In complex areas with multiple SPAs and other non-SPA colonies within range, tagging studies will be challenging and expensive to undertake.
Feeding locations can also be determined by triangulation of radio-tagged birds, though this technique quickly becomes unreliable as foraging distance increases.
Field Surveys to Support Theoretical and Empirical Methods
These survey results could be used to ‘sense check’ the theoretical apportioning, or to add support to tagging studies. As more tagging data are obtained these can be used to describe seabird abundance at sea. Longer attachment periods will bring the added benefit of being able to develop predictive models of densities beyond the breeding season.
a. Gradients of Survey density data
For surveys that cover very large areas (all surveys should cover a buffer area beyond the development site boundaries), then analyses of the gradient of bird densities away from the development site in the direction of each relevant seabird colony SPA (and non-SPA) may yield information on the origin of birds coming from each SPA. The premise is that if an SPA is ‘contributing’ birds to a marine renewables development site, then survey data extending beyond the development site boundary would be expected to show an increasing density of birds in the direction of the relevant colony or colonies. Aerial survey data could be suitable for such analyses, given the ability of planes to cover large areas of sea in short timescales. However, as aerial surveys are usually only samples based on flying transects, it is also probable that geospatial statistical techniques will be needed to underpin the analytical approach, in order to derive a two-dimensional density space.
Key advantages from such an approach are:
- The use of real survey data over a wide spatial area, some of which may have been gathered already as part of other survey work.
- It may be suitable for species that have very large foraging ranges
However, disadvantages include:
- The potential requirement for complex analytical statistical methods.
- The likelihood that, even if connectivity can be established with particular seabird colonies, assessing their relative contribution would be technically demanding and probably subject to a higher degree of uncertainty than from tracking data.
- The need for surveys to extend over a significantly greater area than the usual ‘development area plus buffer’. Although as an additional cost on top of an existing survey programme this would probably prove considerably less expensive than a separate tagging study.
For these reasons we do not consider that this technique can be used to allocate apportioning alone, however it does have a place in providing some evidence of origin of birds at sea.
We are not aware of this method being used in any practical situation. Developers wishing to develop an approach using this method should discuss this with SNH.
b. Flight line data
This makes use of the fact that foraging seabirds generally return to a colony along a direct route after successful foraging. Before the widespread use of cheap, recoverable GPS technology on birds, this was the only suitable technique. Flight lines may be observed visually and small radio-transmitters can provide better data on departure and arrival directions (with the advantage that radio tags don’t need to be retrieved unless there are welfare reasons. Establishing the proportion of birds returning to a colony from the direction of the development area will give an indication of:
- Whether they are likely to be using the development site (i.e. if birds do not return from the development site direction then this may not be an area used).
- The number or proportion of birds using the development site (or general area around the development site).
Flight directions of birds departing from a colony (heading off to forage) are less informative given that birds may have to spend time searching for a patchy and unreliable food resource. However, the recent use of boats (e.g. fast Rigid Inflatable Boats) to follow foraging terns has been very successful in establishing feeding areas for species that do not forage over long distances (Perrow et al. 2011).
Advantages of flight line data analyses are:
- That they may be relatively cheap to undertake, requiring little more than a cliff-top observer with a pair of binoculars and a compass for inshore developments.
- That a large volume of data can be gathered from multiple locations, across the breeding season and in a variety of weather conditions;
The technique also has significant disadvantages, in that:
- The direction of travel cannot identify the actual area over which birds were foraging, though combining directional data with time spent away from the colony by ringed or colour marked individuals, can (using flight speeds) identify potential foraging areas.
- Birds may forage over more than one area before returning to the colony, such that the direction of flight only indicates the last foraging location.
- Birds may not travel in a straight line back from a foraging area.
- Birds may gather in rafts on open water at some distance from the colony before returning to the colony (e.g. shearwaters), or head for loafing/preening/bathing sites before returning to the colony (e.g. kittiwakes at some colonies).
In theory this could produce similar results to tagging studies as flight line data should replicate data from tagged birds which indicate their origins, although surveys would need to identify popular feeding locations to achieve this . However in most cases this is the lowest quality data with large uncertainty attached to estimates of the connectivity to SPAs derived from these methods.
With Digital Aerial Surveys the direction of flight can be determined by the orientation of the bird’s body. However as stated above it is birds travelling back to colonies (after successful provisioning foray) that are most likely to give more accurate indication of true direction of origin. It is thus only birds carrying fish or birds with full crops (if this can be ascertained - some species carry food in the stomach) that should be included in any such data-gathering exercise. This is usually only possible with land-based or boat-based visual surveys.
Given the large uncertainty associated with information from this type of survey, once again we consider it is useful supporting evidence, rather than a method to allocate proportions of birds on development sites.
Integrating Theoretical and Empirical approaches
It is probable that some colonies under consideration for a development will have empirical (tagging) data whereas others will not. Most likely the largest, most important colonies (e.g. SPAs) will get this attention. In this case it would be sensible to try and use this information.
We suggest at present that theoretical apportioning should be applied to all colonies under consideration, and then the apportioning that is allocated to sites with tagging data may be re-apportioned depending on the results of the tracking analysis. For instance, site-specific data may show that the birds’ behaviour grossly violates assumptions e.g. they forage in a completely different direction.
Tracking data can give results for the number of birds entering the development area, the time spent within the development area, or the total number of tracks that reach the development site. To compare with on-site counts which use the mean peak numbers of birds on site, we suggest that a proportion calculated from the mean daily amount of time spent on the development site by tagged birds, weighted by the size of the colony, is used to apply to the proportion already allocated to the sites with tagged birds. This calculation would require knowledge of the time spent by each individual bird within the development area for each day of the tracking / tagging period.
Example Table 2 From Theoretical Apportioning (see Example 1 above) the resulting summed values are:
Non-tagged sites – combined proportion 0.65
Tagged sites - combined proportion 0.35
There are three sites with tracked birds. Using the data from each bird a calculated mean proportion of each day spent within the development area is known.
This requires information from each tag and the length of each foraging day (e.g. daylight period).
For each site the population size is known, the sample size of tagged birds is assumed to be adequate and selected from a random sample of breeding locations within the site. The tracking period should be representative of the breeding season and comparable for all sites.
| Site | Colony population | Mean proportion of day each tagged bird present on development area | Proportion of Day * Colony Population | Final proportion from tagging data | Theoretical proportion * Tagging proportion |
|---|---|---|---|---|---|
| A | 5000 | 0.187465 | 937.325 | 0.566 | 0.198 |
| B | 6000 | 0.11684 | 701.040 | 0.423 | 0.148 |
| C | 10000 | 0.001823 | 18.230 | 0.011 | 0.004 |
| SUM | 1656.595 |
The calculation is made as follows:The table may be edited (to see effects of changing the values of proportion of day each bird is on development) by double clicking in any cell. In the pdf version of this document use the accompanying spreadsheet.
Step 1 – Each Site weight is calculated : Colony Population *(Time birds active in development area /Sum of all time birds active)
Step 2 - Proportion for individual site within tagging group calculated: Site weight / Sum of all site weights
Step 3 – The final proportion calculated : site proportion within group * group proportion
In the first line of the table above (for colony A) the figures are:
Site weight : (5000*(305/1625)) = 937
Within group proportion : 937 /1657 = 0.566
Final proportion: 0.566*0.35 = 0.198
Concluding remarks
The process of apportioning impacts across multiple SPAs is necessarily an approximate one, ideally undertaken with empirically derived site-specific data. However, the data required to account for intra- and inter-annual variation in foraging behaviour will be considerable in most situations and onerous to collect. In the absence of good data, we recommend a theoretical approach though supplementary site-specific data may allow more realistic apportioning. However, tracking studies have shown that some seabirds will travel very long distances, to particular foraging locations. FAME project data have shown that seabirds from some Orkney breeding colonies are known to travel south into the outer Moray Firth, to feed in very specific areas (most likely targeting prey aggregations). Manx shearwaters breeding on Rum travel long distances south into the Irish Sea, whereas many fewer birds venture far north. These studies illustrate such models of feeding distribution are likely to be over-simplifications.
Our collective understanding of the journeys made by foraging seabirds is improving rapidly, and new techniques based on this improving information may soon replace the approach advocated here. At present we recommend that apportioning impacts arising from marine renewables developments are assessed using the methods described above, even if they are relatively simple approximations.
References
Ainsley D G, Ribic C A , Ballard G, Heath S, Gaffney I, Karl B J , Barton K J, Wilson P R, and Webb S (2004) Geographical structure of Adeile penguin populations: Overlap in colony specific feeding areas. Ecological Monographs 70: 159-178
Baron D G et al (2010) Meta-Analysis of transmitter effects on avian behaviour and ecology. Methods in Ecology and Evolution 1: 180 – 187
Cairns D. K. (1989) The Regulation of Seabird Colony Size: A Hinterland Model. The American Naturalist 134: 141-146
Chabrzyk G (1979) The effects of age on the breeding biology and recruitment of the herring gull (Larus argentatus). PhD Thesis, Durham University
Dean B, Freeman R, Kirk H, Leonard K, Phillips R, Perrins C M, and Guilford T (2013) Behavioural mapping of a pelagic seabird combining multiple sensors and a hidden Markov model reveals the distribution of at sea behaviour. J Royal Soc Interface 10 (20130279)
Furness R. W. and Birkhead T. R. (1984) Seabird colony distributions suggest competition for food supplies during the breeding season. Nature 311: 655–656
Elliott K.H., Woo K.J., Gaston A. J., Benvenuti S., Dall’Antonia L., and Davoren G. K. (2009) Central-place Foraging in an Arctic Seabird Provides Evidence for Storer-Ashmole’s Halo. The Auk 126: 613−625
Gaston A J, Ydenberg R C, Smith G E J (2007) Ashmole’s Halo and population regulation in seabirds. Marine Ornithology 35: 119-126
Goulson and Osborne (2009) Foraging Range and the Spatial Distribution of worker bumble bees. In Food Exploitation by social insects eds Jarau and Hrncir CRC Press
Michael P. Harris, Francis Daunt, Mark Newell, Richard A. Phillips and Sarah Wanless (2009) Wintering areas of adult Atlantic puffins Fratercula arctica from a North Sea colony as revealed by geolocation technology. Marine Biology DOI 10.1007/s00227-009-1365-0
Perrow, M. R., Skeate, E. R. and Gilroy, J. J. (2011) Visual tracking from a rigid-hulled inflatable boat to determine foraging movements of breeding terns. Journal of Field Ornithology 82: 68–79
Thaxter C.B., Lascelles B., Sugar K., Cooke A.S.C.P., Roos S., Bolton M., Langston R.H.W and Burton N.H.K. (2012) Seabird Foraging Ranges as a tool for identifying candidate Marine Protected Areas. Biological Conservation doi 10.10161j.biocon.2011.12.009
Wakefield ED; Bodey TW; Bearhop S; Blackburn J; Colhoun K; Davies R; Dwyer RG; Green J; Gremillet D; Jackson AL; Jessop MJ; Kane A; Langston RHW; Lescroel A; Murray S; Le Nuz M; Patrick SC; Peron C; Soanes L; Wanless S; Votier SC; Hamer KC (2013) Space partitioning without territoriality in gannets. Science 341: 68-70
Wakefield, E.D., Owen, E., Baer, J., Carroll, M.J., Daunt, F., Dodd, S.G, Green, J.A., Guilford, T., Mavor, R.A., Miller, P. I., Newell, M.A., Newton, S.F., Robertson, G. S., Shoji, A., Soanes, L. M., Votier, S.C., Wanless, S., and Bolton, M. 2017. Breeding density, fine-scale tracking, and large-scale modelling reveal the regional distribution of four seabird species. Ecological Applications 27.(7) pp. 2074-2091
Vandenabeele, S.P., Shepard, E.L., Grogan, A. & Wilson, R.P. (2012). When three per cent may not be three per cent; device-equipped seabirds experience variable flight constraints. Marine Biology 159: 1-14.
1 Normally using the geometric centre of both and without crossing substantial areas of land
2 Future of the Atlantic Marine Environment (FAME)
3 For Seabird Colony Register data
4 Cheaper GPS tags are widely available now, though require birds to be re-caught and tags removed. Tags that down-load data automatically to a nearby base-station are also available but are more expensive.
Published: 2018
Document downloads
Disclaimer: Scottish Natural Heritage (SNH) has changed its name to NatureScot as of the 24th August 2020.
At the time of publishing, this document may still refer to Scottish Natural Heritage (SNH) and include the original branding. It may also contain broken links to the old domain.
If you have any issues accessing this document please contact us via our feedback form.
Contacts
Marine Protected Areas






