Official Statistics - Scottish Terrestrial Breeding Birds 1994 – 2021
Published: 2022
An Official Statistics Publication for Scotland
Scotland’s terrestrial breeding birds occupy a wide range of habitats including farmland, upland, woodland and urban areas. Some are found in one particular habitat, for example coal tit is a typical woodland bird. Others use a wider range of habitats, such as skylark, which use farmland and upland, or great tit which uses woodland and farmland. Bird populations can respond relatively quickly to drivers, such as changes in habitat extent or condition, which are reflected in breeding success, survival or dispersal (moving from one area to another). Since birds are well surveyed through volunteer-based and professional surveys, and many are widespread and abundant, they are often used as indicators of environmental change.
Evidence
We monitor terrestrial breeding birds primarily through the volunteer-led Breeding Bird Survey (Harris et al. 2021). Randomly located 1km survey squares are visited twice each breeding season (April to July). These squares are intended to be representative of Scotland’s habitats including farmland, woodland, upland and urban areas and we examined bird population trends on each of these habitats. We also examined bird population trends on four types of farmland: arable, pastoral, woody farmland and wet farmland. Ten of the 66 species are assessed using targeted surveys, as they are either too scarce for reliable trends through BBS or are better monitored by specialised surveys.
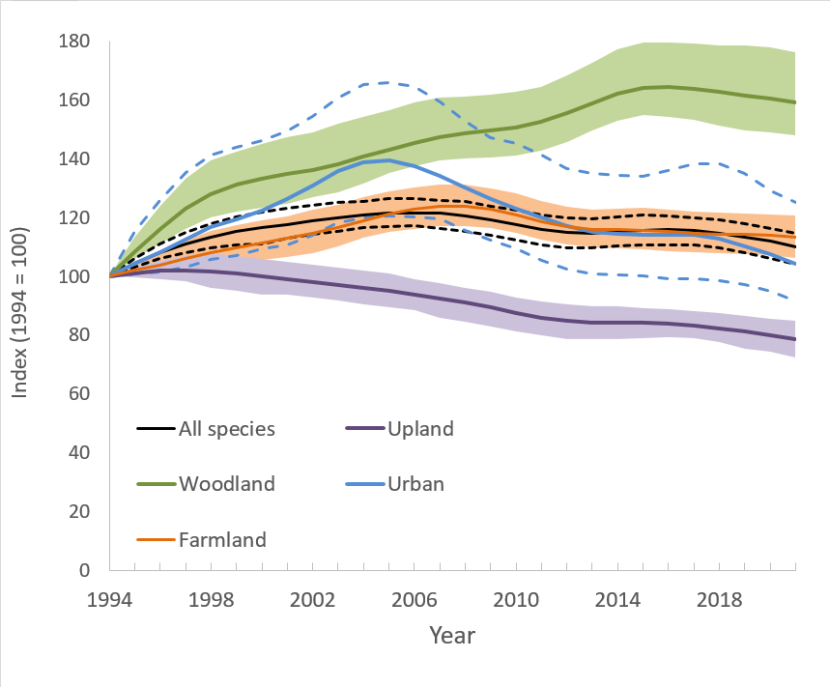
Figure 1. Smoothed Index of Abundance for Scottish Terrestrial Breeding Birds, 1994 to 2021.
Click for a full description
Dashed lines (for all species trend and urban trend) and shaded areas (for other habitat-specific trends) illustrate 95% confidence intervals.
The all-species (smoothed) index increased steadily up to the mid-2000s, but subsequently declined to fluctuate between 10% and 17% above the 1994 index. It had been declining (although only marginally) since 2016 and it is currently 10% higher than in 1994.
The smoothed woodland bird index increased significantly, by 59%, although it has been relatively stable since its peak in 2016.
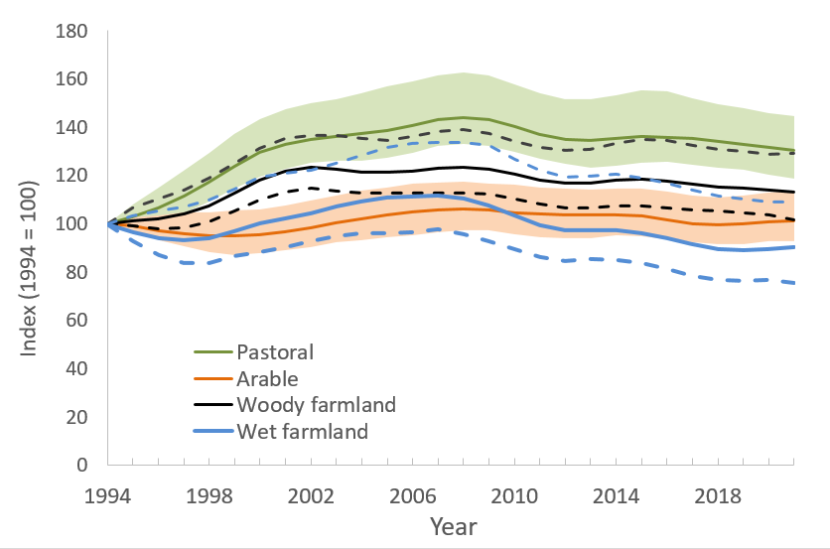
Figure 2. Index of Abundance for Scottish Farmland Breeding Birds, 1994 to 2021.
Click for a full description
Solid lines are smoothed trends. Shaded areas (for arable and pastoral farmland birds) and dashed lines (wet and woody farmland) illustrate 95% confidence intervals.
The smoothed farmland bird index increased steadily up to the late-2000s, peaking at 25% above the 1994 index value. It is currently 13% higher than in 1994. The increase in the farmland bird index was largely due to an increase in the pastoral bird index, which is currently 30% higher than in 1994 and the woody farmland bird index (13% higher than 1994)
Assessment
Since the start of the time series in 1994 to the most recent estimate in 2021:
- Of the 66 bird species 33 increased, 27 declined and 6 remained stable (i.e. within 5% of 1994 value) (Table 1).
- The all-species (smoothed) index increased steadily up to the mid-2000s, but subsequently declined to fluctuate between 10% and 17% above the 1994 index. It had been declining (although only marginally) since 2016 and it is currently 10% higher than in 1994 (Fig. 1).
- The smoothed woodland bird index increased significantly, by 59%, although it has been relatively stable since its peak in 2016 (Fig. 1).
- The smoothed farmland bird index increased steadily up to the late-2000s, peaking at 25% above the 1994 index value. It is currently 13% higher than in 1994. The increase in the farmland bird index was largely due to an increase in the pastoral bird index, which is currently 30% higher than in 1994 and the woody farmland bird index (13% higher than 1994) (Fig. 2). The arable bird index has been largely stable while the wet farmland bird index is currently 9% lower than in 1994.
- The smoothed upland bird index decreased by 21%.
- The smoothed urban bird index increased steadily until the mid-2000s, followed by a steady decline. It is currently 5% higher than in 1994.
- The unsmoothed data show that, between 2019 and 2021 (the short-term change), the all-species index, farmland bird index and upland bird index remained stable (i.e. have not changed more than 5% since last reported). While the all-species index remained stable, more species declined over than last two year than increased (26 species increased, 19 declined). The woodland bird index increased by 7%, while the urban index decreased by 9%.
Understanding the changes
In this commentary, ‘long-term’ refers to the period from 1994 to 2021 while ‘short-term’ refers to changes between 2019 and 2021. Longer-term changes since the 1970s, analogous to those reported for the UK (e.g. DEFRA, 2021; Harris et al., 2021) are not available for Scotland due to low monitoring coverage prior to the start of the BBS in 1994. The main drivers of population change are often unclear, but information is provided, where available, which may help to explain some of the likely causes. Breeding bird populations in Scotland are, as elsewhere, affected by conditions in all parts of their range; in the non-breeding, as well as the breeding season. For example, a summer migrant to Scotland may be affected by conditions in Scotland, along its migration route, and/or on its wintering grounds. Long-term and short-term changes for individual species are assessed using unsmoothed trends. Due to annual variability in the individual species trends, populations are considered to have increased or decreased only when the change is at least 5%. Individual species trends are shown in Table 1. A further 46 terrestrial species regularly breed in Scotland but have generally been too scarce to be monitored by the BBS effectively and are not covered by species-specific surveys. Among these species are several woodland, farmland, upland and urban specialists of current conservation concern (Eaton et al., 2015) such as wood warbler, spotted flycatcher and whinchat.
The latest results show the smoothed all-species index steadily increased since 1994, peaking in 2006 then becoming more variable after this time (Fig. 1). In 2021, it was 11% below the peak in 2006 and 10% higher than in 1994. The all-species indicator correlates positively with the mean annual Scottish temperature (June in the year preceding monitoring – May in the year of monitoring, Met Office et al., 2018). The temperature from the year prior to this, explains 75% of the annual variation (parameter estimate from a linear model of annual index varying with (a) mean annual temperature = 0.097 ± 0.013, t = 7.70, P < 0.001 and (b) mean annual temperature from 2 year – 1 year prior to monitoring = 0.051 ± 0.013, t = 4.07, P < 0.001) (Fig. 3). When temperature effects were accounted for (i.e. the residuals of a model of bird index varying with annual temperature and lagged annual temperature), the all-species index remained relatively stable (Fig. 4). The unexplained variation in the bird indexes have multiple causes but land-use change or other aspects of climate, such as extreme events, are considered major drivers. The woodland, farmland and urban indicators also correlated positively with annual temperature but the upland indicator did not, and instead correlated negatively with summer rainfall.
The two indicators with notable changes since 1994 are woodland which increased by 59%, and upland which declined by 21%. The woodland, farmland, and urban indicators all have positive correlations with temperature (and lagged temperature for farmland birds), but the upland indicator does not. When temperature effects were accounted for, there was still a 39% rise in the woodland indicator, while the urban indicator showed a 14% decline (Fig. 4). The farmland bird index remained stable when temperature effects were accounted for.
In the short-term, the all-species, farmland and upland indicators remained relatively stable while the woodland indicator has continued to rise in the short-term and declines were seen in the urban indicator.
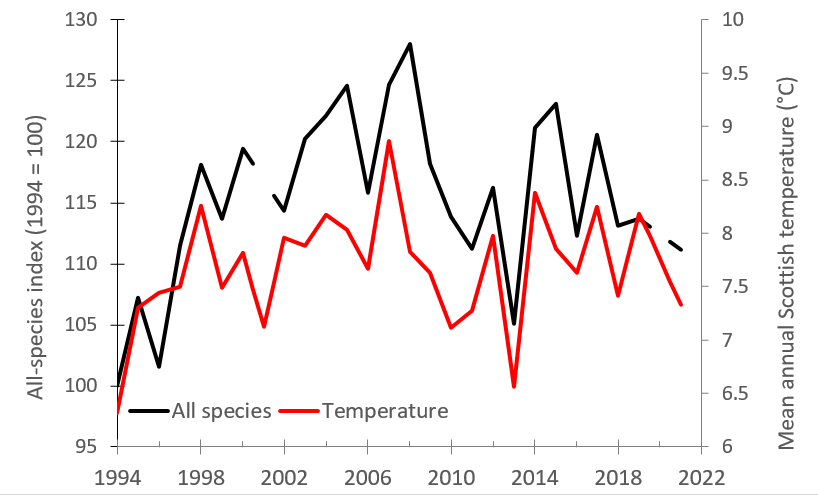
Figure 3. The all-species bird index and mean annual Scottish temperature.
Click for a full description
The all-species bird index (black lines) and mean annual Scottish temperature from June in the previous year to May in the stated year (°C, right-hand axis) from 1994 – 2021 (Met office et al., 2018).
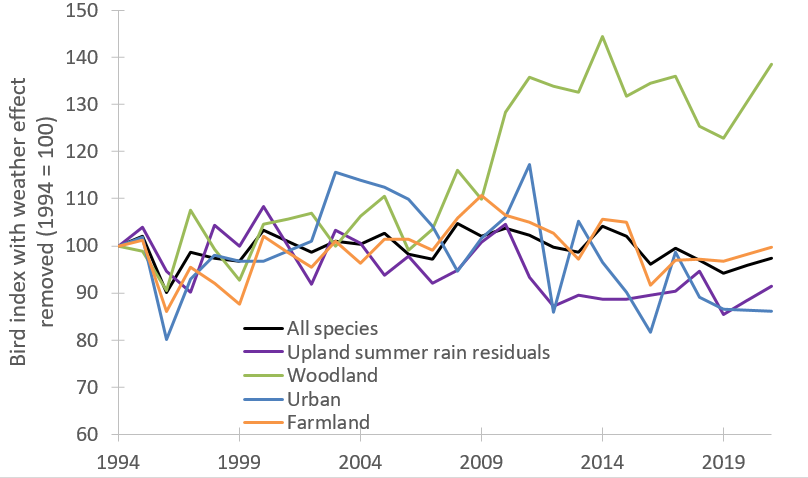
Figure 4. Index of Abundance for Scottish Terrestrial Breeding Birds, 1994 to 2021 with weather effect removed.
Click for a full description
The residuals of linear models of the bird indices varying with mean annual Scottish weather from both years preceding monitoring. The upland bird index was modelled against summer rainfall from both the years preceding monitoring. The residuals of each linear model were subtracted from the 1994 residual value and 100 was added to make the changes comparable to the standard indices.
Woodland
Over the long-term, woodland bird species have increased more than the other groupings. The woodland index correlates with the annual temperature (0.173 ± 0.066, t = 2.62, P = 0.0151) but annual temperatures from the two years prior to monitoring only explain 27% of the variation. Once the temperature effects are accounted for, there is still a 39% increase in the woodland bird index. This contrasts to the English woodland index which declined by 21% over the same period (DEFRA 2021). Twenty of the 23 species in the woodland indicator have increased since 1994. The biggest increases, of over 500%, are for chiffchaff, great spotted woodpecker and blackcap. Bullfinch, treecreeper, cuckoo, tree pipit, willow warbler, dunnock, great tit and mistle thrush have also increased by more than 50%. The biggest long-term decrease (-76%) is for capercaillie. The only other species to decline in woodland was chaffinch (-12%). The chaffinch decline is likely to be due to the disease Trichomonas gallinae (Robinson et al., 2010). Capercaillie declines have been linked to death from collisions with fences and potentially a negative effect of rising temperatures (Moss et al., 2001).
Willow warbler and chiffchaff are among Scotland’s most common woodland migrant birds. Increases in these species are likely to be driven by increasing temperatures (Martay et al., 2022). Most of England has now exceeded the optimum breeding season temperature for willow warbler, linked to the decline of willow warbler populations in England. Average Scottish temperatures are now close to the optimum for willow warbler but still well below the optimum temperature for chiffchaff, so willow warbler may decline in coming years, if the temperature continues to rise (Martay et al., 2022). The impacts of land-use change and climate change outside of the breeding season for migrants are far less understood.
Over the short-term there has been a mixed pattern, with seven species with recent declines. Most of these were small-bodied resident species which are particularly vulnerable to cold weather, including goldcrest, wren, great tit, coal tit and robin. The winter of 2020-2021 was the coldest since 2010 so may have caused these short-term declines. Capercaillie are monitored roughly every 5 years and experienced a substantial drop from around 1100 individuals in 2016 to 500 individuals in 2021. The greatest short-term increases (>30% increase) were seen in treecreeper, mistle thrush, great spotted woodpecker, dunnock and chiffchaff.
Farmland
Overall, there has been a long-term increase of 13% among the species trends that contribute to the farmland bird indicator. This increase was largely due to positive population trends on pastoral farmland. The higher trends on pastoral farmland compared to arable farmland were found across most species: 65% of species that contributed to both the pastoral and arable indicators had more positive trends on pastoral farmland (Table 2). Species that did not fit this pattern and were increasing on arable farmland more rapidly than on pastoral farmland included great tit and whitethroat, while starling and sedge warbler were increasing on arable farmland but declining on pastoral (Table 2).
The greatest long-term increases have been for goldfinch (+400%) and great tit (+154%). Magpie and whitethroat populations on farmland also increased by more than 100% so these species are more than twice as abundant on farmland as they were in 1994. There have been some substantial long-term decreases in other farmland bird populations, however, including declines of more than 50% for kestrel, lapwing and greenfinch, and 25-50% declines in oystercatcher and rook. Some farmland species such as grey partridge, corn bunting and tree sparrow do not contribute to this indicator because of insufficient coverage of these species by the BBS. The 2007-11 Bird Atlas (Balmer et al., 2013) suggests long-term declines for the former two species and substantial increases for the latter in Scotland.
The farmland bird index correlated positively with annual temperature and lagged annual temperature, explaining 45% of variation. When temperature effects were accounted for there was no long-term trend in the farmland bird index (-0.4% change), although a gradual rise until the late-2000s followed by a decline was apparent. Long-term changes in Scottish and UK farmland bird populations have also been driven by a range of factors including agricultural intensification, reduced diversity of crop types at the within-farm scale and reductions in spring-sown crops (Donald et al., 2001; Bell and Calladine, 2017). Agri-environment schemes have been in place in Scotland for around two decades, with results varying between different species. Targeted measures, such as the provision of cover crops has been shown to benefit seed eating birds including goldfinch, linnet and yellowhammer (Foster et al. 2013). A recent study in northeast Scotland found positive associations between bird abundance and specific land management options that met species' ecological requirements, in particular reed bunting and water margins, and yellowhammers and species‐rich grasslands. This highlights the importance of farm‐ and field‐scale targeting of management within AES (Daskalova et al. 2019). In addition, some farmland birds (e.g. goldfinch) may be benefitting from garden feeding (Plummer et al., 2019).
The most surprising short-term changes in farmland bird trends was the 65% increase in kestrel between 2019 and 2021 and the 12% increase in greenfinch, two species with extreme long-term declines. Greenfinch populations have been very badly impacted by the same disease that affects chaffinch (Hanmer et al., 2022) so this recent increase could indicate the start of some recovery from this disease. However, it will require several more years of monitoring to identify whether this is true, or just normal annual fluctuations. Jackdaw, whitethroat and reed bunting all had short-term declines despite large long-term increases. For jackdaw, this was a return to a usual population size after an unusually high 2019 population, while for the others, these were within normal levels of annual fluctuation.
Upland
Upland birds experienced the greatest decline between 1994 and 2021, of 21%. The upland index was the only index that did not correlate with annual temperature. However, when we examined correlations with seasonal temperature and rainfall, we found that the upland index correlated negatively with both summer rainfall in the year prior to monitoring and two years prior to monitoring, explaining 43% of the variation in the upland index. Summer rainfall was the only seasonal weather factor to have a significant change in Scotland between 1993 and 2020, with rainfall increasing from an average of 237mm of rain annually from June-August from 1993 – 1997 compared to an average of 363mm from 2016 – 2020. When the effect of summer rain was accounted for the upland index still declined by 9%.
Seventeen species contribute to the upland bird indicator, and of these twelve are in significant long-term decline. Four species (dotterel, curlew, black grouse and hooded crow) have declined by more than 50%. Long-term changes in upland bird populations have been driven by a number of factors including climate change, forest expansion, and changes in site-based management practices such as grazing and predator control (Buchanan et al., 2017). The link between increasing summer rainfall and upland bird declines supports the hypothesis that climate change is driving some of the long-term changes in upland bird populations. Changes in numbers of golden plover have been linked to climate change, in part due to role of increasing temperatures on the decline of craneflies during the breeding season (Pearce-Higgins et al., 2010). Likewise, curlew abundance and trends have been found to be positively associated with cooler temperatures and higher summer rainfall in a previous study (Franks et al., 2017). Our results indicate that there may be other climate change mechanisms at play - an increase in summer rain would be expected to increase cranefly populations. The negative correlation between summer rainfall and abundance was present for six of the twelve upland species with annual surveys including golden plover and curlew could be due to increased chick mortality with increasing summer rain (Eglington et al., 2010) but would merit further investigation.
One of the biggest findings to emerge from the last Bird Atlas 2007-11 (Balmer et al. 2013) is the disappearance of many wader species from much of their former breeding ranges, consistent with the declines reported by the Breeding Bird Survey in Scotland and across the UK. Concern about both upland and lowland breeding wader declines has resulted in a wide range of research and conservation initiatives, with many of those in Scotland highlighted on the Working for Waders hub. Much research is underway to identify factors affecting the rapidly declining curlew populations, with evidence suggesting interacting effects of climate change, land use change and predation (Douglas et al., 2014; Franks et al., 2017). In contrast, raven and cuckoo populations have increased, by more than 100% since 1994 and two other species (snipe and golden eagle) have increased by more than 25%.
Some other species breeding in the uplands such as whinchat and ring ouzel do not currently contribute to the indicator because of insufficient coverage by the BBS. Both of these species are in long-term decline (Balmer et al., 2013; Henderson et al., 2014; Wotton et al., 2016).
The only short-term increases of more than 20% were for snipe and dipper, possibly indicating a bounce-back from large short-term declines in the previous year. Snipe population have shown long-term increases while dipper populations have declined.
Urban areas
Overall, there has been a small long-term increase of 5% among the species trends that contribute to the urban bird indicator. This is largely due to an increase until the mid-2000s followed by a slow decline. The greatest long-term increases have been for blackcap (+176%), jackdaw (+132%), magpie (+131%) and woodpigeon (+115%). There have been some substantial long-term decreases in other urban bird populations, however, including declines of more than 50% for greenfinch and swift, and 25-50% declines in starling, blackbird and collared dove.
The urban bird index correlated positively with annual temperature, explaining 32% of variation. After accounting for weather effects, the urban index declined by 14% between 1994 – 2021, largely occurring since 2010. This could indicate that climate change has benefitted urban birds but other changes have been detrimental. Possible drivers of the decline include changes in bird communities due to increased garden bird feeding (Plummer et al., 2019) or loss of green space.
The urban bird index declined substantially in the past two years, by 9%. This is partly driven by large declines in the two migratory urban species – swift and house martin. Swift are experiencing a substantial long-term decline (-72%) which may be related to the reduced availability of suitable nesting sites on buildings (Woodward et al., 2018), though a reduction in availability of flying insects may also play a role (Hallmann et al., 2017). Swifts are long-distance migrants and there may also be factors that influence their survival outside the breeding season during migration or on their wintering grounds. Tracking studies in the UK and elsewhere in Europe have revealed that swifts from northern latitudes overwinter in different locations in sub-Saharan Africa than those that breed further south (Åkesson et al. 2020). In contrast, there have been long-term increases for house martin. As these species are both aerial insectivores, there may have been low insect availability due to the dry conditions over the monitoring period (April – June) in 2021. There were also short-term declines in many of the smaller species such as blue tit, house sparrow, wren, great tit and goldfinch, which may be associated with the relatively cold winter of 2020-2021.
Source data and updates
Data for 56 of the 66 species come from the Breeding Bird Survey (BBS) run by British Trust for Ornithology (BTO) on behalf of a partnership that also includes the Joint Nature Conservation Committee (JNCC) and Royal Society for the Protection of Birds (RSPB). The data for BBS consist of annual counts made over a period of years at a series of sites. Generalised Linear Models were used to generate trends. To prevent short-term population variability and sampling error having an undue influence, trends were smoothed and long-term trends assessed using techniques recommended by Fewster et al. (2000). The smoothed indices were used to assess the significance of long-term changes and the unsmoothed indices were used to assess the significance of short-term changes. Details of the methods used to calculate the indices are available from http://www.bto.org/birdtrends2010/methodology.htm. Trends for common sandpiper and dipper are derived from the BTO Waterways Bird Survey and the Waterways Breeding Bird Survey. Grey heron trends are derived from the Heronries Census. Trends for seven species (golden eagle, hen harrier, peregrine, dotterel, corncrake, black grouse and capercaillie) are estimated from single-species surveys carried out periodically, during the period 1994 to 2021 as part of the SCARABBS (Statutory Conservation Agency/RSPB Annual Breeding Bird Scheme) programme. A further 46 terrestrial species regularly breed in Scotland (being found in at least 100 10km squares in Bird Atlas 2007-11), but insufficient data are available through current monitoring schemes to permit their inclusion in these indicators. In Scotland, the production of multi-species indicators is limited to a start date in 1994 as this is when BBS was first implemented, and achieved broad-scale representative coverage of many terrestrial species in Scotland. Changes reported through these indicators therefore exclude any changes that occurred prior to 1994, which based on UK data show marked declines in many farmland and woodland species.
Official Statistics are produced by professionally independent statistical staff in accordance with the Code of Practice for Statistics.
|
Species |
All species index 1994 - 2021 |
All species index 2019 - 2021 |
Woodland index 1994 - 2021 |
Woodland index 2019 - 2021 |
Farmland index 1994 - 2021 |
Farmland index 2019 - 2021 |
Upland index 1994 - 2021 |
Upland index 2019 - 2021 |
Urban index 1994 - 2021 |
Urban index 2019 - 2021 |
|---|---|---|---|---|---|---|---|---|---|---|
|
Chiffchaff |
1192 |
31 |
1192 |
31 |
- |
- |
- |
- |
- |
- |
|
Great Spotted Woodpecker |
651 |
41 |
651 |
41 |
- |
- |
- |
- |
- |
- |
|
Blackcap |
541 |
-4 |
500 |
-11 |
- |
- |
- |
- |
- |
- |
|
Goldfinch |
278 |
7 |
- |
- |
401 |
9 |
- |
- |
176 |
-10 |
|
Bullfinch |
152 |
13 |
152 |
13 |
- |
- |
- |
- |
- |
- |
|
Cuckoo |
118 |
8 |
84 |
10 |
- |
- |
134 |
11 |
- |
- |
|
Whitethroat |
112 |
-15 |
- |
- |
112 |
-15 |
- |
- |
- |
- |
|
Magpie |
109 |
-1 |
- |
- |
123 |
3 |
- |
- |
131 |
3 |
|
Raven |
105 |
-23 |
- |
- |
- |
- |
105 |
-23 |
- |
- |
|
House Martin |
98 |
-31 |
- |
- |
- |
- |
- |
- |
98 |
-31 |
|
Treecreeper |
87 |
57 |
87 |
57 |
- |
- |
- |
- |
- |
- |
|
Corncrake |
86 |
-2 |
- |
- |
86 |
-2 |
- |
- |
- |
- |
|
Stonechat |
83 |
-25 |
- |
- |
- |
- |
- |
- |
- |
- |
|
Tree Pipit |
80 |
-3 |
80 |
-3 |
- |
- |
- |
- |
|
|
|
Great Tit |
73 |
-4 |
60 |
-12 |
154 |
-6 |
- |
- |
68 |
-19 |
|
Willow Warbler |
72 |
6 |
72 |
6 |
- |
- |
- |
- |
- |
- |
|
Mistle Thrush |
57 |
49 |
57 |
49 |
- |
- |
- |
- |
- |
- |
|
Reed Bunting |
54 |
-10 |
- |
- |
54 |
-10 |
- |
- |
- |
- |
|
House Sparrow |
48 |
2 |
- |
- |
- |
- |
- |
- |
-21 |
-22 |
|
Siskin |
48 |
26 |
48 |
26 |
- |
- |
- |
- |
- |
- |
|
Jackdaw |
45 |
-27 |
- |
- |
52 |
-30 |
- |
- |
132 |
11 |
|
Buzzard |
44 |
4 |
18 |
6 |
61 |
0 |
- |
- |
- |
- |
|
Lesser Redpoll |
38 |
-3 |
38 |
-3 |
- |
- |
- |
- |
- |
- |
|
Song Thrush |
37 |
4 |
47 |
8 |
63 |
-4 |
- |
- |
-15 |
14 |
|
Wren |
34 |
-12 |
34 |
-12 |
- |
- |
- |
- |
48 |
-10 |
|
Linnet |
32 |
47 |
- |
- |
32 |
47 |
- |
- |
- |
- |
|
Dunnock |
32 |
16 |
65 |
37 |
18 |
28 |
- |
- |
22 |
6 |
|
Blue Tit |
29 |
-4 |
15 |
-3 |
51 |
0 |
- |
- |
34 |
-17 |
|
Snipe |
29 |
29 |
- |
- |
- |
- |
29 |
29 |
- |
- |
|
Golden Eagle |
28 |
2 |
- |
- |
- |
- |
28 |
2 |
- |
- |
|
Yellowhammer |
24 |
-8 |
- |
- |
24 |
-8 |
- |
- |
- |
- |
|
Blackbird |
17 |
4 |
15 |
11 |
46 |
4 |
- |
- |
-51 |
-9 |
|
Goldcrest |
5 |
-34 |
5 |
-34 |
- |
- |
- |
- |
- |
- |
|
Collared Dove |
4 |
19 |
- |
- |
- |
- |
- |
- |
-42 |
9 |
|
Mallard |
4 |
2 |
- |
- |
- |
- |
- |
- |
- |
- |
|
Robin |
2 |
0 |
5 |
-6 |
- |
- |
- |
- |
42 |
10 |
|
Coal Tit |
0 |
-12 |
0 |
-12 |
- |
- |
- |
- |
- |
- |
|
Sedge Warbler |
-1 |
-12 |
- |
- |
-1 |
-12 |
- |
- |
- |
- |
|
Carrion Crow |
-4 |
7 |
- |
- |
8 |
3 |
- |
- |
-23 |
10 |
|
Hen Harrier |
-6 |
-3 |
- |
- |
- |
- |
-6 |
-3 |
- |
- |
|
Skylark |
-8 |
0 |
- |
- |
-17 |
0 |
-2 |
-9 |
- |
- |
|
Woodpigeon |
-8 |
-1 |
- |
- |
-17 |
-8 |
- |
- |
115 |
-3 |
|
Wheatear |
-8 |
16 |
- |
- |
- |
- |
-8 |
16 |
- |
- |
|
Chaffinch |
-9 |
-1 |
-12 |
-2 |
-2 |
-1 |
|
|
- |
- |
|
Meadow Pipit |
-11 |
0 |
- |
- |
|
|
-11 |
0 |
- |
- |
|
Pied Wagtail |
-15 |
0 |
- |
- |
-15 |
0 |
- |
- |
- |
- |
|
Willow/Red Grouse |
-15 |
-29 |
- |
- |
- |
- |
-15 |
-29 |
- |
- |
|
Golden Plover |
-18 |
-6 |
- |
- |
- |
- |
-18 |
-6 |
- |
- |
|
Swallow |
-19 |
-7 |
- |
- |
-19 |
-7 |
|
|
- |
- |
|
Peregrine |
-20 |
-2 |
- |
- |
- |
- |
-20 |
-2 |
- |
- |
|
Grey Wagtail |
-23 |
-32 |
- |
- |
- |
- |
- |
- |
- |
- |
|
Starling |
-23 |
-16 |
- |
- |
-6 |
-17 |
- |
- |
-40 |
-12 |
|
Grey Heron |
-25 |
-19 |
- |
- |
- |
- |
- |
- |
- |
- |
|
Rook |
-30 |
-7 |
- |
- |
-30 |
-7 |
- |
- |
- |
- |
|
Oystercatcher |
-36 |
8 |
- |
- |
-36 |
8 |
- |
- |
- |
- |
|
Dipper |
-37 |
24 |
- |
- |
- |
- |
-37 |
24 |
- |
- |
|
Common Sandpiper |
-42 |
-8 |
- |
- |
- |
- |
-42 |
-8 |
- |
- |
|
Hooded Crow |
-54 |
-7 |
- |
- |
- |
- |
-54 |
-7 |
- |
- |
|
Black Grouse |
-61 |
-7 |
- |
- |
- |
- |
-61 |
-7 |
- |
- |
|
Curlew |
-63 |
-18 |
- |
- |
- |
- |
-63 |
-18 |
- |
- |
|
Greenfinch |
-65 |
5 |
- |
- |
-61 |
12 |
- |
- |
-78 |
14 |
|
Lapwing |
-66 |
-18 |
- |
- |
-66 |
-18 |
- |
- |
- |
- |
|
Dotterel |
-69 |
-9 |
- |
- |
- |
- |
-69 |
-9 |
- |
- |
|
Kestrel |
-71 |
65 |
- |
- |
-71 |
65 |
- |
- |
- |
- |
|
Swift |
-72 |
-49 |
- |
- |
- |
- |
- |
- |
-72 |
-49 |
|
Capercaillie |
-76 |
-31 |
-76 |
-31 |
- |
- |
- |
- |
- |
- |
|
Species |
Arable farmland index 1994 - 2021 |
Arable farmland index 2019 - 2021 |
Pastoral farmland index 1994 - 2021 |
Pastoral farmland index 2019 - 2021 |
Woody farmland index 1994 - 2021 |
Woody farmland index 2019 - 2021 |
Wet farmland index 1994 - 2021 |
Wet farmland index 2019 - 2021 |
|---|---|---|---|---|---|---|---|---|
|
Goldfinch |
266 |
-3 |
-7 |
420 |
10 |
17 |
457 |
9 |
|
Great Tit |
261 |
-5 |
0 |
96 |
10 |
-11 |
116 |
7 |
|
Whitethroat |
157 |
7 |
12 |
75 |
13 |
-30 |
94 |
15 |
|
Song Thrush |
72 |
9 |
-7 |
69 |
18 |
-6 |
77 |
18 |
|
Starling |
54 |
-28 |
53 |
-15 |
16 |
-27 |
21 |
19 |
|
Buzzard |
53 |
2 |
-12 |
67 |
-3 |
18 |
53 |
4 |
|
Blue Tit |
52 |
17 |
-10 |
51 |
7 |
7 |
50 |
3 |
|
Blackbird |
40 |
1 |
4 |
55 |
1 |
3 |
44 |
1 |
|
Yellowhammer |
26 |
24 |
-24 |
28 |
-1 |
12 |
13 |
14 |
|
Linnet |
26 |
10 |
59 |
33 |
-22 |
45 |
9 |
2 |
|
Dunnock |
16 |
11 |
20 |
33 |
3 |
28 |
4 |
13 |
|
Jackdaw |
7 |
9 |
-24 |
102 |
88 |
-38 |
93 |
25 |
|
Skylark |
6 |
-4 |
8 |
-36 |
-17 |
-4 |
- |
- |
|
Chaffinch |
-11 |
2 |
-5 |
8 |
-4 |
1 |
-8 |
-3 |
|
Rook |
-13 |
-29 |
2 |
-24 |
42 |
-20 |
-3 |
36 |
|
Carrion Crow |
-13 |
-17 |
29 |
30 |
13 |
-11 |
4 |
13 |
|
Pied Wagtail |
-26 |
15 |
-11 |
8 |
5 |
10 |
-28 |
14 |
|
Woodpigeon |
-26 |
-25 |
-13 |
-14 |
-9 |
-10 |
-30 |
-18 |
|
Sedge Warbler |
-34 |
15 |
-18 |
25 |
-17 |
2 |
-18 |
13 |
|
Swallow |
-35 |
-13 |
-11 |
-5 |
-17 |
9 |
-31 |
-8 |
|
Oystercatcher |
-61 |
-5 |
-10 |
-31 |
-10 |
20 |
- |
- |
|
Greenfinch |
-63 |
3 |
49 |
-66 |
-23 |
2 |
-74 |
-22 |
|
Lapwing |
-65 |
45 |
8 |
-58 |
-21 |
-12 |
-74 |
-22 |
|
Corncrake |
- |
- |
- |
86 |
-3 |
-2 |
- |
- |
|
Magpie |
- |
- |
- |
97 |
33 |
1 |
107 |
16 |
|
Reed Bunting |
- |
- |
- |
231 |
-24 |
9 |
138 |
-37 |
References
Balmer, D.E., Gillings, S., Caffrey, B.J., Swann, R.L., Downie, I.S., & Fuller, R.J. 2013. Bird Atlas 2007-11: the breeding and wintering birds of Britain and Ireland (Thetford: BTO Books).
Bell, M.V. & Calladine, J. 2017. The Decline of a Population of Farmland Breeding Waders: a twenty-five year case study. Bird Study 64: 264-273.
Buchanan, G.M., Pearce-Higgins, J.W., Douglas, D.J.T. & Grant, M.C. 2017. Quantifying the importance of multi-scale management and environmental variables on moorland bird abundance. Ibis 159: 744-756.
Daskalova G.N., Phillimore A.B., Bell M., Maggs H.E. & Perkins A.J. 2019. Population responses of farmland bird species to agri-environment schemes and land management options in North eastern Scotland. Journal of Applied Ecology 56: 640– 650.
DEFRA 2021. Wild Bird Populations in England, 1970 to 2019 (Updated for Wintering Waterbirds). National Statistics, 9 December 2021.
Department for Environment, Food and Rural Affairs. 2019. Wild bird populations in the UK, 1970 to 2019 (Updated for Wintering Waterbirds): Annual statistical release. Defra National Statistics, London.
Donald, P.F., Green, R.E. & Heath, M.F. 2001. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proceedings of the Royal Society B: Biological Sciences 268: 25-29.
Douglas, D.J.T., Bellamy, P.E., Stephen, L.S. Pearce–Higgins, J.W., Wilson, J.D. & Grant, M.C. 2014. Upland land use predicts population decline in a globally near-threatened wader. Journal of Applied Ecology 51: 194-203.
Eaton, M., Aebischer, N., Brown, A., Hearn, R., Lock, L., Musgrove, A., ... & Gregory, R. 2015. Birds of Conservation Concern 4: the population status of birds in the UK, Channel Islands and Isle of Man. British Birds 108: 708-746.
Eglington, S.M., Bolton, M., Smart, M.A., Sutherland, W.J., Watkinson, A.R. & Gill, J.A. 2010. Managing water levels on wet grasslands to improve foraging conditions for breeding northern lapwing Vanellus vanellus. Journal of Applied Ecology 47: 451-458.
Fewster, R.M., Buckland, S.T., Siriwardena, G.M., Baillie, S.R. & Wilson, J.D. 2000. Analysis of population trends for farmland birds using generalized additive models. Ecology 81: 1970-1984.
Foster, S., Harrison, P., Buckland, S., Elston, D., Brewer, M., Johnston, A., Pearce‐Higgins, J. & Marrs, S. 2013. Trends of Breeding Farmland Birds in Scotland. Trend Note 022. Scottish Natural Heritage.
Franks, S.E., Douglas, D.J., Gillings, S. & Pearce-Higgins, J.W. 2017. Environmental correlates of breeding abundance and population change of Eurasian Curlew Numenius arquata in Britain. Bird Study 64: 393-409.
Hanmer, H.J., Cunningham, A.A., John, S.K., Magregor, S.K., Robinson, R.A., Seilern-Moy, K., ... & Lawson, B. 2022. Habitat-use influences severe disease-mediated population declines in two of the most common garden bird species in Great Britain. Scientific reports 12: 1-13.
Harris, S.J., Massimino, D., Balmer, D.E., Eaton, M.A., Noble, D.G., Pearce-Higgins, J.W., ... & Gillings, S. 2021. The breeding bird survey 2020. BTO research report, 736.
Henderson, I., Calladine, J., Massimino, D., Taylor, J. & Gillings, S. 2014. Evidence for contrasting causes of population change in two closely related, sympatric breeding species the Whinchat Saxicola rubetra and Stonechat Saxicola torquata in Britain. Bird Study 61: 553-565.
Martay, B., Pearce‐Higgins, J.W., Harris, S.J., & Gillings, S. 2022. Breeding ground temperature rises, more than habitat change, are associated with spatially variable population trends in two species of migratory bird. Ibis.
Met Office, Hollis, D., McCarthy, M., Kendon, M., Legg, T. & Simpson, I. 2018. HadUK-Grid gridded and regional average climate observations for the UK. Centre for Environmental Data Analysis, accessed 20/10/2022.
Moss, R., Oswald, J., & Baines, D. 2001. Climate change and breeding success: decline of the capercaillie in Scotland. Journal of Animal Ecology, 2001: 47-61.
Pearce‐Higgins, J.W., Dennis, P., Whittingham, M.J., & Yalden, D.W. 2010. Impacts of climate on prey abundance account for fluctuations in a population of a northern wader at the southern edge of its range. Global Change Biology 16: 12-23.
Plummer, K.E., Risely, K., Toms, M.P. & Sirawrdena, G. M. 2019. The composition of British bird communities is associated with long-term garden bird feeding. Nature Communications 10: 2088.
Robinson, R.A., Lawson, B., Toms, M.P., Peck, K.M., Kirkwood, J.K., Chantrey, J., ... & Cunningham, A.A. 2010. Emerging infectious disease leads to rapid population declines of common British birds. PLoS one 5: e12215.
Tyler, G.A. & Green, R.E. 2004. Effects of weather on the survival and growth of Corncrake Crex crex chicks. Ibis 146: 69-76.





